
Plate I.—Illustrating the results of Artificial Selection. Within historic times the small Wild Rock Pigeon has been evolved into the large Pouter. The figure illustrates the plasticity of a living organism.
Title: A first book in organic evolution
Author: D. Kerfoot Shute
Release date: June 15, 2024 [eBook #73834]
Language: English
Original publication: Chicago: The Open Court Publishing Company
Credits: Charlene Taylor and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive)
Plate I.—Illustrating the results of Artificial Selection. Within historic times the small Wild Rock Pigeon has been evolved into the large Pouter. The figure illustrates the plasticity of a living organism.
BY
D. KERFOOT SHUTE, A.B., M.D.
OPHTHALMIC SURGEON TO THE UNIVERSITY HOSPITAL (COLUMBIAN)
PROFESSOR OF ANATOMY IN THE COLUMBIAN UNIVERSITY

CHICAGO
THE OPEN COURT PUBLISHING COMPANY
London: Kegan Paul, Trench, Trübner & Co., Ltd.
Paternoster House, Charing Cross Road
1899
COPYRIGHT BY
The Open Court Publishing Co.
CHICAGO, U. S. A.
1899
All rights reserved.
THIS LITTLE BOOK IS INSCRIBED TO
DR. THEODORE GILL,
NOT ONLY IN ADMIRATION FOR HIS HIGH SCHOLARSHIP
AND EMINENT SCIENTIFIC ATTAINMENTS, BUT
ALSO IN APPRECIATION OF MANY
ACTS OF COURTESY AND
KINDNESS TO THE
AUTHOR.
This little book has been written chiefly for the use of students in the Medical Department of the Columbian University. It is designed to serve only as an introduction to the study of the Development Theory, and the subject has been presented, it is hoped, in a manner that will render it interesting and easily intelligible to the general reader.
The doctrine of Evolution itself enters so largely into all those departments of knowledge that especially concern the human race, and it has so profoundly modified our ideas with regard to the origin and destiny of man, that it has attained a commanding interest and become an almost necessary ingredient in what is called a liberal education.
An overwhelming majority of Anthropologists, Zoölogists, and Botanists, and a goodly number—constantly increasing—of Christian clergymen and laymen, have been almost compelled to believe in the truth of the Evolution Theory, whether they would or not, and they cannot but realize how very widely the theory extends into almost every department of human knowledge. No one, therefore, who aspires even to a moderate degree of intellectual culture, can well afford to exclude a clear understanding of what this Doctrine of Evolution[vi] really is. It is hoped this little work will render such a conception easily attainable.
The author makes no claim for originality, unless it be in the manner of presenting the subject. He has utilized the facts collated by other observers, and sometimes quoted the exact language and expressions of well-known writers on Evolution, and has endeavored to put them together in a way that may be helpful to those who are beginning the study of the Evolution Theory.
No attempt has been made to prove the truth of the theory: this is assumed. The arguments in support of it are coextensive with our knowledge of Comparative Anatomy, Embryology, Physiology, Psychology, and many other sciences.
In the preparation of the book the author is especially indebted to his friend Prof. Theodore Gill, the eminent ichthyologist, for many valuable suggestions, and more particularly for his aid in constructing the Diagram of Development. He also desires to thank his friend Dr. A. F. A. King for his kindly assistance in preparing the manuscript for the press; and also his friend Dr. L. O. Howard, Chief of the U. S. Bureau of Entomology, for much valuable information and assistance.
D. K. Shute.
August 1, 1899, 1318 L Street, N. W., Washington, D. C.
| PAGE | |
| Introduction | xiii |
| System of Classification | xiv |
| Definition of Evolution | xv |
| SECTION I. | |
| Organic Cells: The Visible Units of Life | 3 |
| Structure and Composition of Cells | 4 |
| Activities of Cells | 7 |
| Examples of Unicellular Animals and Plants | 10 |
| Mitosis | 25 |
| Maturation of the Human Ovum | 29 |
| Fertilization of the Human Ovum | 29, 42 |
| Segmentation of the Oösperm | 31 |
| Gastrulation | 32 |
| SECTION II. | |
| Heredity with Variation | 39 |
| Examples of Variations | 39 |
| Illustrations of Heredity | 40 |
| Nucleus in Heredity | 41 |
| Heritages | 47 |
| Pseudo-Heredity | 53 |
| Environment and Variations | 55 |
| Acquired Characters | 65 |
| Pangenesis | 66 |
| Continuity of Germ-Cells | 67 |
| Modified Pangenesis | 68 |
| Heredity Stronger than Environment | 75 |
| SECTION III. | |
| Unstable Environment | 81 |
| Development of North America | 85 |
| Archæan Era | 85 |
| Palæozoic Era | 86 |
| Silurian Period | 87 |
| Devonian Period | 87 |
| Carboniferous Period | 87[viii] |
| Jurassic Period | 88 |
| Cretaceous Period | 88 |
| Tertiary Period | 89 |
| Quaternary Period | 89 |
| SECTION IV. | |
| Transmutations of Living Forms | 93 |
| Archæan Era | 94 |
| Table of Stratified Rocks and the Successive Appearance of Typical Life-Forms | 95 |
| Cambrian Period | 96 |
| Lower Silurian Period | 96 |
| Upper Silurian Period | 98 |
| Devonian Period | 100 |
| Carboniferous Period | 102 |
| Permian Period | 104 |
| Triassic Period | 105 |
| Jurassic Period | 106 |
| Cretaceous Period | 108 |
| Tertiary Period | 110 |
| Quaternary Period | 113 |
| SECTION V. | |
| Natural Selection | 119 |
| Artificial Selection | 120 |
| Multiplication of Animals | 122 |
| Elimination of the Unfit | 125 |
| The Coloration of Animals and Environment | 127 |
| Protective Coloration | 128 |
| Protective Resemblance | 138 |
| Alluring Coloration | 141 |
| Warning Coloration | 144 |
| Mimicry | 147 |
| Recognition Marks | 150 |
| Sexual Selection | 152 |
| Insect Selection | 160 |
| Isolation of Varieties | 166 |
| SECTION VI. | |
| Evolution of Man | 173 |
| Development of the Frog | 173 |
| Development of Man | 180[ix] |
| Useless Scaffolding in Man | 191 |
| Development of the Brain | 199 |
| The Brain and Psychic Phenomena | 206 |
| Evolution and Social Problems | 222 |
| SECTION VII. | |
| Classification of Animals and Plants | 227 |
| Protozoa | 229 |
| Porifera | 230 |
| Cœlenterata | 230 |
| Echinodermata | 231 |
| Vermes | 232 |
| Arthropoda | 233 |
| Mollusca | 233 |
| Vertebrata | 234 |
| Tunicata | 234 |
| Leptocardii | 235 |
| Marsipobranchii | 235 |
| Pisces | 236 |
| Amphibia | 237 |
| Reptilia | 238 |
| Aves | 238 |
| Mammalia | 239 |
| Primates | 239 |
| Lemuroidea | 240 |
| Anthropoidea | 241 |
| Hapalidæ | 242 |
| Cebidæ | 242 |
| Cercopithecidæ | 242 |
| Simiidæ | 243 |
| Hominidæ | 243 |
| Flowerless Plants | 244 |
| Flowering Plants | 244 |
| SECTION VIII. | |
| Works of Reference | 247 |
| SECTION IX. | |
| Glossary | 255 |
| SECTION X. | |
| Index | 277 |
| FIG. | PAGE | |
| 1. | Diagram of a Cell. (Drawn by Mr. E. P. Copeland from a sketch by the Author.) | 5 |
| 2. | Stylonychia. (Drawn by Mr. E. P. Copeland.) | 9 |
| 3. | Amœba. (Drawn by Mr. E. P. Copeland.) | 11 |
| 4. | Rotalia. (Drawn by Dr. A. L. Lawrence.) | 12 |
| 5. | Difflugia. (Drawn by Dr. A. L. Lawrence.) | 17 |
| 6. | Noctiluca. (Drawn by Dr. A. L. Lawrence.) | 18 |
| 7. | Gromia. (Drawn by Dr. A. L. Lawrence.) | 21 |
| 8. | Diagram Illustrating Mitosis. (Drawn by Mr. E. P. Copeland from a sketch by the Author.) | 26 |
| 9. | Diagram Illustrating the Maturation and Fertilization of the Ovum. (Drawn by Mr. E. P. Copeland from a sketch by the Author.) | 30 |
| 10. | Diagram Illustrating Segmentation and Gastrulation. (Drawn by Mr. E. P. Copeland from a sketch by the Author.) | 34 |
| 11. | Archæan North America. (From a drawing in Shaler’s “First Book in Geology.”) | 84 |
| 12. | Cretaceous North America. (From a drawing in Shaler’s “First Book in Geology.”) | 86 |
| 13. | Tertiary North America. (From a drawing in Shaler’s “First Book in Geology.”) | 89 |
| 14. | Genesis of Horse’s Feet. (Drawn by Mr. E. P. Copeland.) | 112 |
| 15. | Bipes, Cheirotes, and Snake. (From a drawing in Shaler’s “First Book in Geology.”) | 114 |
| 16. | Domesticated Pig and Wild Boar. (From Romanes’ “Darwin and After Darwin.”) | 121 |
| 17. | Tadpoles and Frog. (Drawn by Dr. A. L. Lawrence.) | 175 |
| 18. | Diagram of Development. (Drawn by Mr. B. H. Esterly from a sketch by the Author.) | 183 |
| 19. | Brain of Fish. (Drawn by Mr. B. H. Esterly from specimens of the Author.) | 198[xi] |
| 20. | Brain of Reptile. (Drawn by Mr. B. H. Esterly from specimens of the Author.) | 199 |
| 21. | Brain of a Marsupial. (Drawn by Mr. B. H. Esterly from specimens of the Author.) | 200 |
| 22. | Brain of a Lemur. (Modified by Mr. B. H. Esterly from Flower.) | 201 |
| 23. | Ideal Sections of Brains. (Modified from Le Conte. Drawn by Dr. A. L. Lawrence from a sketch by the Author.) | 202 |
| 24. | Brain of Human Fœtus—Fish Phase. (Drawn by Mr. B. H. Esterly from specimens of the Author.) | 204 |
| 25. | Brain of Human Fœtus—Reptile Phase. (Drawn by Mr. B. H. Esterly from specimens of the Author.) | 204 |
| 26. | Brain of Human Fœtus—Marsupial Phase. Side View. (Drawn by Mr. B. H. Esterly from specimens of the Author.) | 204 |
| 27. | Brain of Human Fœtus—Marsupial Phase. Dorsal View. (Drawn by Mr. B. H. Esterly from specimens of the Author.) | 205[xii] |
| COLORED PLATES | ||
| PLATE | FACING PAGE | |
| I. | Wild Rock Pigeon and Pouter. (Drawn by Mr. J. L. Ridgway from specimens in the Smithsonian Institution.) | Frontispiece |
| II. | Butterflies. (Drawn by Miss L. Sullivan from specimens in the United States Bureau of Entomology.) | 138 |
| III. | Caterpillar of Geometer Moth. (Drawn by Miss L. Sullivan from specimens in the United States Bureau of Entomology.) | 142 |
| V. | Spider on Flower. (Drawn by Miss L. Sullivan from a plate in McCook’s “American Spiders.”) | 144 |
| VI. | Elaps and Erythrolamprus. (Modified by Mr. J. L. Ridgway from a figure in Romanes’ “Darwin and After Darwin.”) | 146 |
| VII. | Salamandra maculosa. (Drawn by Mr. J. L. Ridgway from specimens in the United States National Museum.) | 148 |
| VIII. | Paradise Birds. (Drawn by Mr. J. L. Ridgway from specimens in the Smithsonian Institution.) | 152 |
| IX. | Primrose Flowers. (Drawn by Miss L. Sullivan from specimens.) | 162 |
| X. | Bee Fertilizing Flower. (Drawn by Miss L. Sullivan from specimens in the United States Bureau of Entomology.) | 164 |
| PLATES ENGRAVED IN BLACK AND WHITE | ||
| IV. | Leaf-hoppers. (Drawn by Miss L. Sullivan from drawings collected by Dr. L. O. Howard.) | 140 |
| XI. | Babies’ Grasping Power. (From a Photograph taken by Dr. Louis Robinson.) | 192 |
| XII. | Brain of Man. (From Carus’ “The Soul of Man.”) | 202 |
It is extremely difficult to realize the variety, wealth, and grandeur of animal and plant life as they exist on the globe to-day. Even if we endeavor to recall, in imagination, all that we have seen in streams and woodland, in ponds and rivers, in meadows, and in the air; even when we call to mind the multifarious specimens we have beheld in all the museums of natural history we have visited, and remember the most vivid descriptions that we have ever read of the wealth of tropical life;—even then we have only the faintest conception of the multitude and variety of living forms, not to mention the vanished hosts of bygone ages.
It has always been the endeavor of students of nature to reduce this great host of living creatures to order by some system of classification. First one system of classification was adopted, and then another, with the progress of Botany and Zoölogy, until eventually, before the theory of evolution was entertained by scientific men, that system was adopted which naturalists likened to a tree. In this system, those lowest organisms which cannot properly be called either animals or plants, may be represented by a short trunk. Soon this short trunk divides into two large trunks, one of which represents[xiv] the animal kingdom and the other the vegetable kingdom. Each of these trunks then sends off large branches representing classes, from which smaller and more numerous branches, representing orders, are given off. From these, other branches and sub-branches are separated, representing families and genera, and finally the terminal twigs or leaves represent species.
In this tree, while there is a general advance in organization from below upwards, there are many deviations in this respect. Some leaves may be growing on different branches at the same level, which means that species belonging to widely divergent classes or orders may still possess an equal grade of organization. On the same branch there may be growing leaves at different levels. This means that one species may be more highly organized than another belonging to the same class. Not only may all living species be classified in the form of a family tree, but all the extinct hosts of species that lived in the ages of the past may be similarly classified. If all the animals that have ever lived on the globe should be represented by a tree, those existing on the earth to-day would be indicated by the topmost twigs and leaves, while the extinct forms would be represented by the trunk and main branches. (Vid. Diagram of Development.)
The detecting of this tree-like arrangement of species in nature is the progressive work of naturalists for centuries past. At about the commencement of the present century, when it was finally detected, naturalists were unable to understand the significance of it. They did not perceive the underlying principle that accounts for the fact that groups[xv] of living forms have such natural affinities that they are arranged like a family tree.
When Darwin came upon the scene and found that his predecessors had already empirically worked out the tree-like system of classification, he convinced naturalists that the great underlying principle of this system was Heredity; and that, therefore, the grouping of living and extinct organisms in a family tree according to their natural affinities is a grouping based upon genetic affinities; and further that the ultimate meaning of classification is the tracing of lines of pedigree. This signifies that all the creatures living on the globe to-day, and all the hosts that lived in the ages of the past, are blood relations in greater or less degree, and that they have all been evolved from simple microscopic creatures that appeared on the globe at the dawn of life. All organisms, then, have undergone an evolution.
The factors of fundamental importance in the study of the theory of Evolution, or the doctrine of the Transmutations of Organisms, are Cells, Heredity and Variation, Environment, Natural Selection, and Isolation.
The evolution of an organism means its descent from preceding organisms with continuous adaptation to its Environment. Its adaptation occurs chiefly, if not entirely, through the Natural Selection of its useful Variations, and the tending of these variations to be transmitted to the offspring by the forces of Heredity. The summation of the variations, by Heredity and Isolation, leads ultimately to specific, generic, ordinal, and other differences in the descendants; leads, in other words, to the transmutations of organisms.
An organism consists of two great groups of structural units called cells,—Germ-Cells and Body-Cells,—harmoniously associated together. As far as Evolution is concerned, the most fundamental parts of an organism are the Germ-Cells.
In order that the reader may have a working knowledge of the wonderful powers of Germ-Cells and their progeny, it is vitally important that some concrete illustrations of the activities of different kinds of cells should be given.
In the sanctuary of S. Vitale at Ravenna, in Italy, is a very interesting representation, in mosaic pictures, and over life-size, of the Emperor Justinian and his Empress Theodora, attended by a numerous suite of ladies and courtiers. The mosaics are small bits of glass, of varying pattern and color, cemented together so nicely as to form beautiful delineations of the Emperor and his attendants.
The bits of glass or mosaics that form the figures may very appropriately be called structural units.
The body of man, a bird, a lizard, an oak tree, and many other animals and plants, may usefully be compared to such mosaic figures; for, just as the mosaic figure has its structural unit, the little bit of glass or stone, called the mosaic; so the bodies of men, birds, and other creatures may be looked upon as infinitely complex figures formed of minute mosaics called cells.
The groups of minute mosaics or cells that make up the bodies of animals and plants differ profoundly from the mosaics that form the figures of the dead Emperor and his companions, inasmuch as each mosaic or cell of the animal or plant body is so small as to require the microscope to reveal it; also[4] each mosaic of the living animal or plant is a living mosaic or cell, in that it can absorb food, digest it, assimilate it, grow, and multiply in numbers. Cells, then, are the morphological or structural units which compose the bodies of all living creatures.
All animals and plants begin life as single cells, which, in the vast majority of cases, are microscopic in size. Those that remain single cells, and dissociated throughout life, are called Unicellular Animals (Protozoa) and Plants (Protophyta), and are to be seen mostly by the microscope alone; but those which, by multiplication and growth, form large numbers of cells that remain associated together as in the body of a bird or lizard, are called Multicellular Animals (Metazoa) or Plants (Metaphyta).
A cell (Fig. 1) is a nucleated lump of protoplasm, or cytoplasm, and most often of microscopic size and more or less covered on its exterior by, and holding in its interior, various products and formations resulting from its activity, which are called metaplasm. Since the protoplasm of a cell, under the microscope, presents a superficial resemblance to a minute speck of that jelly-like substance (albumen) which forms the white of an egg, it is often called an albumenoid substance. But it is very misleading to use such an expression, for protoplasm is not a single chemical substance of great complexity; but it is rather composed of a large number of different chemical substances of great complexity. Many of these substances, it is true, are albumenoid in character. The same is true as to the chemical complexity of the nucleus, which is a physically and chemically differentiated part of the protoplasm.
The protoplasm contains certain globulins, and also albumins and peptones; it also contains large quantities of nucleo-albumins, with other substances. The nucleus not only contains these same substances, but also nuclein and nucleo-proteids. It is important to state that nuclein consists of an albumin and nucleic acid.
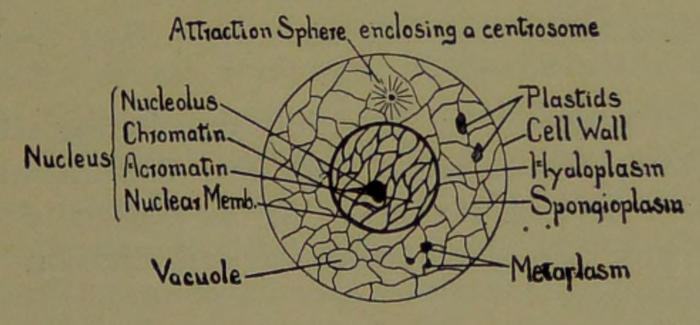
Fig. 1.—Diagram of a Cell, highly magnified.
The protoplasm, structurally, is made up of threads forming a complex, sponge-like substance, or reticulum, called spongioplasm; and in the meshes of the spongioplasm is a more or less fluid-like substance known as hyaloplasm: suspended in the hyaloplasm are various kinds of living bodies known as plastids, besides various products resulting from the activity of the protoplasm and which are designated metaplasm.
In many cells the protoplasm has formed on its periphery a layer of metaplasm which is frequently called a cell-wall. This cell-wall prevents amœboid movements of the protoplasm, and a cell possessing it is said to be encysted.
In many cells, especially vegetable ones, will also be observed clear spaces termed vacuoles. These vacuoles contain water with various chemical substances[6] held in solution, which serve the purpose chiefly of food-reservoirs.
The nucleus also is formed of threads called nuclear or chromatin threads (chromosomes), the interstices of which are filled with hyaloplasm or achromatin. In the nucleus can also be observed the nucleolus.
The protoplasmic and nuclear threads show various structural modifications in different regions and under different physiological states of the cell.
As will be observed later on, the nuclear threads are of special interest to the student of heredity. They may in one phase of cell-activity look like one thread forming an inextricable network, while in other phases they may look like thick, short, distinct rods.
The centrosome (Fig. 1), with its enveloping attraction-sphere, constitutes another fundamentally important part of the cell. It is especially concerned with the phenomena of cell division and multiplication.
Just as the living body consists of an infinitely complex figure of living mosaics termed cells, so the cell itself consists of an infinitely complex figure of still smaller living mosaics called, by Spencer, Physiological Units. These units have been given different names by various writers, viz.: by Darwin, gemmæ (gemmules); by de Vries, pangennæ; by Hertwig, idioblasts; by Weismann, biophors, etc., etc.
Like the atom of the chemist and the molecule of the physicist, the physiological unit of the biologist is merely at present an intellectual conception, yet it is, at the same time, an intellectual necessity and[7] plays a very important part as the theoretical component of many vital questions. Just as the cells are the visible units of life, so the physiological units are the invisible units.
The physiological activities of cells are those that pertain to their nutrition and reproduction.
The nutrition of cells includes all processes that are subservient to their life and well-being, such as irritability, contractility, absorption of food, its digestion and assimilation, secretion, etc.
In consequence of the wonderful nutritive activities of cells, we may well speak of them as marvelous magicians. Hertwig, following Haeckel, speaks of many cells as being builders. In the same spirit, we can say that multitudes of cells are expert chemists, artists, sculptors, mathematicians, and so on, in that they make all the myriad chemical products of organic nature, such as spices, pigments, sugars, starches, acids, perfumes, and numerous other substances; they paint in colors that rival the hues of the rainbow; they construct all of the beautiful forms in the animal and plant worlds; and they draw lines as straight and curves as graceful as the most expert mathematician.
One of the most important reproductive activities of a cell is mitosis (see below). Mitosis essentially consists of a series of processes by which each nuclear thread of the nucleus splits longitudinally into two equal parts, and then these equivalent parts separate from each other, so that from the one nucleus we get two smaller nuclei. Then each of these smaller nuclei appropriates its share of the enveloping protoplasm, finally splitting it into two parts. Thus from the larger cell (nucleated piece of[8] protoplasm) we get two smaller cells (two smaller nucleated pieces of protoplasm). In technical language, we say that the larger cell is the mother cell, and the two smaller cells that it has divided into are the daughter cells. In consequence of the method of mitosis, the two daughter cells very frequently are exactly like the mother cell, except in size. But by the absorption of nutriment, and through digestion and assimilation, they grow and finally become exactly like the mother cell. This is the simplest illustration of heredity. The reproductive process may be repeated very many times, so that from one cell we may get millions of cells.[1]
It is necessary to assume that the nutritive and reproductive activities of cells are based upon and controlled by the nutritive and reproductive activities of the physiological units, inasmuch as these are the ultimate living units.
In the activities of a cell the nucleus and protoplasm are intimately correlated with one another.
The nucleus is looked upon by the majority of cytologists as the formative center of the cell in a chemical, and also, consequently, in a morphological, sense. Active exchanges of material take place between the nucleus and the protoplasm during the nutritive processes of the cell. Possibly this may be altogether a chemical process, or possibly it may be due, as Hertwig suggests, to the migrations of the physiological units as carriers and elaborators.
In these exchanges, and in the upbuilding chemical activities (anabolism) of the cell, the nucleic acid plays a leading part. Here the nucleic acid in[9] the physiological units of the nuclear threads, combines with albumins from the protoplasm, forming nuclein. Much of this nuclein, undergoing further elaboration, is passed into the protoplasm as one of its finished products (metaplasm). The more purely nutritive the activity of a cell, the more nuclein its nuclear threads contain; on the other hand, when the cell is in the phase of reproductive activity, the nucleus contains little nuclein, and is almost entirely composed of pure nucleic acid.
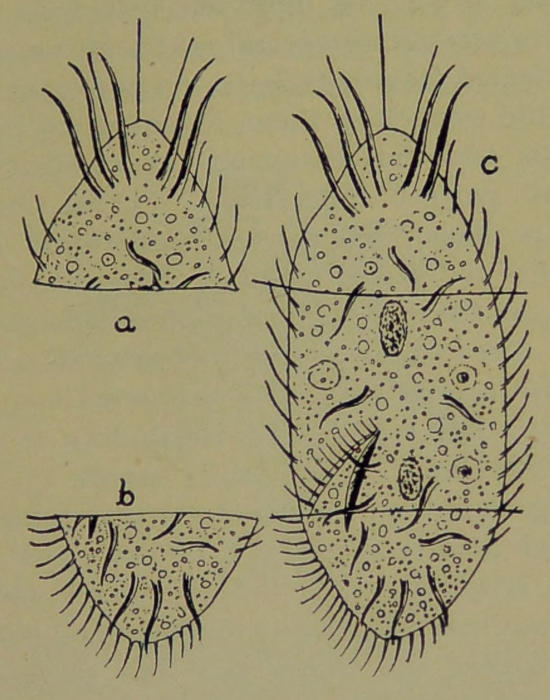
Fig. 2.—Stylonychia: c, an entire animal, showing planes of section; the middle piece of c contains two nuclei and can regenerate a perfect animal; a, and b, contain no nuclei,—they live and swim about for a while and then die.
That the nucleus is the formative center of the[10] cell is indicated by the following, among many facts: If a unicellular animal, such as Stylonychia (Fig. 2), for instance, be broken up into several fragments, it will be observed that some of the fragments are nucleated and others non-nucleated. The nucleated fragments have the power of quickly healing the wounds on them, regenerating the missing parts, and thus restoring the mutilated fragments to perfect individuals. These nucleated fragments have the power to perform all the activities of the perfect animal. The non-nucleated portions, on the other hand, cannot undergo regeneration. They cannot digest food, or grow or secrete substances as the nucleated fragments can. They can simply live for awhile, responding to stimuli and moving about. They finally perish.
Having mentioned in a general way some of the wonderful powers of cells, it will be well now to describe briefly a few of the unicellular plants and animals that can be so easily obtained and studied in warm weather, and which may thus serve as illustrations of the powers of nucleated pieces of protoplasm or cells. Many unicellular plants and animals can be obtained in summer from the superficial ooze on the bottom of slow-running streams and also on the under surfaces of the leaves of water plants, a study of which will be of the greatest value and interest.
Amœba proteus (Fig. 3). This little unicellular animal, which belongs to the Rhizopod type, is very common in ponds and streams in warm weather. In the resting state it is spherical in form, but when active its form is as changeable as the fabled Proteus, hence its name, Amœba proteus. This[11] little creature is a naked piece of protoplasm, with its outer layer differentiated into a firmer and pellucid part called the ectoplasm (Fig. 3, ec); its interior, the endoplasm (en), is quite granular and much more fluid, the granular particles moving quite freely upon one another when the animal changes its shape. The superficial portion of the endoplasm is firmer than its more central parts, and graduates insensibly into the more consistent ectoplasm.
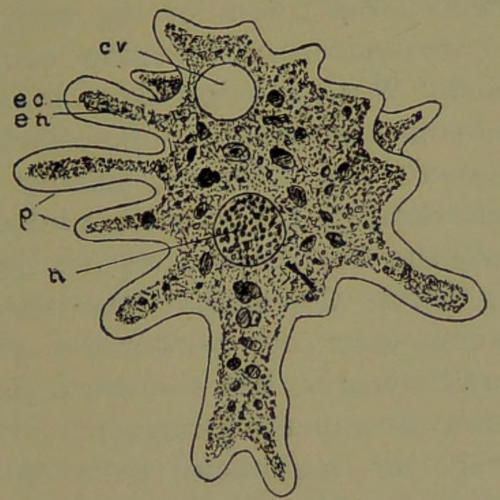
Fig. 3.—Amœba proteus: n, nucleus; cv, contractile vesicle; ec, ectoplasm; en, endoplasm; p, pseudopodia.
In the periphery of the granular endoplasm, and adherent to the inner surface of the ectoplasm, is a clearly defined nucleus (n). When most distinctly seen, it presents the appearance of a clear vesicle surrounding a solid and more or less spherical nucleolus. A contractile vacuole (cv) is also uniformly present, located in the endoplasm. The creature has the power of putting out projections (p)[12] from the surface called false feet (pseudopodia). Sometimes the protrusion consists of ectoplasm alone, but more commonly endoplasm extends into it, when a current of granules will be observed moving from the more central portions of the Amœba into its protrusion, whilst from some other protrusion that is being withdrawn a similar current may set towards the center of the body, and thus the animal moves, in a creeping manner, from place to place. While moving about in this way the little animal comes across other one-celled creatures, such as Desmids and Diatoms, seizing them and forcing them through its ectoplasm into the endoplasm, where the nutritious parts are digested and assimilated. After the animal has taken its prey through its ectoplasm, no break in the continuity of the ectoplasm remains, but the parts immediately come together in a perfect manner. After it has abstracted all the nutriment from its prey, the Amœba casts away from it the parts that are indigestible.
Fig. 4.—Rotalia Freyeri: a many-shelled Foraminifer, or a colony of many single-shelled Foraminifera, with pseudopodia extended.
Foraminifera. These are little protoplasmic unicellular animals that have the power of secreting[13] for themselves more or less complex envelopes composed of limestone. They may be single, as in Lagena, or composed of a number of individuals with the shells cemented together as in Globigerina or Rotalia (Fig. 4). They have played a part of vast importance in the geological development of the world. Their myriads of shells remaining at the bottom of seas millions of years after the little protoplasmic bodies have perished, they have been consolidated into vast expanses of limestone rocks, and finally uplifted into such formations as the huge chalk cliffs of England.
Osteoblasts and Osteoclasts. These cells are naked pieces of protoplasm, the latter much the larger and having many nuclei. They are concerned in some of the most interesting phenomena of many growing animals. Just as the Foraminifera have the power of forming complex aggregations of limestone shells, so the Osteoblasts have the power to construct the bones of animals. And when a bone is broken as the result of accident, these little cells do the mending. While the Osteoblasts are bone-formers, the Osteoclasts are bone-destroyers. It is very curious that little specks of living jelly, like these Osteoclasts, should have the power of destroying hard tissue like bone, but such is the fact. These Osteoclasts can, by their wonderful chemical processes, liquefy and absorb, and by these means destroy, ivory pegs that are driven into living bone. They are the agents by which the roots of children’s milk teeth are destroyed, so that the crowns of the teeth are shed and the way paved for the appearance of the permanent teeth. The wonderful activity of these little Osteoblasts and Osteoclasts is[14] well exemplified in the growth and shedding of the antlers of deer. While these antlers are growing in the spring, they are covered with a delicate skin, technically called “velvet.” This velvet is very sensitive and quite warm from the nutrient blood circulating through it. In it are hundreds and thousands of busy, living Osteoblasts that work together under some mysterious, directing or coördinating agency, to build up the splendid beams, tynes and snags that constitute the antlers, which in many deer of the Rocky Mountains reach such a size that a man may walk under the archway made by setting the shed antlers up on their points. No hive of bees is busier or more replete with active life than the antler of a stag as it grows beneath the warm, soft velvet, through the agency of the Osteoblasts.
The building of the antlers by these little agents continues through the spring and summer. In the autumn the Osteoblasts cease their activity and die; the delicate, sensitive velvet dries and peels off, leaving the dead, hard, bony substance exposed, and they now become weapons adapted for fighting. This is the season when the stags challenge one another to single combat, the hinds standing timidly by to be taken by the victor as his mates. When the loves and battles of the autumn are over and the mating is completed, the antlers no longer serve a useful purpose, and they are shed. The shedding is accomplished through the agency of the bone-destroyers, the little jelly-like cells called Osteoclasts.
Bacteria are exceedingly minute specks of naked protoplasm. They are unicellular plants. Some of them are harmless to mankind; some are very useful[15] to him, and others are his deadly enemies. Many of them are concerned in the production of the infectious diseases. They do so by elaborating various chemical products that are virulent poisons, hence these products are called toxines; when taken up by the blood, they are carried to various parts of the body. In this manner they cause the particular symptoms that are characteristic of a special infectious disease. Why is it that some persons, on exposure to an infectious disease, contract the malady while others similarly exposed do not? In other words, what gives immunity to disease? The explanation is probably as follows: Just as the invading bacteria have the power of secreting toxines, so the cells of the body, normally, have the power of elaborating chemical products that are antidotes to the toxines, and are appropriately called antitoxines. Infectious diseases and immunity from them, are the result of a contest between the invading bacteria and the protecting cells of the body. If the bacteria secrete toxines in greater quantities than can be neutralized by the cells of the body, we have disease; if the reverse occur, we have immunity.
The white blood-corpuscles (cells) also take part in this warfare. They have the power of traveling, in virtue of their amœboid movements, from the blood to the part invaded by the bacteria. Here a contest takes place between them, the corpuscle takes the bacteria into its interior, and either kills them or is itself killed. The result of this contest helps to produce either immunity or infection.
Tetanus bacillus is a cell shaped like a slender rod. It has the power of secreting a poison which, when[16] introduced into the body, produces convulsions and other symptoms of lockjaw. These much resemble those induced by strychnine poisoning.
Bacillus diphtheriæ is an exceedingly small unicellular plant, and has the power of producing a poison called toxalbumin, which is analogous to the poison of certain venomous serpents. It is the speck of protoplasm through whose activity diphtheria is caused.
Many useful bacteria have the power of so acting on dead organic bodies as to decompose them, the three most conspicuous end-products of this decomposition being water, carbonic oxide and ammonia. When the dead bodies are decomposed in the soil there are other bacteria, in addition, that have the power of further acting on the ammonia, causing its oxidation and producing nitrous and nitric acids and their salts. The unicellular plants that bring about these changes are the nitrifying bacteria. Conspicuous illustrations of the functional activity of these little naked pieces of protoplasm are seen in the immense saltpeter beds of Peru and Chili, where, from the enormous fecal accumulations of sea-fowls, the immense quantities of nitrates are produced that supply the commercial world.
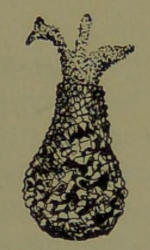
Fig. 5.—Difflugia Pyriformis.
Arcella, of which there are many species, is a unicellular animal whose protoplasmic body has secreted from its surface an enclosing “test” that is composed of a horny membrane, resembling very much in constitution the chitin which gives firmness to the integuments of insects. This creature is commonly discoidal in shape, with one face arched and the other flat, an aperture being situated in the center of the flat side through which the creature[17] may thrust its pseudopodia or withdraw them. The surface of the testaceous covering is often marked with a regular but minute and attractive pattern. In Difflugia (Fig. 5), the test is somewhat pitcher-shaped, and is mostly made up (by the constructive activity of the protoplasm) of exceedingly small particles of shell and gravel cemented together. Many testaceous amœbans form tests of singular beauty and remarkable regularity. In some of the animals the minute plates of which the tests are formed have been picked up from the surface over which the animals crawl, and are cemented into various charming patterns; and in other cases they are formed by secretion from their own bodies. In Quadrula symmetrica the protoplasmic body has constructed a pear-shaped testaceous covering, of complete transparence-like glass, composed of a great number of square plates touching each other by their edges. The protoplasmic body of the animal does not entirely fill the test, the intervening space being occupied by a clear liquid and traversed by bands of protoplasm. A clear, large spherical nucleus is seen in the part farthest from the pseudopodia. It contains a dark and well-defined[18] nucleolus. In front of the nucleus two contractile vesicles are to be observed. The pseudopodia in these creatures, it must be remembered, are not appendages, but lobate protrusions of the protoplasmic body, are few in number, rounded, short and broad.
Diatoms are unicellular plants, isolated or aggregated together, that have the power of constructing flint coverings, often of great complexity and charming pattern. The tracings on many of these flint coverings are so constant and small, that they are frequently employed for the purpose of testing the power of modern compound microscopes. In various parts of the world vast deposits of Diatoms have been discovered. The most remarkable of these for extent, as well as for the beauty and number of the species contained in it, is that on which the city of Richmond, in Virginia, is built, which is over thirty feet deep and extends for many miles.
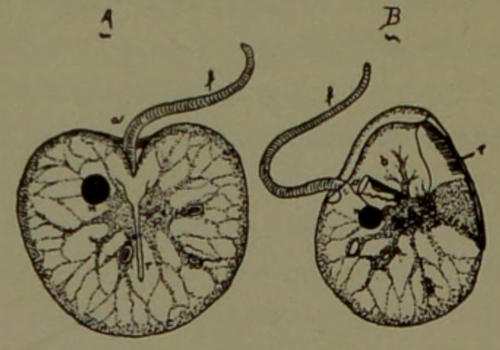
Fig. 6.—Noctiluca miliaris. A, dorsal view; B, side view; n, nucleus; f, flagellum; a, entrance to atrium; b, atrium; o, œsophagus; r, superficial ridge.
Noctiluca miliaris (Fig. 6) is a very large unicellular, flagellate animal. It is spheroidal in form, and has an average diameter of not quite one-half a millimeter. It is just large enough to be observed by[19] the unaided eye when the water in which the animal may be swimming is contained in a glass jar held up to the light. It has a tail-like appendage (flagellum) by which the animal moves about. Along one side of the cell is a meridional groove resembling that of a peach, and leading into a deep depression of the surface termed the atrium (Fig. 6, B, b). It is from the shallow commencement of this depression that the flagellum (Fig. 6, f) originates. At the base of the flagellum the depression sinks down to the mouth (o). A slightly elevated ridge (r) extends along the opposite meridian and commences with a bifurcation at that end of the atrium farthest from the flagellum. The mouth opens into a short œsophagus, which leads down directly to the central protoplasmic mass. The central protoplasmic mass sends off branching prolongations of its substance in all directions, the ramifications of which freely inosculate. The farther these ramifications extend out to the periphery, the thinner they become, until finally a protoplasmic network of extreme tenuity is formed immediately under the enveloping membrane of the cell. In addition to these ramifying prolongations, the central protoplasmic mass sends off a thin, broad, irregular extension to the superficial ridge and coalesces with it. Near the central protoplasmic mass is seen the nucleus (n).
The flagellum is a flattened, whip-like filament, having a striated appearance, and gradually tapers from the base to its extremity. It slowly bends over five or six times a minute to the mouth, and then, more slowly still, bends away again. It is through the movements of the flagellum that particles of food are driven into the mouth and down[20] the œsophagus into the central protoplasmic mass. In this mass and its extensions the food is digested and assimilated.
This little one-celled animal has the power, through its special chemical activities, of manufacturing and emitting light. It is through the agency of myriads of these little creatures that the diffused luminosity of some seas is produced and can be observed at night. The Noctiluca is very transparent, and for this reason it is a particularly favorable subject for the study of its luminosity or phosphorescence. They can be obtained by the tow-net in unlimited quantities from the sea and transferred into a jar of sea water. Here they soon rise to the surface, forming a thick layer. If the jar be placed in the dark and agitated in the slightest degree, there is an instantaneous display of light, which is of a beautiful greenish tint. The light emitted by the Noctiluca is so vivid that it can even be observed in ordinary lamp-light. This phosphorescence is only of an instant’s duration, and a short rest is necessary for its renewal. The special locality for the formation of the phosphorescence is in the very fine protoplasmic network, which lines the external structureless membrane or cell wall. These wonderful little Noctilucæ may well be figuratively called the fire-flies of the ocean.
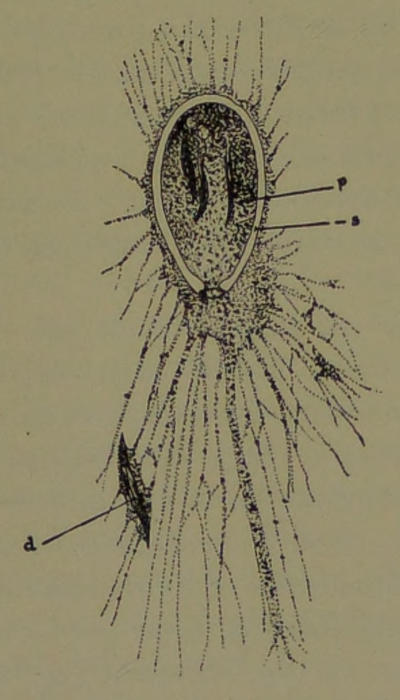
Fig. 7.—Gromia oviformis with protoplasmic threads (pseudopodia) extended and forming an elaborate network in which a captured unicellular organism is seen; d, diatom captured; p, protoplasm containing captured diatoms; s, shell.
Gromia oviformis (Fig. 7) is often found in fresh water adhering to confervæ and other plants of running streams. The protoplasmic body of this animal is enveloped in a chitinous covering that is egg-shaped and of a brownish-yellow color. It is about two millimeters in diameter. When the animal is quiet, no one would suspect its real nature, so much[21] does it look like the seed of an aquatic plant. The testaceous envelope has a single round orifice at its more pointed end. The animal, when in an active state, pushes out the protoplasmic substance, which speedily gives off ramifying extensions, and these by further ramification and inosculation form a complicated network. The protoplasm of the animal also extends itself in such a way as to form a continuous layer on the external surface of the test. From this layer numerous protoplasmic threads may extend out, forming more or less complicated networks. By the alternate contraction and extension of its protoplasmic threads and networks, minute[22] one-celled plants and animals are entrapped like flies in a spider’s web (d). When caught they are carried, by retraction of the protoplasmic thread-like pseudopodia, into the endoplasm in the test. Here the nutritious parts of the entrapped creatures are abstracted and assimilated. In transparent species, the indigestible parts, such as the silicious valves of diatoms, may be distinguished in the midst of the endoplasm, from which they are ultimately extruded.
When gromia oviformis reproduces by mitosis it gives off a bud (small cell), which finally separates from the parent form and constitutes a distinct individual. This process may be repeated many times, so that a great number of separate individuals may be formed, all of which lead detached and independent lives.
Most of the protozoa, which are produced by fission (cell division by mitosis), separate entirely from each other, as in Gromia; but in many of these unicellular animals, the new creatures produced by fission do not separate from one another, but remain more or less closely connected, and thus form colonies of Protozoans. These colonies are of the greatest interest, for they represent a lower stage of the cell colonies of the Metazoa (multicellular animals). They reproduce, in many cases, in a way which is strongly suggestive of reproduction in the Metazoa.
Microgromia socialis is a little unicellular animal, having a thin, nearly globular, calcareous shell that it secretes upon its surface. It multiplies by fission, and forms a number of distinct individuals which have the curious habit of fusing their pseudopodia and uniting into a more or less closely associated[23] colony. The individuals sometimes remain at a distance from one another, but sometimes associate themselves together into a compact colony. These individuals are all alike, performing the same functions. There is no division of labor among the units, but they live practically an independent life. If the individual animals were detached from one another, they would live and build new colonies.
Codosiga umbellata is another unicellular animal—a flagellate Protozoan. It has a collar-like extension of its ectoplasm from the anterior extremity of its body, forming a sort of funnel from the bottom of which the thread-like structure (flagellum) arises. The vibrations of this flagellum cause a current of the surrounding fluid to set into the funnel so that particles of food reach the soft protoplasmic substance which serves as a mouth. The nucleus is seen near the base of the collar. Near the posterior extremity of the body two contractile vesicles are to be observed. This posterior extremity of the animal has a cylindrical extension of its ectoplasm by which it attaches itself to an object. This protozoan multiplies by longitudinal fission. In some species the animals separate completely from one another and lead entirely independent lives. But in Codosiga umbellata the fission does not extend through the cylindrical extension, so that a group of animals are associated in a colony.
Rotalia or Globigerina. The shelled amœba (Lagena) gives off a bud (unseparated cell) which grows to the full size, secretes its calcareous shell and remains connected with the parent form. This process may be repeated a number of times until a colony of shelled amœbæ, of varying pattern in[24] different species, may be formed and permanently associated together (Fig. 4). The individual amœbæ are all alike and perform the same functions. There is no division of labor, no specialization, among them. If the individual animals could be separated from one another they would live and build new colonies.
Pandorina morum forms a small colony of sixteen cells (solid sphere) of mulberry-like shape and enclosed in a common gelatinous envelope. Each cell in the mulberry mass bears two flagella on its peripheral end. These project out beyond the surface of the gelatinous envelope, and are agents for locomotion of the colony. The cells in the colony are all alike. There is no division of labor among them. They all act alike. The cells (flagellate protozoans) of the colony may reproduce in two ways. Each animal in the colony may subdivide into sixteen smaller units, each of which by growth and multiplication may form a new mulberry mass, a new colony, each unit of which acquires two flagella. Or two of the small units may amalgamate (conjugate), and then develop (by fission) into a new colony. The conjugating units are nearly of the same size and look very much alike.
Volvox globator is a spheroidal shaped colony (hollow sphere) of unicellular flagellate animals, about one-half a millimeter in size. It was formerly supposed to be a fresh-water Alga. It is now known to be a colony of Protozoans. All the animals in this colony are not alike. There is a division of labor among the cells, for some are merely vegetative, serving purposes of nutrition, and having no reproductive powers; while other[25] members of the colony are purely reproductive animals. Furthermore, there is quite a marked specialization of the reproductive cells. Those reproductive cells that may be spoken of as the female cells are large and non-motile encysted cells. The male cells are small and actively motile, in that they have two flagella developed on them. A small flagellate male cell penetrates the large encysted female cell, and as the result of this conjugation, fission takes place repeatedly, and a new colony of flagellate protozoans (volvox) is formed. Volvox approaches very suggestively towards the type of animals known as Metazoan.
In order to comprehend, in some measure, the transition from Colonial Protozoa to Metazoa, it will be well for the reader to study a typical sponge. For a long time the Porifera (Sponges) were looked upon as compound Protozoa (colonial Protozoa), but while they are nearer the Protozoa than any of the other types of Metazoa, their position in the animal series is unquestionably among the Metazoa. The Sponge, like the rest of the Metazoa, develops from a fertilized egg by a process of cell multiplication, differentiation, gastrulation, etc.
The multiplication of cells plays a part of such fundamental importance in Evolution, and therefore in Embryology and studies in Heredity, that it is necessary to study the subject somewhat in detail. It is a wonderful process, and is worthy of very careful attention.
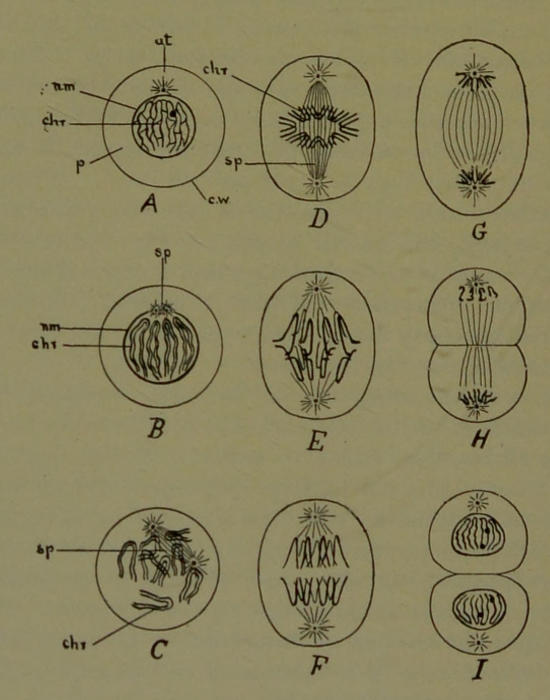
Fig. 8.—Diagram illustrating Mitosis. A, the cell commencing activity; B, C, D, phases in the formation of the spindle and the chromatin loops or V’s, also showing that the mother V’s have split into daughter V’s; D, the chromatin loops forming the equatorial plate, chr; E, F, G, separation of the daughter loops (daughter chromosomes) and their passage towards the poles of the spindle, thus forming daughter nuclei; H, I, division of the protoplasm so as to form two daughter cells; at, attraction sphere enclosing a centrosome; n m, nuclear membrane; chr., chromatin threads; p, protoplasm; c w, cell wall; sp, spindle.
The process by which one cell (a mother cell) divides into two cells (daughter cells) is called mitosis, and is inaugurated by the centrosome (Fig. 8, A, at). The centrosome divides into two centrosomes, which at first remain close together (Fig. 8, B, sp), and then gradually separate from one another. Each centrosome becomes the center of a system of fine achromatin fibers arranged round it in a radiating manner and forming what is called the attraction sphere; also, at the same time, a spindle-shaped bundle of achromatin fibers, called the spindle (Fig. 8, B, sp), extends between the centrosomes. In the meantime, important changes have been taking place in the chromosomes (hereditary threads) of the nucleus. The chromosomes, which at first are arranged in an apparently inextricable tangle or network, frequently assume U-shaped or V-shaped forms (Fig. 8, C, chr), and the nuclear membrane disappears. Sooner or later each chromosome splits longitudinally into two daughter chromosomes, with which the achromatin fibers of the spindle become connected (Fig. 8, D). In this phase of mitosis the split V-shaped chromosomes form a single group called the equatorial plate (chr), and extend across the axis of the spindle. It is to be observed from the diagrams in the figure, that one of the centrosomes has traveled to the opposite pole of the nucleus, thus causing the achromatin fibers of the spindle to extend across the original site of the nucleus. The equatorial plate of split V-shaped mother chromosomes (hereditary threads) thus divides the fibers of the spindle into two parts, one half extending from one centrosome to one group of daughter chromosomes, while the remaining half extend from the other centrosome to the[28] other group of daughter chromosomes. Soon the achromatin fibers of the spindle contract, and in this way separate the two groups of daughter chromosomes, so that one group is drawn towards one centrosome, and the other group to the other centrosome (Fig. 8, E, F, G, H). After the two groups of daughter chromosomes have been drawn to their respective centrosomes, each group assumes the tangle or network phase like the nucleus of the mother cell, and an investing nuclear membrane reappears for each (Fig. 8, I). Thus from the mother nucleus of the mother cell we get two daughter nuclei (I). In a further phase of the mitotic process, a furrow appears on the surface of the protoplasm and surrounds it in the form of a ring. This furrow is in a plane at right angles to the long axis of the spindle, and gradually deepens until the protoplasm is divided into two parts, each segment of protoplasm containing its own nucleus and centrosome; in short, the mother cell has divided into two daughter cells (I).
It will thus be observed that the centrosomes and their achromatin fibers are a beautiful mechanism by which the heredity threads (chromosomes) are exactly divided into two equivalent halves.
There are some cells (Amœba proteus, for instance,) which divide in a much simpler manner than by mitosis; in these there is no complicated rearrangement of the chromosomes and no disappearance of the nuclear membrane, the nucleus simply becoming separated into two parts (Amitosis).
The Human Ovum. The human ovum is a typical cell about one-fifth of a millimeter in size and spherical in shape. It is a nucleated piece of protoplasm[29] possessing an enveloping cell-wall (metaplasm). The protoplasm contains nutrient material or yolk (metaplasm). The maturation of the ovum essentially consists in throwing out half of its chromosomes. In doing this the nucleus (mother) approaches the surface of the protoplasm (Fig. 9, A), and divides, by mitosis, into two daughter nuclei; then the unripe ovum divides into two cells, but of very unequal size. This process is repeated a second time. Thus two small cells are formed which are known as the polar bodies (Fig. 9, A, B, pol. b). The large cell remaining after the formation of the two polar bodies is the mature ovum (B). Its nucleus, which recedes towards the center of the protoplasm, is called the female pronucleus (Fig. 9, B, f. pr.). The pronuclei of mature ova differ from the nuclei of all the other cells of the body in that they only contain half as many chromosomes (hereditary threads).
The ripe spermatozoid (a flagellate sexual cell of the male) corresponds to the ovum of the female. It also contains a pronucleus having only half the number of chromosomes that the other cells of the adult body possess.
Fertilization. The male sexual cell (spermatozoid) is vastly smaller than the female sexual cell (ovum). Having a flagellum (Fig. 9, B, s), it moves about, like a tadpole in water, and seeks the ovum. When it comes in contact with the ovum it penetrates into its interior (usually only one doing so), as indicated at (Fig. 9, B, s). The tail or flagellum of the spermatozoid fuses with the protoplasm of the ovum, and disappears from view. Its pronucleus (C, m. pr.), accompanied by its centrosome (C, m. c.),[30] approaches the female pronucleus (f. pr.) of the ovum (Fig. 9, C, D). Finally the male and female pronuclei coalesce to form a single nucleus (Fig. 9, E, n. os.). The centrosome of the ovum persists for awhile and then disappears; that of the spermatozoid remains in the ovum, and is the agency by which cell multiplication, through mitosis, takes place.
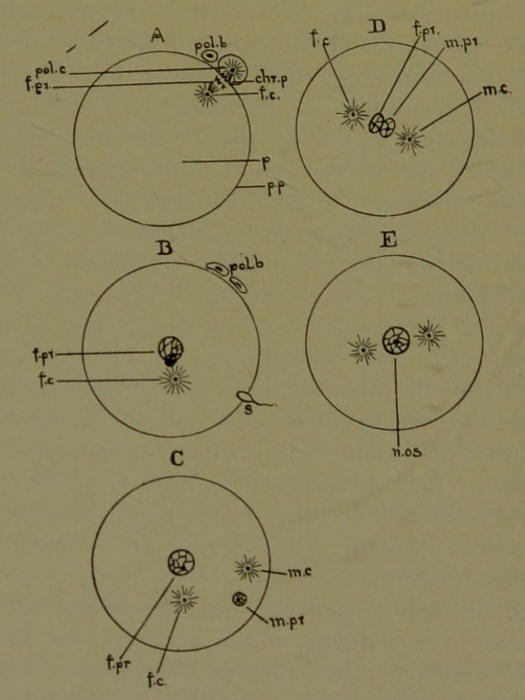
Fig. 9.—Diagram illustrating the maturation and fertilization of the human ovum. A, one polar body is formed and a second is in process of formation; B, both polar bodies are formed and a spermatozoid is penetrating the ovum; C and D represent the approach of the male pronucleus towards the female pronucleus; E indicates the amalgamation of the two pronuclei to form the nucleus of the oösperm (segmentation nucleus); pol. b, polar bodies; pol. c, centrosome of the polar body; chr. p, chromatin of the polar body; f pr, female pronucleus; p, protoplasm; p p, peripheral protoplasm (but not cell wall); f c, female centrosome; m c, male centrosome; m pr, male pronucleus; n, os, nucleus of the oösperm (first stage of a human being).
The cell resulting from the coalescence of the male pronucleus with the pronucleus of the ovum, and which is only one fifth of a millimeter in size, is the first stage in the existence of a human being. Man thus starts his career as a Protozoan-like creature,—as a unicellular animal. The fertilized ovum is called the sperm-egg (oösperm), and contains now the normal number of hereditary threads (chromosomes); for those of the male have been added to those of the female.
Segmentation of the Oösperm. The fertilization of the ovum imparts to it a wonderful stimulus, so that the oösperm divides, by mitosis, into two cells, these two into four, the four into eight, the eight into sixteen, these into thirty-two, and so on repeatedly, until a large number of comparatively small cells are formed (Fig. 10). This mass of cells is spherical in shape, and the little round cells towards the surface project in such a way as to give to the mass an appearance somewhat similar to the fruit of the mulberry, whence it is termed the mulberry body or morula (Fig. 10, 4). In the morula stage of his existence man resembles the solid colony of protozoans represented by Pandorina. The cells of the morula next become arranged regularly in a single layer at the circumference, by which the embryo[32] assumes the form of a hollow sphere, and is known as the blastula (Fig. 10, 5). This phase of man’s existence is quite suggestive of Volvox.
Soon one side of the blastula is invaginated or pushed in, as one would push in one side of a hollow india-rubber ball. The result of this invagination (called gastrulation technically) is the formation of a sort of cup. This is the gastrula (Fig. 10, 6) phase of man’s existence. It is a higher and fundamentally different phase of existence than either the morula or the blastula. It corresponds, not to a Protozoan, but to the higher Metazoan. It possesses the fundamental anatomical qualities of a low Cœlenterate (Polyp).
The mode of gastrulation is different in man from that just described, and varies in different animals, but the essential point common to all is the formation of a double cellular-membrane; the outer membrane being called epiblast or ectoderm, and the inner one hypoblast or endoderm, the enclosed cavity being the primitive digestive cavity. These two layers are the primary germinal layers. A third layer is subsequently formed between them, by the agency of one or both of them, and is called the mesoblast. From these three simple membranes, which are composed exclusively of cells, are formed all the complex tissues and organs of the adult man. The epiblast develops into the nervous system (brain, spinal cord, and nerves), and the cuticle, hair, etc. The hypoblast develops into the cellular parts of the digestive canal, the liver, lungs, etc. The mesoblast develops into the muscles, bones, ligaments, blood-vessels, etc. To trace the details of this evolution of a human being from the microscopic[33] oösperm is a fascinating and instructive study, but is beyond the limits and purpose of this little book. It will well repay further careful study by the reader.
A careful study of the life-histories of the few unicellular animals and plants mentioned in the preceding pages will help to let one realize the wonderful powers of nucleated pieces of protoplasm (cells). If isolated pieces of protoplasm can accomplish so much, one will not be astonished to learn that many diverse cells associated together in intimate correlations, as occurs in the higher animals, may accomplish results that are profoundly interesting and marvelous. As far as anatomy and physiology alone can reveal, it is the result of millions of cells acting together that makes possible the existence of such a living, sentient, thinking creature as man or the highly intelligent elephant or any other multicellular animal. It is due to the mysterious powers of protoplasm that one little microscopic cell, like the fertilized ovum of a woman, is able to hold all the heritages of the race, and gradually unfold them as it builds up the body into myriads of diverse cells intimately associated together.
All of the wonderful results of Embryology are accomplished through cell-multiplications, cell-differentiations, cell-associations, invagination and evaginations of cell-groups (tissues or organs), and unequal growth of parts (cells or groups of cells).
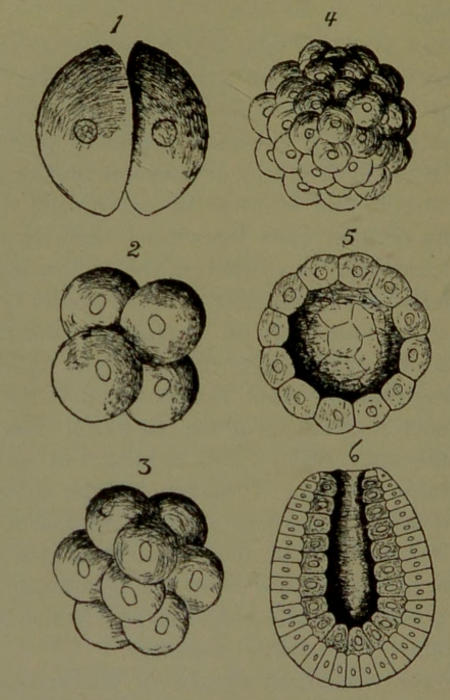
Fig. 10.—Segmentation of the fertilized ovum and Gastrulation: 4, morula; 5, section through blastula showing hollow sphere; 6, gastrula showing outer layer of cells (epiblast) and inner layer (hypoblast); the 6 is at the mouth of the cavity (enteron) of the gastrula.
In concluding this brief but, we hope, useful study of a few selected cells, we may say that an eminent English physiologist has made the statement that a student who has not looked through the microscope and observed the circulation of the blood in the[34] web of a frog’s foot is not fit to study medicine. However beautiful, fascinating and instructive the sight of this circulation may be, we are tempted to make the assertion that the student who has not looked through the microscope at some of the superficial ooze from the bottom of any slow-running stream, in summer, and observed the structure[35] and actions of that wonderful little unicellular animal, the Amœba proteus, is still less prepared to study medicine. In this little speck of protoplasm the problems of life are reduced to their simplest forms, for all higher plants and animals may be regarded as groups of more or less modified amœbæ peculiarly associated together. In our studies of the amœba we will be forcibly reminded of a very clever trick which is practiced in India and is called the mango-trick. In this trick a seed is put into the ground and covered up, and after divers incantations a full-blown mango-bush appears within five minutes. We have never met any one who knew how this thing was done, nor have we ever seen a person who believed it to be anything else than a conjuring trick. So it is with the amœba, a beautiful and fascinating trickster of nature. We understand some of its activities, interesting and exceedingly instructive, but there are many others beyond our ken. We see the commencement and ending of many of its chemical activities, but there are numerous other intermediary processes that take place in the hidden recesses of the protoplasm, and concerning which we know nothing. It may fall to the lot of some reader of this little book, as a patient and keen observer, to unravel some of these mango-like tricks of the amœba or other unicellular creature.
That an offspring always inherits from its parents many of their characteristics is well known; that it always varies, more or less, from them is also equally well known. Heredity and variation are twin forces that play upon every creature, holding it rigidly true to the parental type or compelling more or less divergence therefrom, according to the strength of the one or other power; so that every creature is the resultant of the activities of these two great parallel forces. Variation is coextensive with heredity, and every living creature gives evidence of the existence of variations.
Examples of Variations. No two leaves on a plant are exactly alike; no two children of the same parents give a perfect resemblance; no two individuals of the same species are molded in precisely the same pattern; of the thousands and thousands of faces that we observe in a city in the course of a year, each has some distinctive peculiarity.
The trained eye of the gardener recognizes each hyacinth among hundreds of bulbs; of the shepherd, each sheep in his flock; of the Laplander, each reindeer crowded in his herd like ants on the anthill. In a flock of 1,000 sheep each mother can even recognize a variation in the voice of her own lamb, all alike to us.
Every part of an animal is subject to variations, not only in bodily structure, but also in habits and instincts, and these variations are large in amount, numerous and diverse in character. Many observations, experiments and measurements that have been made at various times attest the truth of this assertion. Not only do variations take place in animals and plants under domestication, but also in the wild state.
Illustrations of Heredity. Mental heredity can be illustrated by studying the genealogies of such persons as Aristotle, Goethe, Darwin, Coleridge, Milton, etc. Probably the Bach family, of Germany, supply one of the best illustrations of the inheritance of intellectual character that we know of. The record of this family begins in 1550, lasting through eight generations to 1800. For about two centuries it gave to the world musicians and singers of high rank. The founder was Weit Bach, a baker of Presburg, who sought recreation from his routine work in song and music. For nearly two hundred years his descendants, who were very numerous in Franconia, Thuringia, and Saxony, retained a musical talent, being all church singers and organists.
When the members of the family had become very numerous and widely separated from one another, they decided to meet at a stated place once a year. Often more than a hundred persons—men, women and children—bearing the name of Bach were thus brought together. This family reunion continued until nearly the middle of the eighteenth century. In this family of musicians twenty-nine became eminent.
Inheritance of moral character is well established.[41] Heredity, in its relation to crime and pauperism, has been thoroughly investigated by Mr. Dugdale in his most instructive little work entitled The Jukes. In this work the descendants of one vicious and neglected girl are traced through a large number of generations. It reveals that a large proportion of the descendants of this woman became licentious, for, in the course of six generations, fifty-two per cent. of the females became harlots and twenty-three per cent. of the children were illegitimate. It shows also that there were seven times more paupers among the women than among the average women of the State, and nine times more paupers among the male descendants than among the average men of the State.
The inheritance of physical peculiarities is so obvious as to need no illustration. Among the ancients the Romans stereotyped its truth by the use of such expressions as the labiones, or thick-lipped; the nasones, or big-nosed; the capitones, or big-headed; and the buccones, or swollen-cheeked, etc. In more recent times we read of the Austrian lip and the Bourbon nose.
Questions of heredity and variation are cytological ones—that is, questions of the anatomy, physiology, physiological chemistry, and pathology of cells. The most important part of a cell, as far as these questions are concerned, is the nucleus. The nucleus is the physical basis of all the heritages of an organism, from the simplest to the most complex. The nuclear threads may, therefore, very appropriately be termed the hereditary threads, or, collectively, the hereditary mass; and the physiological units in them the hereditary units. The nucleus is[42] of fundamental importance in the reproduction or multiplication of both unicellular and multicellular animals and plants.
In unicellular creatures multiplication may take place by fission and by conjugation. Both of these processes can be studied by observation of the infusorians. Maupas’s beautiful investigations on these unicellular animals have demonstrated that multiplication by fission may proceed to a prodigious extent for many generations, but that a time comes when the process fails, and the species will become exhausted and die out unless there is a rejuvenation of it by conjugation of individuals. In conjugation two individual infusoria come in apposition with each other, the nucleus in each undergoes subdivision. They reciprocally exchange part of their nuclear contents so that each infusorian comes to contain hereditary threads of two distinct individuals. From these rejuvenated (or fertilized) individuals multitudes of others may be derived by fission until exhaustion again takes place.
Multiplication in multicellular creatures may be accomplished by budding (which is allied to fission), and is exemplified in the plant, hydra, the queen bee (parthenogenesis), etc., and by fertilization (which is allied to conjugation). A knowledge of the phenomena of fertilization of the ovum by the spermatozoid is essential to any understanding of the problems of heredity and variation in mankind. The nuclear threads of the ovum are its hereditary threads—the groups of maternal hereditary units; likewise, the nucleus of the spermatozoid contains the paternal groups of hereditary units.
Fertilization. In fertilization, the spermatozoid[43] (a nucleated flagellate cell) penetrates the ovum (a nucleated, encysted cell), its protoplasm mixes with that of the ovum, and its nuclear threads come into relation with the nuclear threads of the ovum; so that the fertilized ovum (a new creature, a veritable microcosm) is still a nucleated cell, but one in which the nucleus is compound, is hermaphroditic, in that it contains maternal and paternal threads—that is, maternal and paternal hereditary units which constitute its hereditary mass.
It will be convenient to speak of the maternal and paternal hereditary units in the fertilized ovum as ancestral hereditary units.
This hermaphroditic cell passes through complex phases, illustrated by embryology, to the adult. In doing so this hermaphroditic cell (mother cell) first divides into two smaller cells (daughter cells). The mother cell divides in such a way (by mitosis) that one-half of its nucleus and part of its protoplasm goes to one daughter cell and the other half of the nucleus, with the remainder of the protoplasm, goes to the other daughter cell. It is an interesting fact that although the amount of protoplasm which goes from the mother cell to the two daughter cells may be unequal at times, yet the amount of the nucleus in one daughter cell is always exactly equal[2] to that in the other; so that each daughter cell contains maternal and paternal hereditary masses of equal quantity and quality, being in fact one-half that of the fertilized ovum. In consequence of the fact that each hereditary unit in the nucleus of the daughter cell can absorb nutriment and grow, it[44] comes about that the nucleus of each daughter cell attains to the size of that of the mother cell. The enveloping protoplasm of the nuclei also grows to a greater or less extent, so that the cells as a whole grow. These two daughter cells go through the same process and form other daughter cells, and so on through all the mitoses of development, until all the myriads of cells of a living organism are produced, each of which contains maternal and paternal hereditary masses of equal quality and quantity, and also of the same character as that of the fertilized ovum whence they are all derived.
Apart from their activity in absorbing nutriment and growing, the great majority of the hereditary units in the nuclei of the forming cells remain latent. But some of the hereditary units in each cell produced are active. They multiply, grow and migrate out of the nucleus, and get among the units of the enveloping protoplasm. During this activity they undergo physical and chemical changes and effect corresponding differentiations in the protoplasm of the cell. Thus, through many mitoses and many differentiations of the protoplasm of cells, we finally derive from the fertilized ovum all the cells that constitute the adult body, such as muscle cells, glandular cells (as liver cells, kidney cells, etc.), nerve cells, skeletal cells (as bone, cartilage, and connective-tissue cells), and the ova and spermatozoids. According to this theory, the nucleus of each cell in the adult animal or plant is pure hereditary mass, exactly like that in the nucleus of the fertilized ovum; but the protoplasm of each adult cell that envelopes the nucleus may differ greatly in different cell groups, as in muscle, nerve, cartilage,[45] and the like. Of course, the protoplasm of those cells that develop into the ova and the spermatozoids has differentiated along such lines as to become like that of the ovum and spermatozoid, the junction of which formed the fertilized ovum. These statements hold true for plants and the lower animal organisms, although they cannot be verified for the higher animals. More than likely the pure hereditary masses are present in the body (somatic) cells of the higher organism in latent conditions, but are unable ever to be developed owing to the greater specialization of these cells. It is thus seen that all the cells of many animals and plants can perform their own special functions and at the same time contain all of the hereditary units of the complex organism in a state ready to develop under favoring conditions.
Since the ova and spermatozoids are cells specially differentiated for the purpose of propagating the species by sexual generation, and since their conjugation produces the germ of a new creature, they may very appropriately be spoken of as germ cells. Since all the other cells of the adult form the great bulk of the body that envelopes and protects the germ cells, they may be termed the body cells, or somatic cells.
Suppose that ova, containing maternal and paternal groups of hereditary units, are fertilized by similarly complex spermatozoids, and the process is repeated generation after generation. There will come a time when the fertilized ovum will have a highly complex nucleus composed of many different ancestral groups of hereditary units.
One often hears the expression that a child is a[46] chip of the old block; but this is only a very partial truth, for a child is preëminently a composite chip of many old blocks.
The complex nucleus of the fertilized ovum may be compared to a modern Italian building which has been constructed of material—a column here, a cornice there, a lintel yonder—gathered from different classic buildings of varying antiquity. In view of the increasing number of ancestral groups of hereditary units that must have accumulated in the nuclei of ova in the course of time, there must necessarily, for mechanical reasons, have arrived a period when these nuclei could receive no more of them by fertilization, unless natural selection should develop some saving device; hence we have, possibly, an explanation of the phenomena of maturation in ova (the reducing process of Weismann and Hertwig). Here the ovum, prior to fertilization, undergoes mitosis twice in succession, by which the polar bodies are formed and the hereditary mass is diminished by one-half (the mature germ cells having only one-half the number of nuclear threads that the body cells possess). A homologous process takes place in the maturation of spermatozoids. Fertilization increases the amount of the hereditary mass in the ovum to the original quantity, and thus restores the number of nuclear threads to the specific number. All the body cells derived from the fertilized ovum possess, also, the specific number of threads.
This union of two distinct hereditary masses is called amphimixis as well as fertilization.
Maturation and amphimixis or fertilization are the source of many variations in the body, good and bad,[47] beautiful and ugly, geniuses and monstrosities; because, in the commingling of distinct hereditary masses, there is a struggle for existence between the hereditary units and a survival of the fittest.
In this struggle some of the hereditary units are strengthened so that heritages may be augmented; some mix so that there may be a blending of characteristics; some are mutually exclusive; some are prepotent; some are neutralized; some are destroyed; some lie dormant (latent) for varying lengths of time, and some are so altered as to produce much modified forms; and thus the possibilities of combination, reactions and modifications of the hereditary units, and therefore heritages, are almost endless.
The augmentation of heritages in the fertilized ovum is well displayed, for instance, where fleet horses are bred with fleet ones, until, by careful selection, generation after generation, a progeny may be secured much more swift than the original stock from whence they were derived. In the same way good milch cows have been produced.
The mixing of hereditary units and blending of heritages is shown in the color of the skin, as where a mulatto child is born to a negress by a white father; mutually exclusive heritages are well illustrated in the color of the eyes, as where a child has either the blue eyes of one parent or the black of the other, but never any blending of the colors; this may also be illustrated where the white game bird and black one are crossed, the young being either white or black, but never blended. Prepotency is illustrated where the silky variety of fan-tailed pigeon is mated with any other small-sized variety of pigeon, for the silkiness is invariably transmitted.[48] A most interesting case of prepotency in mankind, mentioned by Ribot, is that of Lislet-Geoffrey, an engineer in Mauritius. He was the son of a very stupid negress and an educated white man. In physical constitution he was as much a negro as his mother; he had the woolly hair, the features, the complexion, and the peculiar odor of his race. He was so thoroughly a white man as regards intellectual development that he succeeded in vanquishing the prejudices of race, so strong in the French colonies, and in being admitted into the most aristocratic houses. At the time of his death he was Corresponding Member of the Academy of Sciences.
In this, it will be observed, we have prepotency in the mother’s physical constitution, and in the father’s intellectual characteristics.
The struggle of heritages in the impregnated ovum may lead to such structural changes of the nucleus, and therefore of the cell, as to develop the most marked variations—such variations as the biologists call sports.
In the latter part of the eighteenth century the farmers of Massachusetts had flocks of ordinary sheep on their farms. These sheep were continually jumping fences and getting on neighboring farms. They were the source of many disputes and much irritation between neighboring farmers. Finally, one of the sheep had a lamb which, when grown, displayed well-marked peculiarities (a sport). It had a longer body than the ordinary sheep and shorter legs, which were bowed. It was noticed that this sheep could not get over the fences. The cute Yankee farmer, noticing this valuable peculiarity, carefully preserved this peculiar sheep, and from it[49] was ultimately derived, by careful selective breeding, a special variety known as the Ancon sheep.
The germinal variations resulting from the mixing of two separate hereditary masses by impregnation find their expression in the most varied qualities of the minds and bodies of developing children. If the variations are not especially marked, they are looked upon as normal and attract no special attention.
But if the variations are so pronounced as to compel attention, and at the same time it is known that they are useful, they are spoken of as talents, or, on the other hand, if they are harmful or useless, they are designated as pathological or monstrosities.
These are truly what the biologists call sports; and to those classes of sports that occur as specially gifted in human culture, in the varied fields of science, art, or literature, we assign such a person as Shakespeare, and call the remarkable variations embodied in him genius. On the other hand, such variations as lead to certain forms of pigmentary degeneration of the retina, and to Daltonism, to dyschromatopsia and achromatopsia, to certain supernumerary glands, polydactylism, and such like, which are either useless or harmful, we designate as pathological cases or monstrosities.
Hereditary units (the carriers of heritages) may be latent—that is, they may appear late in life, or in the offspring, or, still again, in remote descendants; in the latter cases the heritages are spoken of as reversional or atavistic. Latent hereditary units may very usefully be compared to dormant seeds buried in the ground. It is stated that buried seeds may lie dormant for many years, so that when a[50] plot of ground is plowed deeply and upturned, plants that have not been seen there within the memory of one will often make their appearance and flourish. The hereditary units are veritable living seeds, that, under certain and often unknown stimuli, grow and unfold their heritages as do the buried seeds.
Latent heritages are well illustrated by a study of secondary sexual characters as developed at puberty.
Among our barnyard fowls, the hens often, when they have atrophy or degeneration of the ovaries, although up to this time they have laid eggs for years, stop this function, put aside the plumage and appearance proper to their sex, and don more or less completely the garments of the rooster. Thus females have latent in them many secondary sexual characters of the male. For similar reasons the male develops, occasionally, female characters.
This latency is illustrated again in deer. In most species of the deer tribe the males alone possess antlers, yet it is a well-known circumstance that in females with degenerations of the ovaries rudimentary horns that are never shed appear. A study of congenital color-blindness illustrates beautifully latent heritages, showing how the females of one generation may be free from the malady and the males of the next afflicted.
A study of the regeneration of lost parts in various animals and plants illustrates well the latency of many hereditary units. A cutting made from the willow and planted sends out roots and finally reaches the dimensions of the adult tree. Here the body cells of the stem evidently contained, in a latent condition, many hereditary units in their[51] nuclei, which became active through the special stimulus of being planted as a cutting. Adult plants can be raised from the cuttings of many other plants.
If the garden worm is cut in two, the head-part will reproduce the tail-portion. If the little fresh-water polyp, Hydra, be cut into a number of pieces, each segment will reproduce a perfect animal. Many lizards, after losing their tails by violence, manufacture new tails through the agency of the latent hereditary units contained in the body cells of the stump left. If the tentacle or “horn” of a snail, which contains an eye with a perfect lens and retina, be cut off, the animal can reproduce another one with a perfect eye, and this can be repeated a number of times. Often newts, when fighting with one another, or lobsters when fighting, lose a leg or a claw. These highly organized animals have the power of creating new limbs, making bones, ligaments, muscles, nerves, cuticle, and so on. All of this is done through the hereditary masses in the nuclei of the body cells at the site of the injury.
A study of the phenomena of polymorphism in hydroids and insects will beautifully and most interestingly illustrate latent hereditary units.
Much that is speculative and fanciful is included under the subject of atavism, and the safest plan for pathologists and biologists, in considering any abnormality, is to remember a golden rule of Gegenbaur, that only those structures are reversional which are taxonomically not far distant or phylogenetically not very old. Embryology is also a very important check in considering such subjects.
In mankind supernumerary limbs and digits,[52] microcephalia and micrencephalia, have been looked upon as reversions to the simian type.
Lombroso, in contrasting the criminal with normal man, looks upon his homo delinquens as an illustration of atavism, contrasting with homo sapiens. But, as Ziegler, in his Pathology, well observes, many writers have gone too far in this respect, and have characterized as atavistic formations various acquired pathological formations and fresh variations of germ cells.
I think one can safely say that supernumerary ribs and those supernumerary nipples and mammary glands along the line of the deep epigastric and internal mammary arteries are truly atavistic structures; also certain muscles normally belonging to those mammalia which come near to man in the scale of relationship, and which appear in man as muscular variations, are reversional.
Children are often born with pigmented hairy patches on their bodies known as moles; sometimes these hairy moles are only of the size of a split pea, in other cases they are several square inches in area, while in rare cases almost all of the trunk may be thus covered. Although many similar pathological cases are often but marked variations called sports, yet the illustrations mentioned are undoubtedly reversional. Of the multitudinous illustrations of atavism that could be mentioned I wish to refer to but one more case.
The conjunctiva is a modification of skin, and frequently proclaims its ancestry by reverting to its original form. It is by no means a very rare event to see a patient having a patch of hair-covered skin growing upon the ocular conjunctiva. While a[53] clinical assistant at the Royal Ophthalmic Hospital in London, we saw one such case, and Dr. Treacher Collins, the pathologist of that eye hospital, has stated that about twelve cases are seen there annually of this pathological condition, which is atavistic, according to Sutton, although it seemingly violates Gegenbaur’s rule about phylogenetic remoteness, and may be looked upon by some as a pathological illustration of a sport.
In speaking of inheritance, we should carefully discriminate between heredity and pseudo-heredity. Physicians constantly write of tuberculosis, lepra, smallpox, and syphilis as hereditary; but it is incorrect and misleading to do so. When a person has syphilis, say, from the earliest existence—that is, from the fertilized ovum by transmission of a syphilitic microbe through the germ cells of the parents—this should be designated by its proper name as congenital bacterial infection. This is totally different from the hereditary qualities that flow from the structural equilibrium following the commingling and struggle for existence of multitudes of hereditary units.
The one set of hereditary qualities is purely germinal, while the other is germinal profoundly modified by the presence of an infecting microbe. Of course, to the extent that any toxines that are secreted by the bacteria may cause permanent structural changes in the germ-cells, to that extent may the germinal characteristics be transmitted and become hereditary.
Many instances of infection of the child in utero have been reported in cases of endocarditis, scarlet fever, and smallpox; and there can no longer be[54] any doubt, from experimental investigation and recent observation, that pneumococci, typhoid bacilli, anthrax bacilli, and pus cocci are able to pass to the fœtus through the placenta. But the diseases that develop in this way can be called hereditary with even less semblance of correctness than in the case of the fertilized ovum that is invaded with a microbe.
All of these cases are illustrations of pseudo-hereditary transmission, and should, for the sake of clearness and accuracy, be spoken of as prenatal infections.
So far as the problems of heredity and variation are concerned, we may say that the life cycle begins and ends with the germ cell. Insects lay their eggs in old age; among plants the annuals flower but to die; in higher creatures the cessation of the procreative power often marks the beginning of bodily decline.
Bearing in mind that the human body consists of two great classes of cells, germ cells and somatic cells, the following scheme will be found very useful in discussing heredity with variation—viz.:
| Germ-Cell | { | Stable (Heredity) | ||||||
| { | Unstable (Variation) | { | Blastogenetic | { | Temperature. | } | Environment | |
| { | Chemical substances in solution in the fluids that bathe the germ-cells, as food, drugs, poisons, etc. | } | ||||||
| { | Somatogenetic | { | Habitat | } | ||||
| { | Temperature | } | ||||||
| { | Climate | } | ||||||
| { | Air | } | ||||||
| { | Food | } | ||||||
| { | Soil | } | ||||||
| { | Water | } | ||||||
| { | “Use” | } | ||||||
| { | “Disuse” | } | ||||||
Just in proportion as fertilized germ cells during the mitoses of ontogeny give origin, among the somatic cells, to other germ cells that are structurally, and therefore physiologically, like themselves, just to that extent do we have heredity; on the other hand, just to the degree that the new germ cells which are produced are unstable, to that degree also do we meet with variations.
In zoölogy the environment of an organism means the sum-total of the conditions of life that surround and affect it, such as food, air, water, climate, etc.
We have already stated that as far as evolution is concerned the structures of fundamental importance in an organism are the Germ-Cells; therefore, for our purposes, we will define environment to be the sum-total of the conditions that directly or indirectly influence in any way the germ-cells, by which variations in them may be produced, or through which stability may be maintained.
There are two great classes of environmental factors that bring about variations in the germ-cells. One of these classes acts directly on the germ-cells, and is therefore called blastogenetic; the other acts indirectly through the body cells, and is therefore designated somatogenetic. Many of the blastogenetic factors bringing about structural changes in the delicate mechanism of germ-cells are entirely unknown, and are therefore designated as fortuitous. Many other causes, such as poisons in solution in the fluids that bathe them, can readily enough be appreciated.
Blastogenetic Factors. It has been demonstrated that various chemical substances, such as chloroform, morphia, chloral, etc., have a pronounced influence upon the vital activities of cells. It is well known that microscopic unicellular plants constitute the essential part of yeast. These little cells have the power of causing fermentation in solutions of grape sugar by which alcohol and carbon dioxide are formed, the latter being a gas and escaping as bubbles. If chloroform or ether be added to the solution of sugar, before adding the yeast, no fermentation takes place, for the yeast-cells are paralyzed. But when the yeast is separated from the chloroform solution and rinsed with distilled water, it soon regains the power of causing fermentation in pure solutions of sugar.
Ova and Spermatozoids are subject to the action of drugs in a similar manner. If actively motile spermatozoids of a sea-urchin be placed in a one-half of one per cent. solution of chloral in sea water, it will be found that after five minutes their action will be completely arrested. These motions can soon be restored if the chloral solution be sufficiently diluted with pure sea water. These temporarily paralyzed spermatozoids, when completely recovered, will unite as quickly with ova as fresh spermatozoids. When spermatozoids are kept for half an hour in the chloral solution, a more decided paralysis will be observed, which persists for some time after the removal of the poisonous agent. A few minutes elapse before some of the spermatozoids exhibit feeble movements which finally become active. Even when placed near ova, it is some time before they fertilize them, although several may[57] attach themselves to the egg’s surface. But, finally, fertilization does take place by the penetration of one spermatozoid, and the egg normally develops.
In like manner, if ova are subjected to chloral solution of varying strength, they also are influenced in a marked degree; for, when fertilized, they develop in an abnormal manner. Ordinarily, normal ova are fertilized by one spermatozoid. If fertilized by two or more, they become diseased, and develop pathologically. The chloral solution favors this fertilization by several spermatozoids. The stronger the solution of chloral, the larger the number of spermatozoids that fertilize the ovum. Experiment and observation show that the behavior of the nuclear hereditary mass is modified, during mitosis, by the chloral and other solutions. It is thus seen that the germ-cells of the lower animals can be profoundly modified by various substances.
Equally true is it that the man or woman who makes use of such drugs as alcohol, opium, chloral, and such like, in an intemperate manner, contains these poisons in solution in the blood, circulating to every part of the body, and thus bathing and profoundly influencing the germ-cells. In consequence of this fact an acquired and habitual intemperance will seldom fail to leave its impress upon one or more of the offspring, either like the original vice or one very closely allied to it. Intemperate people not only profoundly impair the health, the intelligence, and the morals of their offspring, by poisoning these delicate germ-cells, but they also transmit the fatal tendency to crave for the very substances that have acted as poisons on these germ-cells before and after fertilization. And one of the saddest[58] features of this great medical truth is that the hereditary units which are concerned in transmitting these grave abnormal tendencies may lie dormant in the germs of one generation, to become active in those of the next; so that children of intemperate parents may lead honorable and temperate lives, and take every pains to rear, in turn, their own children in a wholesome and refining atmosphere, and yet these children of good environment may become intemperate through heredity, so that the sins of the grandparents may be visited, not on the children, but on the grandchildren.
These profound truths should lead all, and especially law-makers, to remember that “the man who inherits from his parents an impulsive or easily tempted nature and an inert will and judgment, and commits a crime under the influence of strong emotion, can no more be placed in the same category of responsibility with a man of more favorable constitution and temperament than can a man who steals a loaf under the pangs of starvation with a merchant who commits a forgery to afford him the means of prolonging a guilty career.”[3]
Not only do certain known poisons circulating in the blood, or other fluid that may bathe the germ-cells of living creatures, profoundly affect the germ-cells, but many other substances probably have great influence upon them. Certainly, the amount and character of the food have a very decided influence on them, as will be understood from the following facts.
According to Yung, who has experimented very extensively upon tadpoles, all tadpoles pass through[59] a bisexual (hermaphroditic) stage, as is the case probably with most animals. During this tadpole phase external influences, and, more particularly, food, determine their fate as regards sex. In Yung’s experiments it was found that when tadpoles were left to themselves, the percentage of females was in the majority, the average being probably about 57 per cent. females and 43 per cent. males. In experimenting with three broods, those fed on beef gave 78 per cent. females; those fed on fish gave 80 per cent. females; and those fed on the highly nutritious flesh of frogs gave 92 per cent. females.
In Mrs. Treat’s interesting experiments on moths and butterflies, it was observed that if caterpillars were confined and starved before they entered the chrysalis state, the resultant moths or butterflies were males, but others of the same brood that had been highly nourished came out females.
The study of bees illustrates the same conclusions. It is well known that in a beehive there are three kinds of inmates, as the queen, the drones, and the workers,—the last-mentioned being females whose reproductive organs are imperfectly developed. It is believed that the eggs that give rise to queens and workers are fertilized and developed normally. But it is a very curious fact that the eggs which develop into drones do so without fertilization (parthenogenesis). What factor or factors decide the destiny of the two former, determining whether a given ovum will develop into a queen, and thus be the possible mother of a new generation, or stay at the lower grade of a working, non-fertile female? These factors are the quality and quantity of the[60] food. An abundance of what is called royal food causes the development of the larva in such a way that the queen with her reproductive organs is formed. If a larva on the road to develop into a worker (non-fertile female) “receive by chance some crumbs from the royal superfluity,” it is found that the reproductive organs may develop to such an extent that workers partially fertile may be formed. A worker larva may, by this royal food, be intentionally reared into a queen bee.
It is thus seen how profoundly the germ-cells, in their growth, may be affected and made to vary by such a blastogenetic factor as food.
Somatogenetic Factors. As to somatogenetic factors—granting that structural changes in the body (body-cells) of an animal or plant can profoundly influence in some way the germ-cells, and that, therefore, acquired characters can be transmitted—they are many and well defined. Some of them are the habitat of an animal or plant, the temperature, climate, air, food, soil, water, structures in use or disuse (so-called “Use” and “Disuse”), etc.
The following brief descriptions will enable the reader to understand that change in the surroundings (environment) of a living creature may cause its body (body-cells) to vary.
A certain species of snail was introduced into Lexington, Virginia, a few years ago from Europe. In its new habitat it varied very much. One hundred and twenty-five varieties have been discovered there, sixty-seven of which are new and unknown in Europe, the native home of the species.
The common ringed snake, when living in its natural habitat, deposits eggs in the sand, which are[61] hatched by the heat of the sun; but when this snake is confined in a cage in which no sand is strewn, it gives birth to little living snakes.
In experimenting on moths it has been found that the variations of temperature to which the pupæ, and probably also the larvæ, are subjected, tend to bring about very pronounced differences in the moths. Cold has a tendency to develop a darker hue in the perfect insect.
English dogs when taken to hot climates, like that of India, are known to degenerate in a few generations. It is well known how climate affects the hairiness of animals. When greyhounds are taken to the uplands of Mexico they are unable to course on account of the rarity of the air.
In 1870 a number of pupæ of a certain species of moth (Saturnia) were taken from Texas to Switzerland. After passing the winter there, the pupæ emerged from their cocoons as moths, and resembled the Texan species entirely. The young of these moths were fed on the leaves of a plant different from that the moths in Texas feed on, and they developed into moths so different in form and color-pattern from their parents that entomologists classified them as a distinct species.
We have seen how certain foods affect the germ-cells and act as blastogenetic factors; the preceding case and the following show how certain foods act as somatogenetic factors and modify the body-cells. If the bullfinch be fed on hemp seed, its color is changed to black; if the canary be fed on cayenne, its plumage becomes darker; if the common green Amazonian parrot be fed on the fat of siluroid fishes, it assumes a beautiful variegation of red and yellow.
The character of the soil has a marked influence in inducing somatogenetic variations. In France an experimenter collected seed from the wild radish and sowed one lot in heavy soil in the country, while another lot was sown by him in the dry, light soil near the Museum of Natural History in Paris. The radish “roots” grown in these two places presented marked differences in color and form. Those grown in Paris were either of a rose or white color and elongated; while those from the country were violet, dark-brown or nearly black in color, and more rounded than the former.
In the summer of 1847 Professor Buckman gathered seed from wild parsnips, and sowed them in the spring of 1848 under changed conditions of life. Most of the plants grown from these seeds were like the wild parsnips, but some of them developed the light-green color and hairless, smooth appearance characteristic of the cultivated plant. The roots also were found to be more fleshy than those of the wild variety.
Peas and squashes, when grown in different soils, often show remarkable variations.
There is one species of shrimp that inhabits brackish water, and another that lives in water which is much more salt. These crustaceans differ from one another in the character of the spines they bear and in the form of the tail-lobes. They have been regarded as distinct species, and yet either of them can be transformed into the other in the course of a few generations, by gradually altering the saline conditions of the water.
For a long while the siredon and amblystoma were regarded as being distinct genera of amphibians.[63] Siredon was looked upon as a permanent gill-breather, while amblystoma passes through a metamorphosis and becomes a permanent lung-breather. It is now known that the former can change into the latter. If there is plenty of water the siredon remains indefinitely a gill-breather and reproduces freely; but when the water dries up it changes into the lung-breathing amblystoma. These two cases illustrate very well the power of environment to modify the development of organic forms.
As to “use” and “disuse”: It can readily be observed that exercise increases the size of muscles; that by steady application the capacity for thinking can be developed; that the oarsman’s constant use of his hands leads to the hardening and thickening of the cuticle; that the arm of the blacksmith and the legs of the mountaineer are much enlarged, etc.
When an organ is exercised properly, there is an increased blood supply to it, and, consequently, stimulated nutrition and growth in various parts, such as in the muscular, nervous or other tissues.
When an organ is disused there is diminished blood supply, and, consequently, diminished growth and functional capacity. In man it is known that certain activities, such as coal-heaving, shoemaking, etc., produce recognizable effects upon the muscular system, the skeleton, and other parts of the body.
The peculiar habits of a tribe, such as tree-climbing among those natives of the interior of New Guinea, who build their houses in the upper limbs of lofty trees, modify the body in ways that are readily recognizable.
After considering many facts in connection with the brains of rabbits, Darwin announced that this[64] most complicated and important organ in an animal is subject to the law of decrease in size from disuse. We have very interesting illustrations of the effects of “use” and “disuse” in causing somatogenetic variations, in the differences between domestic ducks and the wild ones from which they have been undoubtedly derived. The wild duck, which must constantly be on the alert for enemies, and uses its wings so much more extensively and its legs comparatively less than the domestic duck, is a much more intelligent fowl than the stupid, well-protected domestic one. The wings of the wild duck are stronger and its legs shorter than those of the barnyard duck. It has been shown that in the wild duck the brain is nearly twice as heavy in proportion to the body as it is in the comparatively imbecile domestic duck.
Many other useful illustrations of disuse, such as the cattle and goats in India, that have dependent ears; also cats in China, and horses in parts of Russia, whose ears are dependent, could be referred to. Use and disuse are included among the factors of environment, because by those terms we mean certain groups of body-cells that are functionally active or inactive; for body-cells on any theory of modified pangenesis constitute an exceedingly important environment of the germ-cells.
The surrounding conditions (environment) of an animal or plant having the power to cause variations in the living creature by affecting its germ-cells or its body-cells, the environment may be spoken of as blastogenetic and somatogenetic.
Whether it is a fact or not that somatic variations can induce corresponding variations in the germ-cells,[65] and thus be transmitted by heredity, it is certainly true that all heritages must come through the germ-cells. For this reason, it is clearly seen that so far as evolution is concerned the germ-cells are the factors of fundamental importance in organisms. Therefore, we may repeat that environment is the sum-total of the conditions of life that affect the germ-cells directly or indirectly.
All heritages, then, are derived directly through the germ cells. Can there be any heritages indirectly from the somatic cells through the germ-cells, as has hitherto been assumed? In other words, can acquired characteristics be transmitted to the offspring? This question has given origin to the battle royal that is still going on between opposing schools of biology. The contending parties have appealed to such biological evidence as is furnished by a study of use-inheritance, reflex and instinctive actions in animals, etc., and to such experimental evidence as the induction of traumatic epilepsy in guinea pigs, a change in the shape of the ear by cutting the cervical sympathetic nerve, protrusion of the eyeball through injury to the restiform body of the brain, and such like, noting the effects on the offspring, and have drawn very different conclusions.
As to the transmission or non-transmission of acquired characters, some have maintained that only germinal variations are transmitted (because they believe the germ cells are insulated from the body cells, and therefore from somatic influences). For[66] instance, Ziegler, in his work on General Pathology, says: “If a disease, such as nearsightedness, is the product of a special inherited predisposition, plus the effect of harmful influences which have acted upon the body during life, only that part can be transmitted which was received by inheritance, but not that part which was derived from external influences.” In other words, there is no transmission of acquired character. In this belief it will be observed that he follows Weismann.
On the contrary, other investigators, like Darwin and Spencer, teach that somatic variations—the plus element in Ziegler’s illustration of nearsightedness—do influence the germ-cells (through some such agency as Darwin’s theory of pangenesis suggests), and that, therefore, acquired characters can be transmitted. The question is one of fundamental importance, and yet no crucial experiment has been devised or fact observed which can compel the correct answer. The evidence seems to favor the view that acquired characters can be transmitted.
The theories as to the transmission or non-transmission of acquired characters may be better understood by reference to schemes No. 1, 2 and 3. Scheme No. 1 represents the theory of Pangenesis, which teaches that reproductive cells are not formed from pre-existing reproductive cells, but by the body cells themselves. Darwin taught that all the cells of the body, such as skeletal-cells, muscle-cells, nerve-cells, and so on, are continually giving off infinitely small cell germs or gemmules, which have the power of growing and forming cells exactly like themselves. These gemmules have a great affinity for one another, and, circulating in the blood in countless numbers,[67] they finally come together in the reproductive glands and form the reproductive cells. On this theory the fact of the transmission of acquired characters can readily be appreciated, and it can easily be understood how the parent molds the child. Suppose, for instance, that the parent, by exercise, has become a skillful athlete. In him certain muscles have become greatly developed and strengthened. During all the time of the exercise of these muscles, the modifying muscle cells have been continually giving off to the blood modified gemmules, which collect in the reproductive cells and make it possible for the offspring to develop into an athlete because the modified gemmules develop into modified muscles like those of the athlete.
Scheme 1 shows the absence of any arrow like those shown in schemes 2 and 3, directly connecting germ-cell with germ-cell; this means that in this theory there is no continuity of the germ-cells. But arrows are seen extending from the various body-cells (skeletal, glandular, etc.) to the germ-cell; this means that the germ-cells are formed by influences or gemmules emanating from the various body cells.
Scheme 2 teaches that a germ-cell (when fertilized, of course) can produce many cells, some of which differentiate, finally, into skeletal cells, some into glandular, some into muscle and nerve cells, and some into new germ-cells; so that an animal or plant, I, is formed. In like manner a germ-cell of animal, I, can give rise to the germ and body cells of animal, II, and so on indefinitely. This scheme shows that there is a direct continuity of the germ-cells; and it also shows that the germ-cells are[68] entirely insulated, as it were, from the body-cells (skeletal, glandular, etc.), inasmuch as no influences (arrows) extend from the body-cells to the germ-cells. This means that the transmission of acquired characters, bodily, mental, moral, etc., is impossible. It means, in other words, that none of the advantages gained by a parent in the course of his life can be handed on to his offspring by heredity. There are many biologists and pathologists who teach this theory as the correct one.
The majority of biologists accept the theory illustrated in Scheme 3. This is the theory of modified Pangenesis, which teaches that there is a direct continuity of the germ-cells and that these germ-cells are not insulated from the body-cells, but that the latter, when modified as the result of experience, can send off influences that correspondingly modify the germ-cells; so that the latter, when developing into a new individual, may cause the same body variations that exist in the parent. In short, this scheme illustrates not only that there is a germinal inheritance, but also an inheritance of acquired characters. In this Scheme 3, the oblique arrows show that germ-cells produce other germ cells; the perpendicular arrows show that the germ-cells are modified by influences that proceed from the body cells.
Germinal characteristics are transmitted with vastly greater amplitude and swiftness than merely body (acquired) characteristics. If, for instance, a man were born with that physical constitution that makes with ease a first-class pianist out of him, his sons may easily, through heredity, be first-class pianists. But if a man be born without such a congenital tendency and has by constant labor and[69] practice so developed the muscles of his forearm, his nerves, his brain, etc., that he becomes a very good pianist (acquired characters); and, further, if his male descendants for thousands of generations, in succession, have become very good pianists by constant practice, we may expect that the sons of these last generations may obtain a congenital tendency to become first-class pianists quite easily. The constant improvement, by practice, of groups of body-cells (muscle-cells, nerve-cells, etc.) for generations, has, in each generation, tended to so correspondingly modify the germ-cells that they have acquired the power to develop into men who may become very good pianists with very little practice. This illustrates that there may be a continuous summation of feeble germ-cell variations that have been induced by prolonged influences emanating from somatic variations, so that, in the course of many generations, robust acquired characters may ultimately be translated into strong congenital characters (Scheme 3).
Scheme 1. Illustrating the theory of Pangenesis. Here the germ-cell (a) develops into the body-cells, e, e, e, e, of animal I, as indicated by the oblique arrows, but not into any germ-cells, as indicated by absence of arrow between germ-cell (a) and germ-cells (b). The germ-cells (b) in animal I are formed by the aggregation of infinite numbers of gemmules from the various groups of body-cells, e, e, e, e, as indicated by the perpendicular arrows. The germ-cell (a) transmits germinal heritages to the body-cells e, e, e, e; these body-cells transmit the heritages to the germ-cells (b) by means of the gemmules. If the body-cells are modified in any[70] way, correspondingly modified gemmules are sent to the germ-cells (b), and these germ-cells are modified and thus transmit acquired characters to animal II, and so on.
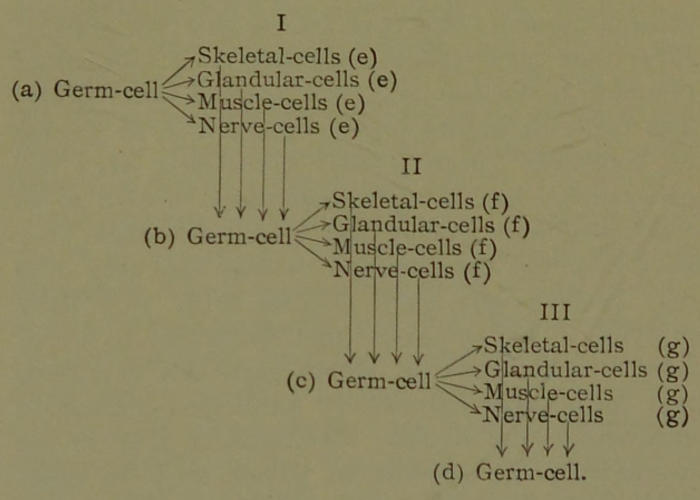
Scheme 2. Illustrating the theory of Continuity of the Germ-Cells; pure germinal inheritance; and the non-transmissibility of acquired characters. The germ-cells are insulated from the body cells. The germ-cell (a) develops into the body-cells, e, e, e, e, and the germ-cells (b), in animal I. The body-cells, e, e, e, e, do not influence in any way the germ-cells (b), as indicated by the absence of perpendicular arrows. The germ-cells (b) get all their heritages from the antecedent germ-cell (a), as indicated by the oblique arrow from (a) to (b). All heritages are purely through the germ-cells. The same with the animals II and III. Germ-cells (a), (b), (c), (d), are connected together by obliquely[71] placed arrows, indicating the continuity of the germ-cells.

Scheme 3. Illustrating the theory of Continuity with Modified Pangenesis. A germ-cell (a) develops into the body-cells, e, e, e, e, and the germ-cells[72] (b) of animal I. The germ-cells (b) get their heritages directly from the germ-cell (a), as indicated by the long, obliquely-situated arrow (continuity of the germ-cells). The germ-cells (b) are, moreover, modified by influences extending from the body-cells, e, e, e, e, as indicated by the perpendicular arrows. A modified germ-cell (b) can develop into a modified animal II, and the body-cells of this animal can influence and modify the germ-cells (c); and so on, indefinitely. The perpendicular arrows indicate that acquired characters are transmitted, and that, too, through the germ-cells.
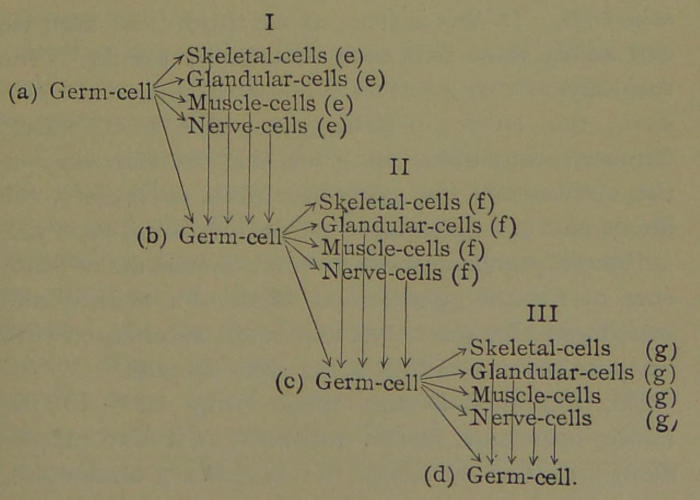
Professor Morgan, of England, has advanced the ingenious theory, which may reconcile the above-mentioned antagonistic views, that somatic variations, in the direction of adaptation, pave the way for germinal variations, so that, while somatic modifications as such are not inherited, they are yet the favoring conditions under which germinal variations are preserved by the great principle of natural selection. If this is true, as we think it is, then we can safely state that each man in his totality is the resultant of two great factors—heredity and environment, the latter including not only food, water, climate, occupation, etc., but also the character of the civilization, the state of morals in society, the ideals and examples most frequently seen, etc., etc.
Heredity brings down to him the streams of tendency of former generations, often of a healthy and beneficent character, but also often surcharged with lust and passion, and reeking with disease.
Environment is the coöperating and, to us, vitally important factor, inasmuch as it may supplement and thus reënforce the hereditary tendencies,[73] whether good or bad; or it may even tend to turn them into new channels, correcting the evil or vitiating the good.
Man is not simply a creature of the present, but profoundly a product of the past. Bodily structure, moral and intellectual tendencies, disease, vices, and virtues are all in the marvelous stream of heritage that comes to him from the past. “Diseases that no facts in the individual life can account for point gaunt fingers of blame from one generation to another. Not a murderer is hung, not a daughter starts on the downward way, but a great company, like those who were present at the stoning of Stephen, stand by inaugurating and consenting to the ruin.”[4]
Truly has it been said that the past is at work in the present, its powers reaching down through the ages, to all the race, largely molding every human life, touching and influencing every individual’s thought and will, and, more than any other force, coloring history.
Studies in heredity illustrate most luridly that the continuity of the human race is a terrible but remorseless reality.
If the ignorance and the perverted pleasures of one generation may produce the vices and the crimes and the diseases of another, a question of tremendous import arises: Is heredity as potent in the direction of virtue and health as of vice and disease? At the first look one is almost tempted to answer Nay! for the most striking examples of heredity seem to be in the direction of evil. But this is perfectly natural. Decay is always more[74] rapid than growth. A cherry rots much more quickly than it ripens. Vice and disease spread much more quickly and widely than virtue and health. But all history and all social and medical science teach that vice and disease carry within themselves the seeds of decay, and virtue and health the seeds of endurance and growth.
Through the great Darwinian principle of natural selection, or survival of the fittest, vice and disease will become less and less predominant, and virtue and hygienic constitutions more and more disseminated.
As influencing a man’s life and character,[5] which is the stronger factor, heredity or environment? Fatalism or choice? In our opinion, as the result of long study and reading, where we have an average man of “mens sana in corpore sano,” environment will be the stronger factor whether for good or for evil—that is, in men in general, who have no organic defect, such as insanity or idiocy, and allied affections, the stronger force is environment; but in those having such defect, heredity is the controlling power, and, we may add, the destroying power.
It must be recalled, though, that the average man with a “sound mind in a sound body,” in his development to his present estate, has become possessed of a vast aggregate of diverse heritages, of varying age, strength and dignity. Some of them are so old and strong that they seem to be cast in unyielding molds, while others are so weak and recent that they fluctuate with every passing circumstance. The most dignified and important of all his heritages is that of rational volition. It is[75] the play of this volition upon many of his other heritages that gives him the power of selecting, to a limited extent, his environment.
Every man is born into the world with a certain physical constitution, and, therefore, with a given temperament; with certain passions; with the power of judgment; and with a certain strength of will. If the power of his will be not equal to the strength of his passions, the latter will surely predominate and will display him as the slave of heredity. If he has such an organization of his nervous system that his volition is superior to his passions, he will be none the less the servant of heredity, though a being now possessed of the power of Free-Will.
Man is, to a far greater degree than is ordinarily realized, the servant of heredity. It seems to us an incontrovertible fact that every living creature, at any given moment, is swayed infinitely more by the totality of its heritages than by its environment. No one can possibly deny this so far as plants and most animals are concerned. Nor, if one look below the surface, can it be denied of the higher animals and of man. Happily, the average man, with his present constitution, has his diverse heritages so proportioned that we may repeat that his life and character (in customs, morals, and religion) are vastly more influenced by environment than by heredity.
The standards for estimating the life and character of men, namely, human customs, morals, and religions, are such recent acquisitions, geologically speaking, that they have, as yet, very slightly if at all influenced the germ-cells. They are acquired[76] (somatic) characteristics, and not congenital (germinal) qualities. They are preëminently the creations of environment. If the infants of a Catholic family which is descended from a long line of Catholic ancestors were to be placed and retained in a purely Mohammedan environment, heredity would carry no Christian customs, morals or religion into that environment, but, on the contrary, the Mohammedan surroundings would instill new customs, different ethical ideas, and a different religion. This illustrates how very feebly indeed are germ-cells correspondingly impressed by pure acquired characters. It is almost certain that the translation of somatic changes into germinal changes is appallingly slow. As far, then, as social customs, morals, and religion are concerned, the average man is, in our opinion, infinitely more the creature of nurture than of nature. But, as far as his temperament, his emotional nature, his judgment, his strength of will, in short, his physical and therefore his mental constitution, are concerned, he is almost absolutely the creature of heredity. The equilibrium of qualities or heritages in the average man, resident in a given, stable community, is in harmony with the average customs, ethical ideas, and religious beliefs of that community. But in all stable communities there are men whose resultant of heritages, some in one direction and some in another, places them out of harmony with the average of their social environment, and they are looked upon, some as idiots, some as geniuses, some as criminals, and others as saints, and so on. So that again we may say that a man’s character in a community is the resultant of an hereditary physical constitution, and his environment.[77] Some men may inherit such a physical constitution that in spite of the best environment they are much debased below the average man; others may possess such heritages that, notwithstanding adverse circumstances, they reach a level of character much above the average man. And there are all gradations between the two extremes.
Where living creatures are in harmony with their surroundings,—where, in other words, they are adapted to their environment,—and where, further, this environment is apparently in a state of equilibrium; there we find the fewest and least marked variations in the living creatures. To the casual observer the face of nature maintains the same guise from year to year. The earth seems solid and unyielding; the mountains appear to be everlasting; the restless waters of rivers and brooks seemingly move and throb in the same channel; the tides ebb and flow in apparently unchanging ocean beds; the birds and flowers and woodlands look alike from year to year; and all the varied phenomena of nature appear completed and permanent, as if the present world were constructed in an unyielding mold.
But nothing is fixed and rigid in nature. The earth itself travels rapidly through space and brings in due season spring, summer, autumn and winter; revolves upon its axis and alternates the starry night with sunshine; and periodically changes its orbit so that at one time the northern pole has a temperate climate where water lilies may grow, and at another period presents an arctic climate with impassable barriers of ice. Ice and frost and other forces are breaking up the rocks of mountains, making[82] larger and smaller fragments and even powder; the rains descend and the mountain brooks are swollen to resistless torrents which carry the fragments and mud to the rivers, and these latter take the mud on to the ocean.
Thus, by degrees, the mountains, hills and all the earth are being eroded and the great bulk of the detritus carried by the rivers to the sea and deposited along the sea margins. Thus sedimentation goes along with erosion, and gradually marginal sea bottoms of immense thickness are formed, which will in time be consolidated into rocks and uplifted as dry land. The ceaseless grinding of waves and tides erodes the coast line and adds débris to the marginal sea bottom. The finest sediment is carried out by the tides so far as to reach the ocean currents, and thus is strewn broadcast over portions of deep-sea bottom, and will also in due time be consolidated into rock.
Myriads of animals that form calcareous shells live and die in the ocean. The shells of the dead animals are falling like a perpetual shower on certain ocean bottoms, year after year, so that immense accumulations of calcareous substances occur there, which will also in time be consolidated into limestone rocks, and uplifted as dry land.
Deltas are forming at present at the mouths of certain rivers, and estuaries at others. Lands are now gradually emerging from the sea in some places and at others sinking into the sea. The entire coast of Scandinavia, both on the Baltic and Atlantic sides, is rising out of the sea, and has been doing so for a long time. It is rising at the rate of more than two feet in a hundred years. During an[83] immense period of time there has been a gradual elevation of all the southern part of the South American continent. Sometimes a large elevation takes place rapidly. In 1822, and again in 1835, the southwest coast of South America, after severe earthquakes, was elevated several feet along a distance of several hundred miles. It is known that the coast of Greenland, for five hundred miles, is subsiding. From a study of coral barriers and atolls it is believed that an area of the mid-Pacific sea bottom covering over ten million square miles is sinking and has been doing so for a long time.
The foregoing facts tend to illustrate the truth that nothing is permanent in the environment of living creatures at present. All surroundings are perpetually changing, though apparently ever so slowly. The changes that are now going on were also taking place yesterday, last week, last century, last æon, and so on throughout geologic time. Gradual oscillations of the earth’s crust on a grand scale and affecting whole continents, but usually so slowly as to escape popular observation, have been taking place ceaselessly through inconceivable ages. These oscillations have produced all the great inequalities of the earth’s surface, such as ocean basins, continents and mountain chains. The oscillations are probably due to the slow cooling and unequal shrinking of the whole earth which has been progressive through all geologic time.
The state of the contest between the eroding and the uplifting agencies of the world at any time determines the height of mountains and continents, the depths of seas, the distribution of land and water, for that period.
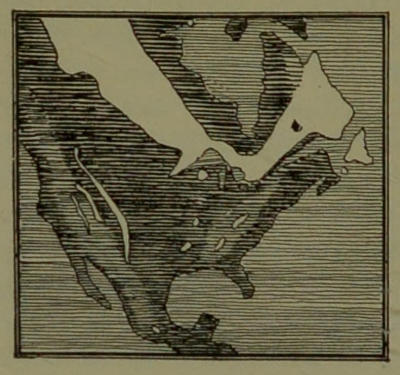
Fig. 11.—Archæan North America. The white part of the drawing indicates the emerged land; the dark shading indicates the submerged land covered by a shallow sea; the light shading indicates the deep sea.
From Shaler’s First Book in Geology. By courtesy of the publishers, D. C. Heath & Co.
Knowing that the present physical agencies at work on the globe have been acting through long ages,[6] it can readily be appreciated how small effects have been accumulated and low elevations, for instance, have become immense, high mountain ranges. The growing mountain ranges alter the climate and the meteorological conditions. The rainfall on one side differs from that on the other. The temperature varies with the altitude, and so on.
Although continents have gradually and steadily grown from the earliest times, there have been many local alterations of land and sea. Marginal sea bottoms have become great mountain ranges. Islands have appeared and sunk from view. Lakes have been gradually converted into solid land or[85] into peat bogs. Fresh water bodies have become brackish. Dry lands have become marshes, and forests have been buried beneath the waves. Geologic changes have caused great alterations in climate at given times and in given areas.
These statements may be illustrated and emphasized by a brief reference to the development of the Continent of North America. This Continent has grown from comparatively a small beginning to its present great proportions. In doing so it has passed through eras of stupendous duration. These eras in the order of their occurrence are as follows:
(1) Archæan era; (2) Palæozoic era (subdivided into Cambrian, Silurian, Devonian and Carboniferous periods); (3) Mesozoic era (subdivided into Triassic, Jurassic and Cretaceous periods); (4) Cenozoic era (subdivided into Tertiary and Quaternary periods); the Tertiary is subdivided into Eocene, Miocene and Pliocene epochs; the Quaternary is subdivided into Glacial, Champlain and Terrace epochs; (5) Psychozoic era, or recent epoch.
The physical geography of the continent at the close of that early geologic era known as the Archæan is shown on the map (Fig. 11). At this time the vast portion of the continent, whose outlines nevertheless existed, was submerged under a shallow sea, as indicated by the dark shading on the figure. The white V-shaped mass, starting just above the site of the great lakes and extending on the one hand in a northeasterly direction to Labrador, and on the other in a northwesterly direction to the Arctic Ocean, is the emerged land of this Archæan time. Smaller masses of Archæan land are also seen at the site of the Blue Ridge[86] Mountains in the east and at that of the Rocky Mountains in the west. Around these lands as a nucleus the North American Continent has been built. Therefore, at the close of the Archæan or beginning of the next, or Palæozoic era, the whole interior portion of the continent was covered by a shallow sea which beat against the Canadian Archæan land on the north, the Blue Ridge Archæan land on the east, and the Rocky Mountain Archæan land on the west. This shallow sea is known as the Palæozoic Sea. Throughout the vast ages of the Palæozoic era, immense sediments were being deposited along the marginal sea bottoms. The deposition of these sediments was simultaneous with a further sinking of the submerged continent, so that the shallowness of the Palæozoic Sea was maintained; finally, the uplifting forces predominated, and the submerged land along the margins of the Canadian Archæan appeared as dry land, and[87] thus increased the area of the infant continent. During all this period there was a steady and slow growth of the land southward from the Canadian Archæan, so that towards its close the visible continent had increased nearly, though not exactly, to the proportions attained in a still later (Cretaceous) period (Fig. 12).
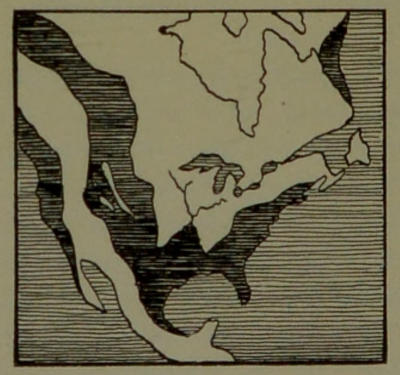
Fig. 12.—Cretaceous North America. The white portion of the figure indicates emerged land—the growing continent.
From Shaler’s First Book in Geology. By courtesy of the publishers, D. C. Heath & Co.
At the close of the Palæozoic era the slow, steady changes that had been going on were replaced by more rapid and comparatively revolutionary changes, which caused great alterations in the physical geography and climate. Hitherto the continent had been comparatively low. Now the vast sedimentary accumulations constituting the marginal sea bottom of the eastern portion of the Palæozoic Sea, which had been accumulating through all Palæozoic time, were uplifted into the great Appalachian chain of mountains.
During the earlier ages (Silurian and Devonian) of this Palæozoic era, the place of the Appalachian chain of mountains was marginal sea bottom; but during the later ages (Carboniferous) it was, through repeated oscillations, in an uncertain state, being sometimes swamp land, sometimes covered with river sediment, and sometimes covered by the sea. It was during this Carboniferous age that the great coal measures were formed; at this time also the climate was probably very uniform, warm and moist, loaded with carbonic acid gas and deficient in oxygen. This period was undoubtedly a paradise for the great coal-forming plants, but was very unsuitable for the hot-blooded air-breathing animals, such as mammals and birds, none of which existed at that period. Throughout all geological[88] time the excessive amount of moisture in the air has been gradually removed by the growth of continents in size and height; also the superabundant carbon dioxide in the atmosphere has been removed in many ways, especially by the plants in the coal period appropriating the carbon. Many ages later, at the close of the Jurassic period, the Sierra Nevada range of mountains was uplifted. Up to this time the site of these mountains was a marginal sea bottom receiving vast amounts of sediment, and the Pacific coast-line was east of the site of the Sierra range. Naturally vast changes in physical geography and climate occurred in consequence. During these and the following Cretaceous ages that the continent was growing, the great interior Palæozoic sea and what may be called the Gulf of Mexico were more and more restricted, as shown in the map of North America in the Cretaceous period of its growth (Fig. 12). This great inland sea, separating the continent into an eastern and western portion, is now called Cretaceous Sea instead of Palæozoic. This Cretaceous Sea covered the whole plains and plateau region of the continent, and extended from the Gulf of Mexico to the Arctic Ocean. At the end of the Cretaceous period of the continent this sea was obliterated by the gradual upheaval of this region and replaced by great lakes. At the same time the western marginal bottom of the sea was uplifted into the Wahsatch range of mountains; also at this time a line of islands in the Cretaceous Sea was uplifted into the Colorado mountains. All these events were entailing tremendous changes in physical geography and climate. Fig. 13 is a representation of the map of North America[89] in the early Tertiary period, the time succeeding the Cretaceous period. In this period, the continent continuing to uplift, the lakes that occupied the site of the Cretaceous Sea are obliterated; the Coast Range mountains of California and Oregon are uplifted from marginal sea bottom (Fig. 13, dark shading); the Atlantic and Gulf borders are extended (dark shading), so that at the close of the Tertiary period the North American continent had attained its present form, except the southern portion of Florida and its keys. Since then the latter have grown and are still growing.
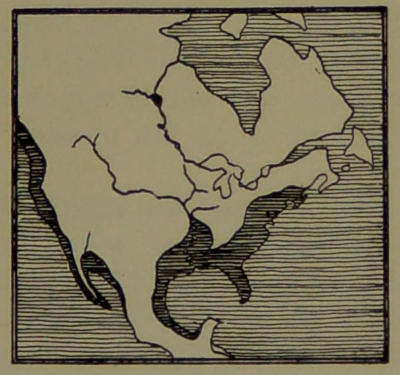
Fig. 13.—Early Tertiary North America.
From Shaler’s First Book in Geology. By courtesy of the publishers, D. C. Heath & Co.
A later epoch still in the history of the globe is known as the Quaternary period, the period that immediately preceded our present epoch. The great features of this period, which is divided into Glacial, Champlain and Terrace epochs, are the wide-spread oscillations of the earth’s crust in high latitudes towards the north and south poles, attended[90] with great changes of climate from temperateness to extreme cold.
The Glacial epoch was characterized by upward crust movements, the land becoming over one thousand feet higher than at present. The land was covered with ice, and an arctic severity of climate extended almost to the Gulf of Mexico. The Champlain epoch was characterized by a downward movement of the coast until it became five hundred feet or more below the present level, so that many lower portions of the continent became covered with sea. At this time there was a moderation of the temperature, a melting of the vast sheets of ice, and consequently a flooding of rivers and lakes, with many icebergs floating in them. The last or Terrace epoch of the Quaternary period was characterized by a crust movement up to the present condition of things.
What is true of the instability of the North American Continent is true of all the continents of the globe. They have all grown from small beginnings to their present huge proportions, and are now undergoing slow but irresistible changes. When these facts are held in mind, one may form a faint conception of the colossal changes that have taken place throughout the sweep of bygone ages. Environment means a complexity of conditions almost infinite in their number and character, and almost infinite in their variations.
It ought now to be understood that not only is the present environment changing, but also that it has been changing from the earliest geologic times. What, then, is to be said about the living creatures that have existed in the changing environment during all these geologic ages? Have they been rigid, unyielding forms? By no means! We know that they can be modified by altering the conditions at present; and a study of the fossils in the rock formations of the different ages of the world shows conclusively that animals and plants have altered in the past with the changing environment. The living creatures in the Silurian ages differ from those in the succeeding Devonian ages, and these latter differ from those in the still later Carboniferous ages; and so on, to the present. Changing physical geography and climate are associated with changing forms in animal and plant life. The growing amplitude and complexity of a continent are associated with increasing complexity and specialization of its living forms. Just as the North American Continent of the Tertiary period differs from that of the Silurian ages, so also do the animal and plant forms of the Tertiary period differ from those of the Silurian ages. Just as there has been a continuity in[94] the growth of the Silurian continent to that of the Tertiary ages, and the present, so, also, there has been a continuity of living creatures from Silurian to Tertiary and present times. Changing conditions of life have compelled modifications in living forms, and those creatures that were unable to adapt themselves to the altering conditions of life have perished, while those that did adapt themselves, through useful variations, lived and progressed in organization.
A study of the fossils in the rock formations (see page 95) of different ages reveals the fact that thousands of species have lived and flourished in one age and then perished, never to appear in succeeding ages. A study of the fossils also reveals that life-forms have passed on from age to age, ever changing with the changing continent, some advancing to higher and higher levels, while others remained lowly.
The facts enumerated above may be instructively illustrated by a hasty reference to the history of organic life as unfolded by the rocks of different ages.[7]
Archæan Era. No evidence of life has been discovered in the Archæan rocks, but, inasmuch as with the dawning of the Palæozoic time, the waters of the sea were peopled with plants and animals living in great numbers and considerable variety, it is evident that the ancestors of these creatures must have lived during the Archæan ages.
| Cenozoic. | Quaternary. | Man, Mastodon, Saber-toothed Tiger and other Vertebrates. Invertebrates. | ||
| Tertiary. | Pliocene. | Equus and other Vertebrates. Pliohippus. Protohippus. Invertebrates. | ||
| Miocene. | Miohippus and other Vertebrates. Mesohippus. Invertebrates. | |||
| Eocene. | Monkeys. Lemurs. Primitive Carnivora and Herbivora. Orohippus. Invertebrates. Eohippus. Fishes, Amphibians, Reptiles. Land Birds. Monotremes. Marsupials. Prim. Monodelphs. | |||
| Mesozoic. | Cretaceous. | Marsupials. Monotremes. Reptiles. Reptilian Birds. Amphibians. Teleosts and other Fishes. Belemnites and other Invertebrates. | ||
| Jurassic. | Reptilian Birds, Reptilian Mammals. Rays, Chimæroids and other Vertebrates. Invertebrates. | |||
| Triassic. | Primitive Marsupials, Primitive Monotremes, Reptiles, Amphibians, Fishes. Ceratites and other Invertebrates. | |||
| Palæozoic. | Carboniferous. | Permian. | Primitive Reptiles, Amphibians, Fishes. Invertebrates. | |
| Primitive Amphibians. Fishes, Invertebrates. | ||||
| Devonian. | Primitive Crossopterygii, Ganoidei, Dipnoi. Goniatites and other Invertebrates. Sharks. | |||
| Silurian. | Upper. | Primitive Sharks. Ostracoderms. Invertebrates. | ||
| Lower. | Blastids, Scorpions, Centipedes, Sea-urchins and other Invertebrates. | Invertebrates. | ||
| Cambrian. | Marine Worms, Molluscs, Trilobites, Brachiopods, Crinoids, Star-fishes, Corals, Graptolites, Cystids, Sponges, Foraminifera. | |||
| Archæan. | Protozoans, though there is no evidence of life.| | |||
Cambrian Period. The Cambrian rocks have furnished many different species of marine animals. Seaweeds are the only plants found fossil in the Cambrian rocks. Some plants and animals may possibly have dwelt upon the land, but if so they have failed to leave any record of their existence. The animals are all Invertebrates, but not of extremely low forms; they have progressed since their appearance in an earlier period. Siliceous Sponges are not uncommon. The Cœlenterates are represented by Graptolites and Corals. Echinoderms are rare, and are principally represented by Cystids, a very primitive type; true Crinoids and Star-fishes appear before the close of the period. That marine worms existed is indicated by borings and tracks in the sands, which have since consolidated into rocks. Mollusca burrowed in the mud or crawled over the ocean bed. Brachiopods existed, and Arthropods were represented by primitive types. The most characteristic of Cambrian fossils are Trilobites and Lampshells.
The former are extinct, and are not represented in the modern ocean; the latter, although greatly reduced in variety and numbers, are still found in various parts of the sea. During all this period, no backboned animals existed; then there were no fishes or amphibians, no reptiles or birds, and no mammals; nothing but invertebrates. The Cambrian fauna shows steady progress, being decidedly more advanced in the upper divisions than in the lower ones.
Lower Silurian (Ordovician) Period. The life characteristics of this period are very similar to those of the Cambrian. Brachiopods have developed[97] extensively, though they have not yet reached their height of development. The Trilobites attained their greatest development in this period; but after the Lower Silurian these creatures commenced to decline in variety and numbers, and finally underwent extinction near the close of the Palæozoic era. In America no plants have been discovered above the grade of seaweeds, but a few of the higher Cryptogams are doubtfully reported in Europe. The character of the flora in a later (Devonian) period makes it highly probable that land plants were well advanced in the Lower Silurian period. Professor Scott states that the remains of land plants may be discovered at any time, though this must remain a matter of chance, inasmuch as all known Ordovician rocks are marine, and therefore not a favorable circumstance for the preservation of land plants. Foraminifera and Radiolaria were abundant in the seas of the Lower Silurian period. Sponges and Brachiopods are numerous and varied. Among Cœlenterates the Graptolites are very numerous and varied; the few and doubtful Cambrian Corals are succeeded by a considerable number of Ordovician genera and species. They were characteristically different from the reef-builders of modern seas. The Echinodermata have greatly increased in importance, being more numerous and varied. The Cystids reach their maximum development in this period. The Crinoids greatly increase in numbers and variety. The Star-fishes greatly expand; and a new and higher order of Echinoderms, the Sea-Urchins, make their first appearance in the later Ordovician period, but under very primitive forms.
Among Arthropods the Trilobites increase greatly in numbers and variety, and attain their maximum development in this period. The occurrence of a Centipede in Ordovician rocks indicates the interesting fact that terrestrial animal life had already begun.
One of the most striking differences between the Lower Silurian and Cambrian periods is the great advance made by the Molluscs, in variety and numbers, during the former period. Molluscs exhibit the most significant change in the great expansion of the Cephalopods, a few of which had appeared in the uppermost Cambrian rocks. In the Lower Silurian period the Cephalopods became one of the predominant types in the marine life. In modern times nearly all Cephalopods are naked (Cuttlefish and Squid), only a few having a shell (Nautilus and Argonaut). The naked forms are higher creatures than those with shells. In the Lower as well as in the Upper Silurian periods no naked forms existed, but only the lower forms with shells (Nautiloids). In modern times and throughout later geological periods only Nautiloids with coiled shells were in existence. In both of the Silurian periods the shells were all straight, and the animals were called Orthoceratites. Such animals were extremely abundant in those times, and often reached an enormous size. Specimens have been found which were ten inches in diameter and over fifteen feet long. They were the most formidable animals of those early ages, and were the rulers and scavengers of the seas; so that the Lower and Upper Silurian periods are known as the Age of Molluscs.
Upper Silurian Period. In this period great progress[99] is made in the history of life on the globe. Upper Silurian life is the continuation and advance of the organic system that flourished in the Ordovician, certain groups expanding, others diminishing; and some new groups now appear for the first time. Sponges are still common. Among Cœlenterates the Graptolites have greatly diminished; Hydroid Corals have become important features of the seas and in the formation of the reefs. True Corals increase largely and play a more important part than in the preceding period. Honeycomb and Chain Corals are quite characteristic of this period.
Among Echinoderms there is a diminution of the Cystids, and a marked increase of the Crinoids. A new class of Echinoderms, the Blastids, now make their first appearance. The Sea-urchins and Star-fishes have increased in variety and abundance. Brachiopods still exist in multitudes, but under changed forms. Among Molluscs the Orthoceratites are still the rulers of the seas. Among Arthropods the Trilobites are still numerous, though decidedly less so than in the preceding period. The land animals are insects of low types, mainly allied to cockroaches. Scorpions are also present as fossils in the Upper Silurian rocks. These animals prove the existence of a contemporaneous land vegetation.
During all the many millions of years that constitute the Silurian periods, hosts of species lived and died; hosts of variations were induced in the living creatures by the ever-changing environment; so that at the close of this time the Silurian rocks show that some of the life-forms, lowly as they were, had yet climbed higher in the scale of organization.
The later Silurian rocks reveal the remains of an[100] entirely new branch of living forms, a higher branch of animals (Vertebrates) than had ever before existed. These remains are those of Ostracoderms and primitive Sharks. But at that early time they held a very subordinate position among the hosts of living creatures; the Molluscan Orthoceratites were still the rulers of the seas; it was still the Age of Molluscs.
Devonian Period. The next set of ages succeeding the Silurian time is known as the Devonian period. Many of the kinds of creatures living in the Silurian ages are also found in these ages, but under changed forms. The Devonian seas had a great abundance and variety of Corals and Crinoids. The Chain Corals have perished, but the Cup and Honeycomb Corals still live on with modified appearance. The Graptolites are almost extinct. The Orthoceratites still live on, much reduced in numbers and size. The Molluscan Cephalopods have been revolutionized. New forms have grown out of the variations of the past, so that now in Devonian times we have the introduction of the great Molluscan Ammonite family, under the forms known as Goniatites, which are characteristic of this and the succeeding Carboniferous ages. The Trilobites still continue under new forms, but much reduced in size and number. Brachiopods are abundant and diversified.
In this age, for the first time, land plants become conspicuous. The Devonian forests consisted of the highest flowerless plants, such as ferns, horsetails, and club mosses; and also the lowest flowering plants, such as cycads, pines, cypresses, etc.—plants which have imperfect and inconspicuous flowers. In all ages, as now, land vegetation has been closely related with insects. Insects, though rare as fossils,[101] are found in connection with the forests in the Devonian period. They are among the lower orders of the class, and are somewhat allied to cockroaches and dragon flies.
The most characteristic feature of the Devonian age is the expansion of that new and higher class of animals, the backboned animals; for during the earliest Devonian or latest Silurian times the Fishes made their appearance. At first many of them were comparatively small in size, they were few in numbers, and of strange, unfishlike forms. Such were the Ostracoderms, which, though generally called fishes, belong to a type much below the true fishes, and more nearly allied to the Lampreys, for they were devoid of jaws and paired fins. There was a great variety and wealth of true fishes in the Devonian period. The Elasmobranchii (Sharks) were well represented, though very generalized ones. Crossopterygii, Dipnoi and Ganoidei were important elements of the fish fauna of this period. At this period there was an entire absence of those highly specialized fishes (Teleosts) which in modern times make up the vast majority of fishes, both marine and fresh-water.
The true fishes soon developed to so great an extent in size and numbers that they swarmed in the Devonian seas and quickly became the rulers of the age; hence Devonian time is known as the Age of Fishes.
These early Devonian Fishes were of a more lowly organization than modern ones, and were what is called generalized forms; that is, they combined in themselves the characters of two distinct classes. They had distinct amphibian characters. From these[102] generalized Fishes were afterwards formed, through many transitions, the Amphibians and the Fishes as branches from a common trunk. This illustrates what is a very general law, viz.: that the first-introduced examples of a class are not typical forms of that class, but intermediate forms or connecting links with other classes. Certain footprints recently discovered in the upper Devonian rocks of Pennsylvania indicate that the Amphibia, the lowest of air-breathing vertebrates, commenced their career in the latter part of the Devonian period; but they were inconspicuous among the monarch fishes.
Carboniferous Age. As in the Devonian Age so in the Carboniferous times, we find flowerless plants, such as ferns, horsetails, and club mosses; but in this coal-forming age they culminate, and have become gigantic in size, especially the two latter. At this time we again meet with the next higher order of plants, as cycads, pines, cypresses, etc.
These plants of the coal period were remarkable, generalized types, connecting classes now widely separated. During this period the insects increase in variety and numbers along with the advancing vegetation. It is interesting to find that as the highest flowering plants are not yet in existence, so those highest orders of insects, the flower-loving and honey-loving ones, such as bees, butterflies and moths, are not yet in existence. Corals and Echinoderms, Molluscs and Crustaceans, etc., continue through the ages with ever shifting forms. Brachiopods have greatly diminished. Foraminifera for the first time assume considerable importance in the earth’s economy.
The Molluscan Goniatites continue through this[103] age with changing forms and advancing organization, and are very numerous. The Trilobites, which are characteristic of Palæozoic times, continue under new forms through this age, and then perish. In this age are introduced for the first time typical Crustaceans of the long-tailed kind, such as shrimps.
Devonian fishes still prevail. The Elasmobranchii are numerous and varied, and some of them are highly specialized. Pleuracanthus is a remarkable shark which has many features in common with the lung-fishes (Dipnoi), such as the character of the pectoral fins, the shape of the tail, the bones which form a roof for the skull, while the skin is naked.
The Dipnoi continue, though in diminished numbers.
The Crossopterygians are much less abundant. The Ganoidei increase in numbers and varieties.
The fact of greatest interest concerning the Carboniferous period is the expansion of the true land-breathing, backboned animals, such as the Amphibians. Their expansion marks a distinct step forward in the scale of life.
The Carboniferous Amphibia all belong to the extinct order of Labyrinthodonts (Stegocephala), in which the skull is well covered with a roof of sculptured bones. The Amphibia of the lower Carboniferous rocks are of small or moderate size, not exceeding eight feet in length and mostly much smaller. A great number of these Amphibians are known, most of them like the Salamanders in shape, but some are snake-like in form, being long and slender. An example of the Stegocephala is Archegosaurus.
The upper Carboniferous rocks represent the later[104] Carboniferous period (Permian Period). The life of this period is transitional between that of the Palæozoic and of the Mesozoic eras. Here we meet with the last of many types which had persisted from Cambrian times, associated with forms which are prophetic of the characteristic types of the Mesozoic era. We also meet with types in the Permian that are peculiar to the period. In this section of the Carboniferous period many of the genera of the fishes are the same, while new ones are introduced. Among the lung-fishes the genus Ceratodus is introduced, a creature very closely allied to the modern lung-fish of Australia. The Amphibia are still represented by the Stegocephala, several of the older genera persisting, while many new forms appear for the first time; several of the latter surpass the earlier Carboniferous genera greatly in size. The transitions from many of the Devonian fishes to the Carboniferous Labyrinthodonts are so gradual that it is sometimes difficult to say whether we are dealing with Fish or Amphibian. In other words, the Devonian fishes are generalized fishes, that is, connecting links between Fish and Amphibian. When the Amphibians finally separated from the fishes they were not the highly specialized forms of more recent times; but they were generalized Amphibians, having some reptilian characteristics. As time passed on and the creatures continued to modify, some varied more and more in the direction of true amphibians, and others more and more in the direction of reptiles, until in the Permian stage early, generalized Reptiles, of lizard-like form, appeared in large numbers. The most interesting of these primitive Reptiles are the Theromorpha[105] (beast forms), which present many remarkable approximations to the structure of the mammals. As yet no snakes or turtles, no alligators or crocodiles had come into existence. In spite of the fact that great advance in animal life was made during Carboniferous times, there was little of the life in those ancient woods that we associate with the forests of the present; they were gloomy wastes of shade, without the presence of bright flowers, no humming of the bees, no song of birds, and few sounds save the gurgling of running streams, the sighing of the wind through the leaves, the splash of waves upon the shore, and the bursting of the thunder clouds. The interest and importance of the Carboniferous Amphibians as the first land backboned animals, is so great as to cause the Carboniferous period to be spoken of as the Age of Amphibians.
Triassic Period. In this period the ferns and horsetails continue under new forms; but the next higher orders of plants, the Cycads and Conifers, now predominate. The Goniatites are replaced by the Molluscan Ceratites, the latter being characteristic of the Triassic period.
One of the most characteristic changes from the Palæozoic to the Mesozoic era consists in the great reduction of the Brachiopods.
The Vertebrata of this period are of extraordinary interest and show great progress. The fishes exhibit the least progress. The Dipnoan Ceratodus is very characteristic, continuing from the Permian. The Crossopterygians have greatly declined. The Ganoidei continue to be the dominant fish-type, and are most like the existing gar-pikes. The Amphibia[106] (Stegocephala) culminate in this period, multiplying and diversifying greatly, and far surpass in size the Carboniferous and Permian genera, and then become extinct.
It is in the Reptilian class that we find the most remarkable changes. The abundance and diversity of the reptiles in this and the two succeeding periods are incomparably greater than those in the Permian age. The Triassic rocks have representatives of almost all the orders of Mesozoic reptiles, though often these are comparatively small and rare forms.
The reptiles with mammalian characters (Theromorpha), which first appeared in the Permian period, culminate in the Triassic, especially in southern Africa, and then become extinct. No birds, or reptiles which can be regarded as the ancestors of birds, have been found in this period. In this period one of the greatest advances in the progress of life is indicated by the first appearance of low mammals,—mammals having very decided reptilian characters, and belonging to the orders of generalized Monotremes and Marsupials (Dromatherium; Microlestes).
Jurassic Period. The Fishes have advanced greatly beyond those of the Triassic rocks. The Sharks have advanced practically to their modern condition, and a new order of Elasmobranchs, the broad and flat Rays, are introduced.
The Chimæroids occur in this period, and were more numerous than in modern times. Dipnoans have become very scarce. The Crossopterygians are greatly reduced. The Ganoidei are still the dominant type; some of these latter approximate the Teleosts so closely that it seems arbitrary to call[107] them Ganoidei. In Europe the Reptiles culminate in this period and show extraordinary development in variety, huge size, number and degree of organization. They were rulers in every department of nature. They were the rulers in the air in place of birds; rulers on the land in place of mammals; and rulers in the sea instead of sharks and whales.
Immense land reptiles (Dinosauria) as large as our largest mammals, and in some cases larger than the elephant, moved sluggishly over the land. Some walked on all fours; others were occasionally or usually bipedal, and walked upright like birds, and had many structural features in common with the latter; some were herbivorous, feeding on plants and even reaching into the branches of trees for their food, others were carnivorous, feeding on their fellow-creatures. Huge reptiles (Ichthyosauria) swam about in the sea in great numbers. Immense bat-like forms (Pterosauria) sailed through the air like birds, being literally flying dragons.
These reptiles, which had branched off from the generalized Amphibians, were themselves very generalized creatures. It is interesting to remember that the Monotreme and Marsupial mammals found in this age were very low reptilian mammals; they were not typical, specialized Monotremes and Marsupials, like the modern creatures, but very generalized forms, being probably connecting links with low generalized Insectivora, which were the first of the true mammals.
These early Monotremes and Marsupials were quite small animals, varying in size from a mole to a rabbit. They were insignificant creatures among the mighty giant reptiles, but they carried in their[108] warm blood the promise of future mammalian supremacy.
Birds appear for the first time in this period, but very different from modern birds. Not only did many of the reptiles of this time have bird-like characters, but all the birds of this period were distinctly reptilian birds (Archæopteryx). Birds came off from primitive reptiles as branches from a stem (see Diagram of Development). It was only with the passing epochs that these early birds changed more and more from their reptilian characters and assumed more and more the features of modern birds. Many fossil connecting links between reptiles and birds have been discovered, and afford most useful illustrations of those changes in creatures that we call Evolution.
Cretaceous Period. In the Cretaceous period the aspect of plant life has changed greatly. Now, for the first time, we meet with ordinary hard-wood trees, such as beech, oaks, hickory, maples, poplar, etc., but of very different genera and species from those existing to-day.
In this period, for the first time, we find the highest order of Cephalopods, viz., the naked ones, allied to cuttlefishes and squids. They are known as the Belemnites. Sometimes the fossil ink-bags of these creatures are found so that they can be drawn in their own fossil ink.
The Ammonites proper, which were introduced in the Triassic period and culminated in Jurassic time, become extinct at the close of the Cretaceous.
The Vertebrates form the most characteristic features of the Cretaceous fauna. A revolution has[109] occurred among the Fishes. Sharks of modern type are numerous. Crossopterygians and Ganoidei are rare. There has been an immense expansion of Teleosts or Bony Fishes, which now become the dominant fishes. Most of the Cretaceous Teleosts belong to modern families and even genera.
The Reptiles, in this period, continue to be the dominant types of the land, air, and sea, and it is difficult to decide whether the Cretaceous or the Jurassic is to be regarded as the culminating period of Reptilian history. The flying Reptiles (Pterosauria) of this period are remarkable for their great size, far exceeding, in this respect, those of the Jurassic time. The land Reptiles (Dinosauria) are in greater numbers than in the preceding period. The sea Reptiles are less numerous than in the Jurassic, but are of greatly increased size.
Cretaceous Birds are much more numerous and advanced than the Jurassic ones.
The Mammals of the Cretaceous are much more abundant and varied than those of Jurassic times, but they are nearly all of very small size, and continue to play a very modest rôle. The lower Cretaceous mammals differ but little from those of the Jurassic, except for the larger number of genera. The mammals of the latest Cretaceous time are much more numerous and diversified than those of the early Cretaceous. They also show affinities with the mammals of the next succeeding or Tertiary period. The birds and mammals, though they have varied with the passing time and changing surroundings like all other living creatures, are very different from those around us now.[110] They have thrown off some of their reptilian characters, yet they are still distinctly and pronouncedly reptilian.
The Triassic, Jurassic and Cretaceous periods together constitute the Mesozoic Era, or Age of Reptiles; for never before and never in succeeding ages were reptiles so huge, and varied, and masters in all the realms of nature.
Tertiary Period. In reaching Tertiary time we enter upon the threshold of our modern world. The reptiles have dwindled to a few low forms, such as alligators and crocodiles, to the small lizards, to turtles and tortoises, and to those low-grade reptiles, the snakes, which now for the first time are the most numerous of the class. In these ages, as now, the Teleost fishes vastly predominate, and the Ganoid and Crossopterygian fishes are nearly extinct. In Tertiary times, as in the present, all the reptilian birds had disappeared, and only typical birds remain. In other words, the bird-class had now separated from the reptilian class, and the connecting links became extinct.
In this age for the first time the highest flowering plants are abundant, and now for the first time also the highest orders of insects are abundant, such as bees, ants, butterflies, etc. On account of the greater warmth and moisture in the Tertiary period, insect and plant life were fuller then than now. That the climate was warmer in those times is shown by the following facts: In America, during the early Tertiary period, figs, evergreens and palms grew in Dakota, showing a temperature there at that time equal to the temperature in Florida at present. In the middle Tertiary period trees like[111] the Redwood of California, and Magnolias, were abundant in Greenland.
Although Reptilian Mammals (Multituberculata) lived in the preceding era, true or typical Mammals first came to view in the earliest Tertiary times. They soon succeeded the vanished giant reptiles as the rulers of the world; so that this and the succeeding (Quaternary) period are known as the Age of Mammals. At this period, for the first time, appear generalized primitive flesh-eating mammals (Creodonta), and primitive grass-eating ones (Condylarthra and Amblypoda); also primitive Primates (primitive Lemuroids, and, later, primitive types of Monkeys, e. g., Anaptomorphus).
The evolution of mammals, compared with that of other animal groups, has been so rapid that each stage of even the Eocene has its own mammalian fauna, differing from those of the succeeding and preceding stages. All through the Tertiary period, as through all the preceding ages, the species that are advancing in life are undergoing greater and greater specialization. Animals at first closely related finally become more and more separated from one another. This is well illustrated by a study of the hoofed animals. In the earliest Tertiary period all these animals seem to unite into one branch but now they consist of many widely separated sub-branches. As we trace the branch down to the earliest Tertiary period, we find that even in early Eocene times it divides into the even-toed and odd-toed ungulates. In the later Miocene times, each of these again separates,—the former, even-toed ones, into the hog and hippopotamus families with four toes, and the ruminant family with two[112] toes; the latter, odd-toed ones, into the elephant family with five toes, the rhinoceros and tapir families with three toes, and the horse family with one toe.
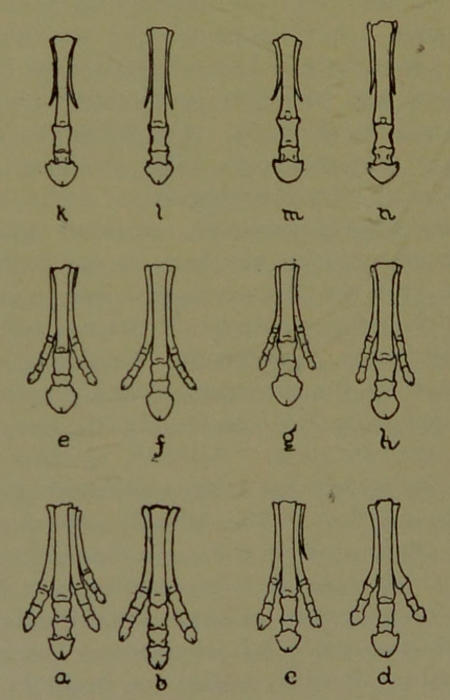
Fig. 14.—Illustrating genesis of horse’s feet. Fore-feet (a) and hind-feet (b) of Orohippus; fore-feet (c) and hind-feet (d) of Mesohippus; fore-feet (e) and hind-feet (f) of Miohippus; fore-feet (g) and hind-feet (h) of Protohippus; fore-feet (k) and hind-feet (l) of Pliohippus; fore-feet (m) and hind-feet (n) of Equus.
It will be exceedingly instructive to trace briefly the progressive specialization of the horse family through the Tertiary period. A wonderful series[113] representing this family has been found in the American Tertiaries. First of all, the Eohippus is found in the earliest Eocene rocks. This little animal was about the size of a fox and had four perfect toes (hoofed) and a fifth, much smaller, imperfect one (splint) on the fore-feet; the hind-feet had three perfect hoofed toes. Later on, in the middle Eocene rocks, was found the Orohippus (Fig. 14), with three toes behind (b), and four in front (a)—the fifth imperfect toe (splint) being lost. This animal was also about the size of a fox. Still later on were found, in the Miocene rocks, the Mesohippus (c, d) and the Miohippus (e, f), with three toes behind (d, f) and three toes and a splint in front (c, e). The splint in the Miohippus is much smaller than in the Mesohippus, and also in the former the two side toes have become smaller and farther removed from the ground. Both of these animals were about the size of sheep. Further on, in the early Pliocene rocks, appears the Protohippus (g, h), with three toes on all the feet, but the middle toe considerably larger and longer than the side toes. This animal was about the size of an ass. Then, finally, came the Pliohippus (k, l) and Equus (m, n) in the latest Pliocene rocks. Here the middle toe is greatly enlarged and the side toes are reduced to useless splints. Thus by degrees, through slight modifications not only of the feet, but also of the skull, teeth, brain, etc., and by adaptations from age to age, was formed the modern horse. In like manner, by gradual changes and adaptation to surroundings, all the early generalized mammals pass into the higher specialized ones.
Quaternary Period. The mammals of this age[114] differ from those of the Tertiary period and from living species. Never before this period, and never since, have mammals been so large. They culminate in this period and then decline. The great mastodon, the great cave-bear, and saber-toothed tiger, the Irish elk, and gigantic sloths and armadillos lived in this age.
Most interesting of all, the remains of man now first appear associated with these extinct animals. Man must have been at first an apparently insignificant creature among the mighty mammals that surrounded him, and must for a time have contended more or less doubtfully with them for mastery. But as he increased in numbers and intelligence he became more and more the ruler of the brute creation; so that this age in which he rules may fittingly be spoken of as the Psychozoic Era, or Age of Man.
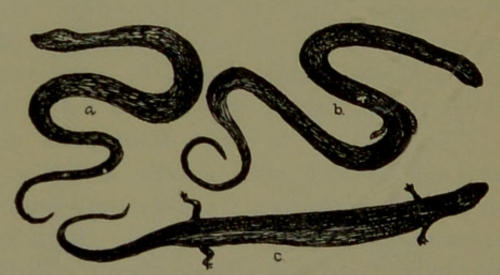
Fig. 15.—A snake and two lizards; a, snake; c, Bipes (lizard); b, Cheirotes (lizard).
From Shaler’s First Book in Geology. By courtesy of the publishers, D. C. Heath & Co.
Fig. 15 is introduced to show how easy may be the transitions from one order of animals to another. Two long, slender Lizards are shown[115] and also a Snake. Bipes has four limbs, but they are very small and weak. Cheirotes has lost, probably through disuse, the hind-limbs and the front ones are small and weak, almost useless. The snake has no limbs at all. The two lizards are quite snake-like in external forms. Just as changing environment, use and disuse, etc., can effect transitions in external forms by slight gradations, so also may the whole internal structures undergo marked alterations by slight modifications through long ages.
The brief references we have made to the changing life-forms during the geologic ages, should be very instructive even to a reader not familiar with zoölogy. They teach the important lesson that life, in the main, has ever advanced to higher and higher levels. At one time in the history of the earth no animals were in existence higher than the Invertebrates; these lived through the ages with ever-changing forms. Later, in the course of geologic history, some of the primitive Invertebrates evolved into the lowest Vertebrates, and Fishes finally appeared, at first very primitive and generalized. For long ages there were no creatures on the globe higher than these. Then as the æons slowly rolled on some of the early fishes evolved finally into a higher group of animals, the Amphibians, and these for long ages were the highest creatures in existence. As time slowly passed, some of the primitive amphibians evolved into primitive Reptiles, and these creatures for long ages were the highest in existence and monarchs of the world. Some of the primitive reptiles branched off in one direction leading to modern Birds, and some advanced in another[116] direction that led through primitive mammals to man and other modern mammals. And now in modern times man is monarch among the animals.
We may now begin to realize that truly with the changing and growing continents during the geologic ages, many life-forms have varied and advanced to higher levels, culminating in the final appearance of man as well as other living creatures.
In briefly outlining the transmutations of living forms that took place during the Geologic Ages, we have said that changing conditions of life—through an ever-shifting environment—have compelled modifications in the form, structure, and habits of living creatures; and that those creatures which were unable to adapt themselves, through useful variations, to the altering conditions of life have perished, while those that did adapt themselves lived and progressed in organization. What, then, is the great agency through which some life forms have been eliminated during the ages, while others have been selected to continue through these ages? This agency is Natural Selection. The phrase “natural selection” is simply a convenient, condensed statement of observable and easily verifiable facts, viz.: that animals and plants are so situated in this world that they can only secure their food and mates by work, by effort, by struggles, whether consciously or unconsciously, and whether directly or indirectly; and that in these struggles those that are best equipped for their life duties are the ones that are most naturally successful in living and procreating their kind. The survival of those best adapted to their environment may be spoken of, in[120] the language of Spencer, as the Survival of the Fittest. Darwin’s phrase—Natural Selection—has precisely the same significance and means that those creatures which are best fitted to their surroundings are the survivors. The working of Natural Selection may be made clearer by a brief reference to the Artificial Selection by Man of various animals and plants. Variations frequently take place in domesticated animals and plants. Some of these variations appeal to man as being of practical value, others as beautiful, and others again as curious or interesting. He selects those individuals whose variations he wishes to preserve, breeding only them together, and in this way accentuates those variations he desires to perpetuate. In course of time the accumulation of these differences becomes so marked as to make the animals differ greatly from the original stock from whence they came. A well-known illustration of this process is the pigeon. All our domestic pigeons, forming a large number of well-marked races, such as the fantail, the tumbler, the pouter, etc., have been produced from the ordinary wild rock pigeon of Europe. The bird fancier, noticing individual differences in the offspring of the wild rock pigeon, selected the peculiar individuals, and bred only them together. By this simple process of artificial selection and isolation of the chosen or selected individuals, all the races of pigeons have been produced. Plate I shows the wild rock pigeon Columba livia and the domesticated pouter. The forms of these pigeons are very different, yet the wild rock pigeon has been transmuted into the pouter through the agency of Artificial Selection. The same is the case[121] with our pigs, dogs, cats, apples, grapes, and other domesticated animals and plants. Fig. 16 shows the domesticated pig that has been derived from the wild boar by Artificial Selection. By this process, such distinct races as the Newfoundland, the Skye Terrier, and the Bulldog have been produced—creatures that have all come from common ancestors, yet so different looking, one from the other, that if they had been found in the wild state, they would not only have been ranked as distinct species, but as even distinct genera. The same method has given us different races of horses, cows, sheep,[122] flowers, grains, etc. The swiftest horses, for instance, are selected to breed together; then the fleetest offspring of these, time after time, until horses are produced whose speed far surpasses that of the originally selected pair from whence they were derived. Darwin has taught us that what man does on a small scale, in a comparatively short time, Nature has been doing on a vast scale for long ages and has thus given rise, from simple forms, to the infinite variety and complexity of animal and plant life, as we behold it on the globe to-day. The selection by man of useful variations in domesticated creatures being appropriately called Man’s Selection, or Artificial Selection; the vastly greater selection by nature of animals and plants with useful variations and on an infinitely grander scale, through inconceivably long ages, is most fittingly called Natural Selection. The struggles—of animals, for instance—that necessarily lead to the survival of the fittest, are intensified and made exceedingly acute and severe by the fact that all animals tend to increase in a geometrical ratio, and by the further fact that the food and place for animals are limited. In other words, the population of the animals in a given area tends greatly to outrun the means of subsistence. And since animals are constantly varying in many directions and are as plastic in the hands of Nature as clay under the chisel of the modeler, those that possess any useful variations, whether congenital or acquired, that give them any advantages in this great battle of life, will most likely come out of the struggle as victors.

Fig. 16.—Wild Boar contrasted with a modern Domesticated Pig. Reproduced from Romanes’ “Darwin, and After Darwin.” By courtesy of The Open Court Publishing Company.
Multiplication of Animals. Even in the slow-breeding[123] elephant, the offspring tend to increase threefold in each generation. Some animals tend to increase twenty or thirty fold in each generation, while still others tend to increase a thousand fold or even ten thousand fold. If all the offspring of the elephant lived, in eight hundred years there would be over nineteen million elephants alive. If the eight million eggs which the roe of certain fishes, such as the cod or the eel, contains, were to develop into adult forms, the ocean would quickly become a solid mass. The aphis or plant louse is so very prolific that it has been estimated that the tenth brood of one female alone would contain more ponderable matter than all the population of China,—estimating this population at five hundred millions. Yet, in spite of this tendency on the part of animals and plants to increase in numbers at such a stupendous rate, it is found that, in any given area, the conditions of which are not changing, the number of the animals and plants remains fairly constant. This is because of the fact that, along with the stupendously large birth rate, there is an equally stupendous death rate. This high death rate is to a large extent indiscriminate, for it involves those that are physically fit to live, as well as those that are unfit to live. At the edge of a coral-reef, free-swimming, active embryos are found in immense numbers. After a while some of these settle at too great a depth in the water or on the muddy bottom, and die; others get into a more suitable position and live. Again, whole nests of bees are destroyed by the badger; tongue loads of ants are engulfed at one gulp by the ant-bear; hundreds of thousands of fry are destroyed by the Greenland[124] whale at one swallow. In all these cases, the destruction is indiscriminate,—the good, bad, and indifferent are alike decimated, but in spite of this wholesale and indiscriminate destruction, keeping down, as it does, the stupendous birth rate of living creatures, more animals and plants are born than are required to keep up the normal number of individuals that can be supported in a given area. Among these creatures there arises a struggle for existence, a struggle for food and place, and those that are the best fitted to live come out of the contest as conquerors. In this struggle those creatures that emerge as victors, on account of having been best adapted to their conditions of life, may be spoken of as having been selected by Nature; or they may be spoken of as chosen by Natural Selection. In this great struggle, over which Natural Selection presides as some inexorable, ever-watchful, sharp-eyed task-master, the victory is to the cunning instead of the stupid; the race is to the swift instead of the slow; and the battle is to the strong. The wolves of keenest scent, the tigers of more supple spring and sharper sight, secure their prey and thrive, while the weaker members fail to get their food and starve. During migration the birds that are strongest on the wing reach the land whither they are flying, while the weaker perish on their course. Thus Natural Selection acts in two ways, eliminating the unfit and selecting the fit, and there are, besides, two special modifications of Natural Selection which are called Sexual Selection and Insect Selection.
The following analysis of Natural Selection may be useful to the reader, viz.:
| Natural Selection | { | Elimination | { | Physical and Climatic Causes. | ||
| { | Enemies. | |||||
| { | Competition. | |||||
| { | Selection | { | Natural Selection Proper. | |||
| { | Sexual Selection. | { | By Preferential Mating. | |||
| { | By Battle. | |||||
| { | Insect Selection. | |||||
Elimination of the Unfit. The elimination of the unfit takes place through the agency of physical and climatic causes and also through enemies, and competition of members of the same species. Elimination through the action of surrounding physical and climatic conditions is shown by the following facts: if certain tropical animals be transferred to sub-Arctic or even temperate regions they are unable to adapt themselves to the requirements of the new climatic conditions, and die sooner or later; many animals are killed if the fresh-water lake in which they live be invaded by the waters of the ocean; fishes which live at great depths in the sea and are, therefore, subject to great pressure, are killed when they are brought to the surface, on account of the expansion of the gases in their tissues; if the water where corals are living becomes too fresh, too muddy, or too cold, they will die. The change of climate to a much colder temperature at the close of the Jurassic Ages was probably the cause of the extinction of the huge reptiles that took place at that time. In the winter of 1854-5, four-fifths of the[126] birds in Darwin’s grounds perished on account of the severity of the cold.
As to elimination by enemies, it is well known to naturalists that throughout nature battle within battle is continually recurring with varying success. When weaker animals are preyed upon by stronger ones, and self-defense is useless, the bulky and slow animals are eliminated, while the swift and agile ones escape; the stupid are destroyed, while the cunning often survive. As to elimination by competition, the stronger animals kill the weaker ones, and then quarrel and fight with one another over the prey, the strongest, etc., getting the food. While weaker animals are being preyed upon by various enemies, and are thus eliminated, these enemies are also competing with one another for the prey. At the same time that the stupid and slow creatures are being destroyed by their captors, thus leaving the more cunning and agile animals in possession of their habitat, the stupider and less active captors are gradually eliminated by competition, through failing to capture their more agile and cunning prey.
The agency of Natural Selection in bringing about the innumerable adaptations of animals and plants, and, therefore, causing transmutations of living creatures, can most interestingly and instructively be illustrated by a study of the coloration of animals and plants.
As a correct idea of the mode of operation of Natural Selection, and the pronounced results attained thereby, is very desirable, it will well repay the reader to study in extenso the relations of color-patterns to environment. The whole subject is[127] nothing but repeated illustrations of one principle, viz.: Natural Selection. This fact and the interest of the subject will justify the numerous details.
The Coloration of Animals and Environment. The colors of animals often harmonize most wonderfully with their surroundings. Thus green is a common color of animals in the evergreen forests of the tropics; white is the prevailing color in the arctic regions; and a yellowish hue in desert places. In the evergreen forests of tropical America, whole groups of birds are found whose fundamental color is green; there the parrots and fruit-eating pigeons are commonly green; the bee-eaters, leaf-thrushes, and many other birds, have so much green in their color as to add greatly to their concealment in the dense green foliage. In the desert places, the lion, the desert antelope and the camel harmonize with the color of the rocks and sand among which they live. In the Arctic regions the polar bear, the Greenland falcon, and the American polar hare are white. We have further the dusky hue of creatures that haunt the night, such as mice, moles, and bats; and the gorgeous tints of fishes that swim among the coral reefs. These local color adaptations of animals are of great use to them, either enabling them to escape the notice of their enemies or to come upon their prey with the least risk of being detected. Certain groups of animals have a local color-adaptation, and may be noticed under the heading of Protective Coloration; others have acquired a wonderful resemblance to surrounding inanimate objects, such as leaves, twigs, bird-droppings, flowers, etc., and may be described under the heading of Protective Imitation of particular objects,[128] or Protective Resemblance; closely allied to the latter are those resemblances to surrounding objects which are not so much for the purpose of protecting the animals from enemies as for attracting their prey, and these resemblances, therefore, will be described under the title of Alluring Coloration. Other animals have such color patterns as to be very conspicuous in their surroundings; these animals are usually very poisonous or possess other deleterious qualities that cause beasts of prey to avoid them; hence these animals may be said to possess Warning Coloration. Many animals that are very desirable food for carnivorous creatures have acquired, in past ages, a remarkable resemblance to these dangerous animals with warning coloration, and are treated of under the title of Mimicry. There are still other groups of animals whose peculiar coloration enables the member of a herd or flock which may have become separated from the herd to readily recognize its companions at a distance as friends, and thus distinguish them from enemies. The color patterns of these animals may be classed under the heading of Recognition Marks.
Protective Coloration, or Local Color Adaptation. In forest-haunting animals of large size, such as forest-cats, and forest-deer, rounded spots are frequently noticed. Animals like the tiger, that spend a great deal of their time among high grasses and reeds, are striped vertically. The combined artistic effects of these spots and stripes, in connection with the lights and shades of the forest and the reeds, are such as quite effectually to conceal the animals from view: for the black stripes of the tiger, for instance, correspond with the black shadows of the reeds or[129] grasses; and his yellow stripes with the yellow of the reeds. In like manner the rounded spots of the forest-deer harmonize with the spotty shadows of the leaves in the forest. An experienced tiger hunter has stated that in following up a wounded tiger the natives saw the animal at a distance of about twenty meters, under a tree among the reeds, and pointed out the animal to him; but the color effects of the stripes of the tiger so harmonized with the artistic effect of the light and shade of the reeds as to effectually prevent his seeing the animal for a minute or so. The zebra, which is such a conspicuous animal in our zoölogical gardens, on its native soil and in a bright star-light night, may be so close to one as to be heard breathing, and yet cannot be seen, so completely are its color patterns in harmony with its native habitat. Marine organisms that float on the surface of the water are beautifully tinged with blue in harmony with the color of the ocean as it appears to their enemies above, the birds; while, looked at from below, they are white, thus harmonizing with the white clouds and the foam as seen by enemies from below.
There are many animals that are very conspicuous when removed from their native haunts, yet when in their proper environment are invisible or detected with the greatest difficulty. Such a large animal as the giraffe is effectually concealed by its form and color when standing among the broken and dead trees that exist on the edges of the thickets where it may be seeking its food. The odd shape of the head, with its horns that resemble broken branches, and the blotchy spots on the skin, so harmonize with its surroundings that even the keen[130] eyes of the natives sometimes mistake giraffes for trees and trees for giraffes.
There is a bat (Kerivoula picta) found in the island of Formosa that has a very conspicuous black and orange color. The body of this bat is of an orange color, and its wings are black and orange-yellow. When resting it suspends itself, head downwards, from the branches of an evergreen tree. During all the year some part of the foliage of this tree is undergoing decay, so that many of the leaves assume tints of orange and black. When the bat is suspended among such decaying leaves its colors and those of the leaves so harmonize that the animal is perfectly concealed, and thus eludes its enemies.
The sunbirds of Africa are very conspicuous when out of their natural environment, being brilliant and gorgeously colored. These birds find their main food supply among plants that have very conspicuous flowers; the aloe-blossoms, especially, which they frequent, are brilliantly colored. The colors of these birds so completely harmonize with the gay colors of the blossoms that even the keen eye of the hawk is unable to detect them. One species of these birds, the black sunbird, is never absent from a forest tree known as the Kaffir boom. This tree has not a single green leaf on it, but consists of a great mass of purplish-black and scarlet blossoms. A dozen of the black sunbirds may be feeding in this tree, and their notes may be heard among its branches all the day, yet their color adaptation to their environment is so complete that they are seen with the greatest difficulty or are entirely invisible.
Those birds whose colors have varied in such a way that they have not harmonized with their surroundings,—that have not become adapted to their environment,—are seen by the hawks and are exterminated; but those whose colors were more favorably arranged and more in harmony with their surroundings have been most often undetected by their enemies, the hawks, and have lived and transmitted their useful color patterns by heredity to their offspring. This process going on, age after age, has developed such perfect adaptations of the sunbirds to their environment as the naturalist observes at present.
These examples are illustrations of the fact that those creatures that are most in harmony with their surroundings are the ones that live and procreate their kind. They are cases of the survival of those animals that are best adapted to their environment,—the survival of the fittest; the selection by nature of favored creatures; in short, Natural Selection.
Among our native birds, the woodcock and snipe have such tints and markings as strikingly to harmonize them with the dead marshy vegetation which constitutes their native haunts. The ptarmigan in winter has a light coloration in harmony with the environment of snow, while in its summer plumage it is tinted and mottled in harmony with the color effects of the lichen-covered stones among which it spends a great deal of its time. Young unfledged plovers are spotted in such a way as very accurately to resemble the beach pebbles among which they remain for protection.
The white-headed fruit pigeon (Ptilopus cinctus) is a very conspicuous bird when taken from its[132] native haunts. It has a pure white neck and head, black back, yellow belly, and well-marked, deeply-curved black band across the breast, and black wings. It is a handsome as well as very conspicuous bird. It frequents trees that are a species of Eucalyptus in the island of Timor, and which have very open foliage and yellowish or whitish bark. The pigeons may be sitting motionless on exposed branches of the tree during the glaring heat of the day. The yellow and white bark of the tree, the deep blue sky seen through the openings of the leaves, with the intense tropical sunlight casting black shadows of one branch upon another, make color effects that harmonize so completely with the color patterns of the pigeons that they are entirely invisible.[8] Here again the complete color adaptation of the pigeons to their environment has been accomplished through the agency of Natural Selection,—Nature selecting those pigeons most frequently that varied most in the direction of useful color patterns, by concealing them from their enemies, the hawks.
There are some birds that are not protectively colored, which therefore do not possess a local color adaptation. The common raven, which inhabits the Arctic regions, is black instead of being white. The raven is found as far north as any known bird or mammal. It is a powerful and fearless bird, and needs no protective coloring; and since it feeds on dead animals it needs no concealing coloration to enable it to secure its food.
The bright and conspicuous coloration of many birds’ eggs have often been looked upon as a[133] difficulty on the theory of adaptive coloration. But Wallace thinks that a careful consideration of the subject in all its bearings shows that in a great number of cases these colorations are instances of protective coloration. He further thinks that when we cannot see the meaning of the particular colors, we may suppose that in some ancestral forms they have been protective, and, being harmless, have persisted under changed conditions which rendered the protection needless. He states, in illustrating the protective coloration of eggs, that “the beautiful blue or greenish eggs of the hedge-sparrow, the song-thrush, the blackbird, and the lesser redpole, seem at first sight especially calculated to attract attention; but it is doubtful whether they are really so conspicuous when seen at a little distance among their surroundings. For the nests of these birds are either in evergreens, as holly or ivy, or surrounded by the delicate green tints of the early spring vegetation, and may thus harmonize very well with the colors around them. The great majority of the eggs of our smaller birds are so spotted or streaked with brown or black on variously tinted grounds, that when lying in the shadow of the nests and surrounded by the many colors and tints of bark and moss, of purple buds and tender green or yellow foliage, with all the complex glittering lights and mottled shades produced among these by the spring sunshine and by the sparkling raindrops, they must have a quite different aspect from that which they possess when we observe them away from their natural surroundings.” We have here, probably, according to Wallace, a similar case, of general protective harmony, to that of the green caterpillars with beautiful[134] white or purple bands and spots, which, though gaudily conspicuous when seen alone, become practically invisible among the complex lights and shadows of the foliage they feed upon.
Eggs that are not protectively colored are usually white or of some uniform pale color, and are concealed by the birds in covered nests or in holes in trees or in the ground. Many birds lay white eggs in open nests, but even here the devices for concealing them are very effective and interesting. In some cases, such as the partridge, the goat-sucker, and others, the birds have the habit of sitting close and almost continuously on the eggs. These birds are protectively colored. Ducks, pheasants, and other birds have the habit of covering their white eggs with dead leaves or other material when they leave the nest. There are some large and powerful birds that lay conspicuous white eggs in open nests. Such are the cormorants, herons, storks, pelicans, and others. But they guard their nests carefully, and are able to drive away any enemies.
It is thus seen that there are many devices by which birds’ eggs are protected, and these have all been developed through the agency of Natural Selection. For example, those exposed eggs of timid birds that varied most in the direction of protectively colored ones were the ones that were most likely to be overlooked by egg-eating creatures, and those that were least protectively colored, and therefore the most conspicuous, were the ones most likely to be destroyed. The protectively colored eggs being thus, in the nature of things, the ones that have suffered less destruction from enemies, have developed into birds that have transmitted, by[135] heredity, their characteristics to succeeding generations of birds’ eggs. And thus in the course of time have many of the colorations of eggs been developed from simple to elaborate patterns.
Many snakes, frogs, butterflies, caterpillars, and so on, are colored green in harmony with the foliage of the trees among which they live. By being colored green these animals are more or less effectually hidden from their enemies, such as carnivorous birds. If they were not concealed by protective coloration they would quickly be exterminated by hungry and greedy enemies. The green frogs are greatly protected by their colors from their enemies, the green snakes. If they were not so colored, they would quickly be exterminated by these snakes. Even protectively colored as they are, large numbers of them are caught by the snakes, which are also protectively colored and keener witted. It may be stated that the protectively colored frogs are hunting for their food, the protectively colored insects; but, at the same time, they try to avoid their protectively colored enemies, the snakes, though they are often caught; the protectively colored snakes are hunting for their food, the frogs, but, in their green surroundings, endeavor to avoid their enemies, the reptile-eating birds, though often unsuccessful. And so on, from day to day, the tragedies of the animal world are enacted.
It may be well to pause a moment here and ask why it is that animals are not completely exterminated which are thus perpetually being persecuted and preyed upon by stronger foes. Why, for instance, are any frogs left in a green locality,[136] when they are so eagerly sought for by their enemies, the snakes? What are the factors that permit the average or normal number of them to exist in the given locality? The most important of them have already been referred to separately, but it will be useful to recapitulate, briefly, a few of them now. One of the most important of them is the tendency of the frogs to increase in geometrical ratio. In the breeding season great numbers of young are brought into the world, thus reinforcing the depleted ranks of the adults. Another factor is the instinctive effort of each frog to avoid its enemies, and their protective coloration greatly facilitates their concealment and escape. Another factor is the circumstance that the snakes also have enemies. They must be wary and careful in hunting prey lest they be unduly exterminated by their foes, the snake-eating animals. Then another factor is the alternating day and night, by which the warfare, offensive and defensive, must be periodically checked. Then again, the hibernating season, winter, checks or rather suspends the life-destroying crusades. It may thus be recognized that many complex factors occur to explain the circumstance that a given locality generally contains the normal number of individuals that constitute the species of frogs.
The complex factors at work in these relations and inter-relations of animals and plants are so nicely balanced that with a comparatively stable environment the number of species of the different classes of animals may remain more or less constant. But suppose one of these factors is profoundly diminished, such, for instance, as a great[137] diminution in the number of the snake-eating birds. Then the snakes, not being much persecuted by their enemies, will be in a most favorable environment; they will increase greatly in numbers; they will be eagerly on the look-out for their food, the frogs; the frogs, being thus unusually persecuted and hard-pressed, will diminish greatly in numbers. Now, this will be a golden opportunity for the insects; their environment becomes much more favorable. The snakes do not bother the insects, but destroy their enemies, the frogs. Therefore, the insects increase greatly in numbers and profoundly affect the vegetation upon which they live and thrive. The injury to the vegetation may thus seriously affect an altogether different class of animals—the grass-eating or foliage-eating animals, like the ruminants.
It may thus be understood how profoundly complex are the relations and correlations of living creatures, and how a disturbance of some of the links in these living chains may very extensively affect the other links. The different creatures referred to above are probably the simplest illustrations that can be given of protectively colored animals. For here we have green animals adapted to a green environment—the green foliage of trees or grass. This adaptation has been brought about by Natural Selection. Suppose, for instance, that the habitat of the non-protectively colored ancestors of the frogs was for any reason unduly crowded by their enemies, the snakes. The frogs would be hard pressed and much persecuted. Suppose that among the young of these frogs there were some that varied in the direction of grass-green or leaf-green colors,[138] while others did not so vary, or possibly varied in the direction of even conspicuous colors, such as black or white. It is evident that the latter groups would be easily detected by the snakes and destroyed, while the green group would frequently escape notice and would thrive and procreate their kind. This process being repeated generation after generation, there would come a time when the given habitat would contain none but frogs with a protective coloration of green. All of these protective colorations of snakes, butterflies, and caterpillars, as well as of frogs, are adaptations. They are illustrations of the survival of the fittest; the survival of those best adapted to their environment—in short, Natural Selection.

Plate II.—Kallima paralekta. A butterfly of Sumatra illustrating the work of Natural Selection: C, butterfly with expanded wings (dorsal surface) which are conspicuously colored through Sexual Selection; B, same butterfly with wings closed (ventral surface) and presenting a close resemblance to a dead leaf, A, through the agency of Natural Selection. B, illustrates the Protective Resemblance of an animal to an inanimate object.
Protective Imitation of Particular Objects. Insects often exhibit a very great amount of detailed resemblance to the leaves, flowers, and twigs of plants among which they live. Those that live on grass are striped longitudinally, while those that feed on ordinary leaves have an oblique striation. There is a larva of a Georgia butterfly (Sphinx fuciformis) which feeds on a plant having small blue flowers and linear, grass-like leaves. This larva has a blue head and a green body striated like the leaves. The resemblance of the insect to its environment is very striking. There is another species that feeds on a plant with small red flowers situated in the axils of the leaves, and this larva has a row of seven red spots of unequal size, which corresponds quite closely with the size and color of the flowers. There is a caterpillar in Borneo that resembles a piece of moss with two exquisite pink-white seed-capsules. Its general hue is greenish,[139] with two little pink spots on its upper surface, and it is covered with hair. Its movements are very slow, and when eating it withdraws its head beneath a mobile fleshy hood that it possesses, so that its motions in feeding are not noticeable. When living in its native haunts it is all but impossible to detect it, so completely does it resemble the surrounding moss. Other insects resemble green or dead leaves in all their varieties of form and color, and to show what a great protection such resemblance affords to insects in concealing them from view the following observation of a naturalist in Nicaragua may be related. In that country there are armies of foraging ants that devour every insect they can catch. Among a multitude of these ants he observed a locust that looked very much like a green leaf. The ants, many of them, were continually running over the body and legs of the insect without detecting its character. In many parts of the world there are many butterflies (Kallima, for example) the under surfaces of whose wings very closely resemble dead leaves. They frequent dry forests and are rapid flyers. They are rather large, and the upper surfaces of their wings are quite showy, having bluish and orange colors (Plate II). It is their habit always to settle on some twig where there are decaying or dead leaves. In doing so it folds its wings together over the back, thus concealing the gay upper surfaces and presenting the protectively colored under surfaces. The resemblance to a dead leaf is much more striking from the fact that the short tails of the hind wings just touch the branch on which the insect rests and look very much like the stalk of a leaf. From this stalk a[140] dark curved line extends to the elongated tip of the upper wings, thus imitating the midrib of a leaf. On both sides of this midrib are oblique lines that are partly markings and partly nervures, which give the appearance of a leaf with its veining. The head and antennæ fit in such a way between the closed upper wings as not to interfere with that irregular outline which is characteristic of the withered and dry leaves. Often the closed wings are covered with small black dots gathered into circular groups that exactly resemble the minute fungi found on decaying leaves, and it is sometimes difficult to believe that the insects themselves are not attacked by a fungus. Wallace states that this wonderful imitation is most complete, and that in Sumatra he has often seen a butterfly enter a bush and then disappear as if by magic. He states that once he was so fortunate as to see the exact spot on which the insect settled, but even then lost sight of it for some time, and was able to discover it close to his eyes only after persistent and careful search.
The curious and interesting leaf-insects of Java are veined and colored in such a way that, with the leaf-like expansions from various parts of the body, not one person in a dozen can detect them when they are resting upon their food-plants right under one’s eyes. Other insects resemble pieces of stick (Plate III), with all the minute details of branches and knots. An eminent naturalist has stated that after being a practical entomologist for thirty years, he was deceived by one of these stick insects and took out his pruning knife to cut from a plum-tree what seemed to him to be a projecting spur. This spur proved to be the caterpillar of a geometer-moth,[141] about two inches in length. He placed a portion of the plum-tree on a table, and showed it to several members of his family, designating a space of several inches in which the caterpillar was to be found, but none of them could detect the insect until it was pointed out to them. These protective resemblances of living creatures to inanimate objects are beautiful illustrations of adaptation to environment through Natural Selection.
Beautiful illustrations of protective resemblance to particular objects are furnished by leaf-hoppers (insects) in Central America. They resemble the thorny and prickly growths of the plants on which they presumably live. Some of them also resemble gall growths on the plants (Plate IV).

Plate IV.—Central American Leaf Hoppers resembling the prickly and thorny growths of plants on which they presumably live. Certain of them also represent gall growths on the plants. Protective Resemblance. [Figures collected by Dr. L. O. Howard, from various plates published in the Biologia Centrali-Americana.]
Alluring Coloration. Besides those insects which secure protection from enemies by their resemblance to the inert objects among which they live, there are others whose adaptive resemblance, and therefore concealment, is for the purpose of securing their food,—for alluring their prey. A most interesting case of alluring coloration is that furnished by a wingless insect of India, the mantis. Its color and form are such as to closely resemble such a fantastic flower as the pink orchis. The insect rests motionless among the bright green foliage, being very conspicuous on account of its pink color, and looking so much like a flower that it allures and captures the butterflies which settle upon it.
There is a species of spider (Thomisus citreus) of a creamy-white color whose abdomen completely resembles in color and contour the unopened buds of the flowers among which it rests. It has been[142] seen to capture flies that were attracted to the flowers. There is another species of spider that looks exactly like the excreta of birds, and through this alluring resemblance captures certain butterflies. A naturalist has related how, in pursuing a butterfly through a jungle in Java, he was stopped by a dense growth of bushes. Here he observed a leaf with a bird’s dropping upon it, and sitting on this dropping was a beautiful butterfly. Surprised at such a usually dainty and pretty butterfly seeking such inappropriate food, he carefully approached to study the actions of the insect. The insect permitted him to get so close that he seized it by the wings, and to his astonishment a part of the body remained behind as if the bird’s dropping was very adhesive. He touched the dropping to see if it really was sticky, and found that his eyes had been deceived and that what he took for the excreta of a bird was a most artfully colored spider, lying on its back with its feet crossed and depressed closely to the body. The spider had been firmly holding the butterfly.
Plate III.—Caterpillar, B, of a Geometer Moth (Prochœrodes transverrata) on the stem of a plant (Ailanthus), A. Illustrating Protective Resemblance.
A very pretty illustration of alluring coloration is furnished by the common spider (Misumena vatia), which is often found spread out upon the yellow heart of an ox-eyed daisy and in like position upon Coreopsis (Plate V). It so closely resembles the flower upon which it lurks that the ordinary observer might well fail to notice its presence. The coloration facilitates the taking of prey and protects the creature from the assaults of enemies (Rev. H. C. McCook). It will be observed from the illustration that the flower is mostly yellow with some red; the same is true of the spider. This case is a very[143] instructive illustration of alluring coloration by which the insect has become a living bait for entrapping butterflies and other insects. Nature has brought about this complete color adaptation, from the variations of myriads of spiders. For those ancestral spiders that varied most in the direction of color adaptation to their environment were less often shunned by shy insects which could serve as prey. The spiders with useful color variations would thus most likely secure an abundance of food, and, thus living, transmit to many of their offspring their useful variations; while those with inharmonious, and therefore harmful, variations would not be so able to deceive their food insects, and would thrive poorly or starve altogether. This is another illustration of the survival of the fittest, the selection by nature of those best adapted to or in harmony with the environment; in short, it is Natural Selection.
Wallace, from whom many of these illustrations are taken, says that to many persons it will seem impossible that such beautiful and detailed adaptations and resemblances—and these are only samples of thousands that occur in all parts of the world—can have been brought about by the preservation of fortuitous useful variations. Yet this will not seem so surprising, continues Wallace, if we keep in mind the facts of the rapid multiplication of animals, the severe struggle for existence, and the constant variability of these and all other organisms; and, further, that we must remember that these delicate adjustments are the result of a process, Natural Selection, which has been going on for millions of years, and that we now see the small percentage of[144] successes among the myriads of failures. “From the very first appearance of insects, for instance, and their various kinds of enemies, the need for protection arose and was usually most easily met by modifications of color. Hence we may be sure that the earliest leaf-eating insects acquired a green color as one of the necessities of their existence; and, as the species became modified and specialized, those feeding on particular species of plants would rapidly acquire the peculiar tints and markings best adapted to conceal them upon those plants. Then, every little variation that, once in a hundred years perhaps, led to the preservation of some insect which was thereby rather better concealed than its fellows, would form the starting-point of a further development, leading ultimately to that perfection of imitation in details which now astonishes us.” So it is with the beautiful color adaptation of birds, mammals, lizards, and other animals. There is a lizard (Phrynocephalus mystaceus) inhabiting certain sandy districts in Asia, whose body is protectively colored and some of whose mouth-parts have alluring coloration and form. The general surface harmonizes with the sand in which it is found, while the skin at each angle of its mouth is of red color and so folded as to closely resemble a little red flower which grows in the sand. The lizard, being thus in harmony with its surroundings, resembling the sand and the flowers, is hidden from its enemies, the reptile-eating creatures. But at the same time insects, being attracted by what they take to be flowers, approach the lizards and are thus captured, being allured to their destruction.

Plate V.—A spider (Misumena vatia) lurking for prey on the center of a flower (Coreopsis). Illustrating especially alluring Coloration (for attracting prey), but also Protective Resemblance (against enemies). Reproduced from “American Spiders.” By courtesy of Rev. Henry C. McCook.
Warning Coloration. Many animals possess color[145] patterns that render them very conspicuous in their environment. It is a very interesting fact that most of these creatures are the possessors of some deadly weapons, as poison-fangs or stings, or that they are very disagreeable and unpalatable food for other animals. Warning colors are most abundant and best developed among insects. A family of butterflies (Heliconidæ), in tropical South America, possesses very pronounced and conspicuous color patterns, so that they are easily seen in their native haunts. Many of them have deep blue-black with vivid red, white, and yellow spots and bands, totally unlike those butterflies in the same locality that are protectively colored and palatable. Their bodies have juices that exhale a powerful odor. If one kill them by pinching the body, a liquid exudes that stains the fingers yellow and leaves an odor on them that can be removed only by repeated washing of the hands. There is a great deal of evidence to show that this odor is very offensive to insect-loving animals. Protectively-colored butterflies fly with great rapidity and are very wary and seek concealment; while the butterflies with conspicuous colors fly slowly, and do not conceal themselves, as if conscious that they have no enemies.
Many caterpillars have gay and conspicuous colors and do not conceal themselves. Bates noticed one in South America four inches long, striped across the body with yellow and black bands, and with bright red head, tail, and legs. It could be seen by any one who passed by, even at a distance of many meters. All of these conspicuous and brightly-colored caterpillars are unpalatable, and are refused as food by insect-eating creatures.
Grasshoppers and locusts generally possess green protective tints and are very palatable, but in tropical regions there are many species most gaudily decorated with blue, red, and black colors. They are inedible and are invariably rejected as food by lizards and birds.
A spider whose bite is exceedingly poisonous is found in Queensland. Its bite will kill a dog, and produces serious illness in man, with agonizing pain. It is black with a bright red patch on the middle of its body. This warning coloration is so conspicuous that even the spider-hunting wasp avoids it.
In all parts of the world frogs are usually protectively colored with browns or greens; and little tree-frogs are either curiously mottled to imitate dead leaves or bark, or they are green like the leaves they rest upon. These protectively colored frogs are always eagerly sought after by snakes and other enemies. But there are some frogs that are very conspicuously colored and that hop about with impunity, being avoided by the snakes and birds of prey. Such is a little frog in Nicaragua which with its “scarlet vest and stockings of blue” is very conspicuous in its native haunts.[9] Such, also, is a small toad found in South America, which is colored a bright vermilion and an intense black, which crawls about in the sunshine over the sands of arid places. Both of these animals are altogether avoided by the frog-eating creatures, because they have disagreeable properties that make them inedible.
In tropical America the very poisonous snake Elaps (Plate VI) is found abundantly. Its style of[147] coloration is very conspicuous and one that does not occur in any other group of snakes, consisting alternately of rings of red, black, and yellow, or red and black of varying width and arranged in different patterns. Snake-eating birds and mammals have learned, through hereditary experiences, to avoid these snakes with gay livery because they are poisonous and therefore dangerous. In Plate VII the conspicuously-colored black and yellow salamander is an animal with warning coloration. It is inedible and avoided by carnivorous birds. These warning colorations have been evolved through the Natural Selection of fortuitous useful color variations in the ancestors during the Geologic Ages.

Plate VI.—Illustrating Warning Coloration (Elaps) and especially Mimicry (Erythrolamprus). Elaps is a very poisonous reptile and Erythrolamprus is harmless. Reproduced (and modified) from Romanes’s “Darwin and after Darwin.” By courtesy of the Open Court Publishing Company.
Mimicry. Protective resemblance of a harmless animal to another of a different species that is harmful is known as mimicry. Mimicry is bound up with and altogether dependent upon warning coloration. Some beetles are protected by having integuments, etc., of very great hardness. Several genera of weevils are in this way saved from attack by insect-loving birds. These weevils are often closely imitated in appearance by softer and more eatable species of different genera from the weevils. Wasps and bees are often mimicked by insects of other orders.
Insectivorous birds are very active in hunting out the edible beetles (Longicornia), and everywhere in tropical regions these beetles so closely resemble other insects which are avoided by the birds that the longicorns are very frequently avoided and thus protected.
In tropical America many butterflies (Heliconidæ) are found that possess warning coloration. They[148] possess an offensive taste and odor which almost entirely exempt them from the attacks of insect-eating animals. The insectivorous birds have learned, by transmitted experiences (heredity) to avoid the Heliconidæ. It is an interesting fact that in the same locality with these distasteful butterflies are other species that are very palatable to insectivorous creatures; but they so closely resemble the non-edible species that the birds pass them by, not recognizing their character.
There are some cases of mimicry among birds. There is a genus of large honey-suckers known as friar birds found in the Malay Archipelago. They are noisy and powerful birds which go in small flocks. They have sharp beaks which are long and curved, and also powerful grasping claws. They are perfectly able to defend themselves, often driving away such birds of prey as hawks and crows when they approach them too closely. In the same environment are weak and timid birds known as orioles, which trust chiefly to their retiring habits and concealment for protection. The orioles, although an entirely distinct species from the friar birds, very closely resemble the latter. In each of the great islands of the Malayan Archipelago there is a distinct species of friar birds, and always in the same locality is a species of oriole that exactly mimics it. The separate species often look so thoroughly alike that competent naturalists, prior to a very close examination, have considered them as belonging to the same species.[10]
The most remarkable cases of mimicry are those in which poisonous snakes are mimicked by harmless[149] ones. There is an egg-eating snake in South Africa that possesses neither teeth nor fangs and is not poisonous. It very closely resembles the poisonous Berg adder. When alarmed it still more closely resembles the adder by the habit of flattening its head and darting forward as if to strike an enemy, hissing at the same time.

Plate VII.—An Amphibian (Salamandra maculosa) illustrating Warning Coloration.
In tropical America, in the localities where the poisonous genus Elaps is found so abundantly, are several genera of harmless snakes of other families, some species of which so closely resemble or mimic the poisonous species that they are distinguished from them with difficulty. The peculiar color patterns of the poisonous snakes serve as warning colors to snake-eating mammals and birds. The mimicking snakes by flying these danger-flags are protected.
In Plate VI Elaps is the venomous snake, and illustrates warning coloration; Erythrolamprus is the edible, non-venomous reptile that has acquired, through Natural Selection, a protective resemblance to Elaps, and illustrates mimicry. At the first glance these two snakes look very much alike; but a closer inspection will show that the detailed color patterns differ in the two cases. Elaps, though a very poisonous snake, has the reputation of not being venomous. This error has probably originated from the fact that it has a gentle disposition and mild temper, and also from the fact that no doubt it is frequently mistaken for the mimicking non-venomous species.
Both warning coloration and mimicry are interesting illustrations of adaptation to environment through Natural Selection; for the myriads of[150] ancestral forms were continually giving slight variations in color patterns, some of which were useful to the creatures and others harmful. Those that harmonized mostly with the environment gave their possessors an advantage in the fierce struggle for life,—the struggle for food and place and safety; and procreating their kind age after age, led to the perfection of mimicry as we behold it to-day.
Coloration as Recognition Marks. In gregarious animals, whether herbivora or carnivora, and whether mammals or birds, a ready recognition of their own kind at a distance, in the dim twilight, or during rapid motion, is of the greatest use, and probably often leads to the preservation of life. Gregarious animals will not usually permit a stranger in their midst. So long as these animals keep together they are generally safe from enemies; but a single animal straggling off by itself may become an easy prey to enemies. In such cases it is of the highest importance to an animal that it should have every facility for quickly discovering its companions at any distance within the field of vision. Also to the young and inexperienced of each herd some means of easy recognition is of vital importance. Recognition marks also enable the sexes to identify their kind readily. The necessity for easy recognition probably is at the basis of the bilateral symmetry in the coloration of animals.[11] In the struggle for existence those gregarious creatures that have the best color patterns for recognition marks are the most likely to get the scattered members of a herd together with the greatest rapidity, and thus to save them from their enemies. The surviving members,[151] transmitting their useful variations in recognition marks to their offspring, are thus able in the course of ages to bring into existence pronounced color patterns. Thus Natural Selection can account for much of the coloration in animals known as recognition marks.
One or two illustrations of recognition marks in creatures will be sufficient. A rabbit when alarmed and fleeing to its burrow displays a conspicuous, upturned white tail. The rest of the body is protectively colored. This conspicuous white surface of the upturned tail is a signal flag of danger. The rabbit mostly feeds during moonlight nights, or soon after sunset. The white upturned tails of alarmed rabbits serve as signals and guides to the feeble and young, and also to those at a more remote distance. Thus a number of rabbits, each following the one or two in front, are all able in the quickest manner to reach a place of safety.
The spring-bok has a white patch on the face and one on the sides. It also has a curiously well-marked white stripe above the tail. When the animal is at rest this last-mentioned white stripe is very nearly concealed by a fold of skin, but when it is in motion it comes into full view, like the upturned tail of the rabbit, and serves as a guide to friends. There are some animals inhabiting the Arctic regions that are not white,—they are not protectively colored. Such is the musk-sheep. This animal, though living in Arctic regions, is yet brown and conspicuous. Its safety depends upon its association in small herds. It is gregarious. Therefore it is of much more importance to this animal that it should be able to quickly recognize its companions[152] at a distance than that it should be protectively colored and so concealed from its enemies. So long as they keep together in herds they are abundantly able to protect themselves. This is an exception to the rule of local color adaptation that proves the rule.
Sexual Selection. Among most backboned animals it is the rule that both sexes should be alike in color. This is especially true among the fishes, reptiles, and mammals. But in birds the diversity of sexual coloring is very frequent. It is among this class, therefore, that Sexual Selection can best be studied. One of the most fundamental characteristics of birds is the greater conspicuousness of coloration in the males. In the tropical regions especially are found the most striking examples of divergence in sexual color patterns. In humming birds, the pheasants, the peacocks, chatterers, tanagers, and birds of paradise, the females are exceptionally dull-colored and plain, while the male birds are gorgeously colored and conspicuously attractive. The male birds of paradise, for instance, are not only brilliantly colored, but also have remarkable gorgets, plumes, and crests; whereas the female paradise birds are without these decorations and as plain as our thrushes in their ornamental plumage (Plate VIII).
The splendor of plumage which characterizes the male pheasants is entirely wanting in the females. The intense crimsons and pure whites, the gorgeous purples and blues of the male chatterers contrast strikingly with the dull browns or olive greens of the females. The sober hues of the females have been accentuated by Natural Selection.[12] When[153] the females were brooding on the eggs in their nests, those of them that had varied in the direction of conspicuousness would most readily be detected by their enemies, the hawks, and would be exterminated; but those that had quiet and dull ornamentation would most frequently escape discovery, and would pass successfully through the brooding season; thus living and transmitting their color patterns to posterity,—Natural Selection ever eliminating the conspicuous and preserving the sober-hued,—in the course of time the dull ornamentation of the females would become more and more pronounced.

Plate VIII.—Male and Female Paradise Birds (Paradisea minor). Illustrating the effects of Sexual Selection. The upper figure (male bird) is much more beautiful than the lower (female bird).
But another factor has been at work in accentuating the marked differences in the ornamentation of the sexes. This factor is Sexual Selection.[13] In the brooding season there is an intense rivalry among the males for the possession of the females. Among them the art of courting has become, indeed, one of the fine arts. The male birds, like the males of almost all animals, have stronger passions than the females, and with rarest exceptions are much more eager than the females. In courtship they display their adornments and accomplishments most zealously before the females; they strut around them in most eager courtship and pompous vanity, displaying the utmost rivalry. The males charm the females in various ways, such as dancing, or performing fantastic antics either in the air or on the ground; and then again by most melodious song. After man, the female birds appear to be the most æsthetic of all animals; therefore, those male birds that are the most pleasing songsters, or the[154] most attractive in their dances and fantastic performances, are the ones selected by the females for mating. In this selection the female birds have paid the minutest attention to fleeting fashions in strut and dance, in form and color,—the progeny of those males that have been selected by the females for mating, transmitting the inherited peculiarities of the parents, have tended more and more to stamp as fixtures these fleeting fashions, and in this way the males have become endowed with all sorts of decorations and accomplishments.[14] Thus have been produced in them the many forms of topknots, wattles, combs, plumes, and feathers elongated and springing gracefully from many portions of the body; also the naked skin of the head and the beak, frequently colored gorgeously. The feathers, through this means, are often most beautifully tinted in charming patterns. As bearing upon this theory of Sexual Selection, it can be observed that birds pay the closest attention to the songs of each other. A bullfinch, for instance, had been taught to pipe a German waltz, and in doing so was a most excellent performer. He was placed in a room where there were kept some eighteen canaries and linnets, and immediately commenced producing his melodies. The birds all ranged themselves on the sides of their cages nearest the performer, listening to his singing with the greatest interest. Undoubtedly this singing is most often a matter of courtship; the female finch selects that one out of a hundred males whose notes charm her the most; the female canary always chooses the best singer. The soft cooing of pigeons and of turtledoves is a matter of[155] courtship. In the breeding season there is the most intense rivalry between the males in singing; a bird will sometimes sing until he drops down almost dead.
That female birds exercise choice in mating was believed by Audubon. He describes how a woodpecker hen was followed by six gay suitors who continued to perform strange antics until a marked preference was shown for one of them. A study of the Australian bower-birds illustrates both the courting antics of the males and the exercise of choice by the females. These birds build bowers which are sometimes quite large. That of one species is raised on a thick platform of sticks and is nearly four feet in length and eighteen inches in height. These bowers are built on the ground, and are for the sole purpose of courtship, since the nests are formed in the trees. They are highly decorated with leaves, berries, feathers, shells, and kindred objects. Both sexes assist in the erection of the bower, although the male bird is the principal worker. The bower-constructing instinct is so strong that it is practiced even in confinement. A naturalist in describing the habits of some satin bower-birds kept in an aviary says that at times the male will chase the female all over the aviary, then go to the bower, pick up a gay feather or a large leaf, utter a curious kind of note, set all his feathers erect, run round the bower, and become so excited that his eyes appear to start from his head. He continues opening first one wing and then the other, uttering a low, whistling note, and, like the domestic cock, seems to be picking up something from the ground, until at last the female goes gently toward him and the wooing is completed.
Instances of love dances may be taken from all classes of the animal kingdom. “Mr. Peckham has described a very interesting love-dance by a certain species of spider (Saitis pulex). He placed a male in a box with a female. As soon as the former saw the latter, about twelve inches away, he became excited and at once moved towards her; when some four inches from her, he stood still, and then began the most remarkable performances that an amorous male could offer to an admiring female. She eyed him eagerly, changing her position from time to time so that he might always be in view. He, raising his whole body on one side by straightening out the legs, and lowering it on the other by folding the first two pairs of legs up and under, leaned so far over as to be in danger of losing his balance, which he maintained only by sidling rapidly towards the lowered side. The palpus, too, on this side, was turned back to correspond to the direction of the legs nearest it. He moved in a semicircle of about two inches, and then instantly reversed the position of the legs, and circled in the opposite direction, gradually approaching nearer and nearer the female. Now she dashes towards him, while he, raising his first pair of legs, extends them upward and forward as if to hold her off, but withal slowly retreats. Again and again he circles from side to side, she gazing towards him in a softer mood, evidently admiring the grace of his antics. This is repeated until are counted one hundred and eleven circles made by the ardent little male. Now he approaches nearer and nearer, and when almost within reach whirls madly around and around her, she joining and whirling with him in a giddy maze.[157] Again he falls back and resumes his semicircular motions, with his body tilted over; she, all excitement, lowers her head and raises her body, so that it is almost vertical; both draw nearer; she moves slowly under him, he crawling over her head, and the mating is accomplished.”[15]
In addition to that form of Sexual Selection where the female chooses a mate from among a number of competing males, and which may be designated preferential mating, there is another form of selection in which the males fight with one another for the mastery and the possession of the females. Among the higher mammals it is a very general fact that the males fight together for the possession of the females. This leads, especially in polygamous animals, to the better armed or stronger males becoming the parents of the next generation, which inherits the peculiarities of the parents. Thus the offensive weapons and the vigor of the males are continually increased, resulting in the antlers of the stag, the tusks of the boar, the fighting instinct and spurs of the gamecock, and the horns and strength of the bull. Even mammals that are not specially armed fight to the death for the possession of the females, such as beavers, moles, squirrels, and hares. Almost all male birds are especially pugnacious during the breeding season. Battles have been observed in such different groups as ducks, finches, woodpeckers, humming birds, and waders. Among fishes deadly battles occur between the males of sticklebacks. Also the males of salmons engage in deadly contests; among reptiles fighting occurs among the male tortoises, crocodiles, and[158] lizards. Spiders and many butterflies often fight for the females. Thus Sexual Selection through the law of battle occurs widely throughout the animal kingdom. This form of Natural Selection greatly increases the vigor and fighting power of male animals; for, in every case, the weaker males are either driven away, killed or wounded, and the field is left to the most vigorous for procreating their kind.
The male stickleback is a little fish that builds its nest among the weeds, weaving the material together by a secretion from its kidneys. It is a very passionate little animal, and is exceedingly pugnacious in relation to its male rivals. The battles of the males are often very desperate. The combatants fasten tight to each other for a time, tumbling over and over again, until they appear to be completely exhausted. The males of the rough-tailed stickleback, when fighting, swim round and round one another, endeavoring to pierce each other with their raised lateral spines. In fighting they are perfect little furies, and their bite is very severe. Their lateral spines are used with such fatal effect during a battle that a male has been observed to rip open his opponent, so that the latter sank to the bottom in a dying condition. The females are very peaceful. When they come out of their hiding-place and view the nest that the male has made he is mad with delight.
The male salmon is as pugnacious as the little stickleback. Two males have been known to virtually battle with each other all day long. In breeding ponds the males can be seen constantly fighting and tearing one another on the spawning[159] beds, and so many are injured in consequence that they may be seen swimming near the banks in a state of exhaustion, many of them apparently dying.
Among birds the law of battle holds as well as the law of preferential mating. During the breeding season they are exceedingly pugnacious. The humming birds, the smallest of any, are among the most quarrelsome. Two males rarely meet without a fierce fight on the wing.
Darwin gives the following illustration of the invincible courage and fighting instinct of the gamecock. One of these birds had both of its legs broken in a cockpit, and its owner made a wager that if his legs could be supported with splints and bandages so that he could stand upright he would keep on fighting. This was accomplished, and the bird fought on with dauntless courage until he received a death stroke.
It is probable that even with the most pugnacious species of birds the pairing does not depend alone on the courage and strength of the male, for these males are usually decorated with various ornaments. Furthermore, these decorations during the breeding season often become more brilliant, and are eagerly displayed before the females. Darwin states that twenty or more males of the Tetrao cupido (species of grouse) will assemble at a particular spot, keeping up a tremendous chattering and strutting about. At the first response from a female the males take to fighting furiously, and the weaker are vanquished. Both the victors and the vanquished pay court to the female, so that the latter must make a choice or the battle is renewed. Here we have the combined[160] action of selection through battle and by preferential mating.
Among mammals the males win the females much more through fighting than through the display of charms. In the breeding season the most timid male animals, which are not even supplied with any special weapons for fighting, engage in the most desperate conflicts. Two male hares have been seen to fight until one was killed. Those male mammals which are provided with special weapons for fighting enter into the fiercest and most deadly conflicts. The wild male elephant, during the period of love, is one of the fiercest fighters in the world. Lions engage in terrible battles, and a young lion dare not approach an old one.
Male seals fight most desperately during the breeding season, using both their claws and teeth. The conquerors appropriate the females and transmit their qualities to their offspring.
These are all cases of the survival of the fittest; of the survival of the males best adapted to the exigencies of their surroundings—in short, additional illustrations of Natural Selection.
Insect Selection. Flowers usually consist of several parts, such as the stem, the calyx composed of green sepals, the corolla formed of attractively colored petals, the stamens, the pistil, and finally of certain nectar-forming organs situated at the base of the last-named. The upper portions of the stamens are known as antheridia. The free extremity of each pistil is called a stigma, the intermediate stalk is the style, and the base is the ovary. The flowers constitute the sexual parts of the plant. The stamens are the male elements and the pistils are[161] the female structures. In some species of plants the flowers possess the stamens but not the pistils,—they are male flowers; in others the flowers have pistils but not stamens,—they are female flowers. In still other plants the flowers possess both the male and the female structures, and are therefore bisexual or hermaphroditic flowers. The ovary of a pistil has a number of cells in it called ovules (female germ cells); the antheridia on the stamens have cells in them called pollen (male germ cells). When a pollen cell is carried by any means to the stigma of a pistil, it sends down through the latter a tubular prolongation by which the nucleus and protoplasm of the pollen cell unite with the nucleus and protoplasm of the ovule; so that we now have a fertilized egg,—the germ of a new plant. This fertilized ovule by repeated cell multiplication can grow into an adult plant.
There are two agencies by which the pollen is carried to the pistil; first, by the wind, and, secondly, by insects (or, occasionally, by humming birds). Flowers that are fertilized through the agency of insects are the most beautiful in existence, displaying all the varied hues and gorgeous patterns that are found in the organic world. On the other hand, the flowers which are fertilized through the agency of the wind are incomparably less beautiful than those of insect production, and they do not secrete sweet juices or nectar. The earliest flowers in geologic time consisted only of those essential portions, the stamens and pistils, and had no colored whorl of petals within another colored whorl of sepals. The poorly developed nectaries secreted only small quantities of honey.[162] The food-seeking insects visited these primitive flowers for the pollen and nectar, even as they do now. The nectaries in the plants were so situated that the insects could not get at the honey without rubbing off some of the pollen and carrying it to other plants which it visited for honey. In this way the nectar-seeking insects would often carry pollen from the flower of one plant to the stigma of another plant’s flower, and thus secure cross-fertilization. Undoubtedly often the stigma of a plant’s flower was fertilized by pollen from the same flower; but it is known that seeds produced by cross-fertilization are much more vigorous and hardy than those produced by self-fertilization in the same flower; therefore, those plants that varied most in the direction best adapted for securing cross-fertilization would have decided advantages in the struggle for existence. Their fertilized eggs (seeds) would be most vigorous, and would most readily secure nutriment and withstand adverse circumstances. The variations of the primitive flowers most calculated to secure cross-fertilization would be those that made the flower more conspicuous by the appearance of bright colorations and delightful odors, such as would attract the attention and visits of nectar-seeking insects. These variations being transmitted, by heredity, to the descendants and accentuated as the ages passed would ultimately lead to all the wonderful adaptations of flower and insect structure to one another that are found in nature.
Plate IX.—Two Primrose flowers (Primula elatior). A, with a long style; B, with a short one. Vertical sections have been made through the flowers.
Natural Selection has acted in a double manner here, preserving those flowers that had the most delightful odors or the most conspicuous and enticing[163] colorations and patterns; and at the same time selecting those insects that varied most in the direction of keenness of scent, acuteness of vision, and color perceptions. It will be instructive to briefly describe the fertilization of the primrose and an orchis by bees. The primrose (Plate IX) has two sorts of flowers that are never found on the same plant; one has stamens high up the corolla tube and a short style with its stigma below the anthers; the other has the stamens far down the tube and a long style with the stigma above the anthers. In both cases the nectar is at the very bottom of the corolla tube. When a bee visits a short-styled flower, it extends its proboscis to the bottom of the tube and in withdrawing it brings away some pollen cells at its base. If it should next visit another short-styled primrose, it would not fertilize it because the pollen on the base of the proboscis could not reach the stigma; it would only gather still more pollen. But when it visits a long-styled primrose, it will necessarily deposit some pollen cells on the stigma, inasmuch as that is at the commencement of the corolla tube. If the bee should first visit a long-styled form of flower, it will carry off pollen on the tip of its proboscis, and when it visits a short-styled primrose will deposit the pollen on the stigma.
In the orchis the stigma is placed below the anther. The stigma is in the front part of the flower and at the base of the lip, the latter being prolonged into a long tube full of nectar. The stigmatic surface is composed of very viscid matter. A bee when seeking the nectar pushes its head against the anther, and in so doing detaches the two sticky glands to which the club-shaped masses of[164] pollen cells are attached (Plate X). It carries these away on the front part of its head. So long as the masses of pollen cells remain erect on the bee’s head, they cannot reach the stigma of any other orchis that it visits. As the sticky glands on the head contract, the pollen masses incline forward and become horizontal, so that they touch the next sticky stigma that is visited. The greater stickiness of the stigma detaches the pollen masses from the bee’s head, so that the flower is fertilized. It takes so long for the pollen masses on the bee’s head to assume the horizontal position that the insect has visited all the flowers on one orchis and then visits another plant. By this time the pollen masses are in the proper position (horizontal) to fertilize the flowers of another plant. In this way cross-fertilization is secured and the vigor of the plant maintained.
It may now perhaps be appreciated how intimate are the relations between the form and habits of insects and the structure and coloration of flowers. Our standards of beauty have largely been created for us through Insect and Sexual Selection. If insects had not been developed on the earth, plants would not be ornamented by beautiful flowers, but would show only such poor and inconspicuous ones as are found on grasses, on oak, ash, and fir trees. Grant Allen has beautifully written that, “while man has tilled only a few level plains, a few great river-valleys, a few peninsular mountain slopes, leaving the vast mass of earth untouched by his hand, the insect has spread itself over every land in a thousand shapes, and has made the whole flowering creation subservient to his daily wants. His[165] buttercup, his dandelion, and his meadowsweet grow thick in every English field. His thyme clothes the hillside; his heather purples the bleak grey moorlands. High up among the Alpine heights his gentian spreads its lakes of blue; amid the snows of the Himalayas his rhododendrons gleam with crimson light. Even the wayside pond yields him the white crowfoot and the arrowhead, while the broad expanses of Brazilian streams are beautified by his gorgeous water lilies. The insect has turned the whole surface of the earth into a boundless flower-garden, which supplies him from year to year with pollen or honey, and itself in turn gains perpetuation by the baits that it offers to his allurement.”
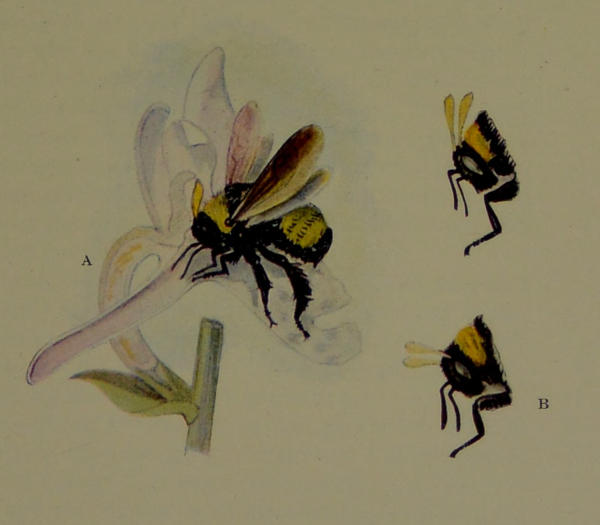
Plate X.—Illustrating the fertilization of an orchis by an insect. A, represents a section of the flower and shows a bee standing on the flower’s lip with its head touching the sticky portion of the pollen masses; C, shows the pollen masses stuck to the bee’s head and erect; B, shows the pollen masses horizontal, the proper position to leave them detached from the bee by the sticky stigma.
Natural Selection is the great agency that accounts not only for the color patterns and forms of living creatures, but also for the great majority, if not all, of the useful characteristics of organic creatures, including their internal organization.
One more illustration of the power of Natural Selection may be given, outside of the subject of color patterns. It may be interesting to readers who are not familiar with the elements of zoölogy to know that whales are not fishes but mammals. They belong to the same class that man does. The embryology of a whale reveals that it is descended from ancestors that were land mammals, and that these mammals had a scanty covering of hair, teeth of different shape, broad tails like beavers, short fore and hind legs, and well developed sense organs. The olfactory organ was especially well developed. It is probable that these ancestors of the whale lived in marshy districts and were omnivorous, eating[166] both plant and animal food. They sought their food in both shallow water and in swamps. As the conditions of life became more and more unpropitious on land, they were slowly modified through the ages under the action of Natural Selection into creatures somewhat like dolphins. At first they lived in fresh water, but finally they found their way into the sea and became the rulers of the ocean, from which the giant sea reptiles of earlier epochs had vanished. Hence are explained the adaptive changes of structure: the fore-limbs were modified into flippers enclosed in a fin-like sac, but retaining the bones corresponding to like structures in other mammals, as in the arm of man, the wing of the bat, and the fore-leg of the horse. Traces of the hind legs may be detected in a few species; the tail, which acted as a powerful swimming organ, became divided into two lobes; the head became fish-like in shape; the seven bones of the neck, common to most mammals, grew together; the skin became hairless; and the teeth, which appear in the young of the true whale, but are never cut, gave place to hanging fringes of whalebone, in the meshes of which the animal entangles the minute organisms it feeds upon.[16]
The following analysis of Isolation will be useful:
| Isolation | { | Sexual. | { | Preferential Mating. |
| { | Cross-Sterility. | |||
| { | Geographical. | |||
It has been stated on an earlier page that the commingling of diverse hereditary units accomplished[167] through the fertilization of an ovum by a spermatozooid is the source of many variations in the offspring. In this fertilized ovum the complexity of chemical substance, and, therefore, the complexity of inheritance, gives instability to the embryo, and thus produces variations in the offspring. In this embryo there is a struggle among the hereditary units,—a struggle among the various qualities inherited from both sides,—and a survival of the fittest, a veritable intra-cellular Natural Selection. It is a well-established law in biology that the union of germ cells of very closely related individuals, that is, of consanguineous individuals (in and in breeding) leads to less vigorous and variable offspring, and the parents are less prolific; while the commingling of diverse heritages by the union of germ cells from individuals belonging to strong but different varieties leads to vigorous and quite variable offspring. The union of such individuals is also most prolific. On the other hand the union of individuals belonging to very diverse varieties becomes less and less prolific until the cross-sterility of species is reached, although there are many exceptions to this rule of cross-sterility. The individuals of a species living in a state of nature are constantly varying. With every generation trivial variations take place in all directions and of all kinds. But these variations are all funded in the common stock, for the varieties freely mingle among one another and cross-breeding is constant not only between them, but with the parent stock. Of the variations that are constantly taking place some are advantageous to the creatures, some are disadvantageous, while many are neutral, being neither useful nor harmful. Natural Selection[168] is ever alert, selecting the advantageous variations and eliminating the disadvantageous ones. The neutral variations are not touched by Natural Selection; among these intercrossing of varieties probably affords Nature an opportunity to make almost endless combinations, some of which might be useful to the animals, and others harmful, and in either case would come under the influence of Natural Selection. The commingling of diverse heritages due to the union of more or less pronounced varieties of the species in nature not only leads to a funding of varietal characteristics, but also increases the instability of the offspring, augmenting their plasticity, so that more numerous and diverse variations take place. According to Romanes the reproductive organs are among the most variable in the body. Of the numerous variations taking place in the individuals of a species under nature, some, therefore, affect the reproductive organs in such a way that certain of these individuals are cross-fertile with one another, but cross-sterile with other varieties and with the parent stock. This interesting and very important kind of variation is known to occur in some individuals of the human species. It is well known that a man and woman have been cross-sterile with one another, being unable to have children; yet when separating and mating with others they have both been cross-fertile, families being reared by both of them.
Variations are commencing species; isolated variations diverge more and more into distinct species. This fact, then, of the occurrence in nature of variations in some of the individuals of a species by which they are cross-fertile with one[169] another, but cross-sterile with other varieties and with the parental stock, shows that Nature has a most effectual means by which varieties may be isolated from one another,—just as effective means as man, the breeder of varieties of domestic animals, possesses in isolating these domestic varieties by physical barriers, as fences, etc. Cross-sterility, therefore, in nature, is a most effective sexual barrier. The special form of it under consideration is what Romanes has called Physiological Selection. The varieties that are isolated by this sexual barrier have got to run the risks of in and in breeding, which Darwin has shown occur in domesticated animals, but which Wallace thinks are much less in a state of nature.
Another very important mode of Nature for isolating varieties is that which arises out of the instinctive preferences of animals. There is a tendency for like to breed with like where varieties are formed. The pale and dark colored herds of fallow deer in the Forest of Dean have never been known to interbreed. In the Falkland Islands all the cattle are known to have descended from the same stock. Here there are differently colored herds of cattle, and those cattle of the same color will interbreed with each other, but not with individuals outside their own color-caste, as Morgan expresses it. When two flocks of heath sheep and merino sheep are mixed together, they do not interbreed. This isolation of varieties by instinctive preferences for those individuals with like color patterns may be spoken of as preferential mating through recognition marks. A very obvious mode of isolation in nature is by geographical barriers (including migration).[170] In treating of environment we learned that during the geologic ages of the globe, the physical geography and climate have repeatedly changed. A very cold temperature, a mountain chain, a body of water, a stretch of desert land, may completely prevent interbreeding, on either side of the barrier, between the individuals of a species.
It should now be understood that Nature, like man, produces divergences from the parental stock, and isolates them by various effective agencies. Natural Selection ever carefully watches over these processes, eliminating the unfit variations and selecting the useful ones. Variation, Heredity, Environment, Isolation, and Natural Selection, having been acting and reacting through the ages, have produced, from a common parental stock, all the innumerable divergent and adaptive forms of living creatures that can be traced through the geologic strata to those on the surface of the earth to-day; have produced, therefore, man also as the inflorescence of the topmost branch of the tree of life.
The detailed study of the development (ontogenetic and phylogenetic) of man is so vastly intricate and extensive a subject that it will be impossible in a work of this character to do more than refer to it in brief outline. One of the best and simplest methods to approach the study of such a subject is to acquire some idea of the development of the frog.
The fertilized frog’s egg, which is the starting point in the life of a new frog, is deposited in water and hatched by the warmth of spring. After fertilization the egg or cell divides into two cells, these two into four, the four into eight, the eight into sixteen, and so on till a large number of small cells, associated together, is formed (Fig. 10). These cell phases represent different stages in the life of the growing frog. It is thus seen that the frog, in its earliest stages, consists of nothing but many small cells. These cells, through the mysterious powers of heredity, are going to differentiate as development proceeds into the various organs (groups of various kinds of cells) that form the tadpole and finally the frog. For further stages in the development of the egg (oösperm) towards the tadpole, works upon Comparative Embryology should be consulted.
At length the tadpole is hatched from the egg, and then soon swims about in the water (Fig. 17).
The creature that comes from the egg looks nothing whatever like a frog. It has no limbs whatever, and consists mainly of a bulky head and tail. This is the tadpole stage in the development of the frog. It can exist only in water, breathing air therefrom by means of gills. Like the fish, it has a two-chambered heart. At this stage it has no lungs, and the gills consist of an external (Fig. 17, a) and an internal pair. The mouth is small, with only horny toothless jaws, with no tongue. The creature is herbivorous, living on decaying vegetable matter. The vertebræ of the spinal column are bi-concave, as in fishes. The tadpole is essentially a fish, and would be so classed if it did not develop further. An evolving fish does not go beyond this stage. But the developing frog does go beyond this stage to a higher one. As its evolution proceeds through the multiplication and differentiation of the cells that form its body, limbs begin to bud out, first posteriorly (Fig. 17, b) and then anteriorly (Fig. 17, c). The lungs now begin to develop, and the external gills dwindle more and more until they soon disappear, the internal ones persisting for a while longer. The tongue, at this stage, also makes its appearance. The creature now can breathe both air and water. This is the permanent condition of many adult amphibians belonging to a lower order than the mature frog, such as the siren, menobranchus, etc. The siren in developing also passes through the fish stage, but does not get beyond the siren stage. But the evolving frog does go beyond this stage, for with the growth of the legs the tail[175] dwindles slowly by its gradual absorption (Fig. 17, d). The internal gills now disappear through absorption, and the lungs develop more thoroughly. Great changes take place in the blood-vascular system, the fish-like, two-chambered heart evolving into the three-chambered, amphibian heart. In spite of its dwindling, the tail is still a very conspicuous organ. In this phase of its development the frog can breathe only air, and must frequently come to the surface of the water for that purpose, and soon leaves the water altogether. Now this stage of the creature’s development corresponds to the permanent adult condition of another order of amphibians, which is higher than that to which the siren belongs but lower than the order of the adult frog. This intermediate order has such creatures in it as the triton. The triton in developing passes[176] through the fish and siren stages, but does not get higher than the triton stage. But the evolving frog goes even higher than this triton-like condition. Its tail is more and more absorbed until it finally disappears, and then the young but perfect frog appears (Fig. 17, e). During this period the teeth develop and the creature becomes carnivorous, feeding on insects. It is thus seen that the developing frog passes by small gradations from one class (the fish class) to an altogether different and higher class (the amphibian class). When it has evolved to this higher class, it then passes from the lower order (“siren” order) to a higher one (“triton” order), and then to the highest order (“frog” order). The bi-concave vertebræ of the fish-like tadpole have now developed into vertebræ with the cup-and-ball joints of the higher amphibian. It is the same with all the complex organs of the adult frog; they evolve from the much simpler structures of the tadpole.
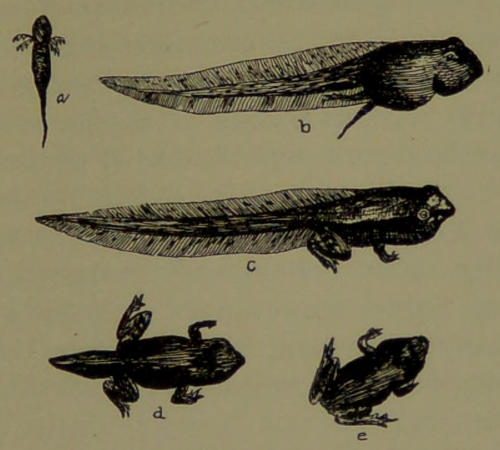
Fig. 17.—Tadpoles and Frog; a, tadpole with branching external gills; b, gills absorbed and hind legs have appeared; c, fore legs have appeared; d, tail shrunk and legs enlarged; e, perfect, young frog,—tail entirely disappeared. The figures represent some stages in the life history of the frog.
This study of the frog’s evolution from the fertilized egg is profoundly instructive. It reveals to us, through direct observation, that a creature varies in its form and structure at succeeding intervals of time. These variations diverge more and more, so that specific, generic, and even ordinal and class distinctions are revealed as the development proceeds. Owen, the distinguished comparative anatomist, in speaking of the transmutation of one species into another in the course of geologic history, says, though with a hostile purpose in view, that in the metamorphoses of the amphibians we seem to have such process carried on before our eyes to its extremest extent. Not merely is one[177] specific form changed to another of the same genus; not merely is one generic modification of an order substituted for another, the transmutation is not even limited by passing from one order (Urodela) to another (Anura); it affects a transition from class to class. The fish becomes the frog (amphibian); the aquatic animal changes to the terrestrial one; the water-breather becomes the air-breather; an insect diet is substituted for a vegetable one. And these changes, moreover, proceed gradually, continuously, and without any interruption of active life. Such is the language of Owen in reference to these remarkable transmutations of the developing frog.
The development of the frog is a brief recapitulation, an epitome, through heredity, of the main transmutations of its ancestral forms in geologic time. It is not true that the embryonic phases in the development of a higher form always resemble the adult stages of lower forms. This may or may not be the case; but what always does occur is that the embryonic phases of a higher form resemble the corresponding phases of the lower forms. So far as the frog’s development is concerned, it is very instructive to know that the order of succession of its embryonic forms undoubtedly parallels the order of succession of corresponding forms in past geologic ages. Fishes appeared in the Upper Silurian rocks with amphibian characteristics. In the succeeding Carboniferous Ages the fishes still continued under new forms; but also the lowest forms of amphibians, the most fish-like forms, now appeared. They were somewhat like the sirens, they were perennibranchs. In the next succeeding[178] rocks, the Permian and Triassic, higher, triton-like forms appeared. They were caducibranchs. Finally, in the Tertiary rocks, the highest forms of amphibians are found, such as the frogs.
In order to understand the relation of Ontogeny to Phylogeny, it must be carefully borne in mind that the simple and lowly organized creatures on the globe at the first appearance of life were performing the two great functions that all living creatures perform, viz.: those of nutrition and reproduction. These functions imply that organisms were reacting to environment, and, therefore, undergoing modifications and adaptations; and at the same time the organisms were giving origin to offspring—they were reproducing their kind through heredity. As these simple organisms lived through the ages and became more and more complex by modifications and adaptations to an ever-changing environment, they still evolved their kind in reproduction. Every new adaptation gained by the parent was transmitted by heredity, in the course of time, to the offspring; every form and structure modified in the parent was modified by heredity in the offspring; and every structure lost by the parent was finally lost in the offspring. Just in proportion as the parents, through the ages, became modified, often becoming more complex by the addition of adaptation to adaptation, retaining some structures of their ancestors by heredity (through use) and losing others, eventually, through disuse; so the offspring of these modifying parents became correspondingly modified, and acquired by heredity the modified structures and habits of the parents, while losing other structures in time that the parents had lost.[179] Just as complex organisms of later ages have been evolved from the simpler organisms of earlier ages by the addition of adaptation to adaptation, in an orderly sequence (Phylogeny); so, therefore, the complex offspring, while growing, unfold these inherited adaptations in the order of their acquisition. This last process is called Ontogeny or Embryology. Ontogeny is undoubtedly an illustration of the results of Natural Selection’s activity; for, during the phylogeny of the frog throughout the incalculable ages of the past its ancestors undoubtedly assumed innumerable forms and structures which were adaptations to the times and surroundings. But with the advancing time and changing environment, some of the old forms and structures continued useful and were retained, while others became useless and were eliminated by Natural Selection. In addition to the old useful structures that were retained changing environment often modified some of the retained structures and added still other adaptations to these. And so on, throughout the ages, in building up a frog, through geologic embryos, geologic “infants,” geologic “children,” and finally geologic adult frogs, Natural Selection has retained during ontogeny many useful structures in the order of their first appearance, and eliminated innumerable others that became useless. The ontogeny of the frog, which has been built up by its phylogeny, reveals the useful structures that have been retained, and in the order of their appearance; often showing structures that have been lost in the parent, but are not yet quite lost in the embryo, while it fails to show innumerable useless structures that have been lost in the past. This is[180] the reason why we say that the ontogeny of a frog is a brief outline recapitulation of the main points in the phylogeny of the frog, with even some main points occasionally omitted altogether. The geologic ancestors of the frog were the scaffoldings by which it climbed from simple creatures up to its present complex organization; just as the embryological phases at present are the scaffoldings by which a simple, unicellular, fertilized ovum climbs up through heredity to the huge complexity of the multicellular adult frog. What is true of the development of the frog, ontogenetically and phylogenetically, is also true of all living creatures, and is therefore true of man.
Man, in his individual development, commences life as a small, microscopic cell—the fertilized ovum—which is only one-fifth of a millimeter in size. His first stage resembles an encysted protozoan animal. As cell-multiplication proceeds he soon gets into the morula stage, which resembles a colony of undifferentiated protozoans. He soon evolves into a stage which may be compared to a colony of protozoans some of the members of which have undergone differentiation. Then comes the gastrula stage, which is distinctly suggestive of a low metazoan, and in which the developing germ assumes fundamental anatomical qualities such as characterize lowly animals like polyps. Then, by gradual transmutations, the vertebrate characteristics appear; but it could not be said at this stage of development, if one did not know, whether one is observing a fish, an amphibian, a reptile, or a mammal. Finally, the developing man passes through his fish and reptile phases and reaches the mammal stage. But as yet[181] it cannot be said to which order the animal belongs. The evolution of the individual continuing, he finally assumes those anatomical characteristics that stamp him as belonging to the order of man.
The theory of evolution, then, teaches that this development of man in the course of a few short months, like the development of the frog, is a very condensed and abbreviated epitome of the evolution of mankind from primitive protozoans during the incalculable ages of the past.
Drummond has prettily written that “the developing human embryo is like a subtle phantasmagoria, a living theater in which a weird transformation scene is being enacted and in which countless strange and uncouth characters take part. Some of these characters are well known to science, some are strangers. As the embryo unfolds, one by one these animal-actors come upon the stage, file past in phantom-like procession, throw off their drapery, and dissolve away into something else. Yet, as they vanish, each leaves behind a vital portion of itself, some original and characteristic memorial, something itself has made or won, that perhaps it alone could make or win,—a bone, a muscle, a ganglion, or a tooth,—to be the inheritance of the race. And it is only after nearly all have played their part and dedicated their gift that a human form, mysteriously compounded of all that has gone before, begins to be discerned as the resultant.”
As has been stated in the introductory part of this book, if all the animals that have ever lived on the globe should be represented by a tree those existing on the earth to-day would be indicated by the topmost twigs and leaves, while the extinct forms[182] would be represented by the trunk and main branches. Just as the leaves, twigs, branches, and trunk of the tree have a common origin, viz., the seed that developed into the tree, so all the different species of animals of the present and the past are the trunk, branches, twigs, and leaves of the “tree of life,” and have had a common origin from a primitive protozoan cell (see Diagram of Development, Fig. 18). Therefore all creatures, living and past, have a more or less blood relationship.
The Diagram of Development will indicate in a very general way the possible track taken by a man as he evolved,—grew higher and higher as the central, straight trunk of the expanding tree of life,—during the geologic ages; and finally appeared as the inflorescence of the topmost branch of this central trunk. It is seen from this scheme that the tree of life commenced in a primitive cell. Without entering into any discussion of the various theories of evolution and epigenesis, we may say that the primitive protozoan contained potentially all the animal forms (each being a cell or group of cells) that have existed on the globe, just as the fertilized egg contains potentially all the tissues and organs (groups of cells) of the adult man.
As the tree of animal life unfolded and expanded—like a germinating seed—from the primitive protozoan, certain of the descendants evolved along the straight and central branch, through the primitive colonial protozoans, on through primitive vermes, and still on through primitive fishes (elasmobranchii), amphibians, reptiles, and on through primitive ornithodelphia (monotremes), and[183] didelphia (marsupials) to a primitive order of monodelphia, viz.: primitive primates. The evolution of man continued through primitive anthropoidea to primitive anthropopithecus. At this point we meet with the common ancestors of the higher anthropoid apes (chimpanzee and gorilla) and man.
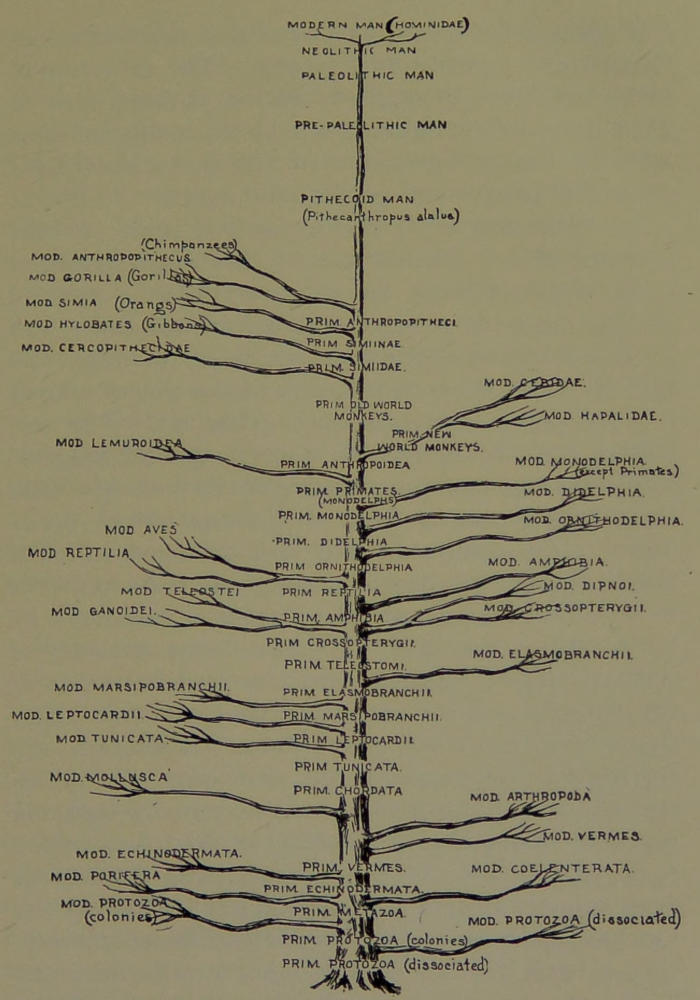
Fig. 18.—Diagram of Development: Portion of the “Tree of Life,” showing approximately the relative places of the great groups of animals. The Central Trunk and Primary Branches represent Primitive (geologic) forms; the Terminal Twigs represent Modern forms.
At each stage of the evolution some of the descendants of the animals of this stage diverged obliquely, modifying the characters they possessed at this stage in a direction that varied more and more from those characters that led on to man. So that all along the central trunk of the tree of animal life collateral branches were given off. The collateral branches given off at each upward stage of evolution represent animals higher in the scale than those that departed from the central trunk lower down. To illustrate what has occurred at each stage in the evolution of man, pause for a moment to consider that phase of progress represented by the primitive reptilia. If we study the anatomy of the specialized reptiles, birds, and monotremes of the present, we will find that they all have many characters in common. These characters are reptilian. Each class has its own distinctive specialized peculiarities in addition to its common reptilian characters. The study of the fossils of the rocks shows that in the Jurassic and Cretaceous ages animals existed that were undoubtedly reptiles, but had also very distinct bird characters; also reptiles existed that had distinct monotreme characters. These reptiles came from those of earlier times that were still more generalized. As the ages passed, some of the generalized reptiles (primitive reptiles) lost more and more the reptilian features[185] and gradually assumed more and more distinct bird characters, until finally the highly specialized modern birds (“glorified reptiles”) were evolved as a branch from primitive reptiles. The specialized reptiles of modern times likewise came from the primitive reptiles. In like manner those primitive reptiles that had mammalian (monotreme) characters, by getting into a suitable environment, gradually lost more and more their reptilian characters and assumed with increasing accentuation the characters of primitive monotremes—the lowest of the mammalian class. But observe particularly that the earliest introduced monotremes were not the specialized monotremes on the globe to-day, but generalized, primitive monotremes. These gave origin to the specialized modern monotremes, and also to the generalized primitive marsupials. The evolution of man continued through the primitive marsupials to primitive anthropoidea.
Here we meet with the common, generalized ancestors of man and the monkeys. These creatures contained, potentially at least, anthropoid as well as pithecoid characters. From them were derived the primitive New World monkeys (primitive Platyrrhines) and the primitive Old World monkeys (primitive Catarrhines). Some of the descendants of the primitive Old World monkeys, migrating into an environment which favored particularly the pithecoid characters, eventually developed into the tailed monkeys of the Old World (Cercopithecidæ). Others of their descendants, migrating into a different environment, found conditions that favored the anthropoid characters especially, and by greater and greater use of these, with[186] the diminished use of the pithecoid ones, the characters of the anthropoid apes (primitive Simiidæ and primitive Simiinæ) became clearer and clearer until, in time, primitive anthropopithecus appeared,—a tailless anthropoid ape of the Old World. More than likely this anthropoid ape bore a close resemblance, as Dr. Theodore Gill long since taught, to the modern chimpanzee. If there were any differences they could scarcely have been of even a generic value. This primitive chimpanzee was undoubtedly a quadrupedal, quadrumanous creature leading an arboreal life. His descendants specialized along two distinct but closely related lines. Those that continued to live in trees specialized along the oblique path that led finally to the gorilla on the one hand and the chimpanzee on the other. Those descendants that abandoned the trees and lived on the ground used the feet more and more for purposes of locomotion and less for grasping; while they employed with increasing frequency the hands for grasping exclusively. Associated with these adaptations were many other correlated adaptations, such as the upright posture, an enlarging brain, a change in the character of the face and of the dentition, etc. As man evolved further and further along the central trunk of the tree of life, he discarded, through disuse, many of the characters that are peculiar to the anthropoid apes; and assumed with increasing emphasis, through use, many of the characters that are distinctive of man. He passed through the phases of pithecoid man and pre-palæolithic (primitive) man, until eventually, in palæolithic man, the visage of humanity is clear and unmistakable.
It is extremely interesting to attempt to form some rough picture of primitive man. It may help us to do so if we recall what Darwin has said about the Fuegians, who are among the lowest of savages. He has written that they are men whose very signs and expressions are less intelligible to us than those of the domesticated animals—men who do not possess the instinct of those animals, nor yet appear to boast of human reason, or at least of arts consequent on that reason.
The Fuegians are much nearer to the ape than to a Shakespeare or Sir Isaac Newton. In the words of Clodd, primitive man was doubtless much lower than the lowest Fuegians. “He was a powerful, cunning biped, with keen sense organs (always sharper, in virtue of constant exercise, in the savage than in the civilized man, who supplements them by science), strong instincts, uncontrolled and fitful emotions, small faculty of wonder, and nascent reasoning power; unable to forecast to-morrow or to comprehend yesterday, living from hand to mouth on the wild products of nature, clothed in skin or bark, or daubed with clay, and finding shelter in trees and caves; ignorant of the simplest arts, save to chip a stone missile, and perhaps to produce fire; strong in his need of life and vague sense of right to it and to what he could get, but slowly impelled by common perils and passions to form ties, loose and haphazard at the outset, with his kind, the power of combination with them depending on sounds, signs, and gestures.”
Through the theory of evolution it can readily be understood why the anatomical characters of the anthropoid apes and of man are so very closely[188] alike. They have a common origin, and are blood relations—the one group of animals having specialized from common ancestors in one direction (obliquely), and the other group having specialized in another (straight) direction. (Vid. Diagram of Development.)
Man, in his individual development from a fertilized ovum, comes from a source infinitely lower than the ape. Why, therefore, should he feel such reluctance to believe that he has passed, during geologic ages, through the phase of generalized simian ancestors? Is there not much more of hope in the knowledge that he has risen higher and higher through the æons of the past than in the belief that he was created an innocent and noble character and then fell to utter wretchedness through great temptation? The motto of evolution is Excelsior. For it shows that the human race, through all the incalculable ages of the past, has risen to higher and higher levels,—to nobler and nobler phases of being. His progress in the almost infinite past suggests the hope that he will mount higher and higher towards perfection during the limitless future. Not only may we hope that there will be boundless improvement of the human race, but boundless evolution of each individual human being as well. Evolution’s motto for each individual may also be Excelsior. And, therefore, may we say with some assurance of hope that love, while kissing the pathetic lips of death, need not entertain in vain the splendid hope of immortality. For if there be no immortality of personal consciousness, then the evolution of the cosmos, of man, of the highest mind in man, have no intelligible meaning for us; they are[189] unfathomable enigmas—idiot stories without meaning.
Man, in specializing along certain lines since separating from the ancestral simian stock, has displayed more and more that structure of his skeleton and of the soft parts molded upon it that is best adapted to the needs of the mind resident within him. His bones are not merely the jointed framework of an animal, but a framework adapted to that erect attitude which so befits his intellectual nature. His feet are not the climbing and grasping feet of the ape, but organs for giving firmness to the tread and dignity to the bearing of a creature capable of high thought. The arms and hands are not for strength alone, for these members are much stronger in many a brute; but they also give greater expression and power to the thoughts that come from within. The hands possess such molding of fingers, thumbs, and palms, such delicacy for tactile impressions, and such capacity for nice adjustments, that they are not alone used for feeding the mouth and fighting antagonists; but they also contribute pre-eminently to the desires of a large mind, and are the efficient servants of its promptings. As Dana well says, “The face, with its expressive features, is formed so as to respond not solely to the emotions of pleasure and pain, but to shades of sentiment and interacting sympathies the most varied, high as heaven and low as earth,—ay, lower, in debased human nature; the whole being, body, limbs, and head, with eyes looking, not towards the earth, but beyond an infinite horizon, is a majestic expression of the divine feature in man and of the infinitude of his aspirations.”
But it is well to remember that man’s structure is riddled with evidences that he passed from an ancestral, quadrupedal condition, through the semi-erect to his present upright posture, slowly and laboriously. His erect attitude, geologically speaking, is a very recent accomplishment, and his anatomy, therefore, reveals many imperfect adaptations to his newly acquired posture. These imperfect adaptations are the sources of many grave diseases in mankind. It would require too technical a knowledge of anatomy to explain these imperfect adaptations, and I will therefore simply mention rupture and uterine displacements as due to imperfect adaptations to the upright attitude.
The common origin of man and the ape accounts for many interesting and otherwise inexplicable facts in anatomy. There is, for instance, a muscle that is normally present in the orang-outang known as the Opponens Hallucis. This muscle enables the orang to oppose his big toe to the other toes, just as we can oppose our thumb to the other fingers of our hand. This muscle is absent from the foot of man ordinarily. But occasionally it is found in man, in the dissecting-room, as a rarity—as an anomaly. The question naturally arises, why should this muscle be present normally in the orang and absent normally in man, occurring in the latter only as an abnormality? The theory of evolution gives the only rational answer. The man-like, ape-like generalized ancestors of man and the orang possessed this muscle, which was useful to them in grasping the branches of the trees among which they lived. These ancestors used the feet and hands alike for purposes of grasping (prehension)[191] and locomotion. But those descendants that evolved more and more man-ward used the feet more for purposes of locomotion and less for grasping, while they used the hands more for grasping and less for locomotion, until, finally, man was created—a creature that uses his feet exclusively for locomotion, and the hands entirely for grasping. Through disuse, therefore, the opponens hallucis gradually disappeared in man; so that now it occurs only as a rare abnormality. The hereditary units that make this muscle still lie dormant in most men are usually so weak, through disuse, that they do not develop. Some unusual stimulus occasionally causes the latent hereditary units to develop and makes it appear in man. The same is the case with many other muscles and structures that are normal in the modern anthropoid apes, and only occur as rarities in man. The appearance of those muscles in man are instances of atavism, i. e., reversions to conditions that were normal in the ancestors of man and the apes, as they are still normal in the latter.
Man, in his post-natal growth, as well as during his embryological development, exhibits reminiscences of his animal ancestry. In the structure and movement of the new-born babe, as well as in the adult frame, we find continuous witnesses to the ancient animal strain.
On the theory that men in bygone ages were closely allied to simian creatures in habit as well as structure; that they led an arboreal life; and that, like the baby-monkeys to-day, the baby-men of other ages clung to their mothers as they climbed among[192] the trees, Dr. Louis Robinson predicted that a baby’s power for grasping would likely be found to equal that of a young monkey which had reached a corresponding period of growth. He tested a large number of new-born infants in reference to this power by extending his finger or a cane, to imitate the branch of a tree, and observed how long they would hang there without any other support (Plate XI). He made experiments on about sixty children under a month old. About thirty of the children experimented upon were not over an hour old. Dr. Robinson states that each of the infants, with two exceptions, was able to hang to the finger or cane by its hands, like an acrobat from a horizontal bar, and sustain the whole weight of its body for at least ten seconds. Twelve of the infants, less than an hour old, held on for half a minute before the grasp relaxed; while four of this age held on for one minute. Over fifty of the infants when four days old could continue the grip for half a minute. Three weeks after birth the faculty for holding on reached its maximum, for at this age several succeeded in hanging on for a minute and a half; two held on for over two minutes; and one infant held on over two minutes and a half. One infant that was less than an hour old hung by both hands to Dr. Robinson’s finger for ten seconds, and then deliberately let go with his right hand, as if to seek a better hold, and continued his grasp with the left hand only, for five seconds longer. In none of these experiments did the limbs of the infants hang down in the attitude of the erect position, but the thighs were invariably in the baby-monkey attitude, at right angles to the body. The doctor says that this[193] attitude and the disproportionately large development of the arms compared with the legs give the photographs of the infants a striking resemblance to a well-known picture of the celebrated chimpanzee, Sally, at the Zoölogical Garden in London. In these experiments the infants very seldom gave any sign of distress, and uttered no cry until the grasp began to give way. The fact that the flexor muscles of the forearm of a new-born infant show such remarkable strength while the other parts of the muscular system are so conspicuously weak and flaccid,—that they are able to perform a feat of muscular strength that will tax the powers of many a healthy adult,—can be explained only on the theory of inherited instinct from simian ancestors that lived in trees. This instinct is no longer useful to an infant. It is a vestigial instinct, a useless scaffolding in its life history.
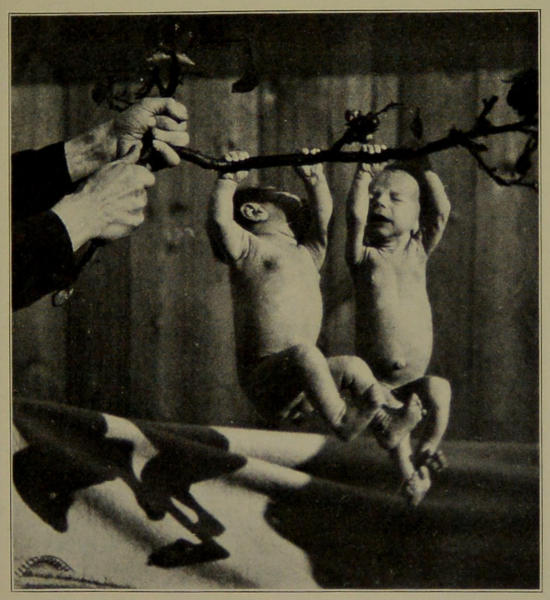
Plate XI.—Illustrating the grasping power of infants. Two infants, ten and thirteen days old, respectively, supporting their weight by the hands only (vestigial instinct.) Reproduced from a photograph taken by Dr. Louis Robinson. By courtesy of the Open Court Publishing Company.
Club-foot. There is an ordinary case of malformation in the foot of a child known as club-foot. The most common kind of this deformity is that where the sole is turned inwards and upwards and the heel is raised. Before birth all children pass through this condition as a perfectly normal and natural one, and only gradually outgrow it (evolve beyond it). But some children fail to evolve beyond this condition and have club-feet throughout life, unless relieved by the surgeon. It is a very instructive fact that this particular form of club-foot is the normal condition of the adult gorilla and orang-outang. The foot of every child passes through this gorilla phase, and if it does not develop beyond this phase it retains the simian characters, and we call it an abnormality. In this[194] abnormality the anatomist finds that those bones that enter into the formation of the ankle joint have the pronounced anatomical characters of the adult orang-outang.
Ribs. Adult man possesses twelve pairs of ribs. The chimpanzee and gorilla possess fourteen pairs. An older comparative anatomy predicted that in an early embryonic condition man would be found to possess thirteen or fourteen pairs. The prophecy has been verified.
Hair. The apes have hair over the entire body. At the sixth month of the embryonic development the human fœtus is thickly covered with a somewhat long, dark hair over all the body, except those parts that are uncovered in the apes, viz.: the palms of the hands and the soles of the feet. This covering of hair is called lanugo. Since it covers all the body except the points noted, it extends, of course, all over the ears, face and forehead. It is usually shed before birth. It is a simian characteristic, and sometimes fails to disappear, but persists and develops greatly. Therefore there are occasionally found such men (“dog-faced men”) as the Russian Jeftichjeff. The Ainos of one of the Japanese Islands also possess this extreme hairiness.
Vermiform Appendix. There are a number of vestigial structures in man that are not only useless but even a menace to life. The most striking of the vestigial structures that come under this category is a portion of man’s large intestine which is called the Appendix Vermiformis. This useless structure is a veritable death trap. In some animals, such as the herbivorous ones, the appendix is very[195] large, sometimes longer than the body itself, and is of great use in digestion. But in man it has shrunken to a small rudiment varying from two to six inches in length, which is very liable to a grave form of disease that frequently causes death unless timely treated by the surgeon. In the early embryo the appendix is equal in caliber to the rest of the bowel, but at a certain date ceases to grow pari passu with it. At birth it has become a small rudiment of the large intestine. In the new-born infant the appendix is often of the same size as it is in the adult. This precocity of an organ is always an indication that it was of great importance to the ancestors of the human species.
Tail. Man, like the anthropoid apes, has no external tail; but, exactly like them, he has a rudimentary one concealed beneath the skin. The embryos of man and the ape at an early stage of growth possess a very conspicuous tail, which is even longer than the limbs. In the embryo of man even the muscles for wagging the tail are still found. In the adult man these muscles are represented, normally, by bands of fibrous tissue. In the dissecting-room one occasionally finds these muscles well developed in the adult man. Man and the anthropoid apes have descended from more primitive simian ancestors that possessed tails.
Hearing. Prominent among vestigial structures, though less easy for beginners to understand, are those that point to piscine ancestors and which, therefore, smack of the sea. Embryology points indubitably to the fact that the ancient, geologic progenitors of man once lived a marine life. In the history of the globe there was a time when all[196] the animals lived in the sea. Land animals appeared as later creations. Man, in evolving from the primitive protozoan, passed through a marine-worm phase and finally, through the ages, attained to the fish stage. The chief characteristic of a fish is its apparatus for breathing the air dissolved in the water. This apparatus consists of gills—strong bars with delicate, highly vascular, fringe-like curtains hung on them, and through which the blood is continually circulating. The circulating blood throws out its impure gases and takes in from the water the pure air, thus breathing. These bars or arches are five or seven in number in many fishes. Slits extend from the surface of the fish between the bars to the throat, so that the water which the fish takes into its mouth is forced out between the bars, thus bathing the delicate curtains on them by which air is breathed from the water. Sometimes the slits between the bars are open and unprotected, as in the sharks; but in the modern fishes (teleosts) they are protected by a lid (operculum). If these slits did not exist in the neck all fishes would quickly perish. They are of so great use to the fish that Natural Selection has taken exceptional care in perfecting their mechanism.
It is one of the most interesting facts in evolution that these slits in the fish’s neck are still represented in the neck of man. One of the most prominent features in every mammalian embryo is the presence of four clefts of the old gill-slits. So persistent are these characters that children are occasionally born with persistent fissures leading to the throat, so that milk, when swallowed, will come out on the neck through an opening. Thus we have[197] a persistent piscine characteristic as an abnormality in the child.
When the fish-like ancestors of man left the water the elaborate breathing apparatus was no longer needed for respiration. Nature, in creating new adaptations for the land animal, did not discard the elaborate gill apparatus that had been evolved through the ages; but utilized this old apparatus for the new adaptations. Nature is exceedingly economical and does not discard old organs when they can be molded for new functions.
In the course of ages, through minute gradations, the first gill-slit and portions of its adjacent bars were molded for purposes of hearing. In man there are two passages leading to the drum or middle ear; one is the external auditory canal (the opening which is seen in what is popularly called the ear), and the other is a canal leading from the throat to the middle ear. In the adult these two channels are partitioned off from each other by the membrane of the drum. These canals are the counterpart or homologues of the spiracle associated in the shark with the first gill-slit. The external ear is developed by the coalescence of six rounded tubercles appearing in the bars or branchial arches that surround the first gill-slit. In the course of ages the remaining gill-bars (branchial arches) were also modified for special uses.
In relation with the external ear of man are found rudimentary muscles that are no longer useful and ordinarily are not under the control of the will. These muscles are the exact counterparts of well developed functional muscles found in great numbers of the lower animals. They are present in[198] man as vestigial structures, because he is descended from animals in whom these muscles were well developed and functional.
The anatomy of man reveals so many additional vestigial structures that we may look upon him as a museum of obsolete anatomies; he is an old curiosity shop containing many discarded tools, many outgrown and aborted organs. The lower animals as well as man contain so many useless (vestigial) structures among their useful organs, and they are so significant of a former state of things in which they were useful, that anatomists are willing to stake the theory of evolution upon their presence alone. Evolution explains a multitude of other facts about man that are inexplicable on any other theory.
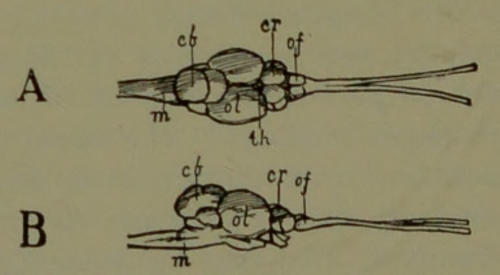
Fig. 19. Brain of Fish (Bluefish). A, dorsal view; B, side view; of, olfactory lobe; cr, cerebrum; ol, optic lobes; cb, cerebellum; m, medulla; th, thalami.
In addition to pointing out the possible track along which man has evolved from a primitive protozoan it would be interesting as well as exceedingly instructive to trace the development of each structure and organ in his body. But the subject is a vast one, and cannot be presented here even in briefest outline. Yet it will be very valuable to trace the unfolding of one organ, and that the highest,[199] as a sample of what occurs with every part of the body. I refer to the development of the brain.
Fig. 19 represents the brain of an average bony fish. It consists of six swellings in a line, one before the other. Beginning from the end towards the spinal cord, they are designated as follows, viz.: a single median lobe, the medulla (Metencephalon), m; then in front of this is another single median lobe, the cerebellum (Epencephalon), cb; then the optic lobes (Mesencephalon), right and left, ol; then the thalami (Thalamencephalon), which are small and hidden from view by the encroachment of the two adjacent segments; then the cerebrum (Prosencephalon), cr; then, finally, the olfactory lobes (Rhinencephalon), of. In this fish the largest of the segments are the optic lobes, ol.
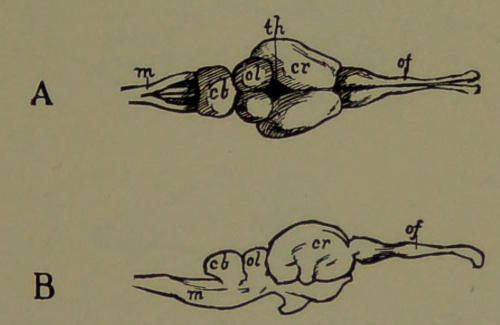
Fig. 20.—Brain of Reptile (Turtle). A, dorsal view; B, side view; of, olfactory lobe; cr, cerebrum; th, thalami; ol, optic lobes; cb, cerebellum; m, medulla.
The reptile’s brain (Fig. 20) shows similar parts with the same serial arrangement. The reptile is a[200] higher creature, a more intelligent animal, than the fish; and in consonance with this fact the cerebrum, cr, is the larger and more dominant part of the brain instead of the optic lobes, ol.
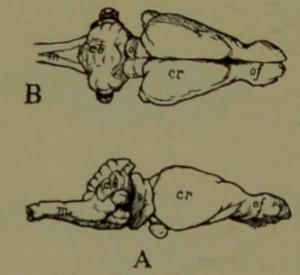
Fig. 21.—Brain of Marsupial (Opossum). A, side view; B, dorsal view; of, olfactory lobes; cr, cerebrum; ol, optic lobes; cb, cerebellum; m, medulla. Thalami concealed from view by the backwardly extended cerebrum; also the optic lobes are partially covered by the cerebrum.
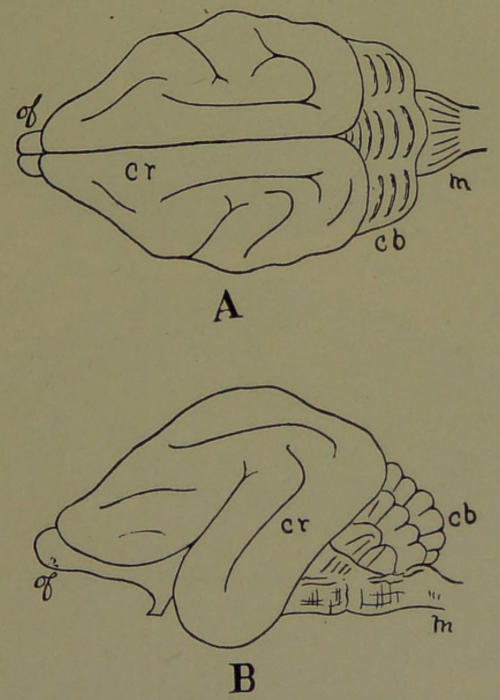
Fig. 22.—Brain of Lemur (Lemur nigrifrons). A, dorsal view; B, side view; cr, cerebrum; cb, cerebellum; m, medulla.
In the marsupial (Fig. 21), a more intelligent animal still, the cerebrum, cr, has grown so large that it extends backwards and partially covers the optic lobes. It is to be observed that in the marsupial the cerebellum, cb (like the cerebrum, cr), has evolved to a higher phase. It consists of a median lobe, cb, which is larger than the median cerebellum of the lower creatures mentioned, and of two lateral lobes, one on either side, which have been acquired in the course of evolution. The median lobe, the homologue of the single, median cerebellum of lower animals, is larger than the lateral ones. The cerebellum of the marsupial has its surface increased by fissures, while that of the fish and reptile is smooth. The fissured cerebellum is a[201] higher evolution than the smooth ones. In the groups of animals referred to so far the cerebrum is smooth and the olfactory lobes are still in front, though much encroached upon in the marsupial by the enlarging cerebrum. In those animals still higher in the scale of life, such as the prosimiæ (Lemurs), the cerebrum has reached yet larger proportions and complexity, and has grown still farther backwards towards the medulla, so that it hides from view a considerable portion of the cerebellum (Fig. 22); it has also grown forward, thus concealing largely the olfactory lobes. The cerebrum is no[202] longer smooth, but has a number of simple fissures and convolutions (the higher animals have numerous complex fissures and convolutions). The lateral lobes of the cerebellum have increased relatively more than the central lobe, and the whole organ has advanced in complexity of fissures. In the higher simiæ (monkeys and apes) the cerebrum has grown so far backwards as to almost completely cover the cerebellum and medulla, and its convolutions have become much more numerous and complex. The cerebellum has also grown greatly, and its lateral[203] lobes are now larger and more complex than the central lobe.
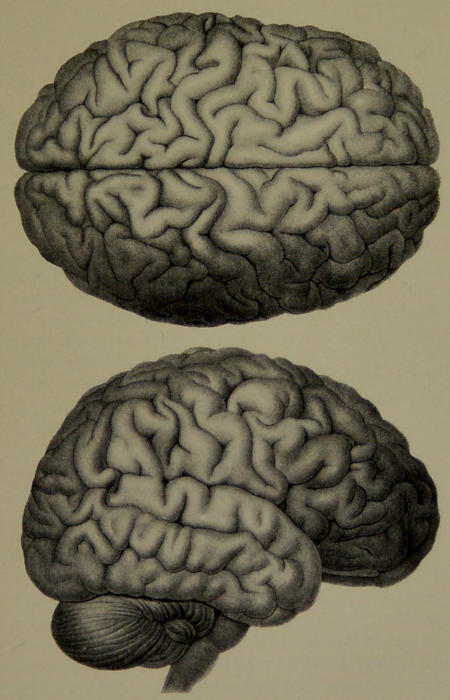
Plate XII.—Brain of man: dorsal and side views. The cerebrum has grown so far backwards and forwards as completely to hide the other segments of the brain when looked at from the dorsal surface. From Carus’s “The Soul of Man.” By courtesy of The Open Court Publishing Company.
Finally, in man (Plate XII), the whole brain has grown so enormously that it is three times larger than the brain of the highest simian creature. The cerebrum, especially, has increased enormously in size. It has grown not only backwards (overlapping cerebellum), upwards, and downwards on the sides, it has grown so far forwards as not only to cover the olfactory lobes, but also to project far beyond them. The cerebellum has also increased in size and complexity, especially the lateral lobes. The ideal vertical section (Fig. 23) shows diagrammatically in one figure all these stages in the evolution of the human brain through the geologic ages.
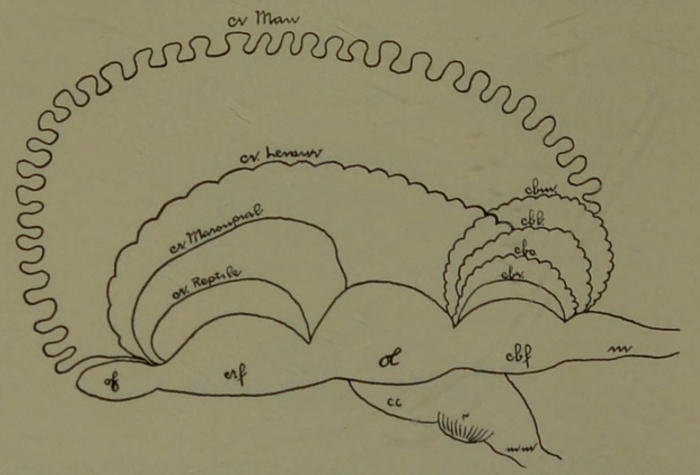
Fig. 23.—Ideal, vertical and sagital section, representing the ontogeny and phylogeny of the human brain. of, olfactory lobe; crf, cerebrum of fish; ol, optic lobes of fish; cbf, cerebellum of fish; m, medulla of fish; cbr, cerebellum of reptile; cbo, cerebellum of opossum; cbl, cerebellum of lemur; cbm, cerebellum of man; cr, cerebrum. Cerebrum convoluted in lemur; much more convoluted in man. Cerebellum convoluted from opossum upwards; mm, medulla of man.
Modified from Le Conte.
It is a very interesting and instructive fact that in the development of the human brain from the fertilized ovum these same stages, which are permanent conditions in the zoölogical (taxonomic) series, are passed through by it as transient stages.
One of the earliest conditions of the human brain is that in which it presents three swellings in a serial arrangement. They are known from behind, forwards as hindbrain, midbrain, and forebrain. For our purposes it is sufficiently accurate to say that the fœtal brain, in developing from this early condition to a later and higher condition, differentiates the hindbrain into the medulla (Fig. 24, m) and the cerebellum (cb); the midbrain becomes the optic lobes (ol); and the forebrain differentiates into the thalami (th) and the cerebrum (cr). A little later the cerebrum buds forth the olfactory lobes (of), so that the human brain will consist of six fundamental segments,—one behind the other.[204] This is the fish stage in the growth of the human brain. (Compare Fig. 24 with Fig. 19.)
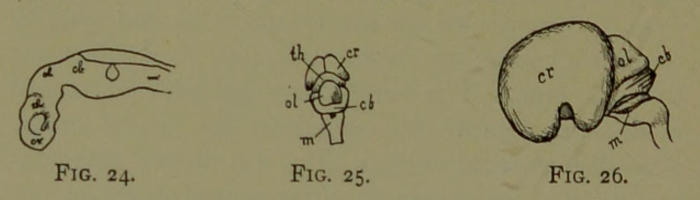
Fig. 24.—Diagrammatic representation of the brain of a human fœtus of the third week. Representing the fish-phase in the ontogeny of the human brain. Side view. cr, cerebrum; th, thalamus; ol, optic lobes; cb, cerebellum; m, medulla. The olfactory lobes at this stage are very small and are not shown.
Fig. 25.—Dorsal view of the brain of a human fœtus of about seven weeks. Representing the reptilian phase in the ontogeny of the human brain. cr, cerebrum; th, thalami; ol, optic lobes; cb, cerebellum; m, medulla.
Fig. 26.—Side view of the brain of a human fœtus of about three months. Representing the marsupial phase in the ontogeny of the human brain. cr, cerebrum; ol, optic lobes; cb, cerebellum; m, medulla. The thalami are completely, and the optic lobes partially, covered by the greatly enlarged cerebrum.
As development proceeds the most conspicuous growth of the brain is observed in connection with the cerebrum and cerebellum. The cerebrum particularly grows relatively and actually larger and larger, but does not yet cover any portion of the optic lobes. This is the reptile stage, represented in Fig. 25. The cerebrum, continuing to grow, finally covers the front portion of the optic lobes. This is the marsupial stage, and is shown in Figs. 26 and 27. Growing further, it soon covers a greater or less portion of the cerebellum. These are the prosimian (Lemur) and simian stages. Finally it grows so far backward as to completely cover the cerebellum, and so far forward as to project much[205] beyond the olfactory lobes. This is the human stage (Plate XII).
Fig. 27.—Dorsal view of the brain (and spinal cord) of a human fœtus of about three months. Representing the marsupial phase of development. cr, cerebrum; ol, optic lobes; cb, cerebellum; m, medulla; bs, brachial enlargement of the spinal cord; ls, lumbar enlargement. The thalami are entirely covered and hidden from view by the cerebrum.
In the study of the phylogeny of the brain we found that the cerebrum in fish, reptile, and lower marsupial is smooth. In the primitive primates (Lemuroidea) it is convoluted; in the simiidæ it is still more convoluted, while in man it reaches the climax of complexity in the size, number, and sinuosity of its convolutions. The object of these convolutions is to increase the surface of the cortex of the brain, the cortex being the seat of psychic[206] phenomena. Other things being equal, the greater the amount of cortex the greater is the intelligence. During its embryonic development the human cerebrum passes also through the stage of smoothness to a convoluted condition; then through stages of increasing complexity of convolutions. Simultaneously with this advance of cerebral organization, there is an unfolding of increasing intelligence.
The cerebellum presides over the co-ordination of the muscular movements of the body. It also, like the cerebrum, passes through the fish, reptile, marsupial, lemur, and simian phases. At first it consists only of the median lobe; then the lateral lobes appear, at first small in size, but getting larger and larger until they greatly surpass in bulk the more primitive median portion. At first the cerebellum is smooth, but as it develops, its fissures become greater and greater, thus increasing its cortex, which presides over the muscular movements. With the developing cerebellum are associated increasing powers of muscular co-ordination; increasing delicacy and complexity of muscular movements. Thus the ontogeny of the brain recapitulates its phylogeny.
The bearing of the theory of evolution on ideas of creation, design, and kindred subjects may briefly be referred to in connection with our views about the relation of psychic phenomena to the brain.
The study of the human brain in its anatomical, physiological and psychological aspects has brought great thinkers, in all ages, into the presence of phenomena that still baffle some of the most subtle[207] philosophers. Here we meet with such realities as self-consciousness, perception, intellection and volition. Are these material entities of such character that we may say they are exclusively products of the activity of the brain, as the secretion of bile is the product of the activity of the liver, as Cabanis taught? To us it seems clear that such is not the case. One cannot take the specific gravity of love or hate, of fear or joy, as one can that of bile; one cannot find a single physical characteristic in any psychic phenomenon. The most physical of all mental processes, viz., perception, has its psychological as well as its physiological phases. The instreaming, through the senses, of impressions from the external world, may be traced by the physiologist along the different nerves of the body to the cortex cells of the brain. All the phenomena that occur at and between these cortex cells and the peripheral endings of the nerves may be formulated in terms of molecular physics. But not so with that consciousness of these impressions which we call perception. In the light of the present knowledge that we possess, it seems to us that the only induction which the physiologist is warranted in making is that, associated with molecular movements in the brain is the phenomenon of perception. This leaves the field clear for each thinker to speculate about the subject in such manner as seems to him most rational. And the history of philosophy shows that many thinkers have formulated theories upon the subject that range in character from the materialism of Büchner to the idealism of Berkeley.
The view which teaches that psychic phenomena are correlated with the physiological phenomena of[208] the brain; that these phenomena have undergone parallel[17] evolution, and “are as inseparable as are the two sides of a sheet of paper” (Dr. Carus), appeals to us as the most comprehensible one and at the same time the one most in consonance with the known phenomena. We accept the view, then, that there is a mind immanent in the brain.[18] The mind is conscious of its personality; conscious of the[209] external world through the innumerable perceptions which reach it through the nervous system; conscious of its power to build its percepts into concepts, and to reason about them; conscious of its power of choice and of causing motion; and conscious of itself, therefore, as a cause in producing effects; and, finally, it is conscious of its power to adapt means to an end,—in short, it knows that it has the power to design.
These facts are at the bottom of much of the philosophy of the present and the past. The untutored savage, knowing that his personality can cause motion, and beholding moving objects in nature, instinctively made the induction that all these objects had personalities behind them. He saw a spirit in his own voice that came back to him as an echo from the rocks; he saw a personality in his shadow; he saw personalities in falling stones, in running brooks, in waving foliage; he beheld them in the raging tempest, in the thunder and the lightning, as well as in the blazing sun and the twinkling stars; he saw spirits in the dead that came back to him in dreams. In short, he recognized a separate personality in every isolated phenomenon in nature. The child talking to its doll, petting it, rebuking it, or whipping it; Xerxes castigating the ocean for wrecking his ships, are illustrations of the strong human tendency to project (or eject)[19] personalities into the inanimate objects of nature. This natural, but lowly, phase of culture and philosophy is known as Fetichism.
As encephalic and psychic evolution advanced; as men, with wider knowledge and broadening experience,[210] ascertained the laws that govern the isolated phenomena of nature, the separate beings in every distinct object and occurrence vanished from thought; but they still beheld a separate personality in every great department of nature. The Romans, for instance, saw Neptune as God of the Ocean, Pluto as God of the lower world, Jupiter as God of the Heavens, and so on. This phase of culture and philosophy, and therefore of religion, is Polytheism.
In the two phases of culture now briefly outlined the personalities were grossly anthropomorphic. They were like human beings, capricious, revengeful, subject to flattery, good and evil, and were therefore to be placated and cajoled by sacrifices and offerings.
Psychic evolution continuing, there appeared from time to time great thinkers who saw one “Infinite Personality” behind the cosmos.[20] This “personality” is still in every phenomenon, though no longer as a separate soul, but only as the separate manifestation of the Soul of the Universe. This is Monotheism, a phase of culture which marks the culmination of philosophy and religion through psychic evolution.
Our knowledge of the universe can be only a shadowy symbol of the reality. The poverty of language is so great and the power of thought so limited that the most subtle philosopher can form only an empty symbol of the cosmic soul. The most ethereal symbols of the greatest thinkers are[211] necessarily incomplete in detail and anthropomorphic, in order to be intelligible. The history of philosophy and religion shows that with the evolution of mind and the acquisition of knowledge, the anthropomorphic ideas of the soul of the cosmos become less crudely coarse and vulgar, until the most elevated and refined ideas of monotheism are reached. But even these refined ideas about the soul of the universe are necessarily anthropomorphic, though in a vastly less degree than in the lower phases of culture. One’s conceptions of this all-pervading soul immanent in the universe are therefore profoundly modified by one’s kind and degree of culture. In the words of Professor Fiske, the great scholar and subtle thinker who has delved in the deepest mines of philosophy and come forth weary and heavy-laden with their boasted treasures, has framed a very different conception of God from that entertained by the priest at the confessional.
A study of the human brain, then, and the soul resident therein prepares us to believe that the cosmos has a soul (God) immanent in it. We can readily grasp the idea that the soul of the cosmos may be self-conscious, wills, thinks, acts, and designs.
This cosmic soul has been and is active in creation. In a low phase of culture every distinct object of nature is looked upon as a separate creation—a manufacture. With the progress of science the conception of separate creative acts becomes greatly modified. The creative acts are judged to be fewer in number and nobler in character. Finally, that phase of highest culture which recognizes the law of universal evolution, formulates the[212] view of one continuous creative act, in which every object is still a creation but not a separate creation,—only a separate manifestation of one eternal act of creative energy. The history of creation, which means the same thing as the history of evolution, shows innumerable adaptations which may surely be considered as the work of a cosmos designer. Evolution has profoundly modified our conceptions of design in nature as it has those of creation. Every separate work of nature, presents a separate, distinct and man-like design to the uncultured. But, with advancing science, all these separate and petty designs are swallowed up into fewer and grander designs, until at last, through evolution, we reach the magnificent and ennobling conception of one infinite and all-embracing design, persisting through infinite time and extending throughout infinite space, which embraces every apparently separate design.
Thus, while evolution destroys low anthropomorphic notions of the mode of working of the Designer and simplifies while it purifies and vastly ennobles our conceptions of this Designer, it yet replaces as much teleology as it destroys. But the highest conceptions that the subtlest philosophers are able to form of a cosmic Designer are necessarily anthropomorphic in some degree, for they can only think in man-like ways.
We have seen in earlier pages of this book how, throughout the incalculable ages of geologic time, innumerable living forms have come upon the stage at different epochs, the forms of one epoch being transmutations from those of an earlier one, and so on, back to the beginning of life. The theory of[213] universal evolution teaches that in the abysmal depths of still earlier æons there was a time when no life existed on the globe, for the globe was then a whirling ball of intensely hot vapor; still further back there was a time when this ball of vapor had not yet been born from that giant nebula—the primitive sun. Through all the sweep of infinite time we see the multitudinous objects of nature coming into existence, one after another, from primeval vapor, and in accordance with laws the character and scope of which we begin to partially understand. It is after the recounting of such well-known facts as these that Professor Fiske makes the statement that Paley’s simile of the watch is no longer applicable to such a world as ours. It must be replaced, he says, by the simile of the flower; for the universe is not a machine, but an organism with an indwelling principle of life; the world was not made offhand, it has grown from more primitive conditions.
In studying the Diagram of Development (Fig. 18) it will be observed that man is the highest and greatest fruitage of the tree of animal life. He is the highest animal in the taxonomic series, as he is in the phylogenic series. He has been the goal, and is the completion of organic evolution. As Dana says, “there is a prophecy of man which runs through the whole of geologic history, which was uttered by the winds and waves at their work over the sands, by the rocks in each movement of the earth’s crust, and by every living thing in the long succession, until man appeared to make the mysterious announcements intelligible.”
The vital path from primitive protozoan to man[214] has been a straight and narrow one, and innumerable groups of animals have branched off laterally. In so doing they departed forever from the man-ward path, and developed obliquely along the diverging roads and bypaths of lower life organizations. They may diverge still farther from the original parting point, but can never get back into the man-ward road. They have lost the golden opportunity and can never regain it.
Man is not only the highest creature that has ever appeared on the globe, but it seems a safe induction to say that he is also the highest animal that evolution will ever develop here.[21] Evolution, through Natural Selection and other agencies, having spent most of its force in creating the innumerable species of animals and plants that have lived in the past and that are now living on the globe to-day, and having had as its goal the creation of that highest and noblest of all creatures—man—is now concentrating its force in further evolving man. Anatomists have reasons to believe that man is now evolving, in many portions of his body, as rapidly as did the horse through Tertiary Ages. Evolution is pushing him on to higher and higher planes, along the straight and perpendicular man-ward track that he has traveled from his protozoan ancestors; while his simian relatives are diverging obliquely more and more from the man-ward track. Through Natural Selection and rational selection evolution seems now to be spending its main force especially on one particular part of man’s body, viz.: his brain and its immanent mind.
The brain of a living, highly civilized man is[215] larger than the brains of men of the tenth century; the brains of these latter are larger than those of palæolithic men. Evolution, having raised the body of man to nearly its highest possible level, is now perfecting more and more his brain, and therefore his thinking power, or, better, his mind. Through his intelligence he is eliminating more and more the noxious plants and dangerous animals that surround him, and is preserving and improving those that are useful to him, and thereby making the organic world more and more subservient to his purposes. He is even getting larger and larger control over the mechanical, physical, and chemical forces of nature, and the possibilities of his improvement in these directions are almost boundless. Evolution for man now means psychic evolution, social evolution; in short, civilization.
From what has been said it can readily be perceived that man, zoölogically and psychologically, is by far the most important creature on the globe. He seems to be the goal towards which evolution has been steadily advancing throughout the geologic ages. It is for these reasons that we believe no higher animal than man will be evolved on the earth. Man himself will continue to evolve higher and higher. Well may we say, with Sir William Hamilton, that there is nothing great in the world but man, and nothing great in man but mind. Is it a shallow philosophy which teaches that it was through design that this most important creature was evolved as the topmost flower on the highest and straightest branch of the tree of life? We do not think so. One of the most profoundly interesting facts to be observed in that higher evolution—psychic[216] evolution—which is now mostly molding man, is the fact of rational selection supplementing and largely replacing Natural Selection. With the creation of man, choice or will comes in as a factor of ever-increasing importance. The active will to use certain capacities and disuse others will play a part in the further development of man of ever-increasing importance and widening influence. Use and disuse have been factors of commanding importance in modifying the bodies and minds of the animal forms that led up to man. Use and disuse will be factors of commanding influence in profoundly modifying the brain, and therefore the mental constitution of man as he advances in social evolution. The use of the brain along chosen lines will, on well-known physiological principles, increase its organization and therefore its power for manifesting psychic phenomena. These two factors will continue to act and react in the future as they have in the past, increased psychical activity enlarging the brain, and the more highly organized brain augmenting the psychic phenomena. What is true of the mind in general is also true of its varied manifestations. The history of psychic evolution gives reason to believe that not only will the capacity for thought be augmented and the power of the will increased in future, but also the strength of selfishness will still further be weakened by disuse and the power of sympathy augmented by practice. As our half-human ancestors were evolving man-ward, and Natural Selection was augmenting their brains, thus increasing their capacity for thought and, therefore, their capacities for more varied experiences through life, there was a concomitant[217] increase in the period of infancy. The activities of the lower animals are mostly of a simple character. They are for the purpose of securing food, escaping enemies, and reproducing their kind. These activities are comparatively so simple and have been repeated so often, generation after generation for ages, that they have become thoroughly organized, by heredity, in the offspring before they are born. When the offspring are born they seek their food, they endeavor to avoid enemies, and in due time procreate their kind without any teaching. With them heredity is almost everything, and experience exceedingly small. These facts can well be exemplified in studying the young of such animals as fishes, amphibians, and reptiles. In the higher birds and mammals Natural Selection has so augmented the size of the brain that their psychic capacities are greatly increased. This increased intelligence is accompanied with an augmented variety and complication of experiences. The acts performed by animals now become so complex, numerous, and varied that they are repeated with much less frequency than are the acts of animals lower in the scale. Consequently, heredity has not had sufficient time to so mold them into the germ-cells that they unfold as perfect reflex or instinctive acts at birth. The hereditary units that carry these acquired experiences of the parents in the developing embryo lie dormant for a while and unfold slowly under the teaching and protection of the parents for a varying period known as infancy. As Natural Selection still further evolved the brains of our advancing half-human ancestors, thus increasing their intelligence and making their[218] lives more replete with complex and varied experiences, there was a concomitant prolongation of the period of infancy—the period of helplessness and dependency. During this evolution of infancy Natural Selection compelled the parents, especially the mother, to possess feelings other than those of utter selfishness. They had to give thought not only to themselves but also to the helpless creatures they brought into the world. The offspring increasing in numbers, all associated together in varying degrees of helpless infancy and dependent upon the care and protection of common parents, the relationships of mother and father, brother and sister, must by degrees have become more and more intimate as evolution proceeded, until finally that social unit appears—the family. In the family personal selfishness can no longer be the exclusively dominant motive to action. Rudimentary sympathies appear. The individuals must conduct themselves so as not to jeopardize the interests of the family. Thus other interests than those of a purely personal character must influence their actions. And thus, finally, the adumbrations of right and wrong conduct appear, and we now find in the newly-created human species the germs of morality and conscience. As social evolution proceeded, the self-regarding faculties were more and more curtailed, and the other-regarding sentiments were extended with ever-enlarging amplitude. Sympathy and helpfulness for others were broadened more and more, including first the clan, then the tribe, then the nation, and, finally, groups of the latter were welded into empires. And the writing on the wall seems to indicate the future federation of all the nations.
Among primeval men, who obtained their food by hunting out such edible objects as were already in existence, war was universal. The supply of fruit, fish, and game being strictly limited, men were compelled to fight under penalty of starvation. As intelligence advanced and men learned to cultivate useful plants and to domesticate animals, and as they learned further to exchange by barter the products of their labor, a much greater population could live upon a given area. These tribes would be more powerful than their neighbors who still lived by hunting, fishing, and such like, and would flourish at their expense. Through agriculture and commerce men slowly learned that one man’s interest was not necessarily opposed to another’s; they also learned, though it may be ever so feebly, that fighting and plundering one another hindered rather than promoted their welfare. Thus man slowly evolved from a primitive, predatory civilization, in which war was universal and chronic, to the higher industrial civilization, in which war is much less frequent and less universal. Out of this primitive industrial civilization, which has grown more and more complex with the passing years, have come the arts and sciences, which give such added interest and value to modern life. This evolving industrial civilization, by furnishing a wider basis for political union through community of interest instead of mere blood-relationship, has greatly extended the field over which moral obligations are recognized as binding.[22] Social evolution is tending to eradicate more and more, through disuse, the brutish instincts of man; weakening his fighting[220] propensities, his cruelty, his selfishness, his passions; and strengthening, by use, his sympathies, his kindness, his mercy, his sense of justice and honor, and his charity. The goal of social evolution seems to be men of character,—men with the widest possible knowledge of the laws of nature, physical, intellectual, and moral; and with the desire and will to rightly obey these laws. Such men will be both loving and lovable characters. In view of this may we not supplement Sir William Hamilton’s aphorism, and say that there is nothing great in mind but character? Since evolution is producing such characters, though it may be seemingly ever so slowly, is it again a shallow philosophy which teaches that there is a designer unfolding these characters? We do not think so. And if there is a designer who has been making towards this goal throughout the infinite sweep of bygone ages, do we not have at least some faint adumbration of knowledge as to the character of this designer? It seems to us that we do. Well may we say, with Matthew Arnold, that there is immanent in the cosmos an eternal soul, not ourselves, that makes for righteousness. This double assertion, that there is a soul in the universe outside of ourselves, and that this soul makes for right conduct, is the basis of fundamental importance in all religions. There are many religions in the world, and many creeds of the one great religion of christendom. They differ in many of the transcendental doctrines that they teach, and in many of the rules of conduct that they prescribe for their adherents; but they all contain as their most fundamental and vitally important basis the double assertion that there is a soul of the universe, and[221] that this soul makes for right conduct. The assertion may be thickly overlaid with superstitions and petty rites by the untrained and dull intelligence of low races, as in the Eskimos; or it may attain a high degree of development and perfection, as among the Jews. The refinement and beauty of the double conception are more and more enhanced with social evolution. Just in proportion as civilization advances, and men come to reason more carefully and entertain wider views of life, just to that extent do they come to value more highly the essential truths of religion, while they attach less and less importance to many superficial details. It is of vastly greater moment to us that there is a cosmic soul in the universe that makes for righteousness than that this soul is threefold or onefold in its transcendental nature. Also of vastly more moment to us is a belief in this soul than any opinions we may entertain about eating meat on Friday or listening to attractive music on Sunday. A thoughtful mind, penetrated with the conviction of the truth of evolution, entertains views on all subjects pertaining to man, very different from those held by one not familiar with the great theory. His conceptions of the first Adam are profoundly modified by a flood of facts. If this flood sweep him on irresistibly, and equally profoundly modifies his conceptions of the second Adam, can it not be seen that even this is a fact of small significance compared with that other fact of overwhelming importance, viz.: the fact of the existence in the universe of a cosmic soul that makes for righteousness? Man is essentially a religious animal, and there is a very[222] substantial philosophical basis for his religion.[23] His religion may be highly colored with emotion, or it may be coldly philosophical. When Herbert Spencer speaks of the eternal Power in the universe which makes for righteousness and is manifested in every event of the universe as the Unknowable, does he not do what Holy Writ has already done? “Canst thou find out the Almighty unto perfection?” When Carlyle speaks of the Universe as in very truth the star-domed city of God, and reminds us that through every crystal and through every grass blade, but most through every living soul, the glory of a present God still beams, he means much the same thing that Mr. Spencer does when he speaks of a Power that is inscrutable in itself, yet is revealed from moment to moment in every throb of the mighty rhythmic life of the universe. The only difference is that Mr. Spencer speaks in the colorless, precise, and formal language of science, while Carlyle’s language is colored by emotion; is, in fact, poetical.[24]
The relation of evolution to many social problems of vital importance is a fascinating as well as very extensive subject. We have only space to say that in order to understand the normal actions, as well as the abnormal ones, of the members of society, and in order, therefore, to understand and inaugurate rational methods of conducting education, minimizing pauperism, vice, disease, and crime, it must[223] constantly be borne in mind that two great streams of tendencies have come down from the ages in the germ cells—what we may call the diseased and animal tendencies on the one hand, and the distinctively human and healthy tendencies on the other.
The most characteristic of the human tendencies are abstract thought and reflection, and therefore the power of choice or will, and altruism.
Also it must be borne in mind that environment is a force of commanding influence. This environment (which the individual may make for himself to a limited extent) may be propitious or adverse to the best human and normal tendencies. The relative preponderance of the animal or the human, the healthy, or the diseased tendencies, taken in conjunction with the character of the environment, stamp man’s actions as normal (and therefore right or wrong) or as abnormal, and therefore irresponsible. Not to discriminate between such normal and abnormal persons is not in accordance with either common morality or common sense. Neither is it in accord with common sense, or morality, or humanity, for society to deal with its habitual criminals and paupers, and subjects of hereditary disease, in the utterly irrational manner that it does. When society takes away from the criminal his personal liberty and places him in an environment that theoretically reforms him and protects itself, why does it not take cognizance of the fact that its theories are often woful failures in practice? The criminal is often not reformed and he gets into the category of habitual offenders; but society permits him, during his intervals of freedom, to procreate[224] his kind and send his polluted cargoes of vicious heritages to helpless offspring. Is this humanity to these offspring? It is the grossest inhumanity! Does society protect itself by its intermittent detentions of habitual criminals? It probably breeds three habitual criminals while it is failing in its efforts to reform one. It is mostly by Nature’s prematurely killing off incorrigible criminals by their diseases and intemperance, that these social pests are kept within due bounds, and not through reformations accomplished in improperly conducted prisons. It seems to us that every consideration of justice and humanity cries aloud for the destruction of the procreating glands in habitual criminals.[25] Castration should go hand in hand with detention behind prison bars. Why should the habitual drunkard, for instance, be permitted to evolve his poisoned germ cells into helpless beings, giving them diseased bodies and vitiated moral characters, thus foredooming them to life-long physical ailments and moral turpitude? Removal of the procreating glands should be the penalty for chronic alcoholism. In objection to this suggestion, some may prate of personal liberty. What a multitude of outrages and brutalities the broad mantle of personal liberty is often made to cover! In allowing personal liberty to an undeserving individual, which more often means unbridled license to that individual, a whole generation of offspring are frequently enslaved by poverty, vice, crime, and[225] disease in its manifold manifestations. During organic evolution Natural Selection has been incessantly on the watch for weaknesses of any kind, ruthlessly exterminating the helpless, the weak, the sick, and those that in any way are unfit. In social evolution Natural Selection has often been of necessity no less ruthless. But during social evolution characters that are unfolding more and more loving and lovable traits have so largely subordinated Natural Selection as to permit the helpless, the old, the sick, and the unfit, to live, thus strengthening those highest attributes of the greatest minds, viz.: intelligent sympathy, pity and love.
But it seems to us that the highest altruism, in dealing kindly with an abnormal, possible parent, will not continue long to stupidly overlook the weighty rights of unborn children. Human selection of the socially unfit will be dominated more and more, as social evolution unfolds its fruits, by those minds that are advancing to the highest goals of evolution, viz.: large minds of high character—widely informed minds, of strong will and broad sympathies. And under these circumstances we may hope that unborn generations will not be given over to total oblivion.
Well may we repeat, before concluding this little book, that man is not only a creature of the present, but profoundly a product of the abysmal ages of a bygone eternity. He is not only a composite chip of many old human blocks, but of innumerable geologic ancestral blocks. He has in his constitution simian, reptilian, piscine, and innumerable other chips, so to speak, and is indeed of the earth earthy; for studies in heredity not only illustrate[226] the continuity of the human race, but also clearly indicate the continuity of this race with more lowly animals. Man has in his structure the indelible impress of the handiwork of these lowly relatives. Upon him, as upon them, and upon all living creatures, the forces of heredity and variation, of use and disuse, of environment and Natural Selection, have been and are perpetually playing, evolving him in one direction and innumerable creatures in other directions.
The goal of evolution seems to be men with Great Minds of High Character. There is nothing great in the world but man, nothing great in man but mind, and nothing great in mind but character.
In order that the reader may appreciate to some extent the relative positions of the different groups of animals, extinct and living, in the scale of life, the following brief classification may be useful. This synopsis is especially intended to help readers who are not familiar with elementary zoölogy to understand the significance of the different types of animals found in the strata of different geologic periods, and more particularly to grasp the meaning of the Diagram of Development. The animals are usually mentioned in the order of their position in the scale of life, commencing with the lowest and simplest.
A. Protozoa. Unicellular animals, dissociated, or associated in simple, loose colonies of similar organisms. Many have an exoskeleton of lime, flint, or other material. Reproduction by fission and by temporary or permanent conjugation.
B. Metazoa. Multicellular animals, consisting of a large number of cells associated together to form single, complex individuals. The cells of an individual are usually differentiated into several[230] kinds performing special functions. Reproduction in the higher forms is sexual; in the lower forms it is often by budding as well as sexual.
Branch I.—Protozoa. Characteristics already defined.
Branch II.—Porifera. Sponges are animals with their cells arranged in two well-defined layers, one of which is internal (endoderm) and the other external (ectoderm). There is a middle stratum of cells (mesoglœa) which does not attain to the definiteness of a mesoderm. Sponges do not possess a body cavity (cœlome); they possess, essentially, a bilateral symmetry. The body varies greatly in shape, and is traversed by canals having large openings (oscula) and numerous small openings (pores) on the surface. The ectoderm is continued through the pores into afferent canals; the endoderm cells line most of the internal (efferent) cavities, and are mostly flagellate; the mesoglœa contains a skeleton of flint, lime, or spongin. Reproduction may be by budding or sexual (either hermaphroditic or unisexual).
Branch III.—Cœlenterata. Animals possessing[231] radial symmetry. There is no body cavity (cœlome), but there is a primitive digestive cavity (enteron). The body consists of two layers, an ectoderm and endoderm; between these two layers there is a mesoglœa of jelly-like consistency; in the simplest cases there are no cells in the mesoglœa, but secondarily endodermal cells may migrate into it. Stinging cells are generally present.
There are two divergent types of structure. The more primitive one is the sessile tubular hydroid, which may be compared to a gastrula furnished at one end with a crown of tentacles surrounding the opening of the enteron, and fixed at the opposite end. The less primitive, derived form, is the active jellyfish (medusoid) type. The hydroid type often constructs a calcareous skeleton known as coral. One life history may present both types (alternation of generations). Reproduction is sexual or by budding, in the latter case often resulting in the formation of colonies.
Branch IV.—Echinodermata. Animals, the larvæ of which possess bilateral symmetry, while the adults have radial symmetry. Even the adults exhibit to a varying extent a tendency to bilateral symmetry. Lime is always deposited in the mesodermic substance (mesenchyme). From the primitive digestive canals of the larvæ pouches grow out to form the usually spacious body cavity and the characteristic water vascular system. The latter and the nervous system exhibit generally[232] a typical five-rayed arrangement. During development there is a distinction between mesoblast derived from gut pouches and mesenchyme produced from migrated amœboid cells. Complicated metamorphosis.
Branch V.—Vermes. The term “worms” includes a “heterogeneous mob,” a collection of classes whose relationships are poorly understood. But they are of great zoölogical interest, for, amid the diversity, there can be discerned affinities with Cœlenterata, Echinodermata, Arthropoda, Mollusca, and Vertebrata. They possess a well-defined mesoderm. They possess bilateral symmetry and have, as a rule, a head, tail, dorsal and ventral surfaces, and right and left sides. The lower worms are unsegmented. In the higher ones the digestive tract extends from the head to the end of the body; a dorsal vessel is usually present above the digestive tract; the nervous system consists of a supra-œsophageal ganglion (brain) and a simple or more commonly double, ventral, ganglionated cord; there is a body cavity (cœlome) lined with mesoderm; true jointed appendages are never present.
Branch VI.—Arthropoda. Animals with bilateral symmetry. Bodies segmented, but not uniformly so. Several or all of the segments bear paired jointed appendages variously modified. Chitinous exoskeleton. Digestive canal beneath the heart. Supra-œsophagel ganglion (“brain”) connected by a nerve ring round the gullet with a double chain of ventral ganglia. Cœlome small in the adult. Sexes almost always separate. Often some metamorphosis.
Branch VII.—Mollusca. Unsegmented animals, possessing, fundamentally, a bilateral symmetry. A very characteristic structure is the “foot”—a muscular protrusion of the ventral surface. Typically, a projecting fold from the dorsal surface of the body forms a mantle, or pallium; the mantle often secretes a single or double shell covering the viscera. The mantle and shell may both be absent. The central nervous system consists of paired ganglia with connecting commissures, viz.: cerebral, pleural, pedal, and visceral ganglia. Heart possesses two auricles and one ventricle. Respiration generally by gills. Frequently there are two larval stages. Development is often direct.
Branch VIII.—Vertebrata or Chordata. Vertebrates are animals having a distinct body cavity (cœlome) and a segmental arrangement of parts. A hypoblastic skeletal notochord is always present in the embryo, but tends to be replaced by an axial skeleton of mesoblastic origin (backbone). Gill slits are always present in the embryo and may or may not persist in adult life. Gill-lamellæ do not occur above Amphibians. Heart is ventral. Central nervous system is dorsal. Eye begins to develop as an outgrowth from the brain.
Class 1.—Tunicata. The tunicates are remarkable animals, and seem to stumble on the border line between Invertebrates and Vertebrates. Their vertebrate characteristics—gill slits, notochord, dorsal nervous system, and brain eye—are generally discernible only in the free-swimming larval state. They generally degenerate as they progress towards the adult condition, and diverge greatly from the vertebrate type. They are mostly stationary.[235] They are multicellular animals possessing bilateral symmetry. The body is enveloped by a thickened cuticle containing cellulose. The pharynx, perforated with the gill slits, is surrounded by a peribranchial chamber (atrium) which communicates with the exterior by an atrial opening. Heart is simple and tubular. The nervous system is generally reduced to a single ganglion. Nephridia are absent. Hermaphrodites. Usually a metamorphosis.
Class 2.—Leptocardii. Simple, worm-like vertebrate animals represented by a single distinct type—amphioxus or lancelet. The central nervous system consists of a spinal cord, and a very ill-defined, rudimentary brain. No skeleton other than an unsegmented and persistent notochord which projects beyond the anterior end of the nerve cord. No scales. Gill slits are very numerous in the adult. Amphioxus is widely removed from the fishes by the absence of skull, jaws, definite brain, sympathetic nervous system, ear, eye, genital ducts, spleen, and definite heart, the latter being simple and tubular. No pectoral or pelvic limbs (fins). Blood colorless; gastrula ciliated and free-swimming. Metamorphosis.
Class 3.—Marsipobranchii. Worm-like vertebrates having round mouths without distinctly developed jaws; no jaw-bones. Without scales and without paired limbs (fins). Six or seven gill pouches. Skeleton consists of persistent, unconstricted, cartilaginous notochord. No sympathetic nervous system. Single nasal sac. No conus arteriosus, no spleen or pancreas. Undivided segmental duct.
Class 4.—Pisces.[26] Aquatic, cold-blooded vertebrates, with a movable lower jaw. Cartilaginous or osseous skeleton, with paired pectoral and pelvic fins, supported by fin rays (radials diverging from several basal pieces); also unpaired fins. No sternum. Exoskeleton of bony plates or scales. Skull has usually one occipital condyle. Breathing by permanent gills attached to gristly or bony arches on the sides of the gill clefts. In most fishes the digestive cavity gives origin to an air bladder which may or may not remain permanently connected with the digestive cavity. It mostly serves a hydrostatic purpose, but in some of the more primitive fishes (Amia, Lepidosteus) it is occasionally of some slight use in respiration. Heart consists of single auricle and single ventricle, and contains only venous blood; there is a sinus venosus and often a conus arteriosus. No inferior vena cava. No allantois. Most fishes lay eggs which are fertilized in the water.
Class 5.—Amphibia. In the evolution of vertebrates the Amphibians represent those forms which made the transition from aquatic to terrestrial life, but have lingered near the water. Certain acquisitions gained by the Dipnoi, such as a three-chambered heart and lungs, have been accentuated and firmly established by the Amphibians.
Amphibia are cold-blooded vertebrates whose larval forms always have gill arches bearing gills. In some forms the gills are retained throughout life, though the adults always possess functional lungs. Nasal sacs open posteriorly into the mouth. In existing forms there is rarely any exoskeleton. There are two occipital condyles. Unpaired fins are frequently present, though they possess no fin rays. When limbs are present they possess distinct digits and conform to the same type as those of the higher vertebrates. Heart has two auricles and one ventricle. There is an inferior vena cava. The intestine ends in a cloacal chamber, as do also the urinogenital ducts. Eggs almost always laid in the water. Often a marked metamorphosis.
Class 6.—Reptilia. Cold-blooded, air-breathing vertebrates. Limbs usually ending in claws; limbs sometimes absent; exoskeleton of scales; ribs well developed; incomplete double circulation; heart is four-chambered in the highest forms; oviparous; no metamorphosis; embryo with an amnion and allantois.
The following are some of the orders that have become extinct, viz.:
The following are the living orders, viz.: a, Ophidia; b, Lacertilia; c, Chelonia; d, Crocodilia.
Class 7.—Aves. Feathered vertebrata; heart four-chambered; warm-blooded; lungs with accessory air-sacs; bones dense and hollow; jaws encased in horny beaks in modern forms; the fore-limbs form wings; oviparous; eggs are very large and covered by calcareous shell.
Class 8.—Mammalia. Animals having the body covered with hair; warm-blooded; young nourished with milk secreted from mammary glands; lower jaw articulating directly with the skull, the quadrate becoming the malleus (ear-bone); heart four-chambered and with the aorta reflected over the left bronchus; red blood-corpuscles non-nucleated; complete diaphragm; brain large, especially the cerebrum and cerebellum; uterine gestation; viviparous.
Primates are an order of Monodelphia or Placentalia, including lemurs, monkeys, and man. They are nearly all adapted to an arboreal life. The hand (manus) and the foot (pes) nearly always have five[240] digits provided with flat nails. The pectoral and pelvic limbs are prehensile, owing to the fact that the thumb (pollex) and big toe (hallux) are more or less completely opposable to the other digits. The orbital fossa is completely surrounded by a bony rim. The femur does not have a third trochanter. The internal condyle of the humerus does not have a foramen above it. The clavicles are always well developed. The testes descend into a scrotum. On the thoracic region there are nearly always two teats. The placenta may be non-deciduate, or deciduate and metadiscoidal.
The order of Primates is subdivided into the following sub-orders, viz.:
Lemuroidea. The Lemurs are small, monkey-like quadrupeds. They are mostly nocturnal, arboreal creatures of comparatively low organization. The body is furry. The orbital and temporal fossæ freely communicate. The lachrymal foramen is situated outside the orbital fossa. The dentition of the Lemurs varies greatly; in some of them it is 2/2 1/1 3/3 3/3 for both jaws. In nearly all of them the median incisors of the upper jaw are separated by a median space. Both feet have flat nails on all the digits, except the second of the hind-foot, which has a claw. The thumb (pollex) and the big toe (hallux) are well developed. The cerebral hemispheres are only slightly convoluted, and do not entirely overlap the cerebellum. The transverse colon is almost always folded on itself. There may be a pair of teats on the abdomen. The uterus is two-horned, and the placenta is diffuse.
This sub-order comprises the Lemurs proper (Tarsius, Lemur, and other genera) and the Aye-Ayes (Chiromys).
Anthropoidea. The Anthropoidea are the most highly organized Primates. They are chiefly modified for an arboreal life. The body is hairy instead of furry, but only slightly so in man. The incisors do not exceed 2/2; the molars are 3/3, except in the marmosets, where they are 2/2. The upper, median incisors are in close contact. The orbital and temporal fossæ are separated by a broad vertical plate. The lachrymal foramen is situated inside the orbital fossa. All of the digits are provided with flat nails, except in the Hapalidæ, in which all except the big toe (hallux) are provided with a claw. In some the thumb (pollex) is rudimentary or absent. The clavicle is well developed. The cerebrum is usually richly convoluted, and more or less completely covers the cerebellum. The uterus has no horns. The placenta is deciduate and metadiscoidal.
The Anthropoidea are subdivided into the two following groups:
The Platyrrhinæ or New-World Monkeys are subdivided into the following families, viz.:
The Catarrhinæ or Old-World Monkeys are subdivided into the following families, viz.:
Hapalidæ. The Hapalidæ or Marmosets are no larger than squirrels, being the smallest monkeys. They are found mostly in Brazil.
There is a broad septum between the nostrils. Their dentition is 2/2 1/1 3/3 2/2, and is distinctive, for the remaining Anthropoidea have 3/3 molars. The tail is long, hairy, and non-prehensile. The arms are not longer than the legs. The thumb (pollex) is long, but is not opposable. All the digits except the great toe (hallux), which is small, have curved, pointed claws. There is no bony external auditory meatus. There are no cheek pouches, or ischial callosities. The Hapalidæ often bear three young ones at a birth, whereas the other Anthropoidea commonly bear but one. There are two genera of Marmosets, viz.: Midas and Hapale.
Cebidæ. The Cebidæ are most at home in Brazil. They have a broad septum between the nostrils. Many of them have prehensile tails. The dental formula is 2/2 1/1 3/3 3/3, and is characteristic. There is no bony external auditory meatus. There are no cheek pouches, or ischial callosities. All the digits are provided with flat nails. The thumb (pollex) is not opposable.
This family includes the following genera, viz.: Tee Tees (Callithrix), Howling Monkeys (Mycetes), Spider Monkeys (Ateles), Squirrel Monkeys (Chrysothrix), and Capuchin Monkeys (Cebus).
Cercopithecidæ. This family includes the Old-World dog-like Apes. They are quadrupeds, and the muzzle or snout is quite dog-like. There is a narrow septum between the nostrils. The dental[243] formula is 2/2 1/1 2/2 3/3. The sternum is narrow. The tail is not prehensile. The cæcum has no appendix vermiformis. There is a bony external auditory meatus. There may or may not be cheek pouches. Over the rough surface of the everted ischia the skin forms callosities (ischial callosities) which are often brightly colored. The thumb (pollex), when present, is opposable. This family includes two genera, Macaques (Macacus) and Baboons (Cynocephalus).
Simiidæ. This family includes the Anthropoid Apes of the Old World. They are less quadrupeds than the former, often walking in a semi-erect position. The dental formula is 2/2 1/1 2/2 3/3. The sternum is broad. The cæcum has an appendix vermiformis. The nasal septum is narrow. The pectoral limbs are much longer than the pelvic ones. There is a bony external auditory meatus. There are no cheek pouches. Only in the Gibbons are there ischial callosities, in whom they are small. The thumb (pollex) is opposable. No visible tail. This family includes the following genera, viz.: Chimpanzees (Anthropopithecus), Gorillas (Gorilla), Orangs (Simia), and the Gibbons (Hylobates).
Hominidæ. This family of Anthropoidea includes only the human species (Homo sapiens). The dental formula is 2/2 1/1 2/2 3/3. The tail is not visible. The Hominidæ differ from the Simiidæ, structurally, mainly in the more perfect assumption of the erect attitude. There is a more complete adaptation of the pelvic limbs to bearing the weight of the body, correlated with compensating alteration in the curvature of the spinal column. The big toe (hallux) is not opposable, and also it is often longer, never shorter than the other toes, and is not[244] abducted from them. He has a far better heel than the Simiidæ. The thumb is far more opposable in man. There is a greater length of the pelvic limbs compared with the pectoral limbs. The canine teeth are much smaller and do not project beyond the level of the others. The brain has a much greater size and complexity. Man has a larger forehead, smaller cheek bones, smaller supraorbital ridges, a less protrusive face, and a true, well-marked chin.
The Simiidæ are subdivided into the two following groups, viz.:
The Metaphyta are subdivided into:
The Cryptogams are subdivided into:
The Phanerogams are subdivided into:
1. Gymnospermæ:
2. Angiospermæ:
The following books have been the constant companions of the author during the preparation of his “First Book in Organic Evolution”:—
A-chro´ma-tin [Gr. a-priv.; chroma, color.] The nuclear hyaloplasm.
A-chro-ma-top´si-a [Gr. a-priv.; chroma, color; opsis, sight.] Color-blindness.
Ac-ti-no-zo´a [Gr. aktis, ray; zoon, animal.] A division of Cœlenterates embracing the sea-anemones and coral-polyps.
Al-lan´to-is [Gr. allas, sausage; eidos, form.] A membranous, sack-like appendage developed from the hinder part of the embryonic alimentary tract and which serves the purpose of effecting oxygenation and other changes in the blood in reptiles, birds, and mammals.
Am-blyp´o-da [Gr. amblys, blunt; pous, foot.] An extinct order of ungulates having elephantine feet and whose navicular bone was supported by the cuboid.
Am-blys´to-ma [Gr. amblys, blunt; stoma, mouth.] A genus of amphibians remarkable for the transformations they undergo. When undeveloped they represent the formerly recognized genus Siredon. Some species are called axolotls.
Am´i-a [Gr. amia, a kind of tunny.]
Am-i-to´sis [Gr. a-priv.; mitos, thread.] Direct cell-division; cell-division without the formation of nuclear figures.
Am´mon-ites [Egyptian Amon.] Fossil cephalopod shells.
Am´nion [Gr. amnion, fœtal membrane.] A membranous sack containing fluid which envelopes the embryo in the classes of reptiles, birds and mammals.
A-mœ´ba pro´te-us [Gr. amoibe, change; Proteus.] A unicellular animal; a Protozoan.
Am-phib´i-a [Gr. amphi, double; bios, life.] A class of vertebrates whose young are fish-like and have gills: the skull has two condyles and a parasphenoid.
Am-phi-mix´is [Gr. amphi, double; mixis, mingling.]
Am-phi-ox´us [Gr. amphi, double; oxys, sharp.] A small worm-like vertebrate whose body tapers at both ends. The[258] skeleton is notochordal; skull, brain, auditory, and renal organs are absent.
An-ab´o-lism [Gr. ana, up; ballo, throw.] Constructive metabolism; the series of chemical changes by which a cell builds up simple and stable food into the highly complex and unstable living material.
An-ap-to-mor´phus [Gr. an-priv.; hapto, fasten; morphe, shape.]
An-gi-o-sper´mæ [Gr. angeion, vessel; sperma, seed.] Plants whose seeds are contained in a closed seed-vessel.
An-nu-la´ta [Lat. annulatus, ringed.] A division of Vermes, including marine worms, leeches, and earthworms.
An-the-rid´i-a [NL., < anthera, anther; Gr. dim. idion.] The male sexual organ in cryptogams answering to the anthers in phanerogams.
An-thro-po-pi-the´cus [Gr. anthropos, man; pithekos, ape.]
An´thro-poid [Gr. anthropos, man; eidos, form.] Manlike: said especially of the gibbons, orangs, gorillas, and chimpanzees.
An-ti-tox´ine [Gr. anti, opposed to; toxikon, poison.]
A-nu´ra [Gr. an, not; oura, tail.] An order of amphibians without a tail in the adult, including the toads and frogs.
A´phis [Gr. apheideis, lavish.] In Entomology a genus typical of Aphididæ. An aphid; a plant-louse.
Ap-pen´dix ver-mi-for´mis [Lat. appendo, to hang from; vermis, worm; forma, form.] A vestigial structure attached to commencement of the large intestine.
A-rach´ni-da [Gr. arachne, spider.] A class of Arthropods embracing the spiders, mites, scorpions, etc.
Ar-cel´la [Lat. dim. arca, a box.] Belonging to a group of protozoans having a chitinous shell.
Ar-chæ´an [Gr. archaios, ancient.] Pertaining to the oldest strata of rocks in geological history.
Ar-chæ-op´ter-yx [Gr. archaios, ancient; pteryx, bird.] A fossil bird with reptilian characteristics.
Ar-che-go-sau´rus [Gr. archegos, beginning; sauros, lizard.] A carboniferous amphibian (labyrinthodont.)
Ar´go-naut [Gr. argo, swift; nautes, sailor.]
Ar-ma-dil´los [Sp. dim. of armado, armed.] Edentate mammals having a carapace formed by ossification of the greater part of the skin and the union of the bony scutes.
Ar-throp´o-da [Gr. arthron, joint; pous, foot.] A branch of animals (invertebrates) with jointed legs, as insects, spiders, and crabs.
As´ter [Gr. aster, star.]
As-ter-oi´de-a [Gr. aster, star; eidos, form.] Echinoderms having radiating arms with ambulacral grooves below, embracing the true star-fishes.
At´a-vism [Lat. atavus, grandfather.] Reversion; recurrence to an ancestral type.
At´e-les [Gr. ateles, imperfect.]
Au´ri-cles [Lat. auricula, little ear.] A chamber of the heart which receives blood from the veins and transmits it to the ventricle.
A´ves [Lat. pl. of avis, bird.] Birds, a class of vertebrata.
Ba-cil´lus [Lat. bacillum, a little stick.] A genus of bacteria consisting of rod-like cells. They multiply by transverse division and by the formation of endogenous spores. Unicellular plants.
Bac-te´ri-a [Gr. bakterion, stick.] Microscopic unicellular plants. A genus of Schizomycetes (fission-fungi).
Bal-a-no-glos´sus [Gr. balanos, acorn; glossa, tongue.] Supposed by some biologists to belong to the Chordata.
Beams [AS. beam, tree.] The main stem of a deer’s antler.
Bel-em´nites [Gr. belemnon, dart.] A belemnitoid cuttlefish.
Bi´o-phores [Gr. bios, life; phero, bear.] Biological units of hereditary mass.
Bi´pes [Lat. bis, twice; pes, foot.]
Blas´tid [Gr. blastos, sprout.]
Blas-to-ge-net´ic [Gr. blastos, a germ; genesis, generation.] Originating in the germ cells.
Blas´tu-la [Gr. blastos, sprout.] A stage in the growth of the embryo; a hollow sphere of one layer of cells enclosing a segmentation-cavity.
Brach-i-op´o-da [Lat. brachium, arm; Gr. pous, foot.] Molluscoid animals with bivalve shells, one of which is dorsal and the other ventral; also with brachial appendages coming from the sides of the mouth. Lamp shells.
Brach´i-o-pod [Vid. Brachiopoda.] Lamp shell.
Bry-oph´y-ta [Gr. bryon, moss; phyton, plant.] A division of the vegetable kingdom consisting of liverworts and mosses.
Bry-o-zo´a [Gr. bryon, moss; zoon, animal.] Moss animals.
Ca-du-ci-bran-chi-a´ta [Lat. caducus, falling; branchia, gills.] Urodeles that lose their gills before maturity.
Ca-du-ci-bran´chi-ates [Vid. Caducibranchiata.]
Cal´li-thrix [Gr. kalos, beautiful; thrix, hair.]
Cal-a-mo-ich´thys [Gr. kalamos, reed; ichthys, fish.]
Ca´lyx [Gr. kalyx, cover.] The outermost series of leaves (sepals) of a flower; usually green.
Cam´bri-an [Cambria, Wales.] The name given by Sedgwick to those Palæozoic rocks that underly the Silurian, in Cambria (or Wales).
Car-bon-if´er-ous [Lat. carbo, coal; fero, bear.]
Car-i-na´tæ [Lat. carinatus, keel-shaped.] Birds having a keeled (carinate) breast-bone.
Car-niv´o-ra [Lat. caro, flesh; voro, devour.] Flesh-eating mammals.
Cas-tra´tion [Lat. castratus.] Removal of the glands that bear the Germ-Cells.
Cat-ar-rhi´næ [Gr. kata, down; rhis, nose.] Old World monkeys. The septum of the nose is narrow; the tail is short and not prehensile.
Cat´ar-rhines [Vid. Catarrhinæ.] Old World monkeys.
Ceb´i-dæ [Gr. kebos, long-tailed monkey.] A family of New World monkeys.
Ce´bus [Gr. kebos, long-tailed monkey.]
Cen-o-zo´ic [Gr. kainos, new; zoe, life.] Relating to the Quaternary and Tertiary eras.
Cen´ti-pede [Lat. centum, hundred; pes, foot.] A many jointed myriapod having a pair of legs to each joint.
Cen´tro-some [Gr. kentron, center; soma, body.] A body just outside of the nucleus of a cell and that governs the mitotic phases of a cell.
Ceph-a-lop´o-da [Gr. kephale, head; pous, foot.] Molluscs with a subcentral head, a beaked mouth, and tentacles taking the place of feet, including cuttlefishes, etc.
Cer´a-tites [Gr. keras, horn; ites, like.] A fossil cephalopod with the habitation chamber short and sutural saddles mostly simple.
Cer´a-to-dus [Gr. keras, horn; odous, tooth.] So-called from the horn-like ridges of the teeth. A lung-fish.
Cer-co-pi-the´ci-dæ [Gr. kerkopithekos, a long-tailed ape; eidos form.] A family of catarrhine anthropoidea.
Cer-e-bel´lum [Lat. dim. of cerebrum, brain.] A subdivision of the encephalon.
Cer´e-brum [Lat. brain.] A subdivision of the encephalon.
Cer´e-bral [Vid. Cerebrum.] Pertaining to the cerebrum.
Ce-ta´ce-a [Lat. cetus, whale.] An order of fish-like mammals including the whales and porpoises.
Chei-rop´te-ra [Gr. cheir, hand; pteron, wing.] Including the Bats,—an order of mammals.
Chei-ro´tes [Gr. cheirotos, having a hand.] A native lizard of Mexico.
Che-lo´ni-a [Gr. chelone, tortoise.] An order of reptiles, including the turtles and tortoises.
Chi-mæ´roids [Gr. chimaira, a fabled monster; eidos, form.] An order of elasmobranchii (holocephali) with opercular membrane and smooth skin.
Chi´ro-mys [Gr. cheir, hand; mus, mouse.]
Chim-pan´zee [W. Afr.] Anthropopithecus. A West African arboreal, anthropoid ape; five feet high; distinct eyelashes, eyebrows, and whiskers; very large ears. It approximates to man in its dentition and the length of its arms. Has darker hair than gorilla.
Chi´tin-ous [Gr. chiton, tunic.] Pertaining to chitin, the horny substance in the skin of insects.
Chro´ma-tin [Gr. croma, color.] Nuclear threads.
Chor-da´ta [Lat. chorda, chord.] A subkingdom of animals with a notochord persistent or evanescent.
Chro´mo-somes [Gr. croma, color: soma, body.] Nuclear threads.
Chry´so-thrix [Gr. chrysos, gold; thrix, hair.]
Clo-a´ca [Lat. cloaca, a common sewer.] The common cavity into which the digestive, urinary, and genital canals empty in most vertebrates below placentals, and in all mammalian embryos.
Coc´ci [Gr. kokkos, berry.] Pl. of coccus. Isolated spherical cells (bacteria).
Co-coons´ [F. cocon, dim. of coque, shell; < Lat. concha, shell.] The protective envelopes spun by certain larval insects, as silkworms, in which they are inclosed in the chrysalis state.
Co-do-si´ga um-bel-la´ta [Gr. kodon, a bell; sige, silence; Lat. umbella, dim. of umbra, shade.]
Cœ-cil´i-a [Lat. cæcus, blind.] Footless amphibians of snake-like form; rudimentary eyes; no neck.
Cœ-len-te-ra´ta [Gr. koilos, hollow; enteron, intestine.]
Cœ-len´ter-ate [Vid. Cœlenterata.]
Cϫlome [Gr. koilos, hollow.] The body-cavity of a metazoan between the viscera and the body-wall.
Combs [AS. camb, crest or ridge.] The fleshy crests on the heads of roosters, etc.
Con-dy-lar´thra [Gr. kondylos, knuckle; arthron, joint.] An Eocene group of ungulates in which the astragalus is not interlocked laterally with the tibia and its head rounded.
Co-nif´e-ræ [Lat. conus, cone; fero, bear.] The pine family of trees.
Co´nus ar-te-ri-o´sus [Latin phrase.] The arterial, cone-shaped portion of the left ventricle of the heart.
Co-rol´la [Lat., dim. of corona, crown.] The inner set of leaves of flowers and usually bright-colored. The individual parts are called petals.
Crests [Lat. crista, tuft.] Projecting natural growths on the tops of animals’ heads and usually ornamental, as the cock’s comb, or the lengthened feathers of a bird.
Cre-o-don´ta [Gr. kreas, flesh; odous, tooth.] A group of fossil animals containing forms ancestrally related to existing Carnivora.
Cre-ta´ceous [Lat. creta, chalk.] Pertaining to the latter part of the Reptilian age.
Cri-noi´de-a [Gr. krinon, lily; eidos, form.] Stone-lilies; stalked star-fishes. A division of Echinoderms.
Cri´noids [Vid. Crinoidea.]
Croc-o-dil´i-a [Lat. crocodilus, lizard.] An order of reptiles including crocodiles, alligators, and gavials. They are the highest order of reptiles. In them the heart and brain approximate very closely to that of birds.
Cros-sop-te-ryg´i-a [Gr. krossoi, tassels; pterygion dim. of pteryx, wing.] A group of fishes with paired, lobate fins, and with an endodermal skeletal, axis fringed with dermal rays.
Crus-ta´ce-a [Lat. crusta, crust.] A division of arthropods containing crabs, lobsters, shrimps, crawfish, barnacles, etc.
Cryp´to-gams [Gr. kryptos, hidden; gamos, marriage.] The lower of the two great subdivisions of the plant kingdom.[263] They have no true flowers containing stamens, pistils, and seeds; they propagate by means of spores.
Cten-oph´o-ra [Gr. kteis, comb; phero, bear.] A subdivision of cœlenterates with paddle-like locomotive plates arranged in eight meridional rows on the outer surface of the body.
Cyc-a-da´ce-æ [Gr. kykos, African coco-palm.] The cycad family of plants (gymnospermous), of fern-like or palm-like aspect.
Cys-toi´de-a [Gr. kystis, bladder; eidos, form.]
Cy-no-ceph´a-lus [Gr. kyon, dog; kephale, head.]
Cy´to-plasm [Gr. kyo, be pregnant; plasma, < plasso, form.] Protoplasm; especially that portion of a cell apart from the nucleus.
Dal´ton-ism [Dalton, chemist, who had color-blindness.] Color-blindness, especially red-blindness.
De-cid´u-ate [Lat. deciduus, falling off.] Shed at periodic times.
Des´mids [Gr. desmos, band.] Unicellular plants of minute size; bright-green in color; mainly solitary, fresh-water algæ.
Dev-o´ni-an [Devonshire, England.] The name given by Murchison to Palæozoic rocks in Devonshire, England.
Di´a-toms [Gr. dia, through; tome, cutting.] Microscopic unicellular algæ inhabiting salt or fresh water. Each individual (frustule) consists of two flint valves which are more or less symmetrical. They are either isolated or attached together in a series.
Di-cot-y-le´don-es [Gr. dikotylos, with two hollows.] The most important and largest class of flowering plants. Characterized by having seeds with two cotyledons, exogenous stems, and leaves that are netted-veined; the parts of the flower mostly in fours or fives.
Di-del´phi-a [Gr. di-, two; delphys, womb.] A sub-class of mammals, as the Marsupials, having a double womb and no placenta.
Dif-flu´gi-a pyri-form´is [Lat. diffluere, to flow apart; pyrum, pear; forma, form.]
Di-no-sau´ri-a [Gr. deinos, terrible; sauros, lizard.] Mesozoic land reptiles.
Dip´no-i [Gr. di-, two; pneo, breathe.] Fishes with regular gills, a double or single lung, and nostrils inside as well as outside the mouth.
Dro-ma-the´ri-um [Gr. dromos, running; therion, small wild beast.]
Dys-chro-ma-top´si-a [Gr. dys, bad; chroma, color; opsis, sight.] Partial color-blindness; difficulty in distinguishing colors.
E-chi-no-der´ma-ta [Gr. echinos, hedgehog; derma, skin.]
E-chi´no-derms [Vid. Echinodermata.]
Ech-i-noi´de-a [Gr. echinos, hedgehog; eidos, form.] Echinoderms including sea urchins.
Ec´to-derm [Gr. ektos, outside; derma, skin.]
Ec´to-plasm [Gr. ektos, outside; plasso, form.]
E-den-ta´ta [Lat. e, out of; dens, tooth.] An order of placentals with a small, one-lobed cerebrum; without median, cutting teeth: including armadillos, ant-eaters, and sloths.
El-as-mo-bran´chi-i [Gr. elasmos, metal plate; branchia, gills.] Fish-like vertebrates which have no membrane bones in the skull or the shoulder-girdle; five pairs of strap-like gills attached by their distal ends; claspers to the ventral fins of males; complicated brain with optic nerves forming a decussation.
Em´bry-o [Gr. en, in; bruein, swell.] The term applied to an animal in the earlier stages of development.
Em-bry-ol´o-gy [Gr. embryon, embryo; logos, < lego, speak.] The study of embryos.
En´do-derm [Gr. endon, within; derma, skin.]
En´do-plasm [Gr. endon, within; plasso, form.]
En-ter-op-neus´ta [Gr. enteron, intestine; pneustos, breathing.] Acorn-tongue worms. Smooth-bodied, footless worms, having a large exserted soft proboscis; breathing by a series of respiratory sacs opening into the digestive canal, and communicating externally by spiracles; nervous system situated above a seeming notochord. Balanoglossus. Included by some biologists among the chordata.
E-o-hip´pus [Gr. eos. dawn; hippos, horse.]
E´o-cene [Gr. eos, dawn; kainos, recent.]
Ep-en-ceph´a-lon [Gr. epi, upon; enkephalos, brain.] A fundamental subdivision of the brain (encephalon).
Ep´i-blast [Gr. epi, upon; blastos, bud.]
E´quus [Lat. horse.]
E-ryth-ro-lam´prus [Gr. erythros, red; lampros, shining.]
Eu-ca-lyp´tus [Gr. eu, good; kalypto, cover.] A large genus of evergreen trees of the myrtle family.
Ev-o-lu´tion [Lat. e, out; volvo, roll.]
Ex-o-skel´e-ton [Gr. exo, outside; skeleton, dried body.] External skeleton; bony or horny hardening of the integument.
Fla-gel´lum [Lat., dim. of flagrum, scourge.] A slender protoplasmic extension of a cell, for purposes of locomotion.
Fo-ram-i-nif´e-ra [Lat. foramen, opening; fero, bear.] A division of protozoans secreting a shell perforated by many minute apertures.
Fos´sil [Lat. fodio, dig.] Any organic body so situated in the earth, and so buried in solid rock or in earthy deposits, as to be capable of indefinite preservation.
Gan´gli-on [Gr. ganglion, tumor.] A swelling that consists of an aggregation of nerve-cells. It receives and discharges nervous impulses and serves to stimulate psychical and organic activities.
Ga-noi´de-i [Gr. ganos, brightness; eidos, appearance.]
Gas-ter-op´o-da [Gr. gaster, stomach; pous, foot.] Including all snails and slugs.
Gas´tru-la [Dim. of Lat. gaster, stomach.]
Gem´mæ [Lat. buds.]
Geph-y-re´a [Gr. gephyra, bridge.] A division of worms with an œsophageal nervous ring and ventral chord; no distinct segments or legs; a terminal or dorsal anus.
Go´ni-a-tites [Gr. gonia, corner; lithos, stone.] A genus of fossil Ammonites.
Gor´gets [Fr. gorgette, dim. of gorge, throat.] Throat-patches distinguished by color or texture, especially in humming birds.
Grap´to-lites [Gr. graptos, written; lithos, stone.] Fossil hydroids.
Greg-a-ri´na [Lat. gregarius, < grex, flock.] A genus typical of Gregarinidæ. Gregarina gigantea is sixteen millimeters in length and is one of the largest unicellular animals known.
Greg-a-rin´i-dæ [Vid. Gregarina.] More or less elongated amœba-like Protozoa, having a well-defined cell-wall, and a “subcuticular” system of muscular fibrillæ; nucleus, but no contractile vacuole; reproduction by encystment and subdivision of the central cell mass or protoplasm, by which[266] shelly psorosperms are formed and from which escape the moner-like young, which undergo a metamorphosis.
Gro´mi-a o-vi-form´is [Lat. gromia; ovum, egg; forma, form.] A characteristic imperforate foraminifer.
Gym-no-phi´o-na [Gr. gymnos, naked; ophis, serpent]
Gym-no-sper´mæ [Gr. gymnos, naked; sperma, seed.] Plants whose seeds are not contained in a closed seed-vessel, as Cycads and Conifers.
Hap´a-le [Gr. hapalos, gentle.]
Ha-pal´i-dæ [Gr. hapalos, gentle] A family of New World monkeys including the Marmosets.
Hel-i-co-nid´æ [N., < heliconius, of Helicon; idæ.] A family of butterflies.
Hes-per-or´nis [Gr. hesperos, western; ornis, bird.] Cretaceous carinate birds with rudimentary wings, short tail, and pointed teeth implanted in grooves.
Hip-po-pot´a-mus [Gr. hippos, horse; potamos, river.]
Hol-o-ceph´a-li [Gr. holos, whole; kepale, head.] A subdivision of Elasmobranchii in which the suspensorium of the lower jaw is continuous with the cranium, as chimæroids, etc.
Hol-o-thu-roi´de-a [Gr. holos, whole; thouros, rushing.] Worm-like echinoderms, with skin-like integuments, and circum-oral tentacles; including sea-cucumbers, sea-slugs, etc.
Ho-min´i-dæ [Lat. homo, man; idæ.] A family of primates restricted to mankind.
Hy´a-lo-plasm [Gr. hyalos, glass; plasma, < plasso, form.]
Hy´dra [Gr. hydra, Lernæan serpent, < hudor, water.] A fresh-water polyp, having the form of a cylindrical tube, surrounded at the mouth with a circle of thread-like tentacles, by which it captures its prey.
Hy´droid [Gr. hudor, water; eidos, form.] Resembling the hydra; pertaining to the Hydroidea.
Hy-dro-zo´a [Hydro-; zoon, animal.] Cœlenterates including Hydroids and jelly-fishes.
Hy-lob-a-ti´næ [Gr. hylobates, one who walks the wood.] An Asiatic subfamily of apes; the long-armed apes or gibbons.
Hy-lob´a-tes [Gr. hylobates.] A gibbon,—an Asiatic ape.
Hy-ra-coi´de-a [Gr. hyrax, shrew-mouse; eidos, appearance.]
Ich-thy-or´nis [Gr. ichthys, fish; ornis, bird.] Cretaceous[267] toothed birds of tern-like form, with socketed acute teeth, and bi-concave vertebræ
Ich-thy-o-sau´ri-a [Gr. ichthys, fish; sauros, lizard.]
Id´i-o-blast [Gr. idios, individual; blastema, sprout.]
In-fu-so´ri-a [Lat. in, into; fundo, pour.] So called because including many animalcules that occur in infusions of decaying substances.
In-sec´ta [Lat. in, into; seco, cut.]
In-sec-tiv´o-ra [Lat. insectum, insect; voro, devour.]
In-ver-te-bra´ta [Lat. in, not; vertebratus, jointed.] Including all animals that do not possess a notochord or backbone: opposed to Vertebrata.
In-ver´te-brates [Vid. Invertebrata.]
Ju-ras´sic [Jura mountains.]
Kaf´ir-boom [Erythrina caffra.] A prickly-stemmed tree of South Africa.
Kal´li-ma para-lek´ta [Gr. kallimos, beautiful; para, along side of; lektos, picked out.]
Keri-vou´la pic´ta.
Lab-y-rin´tho-donts [Gr. labyrinthos, labyrinth; odous, tooth.] Extinct amphibians.
Lac-er-til´i-a [Lat. lacertus, lizard.]
La-ge´na [Gr. lagynos, flask.]
La-mel-li-bran-chi-a´ta [Lat. lamella, a plate; branchia, gills.] Bivalve molluscs.
La-nu´go [Lat. lana, wool.]
Lar´væ [Lat. pl. of larva, mask.] The early forms of animals when they are unlike the parent. In insects the first stages after leaving the eggs, preceding the pupa, as maggots or caterpillars.
Lem-u-roi´de-a [Lemur; Gr. eidos, appearance.] A sub order of Primates. Prosimiæ.
Lep-i-do-si´ren [Gr. lepis, scale; siren, a genus of amphibians.] One of the lung-fishes.
Lep-to-car´di-i [Gr. leptos, fine, small; kardia, heart.]
Liv´er-worts.
Lon-gi-cor´ni-a [Lat. longus, long; cornu, horn.] A division of beetles having very long filiform antennæ.
Ma-caques´ [F., < Afr. macaquo.] Cercopithecine monkeys of the genus macacus. Their form is stout; large ischial tuberosities: muzzle considerably produced.
Ma-ca´cus.
Mam´mal [Lat. mamma, breast; suffix -al; in analogy with Animal. Dr. Th. Gill.] Vertebrate animal whose female has mammæ, or milk-secreting organs.
Mam-ma´li-a [Vid. mammal.]
Man´tis [Gr. mantis, prophet.] The mantises are insects noted for the manner in which they carry the large spinous fore legs when waiting for prey. They have the attitude, then, as if praying.
Mar´mo-set. A small South-American monkey.
Mar-si-po-bran´chi-i [Gr. marsipos, bag; branchia, gills.]
Mar-su´pi-al [Lat. marsupium, pouch.] Animals having a marsupium or pouch for retaining the young.
Mas´to-don [Gr. mastos, breast; odous, tooth.] An extinct elephant.
Me-dul´la [Lat. medius, middle.] A subdivision of the brain—that portion especially that is continuous with the spinal chord.
Men-o-bran´chus [Gr. meno, remain; branchia, gills.] A large American aquatic amphibian, of salamander-like form, with persistent gills, as the mud-puppy.
Mens sana in corpore sano. A Latin phrase meaning a sound mind in a sound body.
Mes-en-ceph´a-lon [Gr. mesos, middle; enkephalos, brain.] A fundamental segment of the brain.
Mes´en-chyme [Gr. mesos, middle; enchyma, infusion.]
Mes´o-blast [Gr. mesos, middle; blastos, germ.]
Mes-o-glϫa [Gr. mesos, middle; gloia, glue.]
Mes-o-hip´pus [Gr. mesos, middle; hippos, horse.]
Mes-o-zo´ic [Gr. mesos, middle; zoe, life.]
Met-a-dis-coi´dal [Gr. meta, after; diskos, disk; eidos, appearance.] Resembling a discoidal form.
Met-a-mor´pho-sis [Gr. meta, over; morphe, form.] The series of pronounced external changes through which an animal passes after leaving the egg-envelopes and before reaching sexual maturity.
Met-a-phy´ta [Gr. meta, above, higher; phyton, plant.]
Met´a-plasm [Gr. metaplasmos, transformation.]
Met-a-zo´a [Gr. meta, after, higher; zoon, animal.]
Met-en-ceph´a-lon [Gr. meta, after; enkephalos, brain.] A fundamental segment of the brain.
Mi-cro-ceph-a´li-a [Gr. mikrokephalos, small-headed.] Imperfect development of the cranium.
Mi-cren-ceph-a´li-a [Gr. mikros, small; enkephalos, brain.] Small-brained.
Mi´cro-cosm [Gr. mikros, small; kosmos, world.]
Micro-gro´mi-a so-ci-al´is [Gr. mikros, small; gromia; Lat. socialis < socius, companion.]
Mi-cro-les´tes.
Mi´das. A wealthy king of Phrygia.
Mi-o-hip´pus [Miocene; Gr. hippos, horse.]
Mi´o-cene [Gr. meion, less; kainos, recent.]
Mi-to´sis [Gr. mitos, thread.]
Mol-lus´ca [Lat. mollis, soft.]
Mon-o-cot-y-le´don-es [Gr. monos, single; kotyledon, cup-shaped cavity.] A group of flowering plants in which the first leaves of the embryo are alternate, for which reason they are said to have one seed-leaf or cotyledon.
Mon-o-del´phi-a [Gr. monos, single; delphys, womb.] A sub-class of mammals having a single vagina and uterus; embryo attached by a placenta; brain has a corpus callosum. Includes all animals above Monotremes and Marsupials.
Mon´o-tremes [Gr. monos, single; trema, hole.] An order of ornithodelphians.
Mor´u-la [Lat. dim. of morum, mulberry.]
Mul-ti-tu-ber-cu-la´ta [Lat. multus, many; tuberculum, tubercle.]
My-ce´tes [Gr. myketes, bellower.]
Myr-i-ap´o-da [Gr. myrios, numberless; pous, foot.] Insects with numerous pairs of legs, as centipedes.
Nau´ti-loids [Gr. nautilos, sailor; eidos, like.] A group of cephalopods.
Nau´ti-lus [Gr. nautilos, sailor.] A cephalopod.
Nem-a-thel-min´thes [Gr. nema, thread; helmins, worm.]
Nem-er-ti´na [Gr. nemertes, unerring.] Worms with skin ciliated, proboscis retractile, and nervous, muscular, and vascular systems characteristically developed.
Ne-o-lith´ic [Gr. neos, new; lithos, stone.]
Noc-ti-lu´ca mi-li-a´ris [Lat. nox, night; luceo, shine; miliarius, < milium, millet.]
No´to-chord [Gr. notos, the back; chorda, a chord.] A cartilaginous rod found in the young chordate embryo in a situation[270] that is later on occupied by the centers of the bodies of the vertebræ. It separates a dorsal nervous system from a ventral alimentary (digestive) canal. The notochord is persistent in its entirety in Leptocardians and Marsipobranchs and also in some pisces, as the sturgeon.
O-don-tor´ni-thes [Gr. odous, a tooth; ornis, bird.] Including all those extinct birds having teeth.
Œ-soph´a-gus [Gr. oiso, will bear; phagein, eat.] The gullet; the membranous tube through which food passes from the pharynx to the stomach.
On-tog´e-ny [Gr. on, being; genesis, origin.] The history of the evolution of the individual; the development of the individual.
On-to-ge-net´ic [Gr. on, being; genesis, origin.] Pertaining to ontogeny.
On-y-choph´o-ra [Gr. onyx, claw; pherein, bear.] Arthropods including only a single genus, the curious caterpillar-like Peripatus.
Oösperm [Gr. oon, egg; sperma, semen.] A fertilized ovum. The first stage in the existence of a human being is called oösperm.
O-per´cu-lum [Lat. lid.] The gill-cover.
O-phid´i-a [Gr. ophis, serpent.] Serpents; snakes.
Op-po´nens Hal-lu´cis [Lat. ob, pono, to place over against; hallux, great toe.] The name of a muscle in the foot.
Or-do-vi´cian [ordovices, ancient Celtic tribe in Wales.]
Or-nith-o-del´phi-a [Gr. ornis, bird; delphys, womb.] A sub-class of oviparous mammals.
Or-o-hip´pus [Gr. oros, mountain; hippos, horse.]
Or-tho-cer´a-tites [Gr. orthos, straight; keras, horn.]
Os´te-o-blasts [Gr. osteon, bone; blastano, sprout.]
Os´te-o-clasts [Gr. osteon, bone; klao, destroy.]
Os´tra-co-derms [Gr. ostrakon, shell; derma, skin.]
Ovum [Lat. egg.] Female germ-cell; female, encysted, sexual cell.
O-vip´a-rous [Lat. ovum, egg; pairo, produce.] Producing eggs that mature and are hatched outside the body.
Pal´li-um [Gr. mantle.]
Pa-læ-o-lith´ic [Gr. palaios, ancient; lithos, stone.]
Pa-læ-o-zo´ic [Gr. palaios, ancient; zoe, life.]
Pan-do-ri´na mor´um [Gr. Pandora; moron, mulberry.]
Pan-gen´e-sis [Gr. pas, all; genesis, origin.]
Par-a-me´ci-um [Gr. paramekes, of longish shape.]
Par-then-o-gen´e-sis [Gr. parthenos, virgin; genesis, origin.] Production of a new individual from a virgin female without intervention of a male; reproduction by means of unfertilized eggs.
Per-en-ni-bran-chi-a´ta [Lat. perennis, perpetual; branchia, gills.]
Per-en-ni-bran´chi-ates [Vid. Perennibranchiata.]
Ped´al [Lat. pes, foot.]
Per´mi-an [Perm, Russia.]
Phan´er-o-gams [Gr. phaneros, visible; gamos, marriage.] Flowering plants.
Phos-phor-es´cence.
Phryn-o-ceph´a-lus mys-ta´ce-us.
Phy-log´e-ny [Gr. phylon, tribe; genesis, origin.] The history of the evolution of a species or group; distinguished from ontogeny.
Phy-lo-ge-net´ic [Vid. phylogeny.] Pertaining to phylogeny.
Pith´e-coid [Gr. pithekos, ape; eidos, like.]
Pith-e-can-thro´pus [Gr. pithekos, ape; anthropos, man.]
Pla-gi-os´to-mi [Gr. plagios, oblique; stoma, mouth.] A division of Elasmobranchii, including sharks and rays.
Pla-cen´tal [Lat. placenta, cake.] So called on account of its shape. Pertaining to the placenta, the organ of attachment of higher vertebrate (monodelph) embryos to the wall of the uterus of the female.
Plas´tids [Gr. plastos, formed.]
Plat-y-hel-min´thes [Gr. platys, flat; helminos, worm.]
Plat-y-rhi´næ [Gr. platys, broad, flat; rhis, nose.] Monkeys of the New World having long tails and a wide septum to the nose.
Pleur-a-can´thus [Gr. pleuron, rib; akantha, thorn.]
Pleu´ral [Gr. pleura, rib, side.]
Pli-o-hip´pus [Pliocene; Gr. hippos, horse.]
Pli´o-cene [Gr. pleion, more; kainos, recent.]
Plumes [Lat. pluma, small soft feather.] Feathers, especially when ornamental or large; or tufts of large and ornamental feathers.
Pneu-mo-coc´ci [Gr. pneumon, lung; kokkos, berry.] Bacteria (spheroidal) found in those suffering from pneumonia.
Pol-y-dac´tyl-ism [Gr. polys, many; daktylos, finger, toe.] The possession of an abnormally large number of fingers or toes.
Pol-y-mor´phism [Gr. polys, many; morphe, form.] Exhibition by a group of animals, as a species, of different types of form or structure.
Po-lyp´te-rus [Gr. polypteros, many-winged.]
Po-ly´o-don [Gr. polys, many; odous, tooth.]
Pol´yp [Gr. polys, many; pous, foot.]
Pol´y-the-ism [Gr. polys, many; theos, god.]
Po-rif´e-ra [Lat. porus, pore; fero, bear.]
Pre-Pa-læ-o-lith´ic [Lat. præ, before; Palæolithic.] The period of time preceding the Ancient Stone Age.
Pri-ma´tes [Lat. primus, first.] An order of mammals.
Pro-bos-cid´e-a [Gr. pro, before; bosko, feed.]
Pro-nu´cle-us [Lat. pro, before; nucleus, kernel.]
Pros-en-ceph´a-lon [Gr. pro, before; enkephalos, brain.] A fundamental segment of the brain.
Pro-to-hip´pus [Gr. protos, first; hippos, horse.]
Pro-toph´y-ta [Gr. protos, first; phyton, plant.]
Pro´to-plasm [Gr. protos, first; plasma, < plasso, form.]
Pro-top´te-rus [Gr. protos, first; pteron, wing.]
Pro-to-zo´a [Gr. protos, first; zoon, animal.]
Pseu-do-he-red´i-ty [Gr. pseudes, false; Lat. heredita, heirship.]
Pseu-do-po´di-a [Gr. pseudes, false; pous, foot.]
Psy-cho-zo´ic [Gr. psyche, soul; zoe, life.]
Pter-i-doph´y-ta [Gr. pteris, fern; phyton, plant.]
Pter-o-sau´ri-a [Gr. pteron, wing; sauros, lizard.]
Pti-lo´pus cinc´tus.
Pu´pa, pl. pupæ [Lat. doll, girl, f. of pupus, boy.] The third stage of those insects that undergo a complete metamorphosis. Pupæ are often surrounded by protective envelopes called cocoons.
Quad-ru´la sym-met´ri-ca.
Qua-ter´na-ry [Lat. quaternarius, consisting of four.]
Ra-ti´tæ [Lat. ratis, raft.] A group of birds whose sternum is without a keel; aborted wings. Including ostriches, emus, moas, rheas, etc.
Rhi-noc´e-ros [Gr. rhis, nose; keras, horn.]
Rhi-zop´o-da [Gr. rhiza, root; pous, foot.]
Ro-den´ti-a [Lat. rodo, gnaw.]
Ro-ta´li-a Frey´er-i [Lat. rota, wheel; Freyer.]
Ro-ta-to´ri-a [Lat. rota, wheel.]
Ro´ti-fer [Lat. rota, wheel; fero, bear.]
Ru´mi-nant [Lat. ruminans, ruminate.]
Sai´tis pu´lex.
Saur-u´ræ [Gr. sauros, lizard; oura, tail.] Jurassic birds having lizard-like tails with distichous feathers, as Archæopterygidæ.
Sca-phop´o-da [Gr. skaphos, bowl, skiff; pous, foot.] A division of molluscs.
Sep´al [Lat. separ, separate.] One of the individual leaves of a calyx.
Si-lu´ri-an [Silures, ancient Celts in Wales.]
Sim´i-a [Lat. simia, ape.] A genus typical of Simiidæ and now restricted to the orang.
Si-mi´i-dæ [Lat. simia, ape.] A family of Old World Anthropoidea.
Sim-i-i´næ [Lat. simia, ape.] A subfamily of Simiidæ with robust form and molars tuberculated as in man and no ischial tuberosities, including Chimpanzees, Gorillas, and Orangs.
Si´nus veno´sus [Lat.] Venous cavity. (1) In higher vertebrates the main part of the auricular cavity. (2) In the lower vertebrates, a dilitation at the termination of venous channels, forming a separate chamber.
Si-re´ni-a [Lat. siren, siren.]
Snags [Norw. snag, a projecting point.] A branch on the antler of a deer.
So-mat´ic [Gr. soma, body.] Pertaining to the body.
So-ma-to-ge-net´ic [Gr. soma, body; genesis, generation, origin.] Originating in the body or soma through external influences.
Sper-ma-to-zo´id [Gr. sperma, seed, semen; zoon, animal.] Male germ-cell; male, flagellate, (“ciliated”) sexual cell. At one time supposed to be minute parasites in the semen, hence often called spermatozooa.
Sphinx fu-ci-form´is.
Spon´gi-o-plasm [Gr. spongion, little sponge; plasso, form.]
Steg-o-ceph´ala [Gr. stego, cover; kepale, head.] Another name for Labyrinthodonts.
Sty-lo-nych´i-a.
Su-pra-œ-soph-ag´e-al [Lat. supra, above; œsophagus.]
Tar´si-us [Gr. tarsos, any flat surface.]
Tax-o-nom´ic [Gr. taxis, orderly arrangement; nemein, distribute.] Pertaining to systematic classification.
Tel-e-os´to-mi [Gr. teleos, perfect; stoma, mouth.] A division of fishes with well-developed dentary and maxillary and membrane bones; includes all fishes except Elasmobranchii and lower forms.
Tel´e-osts [Gr. teleos, perfect; osteon, bone.] A subdivision of Teleostomi that includes most fishes; decussation of optic nerves; non-contractile arterial bulb.
Ter´ti-a-ry [Lat. tertius, third.]
Tet´ra-o cu´pi-do [Lat. tetrao, pheasant; Cupido, the God of Love.]
Thal-am-en-ceph´a-lon [Gr. thalamos, chamber; enkephalos, brain.] A fundamental segment of the brain.
Thal´a-mi [Gr. thalamos, chamber.] The lateral boundaries of the thalamencephalon.
Thal´loph´y-ta [Gr. thallos, young branch; phyton, plant.]
The-ro-mor´pha [Gr. ther, beast; morphe, form.]
Tho-mi´sus cit´re-us.
Ti-mor´.
Top´knots. Crests or tufts of feathers growing on the heads of birds.
Toxalbu´min [Gr. toxikon, poison; albumin, < Lat. albus, white.]
Tri-as´sic [Gr. trias, three.]
Tri´lo-bites [Gr. treis, three; lobos, lobe.]
Tu-ni-ca´ta [Lat. tunica, tunic.] A class of Chordata called Sca-squirts or Ascidians.
Tynes [AS. tind, prong.] The spike or prong of an antler.
Un-gu-la´ta [Lat. unguis, nail.] The order of hoofed mammals.
U-ro-de´la [Gr. oura, tail; delos, manifest.] The tailed Amphibians.
U´te-rus [Lat. uterus, womb.] The organ of a female animal in which the young are protected and developed before birth.
Vac´u-ole [Lat. vacuus, empty.] The little cavities in cells containing watery solutions.
Ves´i-cle [Lat. vesicula, little bladder.] Synonymous with vacuoles.
Ve´na ca´va [Lat.] Hollow vein. Either of the two great veins that empty into the right auricle of the heart.
Ven´tri-cles [Lat. ventriculus, little belly.] One of the cavities of the heart or brain.
Ven´tral [Lat. venter, belly.] Applied to the front (under) side of the body.
Ver´mes [Lat. vermis, worm.]
Ver-te-bra´ta [Lat. vertebratus, jointed.]
Ver´te-brate [Lat. vertebratus, jointed.]
Vi-vip´a-rous [Lat. vivus, alive; pario, bear.] Applied to animals which bring forth their young alive.
Vis´ce-ra [Lat. viscera, organs.] The internal organs of the body.
Vol´vox glo-ba´tor [Lat. volvo, roll; globatus, made into a ball.]
Wat´tles [AS. watel, hurdle.] Fleshy, naked processes depending from the neck or head of a bird.
[1] One cubic millimeter of human blood alone, not to mention other tissues of the body, contains over five million cells.
[2] Except in the maturation of ova and spermatozoids.
[3] Bradford.
[4] Bradford.
[5] Socially, morally, and religiously.
[6] Lyell.
[7] Readers not familiar with Elementary Zoölogy will do well to consult the Classification of Animals in Section VII.; and also the Diagram of Development.
[8] Wallace.
[9] Belt.
[10] Wallace.
[11] Wallace.
[12] Wallace.
[13] Darwin.
[14] Darwin.
[15] Lloyd Morgan.
[16] Clodd.
[17] “Feelings and motions run parallel to each other, and where we do not meet with actual feelings we suppose the presence of the elements of feeling. But this parallelism would be most wonderful indeed if it were a true parallelism consisting of two different and distinct lines. The simplest conception of the case is the monistic view, which considers the parallelism as an identity. Both motion and feeling are abstract conceptions. A motion exists of itself no more than a feeling. The reality from which the ideas motion and feeling have been abstracted is one inseparable whole, which if viewed as an objective process appears as motion, and if viewed from the subjective side appears as feeling. Feelings can only be felt, not seen; but if we could see them, we might observe the elements of feeling wherever motion takes place.
“Fechner seems to have hit the mark, when he compared feeling and motion to the inside and the outside curves of a circle; they are entirely different and yet the same. The inside curve is concave, the outside curve is convex. If we construct rules relating first to the concave inside and then to the convex outside, we shall notice a parallelism in the formulas; yet this parallelism will appear only in the abstractions which have been made of one and the same thing from a different aspect. It results from making two different abstractions. The abstract conceptions form two parallel systems, but the real thing can be represented as parallel only in the sense that it is parallel to itself; it is the parallelism of identity. There is but one line and this one line is concave if viewed from the inside, if viewed from the outside convex.” Dr. Carus in “The Soul of Man,” p. 20.
[18] Central Soul (Dr. Carus).
[19] Clifford.
[20] It is more scientific to say a Cosmic Soul immanent in the Cosmos.
“God, as I conceive him to be, is not less than a person, but more than a person.” Dr. Paul Carus.
[21] Fiske.
[22] Fiske.
[23] “Religion is the conception by man of his relation to the infinite universe and its source; and morality is the ever present guide of life proceeding only from this relation.” Tolstoi.
[24] Fiske.
[25] Society also utterly ignores the rights of unborn children in permitting the constant marriages of men and women having constitutions tainted with hereditary diseases, as insanity, syphilis, tuberculosis, etc.
[26] In Dipnoi, which are fishes with Amphibian characters, the paired fins have each a central segmented axis which bears on each side a series of radial pieces. The air bladder is a true lung, and is frequently used as such. The heart shows indications of becoming three chambered (two auricles and one ventricle). The left auricle contains arterial blood from the lungs. There is an inferior vena cava. The nasal sacs open posteriorly into the mouth.
THE PRIMARY FACTORS OF ORGANIC EVOLUTION. By Prof. E. D. Cope. Cuts, 121. Pp., xvi, 547. Cloth, $2.00 (10s.).
DARWIN AND AFTER DARWIN. An Exposition of the Darwinian Theory and a Discussion of Post-Darwinian Questions. By George John Romanes, LL. D., F. R. S., etc.
(For the American market only.)
1. The Darwinian Theory. With portrait of Darwin. Pp., 460. 125 cuts. Second edition. Cloth, $2.00.
2. Post-Darwinian Questions. Heredity and Utility. With portrait of Romanes. Pp., 338. Cloth, $1.50.
3. Post-Darwinian Questions. Isolation and Physiological Selection. With portrait of Mr. J. T. Gulick. Pp., 181. 8vo. Cloth, $1.00.
(The three volumes supplied to one order for $4.00.)
AN EXAMINATION OF WEISMANNISM. By George John Romanes. Pp., ix, 221. Cloth, $1.00. Paper, 35c.
THE PSYCHIC LIFE OF MICRO-ORGANISMS. By Dr. Alfred Binet. Authorised translation. Pp., xii, 120. Cloth, 75c. (3s. 6d.). Paper, 25c. (1s. 6d.).
ON GERMINAL SELECTION. By August Weismann. Pp., xii, 61. Paper, 25c. (1s. 6d.).
ON MEMORY, AND THE SPECIFIC ENERGIES OF THE NERVOUS SYSTEM. By Ewald Hering. 50 pp. Paper, 15c.
A MECHANICO-PHYSIOLOGICAL THEORY OF ORGANIC EVOLUTION. Summary. By Carl von Nägeli. Only original account of Nägeli’s theories in English. Pp., 52. Paper, 15c. (9d.).
ON ORTHOGENESIS. By Th. Eimer. Pp., 56. Paper, 25c. (1s. 6d.).
THE PRINCIPLES OF BACTERIOLOGY. By Dr. Ferdinand Hueppe, Professor of Hygiene in the University of Prague. 28 Woodcuts. Pp., 465+. $1.75 (7s. 6d.).
THE OPEN COURT PUBLISHING CO.
CHICAGO: 324 Dearborn Street.
LONDON: Kegan Paul, Trench, Trübner & Co.
THE PRIMARY FACTORS OF ORGANIC EVOLUTION.
121 cuts. Pp. xvi. 547. Cloth, $2.00 net (10s.).
THREE INTRODUCTORY LECTURES ON THE SCIENCE OF THOUGHT.
128 pages. Cloth, 75c (3s. 6d.).
THREE LECTURES ON THE SCIENCE OF LANGUAGE.
112 pages. 2nd Edition. Cloth, 75c (3s. 6d.).
DARWIN AND AFTER DARWIN.
An Exposition of the Darwinian Theory and a Discussion of Post-Darwinian Questions. Three Vols., $4.00 net. Singly, as follows:
1. The Darwinian Theory. 460 pages. 125 illustrations. Cloth, $2.00.
2. Post-Darwinian Questions. Heredity and Utility. Pp. 338. $1.50.
3. Post-Darwinian Questions. Isolation and Physiological Selection. Pp. 181. $1.00.
AN EXAMINATION OF WEISMANNISM.
236 pages. Cloth, $1.00 net.
THOUGHTS ON RELIGION.
Edited by Charles Gore, M. A., Canon of Westminster. Third Edition, Pages, 184. Cloth, gilt top, $1.25 net.
FIRST BOOK IN ORGANIC EVOLUTION.
Colored plates, and numerous diagrams. Price, $2.00 (10s.).
THE SCIENCE OF MECHANICS.
A Critical and Historical Exposition of its Principles. Translated by T. J. McCormack. 250 cuts. 534 pages. ½ m., gilt top. $2.50 (12s. 6d.).
POPULAR SCIENTIFIC LECTURES.
Third Edition. 415 pages. 59 cuts. Cloth, gilt top. Net, $1.50 (7s. 6d.).
THE ANALYSIS OF THE SENSATIONS.
Pp. 208. 37 cuts. Cloth, $1.25 net (6s. 6d.).
LECTURES ON ELEMENTARY MATHEMATICS.
With portrait of the author. Pp. 172. Price, $1.00 net (5s.).
ON THE STUDY AND DIFFICULTIES OF MATHEMATICS.
New Reprint edition with notes. Pp. viii+288. Cloth, $1.25 net (5s.).
ELEMENTARY ILLUSTRATIONS OF THE DIFFERENTIAL AND INTEGRAL CALCULUS.
New reprint edition. Price, $1.00 (5s.).
MATHEMATICAL ESSAYS AND RECREATIONS.
Pp. 149. Cuts, 37. Cloth, 75c net (3s. 6d.).
TRAVELS IN TARTARY, THIBET AND CHINA.
(1844-1846.) Translated from the French by W. Hazlitt. Illustrated with 100 engravings on wood. 2 vols. Pp. 28+660. Cl., $2.00 (10s.).
THE ETHICAL PROBLEM.
Second edition, revised and greatly enlarged. 351 pages. Cloth, $1.25 (6s. 6d.).
FUNDAMENTAL PROBLEMS.
Second edition, enlarged and revised. 372 pp. Cl., $1.50 (7s. 6d.).
HOMILIES OF SCIENCE.
317 pages. Cloth, Gilt Top, $1.50 (7s. 6d.).
THE IDEA OF GOD.
Fourth edition. 32 pages. Paper, 15c (9d.).
THE SOUL OF MAN.
With 152 cuts and diagrams. 458 pages. Cloth, $3.00 (15s.).
TRUTH IN FICTION. Twelve Tales with a Moral.
White and gold binding, gilt edges. Pp. 111. $1.00 (5s.).
THE RELIGION OF SCIENCE.
Second, extra edition. Pp. 103. Price, 50c net (2s. 6d.).
PRIMER OF PHILOSOPHY.
240 pages. Second Edition. Cloth, $1.00 (5s.).
THE GOSPEL OF BUDDHA. According to Old Records.
Fifth Edition. Pp. 275. Cloth, $1.00 (5s.). In German, $1.25 (6s. 6d.).
BUDDHISM AND ITS CHRISTIAN CRITICS.
Pages, 311. Cloth, $1.25 (6s. 6d.).
KARMA. A Story of Early Buddhism.
Illustrated by Japanese artists. Crêpe paper, 75c (3s. 6d.).
NIRVANA: A Story of Buddhist Psychology.
Japanese edition, like Karma. $1.00 (4s. 6d.).
LAO-TZE’S TAO-TEH-KING.
Chinese-English. With introduction, transliteration, Notes, etc. Pp. 360. Cloth, $3.00 (15s.).
THE PROPHETS OF ISRAEL.
Popular Sketches from Old Testament History. Pp., 200. Cloth, $1.00 net (5s.).
HISTORY OF THE PEOPLE OF ISRAEL.
Pp. vi+325. Cloth, $1.50 (7s. 6d.).
TRUTH AND ERROR; or, the Science of Intellection.
Pp. 423. Cloth, $1.75 (7s. 6d.).
THE PSYCHOLOGY OF ATTENTION.
THE DISEASES OF PERSONALITY.
THE DISEASES OF THE WILL.
Authorised translations. Cloth, 75 cents each (3s. 6d.). Full set, cloth, $1.75 net (9s.).
EVOLUTION OF GENERAL IDEAS.
Pp. 231. Cloth, $1.25 net (6s. 6d.).
A PILGRIMAGE TO BEETHOVEN.
A Story. With portrait of Beethoven. Pp. 40. Boards, 50c net (2s. 6d.).
THE GOSPEL ACCORDING TO DARWIN.
Pp. xii+241. Price, $1.50 (6s.).
THE LOST MANUSCRIPT. A Novel.
2 vols. 953 pages. Extra cloth, $4.00 (21s.). One vol., cl., $1.00 (5s.).
MARTIN LUTHER.
Illustrated. Pp. 130. Cloth, $1.00 net (5s.).
THE FREE TRADE STRUGGLE IN ENGLAND.
Second Edition. 296 pages. Cloth, 75c (3s. 6d.).
WHEELBARROW: Articles and Discussions on the Labor Question.
With portrait of the author. 303 pages. Cloth, $1.00 (5s.).
GOETHE AND SCHILLER’S XENIONS.
Translated by Paul Carus. Album form. Pp. 162. Cl., $1.00 (5s.).
ANCIENT INDIA: ITS LANGUAGE AND RELIGIONS.
Pp. 100. Cloth, 50c net (2s. 6d.).
SOLOMON, AND SOLOMONIC LITERATURE.
Pp. 243. Cloth, $1.50 net (6s.).
THE REDEMPTION OF THE BRAHMAN. A Tale of Hindu Life.
Laid paper. Gilt top. 96 pages. Price, 75c (3s. 6d.).
THE PHILOSOPHY OF ANCIENT INDIA.
Pp. 89. Cloth, 50c net (2s. 6d.).
THE PRINCIPLES OF BACTERIOLOGY.
28 Woodcuts. Pp. x+467. Price, $1.75 net (9s.).
HISTORY OF MODERN PHILOSOPHY IN FRANCE.
With 23 Photogravure and Halt-Tone Portraits of the Chief French Philosophers. Handsomely bound. $3.00 (15s.).
SCIENCE AND FAITH, or Man as an Animal and Man as a Member of Society.
Pp. 374. Cloth, $1.50 net (7s. 6d.).
THE PSYCHOLOGY OF REASONING.
Pp. 193. Cloth, 75c. (3s. 6d.).
THE PSYCHIC LIFE OF MICRO-ORGANISMS.
Pp. 135. Cloth, 75 cents.
ON DOUBLE CONSCIOUSNESS. See No. 8, Religion of Science Library.
A Monthly Magazine Devoted to the Science of Religion, the Religion of Science, and the Extension of the Religious Parliament Idea.
Terms: $1.00 a year; 5s. 6d. to foreign countries in the Postal Union. Single Copies, 10 cents (6d.).
A Quarterly Magazine of Philosophy and Science.
Per copy, 50 cents; Yearly, $2.00. In England and all countries in U.P.U. per copy, 2s. 6d.: Yearly, 9s. 6d.
CHICAGO:
THE OPEN COURT PUBLISHING CO.,
Monon Building, 324 Dearborn St.
LONDON: Kegan Paul, Trench, Trübner & Company.
A collection of bi-monthly publications, most of which are reprints of books published by The Open Court Publishing Company. Yearly, $1.50. Separate copies according to prices quoted. The books are printed upon good paper, from large type.
The Religion of Science Library, by its extraordinarily reasonable price, will place a large number of valuable books within the reach of all readers.
The following have already appeared in the series:
| No. | 1. | The Religion of Science. By Paul Carus. 25c (1s. 6d). |
| 2. | Three Introductory Lectures on the Science of Thought. By F. Max Müller. 25c (1s. 6d.). | |
| 3. | Three Lectures on the Science of Language. F. Max Müller. 25c (1s. 6d.). | |
| 4. | The Diseases of Personality. By Th. Ribot. 25c (1s. 6d.). | |
| 5. | The Psychology of Attention. By Th. Ribot. 25c (1s. 6d.). | |
| 6. | The Psychic Life of Micro-Organisms. By Alfred Binet. 25c (1s. 6d.). | |
| 7. | The Nature of the State. By Paul Carus. 15c (9d.). | |
| 8. | On Double Consciousness. By Alfred Binet. 15c (9d.). | |
| 9. | Fundamental Problems. By Paul Carus. 50c (2s. 6d.). | |
| 10. | The Diseases of the Will. By Th. Ribot. 25c (1s. 6d.). | |
| 11. | The Origin of Language. By Ludwig Noire. 15c (9d.). | |
| 12. | The Free Trade Struggle in England. M. M. Trumbull. 25c (1s. 6d.). | |
| 13. | Wheelbarrow on the Labor Question. By M. M. Trumbull. 35c (2s.). | |
| 14. | The Gospel of Buddha. By Paul Carus. 35c (2s.). | |
| 15. | The Primer of Philosophy. By Paul Carus. 25c (1s. 6d.). | |
| 16. | On Memory, and The Specific Energies of the Nervous System. By Prof. Ewald Hering. 15c (9d.). | |
| 17. | The Redemption of the Brahman. A Tale of Hindu Life. By Richard Garbe. 25c (1s. 6d.). | |
| 18. | An Examination of Weismannism. By G. J. Romanes. 35c (2s.). | |
| 19. | On Germinal Selection. By August Weismann. 25c (1s. 6d.). | |
| 20. | Lovers Three Thousand Years Ago. By T. A. Goodwin. (Out of print.) | |
| 21. | Popular Scientific Lectures. By Ernst Mach. 50c (2s. 6d.). | |
| 22. | Ancient India: Its Language and Religions. By H. Oldenberg. 25c (1s. 6d.). | |
| 23. | The Prophets of Israel. By Prof. C. H. Cornill. 25c (1s. 6d.). | |
| 24. | Homilies of Science. By Paul Carus. 35c (2s.). | |
| 25. | Thoughts on Religion. By G. J. Romanes. 50c (2s. 6d.). | |
| 26. | The Philosophy of Ancient India. By Prof. Richard Garbe. 25c (1s. 6d.). | |
| 27. | Martin Luther. By Gustav Freytag. 25c (1s. 6d.). | |
| 28. | English Secularism. By George Jacob Holyoake. 25c (1s. 6d.). | |
| 29. | On Orthogenesis. By Th. Eimer. 25c (1s. 6d.). | |
| 30. | Chinese Philosophy. By Paul Carus. 25c (1s. 6d.). | |
| 31. | The Lost Manuscript. By Gustav Freytag. 60c (3s.). | |
| 32. | A Mechanico-Physiological Theory of Organic Evolution. By Carl von Nägeli. 15c (9d.). | |
| 33. | Chinese Fiction. By Dr. George T. Candlin. 15c (9d.). | |
| 34. | Mathematical Essays and Recreations. By H. Schubert. 25c (1s. 6d.). | |
| 35. | The Ethical Problem. By Paul Carus. 50c (2s. 6d.). | |
| 36. | Buddhism and Its Christian Critics. By Paul Carus. 50c (2s. 6d.). | |
| 37. | Psychology for Beginners. By Hiram M. Stanley. 20c (1s.). | |
| 38. | Discourse on Method. By Descartes. 25c (1s. 6d.). |
THE OPEN COURT PUBLISHING CO.,
CHICAGO: 324 Dearborn Street.
LONDON: Kegan Paul, Trench, Trübner & Company.