
Title: Bodily changes in pain, hunger, fear, and rage
an account of recent researches into the function of emotional excitement
Author: Walter B. Cannon
Release date: June 28, 2024 [eBook #73932]
Language: English
Original publication: New York: D. Appleton and Company
Credits: Peter Becker, Neil Mercer and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive)
Transcriber's Note
New original cover art included with this eBook is granted to the public domain.
The List of Illustrations following the "Contents" section has been added by the transcriber; the wording of captions in that List has in some cases been condensed from the captions in the body of the book.
BODILY CHANGES
IN PAIN, HUNGER,
FEAR AND RAGE
AN ACCOUNT OF RECENT RESEARCHES INTO THE FUNCTION OF EMOTIONAL EXCITEMENT
BY
WALTER B. CANNON
GEORGE HIGGINSON PROFESSOR OF PHYSIOLOGY IN HARVARD UNIVERSITY
NEW YORK AND LONDON
D. APPLETON AND COMPANY
1915
Copyright, 1915, by
D. APPLETON AND COMPANY
Printed in the United States of America
TO MY COLLABORATORS IN THESE RESEARCHES
DANIEL DE LA PAZ
ALFRED T. SHOHL
WADE S. WRIGHT
ARTHUR L. WASHBURN
HENRY LYMAN
LEONARD B. NICE
CHARLES M. GRUBER
HOWARD OSGOOD
HORACE GRAY
WALTER L. MENDENHALL
WITH PLEASANT MEMORIES OF OUR WORK TOGETHER
[p. vii]
Fear, rage and pain, and the pangs of hunger are all primitive experiences which human beings share with the lower animals. These experiences are properly classed as among the most powerful that determine the action of men and beasts. A knowledge of the conditions which attend these experiences, therefore, is of general and fundamental importance in the interpretation of behavior.
During the past four years there has been conducted, in the Harvard Physiological Laboratory, a series of investigations concerned with the bodily changes which occur in conjunction with pain, hunger and the major emotions. A group of remarkable alterations in the bodily economy have been discovered, all of which can reasonably be regarded as responses that are nicely adapted to the individual’s welfare and preservation. Because these physiological adaptations are interesting both in themselves and in their interpretation, not only to physiologists and psychologists, but to others as well, it has seemed worth while to gather together in convenient form the original accounts of the experiments, which have been published in various American medical and physiological journals. I have, however, attempted to arrange the results and discussions in an orderly and consecutive manner, and I have tried also to eliminate[p. viii] or incidentally to explain the technical terms, so that the exposition will be easily understood by any intelligent reader even though not trained in the medical sciences.
My first interest in the conditions attending pain, hunger and strong emotional states was stimulated during the course of a previous series of researches on the motor activities of the alimentary canal. A summary of these researches appeared in 1911, under the title, “The Mechanical Factors of Digestion.” The studies recorded in the present volume may be regarded as a natural sequence of observations on the influence of emotional states on the digestive process, which were reported in that volume.
W. B. Cannon.
[p. ix]
| CHAPTER I | |
| PAGES | |
| THE EFFECT OF THE EMOTIONS ON DIGESTION | |
| Emotions favorable to normal secretion of the digestive juices—Emotions unfavorable to normal secretion of the digestive juices—Emotions favorable and unfavorable to contractions of the stomach and intestines—The disturbing effect of pain on digestion | 1–21 |
| CHAPTER II | |
| THE GENERAL ORGANIZATION OF THE VISCERAL NERVES CONCERNED IN EMOTIONS | |
| The outlying neurones—The three divisions of the outlying neurones—The extensive distribution of neurones of the “sympathetic” or thoracico-lumbar division and their arrangement for diffuse action—The arrangement of neurones of the cranial and sacral divisions for specific action—The cranial division a conserver of bodily resources—The sacral division a group of mechanisms for emptying—The sympathetic division antagonistic to both the cranial and the sacral—Neurones of the sympathetic division and adrenal secretion have the same action | 22–39 |
| CHAPTER III[p. x] | |
| METHODS OF DEMONSTRATING ADRENAL SECRETION AND ITS NERVOUS CONTROL | |
| The evidence that splanchnic stimulation induces adrenal secretion—The question of adrenal secretion in emotional excitement—The method of securing blood from near the adrenal veins—The method of testing the blood for adrenin | 40–51 |
| CHAPTER IV | |
| ADRENAL SECRETION IN STRONG EMOTIONS AND PAIN | |
| The evidence that adrenal secretion is increased in emotional excitement—The evidence that adrenal secretion is increased by “painful” stimulation—Confirmation of our results by other observers | 52–65 |
| CHAPTER V | |
| THE INCREASE OF BLOOD SUGAR IN PAIN AND GREAT EMOTION | |
| Glycosuria from pain—Emotional glycosuria—The rôle of the adrenal glands in emotional glycosuria | 66–80 |
| CHAPTER VI | |
| IMPROVED CONTRACTION OF FATIGUED MUSCLE AFTER SPLANCHNIC STIMULATION OF THE ADRENAL GLAND | |
| The nerve-muscle preparation—The splanchnic preparation—The effects of splanchnic stimulation on the contraction of fatigued muscle—The first rise in the muscle record—The prolonged rise in the muscle record—The two factors: arterial pressure and adrenal secretion | 81–94 |
| CHAPTER VII[p. xi] | |
| THE EFFECTS ON CONTRACTION OF FATIGUED MUSCLE OF VARYING THE ARTERIAL BLOOD PRESSURE | |
| The effect of increasing arterial pressure—The effect of decreasing arterial pressure—An explanation of the effects of varying the arterial pressure—The value of increased arterial pressure in pain and strong emotion | 95–109 |
| CHAPTER VIII | |
| THE SPECIFIC RÔLE OF ADRENIN IN COUNTERACTING THE EFFECTS OF FATIGUE | |
| Variations of the threshold stimulus as a measure of irritability—The method of determining the threshold stimulus—The lessening of neuro-muscular irritability by fatigue—The slow restoration of fatigued muscle to normal irritability by rest—The quick restoration of fatigued muscle to normal irritability by adrenin—The evidence that the restorative action of adrenin is specific—The point of action of adrenin in muscle | 110–134 |
| CHAPTER IX | |
| THE HASTENING OF THE COAGULATION OF BLOOD BY ADRENIN | |
| The graphic method of measuring the coagulation time—The effects of subcutaneous injections of adrenin—The effects of intravenous injections—The hastening of coagulation by adrenin not a direct effect on the blood | 135–160 |
| CHAPTER X | |
| THE HASTENING OF COAGULATION OF BLOOD IN PAIN AND GREAT EMOTION | |
| Coagulation hastened by splanchnic stimulation—Coagulation not hastened by splanchnic stimulation if[p. xii] the adrenal glands are absent—Coagulation hastened by “painful” stimulation—Coagulation hastened in emotional excitement | 161–183 |
| CHAPTER XI | |
| THE UTILITY OF THE BODILY CHANGES IN PAIN AND GREAT EMOTION | |
| The reflex nature of bodily responses in pain and the major emotions, and the useful character of reflexes—The utility of the increased blood sugar as a source of muscular energy—The utility of increased adrenin in the blood as an antidote to the effects of fatigue—The question whether adrenin normally secreted inhibits the use of sugar in the body—The vascular changes produced by adrenin favorable to supreme muscular exertion—The changes in respiratory function also favorable to great effort—The effects produced in asphyxia similar to those produced in pain and excitement—The utility of rapid coagulation in preventing loss of blood | 184–214 |
| CHAPTER XII | |
| THE ENERGIZING INFLUENCE OF EMOTIONAL EXCITEMENT | |
| “Reservoirs of power”—The excitements and energies of competitive sports—Frenzy and endurance in ceremonial and other dances—The fierce emotions and struggles of battle—The stimulating influence of witnesses and of music—The feeling of power | 215–231 |
| CHAPTER XIII | |
| THE NATURE OF HUNGER | |
| Appetite and hunger—The sensation of hunger—The theory that hunger is a general sensation—Weakness of the assumptions underlying the theory that hunger is a general sensation—Body need may exist without hunger—The theory that hunger is of general[p. xiii] origin does not explain the quick onset and the periodicity of the sensation—The theory that hunger is of general origin does not explain the local reference—Hunger not due to emptiness of the stomach—Hunger not due to hydrochloric acid in the empty stomach—Hunger not due to turgescence of the gastric mucous membrane—Hunger the result of contractions—The “empty” stomach and intestines contract—Observations suggesting that contractions cause hunger—The concomitance of contractions and hunger in man | 232–266 |
| CHAPTER XIV | |
| THE INTERRELATIONS OF EMOTIONS | |
| Antagonism between emotions expressed in the sympathetic and in the cranial divisions of the autonomic system—Antagonism between emotions expressed in the sympathetic and in the sacral divisions of the autonomic system—The function of hunger—The similarity of visceral effects in different strong emotions and suggestions as to its psychological significance | 267–284 |
| CHAPTER XV | |
| ALTERNATIVE SATISFACTIONS FOR THE FIGHTING EMOTIONS | |
| Support for the militarist estimate of the strength of the fighting emotions and instincts—Growing opposition to the fighting emotions and instincts as displayed in war—The desirability of preserving the martial virtues—Moral substitutes for warfare—Physical substitutes for warfare—The significance of international athletic competitions | 285–301 |
| A LIST OF PUBLISHED RESEARCHES FROM THE PHYSIOLOGICAL LABORATORY IN HARVARD UNIVERSITY | 302–303 |
| INDEX | 305 |
| Figure | Page | |
|---|---|---|
| 1. | Diagram of the more important distributions of the autonomic nervous system | 25 |
| 2. | Diagram of the arrangements for recording contractions of the intestinal muscle | 49 |
| 3. | Intestinal muscle beating in inactive blood | 53 |
| 4. | Alternate application of “excited” blood and “quiet” blood, from the same animal, to intestinal muscle initially beating in Ringer’s solution | 55 |
| 5. | The effect of prolonging the excitement | 55 |
| 6. | Failure of the cava blood to produce inhibition when excitement has occurred after removal of the adrenal glands | 57 |
| 7. | Effect of adding adrenin to formerly inactive blood | 58 |
| 8. | The effect of bubbling oxygen through active blood | 59 |
| 9. | Intestinal muscle beating in normal vena cava blood | 62 |
| 10. | The shielded electrodes used in stimulating the splanchnic nerves | 87 |
| 11. | Contraction of the tibialis anticus and stimulation of the left splanchnic nerves | 89 |
| 12. | Arterial blood pressure with membrane manometer, contractions of tibialis anticus, and splanchnic stimulation | 91 |
| 13–17. | Effect of varying arterial blood pressure upon muscular contraction over time | 98–104 |
| 18. | Threshold stimulus of muscles during an experiment | 116 |
| 19. | Threshold stimulus of muscles during an experiment | 122 |
| 20. | Threshold stimulus of muscles during an experiment | 124 |
| 21. | Effect of adrenin injection upon blood pressure and contractions of the tibialis anticus muscle | 128 |
| 22. | Effect of amyl nitrite injection upon blood pressure and contractions of the tibialis anticus muscle | 128 |
| 23. | Effect of adrenin injection upon blood pressure and contractions of the tibialis anticus muscle when denervated | 131 |
| 24. | Diagram of the graphic coagulometer | 139 |
| 25. | Record of five successive tests of coagulation | 145 |
| 26. | Shortening of coagulation time after injection of adrenin | 151 |
| 27. | Differing effects upon the coagulation time of slow and rapid injections of adrenin | 153 |
| 28. | Persistent shortening of the coagulation time after injection of adrenin when brain and upper cord pithed | 154 |
| 29. | Shortening of coagulation time after stimulation of the left splanchnic nerves | 163 |
| 30. | Shortening of coagulation time after stimulation of the left splanchnic nerves | 164 |
| 31. | Results of stimulating the left splanchnic nerves after removal of the left adrenal gland; and of stimulating the right splanchnic nerves with right adrenal gland present | 169 |
| 32. | Three shortenings of coagulation time after stimulation of the left sciatic nerve | 174 |
| 33. | Shortening of coagulation time during an operation under light anesthesia | 175 |
| 34. | Record of rapid clotting after emotional excitement | 180 |
| 35. | Rapid clotting after emotional excitement, with slowing of the process when the splanchnic nerves were cut in the thorax | 182 |
| 36. | Adrenal secretion produced by asphyxia | 208 |
| 37. | Intragastric pressure, respiration and report of hunger pangs against time | 257 |
| 38. | The same conditions as in Fig. 37 | 259 |
| 39. | Compression of thin rubber bag in the lower esophagus, and report of hunger pangs against time | 260 |
BODILY CHANGES IN PAIN, HUNGER, FEAR AND RAGE
[p. 1]
THE EFFECT OF THE EMOTIONS ON DIGESTION
The doctrine of human development from subhuman antecedents has done much to unravel the complex nature of man. As a means of interpretation this doctrine has been directed chiefly toward the solving of puzzles in the peculiarities of anatomical structure. Thus arrangements in the human body, which are without obvious utility, receive rational explanation as being vestiges of parts useful in or characteristic of remote ancestors—parts retained in man because of age-long racial inheritance. This mode of interpretation has proved applicable also in accounting for functional peculiarities. Expressive actions and gestures—the facial appearance in anger, for example—observed in children and in widely distinct races, are found to be innate, and are best explained as the retention in human beings of responses which are similar in character in lower animals.
[p. 2]
From this point of view biology has contributed much to clarify our ideas regarding the motives of human behavior. The social philosophies which prevailed during the past century either assumed that conduct was determined by a calculated search for pleasure and avoidance of pain or they ascribed it to a vague and undefined faculty named the conscience or the moral sense. Comparative study of the behavior of men and of lower animals under various circumstances, however, especially with the purpose of learning the source of prevailing impulses, is revealing the inadequacy of the theories of the older psychologists. More and more it is appearing that in men of all races and in most of the higher animals, the springs of action are to be found in the influence of certain emotions which express themselves in characteristic instinctive acts.
The rôle which these fundamental responses in the higher organisms play in the bodily economy has received little attention. As a realm for investigation the bodily changes in emotional excitement have been left by the physiologists to the philosophers and psychologists and to the students of natural history. These students, however, have usually had too slight experience in the detailed examination of bodily functions to permit them to follow the clues which superficial observation might present. In consequence our[p. 3] knowledge of emotional states has been meager.
There are, of course, many surface manifestations of excitement. The contraction of blood vessels with resulting pallor, the pouring out of “cold sweat,” the stopping of saliva-flow so that the “tongue cleaves to the roof of the mouth,” the dilation of the pupils, the rising of the hairs, the rapid beating of the heart, the hurried respiration, the trembling and twitching of the muscles, especially those about the lips—all these bodily changes are well recognized accompaniments of pain and great emotional disturbance, such as fear, horror and deep disgust. But these disturbances of the even routine of life, which have been commonly noted, are mainly superficial and therefore readily observable. Even the increased rapidity of the heart beat is noted at the surface in the pulsing of the arteries. There are, however, other organs, hidden deep in the body, which do not reveal so obviously as the structures near or in the skin, the disturbances of action which attend states of intense feeling. Special methods must be used to determine whether these deep-lying organs also are included in the complex of an emotional[*] agitation.
*In the use of the term “emotion” the meaning here is not restricted to violent affective states, but includes “feelings” and other affective experiences. At times, also, in order to avoid awkward expressions, the term is used in the popular manner, as if the “feeling” caused the bodily change.
[p. 4]
Among the organs that are affected to an important degree by feelings are those concerned with digestion. And the relations of feelings to the activities of the alimentary canal are of particular interest, because recent investigations have shown that not only are the first stages of the digestive process normally started by the pleasurable taste and smell and sight of food, but also that pain and great emotional excitement can seriously interfere with the starting of the process or its continuation after it has been started. Thus there may be a conflict of feelings and of their bodily accompaniments—a conflict the interesting bearing of which we shall consider later.
The feelings or affective states favorable to the digestive functions have been studied fruitfully by Pawlow,[1] of Petrograd, through ingenious experiments on dogs. By the use of careful surgical methods he was able to make a side pouch of a part of the stomach, the cavity of which was wholly separate from the main cavity in which the food was received. This pouch was supplied in a normal manner with nerves and blood vessels, and as it opened to the surface of the body, the amount and character of the gastric juice secreted by it under various conditions[p. 5] could be accurately determined. Secretion by that part of the stomach wall which was included in the pouch was representative of the secretory activities of the entire stomach. The arrangement was particularly advantageous in providing the gastric juice unmixed with food. In some of the animals thus operated upon an opening was also made in the esophagus so that when the food was swallowed, it did not pass to the stomach but dropped out on the way. All the pleasures of eating were thus experienced, and there was no necessity of stopping because of a sense of fulness. This process was called “sham feeding.” The well-being of these animals was carefully attended to, they lived the normal life of dogs, and in the course of months and years became the pets of the laboratory.
By means of sham feeding Pawlow showed that the chewing and swallowing of food which the dogs relished resulted, after a delay of about five minutes, in a flow of natural gastric juice from the side pouch of the stomach—a flow which persisted as long as the dog chewed and swallowed the food, and continued for some time after eating ceased. Evidently the presence of food in the stomach is not a prime condition for gastric secretion. And since the flow occurred only when the dogs had an appetite, and the material presented to them was agreeable, the conclusion[p. 6] was justified that this was a true psychic secretion.
The mere sight or smell of a favorite food may start the pouring out of gastric juice, as was noted many years ago by Bidder and Schmidt[2] in a hungry dog which had a fistulous opening through the body wall into the stomach. This observation, reported in 1852, was confirmed later by Schiff and also still later by Pawlow. That the mouth “waters” with a flow of saliva when palatable food is seen or smelled has long been such common knowledge that the expression, “It makes my mouth water,” is at once recognized as the highest testimony to the attractiveness of an appetizing dish. That the stomach also “waters” in preparation for digesting the food which is to be taken is clearly proved by the above cited observations on the dog.
The importance of the initial psychic secretion of saliva for further digestion is indicated when, in estimating the function of taste for the pleasures of appetite, we realize that materials can be tasted only when dissolved in the mouth and thereby brought into relation with the taste organs. The saliva which “waters” the mouth assures the dissolving of dry but soluble food even when it is taken in large amount.
The importance of the initial psychic secretion of gastric juice is made clear by the fact that continuance[p. 7] of the flow of this juice during digestion is provided by the action of its acid or its digestive products on the mucous membrane of the pyloric end of the stomach, and that secretion of the pancreatic juice and bile are called forth by the action of this same acid on the mucous membrane of the duodenum. The proper starting of the digestive process, therefore, is conditioned by the satisfactions of the palate, and the consequent flow of the first digestive fluids.
The facts brought out experimentally in studies on lower animals are doubtless true also of man. Not very infrequently, because of the accidental swallowing of corrosive substances, the esophagus is so injured that, when it heals, the sides grow together and the tube is closed. Under these circumstances an opening has to be made into the stomach through the side of the body and then the individual chews his food in the usual manner, but ejects it from his mouth into a tube which is passed through the gastric opening. The food thus goes from mouth to stomach through a tube outside the chest instead of inside the chest. As long ago as 1878, Richet,[3] who had occasion to study a girl whose esophagus was closed and who was fed through a gastric fistula, reported that whenever the girl chewed or tasted a highly sapid substance, such as sugar or lemon juice, while the stomach was empty, there flowed[p. 8] from the fistula a considerable quantity of gastric juice. A number of later observers[4] have had similar cases in human beings, especially in children, and have reported in detail results which correspond remarkably with those obtained in the laboratory. Hornborg[4] found that when the little boy whom he studied chewed agreeable food a more or less active secretion of gastric juice invariably started, whereas the chewing of an indifferent substance, as gutta-percha, was followed by no secretion. All these observations clearly demonstrate that the normal flow of the first digestive fluids, the saliva and the gastric juice, is favored by the pleasurable feelings which accompany the taste and smell of food during mastication, or which are roused in anticipation of eating when choice morsels are seen or smelled.
These facts are of fundamental importance in the serving of food, especially when, through illness, the appetite is fickle. The degree of daintiness with which nourishment is served, the little attentions to esthetic details—the arrangement of the dishes, the small portions of food, the flower beside the plate—all may help to render food pleasing to the eye and savory to the nostrils and may be the deciding factors in determining whether the restoration of strength is to begin or not.
[p. 9]
The conditions favorable to proper digestion are wholly abolished when unpleasant feelings such as vexation and worry and anxiety, or great emotions such as anger and fear, are allowed to prevail. This fact, so far as the salivary secretion is concerned, has long been known. The dry mouth of the anxious person called upon to speak in public is a common instance; and the “ordeal of rice,” as employed in India, was a practical utilization of the knowledge that excitement is capable of inhibiting the salivary flow. When several persons were suspected of crime, the consecrated rice was given to them all to chew, and after a short time it was spit out upon the leaf of the sacred fig tree. If anyone ejected it dry, that was taken as proof that fear of being discovered had stopped the secretion, and consequently he was adjudged guilty.[5]
What has long been recognized as true of the secretion of saliva has been proved true also of the secretion of gastric juice. For example, Hornborg was unable to confirm in his little patient with a gastric fistula the observation by Pawlow that when hunger is present the mere seeing of food results in a flow of gastric juice. Hornborg explained the difference between his and Pawlow’s results by the different ways in[p. 10] which the boy and the dogs faced the situation. When food was shown, but withheld, the hungry dogs were all eagerness to secure it, and the juice very soon began to flow. The boy, on the contrary, became vexed when he could not eat at once, and began to cry; then no secretion appeared. Bogen also has reported the instance of a child with closed esophagus and gastric fistula, who sometimes fell into such a passion in consequence of vain hoping for food that the giving of the food, after the child was calmed, was not followed by any flow of the secretion.
The inhibitory influence of excitement has also been seen in lower animals under laboratory conditions. Le Conte[6] declares that in studying gastric secretion it is necessary to avoid all circumstances likely to provoke emotional reactions. In the fear which dogs manifest when first brought into strange surroundings he found that activity of the gastric glands may be completely suppressed. The suppression occurred even if the dog had eaten freely and was then disturbed—as, for example, by being tied to a table. When the animals became accustomed to the experimental procedure, it no longer had an inhibitory effect. The studies of Bickel and Sasaki[7] confirm and define more precisely this inhibitory effect of strong emotion on gastric secretion. They observed the inhibition on a dog with an[p. 11] esophageal fistula, and with a side pouch of the stomach, which, as in Pawlow’s experiments, opened only to the exterior. In this dog Bickel and Sasaki noted, as Pawlow had, that sham feeding was attended by a copious flow of gastric juice, a true psychic secretion, resulting from the pleasurable taste of the food. In a typical instance the sham feeding lasted five minutes, and the secretion continued for twenty minutes, during which time 66.7 cubic centimeters of pure gastric juice were produced.
On another day a cat was brought into the presence of the dog, whereupon the dog flew into a great fury. The cat was soon removed, and the dog pacified. Now the dog was again given the sham feeding for five minutes. In spite of the fact that the animal was hungry and ate eagerly, there was no secretion worthy of mention. During a period of twenty minutes, corresponding to the previous observation, only 9 cubic centimeters of acid fluid were produced, and this was rich in mucus. It is evident that in the dog, as in the boy observed by Bogen, strong emotions can so profoundly disarrange the mechanisms of secretion that the pleasurable excitation which accompanies the taking of food cannot cause the normal flow.
On another occasion Bickel and Sasaki started gastric secretion in the dog by sham feeding, and[p. 12] when the flow of gastric juice had reached a certain height, the dog was infuriated for five minutes by the presence of the cat. During the next fifteen minutes there appeared only a few drops of a very mucous secretion. Evidently in this instance a physiological process, started as an accompaniment of a psychic state quietly pleasurable in character, was almost entirely stopped after another psychic state violent in character.
It is noteworthy that in both the favorable and unfavorable results of the emotional excitement illustrated in Bickel and Sasaki’s dog the effects persisted long after the removal of the exciting condition. This fact, in its favorable aspect, Bickel[8] was able to confirm in a girl with esophageal and gastric fistulas; the gastric secretion long outlasted the period of eating, although no food entered the stomach. The influences unfavorable to digestion, however, are stronger than those which promote it. And evidently, if the digestive process, because of emotional disturbance, is for some time inhibited, the swallowing of food which must lie stagnant in the stomach is a most irrational procedure. If a child has experienced an outburst of passion, it is well not to urge the taking of nourishment soon afterwards. Macbeth’s advice that “good digestion wait on appetite and health on both,” is now well-founded physiology.
[p. 13]
Other digestive glands than the salivary and the gastric may be checked in emotional excitement. Recently Oechsler[9] has reported that in such psychic disturbances as were shown by Bickel and Sasaki to be accompanied by suppressed secretion of the gastric juice, the secretion of pancreatic juice may be stopped, and the flow of bile definitely checked. All the means of bringing about chemical changes in the food may be thus temporarily abolished.
The secretions of the digestive glands and the chemical changes wrought by them are of little worth unless the food is carried onward through the alimentary canal into fresh regions of digestion and is thoroughly exposed to the intestinal wall for absorption. In studying these mechanical aspects of digestion I was led to infer[10] that just as there is a psychic secretion, so likewise there is probably a “psychic tone” or “psychic contraction” of the gastro-intestinal muscles as a result of taking food. For if the vagus nerve supply to the stomach is cut immediately before an animal takes food, the usual contractions of the gastric wall, as seen by the Röntgen rays, do not occur; but if these nerves are cut after food has been eaten with relish, the contractions which[p. 14] have started continue without cessation. The nerves in both conditions were severed under anesthesia, so that no element of pain entered into the experiments. In the absence of hunger, which in itself provides a contracted stomach,[11] the pleasurable taking of food may, therefore, be a primary condition for the appearance of natural contractions of the gastro-intestinal canal.
Again just as the secretory activities of the stomach are unfavorably influenced by strong emotions, so also are the movements of the stomach; and, indeed, the movements of almost the entire alimentary canal are wholly stopped during great excitement. In my earliest observations on the movements of the stomach[12] I had difficulty because in some animals the waves of contraction were perfectly evident, while in others there was no sign of activity. Several weeks passed before I discovered that this difference was associated with a difference of sex. In order to be observed with Röntgen rays the animals were restrained in a holder. Although the holder was comfortable, the male cats, particularly the young males, were restive and excited on being fastened to it, and under these circumstances gastric peristaltic waves were absent; the female cats, especially if elderly, usually submitted with calmness to the restraint, and in them the waves had their normal occurrence. Once a female with[p. 15] kittens turned from her state of quiet contentment to one of apparent restless anxiety. The movements of the stomach immediately stopped, the gastric wall became wholly relaxed, and only after the animal had been petted and began to purr did the moving waves start again on their course. By covering the cat’s mouth and nose with the fingers until a slight distress of breathing is produced, the stomach contractions can be stopped at will. In the cat, therefore, any sign of rage or fear, such as was seen in dogs by Le Conte and by Bickel and Sasaki, was accompanied by a total abolition of the movements of the stomach. Even indications of slight anxiety may be attended by complete absence of the churning waves. In a vigorous young male cat I have watched the stomach for more than an hour by means of the Röntgen rays, and during that time not the slightest beginning of peristaltic activity appeared; yet the only visible indication of excitement in the animal was a continued quick twitching of the tail to and fro. What is true of the cat I have found true also of the rabbit, dog and guinea-pig[13]—very mild emotional disturbances are attended by abolition of peristalsis. The observations on the rabbit have been confirmed by Auer,[14] who found that the handling of the animal incidental to fastening it gently to a holder stopped gastric peristalsis for a[p. 16] variable length of time. And if the animal was startled for any reason, or struggled excitedly, peristalsis was again abolished. The observations on the dog also have been confirmed; Lommel[15] found that small dogs in strange surroundings might have no contractions of the stomach for two or three hours. And whenever the animals showed any indications of being uncomfortable or distressed, the contractions were inhibited and the discharge of contents from the stomach checked.
Like the peristaltic waves in the stomach, the peristalsis and the kneading movements (segmentation) in the small intestine, and the reversed peristalsis in the large intestine all cease whenever the observed animal shows signs of emotional excitement.
There is no doubt that just as the secretory activity of the stomach is affected in a similar fashion in man and in lower animals, so likewise gastric and intestinal peristaltic waves are stopped in man as they are stopped in lower animals, by worry and anxiety and the stronger affective states. The conditions of mental discord may thus give rise to a sense of gastric inertia. For example, a patient described by Müller[16] testified that anxiety was always accompanied by a feeling of weight, as if the food remained in the stomach. Every addition of food caused an[p. 17] increase of the trouble. Strong emotional states in this instance led almost always to gastric distress, which persisted, according to the grade and the duration of the psychic disturbance, between a half-hour and several days. The patient was not hysterical or neurasthenic, but was a very sensitive woman deeply affected by moods.
The feeling of heaviness in the stomach, mentioned in the foregoing case, is not uncommonly complained of by nervous persons, and may be due to stagnation of the contents. That such stagnation occurs is shown by the following instance. A refined and sensitive woman, who had had digestive difficulties, came with her husband to Boston to be examined. They went to a hotel for the night. The next morning the woman appeared at the consultant’s office an hour after having eaten a test meal. An examination of the gastric contents revealed no free acid, no digestion of the test breakfast, and the presence of a considerable amount of the supper of the previous evening. The explanation of this stagnation of the food in the stomach came from the family doctor, who reported that the husband had made the visit to the city an occasion for becoming uncontrollably drunk, and that he had by his escapades given his wife a night of turbulent anxiety. The second morning, after the woman had had a good rest, the gastric contents[p. 18] were again examined; the proper acidity was found, and the test breakfast had been normally digested and discharged.
These cases are merely illustrative and doubtless can be many times duplicated in the experience of any physician concerned largely with digestive disorders. Indeed, the opinion has been expressed that a great majority of the cases of gastric indigestion that come for treatment are functional in character and of nervous origin. It is the emotional element that seems most characteristic of these cases. To so great an extent is this true that Rosenbach has suggested that as a term to characterize the cause of the disturbances, “emotional” dyspepsia is better than “nervous” dyspepsia.[17]
The advocates of the theory of organic evolution early pointed out the similarity between the bodily disturbances in pain and in the major emotions. The alterations of function of internal organs they could not know about. The general statement, however, that pain evokes the same changes that are evoked by emotion, is true also of these deep-lying structures. Wertheimer[18] proved many years since that stimulation of a sensory nerve in an anesthetized animal—such stimulation as in a conscious animal would induce[p. 19] pain—quickly abolished the contractions of the stomach. And Netschaiev, working in Pawlow’s[19] laboratory, showed that excitation of the sensory fibres in the sciatic nerve for two or three minutes resulted in an inhibition of the secretion of gastric juice that lasted for several hours. Similar effects from painful experience have been not uncommonly noted in human beings. Mantegazza,[20] in his account of the physiology of pain, has cited a number of such examples, and from them he has concluded that pain interferes with digestion by lessening appetite and by producing various forms of dyspepsia, with arrest of gastric digestion, and with vomiting and diarrhea. The expression, “sickening pain” is testimony to the power of strong sensory stimulation to upset the digestive processes profoundly. Vomiting is as likely to follow violent pain as it is to follow strong emotion. A “sick headache” may be, indeed, a sequence of events in which the pain from the headache is primary, and the nausea and other evidences of digestive disorder are secondary.
As the foregoing account has shown, emotional conditions or “feelings” may be accompanied by quite opposite effects in the alimentary canal, some highly favorable to good digestion, some highly disturbing. It is an interesting fact that the feelings having these antagonistic actions are[p. 20] typically expressed through nerve supplies which are correspondingly opposed in their influence on the digestive organs. The antagonism between these nerve supplies is of fundamental importance in understanding not only the operation of conditions favorable or unfavorable to digestion but also in obtaining insight into the conflicts of emotional states. Since a consideration of the arrangement and mode of action of these nerves will establish a firm basis for later analysis and conclusions, they will next be considered.
1 Pawlow: The Work of the Digestive Glands, London, 1902.
2 Bidder and Schmidt: Die Verdauungssäfte und der Stoffwechsel, Leipzig, 1852, p. 35.
3 Richet: Journal de l’Anatomie et de la Physiologie, 1878, xiv, p. 170.
4 See Hornborg: Skandinavisches Archiv für Physiologie, 1904, xv, p. 248. Cade and Latarjet: Journal de Physiologie et Pathologie Générale, 1905, vii, p. 221. Bogen: Archiv für die gesammte Physiologie, 1907, cxvii, p. 156. Lavenson: Archives of Internal Medicine, 1909, iv, p. 271.
5 Lea: Superstition and Force, Philadelphia, 1892, p. 344.
6 Le Conte: La Cellule, 1900, xvii, p. 291.
7 Bickel and Sasaki: Deutsche medizinische Wochenschrift, 1905, xxxi, p. 1829.
8 Bickel: Berliner klinische Wochenschrift, 1906, xliii, p. 845.
[9] Oechsler: Internationelle Beiträge zur Pathologie und Therapie der Ernährungstörungen, 1914, v, p. 1.
10 Cannon: The Mechanical Factors of Digestion, London and New York, 1911, p. 200.
[p. 21]
11 Cannon and Washburn: American Journal of Physiology, 1912, xxix, p. 441.
12 Cannon: The American Journal of Physiology, 1898, i, p. 38.
13 Cannon: American Journal of Physiology, 1902, vii, p. xxii.
14 Auer: American Journal of Physiology, 1907, xviii, p. 356.
15 Lommel: Münchener medizinische Wochenschrift, 1903, i, p. 1634.
16 Müller: Deutsches Archiv für klinische Medicin, 1907, lxxxix, p. 434.
17 Rosenbach: Berliner klinische Wochenschrift, 1897, xxxiv, p. 71.
18 Wertheimer: Archives de Physiologie, 1892, xxiv, p. 379.
19 Pawlow: Loc. cit., p. 56.
20 Mantegazza: Fisiologia del Dolore, Florence, 1880, p. 123.
[p. 22]
THE GENERAL ORGANIZATION OF THE VISCERAL NERVES CONCERNED IN EMOTIONS
The structures of the alimentary canal which are brought into activity during the satisfactions of appetite or are checked in their activity during pain and emotional excitement are either the secreting digestive glands or the smooth muscle which surrounds the canal. Both the gland cells and the smooth-muscle cells differ from other cells which are subject to nervous influence—those of striated, or skeletal, muscle—in not being directly under voluntary control and in being slower in their response. The muscle connected with the skeleton responds to stimulation within two or three thousandths of a second; the delay with gland cells and with smooth muscle is more likely to be measured in seconds than in fractions of a second.
The skeletal muscles receive their nerve supply direct from the central nervous system, i. e., the[p. 23] nerve fibres distributed to these muscles are parts of neurones whose cell bodies lie within the brain or spinal cord. The glands and smooth muscles of the viscera, on the contrary, are, so far as is now known, never innervated directly from the central nervous system.[*] The neurones reaching out from the brain or spinal cord never come into immediate relation with the gland or smooth-muscle cells; there are always interposed between the cerebrospinal neurones and the viscera extra neurones whose bodies and processes lie wholly outside the central nervous system. They are represented in dotted lines in Fig. 1. I have suggested that possibly these outlying neurones act as “transformers,” modifying the impulses received from the central source (impulses suited to call forth the quick responses of skeletal muscle), and adapting these impulses to the peculiar, more slowly-acting tissues, the secreting cells and visceral muscle, to which they are distributed.[1]
*The special case of the adrenal glands will be considered later.
The outlying neurones typically have their cell bodies grouped in ganglia (G’s, Fig. 1) which, in the trunk region, lie along either side of the spinal cord and in the head region and in the pelvic part of the abdominal cavity are disposed near the organs which the neurones supply. In some instances these neurones lie wholly within the[p. 25] structure which they innervate (see e. g., the heart and the stomach, Fig. 1). In other instances the fibres passing out from the ganglia—the so-called “postganglionic fibres”—may traverse long distances before reaching their destination. The innervation of blood vessels in the foot by neurones whose cell bodies are in the lower trunk region is an example of this extensive distribution of the fibres.
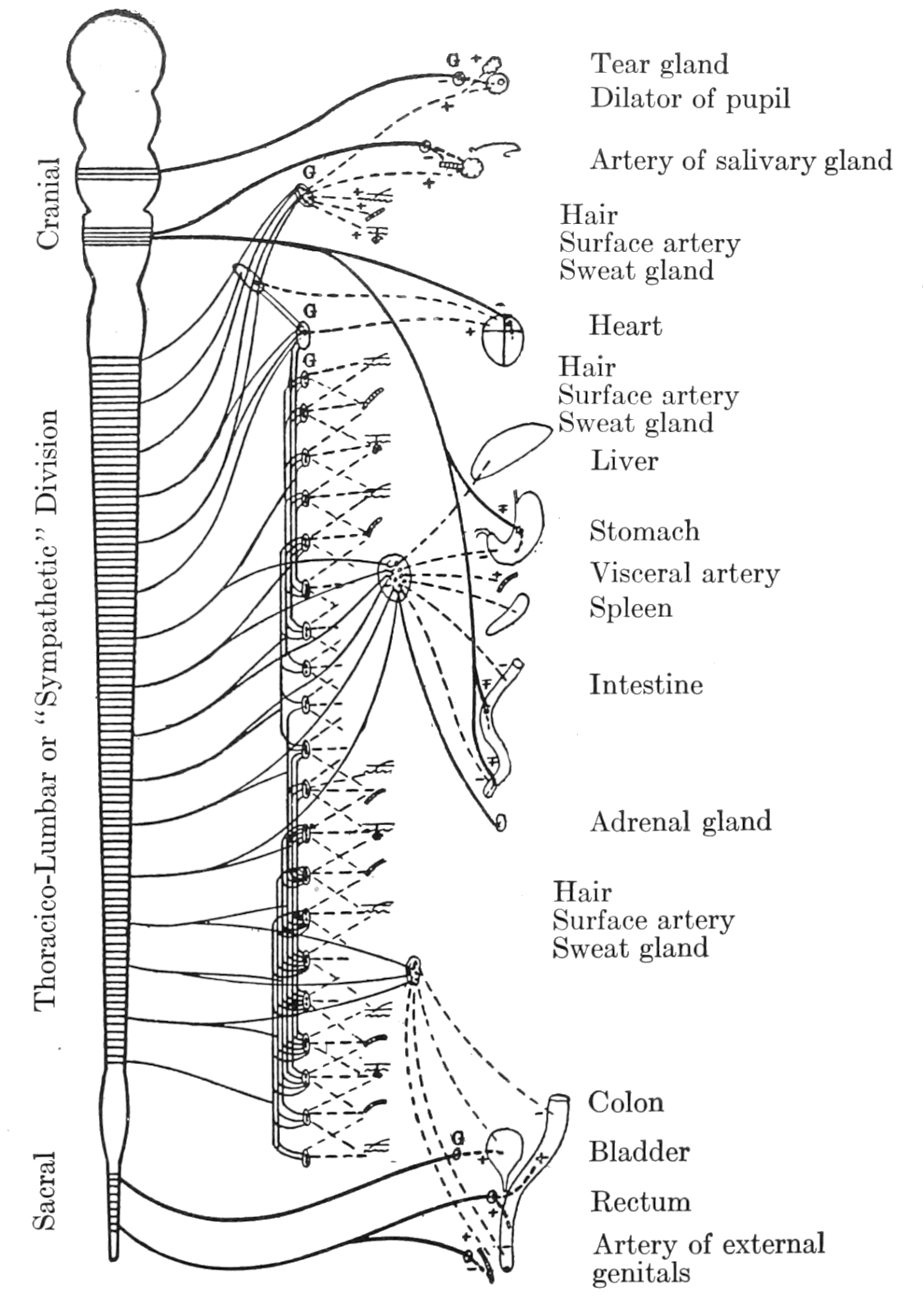
As suggested above, the outlying neurones are connected with the brain and spinal cord by neurones whose cell bodies lie within the central nervous organs. These connecting neurones, represented in continuous lines in Fig. 1, do not pass out in a continuous series all along the cerebrospinal axis. Where the nerves pass out from the spinal cord to the fore and hind limbs, fibres are not given off to the ganglia. Thus these connecting or “preganglionic” fibres are separated into three divisions. In front of the nerve roots for the fore limbs is the head or cranial division, between the nerve roots for the fore limbs and those for the hind limbs is the trunk division (or thoracico-lumbar division, or, in the older terminology, the “sympathetic system”); and after the nerve roots for the hind limbs the sacral division.
This system of outlying neurones, with postganglionic[p. 26] fibres innervating the viscera, and with preganglionic fibres reaching out to them from the cerebrospinal system, has been called by Langley, to whom we are indebted for most of our knowledge of its organization, the autonomic nervous system.[2] This term indicates that the structures which the system supplies are not subject to voluntary control, but operate to a large degree independently. As we have seen, a highly potent mode of influencing these structures is through conditions of pain and emotional excitement. The parts of the autonomic system—the cranial, the sympathetic, and the sacral—have a number of peculiarities which are of prime importance in accounting for the bodily manifestations of such affective states.
The fibres of the sympathetic division differ from those of the other two divisions in being distributed through the body very widely. They go to the eyes, causing dilation of the pupils. They go to the heart and, when stimulated, they cause it to beat rapidly. They carry impulses to arteries and arterioles of the skin, the abdominal viscera, and other parts, keeping the smooth muscles of the vessel walls in a state of slight contraction[p. 27] or tone, and thus serving to maintain an arterial pressure sufficiently high to meet sudden demands in any special region; or, in times of special discharge of impulses, to increase the tone and thus also the arterial pressure. They are distributed extensively to the smooth muscle attached to the hairs; and when they cause this muscle to contract, the hairs are erected. They go to sweat glands, causing the outpouring of sweat. These fibres pass also to the entire length of the gastro-intestinal canal. And the inhibition of digestive activity which, as we have learned, occurs in pain and emotional states, is due to impulses which are conducted outward by the splanchnic nerves—the preganglionic fibres that reach to the great ganglia in the upper abdomen (see Fig. 1)—and thence are spread by postganglionic fibres all along the gut.[3] They innervate likewise the genito-urinary tracts, causing contraction of the smooth muscle of the internal genital organs, and usually relaxation of the bladder. Finally they affect the liver, releasing the storage of material there in a manner which may be of great service to the body in time of need. The extensiveness of the distribution of the fibres of the sympathetic division is one of its most prominent characteristics.
Another typical feature of the sympathetic division is an arrangement of neurones for diffuse[p. 28] discharge of the nerve impulses. As shown diagrammatically in Fig. 1, the preganglionic fibres from the central nervous system may extend through several of the sympathetic ganglia and give off in each of them connections to cell bodies of the outlying neurones. Although the neurones which transmit sensory impulses from the skin into spinal cord have similar relations to nerve cells lying at different levels of the cord, the operation in the two cases is quite different. In the spinal cord the sensory impulse produces directed and closely limited effects, as, for example, when reflexes are being evoked in a “spinal” animal (i. e., an animal with the spinal cord isolated from the rest of the central nervous system), the left hind limb is nicely lifted, in response to a harmful stimulus applied to the left foot, without widespread marked involvement of the rest of the body in the response.[4] In the action of the sympathetic division, on the contrary, the connection of single preganglionic fibres with numerous outlying neurones seems to be not at all arranged for specific effects in this or that particular region. There are, to be sure, in different circumstances variations in the degree of activity of different parts; for example, it is probable that dilation of the pupil in the cat occurs more readily than erection of the hairs. It may be in this instance, however, that specially direct[p. 29] pathways to the eye are present for common use in non-emotional states (in dim light, e. g.), and that only slight general disturbance in the central nervous system, therefore, would be necessary to send impulses by these well-worn courses. Thus for local reasons (dust, e. g.) tears might flow from excitation of the tear glands by sympathetic impulses, although other parts innervated by this same division might be but little disturbed. We have no means of voluntarily wearing these pathways, however, and both from anatomical and physiological evidence the neurone relations in the sympathetic division of the autonomic system seem devised for widespread diffusion of nervous impulses.
The cranial and sacral autonomic divisions differ from the sympathetic in having only restricted distribution (see Fig. 1). The third cranial nerves deliver impulses from the brain to ganglia in which lie the cell bodies of neurones innervating smooth muscle only in the front of the eyes. The vagus nerves are distributed to the lungs, heart, stomach, and small intestine. As shown diagrammatically in Fig. 1, the outlying neurones in the last three of these organs lie within the organs themselves. By this arrangement, although the preganglionic fibres of[p. 30] the vagi are extended in various directions to structures of quite diverse functions, singleness and separateness of connection of the peripheral organs with the central nervous system is assured. The same specific relation between efferent fibres and the viscera is seen in the sacral autonomic. In this division the preganglionic fibres pass out from the spinal cord to ganglia lying in close proximity to the distal colon, the bladder, and the external genitals. And the postganglionic fibres deliver the nerve impulses only to the nearby organs. Besides these innervations the cranial and sacral divisions supply individual arteries with “dilator nerves”—nerves causing relaxation of the particular vessels. Quite typically, therefore, the efferent fibres of the two terminal divisions of the autonomic differ from those of the mid-division in having few of the distributed connections characteristic of the mid-division, and in innervating distinctively the organs to which they are distributed. The cranial and sacral preganglionic fibres resemble thus the nerves to skeletal muscles, and their arrangement provides similar possibilities of specific and separate action in any part, without action in other parts.
The cranial autonomic, represented by the vagus nerves, is the part of the visceral nervous[p. 31] system concerned in the psychic secretion of the gastric juice. Pawlow showed that when these nerves are severed psychic secretion is abolished. The cranial nerves to the salivary glands are similarly the agents for psychic secretion in these organs, and are known to cause also dilation of the arteries supplying the glands, so that during activity the glands receive a more abundant flow of blood. As previously stated (see p. 13), the evidence for a psychic tonus of the gastro-intestinal musculature rests on a failure of the normal contractions if the vagi are severed before food is taken, in contrast to the continuance of the contractions if the nerves are severed just afterwards. The vagi artificially excited are well known as stimulators of increased tone in the smooth muscle of the alimentary canal. Aside from these positive effects on the muscles of the digestive tract and its accessory glands, cranial autonomic fibres cause contraction of the pupil of the eye, and slowing of the heart rate.
A glance at these various functions of the cranial division reveals at once that they serve for bodily conservation. By narrowing the pupil of the eye they shield the retina from excessive light. By slowing the heart rate, they give the cardiac muscle longer periods for rest and invigoration. And by providing for the flow of saliva and gastric juice and by supplying the muscular[p. 32] tone necessary for contraction of the alimentary canal, they prove fundamentally essential to the processes of proper digestion and absorption by which energy-yielding material is taken into the body and stored. To the cranial division of the visceral nerves, therefore, belongs the quiet service of building up reserves and fortifying the body against times of need or stress.
Sacral autonomic fibres cause contraction of the rectum and distal colon and also contraction of the bladder. In both instances the effects result reflexly from stretching of the tonically contracted viscera by their accumulating contents. No affective states precede this normal action of the sacral division and even those which accompany or follow are only mildly positive; a feeling of relief rather than of elation usually attends the completion of the act of defecation or micturition—though there is testimony to the contrary.
The sacral autonomic fibres also include, however, the nervi erigentes which bring about engorgement of erectile tissue in the external genitals. According to Langley and Anderson[5] the sacral nerves have no effect on the internal generative organs. The vasa deferentia and the seminal vesicles whose rhythmic contractions[p. 33] mark the acme of sexual excitement in the male, and the uterus whose contractions in the female are probably analogous, are supplied only by lumbar branches—part of the sympathetic division. These branches also act in opposition to the nervi erigentes and cause constriction of the blood vessels of the external genitals. The sexual orgasm involves a high degree of emotional excitement; but it can be rightly considered as essentially a reflex mechanism; and, again in this instance, distention of tubules, vesicles, and blood vessels can be found at the beginning of the incident, and relief from this distention at the end.
Although distention is the commonest occasion for bringing the sacral division into activity it is not the only occasion. Great emotion, such as is accompanied by nervous discharges via the sympathetic division, may also be accompanied by discharges via the sacral fibres. The involuntary voiding of the bladder and lower gut at times of violent mental stress is well known. Veterans of wars testify that just before the beginning of a battle many of the men have to retire temporarily from the firing line. And the power of sights and smells and libidinous thoughts to disturb the regions controlled by the nervi erigentes proves that this part of the autonomic system also has its peculiar affective states. The fact that one part of the sacral division, e. g., the distribution[p. 34] to the bladder, may be in abeyance, while another part, e. g., the distribution to the rectum, is active, illustrates again the directive discharge of impulses which has been previously described as characteristic of the cranial and sacral portions of the autonomic system.
Like the cranial division, the sacral is engaged in internal service to the body, in the performance of acts leading immediately to greater comfort.
As indicated in the foregoing description many of the viscera are innervated both by the cranial or sacral part of the autonomic and by the sympathetic. When the mid-part meets either end-part in any viscus their effects are antagonistic. Thus the cranial supply to the eye contracts the pupil, the sympathetic dilates it; the cranial slows the heart, the sympathetic accelerates it; the sacral contracts the lower part of the large intestine, the sympathetic relaxes it; the sacral relaxes the exit from the bladder, the sympathetic contracts it. These opposed effects are indicated in Fig. 1 by + for contraction, acceleration or increased tone; and by - for inhibition, relaxation, or decreased tone.[*]
* The vagus nerve, when artificially stimulated, has a primary, brief inhibitory effect on the stomach and small intestine; its main function, however, as already stated, is to produce increased tone and contraction in these organs. This double action of the vagus is marked thus, ∓, in Fig. 1.
[p. 35]
Sherrington has demonstrated that the setting of skeletal muscles in opposed groups about a joint or system of joints—as in flexors and extensors—is associated with an internal organization of the central nervous system that provides for relaxation of one group of the opposed muscles when the other group is made to contract. This “reciprocal innervation of antagonistic muscles,” as Sherrington has called it,[6] is thus a device for orderly action in the body. As the above description has shown, there are peripheral oppositions in the viscera corresponding to the oppositions between flexor and extensor muscles. In all probability these opposed innervations of the viscera have counterparts in the organization of neurones in the central nervous system. Sherrington has noticed, and I can confirm the observation, that even though the sympathetic supply to the eye is severed and is therefore incapable of causing dilation of the pupil, nevertheless the pupil dilates in a paroxysm of anger—due, no doubt (because the response is too rapid to be mediated by the blood stream), to central inhibition of the cranial nerve supply to the constrictor muscles—i. e., an inhibition of the muscles which naturally oppose the dilator action of the sympathetic. Pain, the major emotions—fear and[p. 36] rage—and also intense excitement, are manifested in the activities of the sympathetic division. When in these states impulses rush out over the neurones of this division they produce all the changes typical of sympathetic excitation, such as dilating the pupils, inhibiting digestion, causing pallor, accelerating the heart, and various other well-known effects. The impulses of the sympathetic neurones, as indicated by their dominance over the digestive process, are capable of readily overwhelming the conditions established by neurones of the cranial division of the autonomic system.
Lying anterior to each kidney is a small body—the adrenal gland. It is composed of an external portion or cortex, and a central portion or medulla. From the medulla can be extracted a substance, called variously suprarenin, adrenin, epinephrin or “adrenalin,”[*] which, in extraordinarily minute amounts, affects the structures innervated by the sympathetic division of the autonomic system[p. 37] precisely as if they were receiving nervous impulses. For example, when adrenin is injected into the blood, it will cause pupils to dilate, hairs to stand erect, blood vessels to be constricted, the activities of the alimentary canal to be inhibited, and sugar to be liberated from the liver. These effects are not produced by action of the substance on the central nervous system, but by direct action on the organ itself.[7] And the effects occur even after the structures have been removed from the body and kept alive artificially.
* The name “adrenalin” is proprietary. “Epinephrin” and “adrenin” have been suggested as terms free from commercial suggestions. As adrenin is shorter and more clearly related to the common adjectival form, adrenal, I have followed Schäfer in using adrenin to designate the substance produced physiologically by the adrenal glands.
The adrenals are glands of internal secretion, i. e., like the thyroid, parathyroid, and pituitary glands, for example; they have no connection with the surface of the body, and they give out into the blood the material which they elaborate. The blood is carried away from each of them by the lumbo-adrenal vein which empties either into the renal vein or directly into the inferior vena cava just anterior to the openings of the renal veins. The adrenal glands are supplied by preganglionic fibres of the autonomic group,[8] shown in solid line in Fig. 1. This seems an exception to the general rule that gland cells have an outlying neurone between them and the neurones of the central nervous system. The medulla of the adrenal gland, however, is composed of modified nerve cells, and may therefore be regarded as offering exceptional conditions.
[p. 38]
The foregoing brief sketch of the organization of the autonomic system brings out a number of points that should be of importance as bearing on the nature of the emotions which manifest themselves in the operations of this system. Thus it is highly probable that the sympathetic division, because arranged for diffuse discharge, is likely to be brought into activity as a whole, whereas the sacral and cranial divisions, arranged for particular action on separate organs, may operate in parts. Also, because antagonisms exist between the middle and either end division of the autonomic, affective states may be classified according to their expression in the middle or an end division and these states would be, like the nerves, antagonistic in character. And finally, since the adrenal glands are innervated by autonomic fibres of the mid-division, and since adrenal secretion stimulates the same activities that are stimulated nervously by this division, it is possible that disturbances in the realm of the sympathetic, although initiated by nervous discharge, are automatically augmented and prolonged through chemical effects of the adrenal secretion.
1 Cannon: The American Journal of Psychology, 1914, xxv, p. 257.
[p. 39]
2 For a summary of his studies of the organization of the autonomic system, see Langley: Ergebnisse der Physiologie, Wiesbaden, 1903, ii2, p. 818.
3 See Cannon: American Journal of Physiology, 1905, xiii, p. xxii.
4 See Sherrington: The Integrative Action of the Nervous System, New York, 1909, p. 19.
5 Langley and Anderson: Journal of Physiology, 1895, xix, see pp. 85, 122.
6 Sherrington: Loc. cit., p. 90.
7 Elliott: Journal of Physiology, 1905, xxxii, p. 426.
8 See Elliott: Journal of Physiology, 1913, xlvi, p. 289 ff.
[p. 40]
METHODS OF DEMONSTRATING ADRENAL SECRETION AND ITS NERVOUS CONTROL
As stated in the first chapter, the inhibition of gastric secretion produced by great excitement long outlasts the presence of the object which evokes the excitement. The dog that was enraged by seeing a cat for five minutes secreted only a few drops of gastric juice during the next fifteen minutes. Why did the state of excitation persist so long after the period of stimulation had ended? This question, which presented itself to me while reading Bickel and Sasaki’s paper, furnished the suggestion expressed at the close of the last chapter, that the excitement might provoke a flow of adrenal secretion, and that the changes originally induced in the digestive organs by nervous impulses might be continued by circulating adrenin. The prolongation of the effect might be thus explained. Whether that idea is correct or not has not been tested. Its chief service was in leading to an enquiry as to whether[p. 41] the adrenal glands are in fact stimulated to action in emotional excitement. The preganglionic fibres passing to the glands are contained in the splanchnic nerves. What is the effect of splanchnic stimulation?
It was in 1891 that Jacobi[1] described nerve fibres derived from the splanchnic trunks which were distributed to the adrenal glands. Six years later Biedl[2] found that these nerves conveyed vasodilator impulses to the glands, and he suggested that they probably conveyed also secretory impulses. Evidence in support of this suggestion was presented the following year by Dreyer,[3] who demonstrated that electrical excitation of the splanchnic nerves produced in the blood taken from the adrenal veins an increased amount of a substance having the power of raising arterial blood pressure, and that this result was independent of accompanying changes in the blood supply to the glands. The conclusion drawn by Dreyer that this substance was adrenin has been confirmed in various ways by later observers. Tscheboksaroff[4] repeated Dreyer’s procedure and found in blood taken from the veins after splanchnic stimulation evidences of the presence of adrenin that were previously absent. Asher[5][p. 42] observed a rise of blood pressure when the glands were stimulated in such a manner as not to cause constriction of the arteries—the rise was therefore assumed to be due to secreted adrenin. Dilation of the pupil was used by Meltzer and Joseph[6] to prove secretory action of the splanchnics on the adrenal glands; they found that stimulation of the distal portion of the cut splanchnic nerve caused the pupil to enlarge—an effect characteristic of adrenin circulating in the blood. Elliott[7] repeated this procedure, but made it a more rigorous proof of internal secretion of the adrenals by noting that the effect failed to appear if the gland on the stimulated side was removed. Additional proof was brought by myself and Lyman[8] when we found that the typical drop in arterial pressure produced in cats by injecting small amounts of adrenin could be exactly reproduced by stimulating the splanchnic nerves after the abdominal blood vessels, which contract when these nerves are excited, were tied so that no changes in them could occur to influence the rest of the circulation.
The problem of splanchnic influence on the adrenal glands Elliott attacked by a still different method. Using, as a measure, the graded effects of graded amounts of adrenin on blood pressure, he was able to assay the quantity of adrenin in adrenal glands after various conditions had been[p. 43] allowed to prevail. The tests were made on cats. In these animals each adrenal gland is supplied only by the splanchnic fibres of its own side, and the two glands normally contain almost exactly the same amount of adrenin. Elliott[9] found that when the gland on one side was isolated by cutting its splanchnic supply, and then impulses were sent along the intact nerves of the other side, either by disturbing the animal or by artificial excitation of the nerves, the gland to which these fibres reached invariably contained less adrenin, often very much less, than the isolated gland. Results obtained by the method employed by Elliott have been confirmed with remarkable exactness in results obtained by Folin, Denis and myself,[10] using a highly sensitive color test after adding the gland extract to a solution of phosphotungstic acid.
All these observations, with a variety of methods, and by a respectable number of reliable investigators, are harmonious in bringing proof that artificial stimulation of the nerves leading to the adrenal glands will induce secretory activity in the adrenal medulla, and that in consequence adrenin will be increased in the blood. The fact is therefore securely established that in the body a mechanism exists by which these glands can be made to discharge this peculiar substance promptly into the circulation.
[p. 44]
As we have already seen, the phenomena of a great emotional disturbance in an animal indicate that sympathetic impulses dominate the viscera. When, for example, a cat becomes frightened, the pupils dilate, the activities of the stomach and intestines are inhibited, the heart beats rapidly, the hairs of the back and tail stand erect—from one end of the animal to the other there are abundant signs of nervous discharges along sympathetic courses. Do not the adrenal glands share in this widespread subjugation of the viscera to sympathetic control?
This question, whether the common excitements of an animal’s life might be capable of evoking a discharge of adrenin, was taken up by D. de la Paz and myself in 1910. We made use of the natural enmity between two laboratory animals, the dog and the cat, to pursue our experiments. In these experiments the cat, fastened in a comfortable holder (the holder already mentioned as being used in X-ray studies of the movements of the alimentary canal), was placed near a barking dog. Some cats when thus treated showed almost no signs of fear; others, with scarcely a movement of defense, presented the typical picture. In favorable cases the excitement was allowed to prevail for five or ten minutes, and in[p. 45] a few cases longer. Samples of blood were taken within a few minutes before and after the period.
The blood was obtained from the inferior vena cava anterior to the opening of the adrenal veins, i. e., at a point inside the body near the level of the notch at the lower end of the sternum. To get the blood so far from the surface without disturbing the animal was at first a difficult problem. We found, however, that by making anesthetic with ethyl chloride the skin directly over the femoral vein high in the groin, the vein could be quickly bared, cleared of connective tissue, tied, and opened without causing any general disturbance whatever. A long, fine, flexible catheter (2.4 millimeters in diameter) which had previously been coated with vaseline inside and out, to lubricate it and to delay the clotting of blood within it, was now introduced into the opening in the femoral vein, thence through the iliac and on into the inferior cava to a point near the level of the sternal notch. A thread tied around this tube where, after being inserted to the proper distance, it disappeared into the femoral vein, marked the extent of insertion, and permitted a later introduction to the same extent. This slight operation—a venesection, commonly practised on[p. 46] our ancestors—consumed only a few minutes, and as the only possibility of causing pain was guarded against by local anesthesia, the animal remained tranquil throughout. Occasionally it was necessary to stroke the cat’s head gently to keep her quiet on the holder, and under such circumstances I have known her to purr during all the preparations for obtaining the blood, and while the blood was being taken.
The blood (3 or 4 cubic centimeters) was slowly drawn through the catheter into a clean glass syringe. Care was taken to avoid any marked suction such as might cause collapse of the vein near the inner opening of the tube. As soon as the blood was secured, the catheter was removed and the vein tied loosely, to prevent bleeding. The blood was at once emptied into a beaker, and the fibrin whipped from it by means of fringed rubber tubing fitted over a glass rod. Since this defibrinated blood was obtained while the animal was undisturbed, it was labelled “quiet blood.”
The animal was then exposed to the barking dog, as already described, and immediately thereafter blood was again removed, from precisely the same region as before. This sample, after being defibrinated, was labelled “excited blood.” The two samples, the “quiet” and the “excited,” both obtained in the same manner and subsequently[p. 47] treated in the same manner, were now tested for their content of adrenin.
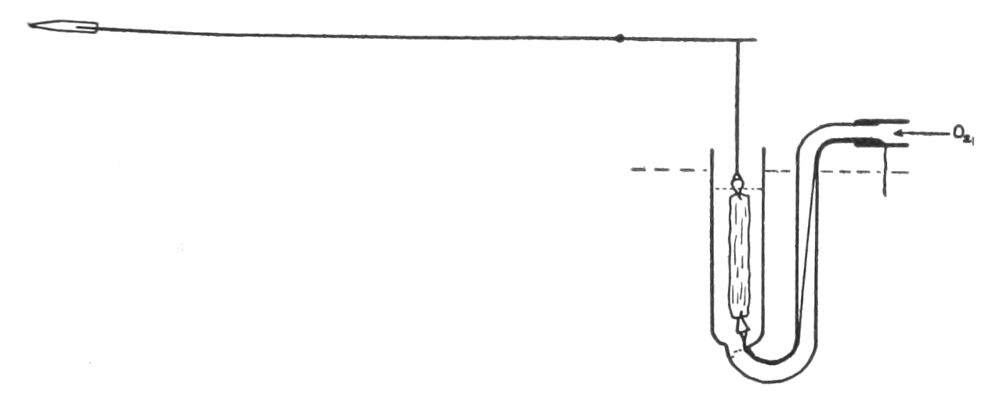
It was desirable to use as a test tissues to which the blood was naturally related. As will be recalled, adrenin affects viscera even after they have been removed from the body, just as if they were receiving impulses via sympathetic fibres, and further, that sympathetic fibres normally deliver impulses which cause contraction of the internal genitals and relaxation of the stomach and intestines. The uterus has long been employed as a test for adrenin, the presence of which it indicates by increased contraction. That isolated strips of the longitudinal muscle of the intestine, which are contracting rhythmically, are characteristically inhibited by adrenin in dilutions of 1 part in 20 millions, had been shown by Magnus in 1905. Although, previous to our investigation in 1910, this extremely delicate reaction had not been used as a biological signal for adrenin, it possesses noteworthy advantages over other methods. The intestine is found in all animals and not in only half of them, as is the uterus; it is ready for the test within a few minutes, instead of the several hours said to be required for the best use of the uterus preparation;[11] and it responds by relaxing. This last characteristic[p. 48] is especially important, for in defibrinated blood there are, besides adrenin, other substances capable of causing contraction of smooth muscle,[12] and liable therefore to lead to erroneous conclusions when a structure which responds by contracting, such as uterus or artery, is used to prove whether adrenin is present. On the other hand, substances producing relaxation of smooth muscle are few, and are unusual in blood.[13]
We used, therefore, the strip of intestinal muscle as an indicator. Later Hoskins[14] modified our procedure by taking, instead of the strip, a short segment of the rabbit intestine. The segment is not subjected to danger of injury during its preparation, and when fresh it is almost incredibly sensitive. It may be noticeably inhibited by adrenin, 1 part in 200 millions!
The strip, or the intestinal segment, was suspended between minute wire pincers (serres fines) in a cylindrical chamber 8 millimeters in diameter and 5 centimeters deep. By a thread attached to the lower serre fine the preparation was drawn into the chamber, and was held firmly; by the upper one it was attached to the short end of a writing lever (see Fig. 2). When not exposed to blood, the strip was immersed in a normal solution of the blood salts (Ringer’s). The blood or the salt solution could be quickly withdrawn from or introduced into the chamber, without[p. 49] disturbing the muscle, by means of a fine pipette passed down along the inner surface. The chamber and its contents, the stock of Ringer’s solution, and the samples of “quiet” and “excited” blood were all surrounded by a large volume of water kept approximately at body temperature (37° C.). Through the blood or the salt solution in the chamber oxygen was passed in a slow but steady stream of bubbles. Under these circumstances the strip will live for hours, and will contract and relax in a beautifully regular rhythm, which may be recorded graphically by the writing lever.
The first effect of surrounding the muscle with blood, whether “quiet” or “excited,” was to send it into a strong contraction which might persist, sometimes with slight oscillations, for a minute or two (see Figs. 4 and 5). After the initial shortening, the strip, if in quiet blood soon began to[p. 50] contract and relax rhythmically and with each relaxation to lengthen more, until a fairly even base line appeared in the written record. At this stage the addition of fresh “quiet” blood usually had no effect, even though the strip were washed once with Ringer’s solution before the second portion of the blood was added. For comparison of the effects of “quiet” and “excited” blood on the contracting strip, the two samples were each added to the muscle immediately after the Ringer’s solution had been removed, or they were applied to the muscle alternately and the differences in effect then noted. The results obtained by these methods are next to be presented.
1 Jacobi: Archiv für experimentelle Pathologie und Pharmakologie, 1891, xxix, p. 185.
2 Biedl: Archiv für die gesammte Physiologie, 1897, lxvii, pp. 456, 481.
3 Dreyer: American Journal of Physiology, 1898–99, ii, p. 219.
4 Tscheboksaroff: Archiv für die gesammte Physiologie, 1910, cxxxvii, p. 103.
5 Zeitschrift für Biologie, 1912, lviii, p. 274.
6 Meltzer and Joseph: American Journal of Physiology, 1912, xxix, p. xxxiv.
7 Elliott: Journal of Physiology, 1912, xliv, p. 400.
8 Cannon and Lyman: American Journal of Physiology, 1913, xxxi, p. 377.
9 Elliott: Journal of Physiology, 1912, xliv, p. 400.
10 Folin, Cannon and Denis: Journal of Biological Chemistry, 1913, xiii, p. 477.
[p. 51]
11 Fraenkel: Archiv für experimentelle Pathologie und Pharmakologie, 1909, lx, p. 399.
12 See O’Connor: Archiv für die experimentelle Pathologie und Pharmakologie, 1912, lxvii, p. 206.
13 Grutzner: Ergebnisse der Physiologie, 1904, iii2, p. 66; Magnus: Loc. cit., p. 69.
14 Hoskins: Journal of Pharmacology and Experimental Therapeutics, 1911, iii, p. 95.
[p. 52]
ADRENAL SECRETION IN STRONG EMOTIONS AND PAIN
If the secretion of adrenin is increased in strong emotional states and in pain, that constitutes a fact of considerable significance, for, as already mentioned, adrenin is capable of producing many of the bodily changes which are characteristically manifested in emotional and painful experiences. It is a matter of prime importance for further discussion to determine whether the adrenal glands are in fact roused to special activity in times of stress.
That blood from the adrenal veins causes the relaxation of intestinal muscle characteristic of adrenal extract or adrenin is shown in Fig. 3. The muscle was originally beating in blood which contained no demonstrable amount of adrenal secretion; this inactive blood was replaced by blood[p. 53] from the adrenal veins, obtained after quick etherization. Etherization, it will be recalled, is accompanied by a “stage of excitement.” Relaxation occurred almost immediately (at b). Then the rhythm was renewed in the former blood, and thereupon the muscle was surrounded with blood from the vein leading away from the left kidney, i. e., blood obtained from the same animal and under the same conditions as the adrenal blood, but from a neighboring vein. No relaxation occurred. By this and other similar tests the reliability of the method was proved.
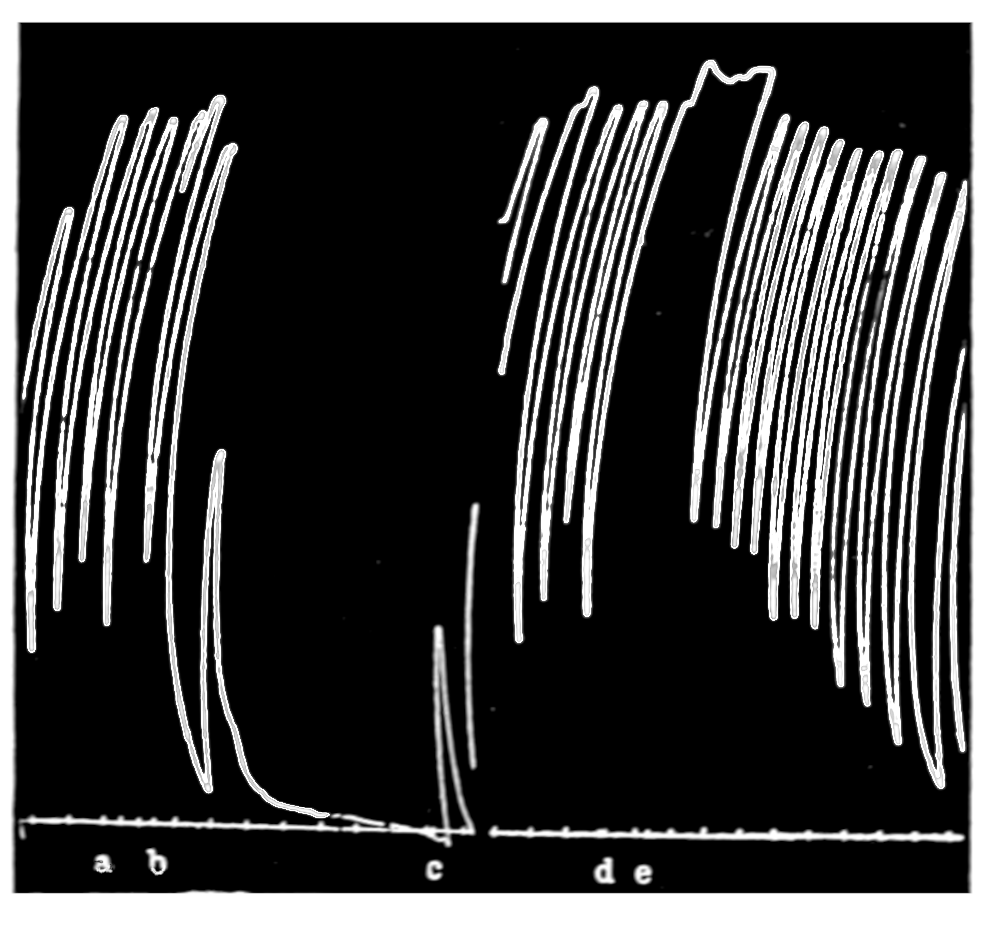
Figure 3.—Intestinal muscle beating in inactive blood, which was withdrawn from the chamber at a. Blood from the adrenal vein of an animal excited by etherization was substituted at b, and withdrawn at c. Contractions were restored in the original inactive blood which was removed at d. Blood from the renal vein (same animal) was added at e.
In this and subsequent records time is marked in half minutes.
[p. 54]
In no instance did blood from the inferior vena cava of the quiet normal animal produce relaxation. On the other hand, blood from the animal after emotional excitement showed more or less promptly the typical relaxation. In Fig. 4 is represented the record of intestinal muscle which was beating regularly in Ringer’s solution. At a the Ringer’s solution was removed, and at b “excited” blood was added; after the preliminary shortening, which, as already stated, occurs at the first immersion in blood, the muscle lengthened gradually into complete inhibition. At c the “excited” blood was removed, and at d “quiet” blood was added in its place. The muscle at once began fairly regular rhythmic beats. At e the “quiet” blood was removed, and at f the “excited” blood was again applied. The muscle lengthened almost immediately into an inhibited state. In this instance the “excited” blood was taken after[p. 55] the cat had been barked at for about fifteen minutes.
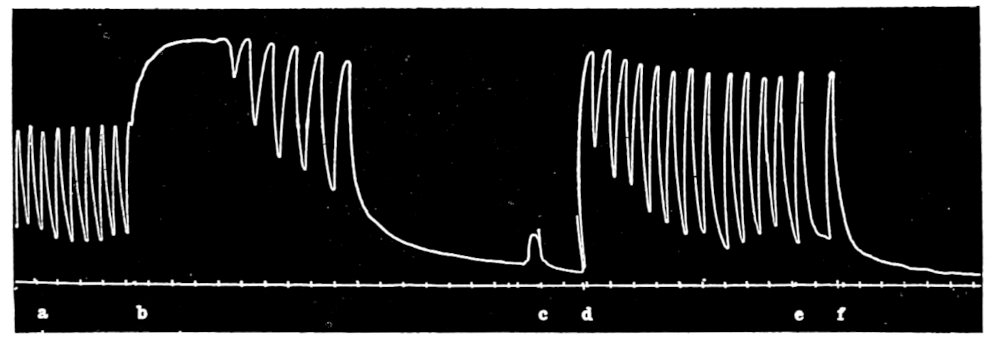
The increase of effect with prolongation of the period of excitement is shown in Fig. 5. A is the record of contractions after the muscle was surrounded with “quiet” blood serum. B shows the gradual inhibition which occurred when the muscle was surrounded with defibrinated blood taken when the animal had been excited eleven minutes. And C is the record of rapid inhibition after fifteen minutes of excitement. In other instances the effect was manifested merely by a lowering of the tonus of the muscle, and a notable slowing of the beats, without, however, a total abolition of them.
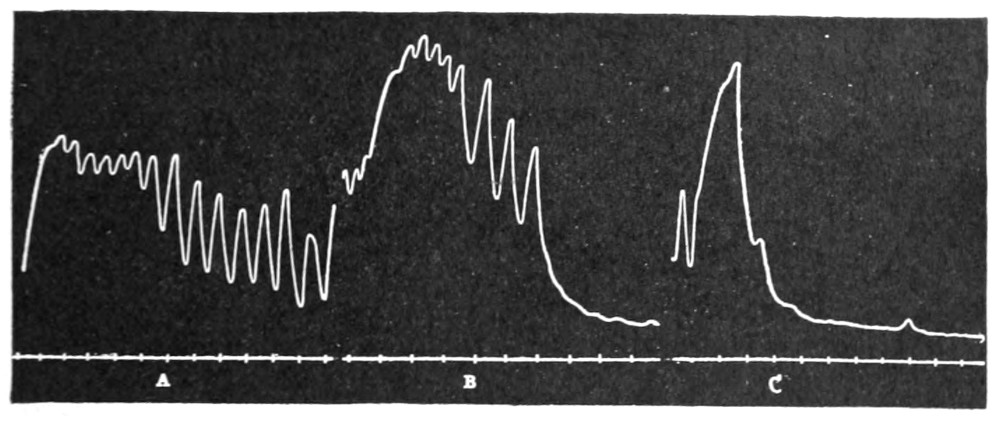
The inference that this inhibition of contraction of the intestinal muscle is due to an increased amount of adrenal secretion in the “excited”[p. 56] blood de la Paz and I justified on several grounds:
(1) The inhibition was produced by “excited” blood from the inferior vena cava anterior to the mouths of the adrenal veins, when blood from the femoral vein, taken at the same time, had no inhibitory influence. Since blood from the femoral vein is typical of the cava blood below the entrance of the kidney veins, the conclusion is warranted that the difference of effect of the two samples of blood is not due to any agent below the kidneys. But that blood from the kidneys does not cause the relaxation is shown in Fig. 3. The only other structures which could alter the blood between the two points at which it was taken are the adrenal glands, and the material[p. 57] secreted by them would produce precisely the inhibition of contraction which was in fact produced.
(2) If in ether anesthesia the blood vessels leading to and from the adrenal glands are first carefully tied, and then the glands are removed, excitement four or five hours later, before the weakness that follows the removal has become prominent, does not alter the blood so that the typical inhibition occurs (see Fig. 6). Thus, although the animal shows all the characteristic signs of sympathetic stimulation, the blood, in the absence of the adrenals, remains unchanged.
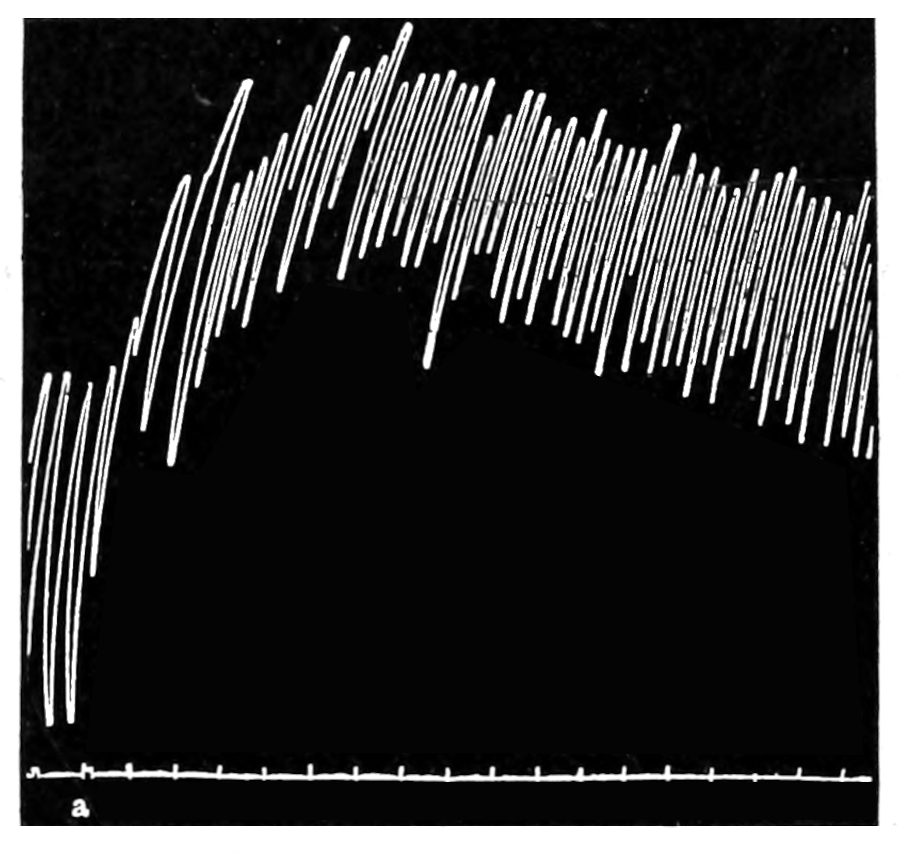
(3) As already shown, sometimes the effect produced[p. 58] by the “excited” blood was prompt inhibition, sometimes the inhibition followed only after several beats, and sometimes a slowing and shortening of contractions, with a lower tone, were the sole signs of the action of adrenin. All these degrees of relaxation can be duplicated by adding to inactive blood varying amounts of adrenin. Fig. 7 shows the effects, on a somewhat insensitive muscle preparation, of adding adrenin, 1:1,000,000 (A), 1:2,000,000 (B), and 1:3,000,000 (C), to different samples of blood previously without inhibitory influence. These effects of adrenin and the effects produced by blood taken near the opening of the adrenal veins are strikingly analogous.
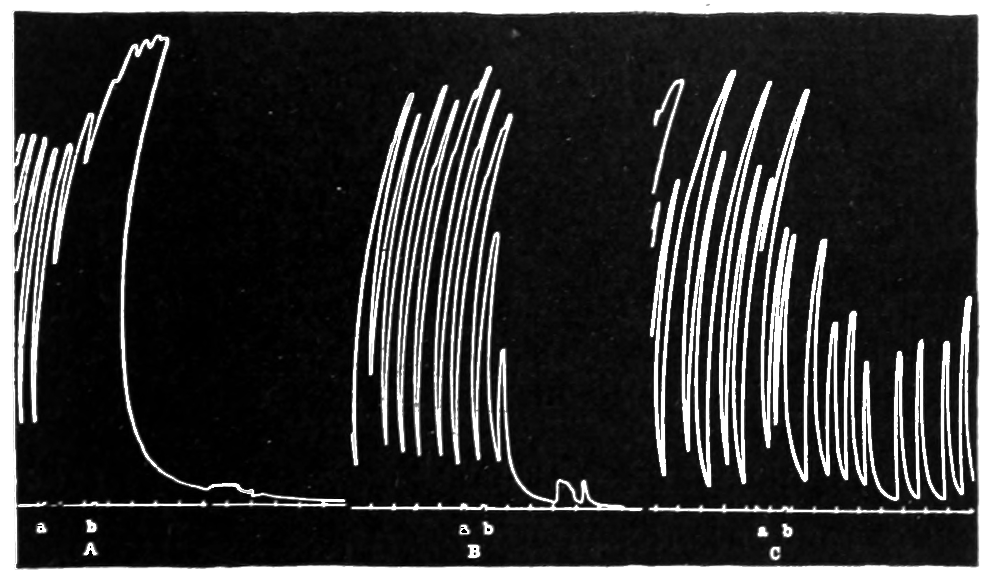
(4) Embden and v. Furth[1] have reported that 0.1 gram of suprarenin chloride disappears almost completely in two hours if added to 200 cubic centimeters of defibrinated beef blood, and the mixture constantly aerated at body temperature. “Excited” blood which produces inhibition loses that power on standing in the cold for twenty-four hours, or on being kept warm and agitated with bubbling oxygen. This change is illustrated in Fig. 8; the power of the “excited” blood to inhibit the contractions of the intestinal muscle when record A was written was destroyed after three hours of exposure to bubbling oxygen, as shown by record B. The destruction of adrenin and[p. 59] the disappearance of the effect which adrenin would produce are thus closely parallel.
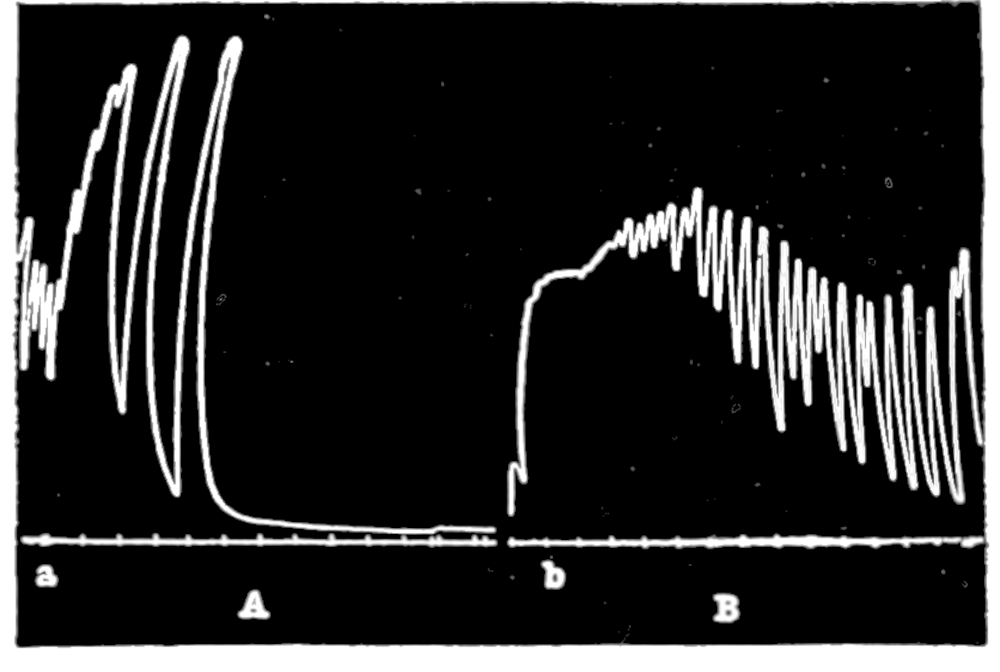
All these considerations, taken with the proof that sympathetic impulses increase secretion of the adrenal glands, and taken also with the evidence that, during such emotional excitement as was employed in these experiments, signs of sympathetic discharges appeared throughout the animal from the dilated pupil of the eye to the standing hairs of the tail-tip, led us to the conclusions that the characteristic action of adrenin on intestinal muscle was in fact, in our experiments, due to secretion of the adrenal glands, and that that secretion is increased in great emotion.
As mentioned in the first chapter, stimulation of sensory fibres in one of the larger nerve trunks[p. 60] is known to result in such nervous discharges along sympathetic paths as to produce marked inhibition of digestive processes. Other manifestations of sympathetic innervations—e. g., contraction of arterioles, dilation of pupils, erection of hairs—are also demonstrable. And since the adrenal glands are stimulated to activity by sympathetic impulses, it was possible that they would be affected as are other structures supplied with sympathetic fibres, and that they would secrete in greater abundance when sensory nerves were irritated.
The testing of this possibility was undertaken by Hoskins and myself in 1911. Since bodily changes from “painful” stimulation can in large degree be produced in an anesthetized animal, without, however, an experience of pain by the animal, it was possible to make the test quite simply. The sensory stimulus was a rapidly interrupted induced current applied to the sciatic nerve. The current was increased in strength as time passed, and thus the intensity of the effect, indicated by continuous dilation of the pupils, was maintained. There was no doubt that such stimulation would have caused very severe pain if the animal had not been anesthetized. Indeed, the stimulus used was probably much stronger than would be necessary to obtain a positive result in the absence of the anesthetic (urethane), which markedly lessens the irritability[p. 61] of visceral nerve fibres.[2] In different instances the stimulation lasted from three to six minutes. Throughout the period there was markedly increased rapidity and depth of breathing.
As Fig. 9 shows, the normal blood, removed[p. 62] from the vena cava before stimulation, caused no inhibition of the beating segment, whereas that removed afterwards produced a deep relaxation. Hoskins and I showed that the increased respiration which accompanies “painful” stimulation does not augment adrenal activity. We concluded, therefore, that when a sensory trunk is strongly excited the adrenal glands are reflexly stimulated, and that they pour into the blood stream an increased amount of adrenin.
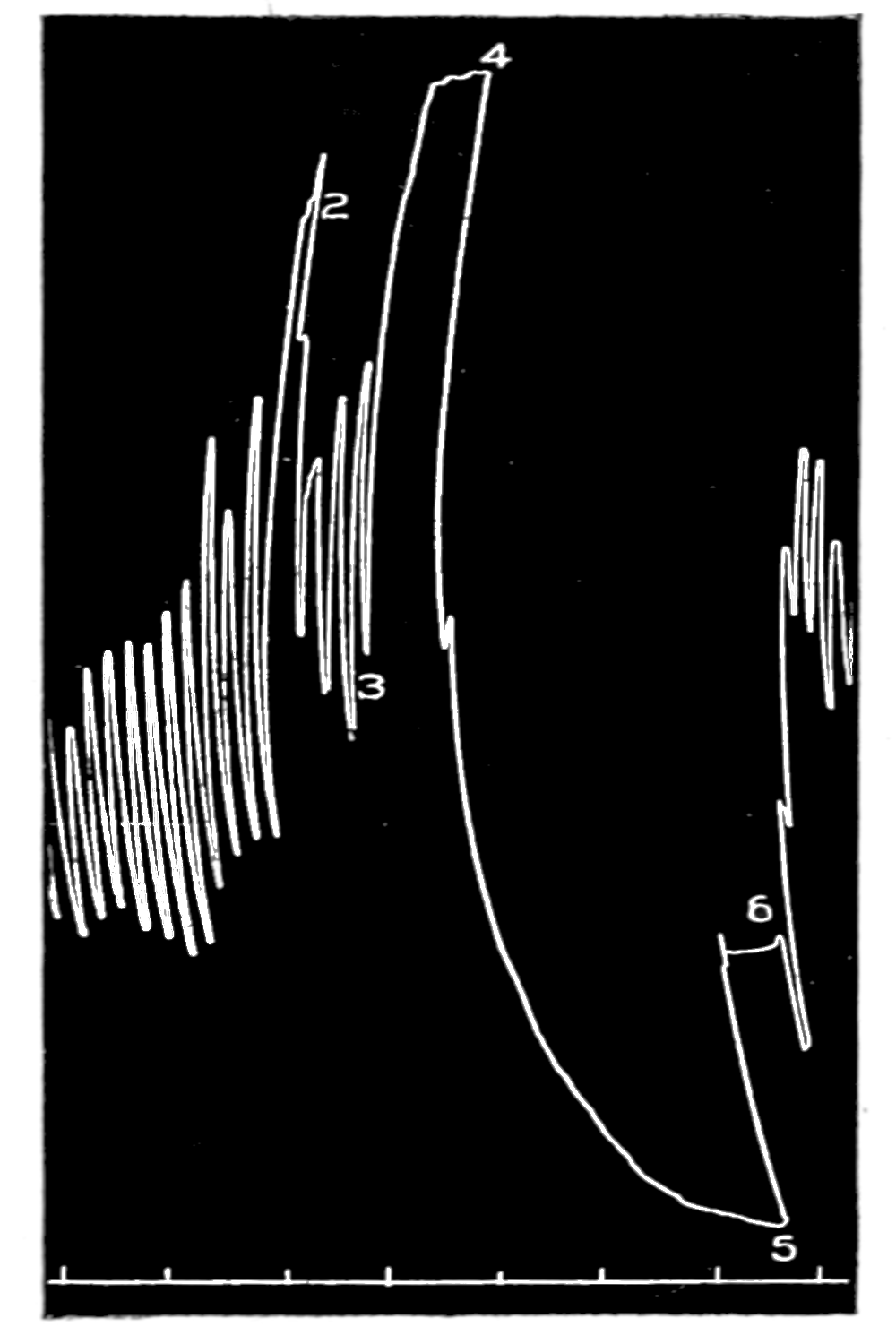
The foregoing experiments and conclusions were reported in 1911. In 1912, Elliott[3] brought confirmatory evidence by use of a method quite different from ours. As previously stated, he studied the effects of experimental procedures on adrenal secretion by a careful comparative quantitative assay of the adrenin content of the glands when one gland was isolated from the central nervous system and the other left connected. He took advantage of the action of morphia and of the substance Β-tetrahydronaphthylamine in evoking in cats all the appearances of great fright. After the animals had thus been “frightened,” he found that the adrenal gland which was still connected with the spinal cord was much depleted of its adrenin content compared with the other, isolated gland. And he observed, further, that animals[p. 63] newly brought to the laboratory, and evidently disturbed by the strangeness of their surroundings, had a considerably smaller amount of adrenin in their glands than other animals grown accustomed to the situation. Elliott also observed that prolonged excitation of a sensory nerve, such as the great sciatic, may cause the adrenin largely to disappear from the gland still connected with the central nervous system and subjected, therefore, to reflex influences.
Our conclusions have also been confirmed more recently (1913) by Hitchings, Sloan and Austin,[4] working in Crile’s laboratory in Cleveland. They used the same method which we had used to obtain blood and to test for adrenin, and found that after great fear and rage had been induced in a cat by the attempt of a muzzled dog to fight it, the adrenin reaction was clearly demonstrable. And just as we had noted that the reaction did not occur if the adrenal glands had been removed, they showed that it did not occur if the nervous connections with the spinal cord were previously severed.
The logic of all these experiments may be briefly summed up. That the adrenal glands are subject to splanchnic influence has been demonstrated anatomically and by the physiological effects of their secretion after artificial stimulation of the splanchnic nerves. Impulses are normally sent along these nerves, in the natural conditions of[p. 64] life, when animals become greatly excited, as in fear and rage and pain. There is every probability, therefore, that these glands are stimulated to extra secretion at such times. Both by an exceedingly delicate biological test (intestinal muscle) and by an examination of the glands themselves, clear evidence has been secured that in pain and deep emotion the glands do, in fact, pour out an excess of adrenin into the circulating blood.
Here, then, is a remarkable group of phenomena—a pair of glands stimulated to activity in times of strong excitement and by such nerve impulses as themselves produce at such times profound changes in the viscera; and a secretion given forth into the blood stream by these glands, which is capable of inducing by itself, or of augmenting, the nervous influences which induce the very changes in the viscera which accompany suffering and the major emotions. What may be the significance of these changes, occurring when conditions of pain and great excitement—experiences common to animals of most diverse types and probably known to their ancestors for ages past—lay hold of the bodily functions and determine the instinctive responses?
Certain remarkable effects of injecting adrenin into the blood have for many years been more or less well recognized. For example, when injected it causes liberation of sugar from the liver into[p. 65] the blood stream. It relaxes the smooth muscle of the bronchioles. Some old experiments indicated that it acts as an antidote for muscular fatigue. It alters the distribution of the blood in the body, driving it from the abdominal viscera into the heart, lungs, central nervous system and limbs. And there was some evidence that it renders more rapid the coagulation of the blood. There may be other activities of adrenin not yet discovered—it may coöperate with the products of other glands of internal secretion. And other glands of internal secretion may be stimulated by sympathetic impulses. But we were not concerned with these possibilities. We wished to know whether the adrenin poured out in pain and emotional excitement produced or helped to produce the same effects that follow the injection of adrenin. Our later researches were concerned with answers to this question.
1 Embden and v. Furth: Hofmeister’s Beiträge zur chemischen Physiologie und Pathologie, 1904, iv, p. 423.
2 Elliott: Journal of Physiology, 1905, xxxii, p. 448.
3 Elliott: Journal of Physiology, 1912, xliv, p. 409.
4 Hitchings, Sloan and Austin: Cleveland Medical Journal, 1913, xii, p. 686; see also Crile and Lower: Anoci-association, Philadelphia, 1914, p. 56.
[p. 66]
THE INCREASE OF BLOOD SUGAR IN PAIN AND GREAT EMOTION
Sugar is the form in which carbohydrate material is transported in organisms; starch is the storage form. In the bodies of animals that have been well fed the liver contains an abundance of glycogen or “animal starch,” which may be called upon in times of need. At such times the glycogen is changed, and set free in the blood as sugar. Ordinarily there is a small percentage of sugar in the blood—from 0.06 to 0.1 per cent. When only this small amount is present the kidneys are capable of preventing its escape in any noteworthy amount. If the percentage rises to the neighborhood of 0.2–0.3 per cent, however, the sugar passes the obstacle set up by the kidneys, and is readily demonstrable in the urine by ordinary tests. The condition of “glycosuria,” therefore, may properly be considered, in certain circumstances, as evidence of increased sugar in the blood. The injection of adrenin can liberate sugar from the[p. 67] liver to such an extent that glycosuria results. Does the adrenal secretion discharged in pain and strong emotional excitement play a rôle in producing glycosuria under such conditions?
In clinical literature scattered suggestions are to be found that conditions giving rise to emotional states may be the occasion also of more or less permanent glycosuria. Great grief and prolonged anxiety during a momentous crisis have been regarded as causes of individual instances of diabetes, and anger or fright has been followed by an increase in the sugar excreted by persons who already have the disease. Kleen[1] cites the instance of a German officer whose diabetes and whose Iron Cross for valor both came from a stressful experience in the Franco-Prussian War. The onset of the disease in a man directly after his wife was discovered in adultery is described by Naunyn;[2] and this author also mentions two cases in his own practice—one started during the bombardment of Strassburg (1870), the other started a few days after a companion had shot himself. In cases of mental disease, also, states of depression have been described accompanied by sugar in the urine. Schultze[3] has reported that in these cases the amount of glycosuria is dependent on the degree of depression, and that the greatest excretion of sugar occurs in the fear-psychoses. Raimann[4] has reported that in both[p. 68] melancholia and mania the assimilation limit of sugar may be lowered. Similar results in the insane have recently been presented by Mita,[5] and by Folin and Denis.[6] The latter investigators found glycosuria in 12 per cent of 192 insane patients, most of whom suffered from depression, apprehension, or excitement. And Arndt[7] has observed glycosuria appearing and disappearing as alcoholic delirium appeared and disappeared in his patients.
Although clinical evidence thus indicates an emotional origin of some cases of diabetes and glycosuria, the intricacies of existence and the complications of disease in human beings throw some doubt on the value of that evidence. Both Naunyn[8] and Hirschfeld, although mentioning instances of diabetes apparently due to an emotional experience, urge a skeptical attitude toward such statements. It is desirable, therefore, that the question of an emotional glycosuria be tested under simpler and more controllable conditions. “Emotional glycosuria” in experimental animals has indeed been referred to by Waterman and Smit[9] and more recently by Henderson and Underhill.[10] Both these references, however, are based on the work of Böhm and Hoffmann,[11] reported in 1878.
[p. 69]
Böhm and Hoffmann found that cats, when bound to an operating board, a tube inserted into the trachea (without anesthesia), and in some instances a catheter inserted into the urethra through an opening above the pubis, had in about half an hour an abundance of sugar in the urine. In three determinations sugar in the blood proved slightly above “normal” so long as sugar was appearing in the urine, but returned to “normal” as the glycosuria disappeared. Since they were able to produce the phenomenon by simply binding animals to the holder, they called it “Fesselungsdiabetes.”
As possible causes of this glycosuria in bound animals, they considered opening the trachea, cooling, and pain. The first two they readily eliminated, and still they found sugar excreted. Pain they could not obviate, and since, without binding the animals, they caused glycosuria by merely stimulating the sciatic nerves, they concluded that painful confinement was itself a sufficient cause. Other factors, however, such as cooling and circulatory disturbances, probably coöperated with pain, they believed, to produce the result. Their observations on cats have been proved true also of rabbits;[12] and recently it has been shown that an operation involving some pain increases blood sugar in dogs.[13] Temporary glycosuria[p. 70] has likewise been noted in association with intense pain in human beings.
Inasmuch as Böhm and Hoffmann did not mention the emotional element in discussing their results, and inasmuch as they admitted that they could not obviate from their experimental procedure pain, which they themselves proved was effective in causing glycosuria, designating what they called “Fesselungsdiabetes” as “emotional glycosuria” is not justified.
The discovery that during strong emotion adrenal secretion is increased, and the fact that injection of adrenin gives rise to glycosuria, suggested that glycosuria might be called forth by emotional excitement, and then that even without the painful element of Böhm and Hoffmann’s experiments, sugar might be found in the urine. The testing of this possibility was undertaken by A. T. Shohl, W. S. Wright and myself in 1911.
Our first procedure was a repetition of Böhm and Hoffmann’s experiments, freed from the factor of pain. The animals (cats) were bound to a comfortable holder, which left the head unfastened. This holder I had used hundreds of times in X-ray studies of digestion, with many different animals, without causing any signs of even so much as uneasiness. Just as in observations[p. 71] on the movements of the alimentary canal, however, so here, the animals reacted differently to the experience of being confined. Young males usually became quite frantic, and with eyes wide, pupils dilated, pulse accelerated, hairs of the tail more or less erect, they struggled, snarling and growling, to free themselves. Females, on the contrary, especially if elderly, were as a rule much more calm, and resignedly accepted the novel situation.
According to differences in reaction the animals were left in the holder for periods varying in length from thirty minutes to five hours. In order to insure prompt urination, considerable quantities of water were given by stomach tube at the beginning of the experiment and in some cases again later. Arrangements were made for draining the urine promptly, when the animal was on the holder or when afterwards in a metal metabolism cage, into a glass receiver containing a few drops of chloroform to prevent fermentation. The diet in all cases consisted of customary raw meat and milk. In every instance the urine was proved free from sugar before the animal was excited.
In our series of observations twelve cats were used, and in every one a well-marked glycosuria was developed. The shortest periods of confinement to the holder which were effective were thirty[p. 72] and forty minutes; the longest we employed, five hours. The average time required to bring about a glycosuria was less than an hour and a half; the average in seven of the twelve cases was less than forty minutes. In all cases no sugar was found in the urine passed on the day after the excitement.
The promptness with which the glycosuria developed was directly related to the emotional state of the animal. Sugar was found early in animals which early showed signs of being frightened or in a rage, and much later in animals which took the experience more calmly.
As cooling may result in increased sugar in the blood, and consequent glycosuria, the rectal temperature was observed from time to time, and it was found to vary so slightly that in these experiments it was a wholly negligible factor. In one cat the rectal temperature fell to 36° C. while the animal was bound and placed in a cold room (about 2° C.) for fifty minutes, but no sugar appeared in the urine.
Further evidence that the appearance of sugar in the urine may arise purely from emotional excitement was obtained from three cats which gave negative results when bound in the holder for varying periods up to four hours. It was noteworthy that these animals remained calm and passive in their confinement. When, however,[p. 73] they were placed, separately, in a small wire cage, and were barked at by an energetic little dog, that jumped at them and made signs of attack, the cats became much excited, they showed their teeth, humped their backs, and growled defiance. This sham fight was permitted to continue for a half hour in each of the three cases. In each case the animal, which after four hours of bondage had exhibited no glycosuria, now had sugar in the urine. Pain, cooling, and bondage were not factors in these experiments. The animal was either frightened or enraged by the barking dog, and that excitement was attended by glycosuria.
The sugar excreted in the twenty-four hours which included the period of excitement was determined by the Bertrand method.[14] It ranged from 0.024 gram to 1.93 grams, or from 0.008 gram to 0.62 gram per kilo body weight, for the twenty-four hours’ quantity.
The presence of sugar in the urine may be used as an indication of increased sugar in the blood, for unless injury has been done to the cells of the kidneys, they do not permit sugar to escape until the percentage in the blood has risen to a considerable degree. Thus, though testing the urine reveals the instances of a high content of blood sugar, it does not show the fine variations that appear when the blood itself is examined. Recently Scott[15] has concluded a thorough investigation[p. 74] of the variations of blood sugar in cats, and has found that merely incidental conditions, producing even mild excitement, as indicated by crying or otherwise, result in a noticeable rise in the amount. Indeed, so sensitive is the sugar-liberating mechanism that all the early determinations of the “normal” content of sugar in blood which has been drawn from an artery or vein in the absence of anesthesia, are of very doubtful value. Certainly when care is taken to obtain blood suddenly from a tranquil animal, the percentage (0.069, Scott; 0.088, Pavy) is much less than when the blood is drawn without anesthesia (0.15, Böhm and Hoffmann), or after light narcosis (0.282, Rona and Takahashi[16]).
Our observations on cats have since been found valid for rabbits. Rolly and Oppermann, Jacobsen, and Hirsch and Reinbach[17] have recently recorded that the mere handling of a rabbit preparatory to operating on it will increase the percentage of blood sugar (in some cases from 0.10 to 0.23 and 0.27 per cent). Dogs are said to be much less likely to be disturbed by the nature of their surroundings than are rabbits and cats. Nevertheless, pain and excitement are such fundamental experiences in animals that without much doubt the same mechanism is operative in all when these experiences occur. Probably, just as the digestion of dogs is disturbed by strong emotion,[p. 75] the blood sugar likewise is increased, for sympathetic impulses occasion both changes.[*] Gib has given an account of a bitch that became much agitated when shut up, and after such enforced seclusion, but never otherwise, she excreted small quantities of sugar in the urine.[18]
* Since the foregoing sentences were written Hirsch and Reinbach have reported (Zeitschrift für physiologische Chemie, 1914, xci, p. 292) a “psychic hyperglycemia” in dogs, that resulted from fastening the animals to a table. The blood sugar rose in one instance from 0.11 to 0.14 per cent, and in another from 0.09 to 0.16 per cent.
The results noted in these lower animals have been confirmed in human beings. One of my former students, W. G. Smillie, found that four of nine medical students, all normally without sugar in their urine, had glycosuria after a hard examination, and only one of the nine had glycosuria after an easier examination. The tests, which were positive with Fehling’s solution, Nylander’s reagent, and also with phenyl-hydrazine, were made on the first urine passed after the examination. Furthermore, C. H. Fiske and I examined the urine of twenty-five members of the Harvard University football squad immediately after the final and most exciting contest of the season of 1913, and found sugar in twelve cases. Five of these positive cases were among substitutes not called upon to enter the game. The only excited spectator of the Harvard[p. 76] victory whose urine was examined also had a marked glycosuria, which on the following day had disappeared.
Other tests made on students before and after important scholastic examinations have been published by Folin, Denis and Smillie.[19] Of thirty-four second-year medical students tested, one had sugar before the examination as well as afterwards. Of the remaining thirty-three, six, or 18 per cent, had small but unmistakable traces of sugar in the urine passed directly following the ordeal. A similar study was made on second-year students at a women’s college. Of thirty-six students who had no sugar in the urine on the day before, six, or 17 per cent, eliminated sugar with the urine passed immediately after the examination.
From the foregoing results it is reasonable to conclude that just as in the cat, dog, and rabbit, so also in man, emotional excitement produces temporary increase of blood sugar.
Since artificial stimulation of the splanchnic nerves produces glycosuria,[20] and since major emotions, such as rage and fright, are attended by nervous discharges along splanchnic pathways, glycosuria as an accompaniment of emotional excitement[p. 77] would naturally be expected to occur. To what extent the adrenal glands which, as already mentioned, are stimulated to increased secretion by excitement, might play a part in this process, has been in dispute. Removal of these glands or cutting of the nerve fibres supplying them, according to some observers,[21] prevents glycosuria after puncture of the fourth ventricle of the brain (the “sugar puncture,” which typically induces glycosuria) and also after stimulation of the splanchnics.[22] On the other hand, Wertheimer and Battez[23] have stated that removal of the glands does not abolish the effects of sugar puncture in the cat. It was questionable, therefore, whether removal of the adrenal glands would affect emotional glycosuria.
Evidence on this point I secured with Shohl and Wright in observations on three animals in which the adrenals were removed aseptically under ether. The animals selected had all become quickly excited on being bound to the holder, and had manifested glycosuria after about an hour of confinement. In the operation, to avoid discharge of adrenin by handling, the adrenal veins were first tied, and then the glands freed from their attachments and removed as quickly and with as little manipulation as possible. In one cat the entire operation was finished in twenty minutes. In two of the cats a small catheter was introduced into the[p. 78] urethra through an incision, so that the bladder could be emptied at any time.
In all three cases urine that was free from sugar was obtained soon after the operation. Although the animals deprived of their adrenals manifested a general lessening of muscular tone, they still displayed much of their former rage or excitement when bound. Indeed, one was more excited after removal of the adrenals than before. That the animals might not be excessively cooled they were kept warm with coverings or an electric heating pad. Although they were now bound for periods from two to three times as long as the periods required formerly to cause glycosuria, no trace of sugar was found in the urine in any instance. The evidence thus secured tends, therefore, to support the view that the adrenal glands perform an important contributory rôle in the glycosuria resulting from splanchnic stimulation.
Possibly the emotional element is in part accountable for the glycosuria observed after painful stimulation, but conditions causing pain alone will reasonably explain it. As we have already seen, strong stimulation of sensory fibres causes the discharge of impulses along the splanchnic nerves, and incidentally calls forth an increased secretion of the adrenal glands. In glycosuria resulting from painful stimulation, as well as in emotional[p. 79] glycosuria, the adrenal glands may be essential factors.
Later the evidence will be given that sugar is the optimum source of muscular energy. In passing, we may note that the liberation of sugar at a time when great muscular exertion is likely to be demanded of the organism may be interpreted as a highly interesting instance of biological adaptation.
1 Kleen: On Diabetes Mellitus and Glycosuria, Philadelphia, 1900, pp. 22, 37–39.
2 Naunyn: Der Diabetes Mellitus, Vienna, 1898, p. 72.
3 Schultze: Verhandlungen der Gesellschaft deutscher Naturforscher und Aerzte, Cologne, 1908, ii, p. 358.
4 Zeitschrift für Heilkunde, 1902, xxiii, Abtheilung iii, pp. 14, 19.
5 Mita: Monatshefte für Psychiatrie und Neurologie, 1912, xxxii, p. 159.
6 Folin, Denis and Smillie: Journal of Biological Chemistry, 1914, xvii, p. 519.
7 Arndt: Zeitschrift für Nervenheilkunde, 1897, x, p. 436.
8 Naunyn: Loc. cit., p. 73; Hirschfeld: Die Zuckerkrankheit, Leipzig, 1902, p. 45.
9 Waterman and Smit: Archiv für die gesammte Physiologie, 1908, cxxiv, p. 205.
10 Henderson and Underhill: American Journal of Physiology, 1911, xxviii, p. 276.
11 Böhm and Hoffmann: Archiv für experimentelle Pathologie und Pharmakologie, 1878, viii, p. 295.
12 Eckhard: Zeitschrift für Biologie, 1903, xliv, p. 408.
13 Loewy and Rosenberg: Biochemische Zeitschrift, 1913, lvi, p. 114.
14 See Abderhalden: Handbuch der biochemischen Arbeitsmethoden, Berlin, 1910, ii, p. 181.
[p. 80]
15 Scott: American Journal of Physiology, 1914, xxxiv, p. 283.
16 Cited by Scott: Loc. cit., p. 296.
17 Rolly and Oppermann: Biochemische Zeitschrift, 1913, xlix, p. 201. Jacobsen: Ibid., 1913, li, p. 449. Hirsch and Reinbach: Zeitschrift für physiologische Chemie, 1913, lxxxvii, p. 122.
18 Cited by Kleen: Loc. cit., p. 37.
19 Folin, Denis and Smillie: Loc. cit., p. 520.
20 See Macleod: American Journal of Physiology, 1907, xix, p. 405, also for other references to literature.
21 See Meyer: Comptes rendus de la Société de Biologie, 1906, lviii, p. 1123; Nishi: Archiv für experimentelle Pathologie und Pharmakologie, 1909, lxi, p. 416.
22 Gautrelet and Thomas: Comptes rendus de la Société de Biologie, 1909, lxvii, p. 233; and Macleod: Proceedings of the Society for Experimental Biology and Medicine, 1911, viii, p. 110 (true for left adrenal and left splanchnic).
23 Wertheimer and Battez: Archives Internationales de Physiologie, 1910, ix, p. 392.
[p. 81]
IMPROVED CONTRACTION OF FATIGUED MUSCLE AFTER SPLANCHNIC STIMULATION OF THE ADRENAL GLAND
In the older literature on the adrenal glands the deleterious effect of their absence, or the beneficial effect of injected extracts, on the contraction of skeletal muscle was not infrequently noted. As evidence accumulated, however, tending to prove an important relation between the extract of the adrenal medulla (adrenin) and the sympathetic nervous system, the relations with the efficiency of skeletal muscle began to receive less consideration.
The muscular weakness of persons suffering from diseased adrenals (Addison’s disease) was well recognized before experimental work on the glands was begun. Experiments on rabbits were reported in 1892 by Albanese,[1] who showed that muscles which were stimulated after removal of the glands were much more exhausted than when stimulated the same length of time in the same animal before the removal. Similarly Boinet[p. 82][2] reported, in 1895, that rats recently deprived of their adrenals were much more quickly exhausted in a revolving cage than were normal animals.
More direct evidence of the favorable influence of adrenal extract on skeletal muscle was brought forward by Oliver and Schäfer.[3] After injecting the extract subcutaneously into a frog they found that the excised gastrocnemius muscle registered a curve of contraction about 33 per cent higher and about 66 per cent longer than the corresponding muscle not exposed to the action of the extract. Similar prolongation of the muscle curve was observed after injecting the extract intravenously into a dog. A beneficial effect of adrenal extract on fatigued muscle, even when applied to the solution in which the isolated muscle was contracting, was claimed by Dessy and Grandis,[4] who studied the phenomenon in a salamander.[*] Further evidence leading to the same conclusion was offered in a discriminating[p. 83] paper by Panella.[5] He found that in cold-blooded animals the active principle of the adrenal medulla notably reënforced skeletal muscle, prolonging its ability to do work, and improving its contraction when fatigued. In warm-blooded animals the same effects were observed, but only after certain experimental procedures, such as anesthesia and section of the bulb, had changed them to a condition resembling the cold-blooded.
* These earlier investigations, in which an extract of the entire gland was used, made no distinction between the action of the medulla and that of the cortex. It may be that the weakness following removal or disease of the adrenals is due to absence of the cortex (see Hoskins and Wheelon: American Journal of Physiology, 1914, xxxiv, p. 184). Such a possible effect, however, should not be confused with the demonstrable influence of injected adrenin (derived from the adrenal medulla alone) and the similar effects from adrenal secretion caused by splanchnic stimulation.
The foregoing evidence indicates that removal of the adrenals has a debilitating effect on muscular power, and that injection of extracts of the glands has an invigorating effect. It seemed possible, therefore, that increased secretion of the adrenal glands, whether from direct stimulation of the splanchnic nerves or as a reflex result of pain or the major emotions, might act as a dynamogenic factor in the performance of muscular work. With this possibility in mind L. B. Nice and I[6] first concerned ourselves in a research which we conducted in 1912.
The general plan of the investigation consisted primarily in observing the effect of stimulating the splanchnic nerves, isolated from the spinal cord, on the contraction of a muscle whose nerve, also isolated from the spinal cord, was rhythmically and uniformly excited with break induction shocks. When a muscle is thus stimulated it[p. 84] at first responds by strong contractions, but as time passes the contractions become weaker, the degree of shortening of the muscle becomes less, and in this state of lessened efficiency it may continue for a long period to do work. The tired muscle which is showing continuously and evenly its inability to respond as it did at first, is said to have reached the “fatigue level.” This level serves as an excellent basis for testing influences that may have a beneficial effect on muscular performance, for the benefit is at once manifested in greater contraction.
In the experimental arrangement which we used, only a connection through the circulating blood existed between the splanchnic region and the muscle—all nervous relations were severed. Any change in muscular ability, therefore, occurring when the splanchnic nerve is stimulated, must be due to an alteration in the quantity or quality of the blood supplied to the laboring muscle.
Cats were used for most experiments, but results obtained with cats were confirmed on rabbits and dogs. To produce anesthesia in the cats and rabbits, and at the same time to avoid the fluctuating effects of ether, urethane (2 grams per kilo body weight) was given by a stomach tube. The animals were fastened back downward, over an electric warming pad, to an animal holder.[p. 85] Care was taken to maintain the body temperature at its normal level throughout each experiment.
The muscle selected to be fatigued was usually the extensor of the right hind foot (the tibialis anticus), though at times the common extensor muscle of the digits of the same foot was employed. The anterior tibial nerve which supplies these muscles was bared for about two centimeters, severed toward the body, and set in shielded electrodes, around which the skin was fastened by spring clips. Thus the nerve could be protected, kept moist, and stimulated without stimulation of neighboring structures. By a small slit in the skin the tendon of the muscle was uncovered, and after a strong thread was tied tightly about it, it was separated from its insertion. A nerve-muscle preparation was thereby made which was still connected with its proper blood supply. The preparation was fixed firmly to the animal holder by thongs looped around the hock and the foot, i. e., on either side of the slit through which the tendon emerged.
The thread tied to the tendon was passed over a pulley and down to a pivoted steel bar which bore a writing point. Both the pulley and this steel writing lever were supported in a rigid tripod. In the earliest experiments the contracting[p. 86] muscle was made to lift weights (125 to 175 grams); in all the later observations, however, the muscle pulled against a spring attached below the steel bar. The tension of the spring as the muscle began to lift the lever away from the support was, in most of the experiments, 110 grams, with an increase of 10 grams as the writing point was raised 4.5 millimeters. The magnification of the lever was 3.8.
The stimuli delivered to the anterior tibial nerve were, in most experiments, single break shocks of a value barely maximal when applied to the fresh preparation. The rate of stimulation varied between 60 and 300 per minute, but was uniform in any single observation. A rate which was found generally serviceable was 180 per minute.
Since the anterior tibial nerve contains fibres affecting blood vessels, as well as fibres causing contraction of skeletal muscle, the possibility had to be considered that stimuli applied to it might disturb the blood supply of the region. Constriction of the blood vessels would be likely to produce the most serious disturbance, by lessening the blood flow to the muscle. The observations of Bowditch and Warren,[7] that vasodilator rather than vasoconstrictor effects are produced by single induction shocks repeated at intervals of not more than five per second, reassured us as to the danger of diminishing the blood supply, for[p. 87] the rate of stimulation in our experiments never exceeded five per second and was usually two or three. Furthermore, in using these different rates we have never noted any result which could reasonably be attributed to a diminished circulation.
The splanchnic nerves were stimulated in various ways. At first only the left splanchnics in the abdomen were prepared. The nerves, separated from the spinal cord, were placed upon shielded electrodes. The form of electrodes which was found most satisfactory was that illustrated in Fig. 10. The instrument was made of a round rod of hard wood, bevelled to a point at one end, and grooved on the two sides. Into the grooves were pressed insulated wires ending in platinum hooks, which projected beyond the bevelled surface. Around the rod was placed an insulating rubber tube which was cut out so as to leave the hooks uncovered when the tube was slipped downward.
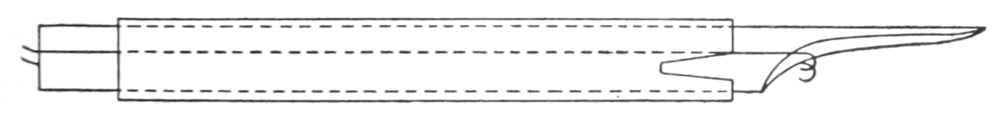
In applying the electrodes the left splanchnic nerves were first freed from their surroundings and tightly ligatured as close as possible to their[p. 88] origin. By means of strong compression the conductivity of the nerves was destroyed central to the ligature. The electrodes were now fixed in place by thrusting the sharp end of the wooden rod into the muscles of the back. This was so done as to bring the platinum hooks a few millimeters above the nerves. With a small seeker the nerves were next gently lifted over the hooks, and then the rubber tube was slipped downward until it came in contact with the body wall. Absorbent cotton was packed about the lower end of the electrodes, to take up any fluid that might appear; and finally the belly wall was closed with spring clips. The rubber tube served to keep the platinum hooks from contact with the muscles of the back and the movable viscera, while still permitting access to the nerves which were to be stimulated. This stimulating apparatus could be quickly applied, and, once in place, needed no further attention. In some of the experiments both splanchnic nerves were stimulated in the thorax. The rubber-covered electrode proved quite as serviceable there as in the abdomen.
The current delivered to the splanchnic nerves was a rapidly interrupted induced current of such strength that no effects of spreading were noticeable. That splanchnic stimulation causes secretion of the adrenal glands has been proved in[p. 89] many different ways which have already been described (see p. 41).
When skeletal muscle is repeatedly stimulated by a long series of rapidly recurring electric shocks, its strong contractions gradually grow weaker until a fairly constant condition is reached. The record then has an even top—the muscle has reached the “fatigue level.” The effect of splanchnic stimulation was tried when the muscle had been fatigued to this stage. The effect which was often obtained by stimulating the left splanchnic nerves is shown in Fig. 11. In this instance the muscle while relaxed supported no weight, and while contracting lifted a weight of 125 grams. The rate of stimulation was 80 per minute.
[p. 90]
The muscle record shows a brief initial rise from the fatigue level, followed by a drop, and that in turn by another, prolonged rise. The maximum height of the record is 13.5 millimeters, an increase of 6 millimeters over the height recorded before splanchnic stimulation. Thus the muscle was performing for a short period 80 per cent more work than before splanchnic stimulation, and for a considerably longer period exhibited an intermediate betterment of its efficiency.
The brief first elevation in the muscle record when registered simultaneously with arterial blood pressure is observed to occur at the same time[p. 91] with the sharp initial rise in the blood-pressure curve (see Fig. 12). The first sharp rise in blood pressure is due to contraction of the vessels in the area of distribution of the splanchnic nerves, for it does not appear if the alimentary canal is removed, or if the celiac axis and the superior and inferior mesenteric arteries are ligated. The betterment of the muscular contraction is probably due directly to the better blood supply resulting from the increased pressure, for if the adrenal veins are clipped and the splanchnic nerves are stimulated, the blood pressure rises as before and at the same time there may be registered a higher contraction of the muscle.
As Fig. 12 shows, the initial quick uplift in the blood-pressure record is quickly checked by a drop. This rapid drop does not appear when the adrenal veins are obstructed. A similar difference in blood-pressure records has been noted before and after excision of the adrenal glands. As Elliott,[8] and as Lyman and I[9] have shown, this sharp drop after the first rise, and also the subsequent elevation of blood pressure, are the consequences of liberation of adrenal secretion into the circulation. Fig. 12 demonstrates that the prolonged rise of the muscle record begins soon after this characteristic drop in blood pressure.
[p. 92]
If after clips have been placed on the adrenal veins so that no blood passes from them, the splanchnic nerves are stimulated, and later the clips are removed, a slight but distinct improvement in the muscular contraction occurs. As in the experiments of Young and Lehmann,[10] in which the adrenal veins were tied for a time and then released, the release of the blood which had been pent in these veins was quickly followed by a rise of blood pressure. The volume of blood thus restored to circulation was too slight to account for the rise of pressure. In conjunction with the evidence that splanchnic stimulation calls forth adrenal secretion, the rise may reasonably be attributed to that secretion. The fact should be noted, however, that in this instance the prolonged improvement in muscular contraction did not appear until the adrenal secretion had been admitted to the general circulation.
Many variations in the improvement of activity in fatigued muscle after splanchnic stimulation were noted in the course of our investigation. The improvement varied in degree, as indicated by increased height of the record. In some instances the height of contraction was doubled—a betterment by 100 per cent; in other instances the contraction after splanchnic stimulation was only a small fraction higher than that preceding the stimulation; and in still other instances there was no[p. 93] betterment whatever. Never, in our experience, were the augmented contractions equal to the original strong contractions of the fresh muscle.
The improvement also varied in degree as indicated by persistence of effect. In some instances the muscle returned to its former working level within four or five minutes after splanchnic stimulation ceased (see Fig. 11); and in other cases the muscle continued working with greater efficiency for fifteen or twenty minutes after the stimulation.
The evidence just presented has shown that splanchnic stimulation improves the contraction of fatigued muscle. Splanchnic stimulation, however, has two effects—it increases general arterial pressure and it also causes a discharge of adrenin from the adrenal glands. The questions now arise—Does splanchnic stimulation produce the improvement in muscular contraction by increasing the arterial blood pressure and thereby flushing the laboring muscles with fresh blood? Or does the adrenin liberated by splanchnic stimulation act itself, specifically, to improve the muscular contraction? Or may the two factors coöperate? These questions will be dealt with in the next two chapters.
[p. 94]
1 Albanese: Archives Italiennes de Biologie, 1892, xvii, p. 243.
2 Boinet: Comptes rendus, Société de Biologie, 1895, xlvii, pp. 273, 498.
3 Oliver and Schäfer: Journal of Physiology, 1895, xviii, p. 263. See also Radwánska, Anzeiger der Akademie, Krakau, 1910, pp. 728–736. Reviewed in Zentralblatt für Biochemie und Biophysik, 1911, xi, p. 467.
4 Dessy and Grandis: Archives Italiennes de Biologie, 1904, xli, p. 231.
5 Panella: Archives Italiennes de Biologie, 1907, xlviii, p. 462.
6 Cannon and Nice: American Journal of Physiology, 1913, xxxii, p. 44.
7 Bowditch and Warren: Journal of Physiology, 1886, vii, p. 438.
8 Elliott: Journal of Physiology, 1912, xliv, p. 403.
9 Cannon and Lyman: American Journal of Physiology, 1913, xxxi, p. 376.
10 Young and Lehmann: Journal of Physiology, 1908, xxxvii, p. liv.
[p. 95]
THE EFFECTS ON CONTRACTION OF FATIGUED MUSCLE OF VARYING THE ARTERIAL BLOOD PRESSURE
That great excitement is accompanied by sympathetic innervations which increase the contraction of the small arteries, render unusually forcible the heart beat, and consequently raise arterial pressure, has already been pointed out (see p. 26). Indeed, the counsel to avoid circumstances likely to lead to such excitement, which is given to persons with hardened arteries or with weak hearts, is based on the liability of serious consequences, either in the heart or in the vessels, that might arise from an emotional increase of pressure in these pathological conditions. That great muscular effort also is accompanied by heightened arterial pressure is equally well known, and is avoided by persons likely to be injured by it. Both in excitement and in strong exertion the blood is forced in large degree from the capacious vessels of the abdomen into other parts of the body. In excitement[p. 96] the abdominal arteries and veins are contracted by impulses from the splanchnic nerves. In violent effort the diaphragm and the muscles of the belly wall are voluntarily and antagonistically contracted in order to stiffen the trunk as a support for the arms; and the increased abdominal pressure which results forces blood out of that region and does not permit reaccumulation. The general arterial pressure in man, as McCurdy[1] has shown, may suddenly rise during extreme physical effort, from approximately 110 millimeters to 180 millimeters of mercury.
What effect the increase of arterial pressure, resulting from excitement or physical strain, may have on muscular efficiency, has received only slight consideration. Nice and I found there was need of careful study of the relations between arterial pressure and muscular ability, and, in 1913, one of my students, C. M. Gruber, undertook to make clearer these relations.
The methods of anesthesia and stimulation used by Gruber were similar to those described in the last chapter. The arterial blood pressure was registered from the right carotid or the femoral artery by means of a mercury manometer. A time marker indicating half-minute intervals was placed at the atmospheric pressure level of the[p. 97] manometer. And since the blood-pressure style, the writing point of the muscle lever, and the time signal were all set in a vertical line on the surface of the recording drum, at any given muscular contraction the height of blood pressure was simultaneously registered.
To increase general arterial pressure two methods were used: the spinal cord was stimulated in the cervical region through platinum electrodes, or the left splanchnic nerves were stimulated after the left adrenal gland had been excluded from the circulation. This was done in order to avoid any influence which adrenal secretion might exert. It is assumed in these experiments that vessels supplying active muscles would be actively dilated, as Kaufmann[2] has shown, and would, therefore, in case of a general increase of blood pressure, deliver a larger volume of blood to the area they supply. The effects of increased arterial pressure are illustrated in Figs. 13, 14 and 15. In the experiment represented in Fig. 13, the rise of blood pressure was produced by stimulation of the cervical cord, and in Figs. 14 and 15 by stimulation of the left splanchnic nerves after the left adrenal gland had been tied off.
The original blood pressure in Fig. 13 was 120 millimeters of mercury. This was increased by 62 millimeters, with a rise of only 8.4 per cent in the height of contraction of the fatigued muscle.
[p. 98]
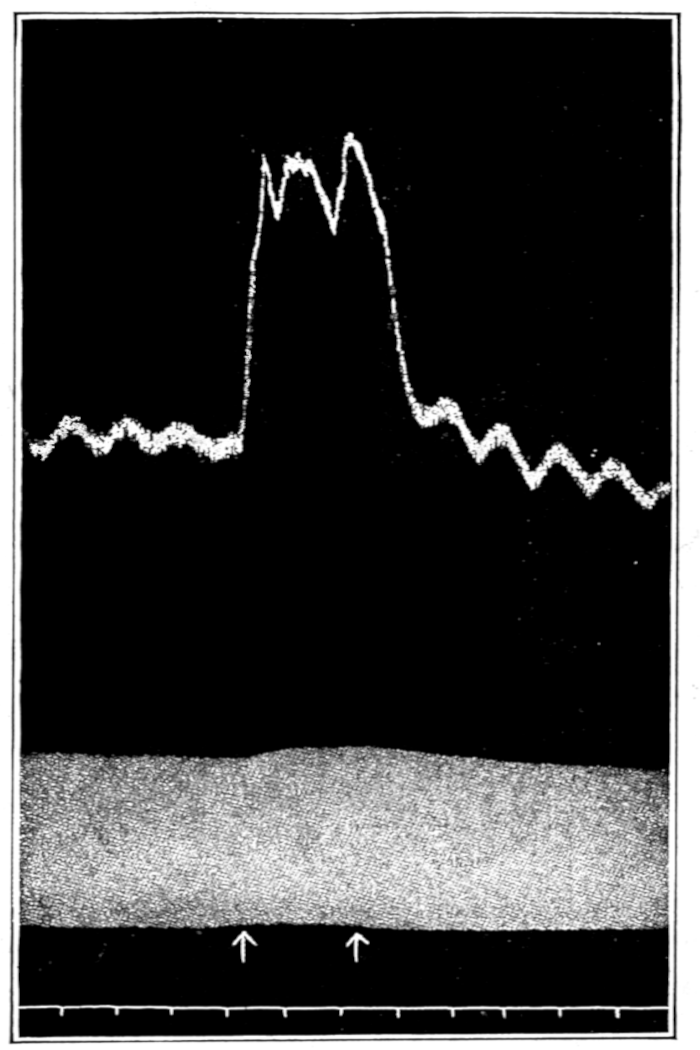
In Fig. 14 the original blood pressure was 100 millimeters of mercury. By increasing this pressure[p. 99] 32 millimeters there resulted simultaneous betterment of 9.8 per cent in the height of muscular contraction. In Fig. 14 B the arterial pressure was raised 26 millimeters and the height of contraction increased correspondingly 7 per cent. In Fig. 14 C no appreciable betterment can be seen although the blood pressure rose 18 millimeters.
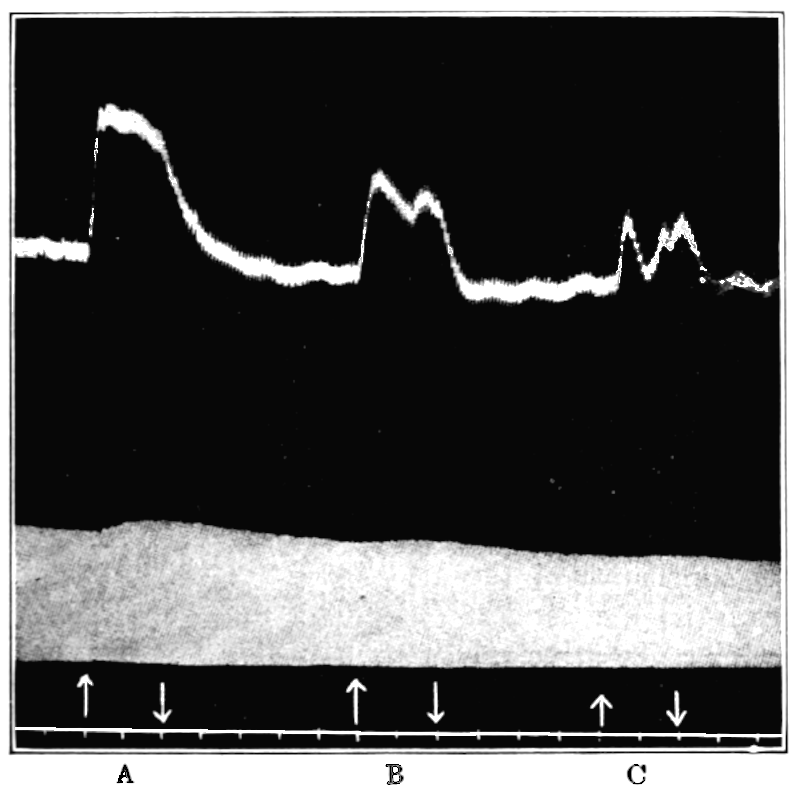
In Fig. 15 the original blood pressure was low—68 millimeters of mercury. This was increased in Fig. 15 A by 18 millimeters (the same as in[p. 100] Fig. 14 C without effect), and there resulted an increase of 20 per cent in the height of contraction. In Fig. 15 B the pressure was raised 24 millimeters with a corresponding increase of 90 per cent in the muscular contraction; and in Fig. 15 C 30 millimeters with a betterment of 125 per cent.
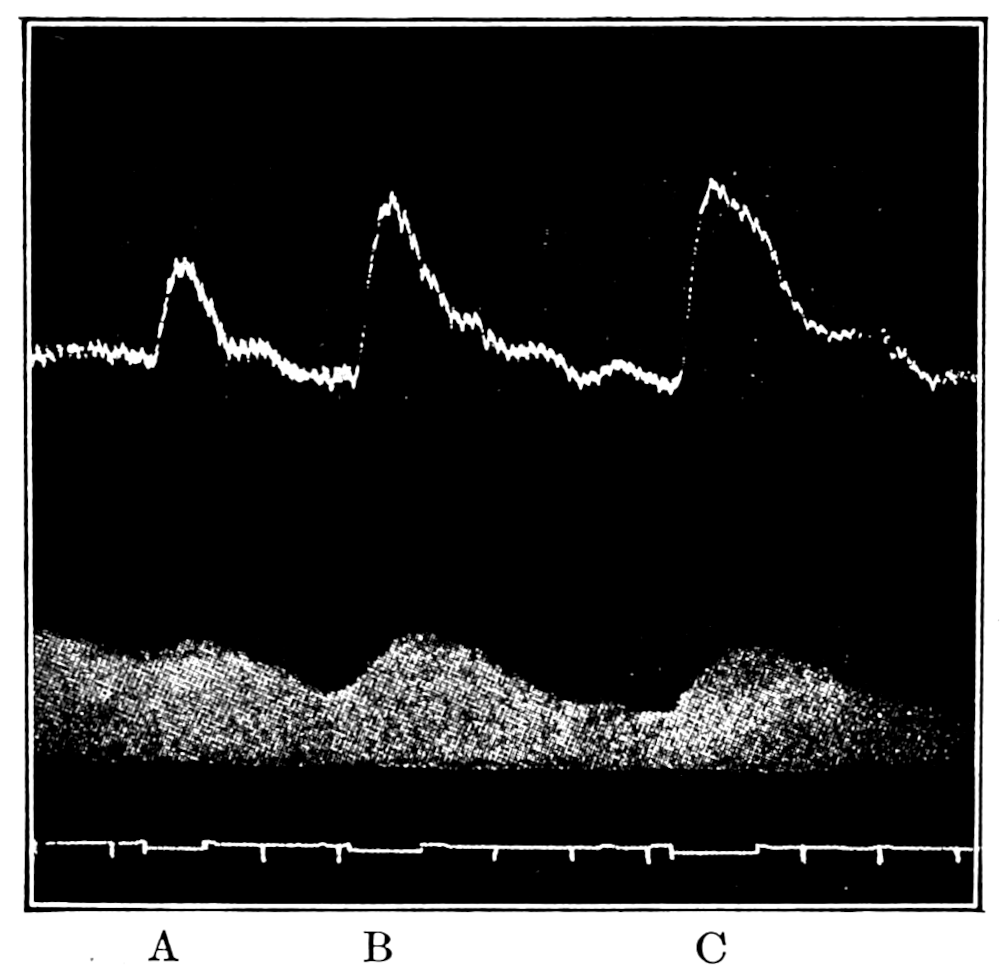
Comparison of Figs. 13, 14 and 15 reveals that the improvement of contraction of fatigued muscle is much greater when the blood pressure is raised, even slightly, from a low level, than when it is raised, perhaps to a very marked degree, from a high level. In one of the experiments performed by Nice and myself the arterial pressure[p. 101] was increased by splanchnic stimulation from the low level of 48 millimeters of mercury to 110 millimeters, and the height of the muscular contractions was increased about sixfold (see Fig. 16).
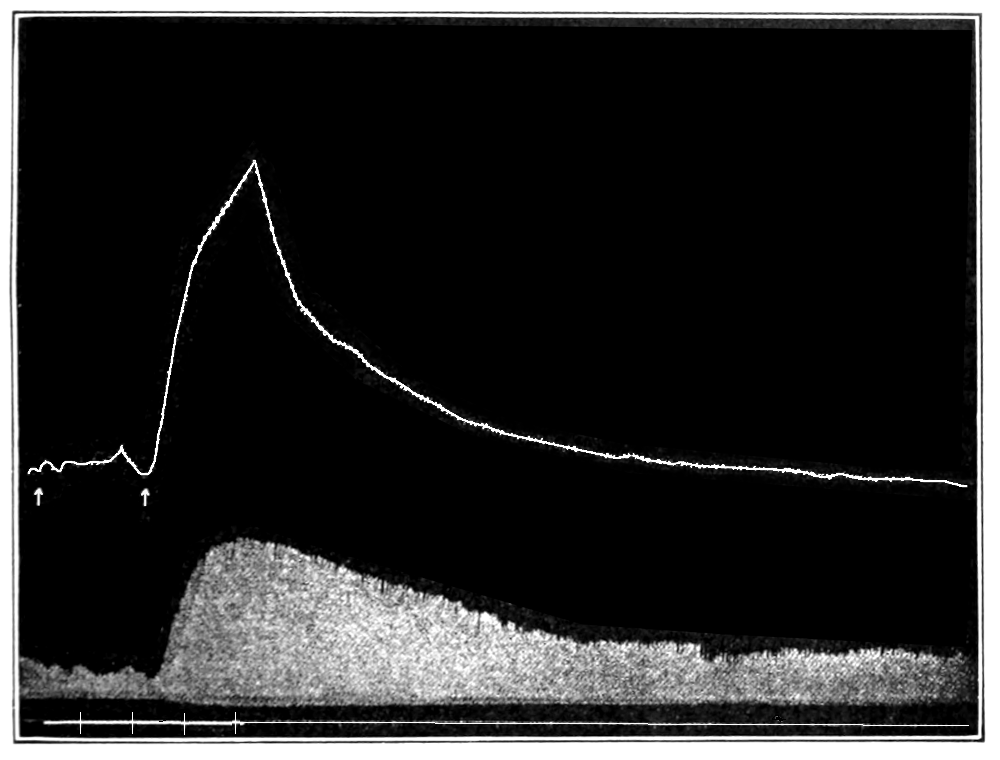
Results confirming those described above were obtained by Gruber in a study of the effects of splanchnic stimulation on the irritability of muscle when fatigued. In a series of eleven observations the average value of the barely effective stimulus (the “threshold” stimulus) had to be increased as the condition of fatigue developed. It[p. 102] was increased for the nerve-muscle by 25 per cent and for the muscle by 75 per cent. The left splanchnic nerves, disconnected from the left adrenal gland, were now stimulated. The arterial pressure, which had varied between 90 and 100 millimeters of mercury, was raised at least 40 millimeters. As a result of splanchnic stimulation there was an average recovery of 42 per cent in the nerve-muscle and of 46 per cent in the muscle. The increased general blood pressure was effective, therefore, quite apart from any possible action of adrenal secretion, in largely restoring to the fatigued structures their normal irritability.
Inasmuch as an increase in arterial pressure produces an increase in the height of contraction of fatigued muscle, it is readily supposable that a decrease in the pressure would have the opposite effect. Such is the case only when the blood pressure falls below the region of 90 to 100 millimeters of mercury. Thus if the arterial pressure stands at 150 millimeters of mercury, it has to fall approximately 55 to 65 millimeters before causing a decrease in the height of contraction. Fig. 17 is the record of an experiment in which the blood pressure was lowered by lessening the output of blood from the heart by compressing the thorax. The record shows that when the pressure[p. 103] was lowered from 120 to 100 millimeters of mercury (A), there was no appreciable decrease in the height of contraction; when lowered to 90 millimeters (B), there resulted a decrease of 2.4 per cent; when to 80 millimeters of mercury (C), a decrease of 7 per cent; and when to 70 millimeters (D), a decrease of 17.3 per cent. Results similar to those represented in Fig. 17 were obtained by pulling on a string looped about the[p. 104] aorta just above its iliac branches, thus lessening the flow to the hind limbs.
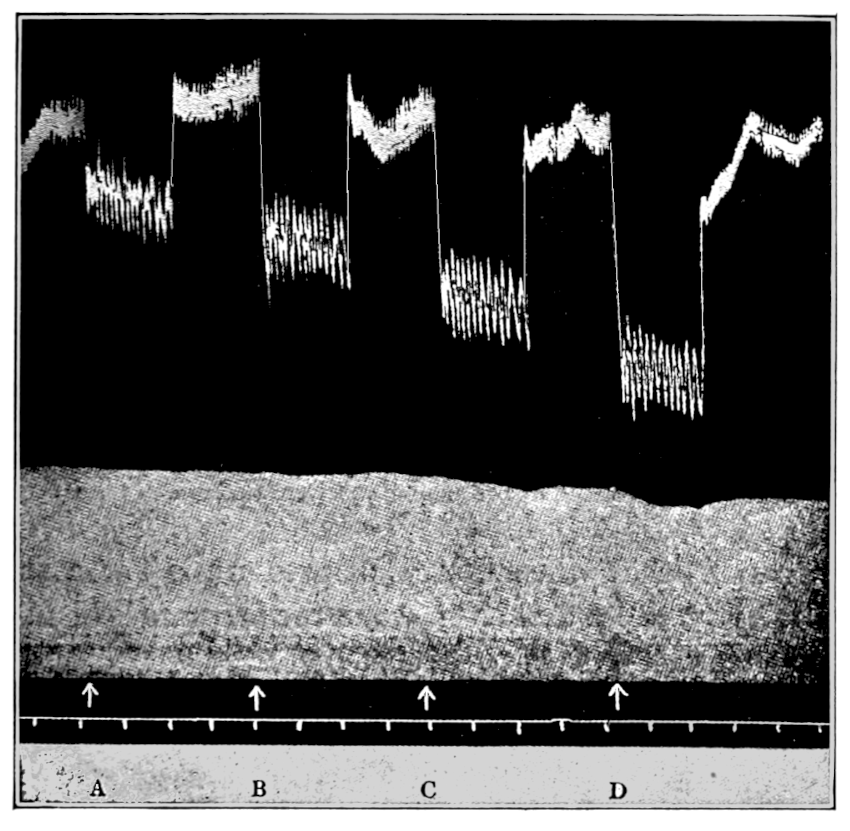
The region of 90 to 100 millimeters of mercury may therefore be regarded as the critical region at which a falling blood pressure begins to be accompanied by a concurrent lessening of the efficiency of muscular contraction, when the muscle is kept in continued activity. It is at that region that the blood flow is dangerously near to being inadequate.
How are these effects of increasing and decreasing the arterial blood pressure most reasonably explained? There is abundant evidence that fatigue products accumulate in a muscle which is doing work, and also that these metabolites interfere with efficient contraction. As Ranke[3] long ago demonstrated, if a muscle, deprived of circulating blood, is fatigued to a standstill, and then the circulation is restored, the muscle again responds for a short time to stimulation, because the waste has been neutralized or swept away by the fresh blood. When the blood pressure is at its normal height for warm-blooded animals (about 120 millimeters of mercury, see Fig. 13), the flow appears to be adequate to wash out the depressive metabolites, at least in the single muscle[p. 105] used in these experiments, because a large rise of pressure produces but little change in the fatigue level. On the other hand, when the pressure is abnormally low, the flow is inadequate, and the waste products are permitted to accumulate and clog the action of the muscle. Under such circumstances a rise of pressure has a very striking beneficial effect.
It is noteworthy that the best results of adrenin on fatigued muscle reported by previous observers were obtained from studies on cold-blooded animals. In these animals the circulation is maintained normally by an arterial pressure about one-third that of warm-blooded animals. Injection of adrenin in an amount which would not shut off the blood supply would, by greatly raising the arterial pressure, markedly increase the circulation of blood in the active muscle. In short, the conditions in cold-blooded animals are quite like those in the pithed mammal with an arterial pressure of about 50 millimeters of mercury (see Fig. 16). Under these conditions the improved circulation causes a remarkable recovery from fatigue. That notable results of adrenin on fatigue are observed in warm-blooded animals only when they are deeply anesthetized or are deprived of the medulla was claimed by Panella.[4] He apparently believed that in normal mammalian conditions adrenin has little effect because quickly destroyed, whereas in[p. 106] the cold-blooded animals, and in mammals whose respiratory, circulatory, and thermogenic states are made similar to the cold-blooded by anesthesia or pithing, the contrary is true. In accordance with our observations of the effects of blood pressure on fatigued muscle, we would explain Panella’s results not as he has done but as due to two factors. First, the efficiency of the muscle, when blood pressure is low, follows the ups and downs of pressure much more directly than when the pressure is high. And second, a given dose of adrenin always raises a low blood pressure in atonic vessels. The improvement of circulation is capable of explaining, therefore, the main results obtained in cold-blooded animals and in pithed mammals.
Oliver and Schäfer reported unusually effective contractions in muscles removed from the body after adrenal extract had been injected. As shown in Fig. 16, however, the fact that the circulation had been improved results in continued greater efficiency of the contracting muscle. Oliver and Schäfer’s observation may reasonably be accounted for on this basis.
As stated in a previous paragraph, there is evidence that the vessels supplying a muscle dilate[p. 107] when the muscle becomes active. And although the normal blood pressure (about 120 millimeters of mercury) may be able to keep adequately supplied with blood the single muscle used in our investigation, a higher pressure might be required when more muscles are involved in activity, for a more widely spread dilation might then reduce the pressure to the point at which there would be insufficient circulation in active organs. Furthermore, with many muscles active, the amount of waste would be greatly augmented, and the need for abundant blood supply would thereby to a like degree be increased. For both reasons a rise of general arterial pressure would prove advantageous. The high pressure developed in excitement and pain, therefore, might be specially serviceable in the muscular activities which are likely to accompany excitement and pain.
In connection with the foregoing considerations, the action of adrenin on the distribution of blood in the body is highly interesting. By measuring alterations in the volume of various viscera and the limbs, Oliver and Schäfer[5] proved that the viscera of the splanchnic area—e. g., the spleen, the kidneys, and the intestines—suffer a considerable decrease of volume when adrenin is administered, whereas the limbs into which the blood is forced from the splanchnic region actually increase in size. The action of adrenin indicates the[p. 108] relative degrees of sympathetic innervations. In other words, at times of pain and excitement sympathetic discharges, probably aided by the adrenal secretion simultaneously liberated, will drive the blood out of the vegetative organs of the interior, which serve the routine needs of the body, into the skeletal muscles which have to meet by extra action the urgent demands of struggle or escape.
But there are exceptions to the general statement that by adrenin the viscera are emptied of their blood. It is well known that adrenin has a vasodilator, not a vasoconstrictor, action on the arteries of the heart; it is well known also that adrenin affects the vessels of the brain and the lungs only slightly if at all. From this evidence we may infer that sympathetic impulses, though causing constriction of the arteries of the abdominal viscera, have no effective influence on those of the pulmonary and intracranial areas and actually increase the blood supply to the heart. Thus the absolutely and immediately essential organs—those the ancients called the “tripod of life”—the heart, the lungs, the brain (as well as its instruments, the skeletal muscles)—are in times of excitement abundantly supplied with blood taken from organs of less importance in critical moments. This shifting of the blood so that there is an assured adequate supply to structures essential for the preservation of the individual may reasonably[p. 109] be interpreted as a fact of prime biological significance. It will be placed in its proper setting when the other evidence of bodily changes in pain and excitement have been presented.
1 McCurdy: American Journal of Physiology, 1901, v, p. 98.
2 Kaufmann: Archives de Physiologie, 1892, xxiv, p. 283.
3 Ranke: Archiv für Anatomie, 1863, p. 446.
4 Panella: Archives Italiennes de Biologie, 1907, xlviii, p. 462.
5 Oliver and Schäfer: Journal of Physiology, 1895, xviii, p. 240.
[p. 110]
THE SPECIFIC RÔLE OF ADRENIN IN COUNTERACTING THE EFFECTS OF FATIGUE
As a muscle approaches its fatigue level, its contractions are decreased in height. Higher contractions will again be elicited if the stimulus is increased. Although these phenomena are well known, no adequate analysis of their causes has been advanced. A number of factors are probably operative in decreasing the height of contraction: (1) The using up of available energy-producing material; (2) the accumulation of metabolites in the fatigued muscle; (3) polarization of the nerve at the point of repeated electrical stimulation; and (4) a decrease of irritability. It may be that there are interactions between these factors within the muscle, e. g., the second may cause the fourth.
The last of the factors mentioned above—the effect of fatigue on the irritability of the nerve-muscle combination, or on the muscle alone—can[p. 111] be tested by determining variations in the least stimulus capable of causing the slightest contraction, the so-called “threshold stimulus.” As the irritability lessens, the threshold stimulus must necessarily be higher. The height of the threshold is therefore a measure of irritability. How does fatigue affect the irritability of nerve-muscle and muscle? How is the irritability of fatigued structures affected by rest? How is it influenced by adrenin or by adrenal secretion? Answers to these questions were sought in researches carried on by C. M. Gruber[1] in 1913.
The neuro-muscular arrangements used in these researches were in many respects similar to those already described in the account of experiments by Nice and myself. To avoid the influence of an anesthetic some of the animals were decerebrated under ether and then used as in the experiments in which urethane was the anesthetic. The nerve (the peroneus communis) supplying the tibialis anticus muscle was bared and severed; and near the cut end shielded platinum electrodes were applied. These electrodes were used in fatiguing the muscle. Between these electrodes and the muscle other platinum electrodes could be quickly applied to determine the threshold stimulus and the tissue resistance. These second electrodes were removed[p. 112] except when in use, and when replaced were set always in the same position. Care was taken, before replacing them, to wipe off moisture on the nerve or on the platinum points.
For determining the threshold stimulus of the muscle the skin and other overlying tissues were cut away from the tibialis anticus in two places about 5 centimeters apart. Through these openings platinum needle electrodes could be thrust into the muscle whenever readings were to be taken. Local polarization was avoided by reinserting the needles into fresh points on the exposed areas whenever new readings were to be taken.
The tendon of the tibialis anticus was attached, as in the previous experiments, by a strong thread passing about pulleys to a lever which when lifted stretched a spring. During the determination of the threshold the spring was detached from the lever, so that only the pull of the lever itself (about 15 grams) was exerted on the muscle.
The method of measuring the stimulating value of the electric current which was used in testing the threshold was that devised by E. G. Martin[*] of the Harvard Laboratory—a method by which the strength of an induced electric shock is calculable in definite units. If the tissue resistance enters[p. 113] into the calculation these are called β units. When the threshold of the nerve-muscle was taken, the apparatus for the determination was connected with the nerve through the electrodes nearer the muscle. They were separated from the fatiguing electrodes by more than 3 centimeters, and arranged so that the kathode was next the muscle. When the threshold of the muscle was taken directly the apparatus was connected with the muscle through platinum needle electrodes thrust into it. The position of the secondary coil of the inductorium, in every case, was read by moving it away from the primary coil until the very smallest possible contraction of the muscle was obtained. Four of these readings were made, one with tissue resistance, the others with 10,000, 20,000, and 30,000 ohms additional resistance in the secondary circuit. Only break shocks were employed—the make shocks were short-circuited. Immediately after the determination of the position of the secondary coil, and before the electrodes were removed or disconnected, three readings of the tissue resistance were made. From these data four values for β were calculated.
* For a full account of Dr. Martin’s method of calculating the strength of electric stimuli, see Martin: The Measurement of Induction Shocks, New York, 1912.
The strength of the primary current for determining the threshold of the nerve-muscle was usually .01 ampere, but in a few cases .05 ampere was used. For normal muscle it was .05 ampere and for denervated muscle 1.0 ampere. The inductorium,[p. 114] which was used throughout, had a secondary resistance of 1400 ohms. This was added to the average tissue resistance in making corrections—corrections were made also for core magnetization.
The threshold for the peroneus communis nerve in decerebrate animals varied from 0.319 to 2.96 units, with an average in sixteen experiments of 1.179.[*] This average is the same as that found by E. L. Porter[2] for the radial nerve in the spinal cat. For animals under urethane anesthesia a higher average was obtained. In these it varied from .644 to 7.05, or an average in ten experiments of 3.081.
* For the detailed data of these and other quantitative experiments, the reader should consult the tables in the original papers.
The threshold for the tibialis anticus muscle varied in the decerebrate animals from 6.75 units to 33.07, or an average in fifteen experiments of 18.8. Ten experiments were performed under urethane anesthesia and the threshold varied from 12.53 to 54.9, with an average of 29.84 β units. From these results it is evident that anesthesia notably affects the threshold.
E. L. Porter proved, by experiments carried on in the Harvard Physiological Laboratory, that the threshold of an undisturbed nerve-muscle remains[p. 115] constant for hours, and his observation was confirmed by Gruber (see Fig. 19). If, therefore, after fatigue, a change exists in the threshold, this change is necessarily the result of alterations set up by the fatigue process in the nerve-muscle or muscle.
After fatigue the threshold of the nerve-muscle, in sixteen decerebrate animals, increased from an average of 1.179 to 3.34—an increase of 183 per cent. In ten animals under urethane anesthesia the threshold after fatigue increased from a normal average of 3.08 to 9.408—an increase of 208 per cent.
An equal increase in the threshold stimulus was obtained from the normal muscle directly. In decerebrate animals the normal threshold of 18.8 units was increased by fatigue to 69.54, or an increase of 274 per cent. With urethane anesthesia the threshold increased from 29.849 to 66.238, or an increase of 122 per cent.
Fig. 18, plotted from the data of one of the many experiments, shows the relative heights of the threshold before and after fatigue. The correspondence of the two readings of the threshold, one from the nerve supplying the muscle and the other from the muscle directly, served as a check on the electrodes. The broken line in the figure represents the threshold (in units) of the nerve-muscle, and the continuous line that of the muscle. The[p. 116] threshold values of the nerve-muscle have been magnified ten times in order to bring the two records close together. In this experiment the threshold of the muscle after fatigue (i. e., at 2) is 167 per cent higher than the normal threshold (at 1), while that of the nerve-muscle after fatigue is 30.5 per cent higher than its normal.
Figure 18.—A record plotted from the data of one experiment. The time intervals in minutes are registered on the abscissa; the value of the threshold in units is registered on the ordinate. The continuous line is the record of the muscle, the broken line that of the nerve-muscle. The values for the nerve-muscle have been magnified ten times, those for the muscle are normal.
(1) Normal values of the threshold.
(2) Fatigue thresholds after one hour’s work, lifting 120 grams 240 times a minute.
(3 and 4) The threshold after rest.
Evidently a direct relation exists between the duration of work and the increase of threshold. For instance, the threshold is higher after a muscle is fatigued for two hours than it is at the end of[p. 117] the first hour. The relation between the work done and the threshold is not so clear. In some animals the thresholds were higher after 120 grams had been lifted 120 times a minute for 30 minutes than they were in others in which 200 grams had been lifted 240 times a minute for the same period. The muscle in the latter instances did almost four times as much work, yet the threshold was lower. The difference may be due to the general condition of the animal.
A few experiments were performed on animals in which the nerve supplying the muscle was cut seven to fourteen days previous to the experiment. The muscle, therefore, had within it no living nerve fibres. The average normal threshold for the denervated muscle in 6 animals was 61.28 units. As in the normal muscle, the percentage increase due to fatigue was large.
That rest decreases the fatigue threshold of both nerve-muscle and muscle can be seen in Fig. 18. The time taken for total recovery, however, is dependent upon the amount of work done, but this change, like that of fatigue, varies widely with different individuals. In some animals the threshold returned to normal in 15 minutes; in others, in which the same amount of work was done, it was[p. 118] still above normal even after 2 hours of rest. This may be due to the condition of the animals—in some the metabolites are probably eliminated more rapidly than in others. There were also variations in the rate of restoration of the normal threshold when tested on the nerve and when tested on the muscle in the same animal. In Fig. 18 (at 3) the nerve-muscle returned to normal in 30 minutes, whereas the muscle (at 4) after an hour’s rest had not returned to normal by a few β units. This, however, is not typical of all nerve-muscles and muscles. The opposite condition—that in which the muscle returned to normal before the nerve-muscle—occurred in as many cases as did the condition just cited. The failure of the two tissues to alter uniformly in the same direction may be explained as due to variations in the location of the electrodes when thrust into the muscle at different times (e. g., whether near nerve filaments or not). The results from observations made on the nerve are more likely to be uniform and reliable than are those from the muscle.
The time required for the restoration of the threshold from fatigue to normal, in denervated muscles, is approximately the same as that for the normal muscle.
[p. 119]
The foregoing observations showed that fatigue raises the normal threshold of a muscle, on the average, between 100 and 200 per cent (it may be increased more than 600 per cent); that this increase is dependent on the time the muscle works, but also varies with the animal; that rest, 15 minutes to 2 hours, restores the normal irritability; and that this recovery of the threshold depends upon the time given to rest, the duration of the work, and also upon the condition of the animal. The problem which was next attacked by Gruber was that of learning whether the higher contractions of fatigued muscle after splanchnic stimulation could be attributed to any influence which adrenal secretion might have in restoring the normal irritability. To gain insight into the probabilities he tried first the effects of injecting slowly into the jugular vein physiological amounts of adrenin.[*]
* The form of adrenin used in these and in other injections was fresh adrenalin made by Parke, Davis & Co.
The normal threshold of the peroneus communis nerve varied in the animals used in this series of observations from 0.35 to 5.45 units, with an average in nine experiments of 1.3, a figure close to the 1.179 found in the earlier series on the effect of fatigue. For the tibialis anticus muscle, in which the nerve-endings were intact, the threshold varied[p. 120] from 6.75 to 49.3 units, with an average in the nine experiments of 22.2. This is slightly higher than that cited for this same muscle in the earlier series. By fatigue the threshold of the nerve-muscle was increased from an average of 1.3 to an average of 3.3 units, an increase of 154 per cent. The muscle increased from an average of 22.2 to an average of 59.6, an increase of 169 per cent. After an injection of 0.1 to 0.5 cubic centimeters of adrenin (1:100,000) the fatigue threshold was decreased within five minutes in the nerve-muscle from an average of 3.3 to 1.8, a recovery of 75 per cent, and in the muscle from an average of 59.6 to 42.4, a recovery of 46 per cent. To prove that this effect of adrenin is a counteraction of the effects of fatigue, Gruber determined the threshold for muscle and nerve-muscle in non-fatigued animals before and after adrenin injection. He found that in these cases no lowering of threshold occurred, a result in marked contrast with the pronounced and prompt lowering induced by this agent in muscles when fatigued.
Figs. 19 and 20, plotted from the data of two of the experiments, show the relative heights of the threshold before and after an injection of adrenin. The close correspondence of the two readings of the threshold, one from the nerve supplying the muscle, the other from the muscle directly, served to show that there was no fault in the electrodes.[p. 121] The continuous line in the Figures represents the threshold (in units) of the muscle, the broken line that of the nerve-muscle. The threshold of the nerve-muscle is magnified 100 times in Fig. 19 and 10 times in Fig. 20. In Fig. 19 (at 2 and 4) the threshold was taken after an intravenous injection of 0.1 and 0.2 cubic centimeter of adrenin respectively.
These examples show that adrenin does not affect the threshold of the normal non-fatigued muscle when tested either on the muscle directly or on the nerve-muscle. In Fig. 19 (at 3) the observation taken after two hours of rest illustrates the constancy of the threshold under these circumstances.
In Fig. 19 the normal threshold was increased by fatigue (at 5)—the muscle had been pulling 120 times a minute for one hour on a spring having an initial tension of 120 grams—from 30.0 to 51.6 units, an increase of 72 per cent; and in the nerve-muscle from 0.62 to 0.89 units, an increase of 46 per cent. The threshold (at 6) was taken five minutes after injecting 0.1 cubic centimeter of adrenin (1:100,000). The threshold of the muscle was lowered from 51.6 to 38.0 units, a recovery of 62 per cent; that of the nerve-muscle from 0.89 to 0.79 units, a recovery of 37 per cent. After another injection of 0.5 cubic centimeter of adrenin the thresholds (at 7) were[p. 122] taken; that of the nerve-muscle dropped to normal—0.59 units—a recovery of 100 per cent, and that of the muscle remained unaltered—26 per cent above its normal threshold.
Figure 19.—A record plotted from the data of one experiment. The time intervals in hours and minutes are represented on the abscissa; the values of the threshold in β units are represented on the ordinate. The continuous line is the record of the muscle, the broken line that of the nerve-muscle. The nerve-muscle record is magnified 100 times; that of the muscle is normal.
(1) Normal threshold stimulus. (2) Threshold five minutes after an intravenous injection of 0.1 cubic centimeter of adrenin (1:100,000) without previous fatigue. (3) Threshold after a rest of two hours. (4) Threshold five minutes after an injection of 0.2 cubic centimeter of adrenin (1:100,000) without previous fatigue. (5) Threshold after one hour’s fatigue. The muscle contracted 120 times per minute against a spring having an initial tension of 120 grams. (6) Threshold five minutes after an injection (0.1 cubic centimeter) of adrenin (1:100,000). (7) Threshold five minutes after another injection of adrenin (0.5 cubic centimeter of a 1:100,000 solution).
In Fig. 20 the threshold (at 5) was taken five[p. 123] minutes after an injection of 0.1 cubic centimeter of adrenin. The drop here was as large as that shown in Fig. 19. The threshold taken from the muscle directly was lowered from 30.6 to 18 units, a recovery of 61 per cent; the nerve-muscle from 1.08 to 0.87 units, a recovery of 51 per cent. That this sudden decrease cannot be due to rest is shown in the same Figure (at 3 and 4). These readings were made after 60 and 90 minutes’ rest respectively. The sharp decline in the record (at 5) indicates distinctly the remarkable restorative influence[p. 124] of adrenin in promptly lowering the high fatigue threshold of neuro-muscular irritability.
Figure 20.—A record plotted from the data of one experiment. The time intervals in hours and minutes are registered on the abscissa; the values of the threshold in units are registered on the ordinate. The continuous line is the record of the muscle, the broken line that of the nerve-muscle. The record of the nerve-muscle is magnified ten times; that of the muscle is normal.
(1) Normal threshold. (2) The threshold after one hour’s fatigue. The muscle contracted 120 times per minute against a spring having an initial tension of 120 grams. (3 and 4) Thresholds after rest; after 60 minutes (3), and after 90 minutes (4). (5) Threshold five minutes after an injection of adrenin (0.1 cubic centimeter of a 1:100,000 solution). (6 and 7) Thresholds after rest; after 60 minutes (6), and after 90 minutes (7).
As stated in describing the effects of arterial blood pressure, an increase of pressure is capable of causing a decided lowering of the neuro-muscular threshold after fatigue. Is it not possible that adrenin produces its beneficial effects by bettering the circulation?
Nice and I had argued that the higher contractions of fatigued muscle, that follow stimulation or injection of adrenin, could not be wholly due to improved blood flow through the muscle, for when by traction on the aorta or compression of the thorax arterial pressure in the hind legs was prevented from rising, splanchnic stimulation still caused a distinct improvement, the initial appearance of which coincided with the point in the blood-pressure curve at which evidence of adrenal secretion appeared. And, furthermore, the improvement was seen also when adrenin was given intravenously in such weak solution (1:100,000) as to produce a fall instead of a rise of arterial pressure. Lyman and I had shown that this fall of pressure was due to a dilator effect of adrenin. Since the blood vessels of the fatigued muscle were dilated by severance of their nerves when the nerve trunk was[p. 125] cut, and, besides, as previously stated (see p. 86), were being stimulated through their nerves at a rate favorable to relaxation, it seemed hardly probable that adrenin could produce its beneficial effect by further dilation of the vessels and by consequent flushing of the muscle with an extra supply of blood.[3] The lowering of blood pressure had[p. 126] been proved to have no other effect than to impair the action of the muscle (see p. 103). Although the chances were thus against an interpretation of the beneficial influence of adrenin through action on the circulation, it was thought desirable to test the possibility by comparing its effect with that of another vasodilator—amyl nitrite.
Figs. 21 and 22 are curves obtained from the left tibialis anticus muscle. The rate of stimulation was 240 times a minute.
The muscle in Fig. 21 contracted against a spring having an initial tension of 120 grams, and that in Fig. 22 against an initial tension of 100 grams. In Fig. 21, at the point indicated on the base line, 0.4 cubic centimeter of adrenin (1:100,000) was injected into the left external jugular vein. There resulted a fall of 25 millimeters of mercury in the arterial pressure and a concurrent betterment of 15 per cent in the height of contraction, requiring two minutes and fifteen seconds of fatigue (about 540 contractions) before it returned to the former level. In Fig. 22, at the point indicated by the arrow, a solution of amyl nitrite was injected into the right external jugular vein. There resulted a fall of 70 millimeters of mercury in arterial pressure and a betterment of 4.1 per cent in the height of muscular contraction, requiring fifteen seconds of fatigue (about 60 contractions) to decrease the height of contraction to its former level. In[p. 128] neither case did the blood pressure fall below the critical region (see p. 104).[*]
* In some cases after injection of amyl nitrite the normal blood pressure, which was high, dropped sharply to a point below the critical region. There resulted a primary increase in muscular contraction due to the betterment in circulation caused by the dilation of the vessels before the critical region was reached. During the time that the pressure was below the critical region the muscle contraction fell. As the blood pressure again rose to normal the muscle contraction increased coincidently.
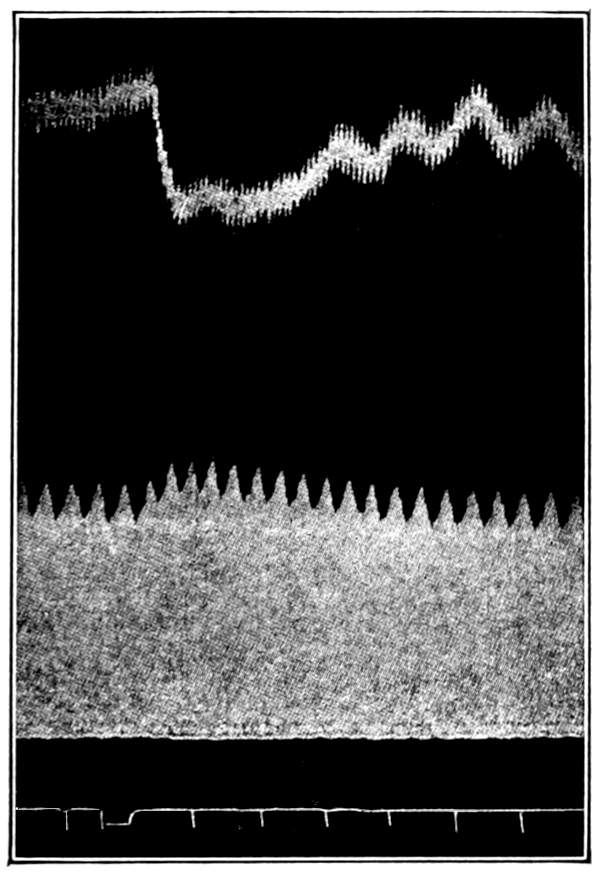
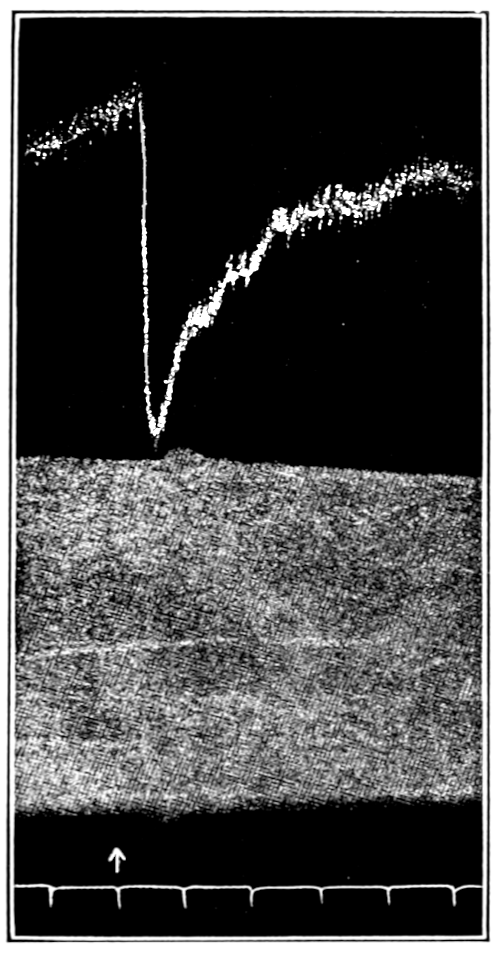
Although the fall in arterial pressure caused by dilation of the vessels due to amyl nitrite was almost three times as great as that produced by the adrenin, yet the resultant betterment was only about one-fourth the percentage height and lasted but one-ninth the time. In all cases in which these solutions caused an equal fall in arterial pressure, adrenin caused higher contractions, whereas amyl nitrite caused no appreciable change.
From the evidence presented in the foregoing pages it is clear that adrenin somehow is able to bring about a rapid recovery of normal irritability of muscle after the irritability has been much lessened by fatigue, and that the higher contractions of a fatigued muscle after an injection of adrenin are due, certainly in part, to some specific action of this substance and not wholly to its influence on the circulation. Some of the earlier investigators[p. 129] of adrenal function, notably Albanese,[4] and also Abelous and Langlois,[5] inferred from experiments on the removal of the glands that the rôle they played in the bodily economy was that of neutralizing, destroying or transforming toxic substances produced in the organism as a result of muscular or nervous work. It seemed possible that the metabolites might have a checking or blocking influence at the junction of the nerve fibres with the muscle fibres, and might thus, like curare, lessen the efficiency of the nerve impulses. Radwánska’s observation[6] that the beneficial action of adrenin is far greater when the muscle is stimulated through its nerve than when stimulated directly, and Panella’s discovery[7] that adrenin antagonizes the effect of curare, were favorable to the view that adrenin improves the contraction of fatigued muscle by lessening or removing a block established by accumulated metabolites.
The high threshold of fatigued denervated muscle, however, Gruber found was quite as promptly lowered by adrenin as was that of normal muscles stimulated through their nerves. Fig. 23 shows that the height of contraction, also, of the fatigued muscle is increased when adrenin is administered. In this experiment the left tibialis anticus muscle was stimulated directly by thrusting platinum needle electrodes into it. The peroneus communis nerve supplying the muscle had been cut and two[p. 130] centimeters of it removed nine days previous to the experiment. The rate of stimulation was 120 times per minute and the initial tension of the spring about 120 grams. At the point indicated by the arrow an injection of 0.1 cubic centimeter of adrenin (1:100,000) was made into a jugular vein. A fall in arterial pressure from 110 to 86 millimeters of mercury and a simultaneous betterment of 20 per cent in the height of contraction[p. 131] were obtained. It required four minutes of fatigue (about 480 contractions) to restore the muscle curve to its former level. Results similar to this were obtained from animals in which the nerve had been cut 7, 9, 12, 14, and 21 days. In all instances the nerve was inexcitable to strong faradic stimulation.
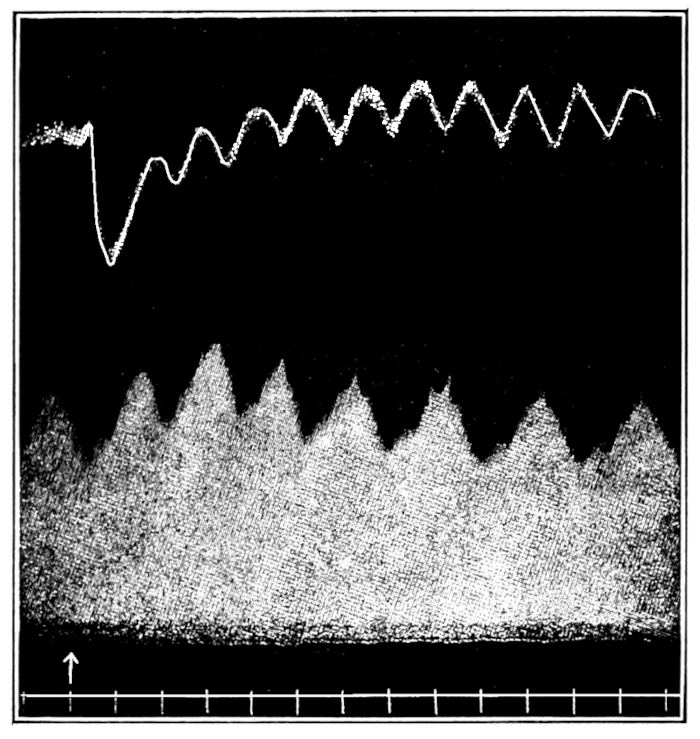
In Radwánska’s experiments, mentioned above, the muscle was stimulated directly when the nerve endings were intact. It seems reasonable to suppose, therefore, that in all cases he was stimulating nerve tissue. Since a muscle is more irritable when stimulated through its nerve than when stimulated directly (nerve and muscle), a slight change in the irritability of the muscle by adrenin would naturally result in a greater contraction when the nerve was stimulated. Panella’s results also are not inconsistent with the interpretation that the effect of adrenin is on the muscle substance rather than on the nerve endings. A method which has long been used to separate muscle from nerve is that of blocking the nervous impulses by the drug curare. Gruber found that when curare is injected the threshold of the normal muscle is increased as was to be expected from the removal of the highly efficient nervous stimulations. And also, as was to be expected on that basis, curare did not increase the threshold in a muscle in which the nerve endings had degenerated. Adrenin antagonizes[p. 132] curare with great promptness, decreasing the heightened threshold of a curarized muscle, in five minutes or less, in some cases to normal. From this observation it might be supposed that curare and fatigue had the same effect, and that adrenin had the single action of opposing that effect. But fatigue raises the threshold of a curarized muscle, and adrenin then antagonizes this fatigue. Langley[8] has argued that curare acts upon a hypothetical “receptive substance” in muscle. If so, probably curare acts upon a substance, or at a point, different from that upon which fatigue acts; for, as the foregoing evidence shows, fatigue increases the threshold of a muscle whether deprived of its nerve supply by nerve section and degeneration or by curare, whereas curare affects only the threshold of a muscle in which the nerve endings are normal.[9] And since adrenin can oppose the effects of both curare and fatigue, it may be said to have two actions, or to act on two different substances or at two different points in the muscle.
The evidence adduced in the last chapter indicated that the greater “head” of arterial pressure produced by the more rapid heart beat and the stronger contraction of many arterioles in times of great excitement would be highly serviceable to the organism in any extensive muscular activity which the excitement might involve. By assuring an abundant flow of blood through the enlarged vessels[p. 133] of the working muscle, the waste products resulting from the wear and tear in contraction would be promptly swept away and thus would be prevented from impairing the muscular efficiency. The adrenin discharge at such times would, as was pointed out, probably reënforce the effects of sympathetic impulses. The evidence presented in this chapter shows that adrenin has also another action, a very remarkable action, that of restoring to a muscle its original ability to respond to stimulation, after that has been largely lost by continued activity through a long period. What rest will do only after an hour or more, adrenin will do in five minutes or less. The bearing of this striking phenomenon on the functions of the organism in times of great need for muscular activity will be considered in a later discussion.
1 Gruber: American Journal of Physiology, 1913, xxxii, p. 437.
2 E. L. Porter: American Journal of Physiology, 1912, xxxi, p. 149.
3 Cannon and Nice: American Journal of Physiology, 1913, xxxii, p. 55.
4 Albanese: Archives Italiennes de Biologie, 1892, xvii, p. 239.
5 Abelous and Langlois: Archives de Physiologie, 1892, xxiv, pp. 269–278, 465–476.
6 Radwánska: Anzeiger der Akademie, Krakau, 1910, pp. 728–736. Reviewed in the Centralblatt für Biochemie und Biophysik, 1911, xi, p. 467.
[p. 134]
7 Panella: Archives Italiennes de Biologie, 1907, xlvii, p. 30.
8 Langley: Proceedings of the Royal Society of London, 1906, lxxviii, B, p. 181. Journal of Physiology, 1905–6, xxxiii, pp. 374–413.
9 See Gruber: American Journal of Physiology, 1914, xxxiv, p. 89.
[p. 135]
THE HASTENING OF COAGULATION OF BLOOD BY ADRENIN
The primary value of blood to the body must have been one of the earliest observations of reasoning beings. When we consider the variety of fundamental services which this circulating fluid performs—the conveyance of food and oxygen to all the tissues, the removal of waste, the delivery of the internal secretions, the protection of the body against toxins and bacterial invasion, and the distribution of heat from active to inactive regions—the view of the ancient Hebrews that the “life of the flesh is in the blood” is well justified. It is naturally of the utmost importance that this precious fluid shall be safeguarded against loss. And its property of turning to a jelly soon after escaping from its natural channels assures a closure of the opening through which the escape occurred, and thus protection of the body from further bleeding. The slight evidence that adrenin hastens the clotting process has already been hinted at. When we[p. 136] found that adrenin is set free in pain and intense emotion, it seemed possible that there might exist in the body an arrangement for making doubly sure the assurance against loss of blood, a process that might nicely play its rôle precisely when the greatest need for it would be likely to arise.
It was in 1903, while tracing in dogs the rise and fall of sugar in the blood after administering adrenin, that Vosburgh and Richards[1] first noted that simultaneously with the increase of blood sugar there occurred more rapid coagulation. In some cases the diminution was as much as four-fifths the coagulation time of the control. Since this result was obtained by painting “adrenalin” on the pancreas, as well as by injecting it into the abdominal cavity, they concluded that “the phenomenon appears to be due to the application of adrenalin to the pancreas.” Six years later, during a study of the effect of adrenalin on internal hemorrhage, Wiggers[2] examined incidentally the evidence presented by Vosburgh and Richards, and after many tests on five dogs found “never the slightest indication that adrenalin, either when injected or added to the blood, appreciably hastened the coagulation process.” In 1911 von den Velden[3] reported that adrenin (about 0.007 milligram per kilo of body weight) decreased the coagulation time in man about one-half—an effect appearing 11 minutes after administration by[p. 137] mouth, and 85 minutes after subcutaneous injection. He affirmed also, but without describing the conditions or giving figures, that adrenin decreases coagulation time in vitro. He did not attribute the coagulative effect of adrenin in patients to this direct action on the blood, however, but to vasoconstriction disturbing the normal circulation and thereby the normal equilibrium between blood and tissue. In consequence, the tissue juices with their coagulative properties enter the blood, so he assumed. In support of this theory he offered his observation that coagulation time is decreased after the nasal mucosa has been rendered anemic by adrenin pledgets. Von den Velden’s claim[3] for adrenin given by mouth was subjected to a single test on man by Dale and Laidlaw,[4] but their result was completely negative.
The importance of Vosburgh and Richards’ observation, the thoroughly discordant testimony of later investigators, as well as the meager and incidental nature of all the evidence that has been adduced either for or against the acceleration of clotting by adrenin, made desirable a further study of this matter. Especially was this further study desirable because of the discharge of adrenin into the blood in pain and emotional excitement. Accordingly, in 1914, H. Gray and I[5] undertook an investigation of the question. In doing so we employed cats as subjects. Usually they were quickly[p. 138] decerebrated under ether, and then continuance of the drug became unnecessary. Body temperature was maintained by means of an electric heating pad. Respiration proceeded normally except in a few instances (in which, presumably, there was hemorrhage into the medulla), when artificial respiration had to be given.
In order to avoid, so far as possible, the personal element in determining when the blood was clotted, the blood was made to record its own clotting. The instrument by means of which this was done was the graphic coagulometer devised by W. L. Mendenhall and myself,[6] and illustrated diagrammatically in Fig. 24. It consists essentially of a light aluminum lever with the long arm nearly counterpoised by a weight W. The long arm is[p. 139] prevented from falling by a support S, and is prevented from rising by a horizontal right-angled rod reaching over the lever at R1 and fixed into the block B which turns on the axis A. Into the same block is fixed the vertical rod R2. When this rod is moved from the post P1, against which it is held by the weight of the horizontal rod R1, towards the other post P2, the check on the long arm of the lever is lifted, and if the short arm is heavier, the long arm will then rise.
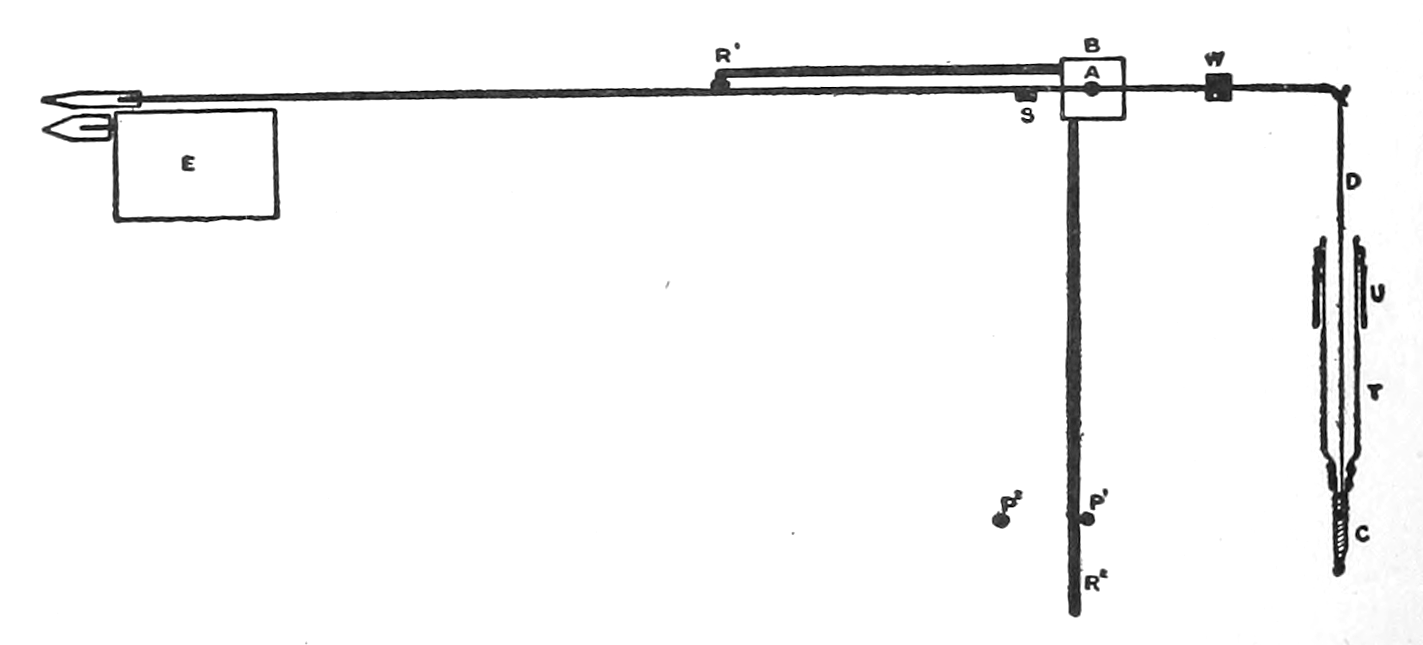
The cannula C, into which the blood is received, is two centimeters in total length and slightly more than two millimeters in internal diameter. It is attached by a short piece of rubber tubing to the tapered glass tube T, five centimeters long and five millimeters in internal diameter. The upper end of this tube is surrounded by another piece of rubber which supports the tube when it is slid into the U-shaped support U, fixed directly below the end of the short arm of the lever.
By drawing the cannulas from a single piece of glass tubing and by making the distance from shoulder to upper end about twelve millimeters, receptacles of fairly uniform capacity are assured. All the dimensions, the reach of the rubber connection over the top of the cannula (2–3 millimeters), the distance of the upper rubber ring from the lower end of the glass chamber (4 centimeters), etc., were as nearly standard as possible.
[p. 140]
A copper wire D, eight centimeters long and 0.6 millimeters in diameter, bent above into a hook and below into a small ring slightly less than two millimeters in diameter, is hung in a depression at the end of the short arm of the lever. The small ring then rests in the upper part of the cannula (see Fig. 24). The weight of the copper wire makes the short arm of the lever heavier than the long arm by 30 milligrams, when the delicate writing point is moving over a lightly smoked drum. Half a dozen of these standard wires are needed.
For accurate determination of the coagulation time Addis[7] has defined the following conditions as essential:
1. The blood must always be obtained under the same conditions.
2. Estimates must all be made at the same temperature.
3. The blood must always come in contact with the same amount and kind of foreign material.
4. The end point must be clear and definite and must always indicate the same degree of coagulation.
The precautions taken to fulfill these conditions were as follows:
1. Drawing the blood.—The blood was taken from the femoral artery. The artery (usually the right) was laid bare in the groin and freed from surrounding tissue. A narrow artery clip, with[p. 141] each limb enclosed in soft rubber tubing (to prevent injury of the tissues), and with its spring exerting gentle pressure, was placed on the artery immediately below the deep femoral branch, thus allowing no blood to stagnate above the clip. Between the clip and a ligature applied about 1.5 centimeters below, an opening was made. The blood was carefully milked out of the vessels between a blunt dissector moved beneath, and a small forceps, twisted into a pinch of absorbent cotton, moved above.
The cannula, cleaned in water, alcohol, and ether, was set in the rubber connection of the glass tube; the point of the cannula was then lubricated with vaseline and slipped into the artery. The pressure of the clip on the artery was next very slightly released and blood was allowed to flow into the cannula up to the lower border of the rubber connection. Only a good-sized drop of blood was needed. Sometimes the blood ran one or two millimeters above or below, but without appreciably changing the result. Since the clip was situated on the femoral immediately below a branch in which the circulation persisted, the blood received in the cannula was always fresh from the moving stream. As soon as the clip gripped the artery again, the cannula was slipped out. A helper then promptly milked the vessel in the manner described above, and covered it with a pad of absorbent cotton[p. 142] smeared with vaseline to prevent drying. Thereby blood was not permitted to stagnate; and when a new sample was to be taken, the vessel was clean and ready for use.
The tip of the cannula was at once plugged by plunging it into a flat mound of plasticine about three millimeters high. It was drawn off sidewise lest the plasticine plug be pulled out again. One of the copper wires D was now slid into the tube and cannula, the tube slipped into the U-support, and the wire lifted and hung on the lever. This procedure, from the moment blood began to flow until the wire was hung, consumed usually about twenty seconds.
2. Uniform temperature.—Under the U-support was placed a large water bath, in which the cannula and the tapering part of the tube were submerged. A thermometer was fixed to the U-support so that the bulb came near the cannula in the bath. The water was kept within a degree of 25° C. This temperature was chosen for several reasons: (a) The cannula has room temperature and rapidly cools the small volume of blood that enters it. To heat blood and cannula to body temperature would take time. A bath near room temperature, therefore, seems preferable to one near body temperature. (b) The test of clotting was conveniently made at intervals of a half-minute, and if the clotting process were hastened by higher temperatures,[p. 143] this interval would become relatively less exact. (c) A temperature of 25° C. rather than lower was selected because, as Dale and Laidlaw[8] have shown, the coagulation time is much slower for a given change in temperature below 25° than for the same change above. And with slowing of the process the end point, when the determination depends on supporting a weight, is less likely to be sharp. (d) The researches undertaken with use of this coagulometer were concerned with factors hastening the process. For that reason and for reason (b), a long rather than a short coagulation time for normal conditions was desirable.
3. Uniformity in the amount and kind of contact with foreign surface.—The capacity of the cannulas was fairly uniform, as stated above; the amount received in them was fairly constant; and the wire hanging in the blood presented approximately the same surface in different observations.
A further condition for insuring consistent treatment of the blood in different cases was that of making the tests for coagulation always at the same intervals. Below the writing point of the lever was set an electromagnetic signal E, which recorded half-minutes. At the moment a record was made by the signal (see first signal mark, Fig. 25) the clip on the artery was opened, the blood taken, and the process thus begun. In about 20 seconds the cannula was suspended in the water[p. 145] bath and the wire was hanging on the lever. At the next record by the signal and at every subsequent record the vertical rod R2 was pushed with the index finger from post P1 to post P2 and allowed to move back. This motion was uniform and lasted about one second. The check R1 on the long arm of the lever was thus raised, and as the wire sank in the blood the writing point rose, recording that coagulation had not taken place (see Fig. 25).

4. Definite end point.—As soon as the blood clotted, the weight of 30 milligrams was supported, and the failure of the lever to rise to the former height in the regular time allowed, recorded that the change had occurred.
Very rarely the swing of the lever would be checked for a moment and would then begin to move rapidly, indicating that a strand of fibrin had formed but not sufficiently strong to support the weight, and that when the strand broke, the weight quickly sank in the blood. If this occurred, the next record almost always was the short line, which signified that the weight was well supported.
A very slight strand of fibrin was able to prevent the weight from dropping, though at different times the amount of support differed, as shown by the varying length of the final lines (compare first and last series, Fig. 25). These variations are probably a rough indication of the degree of coagulation. In our experiments, however, the length of[p. 146] the final line was disregarded, and merely the fact that the lever failed to swing through its usual distance was taken as evidence of a clot, and the consequent short record was taken as the end point.
As soon as this end point was registered, the tube, wire and cannula were lifted out of the bath; the cannula was then separated from the tube and pulled away from the wire. The clot was thus disclosed, confirming the graphic record.
The method, at least when used at half-minute intervals, did not reveal in all instances the same degree of clotting. Usually, when the process was very rapid, the revealed clot was a thick jelly; whereas, when the process was slow, a strand of fibrin or at most a small amount of jelly was found. This difference in the degree of coagulation introduced, of course, an element of inexactness. In our experiments, however, this inexactness was unfavorable to the result we were seeking for, i. e., the acceleration of the process—because the jelly is a later stage than the fibrin strand; and since we nevertheless obtained good evidence of acceleration, we did not in these experiments attempt to determine more accurately differences in the stage of the clotting process.
5. Cleaning of apparatus.—After the wire was removed from the tube, the clot attached to its ring-tip was carefully brushed away under cool running water. Under the running water, also, a[p. 147] trimmed feather was introduced into the cannula and the tube to push out the plasticine and to wash out the blood. Wire, cannula and tube were then dropped into a beaker receiving running hot water (about 80° C.) and there allowed to remain for about five minutes. On removal from this the parts were shaken free from water, passed through 95 per cent alcohol and again shaken free, passed through ether and let dry.
By having a half-dozen cannulas and wires of standard size, it was possible to save trouble by cleaning a number at one time.
Not infrequently the first few samples of blood taken from an animal showed rapid or somewhat irregular rates of clotting. Some causes for these initial variations will be presented in following pages. The fairly uniform rate of clotting in any individual after the initial stage, varied in twenty-one different animals from an average of 3 to an average of 10.6 minutes, with a combined average of 5.9 minutes. The conditions for these variations among the individuals have not been wholly determined.
The first observations were of this class.
Oct. 27. A cat weighing about 3 kilos was given 3 cubic centimeters of adrenin 1:1,000, i. e., 1 milligram per kilo, under the skin. The animal, in this[p. 148] instance, was kept in uniform ether anesthesia. Following is a record showing when blood was taken, and the coagulation time in each instance:
| 2.56 | Injection made | |
| .59 | 6 | minutes |
| 3.07 | 5.5 | “ |
| .13 | 5 | “ |
| .20 | 6.5 | “ |
| —— | ||
| Average | 5.7 | minutes |
| 3.27 | 3.5 | minutes |
| .44 | 2 | “ |
| .55 | 2.5 | “ |
| 4.07 | 3 | “ |
| .20 | 2 | “ |
| —— | ||
| Average | 2.6 | minutes |
| 4.44 | 6 | minutes |
| 5.00 | 4.5 | “ |
| 5.50 | 5 | “ |
| —— | ||
| Average | 5.2 | minutes |
In this case the coagulation time remained at its usual level for about 20 minutes after the subcutaneous injection.[*] Thereafter for about an hour the coagulation time averaged 45 per cent of its previous duration. And widely separated tests made during the following hour indicated that approximately the initial rate of clotting had been regained.
* This period is longer than is expected after the subcutaneous injection of any drug. As will be shown later, strong doses of adrenin, if injected rapidly, may not at first shorten the clotting process. Probably in some instances of subcutaneous injection of these strong doses, the drug enters the circulation more rapidly than in others and in consequence coagulation is not at first accelerated.
The rather long period (nearly 30 minutes), in the case just cited, between the injection and the[p. 149] first appearance of rapid clotting was not the rule. As the following figures show, the coagulation time may become shortened quite promptly after subcutaneous injection.
| Oct. 29. | 3.30 | 5.5 | minutes |
| .36 | 5.5 | “ | |
| .44 | Adrenin, 3 cubic centimeters, 1:1,000, injected subcutaneously. | ||
| .46 | 5.5 | minutes | |
| .53 | 4 | “ | |
| 4.01 | 3.5 | “ | |
| .08 | 3.5 | “ | |
| .16 | 4.5 | “ | |
| .23 | 5 | “ | |
| .30 | 5.5 | “ | |
In this case nine minutes after the injection the change in the rate of clotting had begun, and it continued more rapid for the subsequent half-hour.
We did not attempt to find the minimal subcutaneous dose which would shorten clotting. A dose of 0.01 milligram per kilo, however, has proved effective, as shown by the following figures:
| Feb. 3. | 11.34 | 10 | minutes |
| .45 | 9 | “ | |
| .50 to .52 | Adrenin, 2.8 cubic centimeters, 1:100,000, injected under skin of groin in cat weighing 2.8 kilos. | ||
| .55 | 10 | minutes | |
| 12.06 | 7 | “ | |
| .14 | 4 | “ | |
| .19 | 5.5 | “ | |
| .31 | 6 | “ | |
| .37 | 7 | “ | |
| .45 | 9 | “ | |
As will be shown later, the dose in this instance was ten times the minimal effective intravenous dose. On the basis of these figures, less than a milligram of adrenin given subcutaneously would be necessary to shorten clotting to a marked degree in a man of average weight (70 kilograms).
[p. 150]
Not many observations were made by us on the effects of adrenin administered subcutaneously. The amount reaching the vascular system and the rate of its entrance into the blood could be so much more accurately controlled by intravenous than by subcutaneous introduction that most of our attention was devoted to the latter method.
In this procedure a glass cannula was fastened in one of the external jugular veins and filled with the same solution as that to be injected. A short rubber tube was attached and tightly clamped close to the glass. Later, for the injection, the syringe needle was inserted through the rubber and into the fluid in the cannula, the clip on the vein was removed, and the injection made.
The solutions employed intravenously were adrenin 1:10,000, 1:50,000, and 1:100,000, in distilled water.
The smallest amount which produced any change in clotting time was 0.1 cubic centimeter of a dilution of 1:100,000 in a cat weighing two kilos, a dose of 0.0005 milligram per kilo. Four tests previous to the injection averaged 5 minutes, and none was shorter than 4 minutes. Immediately after the injection the time was 2 minutes, but at the next test the effect had disappeared. Doubling the dose in the same cat—i. e., giving 0.2 cubic centimeter[p. 151] (0.001 milligram per kilo)—shortened the coagulation time for about 40 minutes:
| Dec. 23. | 10.30 | 4 | minutes |
| .35 | 4 | “ | |
| .41 | 4 | “ | |
| .46 | Adrenin, 0.001 milligram per kilo. | ||
| .47 | 2.5 | minutes | |
| .50 | 3 | “ | |
| .53 | 3.5 | “ | |
| 11.00 | 1.5 | “ | |
| .05 | 1.5 | “ | |
| .10 | 3 | “ | |
| .15 | 2 | “ | |
| .20 | 4 | “ | |
| .26 | 4.5 | “ | |
| .31 | 5 | “ | |
From 10.47, immediately after the second injection, till 11.20 the average time for clotting was 2.5 minutes, whereas both before and after this period the time was 4 minutes or longer. At 11.00 o’clock and 11.05, when the end point was reached in 1.5 minutes (a reduction of 63 per cent), a thick jelly was found on examining the cannula. The changes in clotting time in this case are represented graphically in Fig. 26.
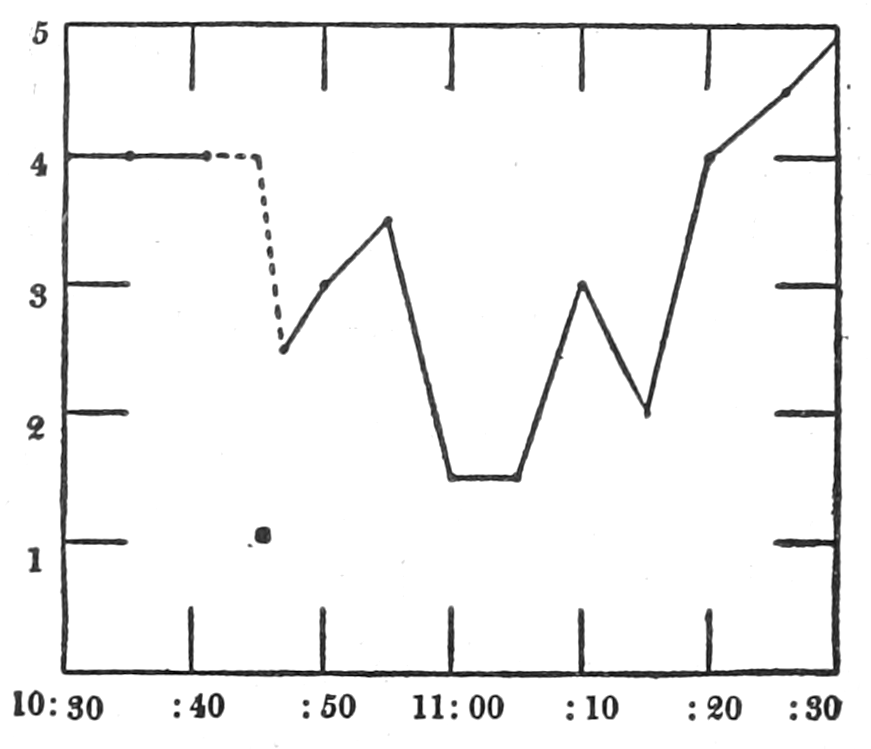
In another case a dose of 0.0005 milligram per kilo failed to produce any change, but 0.001 milligram per kilo (0.28 cubic centimeter of adrenin, 1:100,000, given a cat weighing 2.8 kilos) brought a sharp decline in the record, as follows:
| Jan. 9. | 11.32 | 6 | minutes |
| .40 | 6 | “ | |
| .47 | Adrenin, 0.001 milligram per kilo. | ||
| .48 | 5.5 | minutes | |
| .55 | 4 | “ | |
| 12.00 | 5.5 | “ | |
| .06 | 7 | “ | |
In these instances the animals were decerebrated. For decerebrate cats, the least amount of[p. 152] adrenin, intravenously, needed to produce shortening of coagulation time is approximately 0.001 milligram per kilo.
In the above cases rapid clotting was manifest directly after minute doses. Larger doses, however, may produce primarily not faster clotting but slower, and that may be followed in turn by a much shorter coagulation time. The figures below present such an instance:
| Nov. 25. | 2.36 | 3 | minutes |
| .40 | 3 | “ | |
| .43 | Adrenin, 0.5 cubic centimeter, 1:10,000. | ||
| .44 | 4 | minutes | |
| .49 | 3.5 | “ | |
| .53 | 1.5 | “ | |
| .55 | 1.5 | “ | |
| .58 | 2 | “ | |
| 3.00 | 2.5 | “ | |
| .03 | 1.5 | “ | |
| .05 | 1.5 | “ | |
| .07 | 2.5 | “ | |
| .10 | 1.5 | “ | |
| .14 | 1.5 | “ | |
| .16 | 2.5 | “ | |
| .19 | 3 | “ | |
| .23 | 3 | “ | |
| .30 | 3 | “ | |
[p. 153]
This unexpected primary increase of coagulation time, lasting at least six minutes, is in striking contrast to the later remarkable shortening of the process from 3 to an average of 1.7 minutes for more than 20 minutes (see Fig. 27, A).
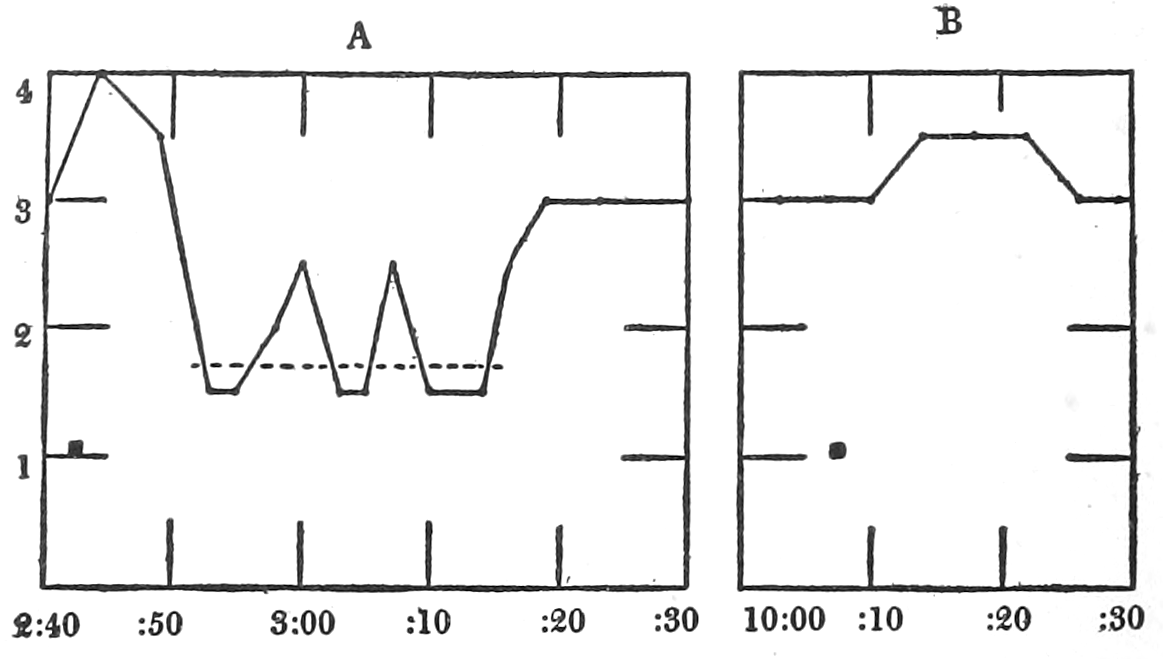
If a strong solution, i. e., 1:10,000, is injected rapidly, the process may be prolonged as above, but not followed as above by shortening, thus:
| Nov. 28. | 9.59 | 3 | minutes |
| 10.03 | 3 | “ | |
| .08 | Adrenin, 0.5 cubic centimeter, 1:10,000. | ||
| .10 | 3 | minutes | |
| .14 | 3.5 | “ | |
| .18 | 3.5 | “ | |
| .22 | 3.5 | “ | |
| .26 | 3 | “ | |
| .29 | 3 | “ | |
| .33 | 3 | “ | |
There was in this case no decrease in coagulation time at any test for a half-hour after the injection, but instead a lengthening (see Fig. 27, B). Howell[9] has reported the interesting observation that repeated massive doses of adrenin given to dogs may so greatly retard coagulation that the animals may be said to be hemophilic. These two instances show that on coagulation large doses have the contrary effect to small, just as Hoskins[10] showed was true for intestinal and Lyman and I[11] showed was true for arterial smooth muscle.
In a few experiments the brain and the cord to midthorax were destroyed through the orbit. Artificial respiration then maintained the animal in uniform[p. 154] condition. Under these circumstances, adrenin intravenously had more lasting effects than when given to the usual decerebrate animals with intact cord. Fig. 28 illustrates such a case. For thirty minutes before injection the clotting time averaged 5.4 minutes. Then, about ten minutes after one cubic centimeter of adrenin, 1:50,000, had been slowly injected, clotting began to quicken; during the next twenty minutes the average was 3.4 minutes, and during the following forty-five minutes the average was 1.9 minutes—only 35 per cent as long as it had been before the injection.
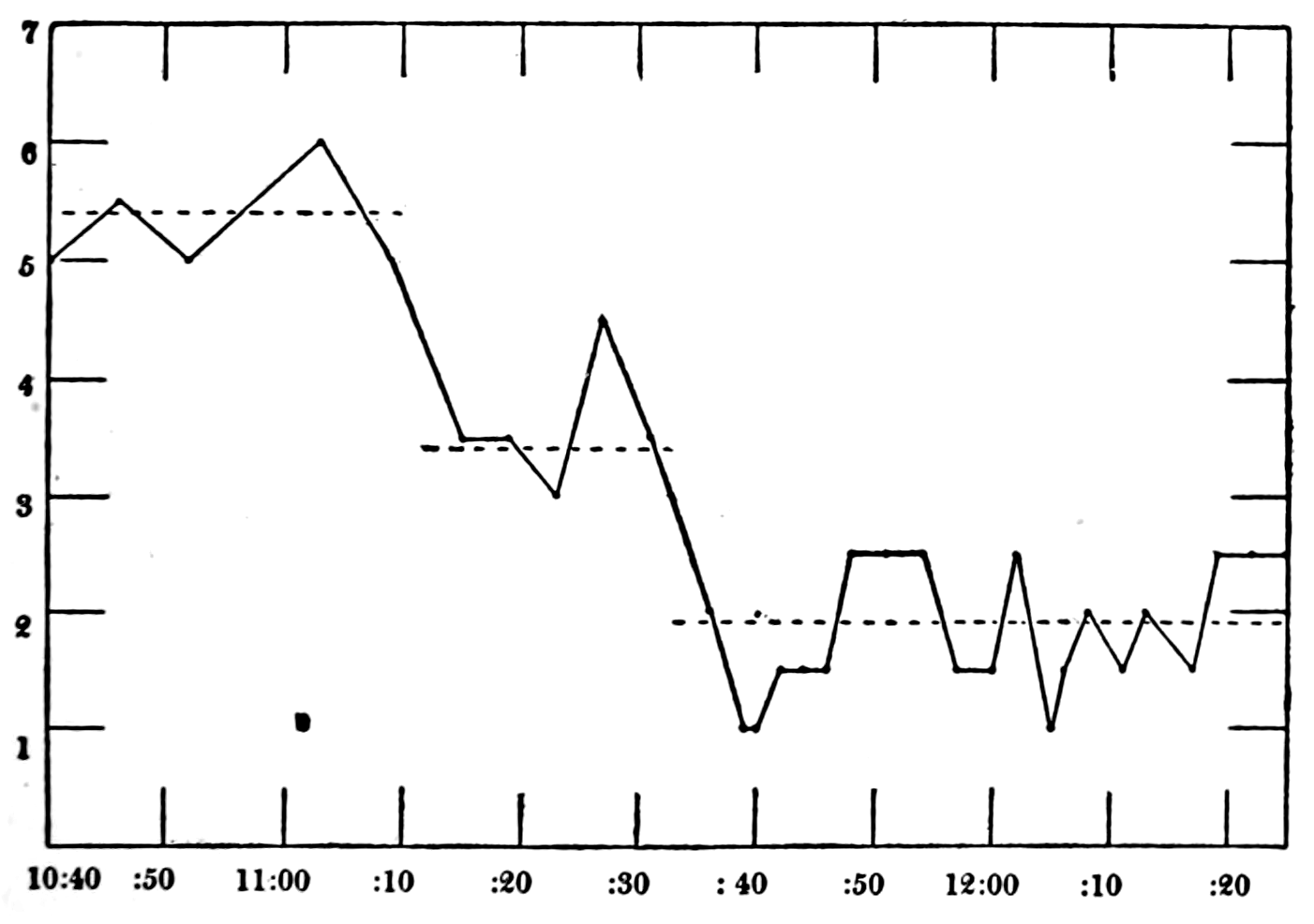
In another case in which the brain and upper cord were similarly destroyed, the clotting time, which for a half-hour had averaged 3.9 minutes, was reduced by one cubic centimeter of adrenin,[p. 155] 1:100,000, to an average for the next hour and forty minutes of 2.3 minutes, with 1.5 and 3 minutes as extremes. During the first forty minutes of this period of one hour and forty minutes of rapid clotting all of eight tests except two showed a coagulation time of 2 minutes or less. The explanation of this persistent rapid clotting in animals with spinal cord pithed is not yet clear.
As indicated in Figs. 26, 27 and 28, the records of coagulation show oscillations. Some of these ups and downs are, of course, within the limits of error of the method, but in our experience they have occurred so characteristically after injection of adrenin, and so often have appeared in a rough[p. 156] rhythm, that they have given the impression of being real accompaniments of faster clotting. It may be that two factors are operating, one tending to hasten, the other to retard the process, and that the equilibrium disturbed by adrenin is recovered only after interaction to and fro between the two factors.
The oscillations in coagulation time after the injections suggest that clotting might vary with changes in blood pressure, for that also commonly oscillates after a dose of adrenin (see, e. g., Fig. 23). Simultaneous recording of blood pressure and determining of coagulation time have revealed that each may vary without corresponding variation in the other. Within ordinary limits, therefore, changes of blood pressure do not change the rate of clotting.
As previously stated, von den Velden has contended that shortening of coagulation time by adrenin is due to exudation of tissue juices resulting from vasoconstriction. The amount of adrenin which produces markedly faster clotting in the cat, is approximately 0.001 milligram per kilo. As Lyman and I[12] showed, however, this amount when injected slowly, as in the present experiments, results in brief vasodilation rather than[p. 157] vasoconstriction. Von den Velden’s explanation can therefore not be applied to these experiments.
He has claimed, furthermore, that adrenin added to blood in vitro makes it clot more rapidly, but, as already noted, he gives no account of the conditions of his experiments and no figures. It is impossible, therefore, to criticise them. His claim, however, is contrary to Wiggers’s[13] earlier observations that blood with added adrenin coagulated no more quickly than blood with an equal amount of added physiological salt solution. Also contrary to this claim are the following two experiments: (1) Ligatures were tied around the aorta and inferior vena cava immediately above the diaphragm, and thus the circulation was confined almost completely to the anterior part of the animal. Indeed, since the posterior part ceases to function in the absence of blood supply, the preparation may be called an “anterior animal.” When such a preparation was made and 0.5 cubic centimeter of adrenin, 1:100,000 (half the usual dose, because, roughly, half an animal), was injected slowly into one of the jugulars, coagulation was not shortened. Whereas for a half-hour before the injection the clotting time averaged 4.6 minutes, for an hour thereafter the average was 5.3 minutes—a prolongation which may have been due, not to any influence of adrenin, but to failure of the blood to circulate through the intestines and liver.[14] In another[p. 158] experiment after the gastro-intestinal canal and liver had been removed from the animal, the average time for coagulation during twenty-five minutes before injecting adrenin (0.23 cubic centimeter, 1:100,000, in an animal weighing originally 2.3 kilos) was 5.5 minutes, and during forty minutes after the injection it was 6.8 minutes, with no case shorter than 6 minutes. In the absence of circulation through the abdominal viscera, therefore, adrenin fails to shorten the clotting time. (2) The cannulas were filled with adrenin, 1:1,000, and emptied just before being introduced into the artery. The small amount of adrenin left on the walls was thus automatically mixed with the drawn blood. Alternate observations with these cannulas wet by adrenin and with the usual dry cannulas showed no noteworthy distinction.
| Feb. 19. | 2.21 | 6 | minutes, | with | usual | cannula |
| .30 | 6.5 | “ | “ | “ | “ | |
| .36 | 6.5 | “ | “ | adrenin | “ | |
| .49 | 6 | “ | “ | “ | “ | |
| .56 | 7 | “ | “ | usual | “ | |
| 3.04 | 6 | “ | “ | adrenin | “ |
The results of these experiments have made it impossible for us to concede either of von den Velden’s claims, i. e., that clotting occurs faster because adrenin is added to the blood, or because adrenin by producing vasoconstriction causes tissues to exude coagulant juices.
Vosburgh and Richards found that coagulation[p. 159] became more rapid as the blood sugar increased. Conceivably faster clotting might result from this higher percentage of blood sugar. Against this assumption, however, is the fact that clotting is greatly accelerated by 0.001 milligram adrenin per kilo of body weight, much less than the dose necessary to increase the sugar content of the blood.[15] And furthermore, when dextrose (3 cubic centimeters of a 10 per cent solution) is added to the blood of an anterior animal, making the blood sugar roughly 0.3 per cent, the coagulation time is not markedly reduced. Adrenin appears to act, therefore, in some other way than by increasing blood sugar.
Since adrenin makes the blood clot much faster than normally in the intact animal, and fails to have this effect when the circulation is confined to the anterior animal, the inference is justified that in the small doses here employed adrenin produces its remarkable effects, not directly on the blood itself, not through change in the extensive neuro-muscular, bony, or surface tissues of the body, but through some organ in the abdomen.
That exclusion of the liver from the bodily economy, by ligature of its vessels or by phosphorus poisoning, will result in great lengthening of the coagulation time has been clearly shown. The liver, therefore, seems to furnish continuously to the blood a factor in the clotting process which is[p. 160] being continuously destroyed in the body. It is not unlikely that adrenin makes the blood clot more rapidly by stimulating the liver to discharge this factor in greater abundance. But proof for this suggestion has not yet been established.
1 Vosburgh and Richards: American Journal of Physiology, 1903, ix, p. 39.
2 Wiggers: Archives of Internal Medicine, 1909, iii, p. 152.
3 Von den Velden: Münchener medizinische Wochenschrift, 1911, lviii, p. 187.
4 Dale and Laidlaw: Journal of Pathology and Bacteriology, 1912, xvi, p. 362.
5 Cannon and Gray: American Journal of Physiology, 1914, xxxiv, p. 321.
6 Cannon and Mendenhall: American Journal of Physiology, 1914, xxxiv, p. 225.
7 Addis: Quarterly Journal of Experimental Physiology, 1908, i, p. 314.
8 Dale and Laidlaw: Loc. cit., p. 359.
9 Howell: American Journal of Physiology, 1914, xxxiii, p. xiv.
10 Hoskins: American Journal of Physiology, 1912, xxix, p. 365.
11 Cannon and Lyman: American Journal of Physiology, 1913, xxxi, p. 376.
12 Cannon and Lyman: Loc. cit., p. 381.
13 Wiggers: Loc. cit., p. 152.
14 See Pawlow: Archiv für Physiologie, 1887, p. 458. Bohr: Centralblatt für Physiologie, 1888, ii, p. 263. Meek: American Journal of Physiology, 1912, xxx, p. 173. Gray and Lunt: Ibid., 1914, xxxiv, p. 332.
15 Cannon: American Journal of Physiology, 1914, xxxiii, p. 396.
[p. 161]
THE HASTENING OF THE COAGULATION OF BLOOD IN PAIN AND GREAT EMOTION
In the foregoing chapter evidence was presented that the intravenous injection of minute amounts of adrenin hastens the clotting of blood. The amounts used did not vary much above or below the amounts discharged by the adrenal glands after brief stimulation of the splanchnic nerves, as found by H. Osgood in the Harvard Laboratory, and may therefore be regarded as physiological. Since injected adrenin is capable of shortening the coagulation time, may not the increased secretion of the adrenals likewise have that effect? The answer to this question was the object of an investigation by W. L. Mendenhall and myself.[1]
The blood was taken and its coagulation was recorded graphically in the manner already described. In some instances the cats were etherized, in others they were anesthetized with urethane, or were decerebrated. The splanchnic nerves always were stimulated after being cut away from connection[p. 162] with the spinal cord. Sometimes the nerves were isolated unilaterally in the abdomen; sometimes, in order to avoid manipulation of the abdominal viscera, they were isolated in the thorax and stimulated singly or together. A tetanizing current was used, barely perceptible on the tongue and too weak to cause by spreading any contraction of skeletal muscles.
That splanchnic stimulation accelerates the clotting of blood, and that the effects vary in different animals, are facts illustrated in the following cases:
Oct. 25.—A cat was etherized and maintained in uniform ether anesthesia. After forty minutes of preliminary observation the left splanchnic nerves were stimulated in the abdomen. Following are the figures which show the effects on the coagulation time:
| 3.00 | 4 | minutes |
| .07 | 5.5 | “ |
| .14 | 4 | “ |
| .32 | 4.5 | “ |
| .39 to .40 | Stimulation of left splanchnic. | |
| .42 | 5 | minutes |
| .49 | 5 | “ |
| .56 | 2 | “ |
| 4.00 | 1 | “ |
| .03 | 2.5 | “ |
| .07 | 2.5 | “ |
| .11 | 3 | “ |
| .16 | 2 | “ |
| .20 | 1.5 | “ |
| .23 | 4 | “ |
| .29 | 5.5 | “ |
| .40 | 5.5 | “ |
| .50 | 5 | “ |
[p. 163]
In this instance at least ten minutes elapsed between the end of stimulation and the beginning of faster clotting. The period of faster clotting, however, lasted for about a half-hour, during which the coagulation time averaged 2.1 minutes, only forty-three per cent of the previous average of 4.8 minutes. It is noteworthy that the curve (see Fig. 29), while lower, shows oscillations not unlike those which follow injection of adrenin (see p. 155).
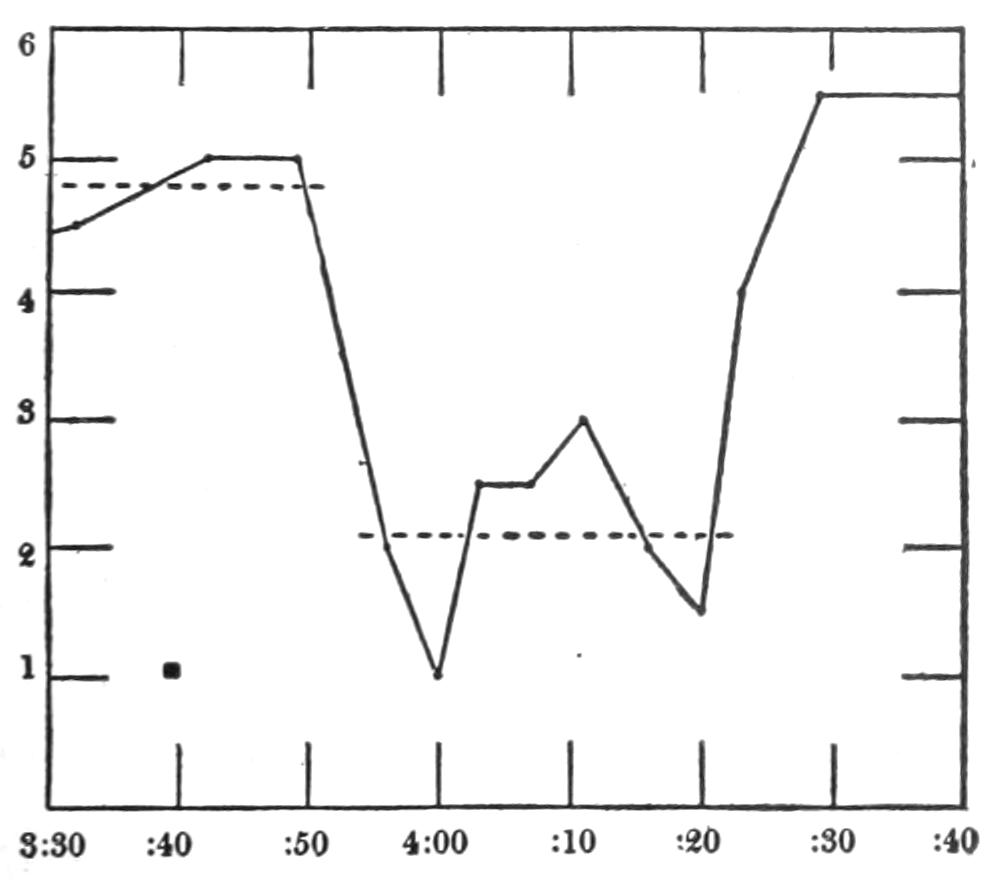
The primary delay of the effect is not always, indeed it is not commonly, present:
Nov. 6.—A cat was anesthetized (1.40 p.m.) with urethane, and later (3.05) its brain was pithed. The following observations on the coagulation time show the prompt effect of splanchnic stimulation:
[p. 164]
| 3.36 | 7 | minutes |
| .46 | 6 | “ |
| 4.02 to .05 | Stimulation of left splanchnic in abdomen. | |
| .08 | 4 | minutes |
| .10 | 3 | “ |
| .18 | 3.5 | “ |
| .23 | 6.5 | “ |
In Fig. 30 is presented the original record of the shortening of the coagulation after stimulation of the left splanchnic nerve (Nov. 8) in a cat with brain pithed.

In the foregoing instances the coagulation time was reduced after splanchnic stimulation to less than half what it was before. The reduction was not always so pronounced.
Nov. 7.—A cat[*] maintained in uniform ether anesthesia with artificial respiration had the following changes in the clotting time of its blood as the result of stimulating the left splanchnic nerve in the thorax:
| 3.40 | 5 | minutes |
| .45 | 5 | “ |
| .51 | 5.5 | “ |
| .58 to 4.00 | Stimulation of left splanchnic. | |
| 4.01 | 4.5 | minutes |
| .06 | 3.5 | “ |
| .11 | 4 | “ |
| .16 | 3.5 | “ |
| .21 | 4 | “ |
| .26 | 4.5 | “ |
| .31 | 5 | “ |
| .36 | 6.5 | “ |
* This animal had just passed through a period of excitement with rapid clotting.
In this case the average for about fifteen minutes before stimulation was slightly over five minutes,[p. 166] and for twenty-five minutes thereafter it was four minutes.
In all cases thus far the period of shortened coagulation lasted from ten to thirty minutes. In other cases, however, the effect was seen only in a single observation. If this had occurred only once after splanchnic stimulation, it might be attributed to accident, but it was not an infrequent result, e. g.:
Oct. 28.—A cat was etherized and decerebrated, and the splanchnic nerves were isolated in the thorax. Following are two instances of brief shortening of coagulation after splanchnic stimulation:
| 3.36 | 4.5 | minutes |
| .42 | 4.5 | “ |
| .47 to .49 | Splanchnic stimulation. | |
| .51 | 4.5 | minutes |
| .57 | 2 | “ |
| 4.01 | 4 | “ |
| .07 | 4.5 | “ |
| .12 | 5.5 | “ |
| .19 to .22 | Splanchnic stimulation. | |
| .23 | 3.5 | minutes |
| .27 | 4 | “ |
| .33 | 5 | “ |
In the foregoing instance it is noteworthy that the degree of acceleration is not so great after the second stimulation of the splanchnics as it was after the first. This reduction of effect as the nerves were repeatedly stimulated was frequently noted. The following case presents another illustration:
Nov. 12.—A cat was etherized (2.35 p.m.) and the medulla was punctured (piqûre) at 3.12. The[p. 167] operation was without effect. The loss or lessening of effectiveness on second stimulation of the left splanchnic nerves is to be compared with the persistence of effectiveness on the right side:
| 3.40 | 4.5 | minutes |
| .45 | 4.5 | “ |
| .54 to .56 | Stimulation of left splanchnic in abdomen. | |
| 4.00 | 3 | minutes |
| .05 | 2 | “ |
| .10 | 5.5 | “ |
| .16 | 5 | “ |
| .22 to .27 | Stimulation of left splanchnic in abdomen. | |
| .30 | 4 | minutes |
| .34 | 4 | “ |
| .39 | 4 | “ |
| .44 | 4 | “ |
| .48 | 4 | “ |
| .55 to .57 | Stimulation of right splanchnic. | |
| .59 | 3 | minutes |
| 5.02 | 2.5 | “ |
| .07 | 3 | “ |
| .11 | 3 | “ |
| .15 | 5.5 | “ |
| .22 | 5.5 | “ |
The experiments above recorded show that stimulation of the splanchnic nerves results immediately, or after a brief period, in a shortening of the coagulation time of the blood—an effect which in different animals varies in duration and intensity, and diminishes as the stimulation is repeated. The next question was whether this effect is produced through the adrenal glands.
The manner in which splanchnic stimulation produces its effects is indicated in the following experiments:
Nov. 28.—A cat was etherized, and through the[p. 168] orbit the central nervous system was destroyed to the midthorax. The blood vessels of the left adrenal gland were then quickly tied and the gland removed. The readings for a half hour before the left splanchnic nerve was stimulated averaged seven minutes, then—
| 4.38 to .40 | Stimulation of left splanchnic (glandless). | |
| .42 | 7 | minutes |
| .50 | 7 | “ |
| 5.02 to .04 | Stimulation of right splanchnic. | |
| .06 | 4 | minutes |
| .10 | 7 | “ |
| .18 | 7 | “ |
| .26 | 7 | “ |
Dec. 4.—A cat was etherized and pithed through the orbit to the neck region. The right and left splanchnic nerves were tied and cut in the thorax. The left adrenal gland was then carefully removed. These operations consumed about a half-hour. The following records show the effect of stimulating the left and right splanchnic nerves:
| 4.10 | 5 | minutes |
| .16 | 4.5 | “ |
| .25 to .28 | Stimulation of left splanchnic (glandless). | |
| .30 | 4.5 | minutes |
| .35 | 4.5 | “ |
| .40 | 7.5 | “ |
| .49 to .51 | Stimulation of right splanchnic. | |
| .55 | 4.5 | minutes |
| 5.00 | 2.5 | “ |
| .14 | 6 | “ |
| .23 to .25 | Stimulation of right splanchnic. | |
| .26 | 6 | minutes |
| .33 | 4.5 | “ |
| .38 | 3.5 | “ |
| .43 | 4.5 | “ |
| .49 | 5 | “ |
| .55 | 6 | “ |
[p. 169]
The results in this experiment are represented graphically in Fig. 31.
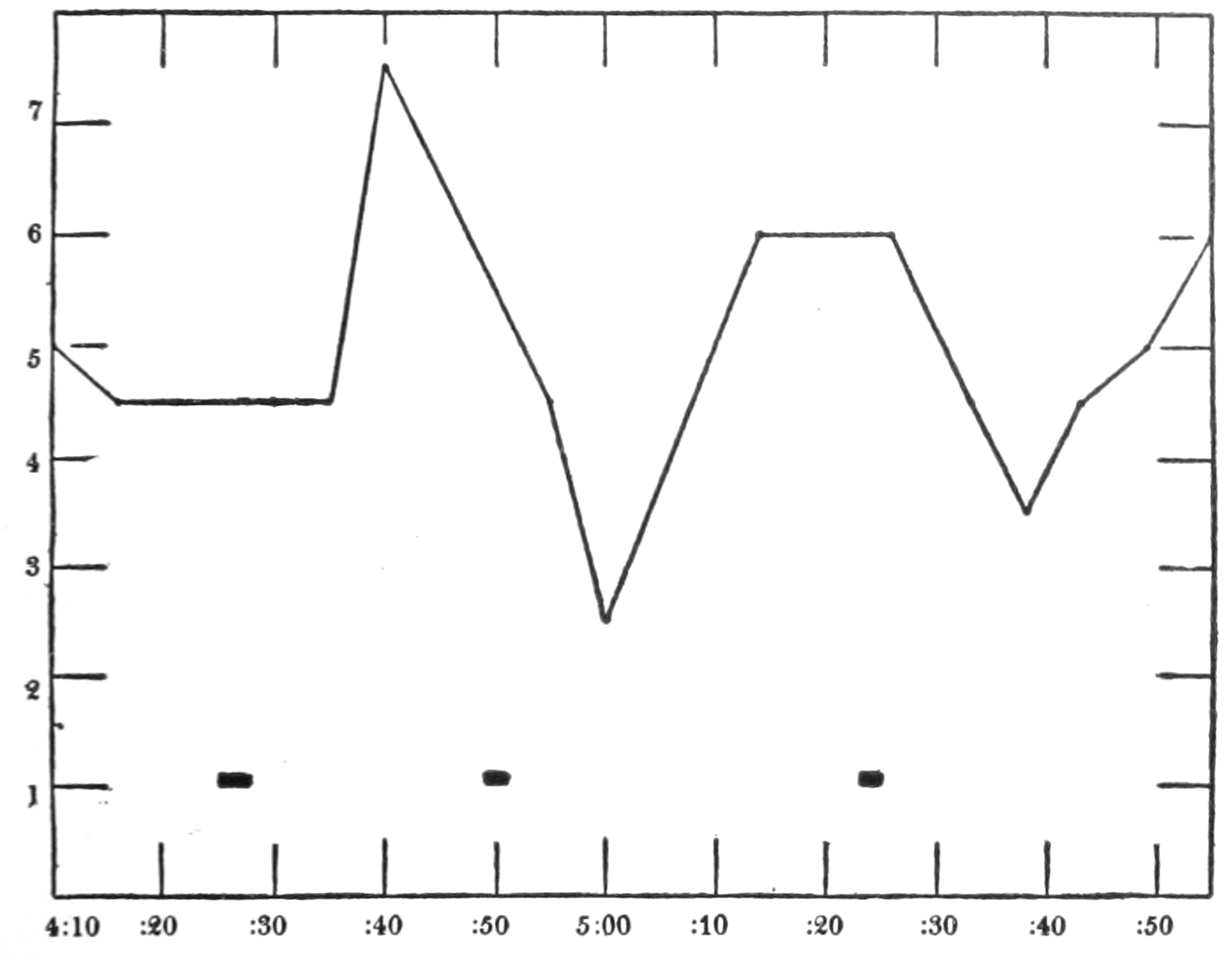
Elliott’s evidence that in the cat the splanchnic innervation of the adrenals is not crossed has already been mentioned. If the gland is removed on one side, therefore, stimulation of the nerves on that side causes no discharge from the opposite gland. As the above experiments clearly show, splanchnic stimulation on the glandless side results in no shortening of the coagulation time; whereas, in the same animals, stimulation of the nerves on[p. 170] the other side (still connected with the adrenal gland) produces a sharp hastening of the clotting process.
The splanchnics innervate the intestines and liver even though the adrenal gland is removed. The foregoing experiments indicate that the nerve impulses delivered to these organs do not influence them in any direct manner to accelerate the speed of coagulation. Indeed, in one of the experiments (Dec. 4, see Fig. 31) a high reading about ten minutes after splanchnic stimulation on the glandless side suggests the possibility of an opposite effect. Direct stimulation of the hepatic nerves on one occasion was followed by a change of the clotting time from 4.5, 5, 4.5, 4.5 minutes during twenty-five minutes before stimulation to 4.5, 7, and 6 minutes during twenty minutes after stimulation.
Since with the adrenals present stimulation of hepatic nerves induces alteration of glycogen in the liver and quick increase of blood sugar,[2] just as splanchnic stimulation does, the failure of the blood to clot faster after stimulation of the hepatic nerves confirms the evidence already offered that faster clotting when adrenin is increased in the blood is not due to a larger amount of sugar present (see p. 159).
The liver and intestines cannot be made to shorten clotting time by stimulation of their nerves, but, as has already been shown (see p. 157),[p. 171] neither can adrenin act by itself to hasten the clotting process. Apparently the effect is produced by coöperation between the adrenals and the liver (and possibly also the intestines). Somewhat similar coöperation is noted in the organization of sugar metabolism; splanchnic stimulation in the absence of the adrenal glands does not increase blood sugar,[3] and in the absence of the liver adrenin is without influence.[4]
The variations of effect noted after splanchnic stimulation can be accounted for by variations in the adrenin content of the glands. Elliott[5] found, as previously stated, that animals newly brought into strange surroundings may have a considerably reduced amount of adrenin in their adrenals. The animals used in our experiments had been for varying lengths of time in an animal house in which barking dogs were also kept, and were therefore subject to influences which would be likely to discharge the glands.
The evidence that stimulation of splanchnic nerves, with accompanying increase of adrenal secretion, results in more rapid clotting of blood is especially interesting in relation to the experiments previously described, which showed that in pain and emotional excitement there is an increased secretion of adrenin into the blood. Does the adrenin thus liberated have any effect on the rate of coagulation? The observations here recorded were[p. 172] made in order to obtain an answer to that question.
In the experiments on the action of stimuli which in the unanesthetized animal would cause pain, it will be recalled that faradic stimulation of a large nerve trunk (the stump of the cut sciatic) and operation under light anesthesia were the methods used to affect the afferent nerves. Elliott[6] found that repeated excitation of the sciatic nerve was especially efficient in exhausting the adrenal glands of their adrenin content, and also[p. 173] that this reflex persisted after removal of the cerebral hemispheres. It was to be expected, therefore, that with well-stored glands, sciatic stimulation, even in the decerebrate animal, would call forth an amount of adrenal secretion which would decidedly hasten clotting. The following case illustrates such a result:
Dec. 12.—A cat was anesthetized with ether at 3.45 and the left sciatic nerve was bared. Decerebration was completed at 3.57. The clotting time of the blood began to be tested six minutes later:
| 4.03 | 4 | minutes |
| .08 | 3.5 | “ |
| .13 | 3.5 | “ |
| .18 | 4.5 | “ |
| .23 to .25 | Stimulation of left sciatic. | |
| 4.26 | 2.5 | minutes |
| .29 | 3.5 | “ |
| .34 | 4 | “ |
| .40 | 5 | “ |
| .45 to .50 | Stimulation of left sciatic. | |
| 4.53 | 2.5 | minutes |
| .57 | 7 | “ |
| 5.06 | 7.5 | “ |
| .15 to .17 | Stimulation of left sciatic. | |
| 5.17 | 4 | minutes |
| .22 | 4.5 | “ |
| .27 | 5.5 | “ |
| .36 | 5.5 | “ |
| .46 | 7 | “ |
The results obtained in this case, which were similar to results in other cases, are represented graphically in Fig. 32. The coagulation time was becoming gradually more prolonged, but each excitation of the sciatic nerve was followed by a marked shortening. The strength of stimulation was not determined with exactness, but it[p. 174] is worthy of note that the current used in the first and the third stimulations was weaker than could be felt on the tongue, whereas that used in the second was considerably stronger, though it did not produce reflex spasms.
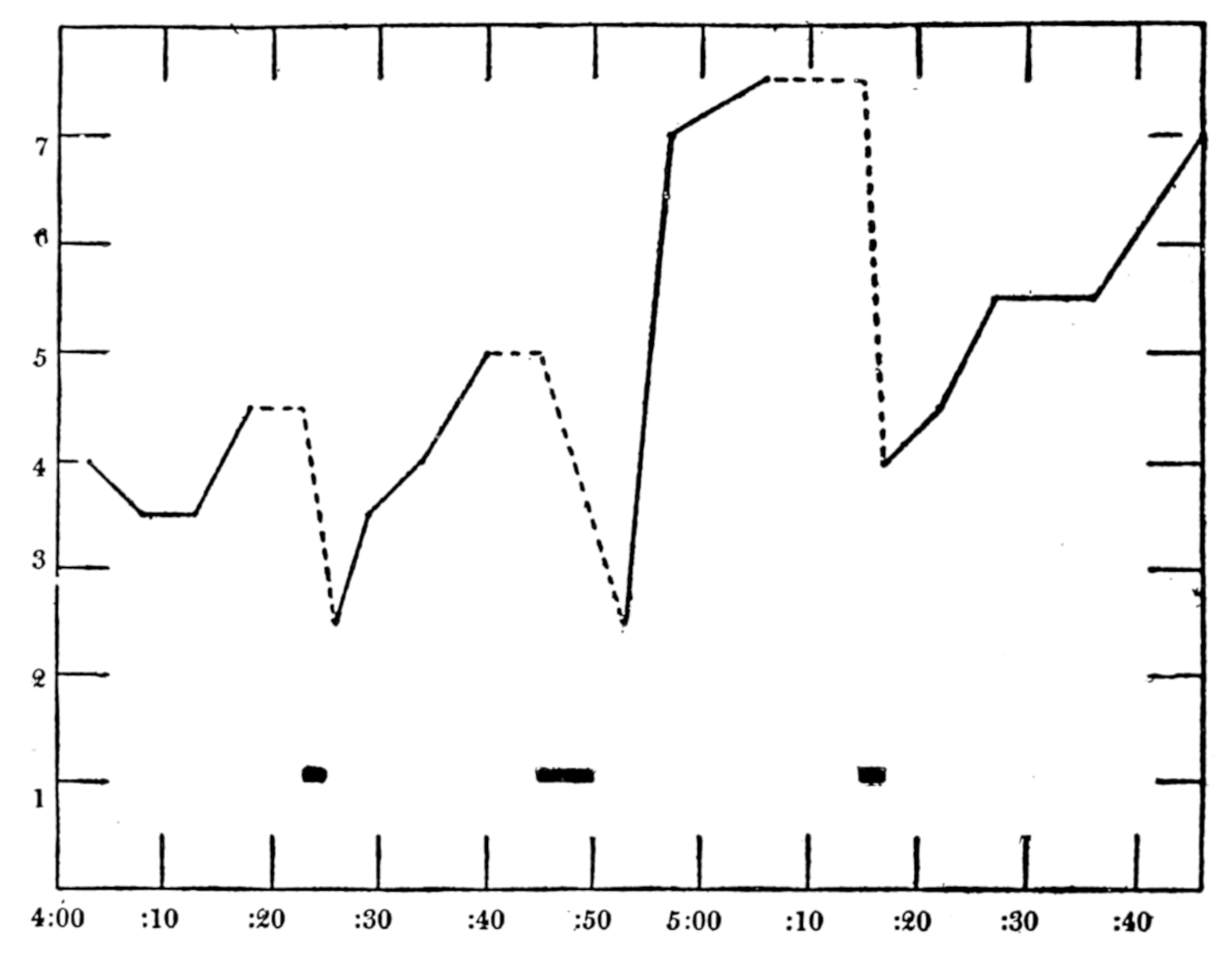
Mere tying of the nerve is capable of producing a marked shortening of coagulation, as the following figures show:
Oct. 21.—10.57 cat under ether, and urethane given:
| 11.11 | 8.5 | minutes |
| .23 | 8.5 | “ |
| .32 to .35 | Left sciatic bared and tied. | |
| .37 | 1.5 | minutes |
| .41 | 5.5 | “ |
| .50 | 7 | “ |
| 12.02 | 8.5 | “ |
Stimulation of the crural nerve had similar effects, reducing the clotting time in one instance from a succession of 3, 3, and 3.5 minutes to 1.5 minutes shortly after the application of the current, with a return to 3.5 minutes at the next test.
Operative procedures performed under light anesthesia (i. e., with the more persistent reflexes still present), or reduction of anesthesia soon after operation, resulted in a remarkable shortening of the coagulation time:
Nov. 8.—A cat was etherized and tracheotomized. The abdomen was then opened and a ligature was drawn around the hepatic nerves. The[p. 175] operation was completed at 2.25. At 2.50 the etherization became light and the rate of clotting began to be faster:
| 2.50 | 6 | minutes |
| 3.00 | 5.5 | “ |
| .10 | 3.5 | “ |
| .15 | 3.5 | “ |
| .20 | 4.5 | “ |
| .30 | 7.5 | “ |
Nov. 11.—A female cat, very quiet, was placed in the holder at 1.55. The animal was not excited. At 2.10 etherization was begun; the animal was then tracheotomized, and the femoral artery was exposed.
| 2.21 | 4.5 | minutes | |
| .26 | 4.5 | “ | Anesthesia lessened. |
| .32 | 3.5 | “ | “ light. |
| .35 | Abdomen opened. | ||
| .47 | 1.5 | minutes. | |
| .52 | 1 | “ | |
| .55 | Ligature passed around hepatic nerves. | ||
| .57 | 1.5 | minutes. | Anesthesia light; corneal reflex present. |
| 3.02 | 3 | “ | |
| .07 | 3 | “ | Some hepatic nerves cut. |
| .12 | 4.5 | “ | Rest of hepatic nerves cut. |
| .22 | 5 | “ | |
The results of this experiment are shown graphically in Fig. 33.
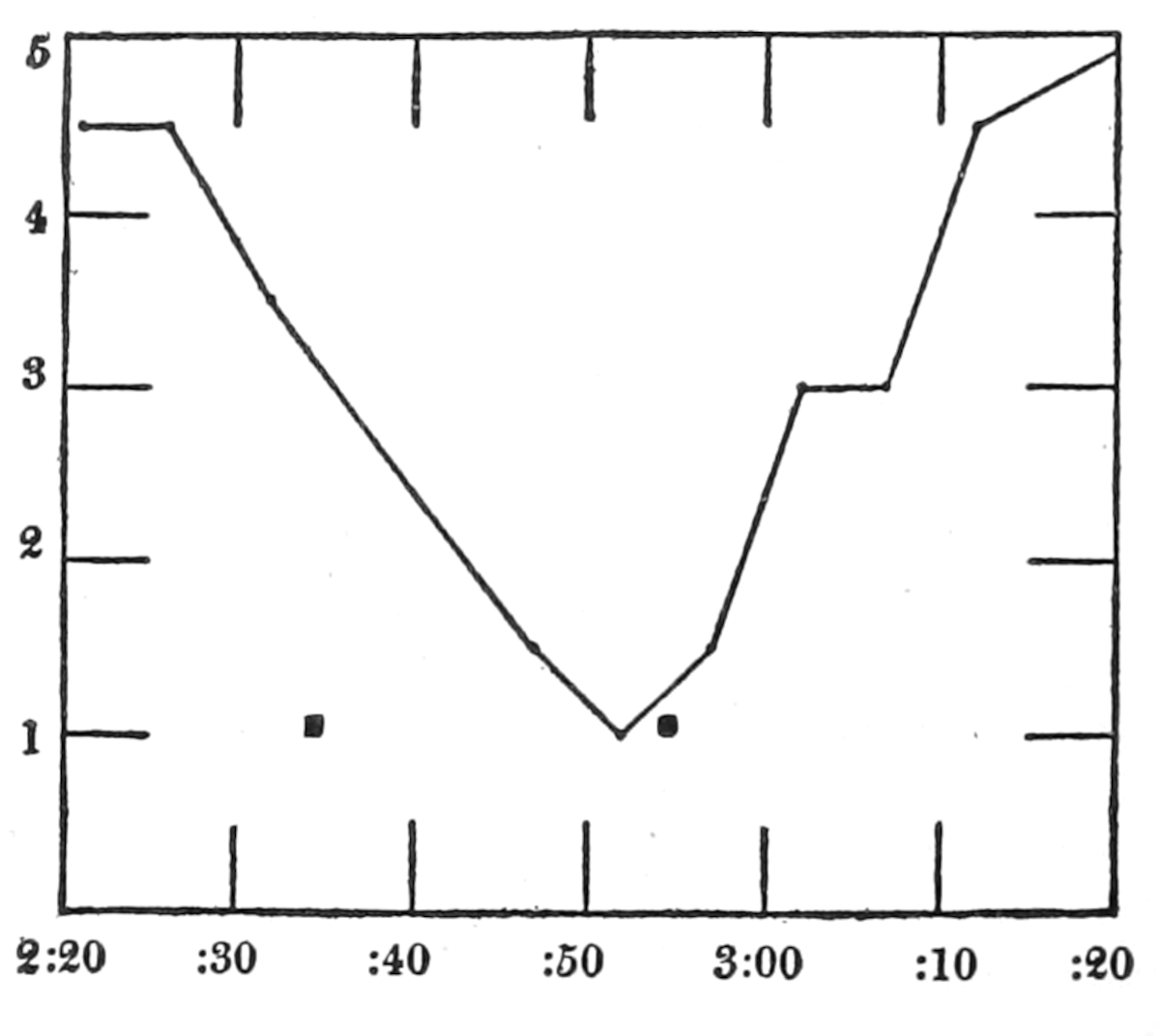
Nov. 13.—A cat was etherized at 1.55, tracheotomized, and the femoral artery laid bare. As soon as these preparations were completed, the ether was removed and anesthesia became light. The blood clotted thus:
[p. 176]
| 2.08 | 6 | minutes | |
| .15 | 4 | “ | Anesthesia light. |
| .20 | 2 | “ | |
| .24 | 1 | “ | Etherization begun again. |
| .27 | 2.5 | “ | |
| .30 | 3.5 | “ | |
| .35 | 5.5 | “ | |
| .50 | 5.5 | “ |
In the foregoing and in other similar instances, a condition of surgical injury, whether just made or being made, was accompanied by more rapid clotting of blood when the degree of anesthesia was lessened. This condition was one which, if allowed to go further in the same direction, would result in pain. Both direct electrical stimulation and also surgical operation of a nature to give pain in the unanesthetized animal result, therefore, in faster clotting.
[p. 177]
It is worthy of note that after decerebration clotting apparently occurred no faster because the abdomen had been opened, although in the decerebrate state etherization was suspended. The mechanism for reflex control of the adrenals may not be higher than the corpora quadrigemina, as Elliott has shown, but the discharge from the glands seems to be more certain to occur when the cerebrum is present and is permitted even slightly to operate.
The evidence for emotional secretion of the adrenal glands has already been presented. As was noted in my earlier observations on the motions of the alimentary canal (see p. 14), cats differ widely in their emotional reaction to being bound; some, especially young males, become furious; others, especially elderly females, take the experience quite calmly. This difference of attitude was used with positive results, the reader will recall, in the experiments on emotional glycosuria; there seemed a possibility likewise of using it to test the effect of emotions on blood clotting. To plan formal experiments for that purpose was not necessary, because in the ordinary course of the researches here reported, the difference in effects on the blood between the violent rage of vigorous young males and the quiet complacency of old females was early[p. 178] noted. Indeed, the rapid clotting which accompanied excitement not infrequently made necessary an annoying wait till slower clotting would permit the use of experimental methods for shortening the process.
The animals used on November 11 and 13 (see pp. 175, 176) are examples of calm acceptance of being placed on the holder; and furthermore, these animals were anesthetized without much disturbance. As the figures indicate, the clotting from the first occurred at about the average rate.
In sharp contrast to these figures are those obtained when a vigorous animal is angered:
Oct. 30.—A very vigorous cat was placed on the holder at 9.08. It at once became stormy, snarling, hissing, biting, and lashing its big tail. At 9.12 etherizing was begun and that intensified the excitement. By 9.15 the femoral artery was tied. The clotting time of the blood for an hour after the ether was first given was as follows:
| 9.18 | 0.5 | minute |
| .19 | 1 | “ |
| .22 | 1 | “ |
| .24 | 1 | “ |
| .26 | 1 | “ |
| .28 | 1.5 | “ |
| .31 | 1 | “ |
| .33 | 0.5 | “ |
| .35 | 0.5 | “ |
| .38 | 0.5 | “ |
| .39 | 0.5 | “ |
| .41 | 1 | “ |
| .43 | 1 | “ |
| .45 | 0.5 | “ |
| .49 | 0.5 | “ |
| .52 | 0.5 | “ |
| .54 | 0.5 | “ |
| .57 | 1 | “ |
| 10.00 | 0.5 | “ |
| .02 | 0.5 | “ |
| .06 | 1 | “ |
| .09 | 0.5 | “ |
| .11 | 0.5 | “ |
| .13 | 1 | “ |
[p. 179]
Twenty-four observations made during the hour showed that the clotting time in this enraged animal averaged three-fourths of a minute and was never longer than a minute and a half. The clots were invariably a solid jelly. The persistence of the rapid clotting for so long a period after anesthesia was started may have been in part due to continued, rather light, etherization, for Elliott[7] found that etherization itself could reduce the adrenin content of the adrenal glands.
The shortened clotting did not always persist so long as in the foregoing instance. The brief period of faster clotting illustrated in the following case was typical of many:
Nov. 18.—A cat that had been in stock for some time was placed on the holder at 2.13, and was at once enraged. Two minutes later etherization was started. The hairs on the tail were erect. The clotting was as follows:
| 2.25 | 1 | minute. |
| .27 | 0.5 | “ |
| .28 | 2 | “ |
| .31 | 4.5 | “ |
| .37 | 3.5 | “ |
| .47 | 4.5 | “ |
It seems probable that in this case just as in some of the cases in which the splanchnic nerves were stimulated (see p. 166), the adrenals had been well-nigh exhausted because of the cat’s being caged near dogs, and that the emotional flare-up practically discharged the glands, for repeated attempts[p. 180] later to reproduce the initial rapid clotting by stimulation of the splanchnic nerves were without result.
Evidence presented in previous chapters makes wholly probable the correctness of the inference that the faster coagulation which follows emotional excitement is due to adrenal discharge from splanchnic stimulation. In this relation the effect of severance of the splanchnics on emotional acceleration of the clotting process is of interest. The following cases are illustrative:
Oct. 29.—A cat was left on the holder for ten minutes while the femoral artery was uncovered under local anesthesia. The blood removed was clotted in a half-minute. The animal was much excited. It was now quickly etherized and the brain pithed forward from the neck. The tests resulted as follows:
| 10.51 | 1 | minute. |
| .53 | 0.5 | “ |
| .55 | 0.5 | “ |
| .57 | 0.5 | “ |
| 11.07 | Cut left splanchnic. | |
| .12 | “ right splanchnic. | |
| .21 | 3.5 | minutes. |
| .26 | 3.5 | “ |
The original record of this case is given in Fig. 34.
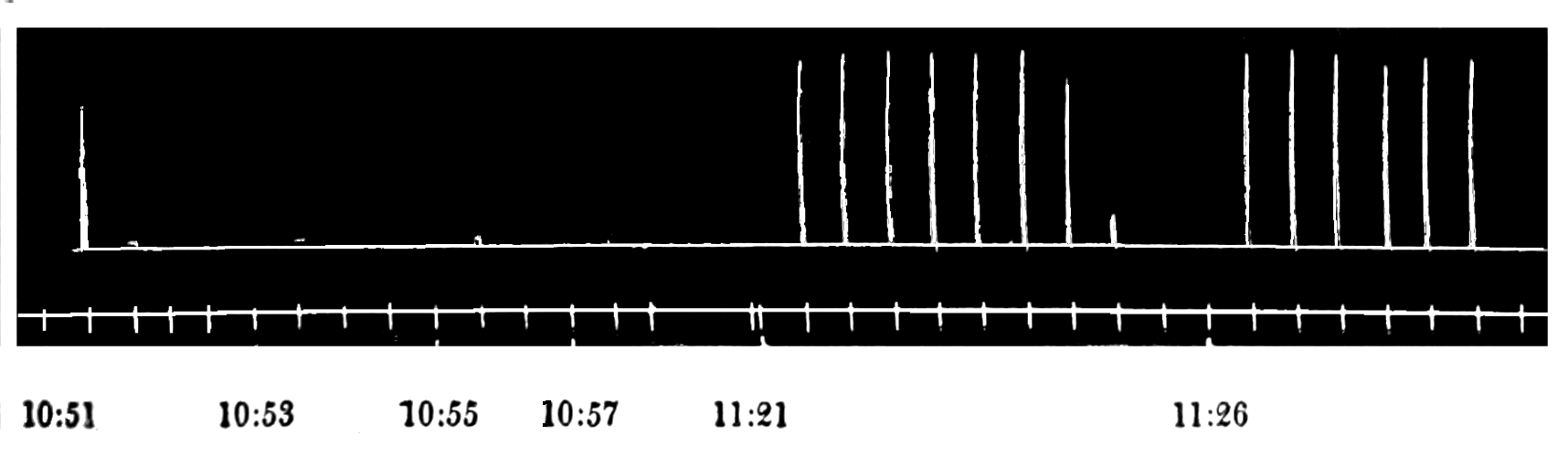
Nov. 5.—A cat was etherized at 2.35. At 2.39 artificial respiration by tracheal cannula was begun,[p. 181] the air passing through an ether bottle. The clotting occurred thus:
| 2.53 | 1.5 | minutes |
| .57 | 1.5 | “ |
| 3.05 | 1.5 | “ |
| .15 | 1.5 | “ |
| .25 | Both splanchnics cut and tied in thorax. | |
| .35 | 4.5 | minutes |
| .55 | 4.5 | “ |
Nov. 7.—A cat was etherized at 1.55 under excitement and with tail hairs erect. At 2.13 the animal was showing reflexes. The figures show the course of the experiment:
| 2.15 | 1.5 | minutes |
| .21 | 1 | “ |
| .26 | 1 | “ |
| .31 | 1 | “ |
| .36 | 1 | “ |
| .41 | 1 | “ |
| .46 | 2 | “ |
| .51 | 2 | “ |
| 3.06 | 2 | “ |
| .11 | 2.5 | “ |
| .26 | Cut left splanchnic in thorax. | |
| .35 | Cut right splanchnic in thorax. | |
| .40 | 5 | minutes |
| .45 | 5 | “ |
| .51 | 5.5 | “ |
[p. 182]
In this instance the subsequent stimulation of the splanchnic nerves resulted again in faster clotting—a reduction from 5.5 minutes to 3.5 minutes (see experiment Nov. 7, p. 164). The results from this experiment are expressed graphically in Fig. 35.
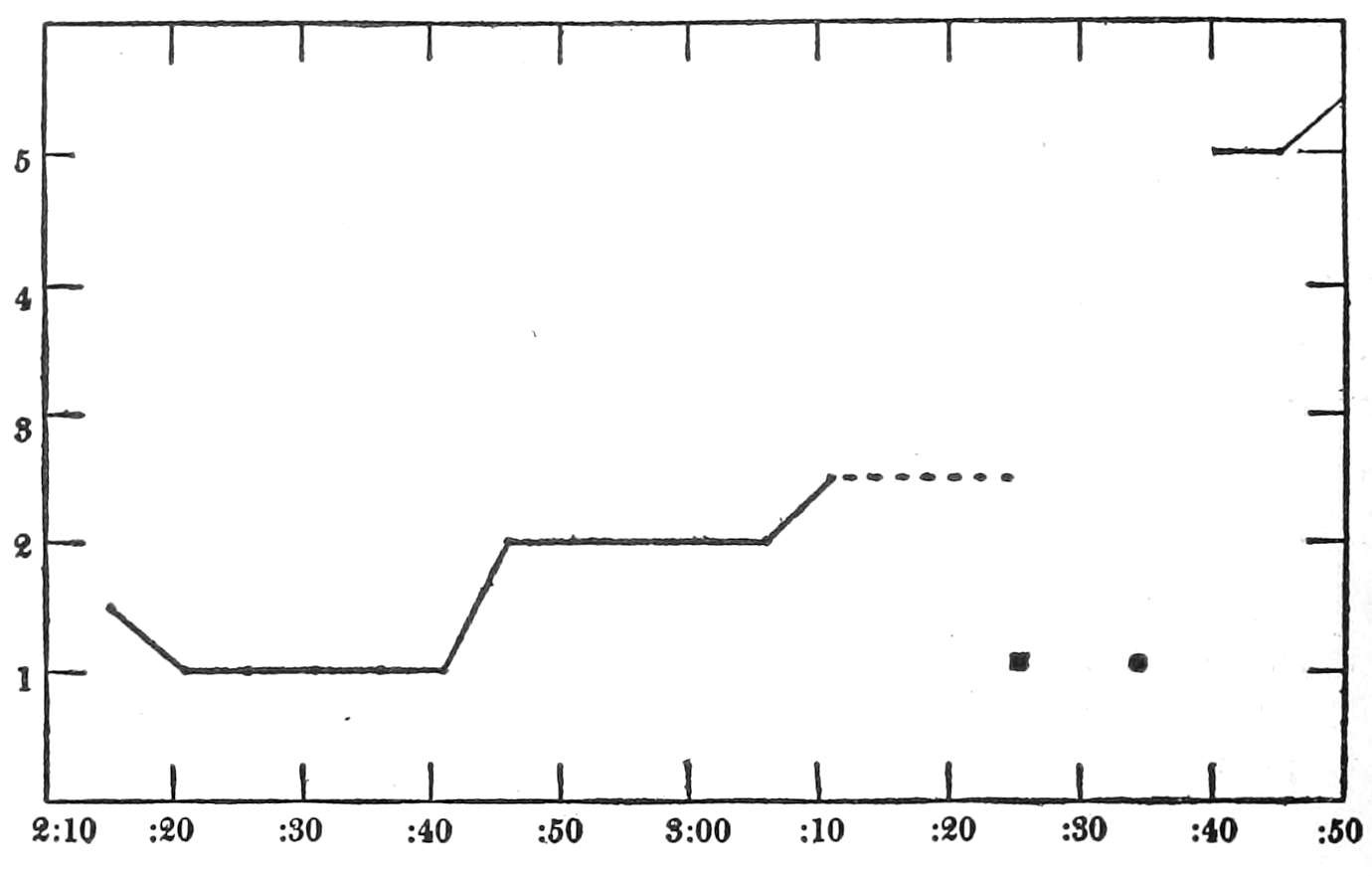
The data presented in this chapter show that such stimulation as in the unanesthetized animal would cause pain, and also such emotions as fear and rage, are capable of greatly shortening the coagulation time of blood. These results are quite in harmony with the evidence previously offered that injected adrenin and secretion from the adrenal glands induced by splanchnic stimulation hasten clotting, for painful stimulation and emotional[p. 183] excitement also evoke activity of the adrenals. Here, then, is another fundamental change in the body, a change tending to the conservation of its most important fluid, wrought through the adrenal glands in times of great perturbation. This bodily change and the others which occur under the same circumstances are next to be examined with reference to their significance.
1 Cannon and Mendenhall: American Journal of Physiology, 1914, xxxiv, p. 251.
2 Macleod: Diabetes: its Pathological Physiology, London, 1913, pp. 68–72.
3 Gautrelet and Thomas: Comptes Rendus, Société de Biologie, 1909, lxvii, p. 233.
4 Bang: Der Blutzucker, Wiesbaden, 1913, p. 87.
5 Elliott: Journal of Physiology, 1912, xliv, p. 379.
6 Elliott: Loc. cit., pp. 406, 407.
7 Elliott: Loc. cit., p. 388.
[p. 184]
THE UTILITY OF THE BODILY CHANGES IN PAIN AND GREAT EMOTION
We now turn from a consideration of the data secured in our experiments to an interpretation of the data. One of the most important lessons of experience is learning to distinguish between the facts of observation and the inferences drawn from those facts. The facts may remain unquestioned; the explanation, however, may be changed by additional facts or through the influence of more extensive views. Having given this warning, I propose to discuss the bearings of the results reported in the earlier chapters.
Our inquiry thus far has revealed that the adrenin secreted by the adrenal glands in times of stress has all the effects in the body that are produced by injected adrenin. It plays an essential rôle in calling forth stored carbohydrate from the liver, thus flooding the blood with sugar; it helps in distributing the blood to the heart, lungs, central nervous system and limbs, while taking it away[p. 185] from the inhibited organs of the abdomen; it quickly abolishes the effects of muscular fatigue; and it renders the blood more rapidly coagulable. These remarkable facts are, furthermore, associated with some of the most primitive experiences in the life of higher organisms, experiences common to all, both man and beast—the elemental experiences of pain and fear and rage that come suddenly in critical emergencies. What is the significance of these profound bodily alterations? What are the emergency functions of secreted adrenin?
The most significant feature of these bodily reactions in pain and in the presence of emotion-provoking objects is that they are of the nature of reflexes—they are not willed movements, indeed they are often distressingly beyond the control of the will. The pattern of the reaction, in these as in other reflexes, is deeply inwrought in the workings of the nervous system, and when the appropriate occasion arises, typical organic responses are evoked through inherent automatisms.
It has long been recognized that the most characteristic feature of reflexes is their “purposive” nature, or their utility either in preserving the[p. 186] welfare of the organism or in safeguarding it against injury. The reflexes of sucking, swallowing, vomiting and coughing, for instance, need only to be mentioned to indicate the variety of ways in which reflexes favor the continuance of existence. When, therefore, these automatic responses accompanying pain and fear and rage—the increased discharge of adrenin and sugar—are under consideration, it is reasonable to inquire first as to their utility.
Numerous ingenious suggestions have been offered to account for the more obvious changes accompanying emotional states—as, for example, the terrifying aspect produced by the bristling of the hair and the uncovering of the teeth in an access of rage.[1] The most widely applicable explanation proposed for these spontaneous reactions is that during the long course of racial experience they have been developed for quick service in the struggle for existence. Earlier writers on organic evolution pointed out the anticipatory character of these responses. According to Spencer,[2] “Fear, when strong, expresses itself in cries, in efforts to hide or escape, in palpitations and tremblings; and these are just the manifestations that would accompany an actual experience of the evil feared. The destructive passions are shown in a general tension of the muscular system, in gnashing of the teeth and protrusion of the[p. 187] claws, in dilated eyes and nostrils, in growls; and these are weaker forms of the actions that accompany the killing of prey.” McDougall[3] has developed this idea systematically and has suggested that an association has become established between peculiar emotions and peculiar instinctive reactions; thus the emotion of fear is associated with the instinct for flight, and the emotion of anger or rage with the instinct for fighting or attack. Crile[4] likewise in giving recent expression to the same view has emphasized the importance of adaptation and natural selection, operative through myriads of years of racial experience, in enabling us to account for the already channeled responses which we find established in our nervous organization. And on a principle of “phylogenetic association” he assumes that fear, born of innumerable injuries in the course of evolution, has developed into portentous foreshadowing of possible injury and has become, therefore, capable of arousing in the body all the offensive and defensive activities that favor the survival of the organism.
Because the increase of adrenin and the increase of sugar in the blood, following painful or strong emotional experiences, are reflex in character, and because reflexes as a rule are useful responses, we are justified in the assumption that under these circumstances these reactions are useful. What, then, is their possible value?
[p. 188]
In order that these reactions may be useful they must be prompt. Such is the case. Some observations made by one of my students, Mr. H. Osgood, show that the latent period of adrenal secretion, when the splanchnic nerve is stimulated below the diaphragm, is not longer than 16 seconds; and Macleod[5] states that within a few minutes after splanchnic stimulation the sugar in the blood rises between 10 and 30 per cent. The two secretions are, therefore, almost instantly ready for service.
Conceivably the two secretions might act in conjunction, or each might have its own function alone. Thus adrenin might serve in coöperation with nervous excitement to produce increase of blood sugar, or it might have that function and other functions quite apart from that. Before these possibilities are considered, however, the value of the increased blood sugar itself will be discussed.
When we were working on emotional glycosuria a clue to the significance of the increase of sugar in the blood was found in McDougall’s suggestion of a relation between “flight instinct” and “fear emotion,” and “pugnacity instinct” and “anger emotion.” And the point was made that, since the fear emotion and the anger emotion are, in[p. 189] wild life, likely to be followed by activities (running or fighting) which require contraction of great muscular masses in supreme and prolonged struggle, a mobilization of sugar in the blood might be of signal service to the laboring muscles. Pain—and fighting is almost certain to involve pain—would, if possible, call forth even greater muscular effort. “In the agony of pain almost every muscle of the body is brought into strong action,” Darwin[6] wrote, for “great pain urges all animals, and has urged them during endless generations, to make the most violent and diversified efforts to escape from the cause of suffering.”[*]
* It is recognized that both pain and the major emotions may have at times depressive rather than stimulating effects. For example, Martin and Lacey have shown (American Journal of Physiology, 1914, xxxiii, p. 212) that such stimuli as would induce pain may cause a fall of blood pressure, and they suggest that the rise of blood pressure commonly reported at times of painful experience is due to the psychic disturbance that is simultaneously aroused. Conceivably there is a relation between recognizing the possibility of escape (with the psychic consequences of that possibility) and the degree of stimulating effect. Thus pains originating from the interior of the body, or from injuries sure to be made more painful by action, would not likely lead to action. On the other hand, the whip and spur illustrate the well-known excitant effect of painful stimuli.
Similarly in the case of the strong emotions, the effect may be paralyzing until there is a definite deed to perform. Thus terror may be the most depressing of all emotions, but, as Darwin pointed out (Loc. cit., p. 81), “a man or animal driven through terror to desperation is endowed with wonderful strength, and is notoriously dangerous in the highest degree.”
[p. 190]
That muscular work is performed by energy supplied in carbonaceous material is shown by the great increase of carbon-dioxide output in severe muscular work, which may exceed twenty times the output during rest. Furthermore, the storage of glycogen in muscle, and the disappearance of this glycogen deposit from excised muscle stimulated to activity,[7] or its reduction after excessive contractions produced by strychnine,[8] and the lessened ability of muscles to work if their glycogen store has been reduced,[9] and the simple chemical relation between sugar and the lactic acid which appears when muscles are repeatedly made to contract, are all indications that carbohydrate (sugar and glycogen) is the elective source of energy for contraction. This conclusion is supported in recent careful studies by Benedict and Cathcart,[10] who have shown that a small but distinct increase in the ratio between the carbon-dioxide breathed out and the oxygen breathed in during a given period (the respiratory quotient) occurs during muscular work, and that a decrease in the quotient follows, thus pointing to a larger proportion of carbohydrate burned during muscular work than before or after—i. e., a call on the carbohydrate deposits of the body.
Whether circulating sugar can be immediately utilized by active muscles has been a subject of dispute. The claim of Chauveau and Kaufmann[p. 191][11] that a muscle uses about three and a half times as much blood sugar when active as when resting, although supported by Quinquaud,[12] and by Morat and Dufourt,[13] has been denied by Pavy,[14] who failed to find any difference between the sugar content of arterial and venous blood when the muscle was contracting; and also by Magnus-Levy,[15] who has estimated that the amount of change in sugar content of the blood passing through a muscle must be so slight as to be within the limits of the error of analysis. On the other hand, when blood or Ringer’s solution is repeatedly perfused through contracting heart muscle, the evidence is clear that the contained sugar may more or less completely disappear. Thus Locke and Rosenheim[16] found that from 5 to 10 centigrams of dextrose disappeared from Ringer’s solution repeatedly circulated through the rabbit heart for eight or nine hours. And recently Patterson and Starling[17] have shown that if blood is perfused repeatedly through a heart-lung preparation for three or four hours, and the heart is continually stimulated by adrenin added to the blood, the sugar in the blood wholly vanishes; or if the supply of sugar is maintained, the consumption may rise as high as 8 milligrams per gram of heart muscle per hour—about four times the usual consumption. When an animal is eviscerated it may be regarded as a preparation[p. 192] in which the muscles are perfused with their proper blood, pumped by the heart and oxygenated by the lungs. Under these circumstances, the percentage of sugar in the blood steadily falls,[18] because the utilization by the tissues is not compensated for by further supply from the liver. Thus, although there may be doubt that analyses of sugar in the blood flowing into and out from an active muscle during a brief period can be accurate enough to prove a clear difference, the evidence from the experiments above cited shows that when the supply of sugar is limited it disappears to a greater or less degree when passed repeatedly through muscular organs.
The argument may be advanced, of course, that the sugar which thus disappears is not directly utilized, but must first be changed to glycogen. There is little basis for this assumption. There is, on the other hand, considerable evidence that increasing the blood sugar does, in fact, directly increase muscular efficiency. Thus Locke[19] proved that if oxygenated salt solution is perfused through the isolated rabbit heart, the beats begin to weaken after one or two hours; but if now 0.1 per cent dextrose is added to the perfusing liquid, the beats at once become markedly stronger and may continue with very slow lessening of strength as long as seven hours. And Schumberg[20] noted that when he performed a large amount of general[p. 193] bodily work (thus using up blood sugar) and then tested flexion of the middle finger in an ergograph, the ability of the muscle was greater if he drank a sugar solution than if he drank an equally sweet solution of “dulcin.” He did not know during the experiment which solution he was drinking. These observations have been confirmed by Prantner and Stowasser, and by Frentzel.[21] In experiments on cats, Lee and Harrold[22] found that when sugar is removed from the animal by means of phlorhizin the tibialis anticus is quickly fatigued; but if, after the phlorhizin treatment, the animal is given an abundance of sugar and then submitted to the test, the muscle shows a much larger capacity for work. All this evidence is, of course, favorable to the view that circulating sugar may be quickly utilized by contracting muscles.
From the experimental results presented above it is clear that muscles work preferably by utilizing the energy stored in sugar, that great muscular labor is capable of considerably reducing the quantity of stored glycogen and of circulating sugar, and that under circumstances of a lessened sugar content the increase of blood sugar considerably augments the ability of muscles to continue contracting. The conclusion seems justified, therefore, that the increased blood sugar attendant on the major emotions and pain would be of direct[p. 194] benefit to the organism in the strenuous muscular efforts involved in flight or conflict or struggle to be free.
The function which the discharged adrenin itself might have in favoring vigorous muscular contraction has already been suggested in the chapter on the effect of adrenin in restoring the irritability of fatigued muscle. Some of the earliest evidence proved that removal of the adrenal glands has a debilitating effect on muscular power, and that injection of adrenal extract has an invigorating effect. For these reasons it seemed possible that increased adrenal secretion, as a reflex result of pain or the major emotions, might act in itself as a dynamogenic factor in the performance of muscular work. It was on the basis of that possibility that Nice and I tested the effect of stimulating the splanchnic nerves (thus causing adrenal secretion), or injecting adrenin, on the contraction of the fatigued tibialis anticus. We found, as already described, that when arterial pressure was of normal height, and was prevented from rising in the legs while the splanchnic was being stimulated, there was a distinct rise in the height of contraction of the fatigued muscle. And we drew the inference that adrenin set free in the blood may[p. 195] operate favorably to the organism by preparing fatigued muscles for better response to the nervous discharges sent forth in great excitement.
This inference led to the experiments by Gruber, who examined the effects of minute amounts of adrenin (0.1 or 0.5 cubic centimeter, 1:100,000), and also of splanchnic stimulation, on the threshold stimulus of fatigued neuro-muscular and muscular apparatus. Fatigue, the reader will recall, raises the threshold not uncommonly 100 or 200 per cent, and in some instances as much as 600 per cent. Rest will restore the normal threshold in periods varying from fifteen minutes to two hours, according to the length of previous stimulation. If a small dose of adrenin is given, however, the normal threshold may be restored in three to five minutes.
From the foregoing evidence the conclusion is warranted that adrenin, when freely liberated in the blood, not only aids in bringing out sugar from the liver’s store of glycogen, but also has a remarkable influence in quickly restoring to fatigued muscles, which have lost their original irritability, the same readiness for response which they had when fresh. Thus the adrenin set free in pain and in fear and rage would put the muscles of the body unqualifiedly at the disposal of the nervous system; the difficulty which nerve impulses might have in calling the muscles into full activity would[p. 196] be practically abolished; and this provision, along with the abundance of energy-supplying sugar newly flushed into the circulation, would give to the animal in which these mechanisms are most efficient the best possible conditions for putting forth supreme muscular efforts.[*]
* If these results of emotion and pain are not “worked off” by action, it is conceivable that the excessive adrenin and sugar in the blood may have pathological effects. (Cf. Cannon: Journal of the American Medical Association, 1911, lvi, p. 742.)
The only evidence opposed to the conclusion which has just been drawn is that which may be found in results recently reported by Wilenko. He injected adrenin into urethanized rabbits, usually one milligram per kilo body weight, and then found that the animals did not oxidize any part of an intravenous injection of glucose. Rabbits supplied with glucose in a similar manner, but not given adrenin, have an increased respiratory quotient. Wilenko[23] concluded, therefore, that adrenin lessens the capacity of the organism to burn carbohydrates. In a later paper he reported that adrenin, when added, with glucose, to physiological salt solution (Locke’s), and perfused through the isolated rabbit heart, notably increases the use of sugar by the heart (from 2.2–2.8 to 2.9–4.3 milligrams[p. 197] of glucose per gram of heart muscle per hour), but that the heart removed after the animal has received a subcutaneous injection of adrenin uses much less sugar, only 0.5–1.2 milligrams per gram per hour. From these results Wilenko[24] concludes that the glycosuria following injection of adrenin is the result of disturbance of the use of sugar—an effect which is not direct on the sugar-consuming organ, but indirect through action on some other organ.
Wilenko’s conclusion fails to account readily for the disappearance of glycogen from the liver in adrenin glycosuria. Furthermore, Lusk[25] has recently reported that the subcutaneous administration of adrenin (one milligram per kilo body weight) to dogs, simultaneously with 50 grams of glucose by mouth, interferes not at all with the use of the sugar—the respiratory quotient remains for several hours at 1.0; i. e., at the figure which glucose alone would have given. In other words, Lusk’s results with dogs are directly contradictory to Wilenko’s results with rabbits. Nevertheless, Wilenko’s conclusion might be quite true for the glycosuria produced by adrenin alone (which must be excessive), and yet have no bearing whatever on the glycosuria produced physiologically by splanchnic stimulation, even though some adrenin is thereby simultaneously liberated.
The amount of injected adrenin used to produce[p. 198] adrenin glycosuria is enormous. Osgood has studied in the Harvard Physiological Laboratory the effects on blood pressure of alternately stimulating the left splanchnic nerves (with the splanchnic vessels eliminated) and injecting adrenin, and by this method of comparison[26] has shown that the amount secreted after five seconds of stimulation varies between 0.0015 and 0.007 milligram. If 0.005 milligram is taken as a rather high average figure, and doubled (for two glands), the amount would be 0.01 milligram. To produce adrenin glycosuria, an animal weighing two kilos would be injected with two hundred times this amount. It is granted that more adrenin would be secreted if the nerves were stimulated longer than five seconds, and that with injection under the skin or into the abdominal cavity (to produce glycosuria), the amount of adrenin in the blood at one time would not be so great as if the injection were into a vein; but even with these concessions the amount of adrenin in the blood, when it has been injected to produce glycosuria, is probably very much above the amount following physiological stimulation of the glands.
Other evidence that the amount of adrenin discharged when the glands are stimulated is not so great as the amount needed to produce glycosuria when acting alone is presented in experiments by Macleod.[27] He found that if the nerve fibres[p. 199] to the liver were destroyed, stimulation of the splanchnic, which would cause increased adrenal secretion, did not increase the blood sugar. The increased blood sugar due to splanchnic stimulation, therefore, is a nervous effect, dependent, to be sure, on the presence of adrenin in the blood, but the amount of adrenin present is not in itself capable of evoking increase.
Furthermore, the increased blood sugar following splanchnic stimulation may long outlast the stimulation period. The adrenals, however, as has been demonstrated by Osgood, are soon fatigued, and fail to respond to repeated stimulation. They seem to be incapable of prolonged action.
Again, as Macleod[28] has shown, a rise in the sugar content of the blood can be induced, if the adrenals are intact, merely by stimulating the nerves going to the liver. The increased blood sugar of splanchnic origin, therefore, is not due to a disturbance of the use of sugar in the body, as Wilenko claims for the increase after adrenin injection, but is a result of a breaking down of the stored glycogen in the liver and is of nervous origin.
We may conclude, therefore, that since the conditions of Wilenko’s observations are not comparable with emotional conditions, his inferences are not pertinent to the present discussion; that when both adrenin and sugar are increased in the blood[p. 200] as a result of excitement, the higher percentage of sugar is not due to adrenin inhibiting the use of sugar by the tissues, and that there is no evidence at present to show that the brief augmentation of adrenal discharge, following excitement or splanchnic stimulation, affects in any deleterious manner the utilization of sugar as a source of energy. Indeed, the observation of Wilenko and of Patterson and Starling, above mentioned, that adrenin increases the use of sugar by the heart, may signify that a physiological discharge of the adrenals can have a favorable rather than an unfavorable effect on the employment of sugar by the tissues.
Quite in harmony with the foregoing argument that sugar and adrenin, which are poured into the blood during emotional excitement, render the organism more efficient in the physical struggle for existence, are the vascular changes wrought by increased adrenin, probably in coöperation with sympathetic innervations. The studies of volume changes of parts of the body, made by Oliver and Schäfer, have already been mentioned. Their observations, it will be remembered, showed that injected adrenin drove the blood from the abdominal viscera into the organs called upon in emergencies—into[p. 201] the central nervous system, the lungs, the heart, and the active skeletal muscles. The absence of effective vasoconstrictor nerves in the brain and the lungs, and the dilation of vessels in the heart and skeletal muscles during times of increased activity, make the blood supply to these parts dependent on the height of general arterial pressure. In pain and great excitement, as we have already noted, this pressure is likely to be much elevated, and consequently the blood flow through the unconstricted or actually dilated vessels of the body will be all the more abundant.
Adrenin has a well-known stimulating effect on the isolated heart—causing an increase both in the rate and the amplitude of cardiac contraction. This effect accords with the general rule that adrenin simulates the action of sympathetic impulses. It is commonly stated, however, that if the heart holds its normal relations in the body, adrenin causes slowing of the beat.[29] This view is doubtless due to the massive doses that have been employed, which are quite beyond physiological limits and which induce such enormous increases of arterial pressure that the natural influence of adrenin on heart muscle is overcome by mechanical obstacles to quick contractions and by inhibitory impulses from the central nervous system. Hoskins and Lovellette have recently shown that when the precaution is taken to inject adrenin into a vein[p. 202] in a manner resembling the discharge from the adrenal glands, not only is there increased blood pressure, but generally, also, an acceleration of the pulse.[30] At the same time, therefore, that a greater amount of work, from increased arterial pressure, is demanded of the heart, blood is delivered to the heart in greater abundance, and the muscle is excited to more rapid and vigorous pulsations. The augmentation of the heart beat is thus coördinate with the other adaptive functions of the adrenal glands in great emergencies.
The urgent need in struggle or flight is a generous supply of oxygen to oxidize the metabolites of muscular contraction, and a quick riddance of the resultant carbon-dioxide from the body. The moment vigorous exercise is begun the breathing at once changes so as to bring about a more thorough ventilation of the lungs. And one of the most characteristic reactions of animals in pain and emotional excitement is deep and rapid respiration. Again the reflex response is precisely what would be most serviceable to the organism in the strenuous efforts of fighting or escape that might accompany or follow distress or fear or rage. It is known that by such forced respirations the carbon-dioxide content of the blood can be so much reduced[p. 203] that the need for any breathing whatever may be deferred for as much as a minute or even longer.[31] And Douglas and Haldane[32] have found that moderately forced breathing for three minutes previous to severe muscular exertion results in greatly diminishing the subsequent respiratory distress, as well as lessening the amount of air breathed and the amount of carbon-dioxide given off. Furthermore, the heart beats less rapidly after the performance and returns more quickly from its increased rate to normal. The forced respirations in deeply emotional experiences can be interpreted, therefore, as an anticipatory reduction of the carbon-dioxide in the blood, a preparation for the augmented discharge of carbon-dioxide into the blood as soon as great muscular exertion begins.[*]
* The excessive production of heat in muscular work gives rise to sweating. The evaporation of sweat helps to keep the body temperature from rising unduly from the heat of exertion. Again in strong emotion and in pain the “cold sweat” that appears on the skin may be regarded as a reaction anticipatory of the strenuous muscular movements that are likely to ensue.
As the air moves to and fro in the lungs with each respiration, it must pass through the fine divisions of the air tubes or bronchioles. The bronchioles are provided with smooth muscle, which, in all probability, like smooth muscle elsewhere in the body, is normally held in a state of[p. 204] tonic contraction. When this tonic contraction is much increased, as in asthma, breathing becomes difficult, and even with the body at rest unusual effort is then required to maintain the minimal necessary ventilation of the lungs. During strenuous exertion, with each breath the air must rush through the bronchioles in greatly increased volume and speed. Thus in a well person “winded” with running, for example, the bronchioles might become relatively too small for the stream of air, just as they are too small in a person ill with asthma. And then some extra energy would have to be expended to force the air back and forth with sufficient rapidity to satisfy the bodily needs. It is probable that even under the most favorable conditions, the labored breathing in hard exercise involves to some degree the work of accelerating the tidal flow of the respiratory gases. This extra labor would obviously be reduced, if the tonic contraction of the ring-muscles in the wall of the bronchioles was reduced, so that the tubules were enlarged. It has been shown by a number of investigators, who have used various methods, that adrenin injected into the blood stream has as one of its precise actions the dilating of the bronchioles.[33] The adrenin discharged in emotional excitement goes to the lungs before entering into relation with any other organ except the right heart chamber; it may, therefore, have as its first[p. 205] effect the relaxation of the smooth muscles of the lungs. This would be another very direct means of rendering the organism more efficient when fierce struggle calls for a bounteous supply of fresh air and a speedy discharge of the carbonaceous waste.
All the bodily responses occurring in pain and emotional excitement have thus far been considered as anticipatory of the instinctive acts which naturally follow. And as we have seen, these responses can reasonably be interpreted as preparatory to the great exertions which may be demanded of the organism. This interpretation of the facts is supported by the discovery that a mechanism exists whereby the changes initiated in an anticipatory manner by emotional excitement are continued or perhaps augmented by the exertion itself.
Great exertion, such as might attend flight or conflict, would result in an excessive production of carbon-dioxide. Then, although respiratory and circulatory changes of emotional origin may have prepared the body for struggle, the emotional provisions for keeping the working parts at a high level of efficiency may not continue to operate, or[p. 206] they may not be adequate. If there is painful gasping for breath in the course of prolonged and vigorous exertion, or for a considerable period after the work has ceased, a condition of partial asphyxia has evidently been induced. This condition, as everyone knows, is distinctly unfavorable to further effort. But the asphyxia itself may act as a stimulus.[34]
In our examination of the influence of various conditions on the secretion of the adrenal glands, Hoskins and I[35] tested the effects of asphyxia. By use of the intestinal segment as an indicator we compared the action of blood, taken as nearly simultaneously as possible from the vena cava above the adrenal vessels and from the femoral vein before asphyxia, with blood taken from the same sources after asphyxia had been produced. The femoral venous blood after passing the capillaries of the leg thus acted as a standard for the same blood after receiving the contribution of the adrenal veins. Asphyxia was caused by covering the tracheal cannula until respiration became labored and slow, but capable of recovery when air was admitted. It may be regarded, therefore, as not extreme.
The results of the degree of asphyxia above described are shown by graphic record in Fig. 36. Blood taken from the vena cava and from the femoral vein before asphyxia (“normal”) failed to[p. 207] cause inhibition of the contractions. Blood taken from the femoral vein after asphyxia produced almost the same effect as blood from the same vein before; asphyxia, therefore, had wrought no change demonstrable in the general venous flow. Blood taken from the vena cava after asphyxia had, on the contrary, an effect markedly unlike blood from the same region before (compare the record after 1 and after 7, Fig. 36)—it caused the[p. 208] typical inhibition which indicates the presence of adrenal secretion.[*]
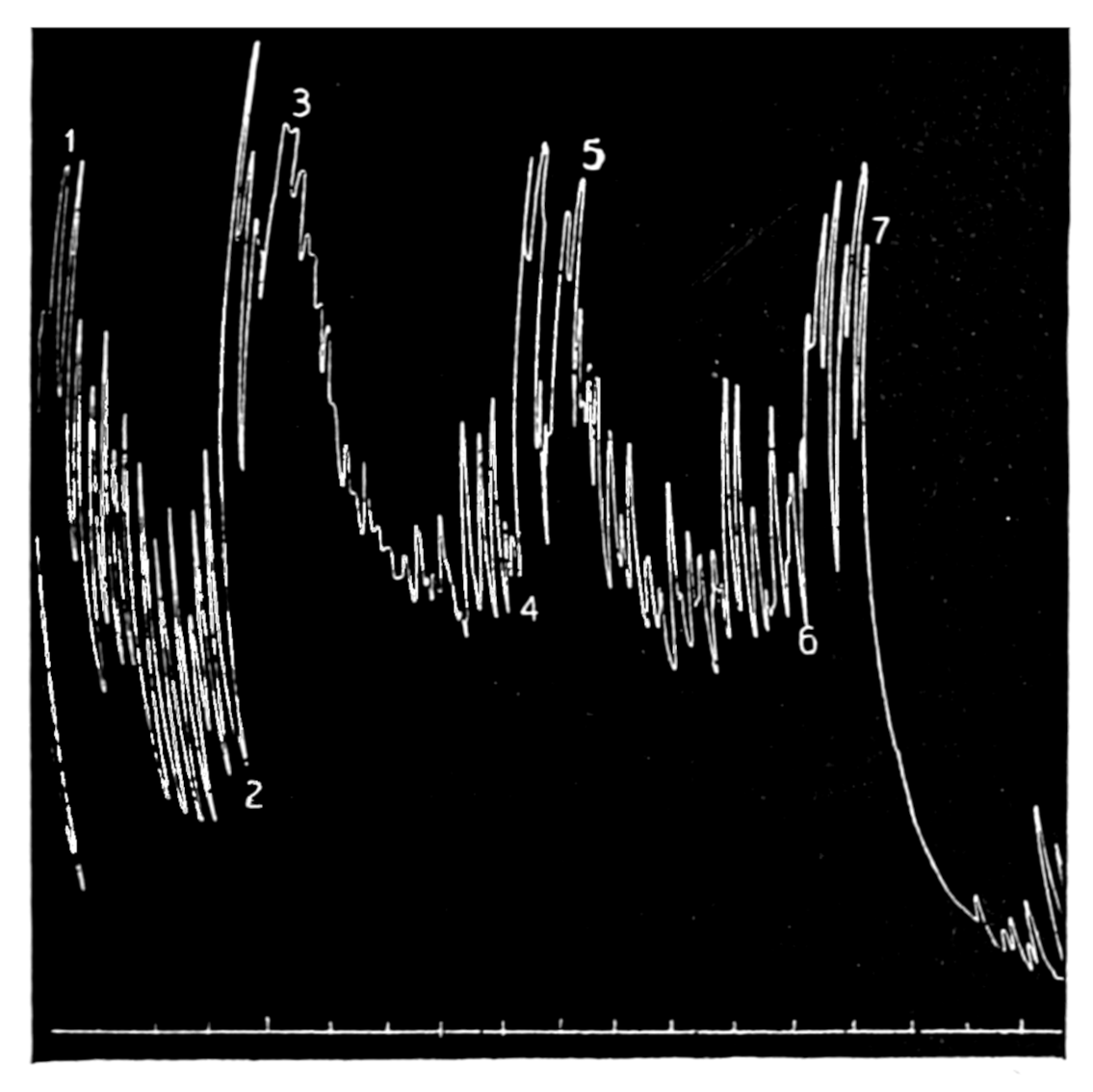
* This positive result might suggest that the comparison of both femoral and vena-cava blood under each condition was unnecessary, and that a comparison merely of vena-cava blood before and after asphyxia would be sufficient. Positive results were indeed thus secured, but they occurred even when the adrenal glands were carefully removed and extreme asphyxia (i. e., stoppage of respiration) was induced. That the blood may contain in extreme asphyxia a substance or substances capable of causing inhibition of intestinal contractions was thus demonstrated. In one instance, after the blood was proved free from adrenin, the aorta and vena cava were tied close below the diaphragm, and the carotids were tied about midway in the neck. Extreme asphyxia was produced (lasting five minutes). Blood now taken from the heart caused marked inhibition of the beating intestinal segment. Probably, therefore, the inhibitory action of blood taken from an animal when extremely asphyxiated cannot be due to adrenin alone.
That the positive result obtained in moderate asphyxia is not attributable to other agencies in the blood than adrenin is indicated by the failure of asphyxial femoral blood to cause inhibition, while vena-cava blood, taken almost simultaneously, brought about immediate relaxation of the muscle. The conclusion was drawn, therefore, that asphyxia results in increased secretion of the adrenal glands.
This conclusion has been supported by Borberg and Fridericia,[36] and also by Starkenstein,[37] who found that an increase of carbon-dioxide in the blood lessens the adrenin in the adrenal medulla.[p. 209] And recently Czubalski[38] also has inferred, from the rise of blood pressure in asphyxia when the adrenals are intact and the absence of the rise if the adrenals are removed, that asphyxia sets free adrenin in the blood.
Asphyxia, like pain and excitement, not only liberates adrenin, but, as might be inferred from that fact, also mobilizes sugar.[39] And, furthermore, Starkenstein[40] has shown that the asphyxia due to carbon-monoxide poisoning is not accompanied by increased blood sugar if the adrenal glands have been removed.
In case strong emotions are followed by vigorous exertions, therefore, asphyxia is likely to result, and this will act in conjunction with the emotional excitement and pain, or perhaps in continuation of the influences of these states, to bring forth still more adrenal discharge and still further output of sugar from the liver. And these in turn would serve the laboring muscles in the manner already described. This suggestion is in accord with Macleod’s[41] that the increased freeing of glycogen from the liver produced by muscular exercise is possibly associated with increased carbon-dioxide in the blood. And it also harmonizes with Zuntz’s statement[42] that the asphyxia of great physical exertion may call out sugar to such a degree that, in spite of the increased use of it in the active muscles, glycosuria may ensue.
[p. 210]
The evidence previously adduced that adrenin causes relaxation of the smooth muscle of the bronchioles, taken in conjunction with the evidence that adrenal secretion is liberated in asphyxia, suggests that relief from difficult breathing may thus be automatically provided for in the organism. The well-known phenomenon of “second wind” is characterized by an almost miraculous refreshment and renewal of vigor, after an individual has persisted in violent exertion in spite of being “out of breath.” It seems not improbable that this phenomenon, for which many explanations have been offered, is really due to setting in operation the supporting mechanism which, as we have seen, plays so important a rôle in augmenting bodily vigor in emotional excitement. The release of sugar and adrenin, the abundance of blood flow through the muscles—supplying energy and lessening fatigue—and the relaxation of the bronchiolar walls, are all occurrences which may reasonably be regarded as resulting from asphyxia. And when they take place they doubtless do much to abolish the distress itself by which they were occasioned. According to this explanation “second wind” would consist in the establishment of the same group of bodily changes, leading to more efficient physical struggle, that are observed in pain and excitement.
[p. 211]
The increase of blood sugar, the secretion of adrenin, and the altered circulation in pain and emotional excitement have been interpreted in the foregoing discussion as biological adaptations to conditions in wild life which are likely to involve pain and emotional excitement, i. e., the necessities of fighting or flight. The more rapid clotting of blood under these same circumstances may also be regarded as an adaptive process, useful to the organism. The importance of conserving the blood, especially in the struggles of mortal combat, needs no argument. The effect of local injury in favoring the formation of a clot to seal the opened vessels is obviously adaptive in protecting the organism against hemorrhage. The injury that causes opening of blood vessels, however, is, if extensive, likely also to produce pain. And, as already shown, conditions producing pain increase adrenal secretion and hasten coagulation. Thus injury would be made less dangerous as an occasion for serious hemorrhage by two effects which the injury itself produces in the body—the local effect on clotting at the region of injury and the general effect on the speed of clotting wrought by reflex secretion of adrenin.
According to the argument here presented the strong emotions, as fear and anger, are rightly[p. 212] interpreted as the concomitants of bodily changes which may be of utmost service in subsequent action. These bodily changes are so much like those which occur in pain and fierce struggle that, as early writers on evolution suggested, the emotions may be considered as foreshadowing the suffering and intensity of actual strife. On this general basis, therefore, the bodily alterations attending violent emotional states would, as organic preparations for fighting and possible injury, naturally involve the effects which pain itself would produce. And increased blood sugar, increased adrenin, an adapted circulation and rapid clotting would all be favorable to the preservation of the organism that could best produce them.
1 See Darwin: Expression of Emotions in Man and Animals, New York, 1905, pp. 101, 117.
2 Spencer: Principles of Psychology, London, 1855.
3 McDougall: Introduction to Social Psychology, London, 1908, pp. 49, 59.
4 Crile: Boston Medical and Surgical Journal, 1910, clxiii, p. 893.
5 Macleod: Diabetes, etc., p. 80.
6 Darwin: Loc. cit., p. 72.
7 Nasse: Archiv für die gesammte Physiologie, 1869, ii, p. 106; 1877, xiv, p. 483.
8 Frentzel: Archiv für die gesammte Physiologie, 1894, lvi, p. 280.
9 Zuntz: Oppenheimer’s Handbuch der Biochemie, Jena, 1911, iv (first half), p. 841.
[p. 213]
10 Benedict and Cathcart: Muscular Work, a Metabolic Study, Washington, 1913, pp. 85–87.
11 Chauveau and Kaufmann: Comptes Rendus, Académie des Sciences, 1886, ciii, p. 1062.
12 Comptes Rendus, Société de Biologie, 1886, xxxviii, p. 410.
13 Morat and Dufourt: Archives de Physiologie, 1892, xxiv, p. 327.
14 Pavy: The Physiology of the Carbohydrates, London, 1894, p. 166.
15 Magnus-Levy: v. Noorden’s Handbuch der Pathologie des Stoffwechsels, 1906, i, p. 385.
16 Locke and Rosenheim: Journal of Physiology, 1907, xxxvi, p. 211.
17 Patterson and Starling: Journal of Physiology, 1913, xlvii, p. 143.
18 See Macleod and Pearce: American Journal of Physiology, 1913, xxxii, p. 192. Pavy and Siau: Journal of Physiology, 1903, xxix, p. 375. Macleod: American Journal of Physiology, 1909, xxiii, p. 278.
19 Locke: Centralblatt für Physiologie, 1900, xiv, p. 671.
20 Schumberg: Archiv für Physiologie, 1896, p. 537.
21 Frentzel: Archiv für Physiologie, 1899, Supplement Band, p. 145.
22 Lee and Harrold: American Journal of Physiology, 1900, iv, p. ix.
23 Wilenko: Biochemische Zeitschrift, 1912, xlii, p. 58.
24 Wilenko: Archiv für experimentelle Pathologie und Pharmakologie, 1913, lxxi, p. 266.
25 Lusk: Proceedings of the Society for Experimental Biology and Medicine, 1914, xi, p. 49. Also Lusk and Riche: Archives of Internal Medicine, 1914, xiii, p. 68.
26 See Elliott: Journal of Physiology, 1912, xliv, p. 376.
27 Macleod: Diabetes, etc., pp. 64–73.
28 Macleod: Diabetes, etc., pp. 68–72.
29 See Biedl: Die Innere Sekretion, 1913, i, p. 464.
30 Hoskins and Lovellette: Journal of the American Medical Association, 1914, lxiii, p. 317.
[p. 214]
31 See Haldane and Priestley: Journal of Physiology, 1905, xxxii, p. 255.
32 Douglas and Haldane: Journal of Physiology, 1909, xxxix, p. 1.
33 See Januschke and Pollak: Archiv für experimentelle Pathologie und Pharmakologie, 1911, lxvi, p. 205. Trendelenburg: Zentralblatt für Physiologie, 1912, xxvi, p. 1. Jackson: Journal of Pharmacology and Experimental Therapeutics, 1912, iv, p. 59.
34 Cf. Hoskins and McClure: Archives of Internal Medicine, 1912, x, p. 355.
35 Cannon and Hoskins: American Journal of Physiology, 1911, xxix, p. 275.
36 Borberg: Skandinavisches Archiv für Physiologie, 1913, xxviii, p. 125.
37 Starkenstein: Zeitschrift für experimentelle Pathologie und Therapie, 1911, x, p. 95.
38 Czubalski: Zentralblatt für Physiologie, 1913, xxvii, p. 580.
39 For evidence and for references to this literature, see Bang: Der Blutzucker, Wiesbaden, 1913, pp. 104–108.
40 Starkenstein: Loc. cit., p. 94.
41 Macleod: Diabetes, etc., p. 184.
42 Zuntz: Loc. cit., p. 854.
[p. 215]
THE ENERGIZING INFLUENCE OF EMOTIONAL EXCITEMENT
The close relation between emotion and muscular action has long been perceived. As Sherrington[1] has pointed out, “Emotion ‘moves’ us, hence the word itself. If developed in intensity, it impels toward vigorous movement. Every vigorous movement of the body ... involves also the less noticeable coöperation of the viscera, especially of the circulatory and respiratory. The extra demand made upon the muscles that move the frame involves a heightened action of the nutrient organs which supply to the muscles the material for their energy.” The researches here reported have revealed a number of unsuspected ways in which muscular action is made more efficient because of emotional disturbances of the viscera. Every one of the visceral changes that have been noted—the cessation of processes in the alimentary canal (thus freeing the energy supply for other parts); the shifting of blood from the[p. 216] abdominal organs, whose activities are deferable, to the organs immediately essential to muscular exertion (the lungs, the heart, the central nervous system); the increased vigor of contraction of the heart; the quick abolition of the effects of muscular fatigue; the mobilizing of energy-giving sugar in the circulation—every one of these visceral changes is directly serviceable in making the organism more effective in the violent display of energy which fear or rage or pain may involve.
That the major emotions have an energizing effect has been commonly recognized.[*] Darwin testified to having heard, “as a proof of the exciting nature of anger, that a man when excessively jaded will sometimes invent imaginary offences and put himself into a passion, unconsciously for the sake of reinvigorating himself; and,” Darwin[2] continues, “since hearing this remark, I have occasionally recognized its full truth.” Under the impulse of fear also, men have been known to achieve extraordinary feats of running and leaping. McDougall[3] cites the instance[p. 217] of an athlete who, when pursued as a boy by a savage animal, leaped over a wall which he could not again “clear” until he attained his full stature and strength. The very unusual abilities, both physical and mental, which men have exhibited in times of stress were dealt with from the psychological point of view by William James[4] in one of his last essays. He suggested that in every person there are “reservoirs of power” which are not ordinarily called upon, but which are nevertheless ready to pour forth streams of energy if only the occasion presents itself. These figurative expressions of the psychologist receive definite and concrete exemplification, so far as the physical exhibitions of power are concerned, in the highly serviceable bodily changes which have been described in the foregoing chapters.
* Russell (The Pima Indians, United States Bureau of Ethnology, 1908, p. 243) relates a tale told by the Indians to their children, in which an injured coyote was chasing some quails. “Finally the quails got tired,” according to the story, “but the coyote did not, for he was angry and did not feel fatigue.”
It would doubtless be incorrect to attempt to account for all the increased strength and tireless endurance, which may be experienced in periods of great excitement, on the basis of abundant supplies provided then for muscular contraction, and a special secretion for avoiding or abolishing the depressive influences of fatigue. Tremors, muscular twitchings, the assumption of characteristic attitudes, all indicate that there is an immensely augmented activity of the nervous system—an activity that discharges powerfully even into parts not directly concerned in struggle, as, for example,[p. 218] into the muscles of voice, causing peculiar cries or warning notes; into the muscles of the ears, drawing them back or causing them to stand erect, and into the small muscles about the lips, tightening them and revealing the teeth. The typical appearances of human beings, as well as lower animals, when in the grip of such deeply agitating emotions as fear and rage, are so well recognized as to constitute a primitive and common means of judging the nature of the experience through which the organism is passing. This “pattern” response of the nervous system to an emotion-provoking object or situation is probably capable of bringing into action a much greater number of neurones in the central nervous system than are likely to be concerned in even a supreme act of volition. The nervous impulses delivered to the muscles, furthermore, operate upon organs well supplied with energy-yielding material and well fortified by rapidly circulating blood and by secreted adrenin, against quick loss of power because of accumulating waste. Under such circumstances of excitement the performance of extraordinary feats of strength or endurance is natural enough.[*]
* If individual neurones obey the law of either supreme action or inaction, the “all-or-none law,” the only means of securing a graded response is through variation of the number of neurones engaged in action—the more, the greater the resulting manifestation of strength.
[p. 219]
In connection with the conception that strong emotion has a dynamogenic value, it is of interest to note that on occasions when great demands are likely to be placed on the neuro-muscular system in the doing of unusual labors, emotional excitement is not uncommonly an accompaniment. In order to emphasize points in the argument developed thus far, I propose to cite some examples of the association of emotional excitement with remarkable exhibitions of power or resistance to fatigue.
Already in an earlier account (see p. 75) I have mentioned finding sugar in the urine in approximately fifty per cent of a group of college football players after the most exacting game of the season’s play. As is well understood, such games are heralded far and wide, loyal supporters of each college may travel hundreds of miles to attend the contest, enthusiastic meetings of undergraduate students are held in each college to demonstrate their devotion to the team and their confidence in its prowess—indeed, the arguments for victory, the songs, the cheering, are likely to be so disturbing to the players, that before an important contest they are not infrequently removed from college surroundings in order to avoid being overwrought when the contest comes.
On the day of the contest the excitement is multiplied[p. 220] manyfold. There is practically a holiday in college and to a large extent in the city as well. The streets are filled with eager supporters of each team as the hosts begin to gather at the field. As many as 70,000 spectators may be present, each one tense and strongly partisan. The student bands lead the singing, by thousands of voices, of songs which urge to the utmost effort for the college; and, in anticipation, these songs also celebrate the victory.
Into the midst of that huge, cheering, yelling, singing, flag-waving crowd, the players are welcomed in a special outburst of these same demonstrations of enthusiasm. Soon the game begins. The position of every player is known, if not because of previous acquaintance and recognition, because card-diagrams give the information. Every important play is seen by the assembled thousands, and the player who makes it is at once announced to all, and is likely to be honored by his multitudinous college mates in a special cheer, ending in his name. Any player who, by infraction of the rules or failure to do his part, loses ground gained by his team is also known. The man who is “played out” in efforts to win for his team and college, and consequently has to leave the field, is welcomed to the side lines by acclamations suited for a great hero. In short, every effort is made, through the powerful incentives of censure[p. 221] and a flaunting recognition, to make each member of the team realize vividly his responsibility, both personal and as one of a group, for the supreme, all-important result—victory for his college.
This responsibility works tremendously on the emotions of the players. In the dressing room before a critical contest I have seen a “gridiron warrior,” ready in canvas suit, cleated shoes, and leather helmet, sitting grimly on a bench, his fists clenched, his jaws tight, and his face the color of clay. He performed wonderfully when the game began, and after it was over there was a large percentage of sugar in his urine! Probably no sport requires a more sustained and extreme display of neuro-muscular effort than American football. And from the foregoing description of the conditions that surround the contests it is easy to realize that they conspire to arouse in the players excitements which would bring forth very efficiently the bodily reserves for use in the fierce struggle which the game requires.
What is true of football is true, though perhaps to a less degree, of the racing sports, as running and rowing. Again great multitudes attend the events, the contests are followed closely from beginning to end, and as the goal is approached the cheering and cries for victory gather in volume and intensity as if arranged for a thrilling climax. The whole setting is most highly favorable to the[p. 222] dramatic development of an acme of excitement as the moment for the last desperate effort to win is put forth.
Dancing, which formed a significant feature of primitive rituals, has always been accompanied by exciting conditions, and not unusually was an exhibition of remarkable endurance. In the transfer of the Ark to Zion there were processions and sacrifices, and King David “danced before the Lord with all his might.” Mooney[5] in his account of dances among the American Indians tells of a young man who in one of the ceremonials danced three days and nights without food, drink or sleep. In such a terrible ordeal the favoring presence of others, who through group action help to stimulate both the excitement and the activities, must be an important element in prolonging the efforts of the individual.
In the history of religious manias[6] there are many instances of large numbers of people becoming frenzied and then showing extraordinary endurance while dancing. In 1374 a mania broke forth in Germany, the Netherlands and France, in which the victims claimed to dance in honor of Saint John. Men and women went about dancing hand in hand, in pairs, or in a circle, on the streets, in the churches, at their homes, or wherever they[p. 223] might be, hour after hour without rest. While dancing they sang, uttered cries, and saw visions. Whole companies of these crazy fanatics went dancing along the public roads and into the cities, until they had to be interfered with.
In 1740 an extraordinary sect, known as the “Jumpers,” arose in Wales. According to the description given by Wesley, their exercises were not unlike those of certain frenzied states among the Indians. “After the preaching was over,” Wesley[7] wrote, “anyone who pleased gave out a verse of a hymn; and this they sung over and over again, with all their might and main, thirty or forty times, till some of them worked themselves into a sort of drunkenness or madness; they were then violently agitated, and leaped up and down in all manner of postures, frequently for hours together.” There were sometimes thousands at a single meeting of the Jumpers, shouting out their excitement and ready to leap for joy.[8] Wesley has also described instances of tremendous emotional outburst at Methodist meetings which he addressed. “Some were torn with a kind of convulsive motion in every part of their bodies, and that so violently that often four or five persons could not hold one of them. I have seen many hysterical or epileptic fits,” he wrote, “but none of them were like these in many respects.”
Among the dervishes[9] likewise the dance is accompanied[p. 224] by intense excitement and apparently tireless movements. “The cries of ‘Yâ Allah!’ are increased doubly, as also those of ‘Yâ Hoo!’ with frightful howlings shrieked by the dervishes together in the dance.”... “There was no regularity in their dancing, but each seemed to be performing the antics of a madman; now moving his body up and down; the next moment turning round, then using odd gesticulations with his arms, next jumping, and sometimes screaming.”... “At the moment when they would seem to stop from sheer exhaustion the sheikh makes a point of exciting them to new efforts by walking through their midst, making also himself most violent movements. He is next replaced by two elders, who double the quickness of the step and the agitation of the body; they even straighten themselves up from time to time, and excite the envy or emulation of others in their astonishing efforts to continue the dance until their strength is entirely exhausted.” Such is the frenzy thus developed that the performers may be subjected to severe pain, yet only show signs of elation.
In all these dances the two most marked features are the intense excitement of those who engage in them and the very remarkable physical endurance which they manifest. Although there is no direct evidence, such as was obtained in examining the football players, that bodily changes favorable to[p. 225] great neuro-muscular exertion are developed in these furies of fanaticism, it is highly probable that they are so developed, and that the feats of fortitude which are performed are to a large extent explicable on the basis of a “tapping of the reservoirs of power” through the emotional excitement.
Throughout the discussion of the probable significance of the bodily changes in pain and great emotion, the value of these changes in the struggles of conflict or escape was emphasized. In human beings as well as in lower animals the wildest passions are aroused when the necessities of combat become urgent. One needs only to glance at the history of warfare to observe that when the primitive emotions of anger and hatred are permitted full sway, men who have been considerate and thoughtful of their fellows and their fellows’ rights suddenly may turn into infuriated savages, slaughtering innocent women and children, mutilating the wounded, burning, ravaging, and looting, with all the wild fervor of demons. It is in such excesses of emotional turbulence that the most astonishing instances of prolonged exertion and incredible endurance are to be found.
Probably the fiercest struggles between men that are recorded are those which occurred when[p. 226] the wager of battle was a means of determining innocence or guilt. In the corners of the plot selected for the combat a bier was prepared for each participant, as a symbol that the struggle was for life or death. Each was attended by his relatives and followers, and by his father confessor.[10] After each had prayed to God for help in the coming combat, the weapons were selected, the sacrament was administered, and the battle was begun. The principals fought to the end with continuous and brutal ferocity, resembling the desperate encounters of wild beasts. A fairly illustrative example is furnished in an incident which followed the assassination of Charles the Good of Flanders in 1127. One of the accomplices, a knight named Guy, was challenged for complicity by another named Herman. Both were renowned warriors. Herman was speedily unhorsed by Guy, who with his lance frustrated all Herman’s attempts to remount. Then Herman disabled Guy’s horse, and the combat was renewed on foot with swords. Equally skilful in fence, they continued the struggle till fatigue compelled them to drop sword and shield, whereupon they wrestled for the mastery. Guy threw his antagonist, fell on him, and beat him in the face with his gauntlets till he seemed to be motionless; but Herman had quietly slipped his hand below the other’s coat of mail and, grasping the testicles, with a mighty effort wrenched them[p. 227] away. Immediately Guy fell over and expired.[11] In such terrific fights as these, conducted in the extremes of rage and hate, the mechanisms for reënforcing the parts of the body which are of primary importance in the struggle are brought fully into action and are of utmost value in securing victory.
It is noteworthy that in all the instances thus far cited—in the great games, in dancing, and in fighting—two factors are present that are well known to have an augmenting effect both in the full development of emotions and in the performance of unusual muscular labors. One of these is the crowd of witnesses or participants, who contribute the “mob spirit” that tends to carry the actions of the individual far beyond the limits set by any personal considerations or prudencies. The other is the influence of music. As Darwin long ago indicated, music has a wonderful power of recalling in a vague and indefinite manner strong emotions which have been felt by our ancestors in long-past ages. Especially is this true of martial music. For the grim purposes of war the reed and the lute are grotesquely ill-suited; to rouse men to action strident brass and the jarring instruments of percussion are used in full force. The influence of martial music on some persons[p. 228] is so profound as to cause the muscles to tremble and tears to come to the eyes—both indications of the deep stirring of emotional responses in the body. And when deeds of fortitude and fierce exertion are to be performed the effectiveness of such music in rousing the aggressive emotions has long been recognized. The Romans charged their foes amid the blasts of trumpets and horns. The ancient Germans rushed to battle, their forces spurred by the sounds of drums, flutes, cymbals and clarions. There is a tradition that the Hungarian troops are the worst in Europe, until their bands begin to play—then they are the best! The late General Linevitch is quoted as saying: “Music is one of the most vital ammunitions of the Russian army. Without music a Russian soldier would be dull, cowardly, brutal and inefficient. From music he absorbs a magic power of endurance, and forgets the sufferings and mortality. It is a divine dynamite.” And Napoleon is said to have testified that the weird and barbaric tunes of the Cossack regiments infuriated them to such rage that they wiped out the cream of his army.[12] A careful consideration of the use of martial music in warfare would perhaps bring further interesting evidence that its function is to reënforce the bodily changes that attend the belligerent emotions.
Only a few instances of the combination of extreme pain, rage, terror or excitement, and tremendous[p. 229] muscular power have been given in the preceding pages. Doubtless in numerous other conditions these two groups of phenomena occur together. In the lives of firemen and the police, in the experiences of escaping prisoners, of shipwrecked sailors, in the struggles between pioneers and their savage enemies, in accounts of forced marches or retreats, search would reveal many examples of such bodily disturbances as have been described in earlier chapters as augmenting the effectiveness of muscular efforts, and such exhibitions of power or endurance as are evidently far beyond the ordinary. There is every reason for believing that, were the conditions favorable to experimental testing, it would be possible to demonstrate and perhaps to measure the addition to the dynamics of bodily action that appears as the accompaniment of violent emotional disturbance.
In this connection it is highly significant that in times of strong excitement there is not infrequent testimony to a sense of overwhelming power that sweeps in like a sudden tide and lifts the person to a new high level of ability. A friend of mine, whose nature is somewhat choleric, has told me that when he is seized with anger, he is also possessed by an intense conviction that he could crush and utterly destroy the object of his hostility. And[p. 230] I have heard a football player confess that just before the final game such an access of strength seemed to come to him that he felt able, on the signal, to crouch and with a jump go crashing through any ordinary door. There is intense satisfaction in these moments of supreme elation, when the body is at its acme of accomplishment. And it is altogether probable that the critical dangers of adventure have a fascination because fear is thrilling, and extrication from a predicament, by calling forth all the bodily resources and setting them to meet the challenge of the difficulty, yields many of the joys of conquest. For these reasons vigorous men go forth to seek dangers and to run large chances of serious injury. “Danger makes us more alive. We so love to strive that we come to love the fear that gives us strength for conflict. Fear is not only something to be escaped from to a place or state of safety, but welcomed as an arsenal of augmented strength.”[13] And thus in the hazardous sports, in mountain climbing, in the hunting of big game, and in the tremendous adventure of war, risks and excitement and the sense of power surge up together, setting free unsuspected energies, and bringing vividly to consciousness memorable fresh revelations of the possibilities of achievement.
[p. 231]
1 Sherrington: The Integrative Action of the Nervous System, New York, 1906, p. 265.
2 Darwin: The Expression of Emotions in Man and Animals, New York, 1905, p. 79.
3 McDougall: Introduction to Social Psychology, London, 1908, p. 50.
4 James: The Energies of Men, p. 227, in Memories and Studies, New York, 1911.
5 Mooney: The Ghost-Dance Religion, United States Bureau of Ethnology, 1892–3, p. 924.
6 Schaff: Religious Encyclopedia, New York, 1908, iii, p. 346.
7 Southey: Life of Charles Wesley, New York, 1820, ii, p. 164.
8 Southey: Loc. cit., i, p. 240.
9 Brown: The Dervishes, London, 1868, pp. 218–222, 260.
10 Majer: Geschichte der Ordalien, Jena, 1796, pp. 258–261.
11 Lea: Superstition and Force, Philadelphia, 1892, p. 178.
12 Narodny: Musical America, 1914, xx, No. 14.
13 Hall: American Journal of Psychology, 1914, xxv, p. 154.
[p. 232]
THE NATURE OF HUNGER
On the same plane with pain and the dominant emotions of fear and anger, as agencies which determine the action of organisms, is the sensation of hunger. It is a sensation so peremptory, so disagreeable, so tormenting, that men have committed crimes in order to assuage it. It has led to cannibalism, even among the civilized. It has resulted in suicide. And it has defeated armies—for the aggressive spirit becomes detached from larger loyalties and turns personal and selfish as hunger pangs increase in vigor and insistence.
In 1905, while observing in myself the rhythmic sounds produced by the activities of the alimentary tract, I had occasion to note that the sensation of hunger was not constant but recurrent, and that the moment of its disappearance was often associated with a rather loud gurgling sound as heard through the stethoscope. This and other evidence, indicative of a source of the hunger sensations in[p. 233] the contractions of the digestive canal, I reported in 1911.[1] That same year, with the help of one of my students, A. L. Washburn, I obtained final proof for this inference.
The sensations of appetite and hunger are so complex and so intimately interrelated that any discussion of either sensation is sure to go astray unless at the start there is clear understanding of the meanings of the terms. The view has been propounded that appetite is the first degree of hunger, the mild and pleasant stage, agreeable in character; and that hunger itself is a more advanced condition, disagreeable and even painful—the unpleasant result of not satisfying the appetite.[2] On this basis appetite and hunger would differ only quantitatively. Another view, which seems more justifiable, is that the two experiences are fundamentally different.
Careful observation indicates that appetite is related to previous sensations of taste and smell of food. Delightful or disgusting tastes and odors, associated with this or that edible substance, determine the appetite. It has, therefore, important psychic elements in its composition. Thus, by taking thought, we can anticipate the odor of a delicious beefsteak or the taste of peaches and cream, and in that imagination we can find pleasure. In[p. 234] the realization, direct effects in the senses of taste and smell give still further delight. As already noted in the first chapter, observations on experimental animals and on human beings have shown that the pleasures of both anticipation and realization, by stimulating the flow of saliva and gastric juice, play a highly significant rôle in the initiation of digestive processes.
Among prosperous people, supplied with abundance of food, the appetite seems sufficient to ensure for bodily needs a proper supply of nutriment. We eat because dinner is announced, because by eating we avoid unpleasant consequences, and because food is placed before us in delectable form and with tempting tastes and odors. Under less easy circumstances, however, the body needs are supplied through the much stronger and more insistent demands of hunger.
The sensation of hunger is difficult to describe, but almost everyone from childhood has felt at times that dull ache or gnawing pain referred to the lower mid-chest region and the epigastrium, which may take imperious control of human actions. As Sternberg has pointed out, hunger may be sufficiently insistent to force the taking of food which is so distasteful that it not only fails to rouse appetite, but may even produce nausea. The hungry being gulps his food with a rush. The pleasures of appetite are not for him—he wants[p. 235] quantity rather than quality, and he wants it at once.
Hunger and appetite are, therefore, widely different—in physiological basis, in localization and in psychic elements. Hunger may be satisfied while the appetite still calls. Who is still hungry when the tempting dessert is served, and yet are there any who refuse it, on the plea that they no longer need it? On the other hand, appetite may be in abeyance while hunger is goading.[3] What ravenous boy is critical of his food? Do we not all know that “hunger is the best sauce”? Although the two sensations may thus exist separately, they nevertheless have the same function of leading to the intake of food, and they usually appear together. Indeed, the coöperation of hunger and appetite is probably the reason for their being so frequently confused.
Hunger may be described as having a central core and certain more or less variable accessories. The peculiar dull ache of hungriness, referred to the epigastrium, is usually the organism’s first strong demand for food; and when the initial order is not obeyed, the sensation is likely to grow into a highly uncomfortable pang or gnawing, less definitely localized as it becomes more intense. This may be regarded as the essential feature of[p. 236] hunger. Besides the dull ache, however, lassitude and drowsiness may appear, or faintness, or violent headache, or irritability and restlessness such that continuous effort in ordinary affairs becomes increasingly difficult. That these states differ much with individuals—headache in one and faintness in another, for example—indicates that they do not constitute the central fact of hunger, but are more or less inconstant accompaniments. The “feeling of emptiness,” which has been mentioned as an important element of the experience,[4] is an inference rather than a distinct datum of consciousness, and can likewise be eliminated from further consideration. The dull pressing sensation is left, therefore, as the constant characteristic, the central fact, to be examined in detail.
Hunger can evidently be regarded from the psychological point of view, and discussed solely on the basis of introspection; or it can be studied with reference to its antecedents and to the physiological conditions which accompany it—a consideration which requires the use of both objective methods and subjective observation. This psycho-physiological treatment of the subject will be deferred till the last. Certain theories which have been advanced with regard to hunger, and which have been given more or less credit, must first be examined.
Two main theories have been advocated. The[p. 237] first is supported by contentions that hunger is a general sensation, arising at no special region of the body, but having a local reference. This theory has been more widely credited by physiologists and psychologists than the other. The other is supported by evidence that hunger has a local source and therefore a local reference. In the course of our examination of these views we shall have opportunity to consider some pertinent new observations.
The conception that hunger arises from a general condition of the body rests in turn on the notion that, as the body uses up material, the blood becomes impoverished. Schiff[5] advocated this notion, and suggested that poverty of the blood in food substance affects the tissues in such manner that they demand a new supply. The nerve cells of the brain share in this general shortage of provisions, and because of internal changes, give rise to the sensation. Thus is hunger explained as an experience dependent on the body as a whole.
Three classes of evidence are cited in support of this view:
1. “Hunger increases as time passes”—a partial statement. The development of hunger as time passes is a common observation which quite accords with the assumption that the condition of the[p. 238] body and the state of the blood are becoming constantly worse, so long as the need, once established, is not satisfied.
While it is true that with the lapse of time hunger increases as the supply of body nutriment decreases, this concomitance is not proof that the sensation arises directly from a serious encroachment on the store of food materials. If this argument were valid we should expect hunger to become more and more distressing until death follows from starvation. There is abundant evidence that the sensation is not thus intensified; on the contrary, during continued fasting hunger, at least in some persons, wholly disappears after the first few days. Luciani,[6] who carefully recorded the experience of the faster Succi, states that after a certain time the hunger feelings vanish and do not return. And he tells of two dogs that showed no signs of hunger after the third or fourth day of fasting; thereafter they remained quite passive in the presence of food. Tigerstedt,[7] who also has studied the metabolism of starvation, declares that although the desire to eat is very great during the first day of the ordeal, the unpleasant sensations disappear early, and that at the end of the fast the subject may have to force himself to take nourishment. The subject, “J. A.,” studied by Tigerstedt and his co-workers,[8] reported that after the fourth day of fasting, he had no disagreeable feelings.
[p. 239]
Carrington,[9] after examining many persons who, to better their health, abstained from eating for different periods, records that “habit-hunger” usually lasts only two or three days and, if plenty of water is drunk, does not last longer than three days. Viterbi,[10] a Corsican lawyer condemned to death for political causes, determined to escape execution by depriving his body of food and drink. During the eighteen days that he lived he kept careful notes. On the third day the sensation of hunger departed, and although thereafter thirst came and went, hunger never returned. Still further evidence of the same character could be cited, but enough has already been given to show that after the first few days of fasting the hunger feelings may wholly cease. On the theory that hunger is a manifestation of bodily need, are we to suppose that, in the course of starvation, the body is mysteriously not in need after the third day, and that therefore the sensation of hunger disappears? The absurdity of such a view is obvious.
2. “Hunger may be felt though the stomach be full”—a selected alternative. Instances of duodenal fistula in man have been carefully studied, which have shown that a modified sensation of hunger may be felt when the stomach is full. A famous case described by Busch[11] has been repeatedly used as evidence. His patient, who lost nutriment through a duodenal fistula, was hungry[p. 240] soon after eating, and felt satisfied only when the chyme was restored to the intestine through the distal fistulous opening. As food is absorbed mainly through the intestinal wall, the inference is direct that the general bodily state, and not the local conditions of the alimentary canal, must account for the patient’s feelings.
A full consideration of the evidence from cases of duodenal fistula cannot so effectively be presented now as later. That in Busch’s case hunger disappeared while food was being taken is, as we shall see, quite significant. It may be that the restoration of chyme to the intestine quieted hunger, not because nutriment was thus introduced into the body, but because the presence of material altered the nature of gastro-intestinal activity. The basis for this suggestion will be given in due course.
3. “Animals may eat eagerly after section of their vagus and splanchnic nerves”—a fallacious argument. The third support for the view that hunger has a general origin in the body is derived from observations on experimental animals. By severance of the vagus and splanchnic nerves, the lower esophagus, the stomach and the small intestine can be wholly separated from the central nervous system. Animals thus operated upon nevertheless eat food placed before them, and may indeed manifest some eagerness for it.[12] How[p. 241] is this behavior to be accounted for—when the possibility of local stimulation has been eliminated—save by assuming a central origin of the impulse to eat?
The fallacy of this evidence, though repeatedly overlooked, is easily shown. We have already seen that appetite as well as hunger may lead to the taking of food. Indeed, the animal with all gastro-intestinal nerves cut may have the same incentive to eat that a well-fed man may have, who delights in the pleasurable taste and smell of food and knows nothing of hunger pangs. Even when the nerves of taste are cut, as they were in Longet’s experiments,[13] sensations of smell are still possible, as well as agreeable associations which can be roused by sight. More than fifty years ago Ludwig[14] pointed out that, even if all the nerves were severed, psychic reasons could be given for the taking of food, and yet because animals eat after one or another set of nerves is eliminated, the conclusion has been drawn by various writers that the nerves in question are thereby proved to be not concerned in the sensation of hunger. Evidently, since hunger is not required for eating, the act of eating is no testimony whatever that the animal is hungry, and, after the nerves have been severed, is no proof that hunger is of central origin.
[p. 242]
The evidence thus far examined has been shown to afford only shaky support for the theory that hunger is a general sensation. The theory, furthermore, is weak in its fundamental assumptions. There is no clear indication, for example, that the blood undergoes or has undergone any marked change, chemical or physical, when the first stages of hunger appear. There is no evidence of any direct chemical stimulation of the gray matter of the cerebral cortex. Indeed, attempts to excite the gray matter artificially by chemical agents have been without results;[15] and even electrical stimulation, which is effective, must, in order to produce movements, be so powerful that the movements have been attributed to excitation of underlying white matter rather than cells in the gray. This insensitivity of cortical cells to direct stimulation is not at all favorable to the notion that they are sentinels set to warn against too great diminution of bodily supplies.
Still further evidence opposed to the theory that hunger results directly from the using up of organic stores is found in patients suffering from fever. Metabolism in fever patients is augmented, body substance is destroyed to such a degree that[p. 243] the weight of the patient may be greatly reduced, and yet the sensation of hunger under these conditions of increased need is wholly lacking.
Again, if a person is hungry and takes food, the sensation is suppressed soon afterwards, long before any considerable amount of nutriment could be digested and absorbed, and therefore long before the blood and the general bodily condition, if previously altered, could be restored to normal.
Furthermore, persons exposed to privation have testified that hunger can be temporarily suppressed by swallowing indigestible materials. Certainly scraps of leather and bits of moss, not to mention clay eaten by the Otomacs, would not materially compensate for large organic losses. In rebuttal to this argument the comment has been made that central states as a rule can be readily overwhelmed by peripheral stimulation, and just as sleep, for example, can be abolished by bathing the temples, so hunger can be abolished by irritating the gastric walls.[16] This comment is beside the point, for it meets the issue by merely assuming as true the condition under discussion. The absence of hunger during the ravages of fever, and its quick abolition after food or even indigestible stuff is swallowed, still further weakens the argument, therefore, that the sensation arises directly from lack of nutriment in the body.
[p. 244]
Many persons have noted that hunger has a sharp onset. A person may be tramping in the woods or working in the fields, where fixed attention is not demanded, and without premonition may feel the abrupt arrival of the characteristic ache. The expression “grub-struck” is a picturesque description of this experience. If this sudden arrival of the sensation corresponds to the general bodily state, the change in the general bodily state must occur with like suddenness or have a critical point at which the sensation is instantly precipitated. There is no evidence whatever that either of these conditions occurs in the course of metabolism.
Another peculiarity of hunger, which I have already mentioned, is its intermittency. It may come and go several times in the course of a few hours. Furthermore, while the sensation is prevailing, its intensity is not uniform, but marked by ups and downs. In some instances the ups and downs change to a periodic presence and absence without change of rate. In my own experience the hunger pangs came and went on one occasion as follows:
| Came | Went |
|---|---|
| 12—37—20 | 38—30 |
| [p. 245]40—45 | 41—10 |
| 41—45 | 42—25 |
| 43—20 | 43—35 |
| 44—40 | 45—55 |
| 46—15 | 46—30 |
and so on, for ten minutes longer. Again in this relation, the intermittent and periodic character of hunger would require, on the theory under examination, that the bodily supplies be intermittently and periodically insufficient. During one moment the absence of hunger would imply an abundance of nutriment in the organism, ten seconds later the presence of hunger would imply that the stores had been suddenly reduced, ten seconds later still the absence of hunger would imply a sudden renewal of plenty. Such zig-zag shifts of the general bodily state may not be impossible, but from all that is known of the course of metabolism, such quick changes are highly improbable. The periodicity of hunger, therefore, is further evidence against the theory that the sensation has a general basis in the body.
The last objection to this theory is that it does not account for the most common feature of hunger—namely, the reference of the sensation to the region of the stomach. Schiff and others[17] who have supported the theory have met this[p. 246] objection by two contentions. First they have pointed out that the sensation is not always referred to the stomach. Schiff interrogated ignorant soldiers regarding the local reference; several indicated the neck or chest, twenty-three the sternum, four were uncertain of any region, and two only designated the stomach. In other words, the stomach region was most rarely mentioned.
The second contention against the importance of local reference is that such evidence is fallacious. An armless man may feel tinglings which seem to arise in fingers which have long since ceased to be a portion of his body. The fact that he experiences such tinglings and ascribes them to dissevered parts, does not prove that the sensation originates in those parts. And similarly the assignment of the ache of hunger to any special region of the body does not demonstrate that the ache arises from that region. Such are the arguments against a local origin of hunger.
Concerning these arguments we may recall, first, Schiff’s admission that the soldiers he questioned were too few to give conclusive evidence. Further, the testimony of most of them that hunger seemed to originate in the chest or region of the sternum cannot be claimed as unfavorable to a peripheral source of the sensation. The description of feelings which develop from disturbances within the body is almost always indefinite. As Head[p. 247][18] and others have shown, conditions in a viscus which give rise to sensation are likely not to be attributed to the viscus, but to related skin areas. Under such circumstances we do not dismiss the testimony as worthless merely because it may not point precisely to the source of the trouble. On the contrary, we use such testimony constantly as a basis for judging internal disorders.
With regard to the contention that reference to the periphery is not proof of the peripheral origin of a sensation, we may answer that the force of that contention depends on the amount of accessory evidence which is available. Thus if we see an object come into contact with a finger, we are justified in assuming that the simultaneous sensation of touch which we refer to that finger has resulted from the contact, and is not a purely central experience accidentally attributed to an outlying member. Similarly in the case of hunger—all that we need as support for the peripheral reference of the sensation is proof that conditions occur there, simultaneously with hunger pangs, which might reasonably be regarded as giving rise to those pangs.
With the requirement in mind that peripheral conditions be adequate, let us examine the state of the fasting stomach to see whether, indeed, conditions may be present in times of hunger which[p. 248] would sustain the theory that hunger has a local outlying source.
Among the suggestions which have been offered to account for a peripheral origin of the sensation is that of attributing it to emptiness of the stomach. By use of the stomach tube Nicolai[19] found that when his subjects had their first intimation of hunger the stomach was quite empty. But, in other instances, after lavage of the stomach, the sensation did not appear for intervals varying between one and a half and three and a half hours. During these intervals the stomach must have been empty, and yet no sensation was experienced. The same testimony was given long before by Beaumont,[20] who, from his observations on Alexis St. Martin, declared that hunger arises some time after the stomach is normally evacuated. Mere emptiness of the organ, therefore, does not explain the phenomenon.
A second theory, apparently suggested by observations on cases of hyperacidity, is that the ache or pang is due to the natural hydrochloric acid of the stomach but secreted while the organ is empty. Again the facts are hostile. Nicolai[21] reported[p. 249] that the gastric wash-water from his hungry subjects was neutral or only slightly acid. This testimony confirms Beaumont’s statement, and is in complete agreement with the results of gastric examination of fasting animals reported by numerous experimenters. There is no secretion into the empty stomach during the first days of starvation. Furthermore, persons suffering from absence of hydrochloric acid (achylia gastrica) declare that they have normal feelings of hunger. Hydrochloric acid cannot, therefore, be called upon to account for the sensation.
Another theory, which was first advanced by Beaumont,[22] is that hunger arises from turgescence of the gastric glands. The disappearance of the pangs as fasting continues has been accounted for by supposing that the gastric glands share in the general depletion of the body, and that thus the turgescence is relieved.[*] This turgescence theory has commended itself to several recent writers. Thus Luciani[23] has accepted it, and by adding the idea that nerves distributed to the mucosa are[p. 250] specially sensitive to deprivation of food he accounts for the hunger pangs. Also Valenti[24] declared a few years ago that the turgescence theory of Beaumont is the only one with a semblance of truth in it. The experimental work reported by these two investigators, however, does not necessarily sustain the turgescence theory. Luciani severed the previously exposed vagi after cocainizing them, and Valenti merely cocainized the nerves; the fasting dogs, eager to eat a few minutes previous to this operation, now ran about as before, but when offered food, licked and smelled it, but did not take it. This total neglect of the food lasted varying periods up to two hours. The vagus nerves seem, indeed, to convey impulses which affect the procedure of eating, but there is no clear evidence that those impulses arise from distention of the gland cells. The turgescence theory, moreover, does not explain the effect of taking indigestible material into the stomach. According to Pawlow, and to others who have observed human beings, the chewing and swallowing of unappetizing stuff does not cause any secretion of gastric juice (see p. 8). Yet such stuff when swallowed will cause the disappearance of hunger, and Nicolai found that the sensation could be abolished by simply introducing a stomach sound. It is highly improbable that the turgescence of the gastric glands can be reduced by either of these[p. 251] procedures. The turgescence theory, furthermore, does not explain the quick onset of hunger, or its intermittent and periodic character. That the cells are repeatedly swollen and contracted within periods a few seconds in duration is almost inconceivable. For these reasons, therefore, the theory that hunger results from turgescence of the gastric mucosa can reasonably be rejected.
* A better explanation perhaps is afforded by Boldireff’s discovery that at the end of two or three days the stomachs of fasting dogs begin to secrete gastric juice and continue the secretion indefinitely. (Boldireff, Archives Biologiques de St. Petersburg, 1905, xi, p. 98.)
There remain to be considered, as a possible cause of hunger pangs, contractions of the stomach and other parts of the alimentary canal. This suggestion is not new. Sixty-nine years ago Weber[25] declared his belief that “strong contraction of the muscle fibres of the wholly empty stomach, whereby its cavity disappears, makes a part of the sensation which we call hunger.” Vierordt[26] drew the same inference twenty-five years later (in 1871), and since then Ewald, Knapp, and Hertz have declared their adherence to this view. These writers have not brought forward any direct evidence for their conclusion, though Hertz has cited Boldireff’s observations on fasting dogs as probably accounting for what he terms “the gastric constituent of the sensation.”
The argument commonly used against the gastric contraction theory is that the stomach is not[p. 252] energetically active when empty. Thus Schiff[27] stated, “The movements of the empty stomach are rare and much less energetic than during digestion.” Luciani[28] expressed his disbelief by asserting that gastric movements are much more active during gastric digestion than at other times, and cease almost entirely when the stomach has discharged its contents. And Valenti[29] stated (1910), “We know very well that gastric movements are exaggerated while digestion is proceeding in the stomach, but when the organ is empty they are more rare and much less pronounced,” and, therefore, they cannot account for hunger.
Evidence opposed to these suppositions has been in existence for many years. In 1899 Bettmann[30] called attention to the contracted condition of the stomach after several days’ fast. In 1902 Wolff[31] reported that after forty-eight hours without food the stomach of the cat may be so small as to look like a slightly enlarged duodenum. In a similar circumstance I have noticed the same extraordinary smallness of the organ, especially in the pyloric half. The anatomist His[32] also recorded his observation of the phenomenon. In 1905 Boldireff[33] demonstrated that the whole gastro-intestinal tract has a periodic activity while not digesting. Each period of activity lasts from twenty to thirty minutes, and is characterized in the stomach by rhythmic contractions ten to twenty in number.[p. 253] These contractions, Boldireff reports, may be stronger than during digestion, and his published records clearly support this statement. The intervals of repose between periodic recurrences of the contractions lasted from one and a half to two and a half hours. Especially noteworthy is Boldireff’s observation that if fasting is continued for two or three days, the groups of contractions appear at gradually longer intervals and last for gradually shorter periods, and thereupon, as the gastric glands begin continuous secretion, all movements cease.
The research, previously mentioned, on the rhythmic sounds produced by the digestive process, I was engaged in when Boldireff’s paper was published. That contractions of the alimentary canal on a gaseous content might explain the hunger pangs which I had noticed seemed probable at that time, especially in the light of Boldireff’s observations. Indeed, Boldireff[34] himself had considered hunger in relation to the activities he described, but solely with the idea that hunger might provoke them; and since the activities dwindled in force and frequency as time passed, whereas, in his belief, they should have become more pronounced, he abandoned the notion of any relation[p. 254] between the phenomena. Did not Boldireff misinterpret his own observations? When he was considering whether hunger might cause the contractions, did he not overlook the possibility that the contractions might cause hunger? A number of experiences have led to the conviction that Boldireff did, indeed, fail to perceive part of the significance of his results. For example, I have noticed the disappearance of a hunger pang as gas was heard gurgling upward through the cardia. That the gas was rising rather than being forced downward was proved by its regurgitation immediately after the sound was heard. In all probability the pressure that forced the gas from the stomach was the cause of the preceding sensation of hunger. Again the sensation can be momentarily abolished a few seconds after swallowing a small accumulation of saliva or a teaspoonful of water. If the stomach is in strong contraction in hunger, this result can be accounted for, in accordance with the observations of Lieb and myself,[35] as due to the inhibition of the contraction by swallowing. Thus also could be explained the prompt vanishing of the ache soon after we begin to eat, for repeated swallowing results in continued inhibition.[*] Furthermore, Ducceschi’s discovery[36] that hydrochloric[p. 255] acid diminishes the tonus of the pyloric portion of the stomach may have its application here; the acid would be secreted as food is taken and would then cause relaxation of the very region which is most strongly contracted.
* The absence of hunger in Busch’s patient while food was being eaten (see p. 239) can also be accounted for in this manner.
Although the evidence above outlined had led me to the conviction that hunger results from contractions of the alimentary canal, direct proof was still lacking. In order to learn whether such proof might be secured, Washburn determined to become accustomed to the presence of a rubber tube in the esophagus.[*] Almost every day for several weeks Washburn introduced as far as the stomach a small tube, to the lower end of which was attached a soft-rubber balloon about 8 centimeters in diameter. The tube was thus carried about each time for two or three hours. After this preliminary experience the introduction of the tube and its presence in the gullet and stomach were not at all disturbing. When a record was to be taken, the balloon, placed just within the stomach, was moderately distended with air, and was connected with a water manometer ending in a cylindrical chamber 3.5 centimeters wide. A float recorder resting on[p. 256] the water in the chamber permitted registering any contractions of the fundus of the stomach. On the days of observation Washburn would abstain from breakfast, or eat sparingly; and without taking any luncheon would appear in the laboratory about two o’clock. The recording apparatus was arranged as above described. In order to avoid any error that might arise from artificial pressure on the balloon, a pneumograph, fastened below the ribs, was made to record the movements of the abdominal wall. Uniformity of these movements would show that no special contractions of the abdominal muscles were made. Between the records of gastric pressure and abdominal movement, time was marked in minutes, and an electromagnetic signal traced a line which could be altered by pressing a key. All these recording arrangements were out of Washburn’s sight; he sat with one hand at the key, ready whenever the sensation of hunger was experienced to make the current which moved the signal.
* Nicolai (loc. cit.) reported that although the introduction of a stomach tube at first abolished hunger in his subjects, with repeated use the effects became insignificant.
Sometimes the observations were started before any hunger was noted; at other times the sensation, after running a course, gave way to a feeling of fatigue. Under either of these circumstances there were no contractions of the stomach. When Washburn stated that he was hungry, however, powerful contractions of the stomach were invariably being registered. As in my own earlier experience,[p. 257] the sensations were characterized by periodic recurrences with free intervals, or by periodic accesses of an uninterrupted ache. The record of Washburn’s introspection of his hunger pangs agreed closely with the record of his gastric contractions. Almost invariably, however, the contraction nearly reached its maximum before the record of the sensation was started (see Fig. 37).
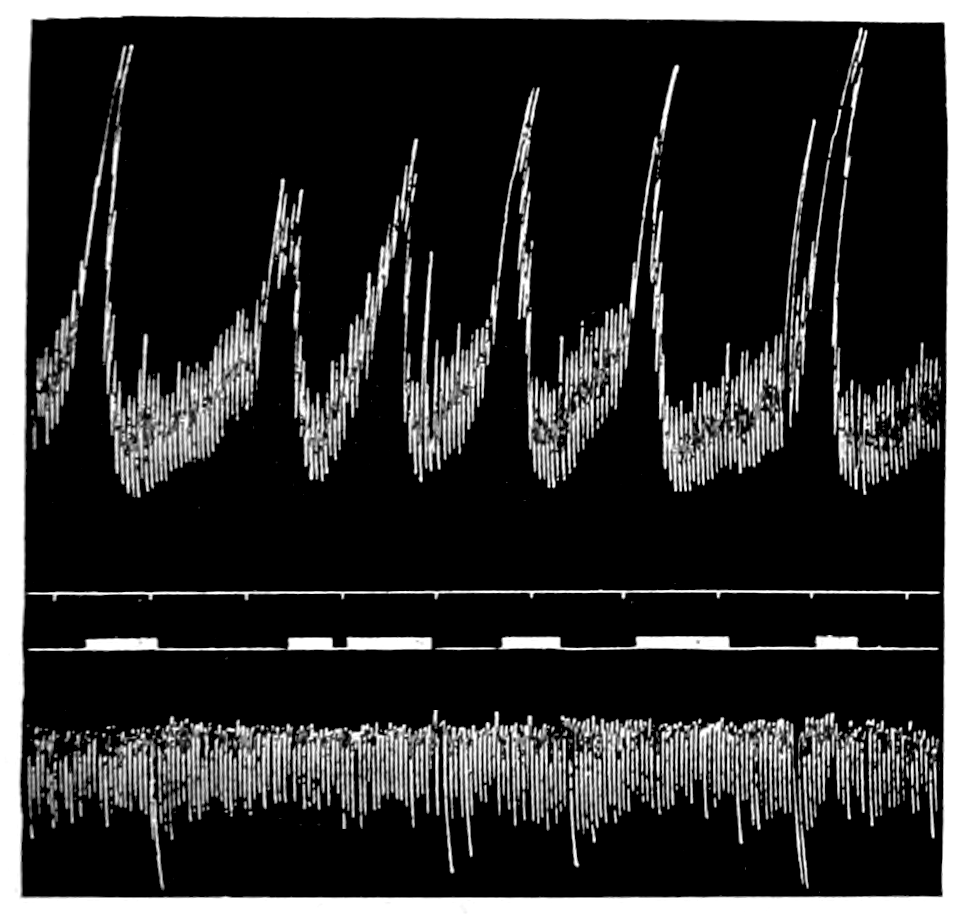
This fact may be regarded as evidence that the contraction precedes the sensation, and not vice versa, as Boldireff considered it. The contractions were about a half-minute in duration, and[p. 258] the intervals between varied from thirty to ninety seconds, with an average of about one minute. The augmentations of intragastric pressure in Washburn ranged between eleven and thirteen in twenty minutes; I had previously counted in myself eleven hunger pangs in the same time. The rate in each of us was, therefore, approximately the same. This rate is slightly slower than that found in dogs by Boldireff; the difference is perhaps correlated with the slower rhythm of gastric peristalsis in man compared with that in the dog.[37]
Before hunger was experienced by Washburn the recording apparatus revealed no signs of gastric activity. Sometimes a rather tedious period of waiting had to be endured before contractions[p. 259] occurred. And after they began they continued for a while, then ceased (see Fig. 38). The feeling of hunger, which was reported while the contractions were recurring, disappeared as the waves stopped. The inability of the subject to control the contractions eliminated the possibility of their being artifacts, perhaps induced by suggestion. The close concomitance of the contractions with hunger pangs, therefore, clearly indicates that they are the real source of those pangs.
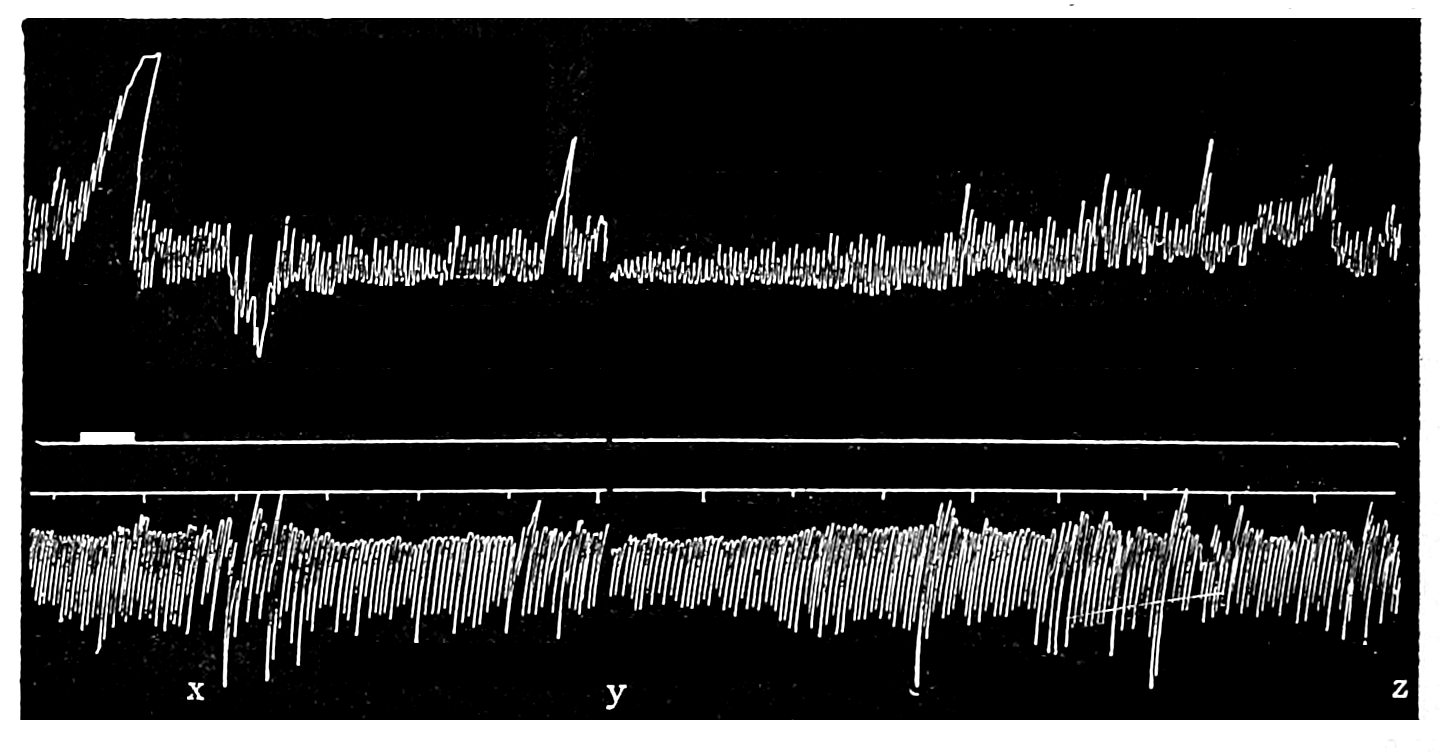
Boldireff’s studies proved that when the empty stomach is manifesting periodic contractions, the intestines also are active. Conceivably all parts of the alimentary canal composed of smooth muscle share in these movements. The lower esophagus in man is provided with smooth muscle. It was possible to determine whether this region in Washburn was active during hunger.
To the esophageal tube a thin-rubber finger-cot (2 centimeters in length) was attached and lowered into the stomach. The little rubber bag was distended with air, and the tube, pinched to keep the bag inflated, was gently withdrawn until resistance was felt. The air was now released from the bag and the tube farther withdrawn about 3 centimeters. The bag was again distended with air at a manometric pressure of 10 centimeters of water. Inspiration now caused the writing lever, which recorded the pressure changes, to rise; and a[p. 260] slightly farther withdrawal of the tube changed the rise, on inspiration, to a fall. The former position of the tube, therefore, was above the gastric cavity and below the diaphragm. In this position the bag, attached to a float recorder (with chamber 2.3 centimeters in diameter), registered the periodic oscillations shown in Fig. 39. Though individually more prolonged than those of the stomach, these contractions, it will be noted, occur at about the same rate.
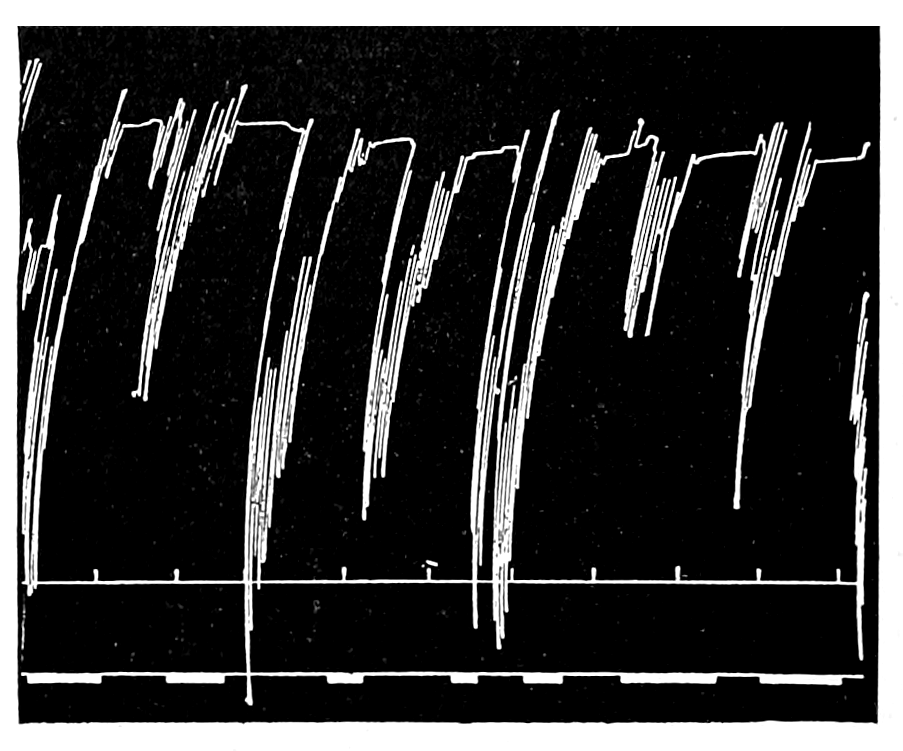
[p. 261]
This study of hunger, reported by Washburn and myself in 1912, has since been taken up by Carlson of Chicago, and in observations on a man with a permanent gastric fistula, as well as on himself and his collaborators, he has fully confirmed our evidence as to the relation between contractions of the alimentary canal and the hunger sensation. In a series of nearly a score of interesting papers, Carlson and his students[38] have greatly amplified our knowledge of the physiology of the “empty” stomach. Not only are there the contractions observed by Washburn and myself, but at times these may fuse into a continuous cramp of the gastric muscle. The characteristic contractions, furthermore, continue after the vagus nerve supply to the stomach has been destroyed, and, therefore, are not dependent on the reception of impulses by way of the cranial autonomic fibres. Recently Luckhardt and Carlson have brought forward evidence that the blood of a fasting animal if injected into the vein of a normal animal is capable of inducing in the latter the condition of cramp or tetanus in the gastric muscle mentioned above—an effect which does not occur when the blood of a well-fed animal is injected. It seems possible that a substance exists in the blood which acts to excite the gastric hunger mechanism. But this point will require further investigation.
With these demonstrations that contractions are[p. 262] the immediate cause of hunger, most of the difficulties confronting other explanations are readily obviated. Thus the sudden onset of hunger and its peculiar periodicity—phenomena which no other explanation of hunger can account for—are at once explained.
In fever, when bodily material is being most rapidly used, hunger is absent. Its absence is understood from an observation made by F. T. Murphy and myself,[39] that infection, with systemic involvement, is accompanied by a total cessation of all movements of the alimentary canal. Boldireff observed that when his dogs were fatigued the rhythmic contractions failed to appear. Being “too tired to eat” is thereby given a rational explanation.
A pathological form of the sensation—the inordinate hunger (bulimia) of certain neurotics—is in accordance with the well-known disturbances of the tonic innervation of the alimentary canal in such individuals.
Since the lower end of the esophagus, as well as the stomach, contracts periodically in hunger, the reference of the sensation to the sternum by the ignorant persons questioned by Schiff was wholly natural. The activity of the lower esophagus also explains why, after the stomach has been removed, or in some cases when the stomach is distended with food, hunger can still be experienced.[p. 263] Conceivably the intestines also originate vague sensations by their contractions. Indeed, the final banishment of the modified hunger sensation in the patient with duodenal fistula, described by Busch, may have been due to the lessened activity of the intestines when chyme was injected into them.
The observations recorded in this paper have, as already noted, numerous points of similarity to Boldireff’s observations[40] on the periodic activity of the alimentary canal in fasting dogs. Each period of activity, he found, comprised not only widespread contractions of the digestive canal, but also the pouring out of bile, and of pancreatic and intestinal juices rich in ferments. Gastric juice was not secreted at these times; when it was secreted and reached the intestine, the periodic activity ceased. What is the significance of this extensive disturbance? I have elsewhere presented evidence[41] that gastric peristalsis is dependent on the stretching of gastric muscle when tonically contracted. The evidence that the stomach is in fact strongly contracted in hunger—i. e., in a state of high tonus—has been presented above.[*] Thus[p. 264] the very condition which causes hunger and leads to the taking of food is the condition, when the swallowed food stretches the shortened muscles, for immediate starting of gastric peristalsis. In this connection the observations of Haudek and Stigler[42] are probably significant. They found that the stomach discharges its contents more rapidly if food is eaten in hunger than if not so eaten. Hunger, in other words, is normally the signal that the stomach is contracted for action; the unpleasantness of hunger leads to eating; eating starts gastric digestion, and abolishes the sensation. Meanwhile the pancreatic and intestinal juices, as well as bile, have been prepared in the duodenum to receive the oncoming chyme. The periodic activity of the alimentary canal in fasting, therefore, is not solely the source of hunger pangs, but is at the same time an exhibition in the digestive organs of readiness for prompt attack on the food swallowed by the hungry animal.
* The “empty” stomach and esophagus contain gas (see Hertz: Quarterly Journal of Medicine, 1910, iii, p. 378; Mikulicz: Mittheilungen aus den Grenzgebieten der Medicin und Chirurgie, 1903, xii, p. 596). They would naturally manifest rhythmic contractions on shortening tonically on their content.
1 Cannon: The Mechanical Factors of Digestion, London and New York, 1911, p. 204.
2 Bardier: Richet’s Dictionnaire de Physiologie, article Faim, 1904, vi, p. 1. See, also, Howell: Text-book of Physiology, fourth edition, Philadelphia and London, 1911, p. 285.
3 See Sternberg: Zentralblatt für Physiologie, 1909, xxii, p. 653. Similar views were expressed by Bayle in a thesis presented to the Faculty of Medicine in Paris in 1816.
4 See Hertz: The Sensibility of the Alimentary Canal, London, 1911, p. 38.
[p. 265]
5 Schiff: Physiologie de la Digestion, Florence and Turin, 1867, p. 40.
6 Luciani: Das Hungern, Hamburg and Leipzig, 1890, p. 113.
7 Tigerstedt: Nagel’s Handbuch der Physiologie, Berlin, 1909, i, p. 376.
8 Johanson, Landergren, Sonden and Tigerstedt: Skandinavisches Archiv für Physiologie, 1897, vii, p. 33.
9 Carrington: Vitality, Fasting and Nutrition, New York, 1908, p. 555.
10 Viterbi, quoted by Bardier: Loc. cit., p. 7.
11 Busch: Archiv für pathologische Anatomie und Physiologie und für klinische Medicin, 1858, xiv, p. 147.
12 See Schiff: Loc. cit., p. 37; also Ducceschi; Archivio di Fisiologia, 1910, viii, p. 579.
13 Longet: Traité de Physiologie, Paris, 1868, i, p. 23.
14 Ludwig: Lehrbuch der Physiologie des Menschen, Leipzig and Heidelberg, 1858, ii, p. 584.
15 Maxwell: Journal of Biological Chemistry, 1906–7, ii, p. 194.
16 See Schiff: Loc. cit., p. 49.
17 See Schiff: Loc. cit., p. 31; Bardier; Loc. cit., p. 16.
18 Head: Brain, 1893, xvi, p. 1; 1901, xxiv, p. 345.
19 Nicolai: Ueber die Entstehung des Hungergefühls, Inaugural Dissertation, Berlin, 1892, p. 17.
20 Beaumont: The Physiology of Digestion, second edition, Burlington, 1847, p. 51.
21 Nicolai: Loc. cit., p. 15.
22 Beaumont: Loc. cit., p. 55.
23 Luciani: Archivio di Fisiologia, 1906, iii, p. 54. Tiedemann long ago suggested that gastric nerves become increasingly sensitive as fasting progresses. (Physiologie des Menschen, Darmstadt, 1836, iii, p. 22.)
24 Valenti: Archives Italiennes de Biologie, 1910, liii, p. 94.
25 Weber: Wagner’s Handwörterbuch der Physiologie, 1846, iii2, p. 580.
26 Vierordt: Grundriss der Physiologie, Tübingen, 1871, p. 433.
27 Schiff: Loc. cit., p. 33.
[p. 266]
28 Luciani: Loc. cit., p. 542.
29 Valenti: Loc. cit., p. 95.
30 Bettmann: Philadelphia Monthly Medical Journal, 1899, i, p. 133.
31 Wolff: Dissertation, Giessen, 1902, p. 9.
32 His: Archiv für Anatomie, 1903, p. 345.
33 Boldireff: Loc. cit., p. 1.
34 Boldireff: Loc. cit., p. 96.
35 See Cannon and Lieb: American Journal of Physiology, 1911, xxix, p. 267.
36 Ducceschi: Archivio per le Scienze Mediche, 197, xxi, p. 154.
37 See Cannon: American Journal of Physiology, 1903, viii, p. xxi; 1905, xiv, p. 344.
38 See American Journal of Physiology, 1913, 1914.
39 Cannon and Murphy: Journal of the American Medical Association, 1907, xlix, p. 840.
40 Boldireff: Loc. cit., pp. 108–111.
41 Cannon: American Journal of Physiology, 1911, xxix, p. 250.
42 Haudek and Stigler: Archiv für die gesammte Physiologie, 1910, cxxxiii, p. 159.
[p. 267]
THE INTERRELATIONS OF EMOTIONS
Emotions gain expression through discharges along the neurones of the autonomic nervous system. The reader will recall that this system has three divisions—the cranial and sacral, separated by the sympathetic—and that when the neurones of the mid-division meet in any organ the neurones of either of the end divisions, the influence of the two sets is antagonistic. As previously stated (p. 35), there is evidence that arrangements exist in the central nervous system for reciprocal innervation of these antagonistic divisions, just as there is reciprocal innervation of antagonistic skeletal muscles. The characteristic affective states manifested in the working of these three divisions have been described. Undoubtedly, these states have correspondents—activities and inhibitions—in the central neurones. The question now arises, are the states which appear in opposed divisions also in opposition?
[p. 268]
The cranial autonomic, as already shown, is concerned with the quiet service of building up reserves and fortifying the body against times of stress. Accompanying these functions are the relatively mild pleasures of sight and taste and smell of food. The possibility of existence of these gentle delights of eating and drinking and also of their physiological consequences is instantly abolished in the presence of emotions which activate the sympathetic division. The secretion of saliva, gastric juice, pancreatic juice and bile is stopped, and the motions of the stomach and intestines cease at once, both in man and in the lower animals, whenever pain, fear, rage, or other strong excitement is present in the organism.
All these disturbances of digestion seem mere interruptions of the “normal” course of events unless the part they may play in adaptive reactions is considered. In discussing the operations of the sympathetic division, I pointed out that all the bodily changes which occur in the intense emotional states—such as fear and fury—occur as results of activity in this division, and are in the highest degree serviceable in the struggle for existence likely to be precipitated when these emotions are aroused. From this point of view these perturbations,[p. 269] which so readily seize and dominate the organs that in quiet times are commonly controlled by the cranial autonomic, are bodily reactions which may be of the utmost importance to life at times of critical emergency. Thus are the body’s reserves—the stored adrenin and the accumulated sugar—called forth for instant service; thus is the blood shifted to nerves and muscles that may have to bear the brunt of struggle; thus is the heart set rapidly beating to speed the circulation; and thus, also, are the activities of the digestive organs for the time abolished. Just as in war between nations the arts and industries which have brought wealth and contentment must suffer serious neglect or be wholly set aside both by the attacker and the attacked, and all the supplies and energies developed in the period of peace must be devoted to the present conflict; so, likewise, the functions which in quiet times establish and support the bodily reserves are, in times of stress, instantly checked or completely stopped, and these reserves lavishly drawn upon to increase power in the attack and in the defense or flight.[*]
* One who permits fears, worries and anxieties to disturb the digestive processes when there is nothing to be done, is evidently allowing the body to go onto what we may regard as a “war footing,” when there is no “war” to be waged, no fighting or struggle to be engaged in.
It is, therefore, the natural antagonism between these two processes in the body—between saving[p. 270] and expenditure, between preparation and use, between anabolism and catabolism—and the correlated antagonism of central innervations, that underlie the antipathy between the emotional states which normally accompany the processes. The desire for food, the relish of eating it, all the pleasures of the table, are naught in the presence of anger or great anxiety. And of the two sorts of emotional states, those which manifest themselves in the dominant division of the autonomic hold the field also in consciousness.
The nervi erigentes are the part of the sacral autonomic in which the peculiar excitements of sex are expressed. As previously stated, these nerves are opposed by branches from the sympathetic division—the division which is operated characteristically in the major emotions.
The opposition in normal individuals between the emotional states which appear in these two antagonistic divisions is most striking. Even in animals as low in the scale as birds, copulation is not performed “until every condition of circumstance and sentiment is fulfilled, until time, place and partner all are fit.”[1] And among men the effect of fear or momentary anxiety or any intense emotional interest in causing inhibition of the act can be supported[p. 271] by cases in the experience of any physician with extensive practice. Indeed, as Prince[2] has stated, “the suppression of the sexual instinct by conflict is one of the most notorious experiences of this kind in everyday life. This instinct cannot be excited during an attack of fear or anger, and even during moments of its excitation, if there is an invasion of another strong emotion the sexual instinct at once is repressed. Under these conditions, as with other instincts, even habitual excitants can no longer initiate the instinctive process.”
When the acme of excitement is approaching it is probable that the sympathetic division is also called into activity; indeed, the completion of the process—the contractions of the seminal vesicles and the prostate, and the subsidence of engorged tissues, all innervated by sympathetic filaments (see pp. 32, 33)—may be due to the overwhelming of sacral by sympathetic nervous discharges. As soon as this stage is reached the original feeling likewise has been dissipated.
The other parts of the sacral division which supply the bladder and rectum are so nearly free from any emotional tone in their normal reflex functioning that it is unnecessary to consider them further with reference to emotional antagonisms. Mild affective states, such as worry and anxiety, can, to be sure, check the activity of the colon and[p. 272] thus cause constipation.[3] But the augmented activity of these parts (contraction of the bladder and rectum) in very intense periods of emotional stress, when the sympathetic division is strongly innervated, presents a problem of some difficulty. Possibly in such conditions the orderliness of the central arrangements is upset, just as it is after tetanus toxin or strychnine poisoning, and opposed innervations no longer discharge reciprocally, but simultaneously, and then the stronger member of the pair prevails. Only on such a basis, at present, can I offer any explanation for the activity and the supremacy of the sacral innervation of the bladder and distal colon when the sympathetic innervation is aroused, as, for example, in great fright.
A summary in few words of the chief functions typically performed or supported by each division of the autonomic would designate the cranial division as the upbuilder and restorer of the organic reserves, the sacral as the servant of racial continuity, and the sympathetic as the preserver of the individual. Self-preservation is primary and essential; on that depends racial continuity, and for that all the resources of the organism are called forth. Analogously the sympathetic innervations, when they meet in organs innervated also by the cranial[p. 273] and sacral divisions, almost without exception predominate over their opponents. And analogously, also, the emotional states which are manifested in the sympathetic division and are characteristically much more intense than those manifested in the other divisions, readily assume ascendancy also in consciousness.
It is obvious that extended action of the sympathetic division, abolishing those influences of the cranial division which are favorable to proper digestion and nutrition, might defeat its own ends. Interruption of the nutritional process for the sake of self-preservation through defense or attack can be only temporary; if the interruption were prolonged, there might be serious danger to the vigor of the organism from failure to replenish the exhausted stores. The body does not have to depend on the return of a banished appetite, however, before its need for restoration is attended to. There is a secondary and very insistent manner in which the requirement of food is expressed, and that is through the repeated demands of hunger.
Unlike many other rhythmically repeated sensations, hunger is not one that anybody becomes accustomed to and neglects because of its monotony. During the period of his confinement in the citadel of Magdeburg, the celebrated political adventurer Baron von Trenck[4] was allowed only a pound and a half of ammunition bread and a jug of water[p. 274] as his daily ration. “It is impossible for me to describe to my reader,” he wrote in his memoirs, “the excess of tortures that during eleven months I endured from ravenous hunger. I could easily have devoured six pounds of bread every day; and every twenty-four hours, after having received and swallowed my small portion, I continued as hungry as before I began, yet I was obliged to wait another twenty-four hours for a new morsel.... My tortures prevented sleep, and looking into futurity, the cruelty of my fate seemed to me, if possible, to increase, for I imagined that the prolongation of pangs like these was insupportable. God preserve every honest man from sufferings like mine! They were not to be endured by the most obdurate villain. Many have fasted three days, many have suffered want for a week or more, but certainly no one besides myself ever endured it in the same excess for eleven months; some have supposed that to eat little might become habitual, but I have experienced the contrary. My hunger increased every day, and of all the trials of fortitude my whole life has afforded, this eleven months was the most bitter.”[*]
* In all probability the continued experience of hunger pangs reported by Baron von Trenck was due to the repeated eating of amounts of food too small to satisfy the bodily demand. The reader will recall that persons who for some time take no food whatever report that the disagreeable feelings are less intense or disappear after the third or fourth day (see p. 238).
[p. 275]
Thus, although the taking of food may be set in abeyance at times of great excitement, and the bodily reserves fully mobilized, that phase of the organism’s self-protecting adjustment is limited, and then hunger asserts itself as an agency imperiously demanding restoration of the depleted stores.
The dominant emotions which we have been considering as characteristically expressed in the sympathetic division of the autonomic system are fear and rage. These two emotions are not unlike. As James[5] has indicated, “Fear is a reaction aroused by the same objects that arouse ferocity.... We both fear and wish to kill anything that may kill us; and the question which of the two impulses we shall follow is usually decided by some one of those collateral circumstances of the particular case, to be moved by which is the mark of superior mental natures.” The cornering of an animal when in the headlong flight of fear may suddenly turn the fear to fury and the flight to a fighting in which all the strength of desperation is displayed.
Furthermore, these dominant emotions are states into which many other commonly milder affective states may be suddenly transformed. As McDougall[p. 276][6] has pointed out, all instinctive impulses when met with opposition or obstruction give place to, or are complicated by, the pugnacious or combative impulse directed against the source of the obstruction. A dog will bristle at any attempt to take away his food, males will fight furiously when provoked by interference with the satisfaction of the sexual impulse, a man will forget the conventions and turn hot for combat when there is imputation against his honor, and a mother all gentle with maternal devotion is stung to quick resentment and will make a fierce display of her combative resources, if anyone intentionally injures her child. In these instances of thwarted or disturbed instinctive acts the emotional accompaniments—such as the satisfaction of food and of sexual affection, the feeling of self-pride, and the tender love of a parent—are whirled suddenly into anger. And anger in one is likely to provoke anger or fear in the other who for the moment is the object of the strong feeling of antagonism. Anger is the emotion preëminently serviceable for the display of power, and fear is often its counterpart.
The visceral changes which accompany fear and rage are the result of discharges by way of sympathetic neurones. It will be recalled that these neurones are arranged for diffuse rather than for narrowly directed effects. So far as these two[p. 277] quite different emotions are concerned, present physiological evidence indicates that differences in visceral accompaniments[*] are not noteworthy—for example, either fear or rage stops gastric secretion (see pp. 10, 11). There is, indeed, obvious reason why the visceral changes in fear and rage should not be different, but rather, why they should be alike. As already pointed out, these emotions accompany organic preparations for action, and just because the conditions which evoke them are likely to result in flight or conflict (either one requiring perhaps the utmost struggle), the bodily needs in either response are precisely the same.
* Obvious vascular differences, as pallor or flushing of the face, are of little significance. With increase of blood pressure from vasoconstriction, pallor might result from action of the constrictors in the face, or flushing might result because constrictors elsewhere, as, for example, in the abdomen, raised the pressure so high that facial constrictors are overcome. Such, apparently, is the effect of adrenin already described (see p. 107). Or the flushing might occur from local vasodilation. That very different emotional states may have the same vascular accompaniments was noted by Darwin (The Expression of Emotions in Man and Animals, New York, 1905), who mentioned the pallor of rage (p. 74) and also of terror (p. 77).
In discussing the functioning of the sympathetic division I pointed out that it was roused to activity not only in fear and rage, but also in pain. The machinery of this division likewise is operated wholly or partially in emotions which are usually mild—such as joy and sorrow and disgust—when[p. 278] they become sufficiently intense. Thus, for instance, the normal course of digestion may be stopped or quite reversed in a variety of these emotional states.
Darwin[7] reports the case of a young man who on hearing that a fortune had just been left him, became pale, then exhilarated, and after various expressions of joyous feeling vomited the half-digested contents of his stomach. Müller[8] has described the case of a young woman whose lover had broken the engagement of marriage. She wept in bitter sorrow for several days, and during that time vomited whatever food she took. And Burton,[9] in his Anatomy of Melancholy, gives the following instance of the effect of disgust: “A gentlewoman of the same city saw a fat hog cut up, when the entrails were opened, and a noisome savour offended her nose, she much misliked, and would not longer abide; a physician in presence told her, as that hog, so was she, full of filthy excrements, and aggravated the matter by some other loathsome instances, insomuch this nice gentlewoman apprehended it so deeply that she fell forthwith a vomiting, was so mightily distempered in mind and body, that with all his art and persuasion, for some months after, he could not restore her to herself again, she could not forget or remove the object out of her sight.”
In these three cases, of intense joy, intense sorrow[p. 279] and intense disgust, the influence of the cranial division of the autonomic has been overcome, digestion has ceased, and the stagnant gastric contents by reflexes in striated muscles have been violently discharged. The extent to which under such circumstances other effects of sympathetic impulses may be manifested, has not, so far as I know, been ascertained.
From the evidence just given it appears that any high degree of excitement in the central nervous system, whether felt as anger, terror, pain, anxiety, joy, grief or deep disgust, is likely to break over the threshold of the sympathetic division and disturb the functions of all the organs which that division innervates. It may be that there is advantage in the readiness with which these widely different emotional conditions can express themselves in this one division, for, as has been shown (see p. 276), occasions may arise when these milder emotions are suddenly transmuted into the naturally intense types (as fright and fury) which normally activate this division; and if the less intense can also influence it, the physiological aspect of the transmutation is already partially accomplished.
If various strong emotions can thus be expressed in the diffused activities of a single division of the autonomic—the division which accelerates the heart, inhibits the movements of the stomach and[p. 280] intestines, contracts the blood vessels, erects the hairs, liberates sugar, and discharges adrenin—it would appear that the bodily conditions which have been assumed, by some psychologists, to distinguish emotions from one another must be sought for elsewhere than in the viscera. We do not “feel sorry because we cry,” as James contended, but we cry because when we are sorry or overjoyed or violently angry or full of tender affection—when any one of these diverse emotional states is present—there are nervous discharges by sympathetic channels to various viscera, including the lachrymal glands. In terror and rage and intense elation, for example, the responses in the viscera seem too uniform to offer a satisfactory means of distinguishing states which, in man at least, are very different in subjective quality. For this reason I am inclined to urge that the visceral changes merely contribute to an emotional complex more or less indefinite, but still pertinent, feelings of disturbance in organs of which we are not usually conscious.
This view that the differential features of emotions are not to be traced to the viscera is in accord with the experimental results of Sherrington,[10] who has demonstrated that emotional responses occur in dogs in which practically all the main viscera and the great bulk of skeletal muscle have been removed from subjection to and from influence[p. 281] upon the brain, by severance of the vagus nerves and the spinal cord. In these animals no alteration whatever was noticed in the occurrence, under appropriate circumstances, of characteristic expressions of voice and features, indicating anger, delight or fear. The argument that these expressions may have been previously established by afferent impulses from excited viscera was met by noting that a puppy only nine weeks old also continued to exhibit the signs of emotional excitement after the brain was disconnected from all the body except the head and shoulders. Evidence from uniformity of visceral response and evidence from exclusion of the viscera are harmonious, therefore, in minimizing visceral factors as the source of differences in emotional states.[*]
* The paucity of afferent fibres in the autonomic system, and the probability of an extremely low degree of sensitiveness in the viscera (for evidence, see Cannon: The Mechanical Factors of Digestion, London, 1911, p. 202), likewise support this conclusion.
If these differences are due to other than visceral changes, why is it not always possible by voluntary innervations to produce emotions? We can laugh and cry and tremble. But forced laughter does not bring happiness, nor forced sobbing sorrow, and the trembling from cold rouses neither anger nor fear. The muscle positions and tensions are there, but the experiencing of such bodily changes does not seem even approximately to rouse[p. 282] an emotion in us. Voluntary assumption of an attitude seems to leave out the “feeling.” It is probable, however, that no attitude which we can assume has all the elements in it which appear in the complete response to a stirring situation. But is not this because the natural response is a pattern reaction, like inborn reflexes of low order, such as sneezing, in which impulses flash through peculiarly coöperating neurone groups of the central system, suddenly, unexpectedly, and in a manner not exactly reproducible by volition, and thus they throw the skeletal muscles into peculiar attitudes and, if sufficiently intense, rush out in diffuse discharges that cause tremors and visceral perturbations? The typical facial and bodily expressions, automatically assumed in different emotions, indicate the discharge of peculiar groupings of neurones in the several affective states. That these responses occur instantly and spontaneously when the appropriate “situation,” actual or vividly imagined, is present, shows that they are ingrained in the nervous organization. At least one such pattern, that of anger, persists after removal of the cerebral hemispheres—the decorticated dog, by growling and biting when handled, has the appearance of being enraged;[11] the decerebrate cat, when vigorously stimulated, retracts its lips and tongue, stares with dilated pupils, snarls and snaps its jaws.[12] On the other hand, stroking the hair,[p. 283] whistling and gently calling to produce a pleased attitude, or yelling to produce fright, have not the slightest effect in evoking from the decorticated dog signs of joy and affection or of fear, nor does the animal manifest any sexual feeling. The absence of bodily indications of these emotions is quite as significant as the presence of the signs of anger. For since expressions of anger can persist without the cerebral cortex, there is little reason why the complexes of other emotional expressions, if their “machinery” exists below the cortex, should not also be elicitable. That they are not elicitable suggests that they require a more elaborately organized grouping of neurones than does anger—possibly what the cortex, or the cortex in combination with basal ganglia, would provide.
The contrast between the brevity of the “pseudo-affective reactions” in the decerebrate cat, though the viscera are still connected with the central nervous system, and the normal duration of emotional expression in the dog with the body separated from the head region, has been used by Sherrington to weigh the importance of the visceral and other factors. And the evidence which I have given above, as well as that which he has offered, favors the view that the viscera are relatively unimportant in an emotional complex, especially in contributing differential features.
[p. 284]
1 James: Principles of Psychology, New York, 1905, i, p. 22.
2 Prince: The Unconscious, New York, 1914, p. 456.
3 Hertz: Constipation and Allied Intestinal Disorders, London, 1909, p. 81.
4 v. Trenck: Merkwürdige Lebensgeschichte, Berlin, 1787, p. 195.
5 James, Loc. cit., p. 415.
6 McDougall: Introduction to Social Psychology, London, 1908, p. 72.
7 Darwin: Loc. cit., p. 76.
8 Müller: Deutsches Archiv für klinische Medicin, 1907, lxxxix, p. 434.
9 Burton: The Anatomy of Melancholy (first published in 1621), London, 1886, p. 443.
10 Sherrington: Proceedings of the Royal Society, 1900, lxvi, p. 397.
11 Goltz: Archiv für die gesammte Physiologie, 1892, li, p. 577.
12 Woodworth and Sherrington: Journal of Physiology, 1904, xxxi, p. 234.
[p. 285]
ALTERNATIVE SATISFACTIONS FOR THE FIGHTING EMOTIONS
The uniformity of visceral responses when almost any feelings grow very intense, and under such conditions the identity of these responses with those characteristically aroused in the belligerent emotion of anger or rage and its counterpart, fear, offer interesting possibilities of transformation and substitution. This is especially true in the activities of human beings. And because men have devised such terribly ingenious and destructive modes of expressing these feelings in war, an inquiry into the basis for possible substitution seems not out of place.
The business of killing and of avoiding death has been one of the primary interests of living beings throughout their long history on the earth. It is in the highest degree natural that feelings of[p. 286] hostility often burn with fierce intensity, and then, with astonishing suddenness, that all the powers of the body are called into action—for the strength of the feelings and the quickness of the response measure the chances of survival in a struggle where the issue may be life or death. These are the powerful emotions and the deeply ingrained instinctive reactions which invariably precede combat. They are the emotions and instincts that sometimes seize upon individuals in groups and spread like wildfire into larger and larger aggregations of men, until vast populations are shouting and clamoring for war. To whatever extent military plans are successful in devising a vast machine for attack or defense, the energies that make the machine go are found, in the last analysis, in human beings who, when the time for action comes, are animated by these surging elemental tendencies which assume control of their conduct and send them madly into conflict.
The strength of the fighting instinct in man has been one of the main arguments used by the militarists in support of preparation for international strife. They point to the historical fact that even among highly civilized peoples scarcely a decade passes without a kindling of the martial emotions, which explode in actual warfare. Such fighting, they say, is inevitable—the manifestation of “biological law”—and, so long as human nature[p. 287] remains unchanged, decision by battle must be resorted to. They urge, furthermore, that in war and in the preparations for war important physical qualities—sturdiness, hardihood, and strength for valorous deeds—are given peculiarly favorable opportunities for development, and that if these opportunities are lacking, lusty youth will give place to weaklings and mollycoddles. In addition the militarists say that war benefits mankind by its moral effects. Without war nations become effete, their ideals become tarnished, the people sink into self-indulgence, their wills weaken and soften in luxury. War, on the contrary, disciplines character, it sobers men, it teaches them to be brave and patient, it renews a true order of values, and its demand for the supreme sacrifice of life brings forth in thousands an eager response that is the crowning glory of the human spirit. As the inevitable expression of a deep-rooted instinct, therefore, and as a unique means of developing desirable physical and moral qualities, war is claimed by the militarists to be a natural necessity.[1]
The militarist contention that the fighting instinct is firmly fixed in human nature receives strong confirmation in the results of our researches. Survival has been decided by the grim law of mortal conflict, and the mechanism for rendering the body more competent in conflict has been revealed in earlier chapters as extraordinarily perfect[p. 288] and complete. Moreover, the physiological provisions for fierce struggle are found not only in the bodies of lower animals, that must hunt and kill in order to live, but also in human beings. Since this remarkable mechanism is present, and through countless generations has served the fundamentally important purpose of giving momentous aid in the struggle for existence, the militarists might properly argue that, as with other physiological processes, bodily harmony would be promoted by its exercise. Indeed, they might account for the periodic outburst of belligerent feelings by assuming that these natural aptitudes require occasional satisfaction.[*]
* Mr. Graham Wallas has made the interesting suggestion (The Great Society, New York, 1914, p. 66) that nervous strain and restlessness due to “baulked disposition” may result from the absence of circumstances which would call the emotional responses into action. And he cites Aristotle’s theory that pent passions may be released by represented tragedy and by music.
In spite of the teachings of history that wars have not grown fewer, and in spite of the militarist argument that war is a means of purging mankind of its sordid vices, and renewing instead the noblest virtues, the conclusion that the resort to arms is unavoidable and desirable is nowadays being strongly contested. The militarists show only[p. 289] part of the picture. No large acquaintance with the character of warfare is necessary to prove that when elemental anger, hate and fear prevail, civilized conventions are abandoned and the most savage instincts determine conduct. Homes are looted and burned, women and children are abominably treated, and many innocents are murdered outright or starved to death. No bland argument for the preservation of the manly virtues can palliate such barbarities. Even when fighting men are held within the rules, the devices for killing and injuring are now made so perfect by devilish ingenuity that by the pulling of a trigger one man can in a few seconds mow down scores of his fellow-creatures and send them writhing to agony or death. War has become too horrible; it is conducted on too stupendous a scale of carnage and expenditure; it destroys too many of the treasured achievements of the race; it interferes too greatly with consecrated efforts to benefit all mankind by discovery and invention; it involves too much suffering among peoples not directly concerned in the struggle; it is too vastly at variance with the methods of fair dealing that have been established between man and man; the human family has become too closely knit to allow some of its members to bring upon themselves and all the rest poverty and distress and a long heritage of bitter hatred and resolution to seek revenge.
[p. 290]
All these reasons for hostility to war imply a thwarting of strong desires in men—desires for family happiness, devotion to beauty and to scholarship, passion for social justice, hopes of lessening poverty and disease. As was pointed out in the previous chapter, the feeling of hostility has no definite object to awaken it. It is roused when there is opposition to what we ardently wish to get. And because war brings conditions which frustrate many kinds of eagerly sought purposes, war has roused in men a hostility against itself. There is then a war against war, a willingness to fight against monstrous carnage and destruction, that grows in intensity with every war that is waged.
Although there is increasing opposition to the display of the fighting emotions and instincts in war, nevertheless the admirable moral and physical qualities, claimed by the militarists to be the unique products of war, are too valuable to be lost. As McDougall[2] has indicated, when the life of ideas becomes richer, and the means we take to overcome obstructions to our efforts more refined and complex, the instinct to fight ceases to express itself in its crude natural manner, save when most intensely excited, and becomes rather a source of increased energy of action towards the end set by any other instinct; the energy of its impulses adds[p. 291] itself to and reënforces that of other impulses and so helps us to overcome our difficulties. In this lies its great value for civilized man. A man devoid of the pugnacious instinct would not only be incapable of anger, but would lack this great source of reserve energy which is called into play in most of us by any difficulty in our path.
Thus the very efficiency of a war against war, as well as struggle against other evils that beset civilized society, rests on the preservation and use of aggressive feeling and the instinct to attack. From this point of view the insistence by the militarists that we must accept human nature as we find it, and that the attempt to change it is foolish, seems a more justifiable attitude than that of the pacifists who belittle the fighting qualities and urge that changing them is a relatively simple process. We should not wish them changed. Even if in the war against war a means should be established of securing international justice, and if through coöperative action the decrees of justice were enforced, so that the occasions which would arouse belligerent emotions and instincts were much reduced, there would still remain the need of recognizing their elemental character and their possible usefulness to society. What is needed is not a suppression of these capacities to feel and act, but their diversion into other channels where they may have satisfactory expression.
[p. 292]
“We must make new energies and hardihoods continue the manliness to which the military mind so faithfully clings. Martial virtues must be the enduring cement; intrepidity, contempt of softness, surrender of private interest, obedience to command, must still remain the rock upon which states are built.” Thus wrote William James[3] in proposing a “moral equivalent for war.” This, he suggested, should consist of such required service in the hard and difficult occupations as would take the childishness and superciliousness out of our youth and give them soberer ideas and healthier sympathies with their fellow-men. He conceived that by proper direction of its education a people should become as proud of the attainment by the nation of superiority in any ideal respect as it would be if the nation were victorious in war. “The martial type of character,” he declared, “can be bred without war. Strenuous honor and disinterestedness abound elsewhere. Priests and medical men are in a fashion educated to it, and we should all feel some degree of it imperative if we were conscious of our work as an obligatory service to the state. We should be owned, as soldiers are by the army, and our pride would rise accordingly. We could be poor, then, without humiliation, as army officers now are. The only thing needed henceforth is to inflame the civic[p. 293] temper as past history has inflamed the military temper.”
Similar ideas have been expressed by others.[4] It has been pointed out that the great war of mankind is that against pain, disease, poverty and sin; that the real heroes are not those who squander human strength and courage in fighting one another, but those who fight for man against these his eternal foes. War of man against man, in this view, becomes dissension in the ranks, permitting the common enemies to strike their most telling blows.
These moral considerations, however, are apart from the main intent of our discussion. Our earlier inquiry confirmed the belief that the fighting emotions are firmly rooted in our natures, and showed that these emotions are intimately associated with provisions for physical exertion. It is particularly in this aspect of the discussion of substitutes for war that these studies have significance.
The idealization of the state and the devotion of service to social welfare, which have been suggested as moral substitutes for military loyalty, leave unanswered the claims of the militarists that in war and in preparations for war opportunities are offered which are peculiarly favorable to the development of important physical qualities—bodily[p. 294] vigor, sturdiness, and ability to withstand all manner of hardships.
In the evidence previously presented, it seems to me there was a suggestion that offers a pertinent alternative to these claims. When the body goes onto what we have called a war footing, the physiological changes that suddenly occur are all adapted to the putting forth of supreme muscular and nervous efforts. That was what primitive battle consisted of, through countless myriads of generations—a fierce physical contest of beast with beast, and of man with man. Such contests, attended as they were by the thrill of unpredictable incidents, and satisfying completely the lust of combat, are to be contrasted with the dull grind in preparation for modern war, the monotonous regularity of subservience, the substitution everywhere of mechanism for muscle, and often the attack on an enemy who lies wholly unseen.[*] As[p. 295] Wallas with nice irony has remarked, “The gods in Valhalla would hardly choose the organization of modern lines of military communication, as they chose the play of sword and spear, to be the most exquisite employment of eternity.”
* Lord Wolseley, while commander-in-chief of the English forces, in 1897, secured sanction for not displaying the regimental colors in battle. “It would be madness and a crime,” he declared, “to order any soldier to carry colors into action in the future. You might quite as well order him to be assassinated. We have had most reluctantly to abandon a practice to which we attached great importance, and which, under past and gone conditions of fighting, was invaluable in keeping alive the regimental spirit upon which our British troops depended so much.” All war has been transformed by the invention of the far-reaching and fate-dealing rifle and automatic gun, with which an enemy kills, whose face is not even seen. War is almost reduced to a mechanical interchange of volleys and salvoes, and to the intermittent fire of rifles and machine guns, with short rushes at the last, in which there is no place for the dignity and grace of the antique battle of the standard. (See London Times, July 31, 1897, p. 12.)
T. F. Millard, the well-known correspondent of the Russo-Japanese War, wrote as follows of the characteristics of present day conflicts: “A large part of modern war is on too great a scale to give much opportunity for individual initiative. Soldiers can rarely tell what is going on in their immediate vicinity. They cannot always see the enemy they are firing at, and where they can see the object of their fire such an important matter as range and even direction cannot be left to them.... Troops are clothed so much alike nowadays that it is very difficult to distinguish friend from foe at five hundred yards, and large bodies of troops rarely get that close to each other in modern war while there is light enough to see clearly.... Battery officers simply see that their guns are handled according to instructions. They regulate the time, speed, objective and range as ordered.... The effects of the fire are observed by officers appointed to that duty, stationed at various parts of the field, often miles and miles apart, and who are in constant communication with the chief of artillery by telephone.” (See Scribner’s Magazine, 1905, xxxvii, pp. 64, 66.)
The testimony of a captain of a German battery engaged against the French and English in 1914, supports the foregoing claims. He is reported as saying: “We shoot over those tree tops yonder in accordance with directions for range and distance which come from somewhere else over a field telephone, but we never see the men at whom we are firing. They fire back without seeing us, and sometimes their shells fall short or go beyond us, and sometimes they fall among us and kill and wound a few of us. Thus it goes on day after day. I have not with my own eyes seen a Frenchman or an Englishman unless he was a prisoner. It is not so much pleasure—fighting like this.” (See Philadelphia Saturday Evening Post, December 26, 1914, p. 27.)
While it is true that physical strength can be developed by any form of hard labor, as, for example, by sawing wood or digging ditches, such labor does not stimulate quickness, alertness, and resourcefulness in bodily action. Nor does it give any occasion for use of the emotional mechanism for reënforcement. If this mechanism, like other physiological arrangements, is present in the body for use—and previous discussion leaves little[p. 296] doubt of that—then as a means of exercising it and, in addition, satisfying the strong instinct for competitive testing of strength and physical skill, some activity more enlivening than monotonous gymnastics and ordered marching is required.
In many respects strenuous athletic rivalries present, better than modern military service, the conditions for which the militarists argue, the conditions for which the body spontaneously prepares when the passion for fighting prevails. As explained in an earlier chapter, in competitive sports the elemental factors are retained—man is again pitted against man, and all the resources of the body are summoned in the eager struggle[p. 297] for victory. And because, under such circumstances, the same physiological alterations occur that occur in anticipation of mortal combat, the belligerent emotions and instincts, so far as their bodily manifestations are concerned, are thereby given complete satisfaction.
For reasons given above, I venture to lay emphasis on a suggestion, which has been made before by others, that the promotion of great international athletic contests, such as the Olympic games, would do for our young men much that is now claimed as peculiar to the values of military discipline. The substitution of athletic rivalries for battle is not unknown. In the Philippine Islands, according to Worcester,[5] there were no athletics before the American occupation. The natives soon learned games from the soldiers. And when the sports reached such development that competition between towns and provinces was possible, they began to arouse the liveliest enthusiasm among the people. The physical development of the participants has been greatly stimulated, the spirit of fair play and sportsmanship, formerly lacking, has sprung into existence in every section of the Islands, and the annual meets between athletic teams from various provinces are recognized as promoting a general and friendly[p. 298] understanding among the different Filipino tribes. The fierce Igarots of Bontoc, once constantly at war with neighboring tribes, now show their prowess not in head-hunting, but in baseball, wrestling, and the tug-of-war.[*]
* It is reported that when these warriors first appeared at the games, each brought his spear, which he drove into the ground beside him, ready for use. As the nature of the new rivalries became known, the spears were left behind.
Is it unreasonable to expect that what has happened in the Philippine Islands might, by proper education and suggestion, happen elsewhere in the world? Certainly the interest in athletic contests is no slight and transient interest. At the time of a great war we know that news of the games is fully as much demanded as news of the war. Already in the United States, without special stimulation, the number of young men engaged in athletic training is estimated as equal to the number in the standing army. And in England, belief in the efficacy of athletics as a means of promoting hardihood and readiness to face stern hazards has found expression in the phrase that England’s battles have been won on the football fields of Rugby and of Eton. With the further promotion of international contests the influence of competitive sports is likely to increase rather than lessen. Within national boundaries emulation is sure to stimulate extensively such games as will bring forth the best representative athletes that the country[p. 299] can produce. In one of the high-spirited European nations, which made a poor showing at the last Olympic meet, thousands of young men began training for the next meet, under a director imported from the nation that had made the highest records.
Training for athletic contests is quite as likely to enure young men to physical hardship and fatigue, is quite as conducive to the development of bodily vigor, the attainment of alertness and skill and the practice of self-restraint, as is army life with its traditional associations and easy license. It may be urged, however, that an essential element is lacking in all this discussion—the sobering possibility that in war the supreme surrender of life itself may be required. Death for one’s country is indeed glorious. But the argument that being killed is desirable has little to commend it. When the strongest and sturdiest are constantly chosen to be fed to the engines of annihilation, the race is more likely to lose greater values than it gains from the spectacle of self-sacrifice, however perfect that may be. Are there not advantages in the conditions of great athletic rivalries that may compensate for war’s most austere demand? The race of hardy men, to secure which the militarists urge war, is much more likely to result from the honoring and preserving of vigorous men in their vigor than it is from the[p. 300] systematic selection of such men to be destroyed in their youth.
There are other aspects of international games which strongly commend them as an alternative to the pursuit of military discipline. The high standards of honor and fairness in sport; its unfailing revelation of excellence without distinctions of class, wealth, race or color; the ease with which it becomes an expression of the natural feelings of patriotism; the respect which victory and pluckily borne defeat inspire in competitors and spectators alike; the extension of acquaintance and understanding which follows from friendly and magnanimous rivalry among strong men who come together from the ends of the earth—each of these admirable features of athletic contests between nations might be enlarged upon. But, as intimated before, these moral considerations must be left without further mention, as being irrelevant to the physiological processes with which we are dealing.
We are concerned with the question of exercising the fighting instinct and thus assuring the physical welfare of the race. The race must degenerate, the militarists say, if this instinct is not allowed to express itself in war. This declaration we are in a position to deny, for the evidence is perfectly clean-cut that the aggressive instincts, which through æons of racial experience[p. 301] have naturally and spontaneously developed vigor and resourcefulness in the body, are invited by elemental emotions, and that through these emotions energies are released which are highly useful to great physical effort. No stupid routine of drill, or any other deadening procedure, will call these energizing mechanisms into activity. War and the preparations for war nowadays have become too machine-like to serve as the best means of preserving and disciplining these forces. The exhilarating swing and tug and quick thrust of the big limb muscles have largely vanished. Pressing an electric contact or bending the trigger finger is a movement altogether too trifling. If, then, natural feelings must be expressed, if the fighting functions of the body must be exercised, how much better that these satisfactions be found in natural rather than in artificial actions, how much more reasonable that men should struggle for victory in the ancient ways, one against another, body and spirit, as in the great games.
1 See Angell: The Great Illusion, New York and London, 1913, pp. 159–164.
2 McDougall: Introduction to Social Psychology, London, 1908, p. 61.
3 James: Memories and Studies, New York, 1911, p. 287.
4 See Perry: The Moral Economy, New York, 1909, p. 32; and Drake: Problems of Conduct, Boston, 1914, p. 317.
5 Worcester: The Philippines, Past and Present, New York, 1914, ii, pp. 515, 578.
[p. 302]
1. The Influence of Emotional States on the Functions of the Alimentary Canal. By W. B. Cannon. American Journal of the Medical Sciences, 1909, cxxxvii, pp. 480–487.
2. Emotional Stimulation of Adrenal Secretion. By W. B. Cannon and D. de la Paz. American Journal of Physiology, 1911, xxviii, pp. 64–70.
3. The Effects of Asphyxia, Hyperpnœa, and Sensory Stimulation on Adrenal Secretion. By W. B. Cannon and R. G. Hoskins. Ibid., 1911, xxix, pp. 274–279.
4. Emotional Glycosuria. By W. B. Cannon, A. T. Shohl and W. S. Wright. Ibid., 1911, xxix, pp. 280–287.
5. A Consideration of Some Biological Tests for Epinephrin. By R. G. Hoskins. Journal of Pharmacology and Experimental Therapeutics, 1911, iii, pp. 93–99.
6. The Sthenic Effect of Epinephrin upon Intestine. By R. G. Hoskins. American Journal of Physiology, 1912, xxix, pp. 363–366.
7. An Explanation of Hunger. By W. B. Cannon and A. L. Washburn. Ibid., 1912, xxix, pp. 441–454.
8. A New Colorimetric Method for the Determination of Epinephrin. By O. Folin, W. B. Cannon and W. Denis. Journal of Biological Chemistry, 1913, xiii, pp. 477–483.
9. The Depressor Effect of Adrenalin on Arterial Pressure. By W. B. Cannon and Henry Lyman. American Journal of Physiology, 1913, xxxi, pp. 376–398.
[p. 303]
10. The Effect of Adrenal Secretion on Muscular Fatigue. By W. B. Cannon and L. B. Nice. Ibid., 1913, xxxii, pp. 44–60.
11. Fatigue as Affected by Changes of Arterial Pressure. By C. M. Gruber. Ibid., 1913, xxxii, pp. 222–229.
12. The Threshold Stimulus as Affected by Fatigue and Subsequent Rest. By C. M. Gruber. Ibid., 1913, xxxii, pp. 438–449.
13. The Fatigue Threshold as Affected by Adrenalin and by Increased Arterial Pressure. By C. M. Gruber. Ibid., 1914, xxxiii, pp. 335–355.
14. The Emergency Function of the Adrenal Medulla in Pain and the Major Emotions. By W. B. Cannon. Ibid., 1914, xxxiii, pp. 356–372.
15. The Relation of Adrenalin to Curare and Fatigue in Normal and Denervated Muscles. By C. M. Gruber. Ibid., 1914, xxxiv, pp. 89–96.
16. The Graphic Method of Recording Coagulation. By W. B. Cannon and W. L. Mendenhall. Ibid., 1914, xxxiv, pp. 225–231.
17. The Hastening or Retarding of Coagulation by Adrenalin Injections. By W. B. Cannon and Horace Gray. Ibid., 1914, xxxiv, pp. 232–242.
18. The Hastening of Coagulation by Stimulating the Splanchnic Nerves. By W. B. Cannon and W. L. Mendenhall. Ibid., 1914, xxxiv, pp. 243–250.
19. The Hastening of Coagulation in Pain and Emotional Excitement. By W. B. Cannon and W. L. Mendenhall. Ibid., 1914, xxxiv, pp. 251–261.
20. The Interrelations of Emotions as Suggested by Recent Physiological Researches. By W. B. Cannon. American Journal of Psychology, 1914, xxv, pp. 256–282.
[p. 305]
The following changes have been made to the text as printed.
1. Illustrations and end-of-page footnotes (marked with an asterisk) have been located in appropriate paragraph breaks. References, marked with numbered indices, are listed at the end of each chapter as in the book.
2. Obvious typographical errors have been corrected.
3. Where a word is used repeatedly in the same way, spelling and hyphenation have been made consistent, preferring the form most often used in the printed work, or failing that the more usual form in general use at the time of publication. No typographical change has been made within direct quotes from other works.
4. Page 25: "or thoradico-lumbar division" has been changed to "or thoracico-lumbar division".
5. Page 58: The name "Emden" in "Emden and v. Furth" has been changed to "Embden" in agreement with Footnote 1 on Page 65. (Gustav Embden, 1874–1933.)
6. Page 62: The Greek letter Β (Beta) has been substituted for Latin B at the start of "Β-tetrahydronaphthylamine".