Transcriber’s notes:
The text of this e-book has largely been preserved in its original
form. However, some spelling
errors have been corrected and some missing punctuation
items inserted (hyphen, space, parenthesis, quotation mark). Inconsistent spellings have not
been changed. Missing footnote markers on pages 96 and 136 have been
inserted at what seemed to be likely locations. A few long paragraphs
have been split to allow repositioning of diagrams closer to the
relevant text. Two diagrams have the same number (Fig. 24.).
Hyperlinks are indicated by a black underline and coloured
highlighting when the mouse pointer hovers over them. Page numbers are shown in the right margin. Footnotes
have been numbered consecutively and are located at the end. Footnotes have been numbered consecutively and are
listed at the end.
The cover image of the book was created by the
transcriber and is placed in the public domain.
YALE UNIVERSITY
MRS. HEPSA ELY SILLIMAN MEMORIAL LECTURES
IRRITABILITY
SILLIMAN MEMORIAL LECTURES
PUBLISHED BY YALE UNIVERSITY PRESS
ELECTRICITY AND MATTER. By Joseph John Thomson, D.Sc.,
LL.D., Ph.D., F.R.S., Fellow of Trinity College, Cambridge, Cavendish
Professor of Experimental Physics, Cambridge.
Price $1.25 net; postage 10 cents extra.
THE INTEGRATIVE ACTION OF THE NERVOUS SYSTEM. By
Charles S. Sherrington, D.Sc., M.D., Hon. LL.D., Tor., F.R.S., Holt
Professor of Physiology in the University of Liverpool.
Price $3.50 net; postage 25 cents extra.
RADIOACTIVE TRANSFORMATIONS. By Ernest Rutherford,
D.Sc., LL.D., F.R.S., Macdonald Professor of Physics, McGill University.
Price $3.50 net; postage 22 cents.
EXPERIMENTAL AND THEORETICAL APPLICATION OF
THERMODYNAMICS TO CHEMISTRY. By Walther Nernst,
Professor and Director of the Institute of Physical Chemistry in the
University of Berlin.
Price $1.25 net; postage 10 cents extra.
PROBLEMS OF GENETICS. By William Bateson, M.A., F.R.S.,
Director of the John Innes Horticultural Institution, Merton Park, Surrey,
England.
Price $4.00 net; postage 25 cents extra.
STELLAR MOTIONS, WITH SPECIAL REFERENCE TO
MOTIONS DETERMINED BY MEANS OF THE SPECTROGRAPH.
By William Wallace Campbell, Sc.D., LL.D., Director of the
Lick Observatory, University of California.
Price $4.00 net; postage 25 cents extra.
THEORIES OF SOLUTION. By Svante August Arrhenius, Ph.D.,
Sc.D., M.D., Director of the Physico-Chemical Department of the Nobel
Institute, Stockholm, Sweden.
Price $2.25 net; postage 14 cents extra.
IRRITABILITY, A PHYSIOLOGICAL ANALYSIS OF THE GENERAL
EFFECT OF STIMULI IN LIVING SUBSTANCE. By Max
Verworn, M.D., Ph.D., Professor at Bonn Physiological Institute.
Price $3.50 net; postage 20 cents extra.
IRRITABILITY
A PHYSIOLOGICAL ANALYSIS OF THE GENERAL
EFFECT OF STIMULI IN LIVING SUBSTANCE
BY
MAX VERWORN, M.D., Ph.D.
Professor at Bonn Physiological Institute
WITH DIAGRAMS AND ILLUSTRATIONS
New Haven: Yale University Press
London: Henry Frowde
Oxford University Press
MCMXIII
COPYRIGHT, 1913
By YALE UNIVERSITY PRESS
First Printed May, 1913, 600 Copies
THE SILLIMAN FOUNDATION.
In the year 1883 a legacy of eighty thousand dollars was left
to the President and Fellows of Yale College in the city of New
Haven, to be held in trust, as a gift from her children, in memory
of their beloved and honored mother, Mrs. Hepsa Ely Silliman.
On this foundation Yale College was requested and directed to
establish an annual course of lectures designed to illustrate the
presence and providence, the wisdom and goodness of God, as
manifested in the natural and moral world. These were to be
designated as the Mrs. Hepsa Ely Silliman Lectures. It is the
belief of the testator that any orderly presentation of the facts
of nature or history contributed to the end of this foundation
more effectively than any attempt to emphasize the elements of
doctrine or creed; and he therefore provided that lectures on
dogmatic or polemical theology should be excluded from the scope
of this foundation, and that the subjects should be selected rather
from the domains of natural science and history, giving special
prominence to astronomy, chemistry, geology, and anatomy.
It was further directed that each annual course should be made
the basis of a volume to form part of a series constituting a
memorial to Mrs. Silliman. The memorial fund came into the
possession of the corporation of Yale University in the year 1901;
and the present volume constitutes the ninth of the series of
memorial lectures.
PREFACE
The lectures on irritability here published were held at the
University of Yale in October, 1911. When the authorities of
that University honored me by an invitation to give a course of
Silliman memorial lectures, I accepted with the more pleasure as
it furnished me with the opportunity of summarizing the results of
numerous experimental researches carried out with the assistance
of my co-workers during the course of more than two
decades in the physiological laboratories of Jena, Göttingen and
Bonn, to unite therewith the results obtained by other investigators
and thus present a uniform exposition of the general effects
and laws of stimulation in the living substance. I have long
entertained this plan and this for the following reason:
The physiologist, the zoölogist, the botanist, the psychologist,
the pathologist, have to deal, day in, day out, with the effects of
stimulation on the living substance. No living substance exists
without stimulation. In the vital manifestations of all organisms
the interplay of the most varied stimuli produces an enormous
and manifold variety of effects. Experimental biological science
employs artificial stimulation as the most important aid in the
methodic production of certain effects of stimulation. The number
of researches in which special effects of stimulation are
treated is endless. Nevertheless the systematic investigation of
the effects of stimulation have, curiously enough, been strangely
neglected. Although countless results of individual effects of
stimulation have been studied, the attempt has never been made
to establish a general physiology of the laws of stimulation and
consider it as an independent problem. This circumstance induced
me to systematically investigate the general laws of the effect of
stimulation. In the fifth and sixth chapters of my book on
general physiology the results of these studies are recorded for
the first time. Since then, especially during our own researches
on the general physiology of the nervous system, a great numberviii
of new facts of importance for the general physiology of the
effects of stimulation have been obtained. All these results I
have endeavored to combine and elucidate in the following
lectures.
The text of the lectures in its present form was written in
German in 1911. The English translation was made by my wife,
with the help of our friend, Dr. Lodholz of the University of
Pennsylvania, who also undertook the reading of the proofs.
We wish here to thank him once again and express our deep
appreciation of the great sacrifice of time and labor involved in
this task. I am likewise much indebted to Dr. Julius Vészi for
his assistance unstintingly given, especially in obtaining a number
of curves. Finally, I wish to take this opportunity to render
warmest thanks to the authorities of Yale University, and especially
to President Hadley and Professor Chittenden, as well as
to my special colleagues, for the hospitality and cordial reception
extended to me in New Haven and for the pleasant hours I was
privileged to spend in their midst.
Max Verworn.
Bonn.
Physiological Laboratory of the University.
I |
| Contents: Introductory. Earliest period. Francis Glisson as founder
of the doctrine of irritability. Albrecht von Haller. The vitalists.
Bordeu and Barthez. John Brown’s system. Johannes Müller
and the specific energy of living substance. Rudolf Virchow’s
doctrine of the irritability of the cell. Discovery of the inhibitory
effects of stimulation. Weber, Schiff, Goltz, Setschenow, Sherrington.
Claude Bernard studies on narcosis. Tropisms. Ehrenberg,
Engelmann, Pfeffer, Strassburger, Stahl. Semon’s speculations
on mneme. | |
II |
| Contents: Principles of scientific knowledge and research. Origin
and meaning of the conception of cause. Cause and condition.
Criticism of the conception of cause. The conditional point of
view. Conception of cause. The conditional point of view applied
to the investigation of life. Conception of vital conditions. Definition
of the conception of stimulation. | |
III |
| Contents: The quality of the stimulus. Positive and negative alterations
of the factors which act as vital conditions. Extent of the
alteration in vital conditions or intensity of the stimulus. Threshold
stimuli, sub-threshold, submaximal, maximal and supermaximal
intensities of stimulus. Relations between the intensity of
stimulus and the amount of response. The Weber and Fechner
law. All or none law. Time relations of the course of the
stimulus. Form of individual stimulus. Absolute and relative
rapidity in the course of the stimulus. Duration of the stimulus
after reaching its highest point. Adaptation to persistent stimuli.
Series of individual stimuli. Rhythmical stimuli. The Nernst
law. | x |
IV |
| Contents: Various examples of the effects of stimulation. Metabolism
of rest and metabolism of stimulation. Metabolic equilibrium,
Disturbances of equilibrium by stimuli. Quantitative and
qualitative alterations of the metabolism of rest under the influence
of stimuli. Excitation and depression. Specific energy of
living substance. Qualitative alterations of the specific metabolism
and their relations to pathology. Functional and cytoplastic
stimuli. Relations of the cytoplastic effects of stimuli to the functional.
Hypertrophy of activity and atrophy of inactivity. Metabolic
alterations during growth of the cell. Primary and secondary
effects of stimulation. Scheme of effects of stimulation. | |
V |
| Contents: Indicators for the investigation of the process of excitation.
Latent period. The question of the existence of assimilatory excitations.
Dissimilatory excitations. Excitations of the partial
components of functional metabolism. Production of energy in
the chemical splitting up processes. Oxydative and anoxydative
disintegration. Theory of oxydative disintegration. Dependence
of irritability on oxygen. Experiments on unicellular organisms,
nerve centers and nerve fibers. Restitution after disintegration by
metabolic self-regulation. Organic reserve supplies of the cell.
The question of a reserve supply of oxygen of the cell. Metabolic
self-regulation as a form of the law of mass effect, and metabolic
equilibrium as a condition of chemical equilibrium. Functional
hypertrophy. | |
VI |
| Contents: Only processes of excitation are conducted, not processes of
depression. Conduction of excitation in its two extreme instances.
Conduction in undifferentiated pseudopod protoplasm of rhizopoda.
Conduction of excitation with decrement of intensity and rapidity.
Conduction of excitation in the nerve. Rapidity of conduction.
Conduction of excitation without decrement. Relation between
irritability and conductivity. Conduction of excitation with decrement
of the nerve after artificial depression of irritability by narcosis.
Theory of the decrementless conduction of the normal
nerve. Proof of the validity of the “all or none law” in the
medullated nerve. Theory of the process of the conductivity of
excitation. Theory of core model (Kernleiter). Electrochemical
theory of conduction based on the properties of semi-permeable
surfaces. | xi |
VII |
| Contents: Conception of specific irritability. Alteration of specific
irritability during and after excitation. Refractory period in
various forms of living substance. Absolute and relative refractory
period. Curve of irritability during refractory period.
Dependence of the duration of the refractory period on the
rapidity of the course of the metabolic processes in the living
substance. Dependence on temperature. Dependence on supply
of oxygen. Theory of refractory period. Refractory period as
basis of fatigue. Fatigue as a form of asphyxiation. Alterations
of irritability and the course of excitation in fatigue. Recovery
from fatigue. The rôle played by oxygen in recovery. Fatigue
as an expression of the prolongation of the refractory period
conditioned by the relative want of oxygen. Fatigue of the nerve. | |
VIII |
| Contents: Examples of effects of interference of stimuli in unicellular
organisms. Interference of galvanic and thermic stimuli in Paramecia.
Interference of galvanic and thermic stimuli and narcotics.
Interference of galvanic and mechanical stimuli. Interference of
galvanotaxis and thigmotaxis in Paramecia and hypotin infusoria.
Real or homotop interference, apparent or heterotop interference.
The two effects of homotop interference of excitations: Summation
and inhibition of excitations. Theory of the processes of
inhibition. Hering-Gaskell Theory. Inhibition as an expression
of the refractory period. Individual possibilities of interference
of two stimuli. Interference of an excitating and a depressing
stimulus. Interference of two depressing stimuli. Interference
of two excitating stimuli. Analysis of the interference of two
excitations. Interference of two single stimuli. Conditions upon
which the result of interference is dependent. Heterobole and
isobole living systems. Intensity of the two stimuli. Interval
between the stimuli. Specific irritability and rapidity of reaction
of the living system. Latent period. Interference of single stimuli
in a series. General scheme of the development of the effect
of interference. Summation and inhibition. Apparent increase of
irritability. Conditions of summation. Tonic excitations. Conditions
of inhibitions. Various types of inhibition. Interference of
two series of stimuli. Relations in the nervous system. Peculiarities
of the nerve fibers. Conversion of the nerve by relative
fatigue from an isobolic into a heterobolic system. | xii |
IX |
| Contents: Necessity of cellular physiological analysis of toxic depressions
by pharmacology. Apparent variety of processes of depression.
Depression of oxydative disintegration as the most extended
principle in the processes of depression. Asphyxiation, fatigue,
heat depression, as a consequence of restriction of oxydative disintegration.
Narcosis. Theories of narcosis. The alteration of
specific irritability and conductivity in narcosis. Depression of
oxydative processes in narcosis. Asphyxiation of living substance
when oxygen is present during narcosis. Persistence of anoxydative
disintegration in narcosis. Increase of the same by stimuli.
Depression by narcosis as a form of acute asphyxiation. Hypothesis
on the mechanism of depression of oxygen exchange by narcotics.
Possibility of combining the facts with the observations of
Meyer and Overton. | |
1
IRRITABILITY
CHAPTER I
THE HISTORY OF THE SUBJECT
Contents: Introductory. Earliest period. Francis Glisson as founder of
the doctrine of irritability. Albrecht von Haller. The vitalists. Bordeu
and Barthez. John Brown’s system. Johannes Müller and the
specific energy of living substance. Rudolf Virchow’s doctrine of the
irritability of the cell. Discovery of the inhibitory effects of stimulation.
Weber, Schiff, Goltz, Setschenow, Sherrington. Claude Bernard
studies on narcosis. Tropisms. Ehrenberg, Engelmann, Pfeffer,
Strassburger, Stahl. Semon’s speculations on mneme.
Irritability is a general property of living substance but not
exclusively so. Irritable systems also exist in inanimate nature.
What characterizes living substances is not irritability as such,
but an irritability of a specific type. The irritability of the living
system can, therefore, not be studied alone, but as the properties
of a living system are dependent upon each other, so this property
must be considered with the others possessed by a living substance.
In this sense irritability presents a problem of fundamental
physiological importance. For if we could analyze the
irritability of living substance to its essence, then the nature of
life itself would be fathomed. The analysis of irritability of
living substance offers us, therefore, a path to the investigation
of life and herein lies the importance of the study of irritability.
I wish to follow this path toward the knowledge of the vital
processes and to endeavor to show in these lectures what information
the analysis of irritability and that of the effect of stimuli
can give us of the mechanism of the processes in living substance.
Before doing so, however, I wish to consider somewhat more in
detail the question as to how we have arrived at the conception
of the nature of irritability.
2
To the thinkers both in the field of physiology and medicine of
ancient and mediæval times the conception of irritability was
quite foreign. Even a comprehension of the nature of stimuli
had not yet begun to crystallize from vague impressions of the
various influences of different agents on the human being.
Nevertheless they knew of such influences of the most varying
kinds upon the human body. The ancients already possessed
a materia medica, founded on the real or supposed influence of
various mineral, vegetable and animal substances upon the organism.
It was also known that heat and cold, light and darkness
had an effect upon disease. They likewise believed in the influence
of certain factors upon the health of man, which in reality
have no effect whatsoever, as the stars and the magnet. But
neither in ancient nor in mediæval times was the state of knowledge
reached wherein generalizations were made from these
agents, which had a real or supposed action upon the organism,
and to combine these to a general conception of stimulation.
The conception of stimulation and irritability cannot however
be separated.
The founder of the doctrine of the irritability of living substance
is Francis Glisson (1597–1677), member of the Collegium
Medicum in London and at the same time Professor in Cambridge.
It is a fact also not altogether without interest, that
Glisson at the same time was in a certain sense a forerunner of
those who interpreted nature from a physical standpoint. Glisson
as an anatomist and physiologist was an excellent observer and
experimenter, but the most prominent trait of his character
was his inclination to philosophic observation and analysis of
nature. His “Tractatus de natura substantiæ energetica”1 must,
therefore, be considered as the chief work of his life. In this
voluminous book Glisson develops an entire system of natural
philosophy, which in accord with the character of the philosophy
of that time is unfortunately of an absolutely speculative nature
and which had hardly emancipated itself from the scholasticism3
of the preceding period of thought. When the ideas of Glisson
are isolated from the wilderness of scholastic phraseology, the
system is somewhat as follows. The basis of all existence,
“substance,” has according to him two general properties, its
“fundamental subsistence,” that is, the essence of its being, and
its “energetic subsistence,” that is, the essence of its activity. To
these are added the properties possessed in specific cases, that is,
its “additional subsistence.” The energetic subsistence forms the
basis of all life. Life is therefore present not only in organic
nature, but in all nature which is characterized by the union of
the general energetic subsistence with the special additional subsistence
of an animal and vegetable nature. In other forms of
life in nature the energetic subsistence is combined with other
special forms of the additional subsistence. The universal
essence of all life, that is the energetic subsistence, has only three
fundamental faculties: the “appetitiva,” the “perceptiva” and the
“motiva.” The modus is the result of a “perceptio,” but the
“perceptio” is not thinkable unless the object has the “appetitus”
to receive the external influence. Glisson’s doctrine of irritability
is based on this conception, which he develops in a second work
already begun before the “Tractatus de natura substantiæ,” but
not finished until later and only published after his death. In
this “Tractatus de ventriculo et intestinis,”2 Glisson dwells in
detail on the physiological properties of animal structures and
develops for the first time his conception of irritability in the
chapter “De irritabilitate fibrarum.” The “irritability” manifests
itself in the appearance of the alteration of movement, which is
brought about by external influences on the animal structure, for:
“Motiva fibrarum facultas nisi irritabilis foret, vel, perpetuo
quiesceret, vel perpetuo idem ageret.” The fundamental factor of
this irritability Glisson attributes to the “perceptio,” which he distinguishes
as a “perceptio naturalis, sensitiva and animalis.”
The want of clearness produced here by Glisson’s artificial distinctions
and mode of expression is in part removed if we endeavor4
to transfer his meaning into our present methods of thought.
This distinction would then simply point out the different means
by which the stimuli can reach the irritable structures. The “Perceptio
naturalis” is that which today we should call “direct
response” to stimulation, that is, the excitation of the fiber by
artificial stimuli applied directly to the tissue. Glisson shows
here, that the intestines and muscles in the body immediately
after death and even when removed from the body can be
stimulated to movement by means of corrosive fluids or cold.
The “Perceptio sensitiva” is, according to Glisson, the excitation
of the fibers by external stimuli which act on the intact body as
a whole by way of the sensory nerves. The “Perceptio ab appetitu
animali regulata” finally is the excitation by inner stimuli
proceeding from the brain. The Perceptio naturalis is possessed
by all parts of the body, even the fluids, the bones and the fat.
All of them are irritable. But a “vitale” and a special “animal”
irritability they do not possess to a perceptible degree. These
forms of irritability belong only to the special parts of the body.
Here, however, the distinctions made by Glisson, are quite vague
and contradictory. In his “Tractatus de ventriculo et intestinis”
Glisson sharply distinguishes the “sensatio” from the “perceptio.”
The perceptio in itself is not a sensation, for although individual
organs of the body are irritable, as they all possess a “perceptio,”
they are not in themselves sensitive. The “sensatio,” the sensation,
only arises when the external “perceptio” of the individual
organs combine through the nerves with the internal “perceptio”
of the brain. “Nisi enim percepto externa ab interna simul percipiatur,
non est cognitio sensitiva completa.” Sensitivity is,
therefore, a special faculty, that is only based upon irritability.
I have treated the views of Glisson somewhat in detail for
on the one hand this seemed to me to be only due to the founder
of the doctrine of irritability, and on the other we have
here for the first time, although in somewhat vague and little
worked out form, the discovery of a general property of all
living substance, and its fundamental importance for the life
of the organisms. One might, therefore, in a certain sense, date
from Glisson the beginning of general physiology, and all the5
more so, because Glisson from the very first connected the irritability
of the living substance through its possessing universal
energy with the phenomena in nature generally, just as we do
today two hundred years after, on the basis of the modern teachings
of energy.
It might appear strange that a teaching of such fundamental
importance as that of Glisson’s theory of irritability was not at
once accepted on all sides and further developed. There were
two reasons, however, which prevented this. Firstly, Glisson did
not devote himself to his post of teacher at the University of
Cambridge with any particular zeal and so consequently did not
establish a school of his own, to further work out and develop
his ideas. Secondly, his doctrines were so speculative and difficult
to understand, his differentiations and definitions so artificial
and labored, that it required the greatest effort to penetrate to
his fundamental conceptions and so it happened that Glisson’s
theory of irritability received attention only at a comparatively
late date. Even then, of his speculative theories hardly more than
the name “doctrine of irritability” was adopted. Since the middle
of the eighteenth century this name, however, was destined to lead
to excited controversies.
The first attempt to give Glisson’s expression “irritability” a
more concrete meaning was made by Haller (1708–1777)3.
Unfortunately, though, he confined this conception solely to
muscles, in that he understood by the term irritability “the capability
of the muscles to contract, when stimulated, as the result
of vital force (vi viva).” He, therefore, applied the term “irritability”
to that which we today refer to as “contractility.”
On the other hand he applied the term contractility solely to a
property possessed by other living and dead animal as well as
vegetable matter, elasticity, that is, the capability to resume its
original form after distortion. He makes a sharp distinction
between “irritability,” which manifests itself by a contraction of
the muscles after stimulation by its own vital force (vi viva),
and the “sensitivity,” which is possessed only by the nervous6
system. “Sola fibra muscularis contrahitur vi viva; sentit solus
nervus et quæ nervos acciperunt animales partes.” By confining
the conception of irritability to a single living substance, the
muscle, Haller’s theory represents a great regression in comparison
to the correct fundamental thoughts of Glisson. This unfortunate
use of the term of “irritability,” “contractility” and
“sensitivity” has opened wide the gates to confusion and misunderstanding.
This confusion was still further augmented by
the fact that the vitalistic school of Montpelier confused the idea
of vital force with that of irritability. In the works of Bordeu
(1722–1776) these views are comparatively clear, if one bears
in mind that he substitutes Glisson’s term of “irritability” with
that of “sensitivity.” He assumes a “sensibilité générale” or a
common property of all living structures, both solid and fluid.
Besides this, each different part has according to him its “sensibilité
propre.” Here in place of the clear conception of irritability
we find one of more or less mythical nature possessing
traces of Stahl’s “anima.” Nevertheless we observe here the
idea that all living organisms possess in common a capability to
respond to stimuli. Even though Bordeu’s differentiation of the
“sensibilité propre” and the “sensibilité générale” is too artificial
and the coexistence of both not justifiable, his discussion of the
“sensibilité propre” shows that he is already on the track of the
characteristics of the effect of stimuli which only later under the
name of “specific energy” was clearly recognized as a fundamental
property of all living substance. On the other hand the
celebrated pupil of Bordeu, Barthez (1734–1806), accepted the
existence of a meaningless vital principle, the “principe vitale,”
governing all vital manifestations. The two forms of vital force
of all living substances, the “forces sensitives” and the “forces
motrices,” were according to his views manifestations of this
vital principle. He differentiates the “force sensitive” into a
“sensibilité avec perception” and “sensibilité sans perception,”
using the term sensibility in the sense adopted by Bordeu and
which today we, with Glisson, call irritability.
In this way serious thinkers of that time trifled with the words
irritability, sensitivity, contractility, perception. This led to7
futile conceptions, which equalled the phantasies of the worst
period of speculative philosophy and which in no way led to
progress. Hence it is easy to understand that numerous attempts
were made in those days to reconcile in some way these different
conceptions. An explanation, which was the beginning of further
development, came from England in the works of John
Brown (1735–1788),4 a man who was as talented as he was dissolute.
Brown was an independent thinker, not without genius,
whose knowledge in practice and theory, however, was limited.
This combination in his mentality enabled him to observe the
problems somewhat differently than through the glasses of the
usual conceptions of that time. In direct opposition to his teacher
Cullen (1712–1790), one of the leading minds in the medical
school of Edinburgh, who considered irritability only as an
effect of sensibility and pronounced the latter a specific property
of the nervous system, Brown took the standpoint that all living
substance, vegetable as well as animal, in contrast to lifeless
matter, possessed a fundamental property which he designated
as excitability, that is to say, the capability of being stimulated
to specific vital manifestations through external factors or
“stimuli,” in which sensitivity and indeed all mental processes as
well as movement are interpreted as specific effects, which the
“stimuli” produce on the irritable organs. This was an important
advance and from a wilderness of trifling conceptions his observations
led to a clearer knowledge of this subject. But Brown
went even further. In his so-called “theory of irritation,” he
has presented a whole system of responsivity to stimulation, which
in the first chapters of his chief work he expounds with wonderful
clearness. The fundamental principles here established must
be accepted even today. The essential basis of this “theory of
irritability” which he worked out especially for his doctrine of
disease, and which has also played an important part in pathology,
is the following: Every living, that is, excitable system, is continually
influenced by stimuli. The stimuli consist of either external
factors, such as heat, food, foreign matter, poisons, etc.,
or inner factors which result from the influence of the activity8
of one organ upon another. Only as a result of the continual
action of stimuli is life maintained, in that the stimuli produce
continual “excitement” in the irritable substance. The degree of
irritability differs in various plants, animals, in different structures
of the body, and even in the same individual at different
times under different circumstances. The strength of the
“excitement” depends on the one hand upon the degree of irritability,
and on the other upon the strength of the stimulus. The
irritability itself is influenced and changed by the action of the
stimuli. If the stimuli are too strong and are of prolonged duration,
the irritability diminishes as a result of exhaustion; if weak
stimuli act during a prolonged time, the irritability increases.
The healthy organism has a mean degree of irritability. Disease
occurs when this state is altered by strong stimuli or by an
absence of stimulation. Disease and health, therefore, differ not
qualitatively but only quantitatively. It is here seen that we have
the first attempt at a systematic interpretation of the effects of
stimulation, and it is astonishing how sharply and successfully
Brown has pointed out the foundations of this important field.
He has in this way not only amply compensated for the great setback
in the history of the teaching of irritability produced by the
confusions of conceptions created by Haller and the vitalists, but
also placed the whole of the physiology of stimulation on a firm
foundation upon which it is possible to build further. Though it
is true that many of his special theories, in particular those on
nature and the origin of disease, are quite erroneous, still a just
critic must judge work in relation to the period in which it was
written, and I question if at the present day the science of medicine
does not contain teachings which in a hundred years will
also prove untenable.
Johannes Müller (1801–1858) then added an important stone
to the building up of our knowledge of irritability. This was the
clear recognition of the specific energy of living substances. We
have already found the germ in Bordeu’s term “sensibilité propre”
or “sensibilité particulière.” Brown was also of the opinion that
different living objects possessed different types of irritability
and that excitation of their special functions was not dependent9
upon the kind of stimulus acting upon them. Johannes Müller,
grasping the idea hidden in this presentation, transformed it into
a clear and fundamental conception. Already in the work written
in his early years treating of optical illusions he says:5 “It is
immaterial by which means the muscle is stimulated, whether
by galvanism, chemical agents, mechanical irritation, inner organic
stimuli or sympathetic response from quite different organs;
to every means by which it is stimulated and an effect produced,
it responds by movement. Movement is, therefore, the
effect and the energy of the muscle at the same time.” “Thus
it is throughout with all reactions in the organisms.” “The sensory
nerve, responding to any stimulus of whatever kind, has its
specific energy; pressure, friction, galvanism and inner organic
stimuli produce in nerves of sight that which is peculiar to them,
light sensation; in the nerves of hearing, that which is peculiar
to them, sound sensation; and in the nerves of touch, touch
sensations. On the other hand, everything which affects a secretory
organ produces change of the secretion; that which affects
the muscle, movement. Galvanism is not superior to any other
methods, of whatever kind, which can bring about stimulation.”
And in his handbook of physiology Johannes Müller6 formulates
the law of specific energy for the sensory structures briefly in the
following words: “The same external factor produces different
sensations in the different senses according to the nature of each
sense, namely, the sensation of the particular sensory nerves;
and the reverse: the characteristic sensations peculiar to every
sensory nerve can be produced by several internal and external
influences.” This doctrine of the specific energy of the sense
substance possesses an importance which extends far beyond the
domain of the physiology of stimulation, for it forms the basis
on which the whole theory of human knowledge must be built
up, no matter how it may be constructed in detail.
As Johannes Müller already clearly emphasizes, it is here not10
the question of a law confined to the sense substance, but one
that applies to all living substances. Every living substance has
its “specific energy,” that is, its characteristic vital phenomena
and this is produced by stimuli of the most varied kind. This
doctrine received an extension of inestimable value for its future
development by the great discovery of Schleiden, that the cell is
the elementary building stone of the plant organism. Subsequently
Schwann at the instigation of Schleiden made further
investigations and found that this discovery applied also to the
animal organism. Irritability having been recognized as a general
property of living substance, it followed that, after the foundation
of the cell doctrine, every cell must possess irritability and
have its own specific energy. It now became necessary to study
the manifestations of irritability of the cells in their specific form.
Strange to say, this was done at an earlier date in pathology than
in physiology. Indeed, since the time of Brown the study of irritability
was furthered far more by pathology than by physiology.
The chief reason for this is probably the great practical interest
that the investigation of disease possesses, Brown having already
quite correctly ascribed the existence of disease to the relations
of the organism or its parts to stimuli. Rudolph Virchow then,
after the establishment of the cell doctrine, arrived at the momentous
conclusion, that disease must be considered as reactions of
the body cells to stimuli. In his epoch-making “Cellular pathologie,”7
he has carried out this idea in a classical manner. By irritability
Virchow understands “a property of the cells, by virtue
of which they are set into activity, when affected by external
influences.” There are, however, various kinds of actions which
can be brought about by external influences. But essentially there
are three kinds. The effects produced are functional, nutritive,
formative. The result of excitation, or if one will, of stimulation
of a living part, can, therefore, according to circumstances, be
either merely a functional process, or there can be a more or less
intense nutritive activity produced without the function being
necessarily at the same time activated, or finally, it is possible11
that a process of formative change may occur which produces
new elements in greater or less numbers. Virchow touches here
for the first time upon a question of extraordinary moment, the
important bearings of which have only now begun to be recognized
and seriously considered. We now know, for example,
that the functional excitation can be separated to a certain degree
from the cytoplastic excitation of the muscle. If the muscle is
acted upon by functional stimuli, the excitation takes place mainly
in the form of functional metabolism, nitrogen-free substances
are broken down in increased quantities, whereas cytoplastic
metabolism, which produces more profound alteration in the living
substance, and which goes so far as to bring about a breaking
down and building up of the nitrogen containing atom groups, is
hardly at all increased. It would be an error, however, to look
upon these different kinds of metabolism as quite independent.
Considering the close correlation which all the phases of metabolism
bear to each other, this idea cannot well be entertained.
If, however, we question in what manner, for instance, the
functional and the cytoplastic metabolism are linked together,
we have a problem before us which does not belong to the past,
but to the present and future. Indeed, Virchow seems already to
have felt that a sharp division between the different phases
and parts of functional metabolism in the cell does not exist, for
he says: “It is true that it cannot be denied that, especially between
the nutritive and formative processes and likewise between
the functional and nutritive, intermediate gradations occur.” Still
they differ essentially in their characteristic action and in the
internal alterations which the stimulated part undergoes, depending
on whether it functionates, nourishes itself, or is the seat of
special growth. Disease consists of the influence of stimuli upon
these physiological processes. The law of the specific energy of
living substance is as clearly expressed in functional disease as it
is in the physiological effects of stimuli. The pathological disturbance
of function is purely quantitative, “nowhere is there a
qualitative divergence.” The function exists or it does not exist.
If it is present, it is either strengthened or weakened. This gives
the three fundamental forms of disturbance: absence, weakening12
and strengthening of the function. No function other than the
physiological, even under the greatest pathological alterations,
exists in any structure of the body. “The muscle does not perceive,
the nerve moves no bone, the cartilage does not think.”
In this way Virchow rediscovered in the domain of pathology
the law that his great teacher, Johannes Müller, had already
clearly established in the field of physiology. But this law can
no longer be applied to all pathological disturbances of the nutritive
and formative activities of the cell. Here processes occur
which do not consist of a quantitative change of the normal
phenomena, but in the appearance of wholly foreign states, as
in the case of amyloid degeneration or heteroplastic tumors.
The question today and for the future arises, therefore, as to
where the limits of the validity of the law of the specific energy
of living substances are to be placed, a question closely connected
with the other before mentioned, of the relations between
functional and cytoplastic metabolism.
By means of cell pathology Virchow has laid the foundations
upon which our modern medical attitude is built and which must
remain essentially forever the basis of all future medical thought.
Certain critics, lacking in appreciation of the interrelations
between things and ignoring the safer and established knowledge,
have considered, in view of the unfoldings of the researches on
immunity and of serum therapy, that the time of cell-pathology
was passed and must be replaced by the humoral-pathological
teaching. These ultramodern critics, however, have here completely
ignored the fact that, on the one hand, the life of our
body is built up from the life of all of the contained cells, for
life in our body exists only in the cells; and on the other, a fact
not considered by them is that the components of the body fluids
originate from vital activity of the cells either directly or indirectly.
No result, indeed, of present serology can alter in the
least degree the fact that every disease represents only a disturbance
of the physiological processes of cell life of the organism
and the harmony in their combined workings. Indeed the more
recent observations of serology and chemotherapy are so little
opposed to cell-pathology that they are in fact only possible when13
based on the latter. They are only comprehensible then from
the unfoldings of cellular pathology.
Until quite recently all those effects of external factors on the
living substance which consist in excitation, that is, in an increase
of their specific vital processes, have always stood in the foreground
of all researches and observations on irritability. It was
gradually, however, more and more recognized that the depressing
influence of stimuli played a great rôle in the vital process of
the organism. Brown was acquainted with exhaustion produced
by stimuli, and the discussion of “asthenic” diseases, in which the
irritability was reduced, occupied an important place in his pathology.
That, however, in the normal activities of the organism
such depression or lessening of vital manifestation could result
from the influence of stimulation, first became clear after the
brothers Weber8 in 1846 discovered the inhibitory effects of the
galvanic stimulation of the vagus upon the heart.
Since then the inhibitory processes in nerves have been frequently
investigated by Schiff (1823–1896), Goltz (1834–1901)
and others, who gave us a theory concerning the same. Only
a small number of inhibitory processes were known at that
time, as for instance the inhibition of the croak reflex of the
frog, or the inhibition of the grasp reflex during copulation
of these animals through skin stimuli, and a few other cases.
They regarded the inhibitory nervous processes as a special state,
of which the inhibition of the heart through the vagus was the
best illustration. Further, the Russian physiologist Setschenow
succeeded by directly stimulating certain parts of the central
nervous system, especially the optic lobes of the frog, in producing
inhibition. It was, therefore, frequently assumed, as Setschenow
did, that in the brain there exist special inhibitory centers, just
as there are motor centers. This view was later shown to be
untenable. It is only quite recently, and especially since Sherrington
has shown that inhibition plays a part in all antagonistic
muscle movements, that we have obtained a broad and more
thorough understanding of the inhibitory processes in the life14
of the organism, and a physiological explanation of this important
group of activities of the central nervous system. This inhibitory
effect of stimulation, brought about by the involvement of the
central nervous system in the normal organism, was studied side
by side with the depressing effects of stimulation. Claude Bernard
(1813–1878)9 first discovered that the excitation of all living
substance could be depressed or totally suspended through the
influence of certain anæsthetics, such as ether or chloroform.
By a series of experiments, as simple as they were convincing,
the French scientist showed that irritability could be depressed
in mimosa leaves, the growth of germinating plant seeds and the
ferment action of yeast cells stopped, likewise the disintegration
of the carbon dioxide in the cells of the green leaf, as well as the
development of the egg cells, and also the movements of the
animal organism and the sensations of man. By this means he
recognized that not only does all living protoplasm possess irritability,
but that it can also by means of certain substances be put
into the condition of “anæsthesia,” a state dependent upon a
change of the protoplasm, which he termed “semi-coagulation.”
Finally, besides the more apparent processes of excitation and
those less so, belonging to the group of inhibition and depression,
in the last century the knowledge of the subject was greatly increased
by the addition of another group, which recently in consequence
of various reasons has met with particular interest. These
being effects of stimuli on the direction of movements of motile
organisms, it became more and more recognized that these curious
manifestations of irritability, which appeared to have such a surprising
likeness to the mysterious attraction and repulsion in the
sphere of electricity and magnetism, occur universally in the vegetable
as well as in the animal world. These movements are of
the greatest biological importance for the obtaining of food,
propagation, protection against disease, etc. Botanists have long
known of the geotaxis of the roots and stems of plants, the
heliotaxis of their leaves and flowers and of the thigmotaxis
of their tendrils. Likewise the phototaxis of freely moving15
protistæ had been often observed, especially by Ehrenberg10 of
Berlin, well known for his researches on infusoria. Then
Engelmann, Pfeffer, Strassburger, Stahl, and many others
discovered and studied more carefully the facts concerning
chemotaxis, thigmotaxis, rheotaxis, geotaxis, phototaxis, etc.,
of bacteria, motile spores, rhizopoda, and so on. The question
arose if one should regard this singular behavior of the unicellular
organisms as an expression of conscious sensations, discrimination
or will. This view was as determinedly denied on the one hand
as it was accepted on the other. Whilst even today certain
scientists still consider the reactions of the unicellular organisms
as a manifestation of conscious sensation, discrimination or
will, others look upon them as unconscious reflex reactions of
cell organism, taking place as purely mechanically as the spinal
cord reflexes of vertebrates. This divergence of opinion would
have practically no value for the development of our knowledge
of irritability had not here, as in the case of the relations between
the mental and physical processes in man, the view been entertained
with more or less fervor, that at some stage or other in
the chain of the purely physiological processes of responsivity,
an intangible factor had been introduced which was considered
as the essential “cause” of the peculiar reactions to stimuli. It
is not here the place to enter into the question if, and in what
degree, animal psychology may be a field of scientific research.
Even if one looks upon conscious processes as effects of stimulation,
in both lower animals and in man, in no case should one
assume them to be factors of an essentially different nature,
interrupting the chain of the mechanical reactions; neither should
one consider the particular characteristic responses observed in
unicellular organisms as effects of non-mechanical “causes.” As
a result, a mysticism, in reality quite foreign to it, would be introduced
into physiology. As a matter of fact the physiological
investigations for the tropic reactions of stimuli, which have
been carried out in great number since the end of the eighties,
have shown more and more clearly that this peculiar behavior
of unicellular organisms towards unilateral stimuli is produced16
by a comparatively simple mechanism. The analysis of this
shows a difference in the intensity of the exciting or depressing
effect produced by the stimulus. The stimulus exerts its influence
unequally upon the specific activity of the motor elements of
different parts of the surface of the cell body. This difference
in response causes the axis of the freely moving organism to
assume a different direction in which to move. It is compelled
to move in a definite direction and so, in this field, the apparently
mysterious attraction and repulsion of living organisms toward
stimuli has, by means of the most simple analysis, been robbed
of its mystical character.
Finally, I should like to touch briefly upon a view of the irritability
of living substance which has recently been brought forward
by Semon.11 It assumes the proportions of a whole system
and is proclaimed as a basis for the comprehension of organic
phenomena. It originated with an idea which Hering12 developed
many years ago and which later was accepted by Haeckel,13
namely that heredity is a species of memory of the living substance.
Semon attributes to living substance, in contrast to non-living,
a “Mneme.” By “Mneme” he understands the capability
of living substance to assume, through the influence of a stimulus,
a permanently altered condition. The latent alteration resulting
from the stimulus he terms “Engramm.” These “Engramms”
can later, however, not only be activated by the reapplication of
the original stimulus, but also by other stimuli, so that the state
of excitation once brought about by the original stimulus reappears.
Semon calls the reproduction of the state of primary
excitation by a later stimulus “Ekphorie.” A great number of
other new word formations, such as “chronogene Engramme,”
“phasogene Ekphorie,” “mnemische Homophonie,” “mnemisches
Protomer” and countless others are supposed to serve for the
better understanding of a series of special facts, chiefly in the17
domain of the processes of heredity. That which is termed
“Mneme” and “Engramm” is not further analyzed. Semon
expressly declines to discuss the kind of alterations in which
the physical or chemical nature of an “Engramm” consists.
Hence physiological analysis has not been advanced in any
way by Semon’s new formation of words applied to long-known
facts. With a series of new expressions the originator
of the “Mneme doctrine” deceives himself, as well as a number of
his readers not endowed with the critical faculty, into supposing
that he has achieved a serious analysis. Of such, however, there
is not a trace. As can be conceived, this way of treating the
manifestations of life has met with no further attention from the
physiological side. For indeed, what physiologist would consider
that the fact of muscle responding by a contraction to an
induction shock, or to any other stimulus, is sufficiently analyzed
by the explanation that we have the “Ekphorie” of a state of
excitation that was once previously produced by an original
stimulus of some unknown kind, and of which the living substance
of the muscle, in consequence of its “Mneme,” has retained
a latent “Engramm”? Here the deep gulf is apparent which
exists between the demands of a physiological analysis and the
futile explanation of the mneme doctrine. Physiological investigation
must reject such a manner of treating its problems.
With this the history of the doctrine of irritability enters into
its present phase of development. To future research remains
then the problem of further analyzing irritability, this common
property of living substance, and finally rendering it into its simplest
chemical and physical components. This last goal can only
be approached very gradually, step by step. With the analysis of
irritability we shall investigate life itself. In the following lectures
it will be my endeavor to show how far, with our present
knowledge, we can penetrate by this path into the great secret.
18
CHAPTER II
THE NATURE OF STIMULATION
Contents: Principles of scientific knowledge and research. Origin and
meaning of the conception of cause. Cause and condition. Criticism
of the conception of cause. The conditional point of view. Conception
of cause. The conditional point of view applied to the investigation
of life. Conception of vital conditions. Definition of the
conception of stimulation.
The common problem of all scientific research is the investigation
and formulation of natural laws. The assumption of a
unity in the happenings and of existence in the world, in accordance
with definite laws, forms the indispensable foundation of all
scientific study and is fully justified by experience. Experience
has taught us, as a result of innumerable individual observations,
the existence of such an accordance, whereas in not a single
instance has it been shown that this is not the case. We are thus
justified in assuming without further discussion that every scientific
research, every new problem which we approach, is likewise
founded on this unity of occurrences in accordance with natural
laws. Only on the firm basis of this assumption has scientific
investigation a purpose, and every success is a new proof of this.
There is an unanimity of opinion concerning this among scientific
investigators in all fields.
Not such complete agreement, however, exists in regard to the
question by what symbols of human thought and speech these
laws can be described in part as well as in toto, so that existing
laws can not only be fully and conclusively defined, but at the
same time without the use of superfluous terms. According to
Ernst Mach, thought is an adaptation to facts. Our speech is19
simply a method of expression of our thoughts and indeed the
most satisfactory form we have. We must, therefore, use those
symbols which are most closely adapted to facts as the most
precise expression of these existing laws. What forms of
expression have we?
It might appear that a discussion of this fundamental question
has not a close connection with our special subject of physiology
of stimulation. This, however, is not the case. Indeed, it is an
irremissibly previous requirement not only for the elucidation,
but also for the understanding itself in this particular field. We
could not come to a clear understanding in this field without
such analysis. The interpretation of the unity of being and happenings
in accordance with natural laws, which today is widely
accepted in the scientific world as the only exact one, implies the
assumption of a “causation” according to which things are explained
by the law of “cause” and “effect.” I14 have already on
various occasions taken the opportunity to criticise this view and
to show the error and confusion to which it leads. I should like
here to enter somewhat more in detail into the reason for this
criticism. It is particularly directed against the scientific use of
the term “cause” on the basis of our best-known theoretical principles.
It is clear that all scientific observations and explanations
are founded on experience. Can it be said that the conception
of “cause” originates from experience?
We can say with absolute certainty that the conception of
cause dates from prehistoric times. Its beginning reaches back
to the stone age, at least to neolithic, possibly to palæolithic culture.
This is demonstrated by the careful reconstruction of these
prehistoric races based on a critical comparison of the remains
of their culture with that of primitive races living today. The
ideas of these primitive races show an inclination to an extraordinary20
degree to explain all happenings in the world anthropomorphously.
All happenings in surrounding nature are given
the same origin as the activities of man himself. To man, on
this plane of phantastic religious speculation, all events in nature
appear as acts of the will of invisible powers, which, having
originally proceeded from the souls of dead human beings, think,
feel and act exactly as he does. This anthropomorphic conception
of the occurrences in the surrounding world is one of the many
conclusions which ensue from the supposition of an invisible
soul, which can be separated from the body. It was this conception
which gave the impetus for the transition of human
thought from the era of the naïvely practical to the era of the
theoretical spirit in that far removed age. In this anthropomorphic
transference of personal subjective impulses of will to
the objectively observed events of the surrounding world, lies
the origin of causal conception, which since then has been generally
used as the explanation of the happenings in the world.
One cannot assert that the formation of the conception of cause is
purely a product of experience, but rather a result of naïve speculation.
Even if a later evolution of human thought shows a continued
endeavor to dismantle the conception of cause of its primitive
trappings and to modernize, as it were, its outer appearance,
we still find today many inner components clinging to it, which
do not agree with the strict demands of critical scientific exactness,
demands which must particularly be made concerning a
conception which has been given such fundamental importance
in theoretical knowledge.
I wish to observe here, however, that the conception of cause,
even though more or less unconsciously so, is still the remains of
a part of the old anthropomorphic mysticism carried over into
our own times. This shows itself especially in the conception
of force, which is nothing more than a form of the conception
of cause. Force is the cause of movement. One has here in
anthropomorphic manner transferred the action of the will of
man, which produces movement of the muscles, into lifeless
nature. The force of the sun attracts the earth, that of the magnet
attracts iron, etc. In short, one has introduced a mysterious21
unknown factor instead of being content with the simple description
of facts, such as Kirchhoff15 has advanced in the field of
mechanics. Although of late natural science has also dispensed
more and more with conception of force as a means of explanation,
it is still today not wholly done away with. That which
applies to the conception of force is likewise true of the conception
of cause.
Another point concerning the application of the conception of
cause seems to me, however, to be of much more importance,
namely that a single cause is held responsible for the taking
place of a process. One endeavors to explain a process in general
by seeking for its “cause.” The cause being found, the
process is considered as fully accounted for. This idea is not
only widely spread in everyday life, but is even found frequently
in natural science, especially in biology, although here, it should
be known, the processes are decidedly more complicated. The
search for the “cause” of development, for the “cause” of heredity,
for the “cause” of death, for the “cause” of the respiration,
for the “cause” of the heart beat, for the “cause” of sleep, for
the “cause” of disease, etc., was for a long time and frequently
even today a characteristic of biological investigation. As if
such a complicated process as development, death or disease
could be explained by a single factor! In reality, one has
obtained very little as a result of the analysis of a process by
discovering its cause; and in addition the false impression arises
that through the finding of this one factor the process has been
definitely explained. It has been generally recognized in the
natural sciences in recent times that no process in the world
is dependent upon one single factor and attempts have been made
to give this fact more consideration.
It is the custom at the present time to hold the view that every
process or state is brought about by its cause, but that a series of
conditions are also necessary to the production of the process.
Such a view, however, which considers that two different factors
existing at the same time are necessary to the accomplishment of22
every happening or state, namely, the cause and the conditions,
leads to new difficulties, for then, upon a more exact analysis
arises the question: Which is the cause and what are the conditions?
It is very soon found, however, that this does not permit
of any strict differentiation, as the two conceptions can not be
sharply separated. This difficulty was brought to my notice with
particular force during an animated discussion with a friend
and colleague about twenty years ago, which I have always
remembered. I had observed at that time the dependence of
pseudopod formation of amœboid cells on the oxygen of the
medium, and had found that the expansion phase of protoplasmic
movement, that is, the extension of pseudopods, the
centrifugal flowing of the protoplasm into the surrounding
medium and with this the enlargement of the surface of the cell
body, only takes place when oxygen is contained in the surrounding
medium and never occurs in its absence. Being at that
time wholly under the influence of the conception of cause, I
believed that oxygen was the cause of the formation of the
pseudopods. To this my friend made the objection: “Yes, I
quite acknowledge the fact of the dependence of the formation
of pseudopods on oxygen, but what informs me that the oxygen
is really the cause? It might be simply a necessary condition.”
This objection led to a long debate, which ended, however, without
our being able to agree. We were not in a position to distinguish
between the conception of cause and that of condition,
and at that time the idea did not occur to us to emancipate
ourselves from the conception of cause deeply implanted in us
as a result of our training. In fact, one is greatly embarrassed
if one attempts to sharply distinguish by a definition the conception
of cause and that of condition. A condition is a factor on
which a state or a process is dependent for its existence or its
taking place. To the conception of condition belongs, besides
the factor of relation, that of necessity. Every condition is necessary
to the existence or taking place of this state or process.
Without the condition in question the state or process does not
occur. The same must be demanded for the conception of cause.
No state exists, no process takes place, without its cause. The23
cause then has itself the specific character of a condition, it is
itself a condition. Has it perhaps then some specific peculiarity
in contrast to the other conditions, which would give it a prominent
place? Experience teaches us that nothing, that is to say,
no state or process in the world, is dependent upon a single factor
alone. There are always numerous factors which bring about
the state or process. Would it be possible to distinguish which
of these particular conditions is of the greatest importance?
First of all, it must here be taken into consideration that the
importance of a condition is not one which is capable of increase
or decrease, for the simple reason that necessity, which forms
an essential component of the conception of cause cannot be
varied. A factor cannot be more than necessary for the existence
of a state or the taking place of a process. If, however,
it is less than necessary, then it is not necessary at all, and
the state or process exists also without it, that is to say, the
factor is not a condition. In other words: all conditions for a
state or process are of equal value for its existence, as they are
all necessary.
If one attempts to prove by means of concrete examples this
statement obtained by purely logical deduction—a control which,
considering the experimental nature of modern thought, never
should be neglected even in the simplest of reasoning—it might
appear that an objection could still be made against its general
validity. From various instances it might be concluded that
there are conditions, which as such are not absolutely necessary
for a state or process, but can be replaced by other factors. An
example may serve to make this clear. I pour diluted hydrochloric
acid on powdered carbonate of sodium, and carbon dioxide
is set free. The addition of hydrochloric acid is here a condition
for the liberation of the carbon dioxide. Without the presence of
the hydrochloric acid the process does not occur. Nevertheless
I can substitute diluted sulphuric acid for the hydrochloric acid.
Here it would appear that one condition can be replaced by
another. But one must not be deceived. A closer observation
soon shows that the process has not been sufficiently analyzed
if we look upon the addition of hydrochloric acid as a condition24
for the liberation of carbon dioxide. It is not the presence of
hydrochloric acid or sulphuric acid, as such, which is a condition
for the process, but rather the separation of the sodium atoms
from their combinations with the oxygen in the molecule of the
carbonate. This reaction can occur as a partial component in
very different complexes of processes. Or to quote another
example, taken from the subject with which we are especially here
concerned. I allow an induction shock to act on the nerve of
a nerve muscle preparation of the frog. The muscle contracts.
The electric stimulus is the condition for the muscle contraction.
But I can substitute for the induction shock a mechanical
stimulus by sudden pressure of the nerve. The muscle again
contracts. The analysis again shows that the induction shock as
such was not the condition for the muscle contraction, but the
excitation of the nerve which it produced and which is conducted
as a specific impulse to the muscle. This excitation of the nerve
can, however, be induced by very different kinds of processes,
namely, by all processes which possess in common the condition
that they suddenly increase certain disintegration processes in the
living nerve substance. Indeed, the further analysis of the whole
process shows in addition that the nerve impulse as such likewise
does not form a condition for the contraction of the muscle, but
it first of all produces the necessary condition for the muscle
contraction by suddenly greatly increasing certain chemical processes,
which take place in the living substance of the resting
muscle. The nerve impulse can, therefore, also be replaced by
other processes, if only these contain the condition for an increase
of disintegration of the muscle substance, as in the case of the
direct stimulation of the curarized muscle, where the influence
of nervous impulses is totally eliminated. In a further analysis
of this process we should penetrate even more deeply into the
differentiation of the individual constituent processes and the
isolating of the special conditions on which each link in the chain
is dependent.
Such an analysis then shows us the following: Every thing,
every state or process, is a complex of numerous components, of
which one always conditions the other in the manner that the25
individual conditioning components are themselves in their turn
contained as constituents of other complexes and are conditioned
here again by other factors. These factors in themselves
as such are not directly necessary to the taking place or existing
of the special component and can, therefore, be replaced by
others. Closer observation shows that there is a constant interdependence
between all things in the world. Every thing in the
world is indirectly dependent upon every other, although often so
remotely that we are not able to trace the connection. Absolute
things, completely isolated and independent of others, do not exist
in the world. In observing and studying complexes individually,
we must not forget that we only think of them as isolated from
the great eternal coherence, from which they are in reality not
separated. The conception of condition, however, only then has
meaning, if we refer to it in connection with the direct dependence
of one factor upon another. Nevertheless if we understand
by conditions those which are connected by multitudinous intermediate
components, then we would render the conception of
conditions useless. For if every thing in the world were the
condition for every other, the conception of relation would lose
its value in special states or processes. Should the conception of
condition have a meaning in regard to a certain state or process,
then we should only look upon that part of a complex upon which
the other is directly dependent as a condition. When, however,
we meet with a factor for a process or state, which can apparently
be replaced by another factor, we have not carried the
analysis far enough. Upon deeper penetration into the subject,
it is found that the essential condition for the process, which
exists, is a component common to both factors, one of which in
consequence can replace the other.
It is the task of all scientific research to penetrate deeper and
deeper into these relations, these connections and the order of
succession of states and processes and to separate them into
their individual components, and in this way gain a more thorough
knowledge of the constancy of existence and happenings in
the world.
This analytical process, it is true, only advances very gradually,26
and we must accept for the present, especially in the complex
biological processes, that a whole complexity of members
appear conditioned, and that a complex aggregate is a condition
of the whole process. We are not yet in the position to
define the special components of the constituent processes. It
is only step by step that we are able to differentiate the necessary
from the accessory parts in these complexes. However, we are
here only concerned for the present with a purely theoretical
question and we may be permitted to say: If we maintain that
the conception of condition has as an integral part the element
of necessity and of relation to a special thing, then there are no
substituting conditions. For then every condition for a state or
process is of equal value. There is no justification to give more
prominence to one condition and place it in the position of being
the “cause.”
If the cause is elevated, then it is done from some superficial
motive. This is confirmed by a glance at the practical use of the
term cause. The cases in which the cause is always at once
clearly recognized and named without doubt or hesitation are
those where a new factor is added to an already existing system
of conditions, which bring about a process. When such a process
is produced, the last added condition is considered as “cause.”
A shock acts on an explosive body, the body explodes: the shock
is considered the cause. An induction shock acts on a muscle, the
muscle contracts; the induction shock is looked upon as the cause
of the muscle contraction. To regard only the last added condition
as being of especial importance to the taking place and the
explanation for a process is, however, a standpoint which could
satisfy only the most superficial of observers.
In a scientific investigation such methods should play no rôle.
For to every careful observer it must appear quite clear from the
beginning, that the previously existing conditions have as great
a value for the taking place of the process and its explanation as
that last added.
The induction shock would not have produced the characteristic
effect had not the other conditions been already previously
combined, had not certain special atoms in the molecule of the27
explosive combination in consequence of former processes
assumed quite a peculiar labile position, had not in the evolution
of the muscle in the growth and metabolism certain combinations
been formed, and certain chemical processes taken place.
Therefore if I do not analyze these previously existing processes
and the conditions brought about by them in the system
of the explosive substances or the muscle, and simply know the
condition added last, then I have learned nothing of the process
itself, have explained nothing. The time of application of a new
condition does not justify in any degree the assignment of a dominant
position to a factor. But more: in many cases there is not
a question at all of the addition of a process to an existing state,
but rather of the simultaneous interference of two or more processes.
Several conditions can appear at the same time. In other
cases the sequence of the combination can be reversed. Which
then is the cause? Has the process several causes, or has it no
cause? Here one sees plainly to what absurd results it leads if
time alone is used as a basis of the conception of cause. To
illustrate this I return to the case of the liberation of carbon
dioxide from carbonate of sodium. I place anhydrous carbonate
of sodium in a beaker and add hydrochloric acid. The carbon
dioxide escapes. Here the addition of hydrochloric acid would
be assumed to be the cause of the freeing of the gas. Then I put
hydrochloric acid in a beaker and add carbonate of sodium. The
same process takes place, but now the addition of carbonate of
sodium would be considered the cause for the formation of gas.
Now I put both simultaneously into a beaker. Again the same
process. Which was now the cause? Has the process now two
or has it no cause at all? Finally I put anhydrous carbonate of
sodium and hydrochloric acid in ether solution into the beaker.
The formation of gas does not take place, and yet both causes for
this formation of gas are present, the carbonate of sodium and the
hydrochloric acid. Only when I add water to the mixture does the
formation of carbon dioxide take place. Here water would be
considered the cause. Hence every condition would be in succession
the cause for one and the same process. Under some
circumstances the same process would have several and in others28
no cause at all. It is scarcely necessary for further comments
upon the value of the conception of cause for the scientific explanation
of a state or process. If we do not seek to introduce
into exact science the antiquated symbols which have become useless
and belong to a primitive phase of development of human
thought, there cannot be a moment’s doubt that a strict scientific
analysis in whatever field of investigation it may be carried on can
consist only in the study of all the conditions concerned in a state
or process. If this is done, then the work of exact research is
accomplished. Further problems do not exist. The use of superfluous
terms or symbols for the definition of things would be in
opposition to the fundamental principle, already brought forward
by Kirchhoff, especially for mechanics, namely, that of formulating
comprehensively and in the simplest manner the processes
which take place in nature.
At first glance one might be tempted to find an incompleteness
in the observation and description, when a conditional standpoint
is adopted. It might be thought that conditionalism were a
purely formal method of observation, and only considered the
interdependence of things, but not the properties, the nature of
the objects themselves. Regarded more closely, however, it is
seen that this objection does not hold good. For what is a
condition?
A condition is in itself a thing of quite distinct properties.
The properties of a thing are, however, determined by the specific
combination of conditions which characterize the thing. The
conditions by which a thing, that is to say, a state or process, is
determined, are identical with its being and nature; in other
words, they are the thing itself. Purely formal relations without
essence would be altogether an absurd fiction not in accord with
reality, and which even the science of mathematics does not
acknowledge, for we cannot have a conception without concrete
content, just as in nature we do not find a form existing independently
of a thing. Every thing is equal to the sum of all its
conditions and depending upon the uniform constancy in accordance
with natural laws is solely determined by its conditions.29
The problem of all scientific research consists wholly in the
ascertaining of the conditional interdependency.
A state or process is solely determined by the sum total of its
conditions. A state or process is identical with all of its conditions
in totality. From this it follows that equal states or processes
are always the expression of equal conditions and wherever
unequal conditions exist, unequal states or processes will
result; and further, a state or process is completely investigated
when the entire number of its conditions is ascertained.
This fundamental statement of conditionism should be engraved
over the portals to the entrance of every scientific investigation.
That there is not the least difficulty in presenting scientific
observations strictly according to these principles of conditionism,
and that one can perfectly well do without the causal conception
in a scientific description, I have shown by a concrete example,
namely, in the fifth edition of my “General Physiology.” In the
whole volume the conception of cause is only mentioned in one
place, where its theoretical value is criticised, elsewhere not at
all, and yet I do not think that any one will miss this conception,
and indeed, if their attention is not especially called to the fact,
even notice the omission.
These principles of an exact conditional investigation must also
guide us in the analysis of the processes of stimulation. The
process of stimulation is especially apt to tempt one to employ
the old conception of cause, for it belongs to that group of processes
which originate from an already existing system by the
addition of a new factor. An electric stimulus acts on the muscle.
The muscle contracts. The stimulus is considered the cause of
the contraction. But what would I explain if I were to prove
that the stimulation is the cause of the contraction?
The history of physiology shows us that this subject has advanced
long since far beyond the stage of being satisfied with
such an explanation. Today the process would only then be
fully investigated if we knew the entire number of its conditions
and had traced the dependency of the individual partial constituents
of the whole complex process upon one another. For30
this, however, it is essential that we study the conditions already
existent in the entire system previous to the action of the
stimulus.
That which we describe with the word life is an exceedingly
complex process. If we analyze life, it is found to be composed
of an immense number of separate constituent processes, each
one being conditioned by the others. These constituent processes
are the vital conditions. A vital process occurs, and must
occur, where and when the whole sum of vital conditions is
realized. It is identical with the sum total of the vital conditions.
If only one condition is absent, then life does not exist.
It is then expedient to reserve the expression “life” for the
entire sum of the vital conditions. When we speak of the individual
constituent processes as “vital processes” in the plural,
we must bear in mind that in reality each is not in itself life.
Only the whole complex “lives,” not an individual constituent
of the same. Living substance is rather the whole system, and
not a constituent part of the same, not a piece of protoplasm, not
a nucleus and not a specific protein combination in the cell.
A property of this system should receive our consideration at
this point. It is a characteristic of every system in the world,
namely, the fact that a system is not isolated from its surroundings.
It is a deception resulting from the selective action of our
sensory organs, if we consider the bodies as separated and isolated
from their environment. This deception disappears upon
further analysis and when we assist our organs of sense, which
only respond to certain parts of the whole process, by experimental
methods of investigation. Our experience then shows
us that an isolated system does not exist, but that there are
instead everywhere connections which extend further and further
into the infinity of the world. An organism is consequently
no delimitated system and the vital process cannot, therefore, be
sharply separated from the processes in the medium. We cannot
draw a sharp line between vital processes and say: on the
right we have factors which are necessary for the maintenance
of life, and on the left factors which are not necessary. The
conditional connection between individual processes extends to31
the entire world, and likewise a great series of constituents, each
influencing the others, extend from the medium into the organism.
The nature of our sense perception, and consequently the knowledge
derived therefrom, is such that we are obliged to arbitrarily
take into consideration merely fragments from the endless interdependence
of all things in the world, and so we separate the
vital conditions of the organisms from their surrounding factors,
as though they were independent. A conscientious theoretical
analysis requires that we should never forget that in reality such
an isolation does not exist. Only with the recognition of this
can we distinguish for practical purposes between internal and
external vital conditions. In such a differentiation the internal
vital conditions which compose the living system conceived to be
isolated, are the organs, the tissues, the cells, the protoplasm and
the cell nucleus, and within the protoplasm and the nucleus the
arrangement and quantitative relations of certain substances,
such as proteins, salts, water and the thousands of special components
with their interactions and continued alterations. On
the other hand, the external vital conditions, which act on the
periphery, are the conditions of the surrounding medium, as
foodstuffs, water, oxygen, static and osmotic pressure, temperature,
light, etc. But this distinction has only a practical value
for the study of the organism as an independent system. Theoretically
it is as impossible to make a sharp distinction between
internal and external vital conditions, as to distinguish between
the vital conditions generally and the more remote conditions of
the environment. All these conditions form a widely branching
system of factors of which one is conditioned by the other reaching
continually from the interior of the vital system into the
surrounding medium, so that on the periphery of the system it
cannot always be said whether or not a component still belongs
to life. Considering these circumstances we can roughly for the
present define the conception of stimulus as follows:
A stimulus is every change in the vital conditions.
The most essential point in this definition is the relation of
the conception of stimulus to that of vital conditions. These
relations, however, call for a brief explanation. Here again the32
conditional method of observation saves us from error, for it
would be wrong to place the conception of stimulus and vital
conditions in contrast to one another, one excluding the other.
On the other hand, this method of observation shows that the
stimuli are likewise only conditions, but conditions producing
certain changes in the vital system. If a stimulus acts, that is,
if there is any change whatever in the vital conditions, the whole
complex of life in consequence of the dependency of the constituent
parts upon each other is also changed, and a new state
of living substance occurs. Stimuli are, therefore, also only
vital conditions, but vital conditions for new vital manifestations.
The relation of one given state to another, forms an indispensable
point in the understanding of vital conditions as well as that of
the stimulus. The stimulus becomes a vital condition for the
new state which it produces. It is only a stimulus relatively to
the original state, which previously existed. The essential point,
therefore, in the conception of the stimulus is that of alteration.
An example will serve to make this clearer. If Amœba limax are
bred in a hay infusion they appear in countless masses. Observed
in water in a watch glass they show at first the well-known form
of Amœba proteus with short, broad, lobate pseudopods. (Figure 1, A.)
After a period of rest, however, they gradually assume the
characteristic elongated limax form. (Figure 1, B.) In this shape
they constantly move about. But if I add to the water only a
faint trace of diluted solution of caustic potash, the amœbæ first
assume the shape of a ball (Figure 1, C), and then after a time,
stretch out long, pointed pseudopods, which give them the characteristic
form of Amœba radiosa. I have observed them for several
hours at a time. (Figure 1, D and E.) They
remain permanently16 in this form. They move in the same manner as Amœba
radiosa. They draw in one pseudopod, stretch out another and
float freely in the water in contrast to their limax state, in which
they are always attached to some support. The long, pointed,
often threadlike pseudopods, yield to every movement of the
water, bending in consequence like whipcords. In this example34
the amœbæ under the vital conditions existing in tap water have
limax form. The vital conditions undergo a change by the addition
of a solution of caustic potash, which acts as a stimulus. The
consequence is a reaction, in which the animal assumes radiosa
form. By the action of the stimulus a new state of the living
substance is produced, and remains as long as the solution of
caustic potash is contained in the medium. The solution of
caustic potash is, therefore, a stimulus for the state of the vital
system, which is manifested in the limax form, whilst for the
state of the system which shows itself in the radiosa form, it is a
vital condition. If I place the amœbæ of the radiosa form once
again in tap water, they assume the proteus and then the limax
form. The withdrawal of the solution of caustic potash, the presence
of which is a vital condition for the radiosa state, acts as a
stimulus, which results in a transition of the vital system to
another state. By altering the medium I can at will bring about
this change of form in the same individuals. In this way one and
the same factor can figure as stimulus and vital condition, according
to the state of the vital system on which it acts. Whilst its
addition acts as stimulus in the one state, its withdrawal acts as a
stimulus in the other state, which it has produced. The same
fact is shown by the well-known example of Artemia salina, which
on being placed in fresh water changes into Branchipus stagnalis
and, when again introduced into sea water, becomes once more
Artemia salina.
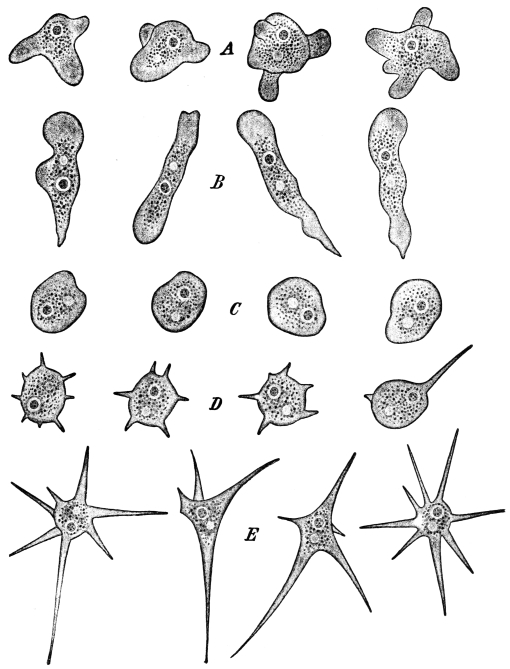
These facts show clearly that some stimuli can also be considered
as vital conditions. In the absence of certain stimuli,
life could not exist for any length of time. In the highly differentiated
cell community of the animal organism, for instance,
as a result of the coexistence of the cells and the tissues, many
parts have forfeited in a measure their independence. An
example of this is the skeletal muscle, which, in the absence of
impulses from the nervous system, reaches a low level of chemical
change and energy transformation. Here the nervous impulses
which act as momentary stimuli, are also in the course of time
indispensable vital conditions. Without them the muscle would
gradually become atrophied from inactivity. The same applies35
to all other tissues of our bodies. The functional stimuli are for
them at the same time vital conditions. These vital conditions
undergo fluctuations and interruptions but at each alteration
from a given state they act as stimuli.
Stimulus is every change in the vital conditions. But is this
definition complete? Are we really justified in regarding every
alteration in the vital conditions as a stimulus?
In considering this question, one point must not be omitted.
This is the fact that one of the chief characteristics of the vital
process is, that it undergoes continuous change. A vital process
involves not simply an alteration in metabolism or transformation
of energy in the sense that the same chemical processes continuously
reoccur in the same manner. Such a view could only
be admissible for the observation of living substance during a
limited period. An investigation over a long period of time
shows rather that every living system alters as long as it exists,
although this alteration is very gradual. The constituent processes,
in short, continuously undergo metabolic change both
quantitative and qualitative in nature.
If we observe the occurrences in a living system at various
moments of the cycle of life, we will find that the condition
differs qualitatively at each period. The progressive alteration
of the system is such that every state of living substance conditions
another, by which it is followed. No state can permanently
exist as such. Every state is the product of the preceding,
as it in turn conditions its successor. Consequently the
relations of the system to the surrounding medium also undergo
alteration, even when the external factors themselves in no
way alter. That which today is still a vital condition, is not
in consequence necessarily one tomorrow. These progressive
changes exist continuously until the death of the system takes
place. They characterize life. It is development, and life
cannot exist without development. Death is only the last phase
of development. The individual constituent processes of metabolism
gradually change to such a degree that they can no longer
work harmoniously together. Then the chain of processes is
interrupted at one point or another. The system develops into36
death or, on the other hand—and this, as Weissman especially
emphasizes, is realized in the case of unicellular organisms—a
corrective process takes place, a process of cell division by which
the original state of the cell is restored and development begins
anew and in a similar manner.
Ought we to designate these constant alterations in the inner
vital conditions as “stimuli”? Usage in this connection has
already answered in the negative, by applying to them the word
“development.” And this use is in a certain sense justified.
Let us imagine an organism or any other object for the purpose
of investigation as isolated from its surroundings. This conception,
which we have already stated, proves untenable on closer
analysis, but it, however, is based on the nature of the methods
of human observation and is indispensable for practical use
within certain limits. Then the inner vital conditions belong to
the organism, the external to the medium. They differ in so far
that the external vital conditions can exist permanently without
alteration, that is, independently of the development of living
systems, whilst the inner vital conditions of every living organism
continuously and progressively undergo alteration. In this sense,
but only in this, there is evidently a difference between the inner
and outer vital conditions, which permits a separation of the two
groups. But we should always bear in mind that this separation
cannot be sharply defined. On the same basis we assume that
the organism for purposes of study is separated from its surroundings
as an independent system, which leads us in consequence
to contrast the alterations in the internal with those in
the external vital conditions, in which we designate the first as
processes of development, the latter as stimuli. This distinction,
as all differentiations and separations in nature, gives us only a
practical working basis.
In this way we confine the conception of the stimulus to all
alterations in the external vital conditions of a living system,
considered as isolated. This view does not exclude the fact
that stimuli can also occur and act within an organism. If a nervous
impulse is conducted from the cerebral cortex through the
pyramidal tract to a skeletal muscle, this impulse acts upon the37
muscle cells as a stimulus. Although the explosion of the impulse
is an alteration within the body, nevertheless, as far as the
muscle is concerned, it may be looked upon as an external vital
condition, therefore as a stimulus. As the conception of stimulus
involves the relation to a given state, it likewise involves at the
same time the relation to a given living system, upon which it acts
from the exterior.
What is the value then of all this theoretical discussion?
In presenting the conception of stimulation from a conditional
standpoint, I desired to show what difficulties stand in the way
of a theoretical isolation of a fundamental conception in the field
of physiology, which indeed is used in our practical research
work at every step. “Natura non facit saltus.” I wished to
demonstrate that the sharp separation of the conception of stimulation,
like all artificial divisions which we make in nature, must
always contain an arbitrary note, as in reality isolated systems
do not exist in the world. I wished to show that, for this reason,
the conception of vital system, the conception of life, the conception
of vital conditions are not sharply defined. I wished
likewise to show that as a necessary consequence of this fact
a sharp separation of the conception of stimulation, which can
only be made in relation to that of vital conditions, cannot be
maintained theoretically. I wished to show further that there
is no sharp line of division between inner and outer vital conditions,
and that we cannot, therefore, make a strictly theoretical
distinction between the conception of stimulation and that of
the processes of development. I wished to show that, for these
reasons, we must not expect from the conception of stimulation,
as we understand it, anything beyond its possibilities. But finally
I wished also to show that, whilst fully conscious of and with
due consideration of all these difficulties, it is possible to work
out a definition of stimulation which is of great practical working
value. The definition in short is: “Stimulus is every alteration
in the external vital conditions.”
This definition gives to the conception of stimulation its most
complete, that is to say, its generally applicable and simplest
form. The great importance from a methodical standpoint of38
this definition of stimulation for the research of life is evident.
Our whole experimental natural science always employs for
investigation of any state or process the same method: the state
or process to be observed is studied under systematically altered
conditions. By stimulating the living substance it is brought
under changed external conditions. A systematic employment
of stimulus is, therefore, the experimental means for the research
of life.
39
CHAPTER III
THE CHARACTERISTICS OF STIMULI
Contents: The quality of the stimulus. Positive and negative alterations
of the factors which act as vital conditions. Extent of the alteration
in vital conditions or intensity of the stimulus. Threshold stimuli,
sub-threshold, submaximal, maximal and supermaximal intensities of
stimulus. Relations between the intensity of stimulus and the amount
of response. The Weber and Fechner law. All or none law. Time
relations of the course of the stimulus. Form of individual stimulus.
Absolute and relative rapidity in the course of the stimulus. Duration
of the stimulus after reaching its highest point. Adaptation to persistent
stimuli. Series of individual stimuli. Rhythmical stimuli.
The Nernst law.
We have found that stimuli are alterations in the external vital
conditions and that the irritability of living substance consists in
the capability to respond to stimuli by changes of the vital processes.
It now behooves us in the interest of experimental
research to investigate the relations between the nature of the
alterations in the external vital conditions on the one hand, and
that of the alterations of the vital process on the other; that is to
say, to systematically study the effects of stimulation on the living
organism. For this purpose it is above all necessary to become
acquainted with the almost countless numbers of alterations
which take place in the external vital conditions of an organism,
and to create a systematic scheme of stimulation which differentiates
and presents in comprehensive order those various elementary
factors which, among the innumerable varieties of stimuli,
would prove effectual. For this purpose it is necessary to
select the various factors which are involved in an alteration of
the external vital conditions.
The first of these factors is the quality of the stimulus. The
external vital conditions are, in short, a series of chemical factors,
such as foodstuffs, water and oxygen; the presence of a certain40
temperature; the existence of a certain light intensity; the
existence of a definite static pressure; and finally the presence of
an equal osmotic pressure. The stimulus according to its quality
can be differentiated into chemical, thermal, photic, mechanical
and osmotic varieties. To these must be added other forms of
stimuli not ordinarily operative, for instance, many uncommon
chemicals, and certain kinds of rays. The form of stimulation,
par excellence, which has acquired the greatest importance for the
experimental investigation of life, is electricity. In its manifold
forms it permits, as no other, of such fine gradations of intensity
and duration that it has become in the hand of the physiologist
an invaluable means of research.
Alterations in those factors which act as vital conditions compose
the great mass of physiological stimuli which act continuously
on every living organism. The first point to be considered
in every alteration is its direction. The alterations produced by
stimuli may be of two different kinds, either positive or negative.
The quantity of foodstuffs, water or oxygen, in the surrounding
medium, can undergo an increase or diminution; as may the
temperature, intensity of light, the atmospheric and osmotic pressure.
The strength of the electric current, which may be applied,
can also be regulated. In accordance with the definition of stimulation
already referred to, we must consider these alterations,
whether negative or positive, as forms of stimulation. Now the
question arises: Is this point of view justifiable? Should one
also consider, for example, the lessening or total removal of a
vital condition as a stimulus? Should one consider the removal
of water or oxygen, cooling or darkening, as a stimulus? It has,
in point of fact, been occasionally attempted not to regard these
negative deviations as forms of stimuli. These observers permitted
themselves to be led by the dogma, that only that which
produces an excitation, that is, an increase of the processes in the
living substance, should be regarded as a stimulus. Such a limitation
of the conception of stimuli would only result from the
one-sided consideration of an all too limited circle of facts. Considered
from the point of view which results from a broader
range of experience, this narrow view becomes untenable.
41
In the first place it does not follow that only positive fluctuations
of a factor, acting as a vital condition, result in excitation
in the existing vital processes. The withdrawal of water produces
a diametrically opposite effect. A muscle, from which
water has been removed, if exposed to dry air or placed in a
hypertonic salt solution, shows violent excitation, which manifests
itself in great increase of irritability and development of fibrillary
contractions. The breaking of a constant current which has for
a long time flowed through a nerve or muscle also elicits a
momentary excitation. Further, the abrupt removal of light
may also bring about stimulation. To cite an example from
the physiology of the single cell, I should like to call to your
attention the interesting observations of Engelmann17 on the
Bacterium photometricum, of which he was the discoverer.
When the field containing these organisms is suddenly darkened,
all the individuals contained in the drop immediately dart forward
for some distance, at the same time, as is usually the case, quickly
rotating around their own axis, and then after a moment of
immobility, swim on quickly in another direction. An analogous
responsivity has also been shown by other single cell organisms,
as has been pointed out by several observers and especially by
Jennings.18 In all these cases the excitation was produced by a
lessening or total withdrawal of the factors which act as vital
conditions; and even those who take the standpoint that only such
factors are to be considered as stimuli which produce an exciting
effect, are compelled to regard these alterations as stimuli, in
spite of the fact that they are negative variations of external
vital conditions.
But further, the restriction of the term stimulation to those
alterations which increase the course of the changes in the living
substance involves the observer in still greater contradictions.
It can easily be shown that one and the same factor in one and
the same form of living substance has now an exciting, now a
depressing effect on the vital processes. This fact can be readily42
demonstrated19 by means of the infusoria Colpidium colpoda,
which can be grown without difficulty in a hay infusion. A
number of individuals in a drop of fluid may be placed in a
warm stage and observed under the microscope; one then sees
that at room temperature they swim about by moving their ciliary
processes at a definite rate. Now if the temperature is raised to
about 35° C., the ciliary movement becomes enormously increased.
The infusoria swim madly through the field of vision. They are
in a state of violent excitement. The increase has, therefore,
acted as a strong, exciting stimulus. But if one allows the temperature
to further increase only a few degrees the ciliary movements
are suddenly greatly retarded. The infusoria now swim
sluggishly through the field of vision and finally remain stationary.
In this case the increase in the temperature has had a depressing
effect. If the infusoria are not quickly removed, the depression
is followed by death. Should the increase in temperature be
regarded in the first instance as a stimulus, and not as such in the
second, in which the temperature rises only a few degrees higher?
Here the change in the vital conditions concerned is in both
instances positive. In all cases of overstimulation we are confronted
by the same question. Nevertheless it is not at all necessary
to refer to such strong or even life-endangering stimuli for
the observation of these conditions. In this connection I would
like to cite an even more striking instance and which is of special
interest for the understanding of the phenomena in nerve centers.
If the posterior spinal roots of a Rana temporara are severed, and
the eighth root stimulated with a faradic current, whilst the musculus
Gastrocnemius of the same side is connected with a writing
lever, one obtains, as Vészi20 has found, at the moment of the
beginning of stimulation a contraction of the muscle. The faradic
stimulus has, therefore, produced an excitation reflexly. If instead
of the eighth the ninth posterior root is stimulated, the result
obtained is also an excitation of the muscle. In this case, however,
the excitation in the form of a tetanic contraction lasts for43
some time, provided that the stimulation is not at once stopped. If
now during tetanic stimulation of the ninth root the eighth is at
the same time stimulated, with a strength of current equal to
that which previously brought about contraction of the muscle,
instead of an increase and a strengthening of contraction there
is, on the contrary, an inhibition which continues throughout the
time during the stimulation of the eighth root. If the stimulation
of the eighth root is discontinued, the tetanic response of the
ninth root reappears. If, on the other hand, the faradic stimulation
of the ninth root is interrupted and the eighth root now
again stimulated, one obtains once more, as in the beginning, with
each stimulation a contraction of the muscle. This fact is illustrated
by the accompanying tracings. (Figure 2.) In this investigation
undertaken in the Göttingen laboratory it was further
shown that a faradic current of the same strength and the same
frequency had at one time an augmenting, at another an inhibitory
effect, and these effects could be produced alternately at will.
Should the faradic current at one time be called a stimulus, at
another not? It is here clearly shown to what absurd consequences
it leads if the conception of stimulation is limited solely
to the cases in which an external factor has an exciting effect;44
and yet an immense number of instances of a like nature could
be cited to show the untenability of this view.
Fig. 2.
Lower thick line shows duration of stimulation of 9th root; upper thick line that of
8th root.
It follows from this, that it is altogether impracticable to define
the stimulus itself in relation to the nature of the effects which
the stimulus has upon the substances in the living system. One
can only appreciate the nature of stimulation in relation to the
vital conditions and without considering the nature of the action
of the stimuli on the living substance. It is true that every
stimulus is followed by an alteration in living processes, but this
is to be expected when one clearly understands the nature of
vital conditions. A stimulus is in all cases an alteration in vital
conditions and, in that each of the vital conditions is necessary for
the continuance of life, it follows of necessity that every alteration
in the vital conditions, so intimately connected with the
living processes, will also be followed by an alteration in the
processes occurring in the living system. In short, response is
produced. Nevertheless, a definite alteration of an external vital
condition, depending upon the state of other vital conditions,
that is, according to the state of living substance at the moment,
can produce quite opposite effects. Although it may appear
expedient to include in the conception of stimulation in given
instances, distinctions between stimuli according to the nature
of their effects upon the living substance, in all cases the conception
must under all circumstances be so formulated that it
comprises all alterations in the external vital conditions, either
positive or negative, that is to say, an increase or decrease, an
augmentation or diminution in those factors, acting as vital
conditions.
Besides the quality there is another highly important factor
to be considered in the study of every alteration in the living
process, namely, its amount. The chemical concentration of the
medium, temperature, amount of light, the static and osmotic
pressure may undergo more or less variation. The electric
stimulus can rise from zero to great intensity and from great
intensity can fall to zero. The extent of the alteration determines
the intensity of the stimulus. In relation to the intensity,
a differentiation of stimulation has been introduced, which is not45
dependent upon the absolute intensity of the stimulus, that is,
upon the extent of the alterations in the external vital conditions,
but the intensity of the response that can be observed. One
refers frequently to threshold stimulation, to stimulation beneath
the threshold, to submaximal, maximal and supermaximal stimulation.
Such a classification is in many ways very valuable. It
is not only of practical value for the establishment of definite
intensities of stimulation, but also for the study of the state of
irritability in the living organisms.
The threshold of stimulation furnishes roughly a standard for
the degree of irritability of a living system. The threshold value
of a stimulus is then that degree of intensity which is just sufficient
to bring about a perceptible response. The threshold of
stimulation is low, that is, the irritability is great, when the intensity
of the threshold stimulus is small; the threshold is high,
that is, the irritability of a system is small, if the intensity of the
threshold stimulus is great. All intensities of stimuli beneath the
threshold are sub-threshold stimuli. Here a point must not be
overlooked, which in older physiology did not generally meet
with sufficient attention. From the fact that the sub-threshold
stimuli produce no apparent effects, the wrong deduction must
not be made, that they have no effect whatsoever. The conception
of the threshold of stimulation originated in the field of
muscle physiology and that of the special senses. Here the indicator
of the response is, on the one hand, contraction of the
muscles, and on the other, conscious sensation. There was a
great temptation to consider the stimulus altogether ineffectual,
if it produced no conscious sensation or no contraction of the
muscle. Today with our finer and more sensitive indicators for
the study of the alterations in the living substance, we know in
reality that sub-threshold stimuli, which produce no apparent
effect in the living substance, can have an effect in reality.
I will call your attention later to the fact that these sub-threshold
stimuli play a very important rôle under certain conditions
in the activities of the central nervous system. It only
depends upon the sensitivity of our special senses, or the indicators
used for this purpose, as to whether the alterations can46
be observed or not. The conception of the threshold of stimulation,
therefore, has meaning only when used in relation to
a certain indicator. The threshold of the same living system
may be different for different indicators. When we use the
term threshold we must necessarily know the indicator employed
in its determination. The threshold stimulus produces only
barely perceptible effects. The amount of response in most
living substances increases with the intensity to a certain limit.
If this limit is reached, that is, if the response is maximal, the
stimulus of the weakest strength necessary to produce this result
is termed the maximal stimulus, whereas all intensities lying
between the threshold and the maximal stimulus are termed
submaximal stimuli. If the intensity of the stimulus is increased
above that of the maximal, the response, as in the case of the
muscle, does not increase, and therefore one could say that all
intensities above the maximal could also be called maximal
stimuli.
In realty, however, the response to stimuli of different intensities
is never equal, even though it may appear so, when measured
by an indicator, as for instance, the height of the maximal
muscle contractions. This is clearly shown, for example, when
the electrical stimulus is increased far beyond that intensity which
is necessary to produce maximal effect. Injury is thereby produced,
which is manifested, for instance, in the muscle contraction
by the nature of its course and also by its height. One is,
therefore, justified in a certain sense in calling the intensities of
the stimulus, which are above the value which barely produces
maximal contraction, “supermaximal stimuli,” notwithstanding
this is logically far from being a happy expression. The term
“maximal stimulus,” then, is limited to the intensity of the stimulus
which just produces a maximal effect. I wish to point out this
distinction between maximal and supermaximal stimulus, as there
is often a lack of clearness in the use of these terms.
In that the nomenclature of intensity of stimulation is based
upon the intensity of response, the question arises as to the relation
between the intensity of stimulus and the amount of response.
It is well known that this question has met in one special field47
of physiology with a very detailed and comprehensive treatment.
I allude to the teaching concerning sensation. Ernst Heinrich
Weber21 first called attention to the relation between increase in
sensation and that of the stimulus in the case of the sense of
touch. His observations, which have been formulated into
“Weber’s law,” have been the object of animated discussion. A
presentation of this law is the following: “The amount of pressure
necessary to produce a perceptible increase of sensation
always bears the same ratio to the amount of the stimulus already
applied.”
If in accordance with Ziehen22 we designate the relative
increase in pressure to that already applied, which is necessary
to produce a perceptible increase in sensation, as the threshold of
relative differentiation, we can formulate the law in the simplest
way thus: The relative threshold of differentiation is constant.
Fechner,23 who indeed attempted to apply this law, applicable to
the sense of pressure, to all the other special senses, has given
us a mathematical formula, based on the assumption that the just
perceptible increase of sensation has the same value at all levels.
By this assumption he was able to establish for the first time a
relation between the intensity of sensation and that of stimulus,
for it follows that “the sensation increases in intensity in arithmetical
progression, whereas the intensity of the stimulus increases
in geometrical progression.” From this Fechner has
worked out a psychophysical formula, which today is generally
termed the Fechner law. This is the law: The intensity of sensation
varies with the logarithm of the intensity of the stimulus.
Soon the Weber as well as the Fechner law had been extended
over the whole field of sensation and stimulation. In this connection
Preyer24 has formulated his “myophysical law,” which
states that there is the same relation between strength of stimulus
and the intensity of response of the muscle as is laid down by the48
Fechner law for stimulation and sensation. Pfeffer25 has found
that Weber’s law applied also to the relations of the chemotaxis
of bacteria, to the intensity of the chemical stimulus, and likewise
the attempt has been made to show that all living substances
respond in the manner laid down by the Weber-Fechner law.
Unfortunately the innumerable investigations in this field have
shown more and more clearly that it is not possible to formulate
a general mathematical law, which strictly fixes the relations of
the intensity of the stimulus and the intensity of response. Even
in the field of the physiology of the special senses many voices
have opposed the general application of the Weber and the
Fechner law. Lotze, G. Meissner, Dohrn, Hering, Biedermann
and Löwitt, Funke and numerous other investigators have already
demonstrated for some decades, partly by means of critical
inquiry, partly by experimentation, that these laws are not strictly
valid. Above all these experiments have shown that logarithmic
relations are not tenable and likewise are not applicable to very
strong stimuli. The assumption made by Fechner, that is, the
acceptance that all barely perceptible increases of sensation have
an equal value, has been set aside as incorrect, and with this his
mathematical formulation within those boundaries of intensity
of the stimulus, in which the Weber law has proven itself valid,
must also be abandoned. That which we can say today with certainty
concerning the relation between the intensity of stimulus
and the amount of response is as follows: A law generally applicable
to the relation between the strength of the stimulus and
the amount of response cannot be mathematically formulated.
For a great number of living systems the rule which holds for
the intensity of stimulation within certain boundaries is the following:
With increase of the intensity of stimulation the response
at first increases rapidly and later more and more slowly.
This rule of course only applies within the boundaries of the
intensity between the threshold of stimulation and maximal
stimulus. The interval, however, between these intensities varies49
considerably in different living substances. In this connection
there are several forms of living substance which call for our
special attention. In these the surprising condition seems to exist,
that the interval between the threshold and the maximal stimulus
is zero; that is, every stimulus which acts at all always produces
a maximal response. Bowditch26 first observed this behavior in
the frog’s heart and this has also been confirmed by Kronecker.27
The induction current produces, as Bowditch says, either a contraction
or nothing. If the former, it is the strongest contraction
which can be produced by an induction shock at the given
time. Here for the first time a constancy of response was discovered
which has been termed the all or none law. McWilliams28
has later verified the same fact for the mammalian heart.
Gotch29 has also arrived at the same conclusion in connection with
the nerve. He states that “the comparison of submaximal with
maximal responses shows that although there is an obvious difference
in the amount of E. M. F., there is little or no difference
between such time relations as the moment of commencement, the
moment of culmination of E. M. F. and the rate at which E. M. F.
disappears.” Further: “the rate of propagation of the excitatory
wave is the same whether this is maximal or submaximal.” He
likewise assumes that the “all or none law” is applicable to the
constituent fibers, and that the variations in the strength of
response with weak and strong stimulation are brought about
in the first instance by stimulation of a few, in the latter by
a greater number of fibers in the nerve trunk. The same conclusion
has been reached by Keith Lucas30 for the single cross-striated
fiber of the skeletal muscle, founded on the fact that50
by direct stimulation of a bundle of curarized muscle fibers,
the contraction only increases inconstantly and not regularly
with the increasing intensity of the stimulus. This is only
comprehensible if one takes into consideration that, with the
increasing intensity of the stimulus, a greater and greater number
of fibers are stimulated. Keith Lucas31 came to the same conclusion
in the case of the muscle stimulated indirectly through
the nerve. He, therefore, sees, because of the nature of the
response of the single muscle cell, no difference between heart
muscle and skeletal muscle. The “all or none law” applies to the
individual muscle cells of both kinds. The difference between
the heart and skeletal muscle, according to him, lies in the fact
that in the heart the individual muscle cells in their totality stand
together as conductors of excitation, whereas in the skeletal
muscle the individual muscle fibers are separated, as far as conduction
of excitation is concerned, by the sarcolemma. Finally,
the recent investigations of Vészi32 with strychnine poisoned ganglia
cells of the posterior horns of the spinal cord, have made it
appear probable that “the all or none law” can be applied likewise
to the individual ganglion cell. He draws this conclusion
not only from the fact that all reflex contractions of a muscle
of a strychninized frog are maximal, whether they are produced
by weak or strong stimuli, but also especially because of the
loss in the strychninized spinal cord of the capacity of the summation
of irritability. The normal spinal cord does not reflexly
respond at all to weak single stimuli, but responds to equally
weak faradic stimulation very readily. Therefore, the threshold
lies very high for the individual induction shock and very low
for faradic shocks. But these differences are equalized in the
strychninized frog. This seems intelligible, when we assume
that the strychninized cell responds to every stimulus, to which
it responds at all, to the maximal extent which is permitted at
that moment by its stored up energy, otherwise the excitation
would necessarily be summated by faradic stimulation.
51
Such are the instances to which one has up to the present
applied the “all or none law.” The question if, as a matter of
fact, such a condition has ever been realized in any living substance
has until now found no final answer. Most authors, who
accept the validity of the “all or none law” for certain living substances,
do so with a certain reserve and speak only of the possibility
or probability of such behavior. The subject has, however,
as will be shown later, a great and even vital interest in
another direction. For this reason I should prefer to postpone the
treatment of the same to a later occasion. Here I wish simply to
say, that if the “all or none law” is valid in a strict sense for
certain structures, then there exists no general constancy of the
relations of the intensity of the stimulation and the amount of
response, applicable to all living organisms.
We will now return from this digression concerning the relations
between the intensity of the stimulus and the response, to
the further characterization of the properties of the stimulus.
Besides the quality, the direction and the intensity of every alteration
in vital conditions, an equally important factor is the duration
of the alteration. The time relations, under which a deviation
of the external vital conditions takes place, present immense
and manifold variations in nature. In many cases the change
is very complicated, as for instance, the alteration of the static
pressure or the temperature under the influence of air or
water currents, the osmotic pressure or chemical factors in
diffusion currents, and the light intensity produced by the movement
of clouds. These very irregular alterations have practically
little interest for us. Here we are concerned rather with the
differentiation of the time alterations of the processes of the
simplest fundamental types, which are of importance in studying
the course of the reaction. For it is of such simple elements
that the complicated and irregular alterations of the above-mentioned
kinds are composed.
The simplest form of an individual change in the external
vital conditions would be a regular and constant alteration of
intensity which can be graphically represented as a straight line,
wherein the intensities are the ordinates and the time the abscissa.52
(Figure 3, A.) A regularly rising pressure would, for instance,
represent a stimulus in its simplest form. But such forms of
stimuli are only very rare in nature and are also experimentally
very difficult to produce. It is, for example, not easy to give the
electrical stimulus, so much used for experimental purposes, this
form. Fleichl and v. Kries have only accomplished this by means
of complicated apparatus. The usual form of the individual
stimulus is not a straight line, but a logarithmic curve. (Figure
3, B.) The alteration hardly ever progresses with equal rapidity
from its beginning until it reaches its highest point, but as a rule,
with decreasing rapidity. This is the usual course of alterations
of concentration, also of chemical and osmotic stimuli, of changes
of temperature and of electric stimulation.
The rapidity of alterations in vital conditions has quite an
important influence on the development of the response to stimulation.
It is well known that if a constant current, which reaches
its highest intensity rapidly, is permitted to act upon a muscle,
the effect differs from that following the application of a current
of the same intensity but in which this is reached very slowly. In
the first case there is a sudden strong twitch, in the second none
at all. In spite of this there can be no doubt whatever of the
current in the last case being effective. That the muscle is also
excited when the current is slowly increased is shown by the
contracture, which grows more and more plainly perceptible with
the increasing intensity of the current and in higher intensities
by the so-called Porret’s phenomenon, which consists in a curious
wave-like movement of the muscle-substance. In reference to53
the rapidity of the alterations in the factors which act as stimuli,
the behavior varies greatly. Many stimuli because of their nature
never have a steep ascent or descent of intensity, as, for instance,
alterations in the concentrations of soluble substances, that is,
chemical or osmotic stimuli; likewise temperature variations may
be mentioned. They always act relatively slowly. On the contrary
there are forms of stimuli which have now a rapid, now a
slow, ascent or descent of their intensity, such as the photic and
mechanical stimuli. Finally, there are other stimuli that nearly
always show a very abrupt change of intensity, such as the
electrical form.
The most important factor to be considered in producing the
response to variations of intensity, is not the absolute rapidity, but
rather the relative rapidity; that is, the rapidity in relation to the
characteristic rapidity of reaction of the particular living substance
concerned. The rapidity of the reaction to stimuli is very
different in various forms of living substance. On the one
hand, we have forms reacting very quickly, as the nerve and the
striated muscle; on the other, those which respond very slowly,
such as a great number of unicellular organisms. Between these
are a great number of living substances which, as far as the
rapidity of the reaction is concerned, occupy intermediate positions
of every varying degree. It is clear that the adequate
stimuli for slowly reacting substances must be those having also
a slow change of intensity; for quickly reacting, those having a
rapid change of intensity.33 If a nerve muscle preparation is
simulated with the single induction shock, the “break” as well as
the “make” shock has effect. But even here a difference is
noticeable. The “make” shock has a weaker effect than the
“break” shock. This difference is due to the difference of
abruptness in its course, which when the current is made is less
than that of opening, for, when the current is made, the ascent
of the primary current is retarded by the extra current flowing
in the opposite direction, whereas, when broken, with the fall
of the intensity of the primary current, the extra current in the54
primary coil flows in the same direction. In consequence of this
there is a perceptible difference in the rapidity of the alteration
of the “make” and “break” shocks. (Figure 4.)
Fig. 4.
Course of induction shocks. 1 and 2 make and break of the primary current.
11 and 21 make and break induction shocks. (After Hermann.)
Now slowly reacting forms of living substance, such as certain
foraminifera, in which the extended pseudopods are stimulated
with single induction shocks, the break as well as the make
shocks are wholly without effect, as both take place far too quickly
for the slow responsivity of these organisms. I have made such
observations on various forms of foraminifera of the Red Sea, on
Orbitolites, Amphistegina and others. The movement of granules
in the pseudopods is not influenced by the induction shocks in the
least. It also continues without interruption when the pseudopods
are extended. Even with the strongest induction shocks at my
disposal I could not induce them to contract; the faradic current,56
also, the intensity of which I found quite unbearable, remained
utterly without effect.34 These two extreme cases, the nerve and
the foraminifera, show plainly that the effect of a stimulus is not
produced by the absolute rapidity of the increase of intensity, but
is solely influenced by the relative rapidity of the same.
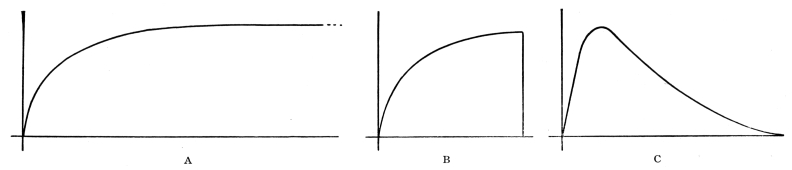
A further point for consideration in the duration of an alteration
in a vital condition in producing a stimulant action is the
length of time the stimulus remains after reaching its highest
point. In the forms of stimuli occurring in nature the duration
of the alteration after reaching its highest level can vary considerably.
The stimulus may remain indefinitely at a certain level,
when this is once reached. (Figure 5, A.) The alteration likewise
persists. This would be the case, for instance, with the
changes of concentration in the transfer of an organism from
fresh into sea water. The alteration can also, however, immediately
after attaining its highest level, return, so that the original
state is at once reestablished. (Figure 5, B and C.)
Here it is a case of a quick deviation in the external vital conditions. A
sudden jar would be a case in point. Between these two extremes
we have all variations in the duration of all natural and experimental
forms of single stimuli.
Now we arrive at the question: Has a prolonged stimulation
really a prolonged effect? This question might seem superfluous,
as from a conditional standpoint it is self-evident that every
alteration in any one of the conditions of a system is followed
by an alteration in the system. But this very question played an
important rôle in older physiology and led to prolonged discussions
for the reason that a special case was taken into consideration
in this connection, which at that time was not clearly understood.
Du Bois-Reymond,35 as a result of his investigations on
the nerve muscle preparation of the frog, formulated a law of
nerve excitation, according to which it is not the absolute value
of the intensity of the constant current which produces an excitation
of the nerve and contraction of its muscle, but an alteration57
of the intensity from one moment to another. The more rapidly
these changes are produced, the greater is the excitation. His
arguments were based upon the fact that a contraction can only
take place on the “making” or “breaking,” or by rapidly strengthening
or weakening the constant current; it is possible to subject
a nerve muscle preparation to a current of considerable strength
without a muscle contraction resulting, provided it is slowly
increased. One might be disposed to conclude from this that
the constant current, when showing no fluctuations, has no stimulating
effect whatsoever. Should this observation be carried
even further and the attempt made to extend it into a general law
of excitation by assuming that the effects of stimulation are only
produced by variations in the intensity, not by its continued
duration, one would commit the error of judging the occurrence
of a stimulus only by the unsatisfactory criterion of an abrupt
muscle contraction. Today we know with positiveness that a
continued effect also exists during the uninterrupted flowing of
a constant current in nerve or muscle, though much weaker, however,
than in the case of the excitations produced by sudden
fluctuations of the intensity. This is shown in the nerve by an
altered excitability, which continues at the poles during the whole
duration of the current. In the region of the anode the excitability
is diminished, in that of the cathode it is increased. An
excitation can also be demonstrated which extends from the
cathode through the nerve, which can easily be detected by
sufficiently delicate methods. Among other effects of prolonged
stimulation is that of cathodal contracture, which remains localized
in the region of the cathode and which excitation persists as
long as the current continues. This permanent excitation can be
particularly well observed in the single cells of the rhizopods.
If a constant current is allowed to flow through an Actinosphærium,36
the straight, smooth, ray-shaped pseudopods of the
cell body at the moment of “making,” show evidence of contraction
by being drawn in, particularly those directed towards58
the anodic and in less degree also those towards the cathodic
pole. This excitation, greatest at the time of “making” of the
current, though diminishing rapidly in intensity during its continuance,
remains, however, to a less degree, and leads to a
progressive disintegration of the protoplasm on the side towards
the anode, which lasts until the current is again broken. (Figure
6.) Thus even though there can be no doubt, on the one hand,
that the effect of stimulation, which appears at the moment of
the entrance, is to produce alterations, which develop very rapidly,
and that by a continuation of this state there is a more or less
rapid fall to a low level; on the other hand, it is just as certain
that the alterations in the living system persist throughout the
duration of the changed external conditions, or to put it more59
concisely: the effect of the stimulus never wholly disappears
unless the changes in the external vital conditions return to their
original state.
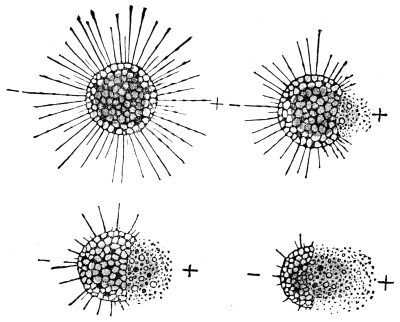
Fig. 6.
Actinosphaerium eichhornii. Four stages showing the progressive influence
of a constant current. Protoplasmic disintegration at
the side toward the anode.
But more, an effect of the stimulus cannot indeed take place
without a certain duration of stimulation, which is related in its
turn to the rapidity of reaction of particular living system. This
can be much more readily observed in more slowly reacting substances.
Fick37 first proved this fact on the muscle of the Anodonta.
I have also been able to demonstrate the same fact in the
slowly reacting sea rhizopods38 by the use of the constant current.
When Orbitolites is stimulated with a constant current lasting
approximately the tenth of a second, no response is seen in its
extended pseudopods, which are directed towards the poles.
The same is the case if the induction current is employed. Only
when the constant current of the uniform strength lasts
approximately .05 seconds, a barely perceptible response occurs,
manifested by the sudden stoppage of the centrifugal flowing of
granules in the anodic pseudopods, which, however, after the
lapse of one to three seconds continues again unaltered. Should
the duration of the constant current be still further prolonged,
typical symptoms of contraction are seen being manifested by a
heaping up of the protoplasm in the pseudopods in the form of
spindles and balls, whilst the protoplasm flows in a centripetal
direction towards the central cell body. (Figure 7.)
Two effects can be realized by the alteration in the living
system as the result of prolonged stimulation. Either a new
state of equilibrium is established by the prolonged action, or
sooner or later death develops. In considering both results, however,
we will ignore for the present the fact that every living
system in the absence of such prolonged stimulation is always in
a state of change, i.e., development. Only with this restriction
can an equilibrium of the living system be spoken of.
60
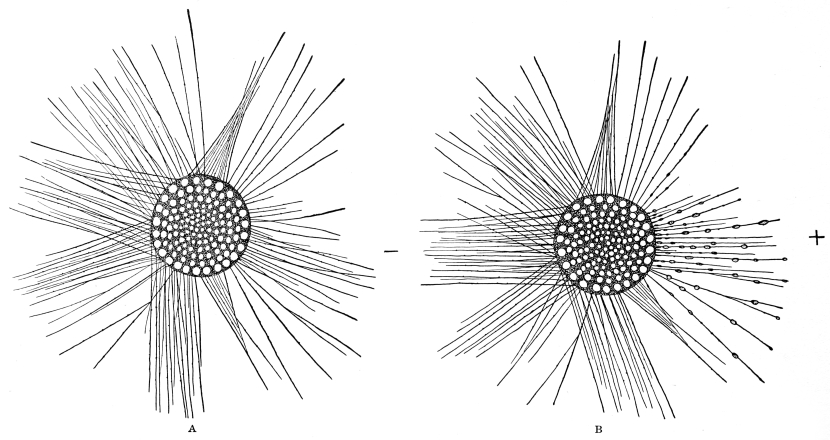
Fig. 7.
Orbitolites complanatus. A—Before stimulation. B—Under influence of a constant current.
61
It is sometimes the case that under the influence of a stimulus
a new equilibrium is developed, which may remain as long as
the stimulus persists. This most frequently occurs as a result
of weak stimuli. That which is usually termed “individual
adaptation” belongs in this category. Likewise some of the natural
and artificial immunizations may also be included. The continued
stimulation in such cases of adaptation as we learned
before in the example of Amœba limax and radiosa or Branchipus
stagnalis and Artemia salina becomes a vital condition for the
living substance in its new state.
The other result, namely, that of death ensuing sooner or
later, is most frequently produced by stronger stimulation.
Through the effect of the prolonged stimulation, the change in
the living system is so great that all harmonious interaction of
the various processes of life become after a time impossible.
The disturbance of this equilibrium after a longer or shorter time
becomes so great that life ceases. By far the greater number of
all diseases furnish examples of this kind. Disease is nothing
else but reaction to stimulation. Should a constant stimulus persist
and if the development of a new equilibrium of this system
is not established, the result is premature death.
In most cases, as, for instance, the nerve impulses which move
toward an organ, or better still the electrical stimuli as used for
experimental purposes, it is not a question of a permanent but
of a temporary alteration in the external vital conditions. The
stimulus starts, then ceases after a longer or shorter period. In
this way there is added to the deviation at the start also the alteration
at its termination. The latter takes place with different
degrees of rapidity, in a manner analogous to that of the initial
alteration, and can bring about response. With this the curve
of the duration of the course of the stimulus becomes somewhat
more complicated and in consequence a like effect is observed in
the response. The “making,” duration and “breaking” of the
constant current furnishes the example of this type. The
“making” of the current being a quick alteration calls forth a
strong and sudden excitation (in the muscle contraction); the
continuation of the current maintains weak excitation of equal62
intensity (in the muscle a continued contraction) and the
“breaking,” being a sudden alteration, is followed again by a
stronger excitation (in the muscle a contraction). The duration
of the change can, however, be so short that its intensity does
not remain at two periods of time at the same height, but instead
the ascent of the intensity is immediately followed by its descent
to zero. Induction shocks of short duration, the duration of
which have been observed more in detail especially by Grützner,39
offer typical examples. Here a single effect of the stimulus results
from the rise and fall of the intensity curve. Hence the induction
shocks as momentary stimuli are universally used for experimental
purposes.
In contrast to the single stimuli, which find their ideal in
induction shocks, another form of stimulation should receive our
attention, namely, the series of stimuli which produce a rhythmical
alteration of vital conditions. These show among their
complex combination of simultaneous and successive actions of
their single stimuli relatively the simplest and most easily understood
regularity in their effects. They are of particular interest,
because they develop in the normal physiological happenings of
the animal body in the form of rhythmical intermittent impulses
of the nervous system.
Here again it is self-evident that with regard to the course of
response, we must first consider the character of the single
stimulus of the series, and this must be done from all those standpoints
already here discussed. However, a new factor is met
with here, that is, the frequency of the single stimuli of the series,
or that which has the same meaning, the duration of the intervals
between them. This is a feature upon which the result of stimulation
depends in a very high degree. But here, too, however,
it is not a case of the absolute frequency of the single stimulus,
but simply of the relative frequency in regard to the rapidity of
reaction of the particular living system. I should like to remark
here that it is of greatest importance whether the interval between
the two single stimuli of the series is sufficiently long or not to63
allow the living system time to completely recover from the effect
of the preceding stimulus. In the cases, for instance, where we
have recovery, we have the same rhythm of stimulation as that
of response. When recovery does not occur, interferences of the
response are developed, which are of great physiological importance,
with the analysis of which we shall later on find occasion
to occupy ourselves in detail. The physiological example for these
stimuli is the rhythmical discharge of impulses of the nerve
centers; the physical method, which is most widely used for
experiments, is the faradic current.
It is apparent that the question of frequency must again be
combined with all those factors previously discussed in connection
with the single stimulus. In consequence another complication
arises and with this another point must be taken into
consideration, namely, the fact that the duration of the single
stimulus in a series undergoes alteration by increasing frequency
beyond a certain limit. Beyond this limit the duration of the
single stimulus must become less and less. As the result of the
fact that stimulation is, as we have seen, dependent on the duration
of stimulus, it is evident that, depending upon the rapidity
of response of the living system, sooner or later the rhythmical
stimulation must become ineffectual. Nevertheless, this effect
of shortening the duration of the single stimulus can be compensated
by a corresponding increase of its intensity. In this connection
Nernst40 showed a very simple relation for induction currents
of higher frequency of interruption, which furnishes a law
according to which such a compensation takes place. In conjunction
with Barratt he found, namely, that the intensity must increase
proportionately to the square root of the number of single
stimuli if the threshold value of the stimulus is to be maintained,
that is, I : √m = const., in which I is the intensity of the current
and m the frequency of interruptions. The limits of the validity
of this law cannot at present be conclusively established.
This exhausts the small number of elementary factors concerned
in the course of the stimulation, and which are of importance64
in considering its effect. The combination of the different
varieties of these single factors, that is, the nature, the direction,
the intensity, the rapidity, the duration and number of alterations
in the external vital conditions of the organism produce
the enormous variety of effects of stimulation which we observe
in the living world.
65
CHAPTER IV
THE GENERAL EFFECT OF STIMULATION
Contents: Various examples of the effects of stimulation. Metabolism
of rest and metabolism of stimulation. Metabolic equilibrium. Disturbances
of equilibrium by stimuli. Quantitative and qualitative
alterations of the metabolism of rest under the influence of stimuli.
Excitation and depression. Specific energy of living substance.
Qualitative alterations of the specific metabolism and their relations
to pathology. Functional and cytoplastic stimuli. Relations of the
cytoplastic effects of stimuli to the functional. Hypertrophy of
activity and atrophy of inactivity. Metabolic alterations during
growth of the cell. Primary and secondary effects of stimulation.
Scheme of effects of stimulation.
In the foregoing lectures we have had occasion to touch more
or less often on the subject of the effects of the stimuli. This
was the case, however, only when it appeared necessary to obtain
a systematic knowledge of the stimuli and the differentiation of
the individual factors. We will now proceed to consider the
effect of stimulation in a more systematic manner. The conditional
method of observation, however, will remain our guide.
We have already pointed out the relations between the conception
of stimulation and that of vital conditions, now we will
consider that of the effect of stimulation with that of vital processes.
Nevertheless, the effect of stimulation being a manifestation
of the vital process is not, therefore, in opposition to the latter
as such. Hence the question presents itself as to the connections
between vital process and the effect of stimulation.
When we study the motile flagellate infusorium Peranema
swimming undisturbed in water, we observe that the swimming
movements are absolutely regular in character. The elongated
cell body remains unaltered in shape. The long flagellum is
extended in a perfectly straight line in the axis of the body and66
only the extreme end lashes with regularity through the water
(Figure 8, A). There is majestic grace in this perfect uniformity
of motion. The picture suddenly alters the moment the Peranema
is influenced by the slightest jar. The whole flagellum at once
executes a few violent movements (Figure 8, B), the body draws
together, soon stretches itself again and swims immediately after,
in another direction, with the same majestic calm as before.
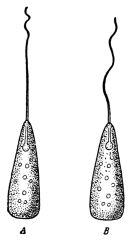
Fig. 8.
Peranema. A—Swimming in non-stimulated condition.
B—Mechanically stimulated at the end of the flagellum.
Another instance. A number of fertilized eggs of the sea
urchin are placed in a watch glass in sea water. The temperature
of the water should correspond with the mean temperature in
which the animals live in the sea, averaging about 15° C. The
eggs begin to form grooves and to develop slowly by progressive
division. In another glass we observe a second sample of fertilized
eggs of the same kind and under the same conditions, but
in this case we increase the temperature to 25° C. The increased
temperature brings about a decided increase of segmentation and
the same stage of development is reached in less than half the
time. The increased temperature, therefore, increases the development.
Further we take a third sample of the same urchin
eggs in a watch glass with sea water of 15° C. and add a little67
sea water mixed with ether. The development of the eggs now
comes to a standstill. The narcotic has produced an inhibition
of development.
To quote another instance. Bacterium phosphorescens having
been bred upon a putrid fish are exposed in the culture fluid to
the air. In the dark the bacteria give forth a phosphorescent
light. Then the culture fluid containing the bacteria is put into
a glass receptacle, which can be rendered air-tight and all oxygen
excluded. After a short time the light formation ceases completely.
The absence of oxygen has here had a depressing effect
and it is only after air has been again introduced that light is once
more produced.
Lastly, an example from the group of mammals may be cited.
The metabolism of a dog in complete rest is examined for a prolonged
length of time and we ascertain the values of the oxygen
consumption, the carbon dioxide production, and the nitrogen
elimination in the urine. Under the same nutritive conditions
the animal is then allowed to work from time to time in a treadmill.
During these working periods impulses of excitation are
continually conducted to the muscles from the nervous system.
It is now found that under the influence of the constantly recurring
stimuli the quantity of nitrogen in the urine has only
very slightly augmented, whereas the consumption of oxygen and
the production of carbon dioxide has markedly increased.
What conclusions can be drawn from these instances of response
to stimuli, of which any number could still be quoted?
They show us, first of all, that a state or process existing under
given conditions, is altered by the influence of the stimulus. This
is a fact, however, which could be expected from the beginning
and is self-evident, for stimuli are alterations in the vital conditions,
and when these are altered the state of the system or the
happenings thereof must also alter. The question with which
we are here more closely concerned, however, is a somewhat
more detailed characterization of the state or process itself, as
well as that of alterations produced by the influence of the stimulus.
The instances of response to stimuli already cited furnish
us with information in both kinds.
68
In all these examples, the living processes occur with equal
constancy and unaltered rapidity, provided a stimulus is not
operative. Here, however, the gradual alterations, the result of
development, must not be overlooked. An excellent example of
this is seen in the eggs of sea urchin, where the development is
readily perceptible. In all these instances, however, the condition
is immediately changed by the influence of the stimulus.
The previous state of constancy in the vital process is disturbed.
The rapidity of its course is changed, being either increased or
decreased, and the specific vital manifestations concerned are,
therefore, augmented or diminished. We will now study the
vital process with the methods of chemical investigation and consider
the problem from the standpoint of metabolism. It may
be noted here, that other methods, such as the transformation of
energy or changes of form of the living system, would serve
equally well as indicators for this purpose. In every instance
there is a uniformity of the processes; the difference, however,
is in the nature of the indicators and the terms used. The methods
and the terms used in chemical investigation and description
reach proportionately much deeper than those employed when
the transformation, energy or the variations of form of the
organisms are studied, and permit of the finest differentiation of
the processes. The atomistic terminology is, for this reason,
preëminently fitted for the description of vital processes. When
we study the vital process metabolically, we can, as shown in the
above-mentioned instance, divide the processes into a metabolism
of stimulation in contradistinction to a metabolism of rest.
The comprehension of the metabolism of rest demands a closer
consideration. On closer observation we must say that this
much-used conception is merely an abstraction nowhere realized
in a strict sense. In truth, there is nowhere in nature a metabolism
of rest. No cell exists which in a mathematical sense
remains for even two successive moments under absolutely the
same external conditions. If we imagine a single living cell of
the simplest kind living in a fluid nutritive medium, and if we
suppose its body and surroundings so magnified that the single
molecules and atoms were respectively of the size of cannon and69
rifle balls, the boundary between cell and medium would represent
a battlefield, on which a heavy bombardment is constantly
taking place. The rain of shot of food and oxygen molecules
penetrating into the cell from the medium, would produce an
explosion in the existing ammunition depots, now at one point,
now at another, creating great breaches through which new
masses of shot would reach the interior. The fragments of these
exploding molecules would be flung out here and there into the
medium and would stem, now at this, now at that point the
besieging masses of shot. In this wild confusion on the whole
boundary line between cell and medium there can be no question
of rest or even equilibrium at any point. The human mind,
superior to the material world as we may deem it, is yet always
dependent upon the results of experience, and even in its highest
flights cannot become wholly emancipated from the concrete
objects. For this reason it is of great purport to conceive processes
whose dimensions cannot be observed even microscopically,
as enlarged and transformed to that method of expression most
familiar to the human mind, namely, in the field of optical presentation.
This method is of great help in aiding our understanding,
and likewise here, even in the resting state, the cell is
constantly exposed to local effects of stimulation, now at one
point, now at the other. The conception of the metabolism of
rest is, therefore, in a strict sense fiction.
Nevertheless, the conception of the metabolism of rest as an
abstraction can be of value provided always that it is strictly
and definitely limited. It must, for instance, not be applied to
short periods of time. The continued local and temporary responses
to stimulation constitute a mean value which, although
composed of numberless small sub-threshold responses, we can
still call a metabolism of rest. Weak stimuli have, however, as
already seen, the property, provided their influence is constant,
of effecting an adaptation to the stimulus on the part of the living
organism, so that the stimulus becomes a vital condition for this
state of the organism. Hence the continued existence of a vital
process resulting from the constant action of stimulation is made
possible. That which we are in the habit of calling metabolism70
of rest, would, therefore, be metabolism of stimulation, but one
that is characterized by a constantly existing metabolic equilibrium.
This “equilibrium of metabolism” distinguishes the metabolism
of rest from that form which is developed in response to temporary
stimulation, in that every temporary stimulation has the
effect that it disturbs the existing metabolic equilibrium for a
longer or shorter time. This disturbance of the equilibrium of
metabolism can in contrast to the metabolism of rest be termed
“metabolism of stimulation.” In this, but only in this sense, can
these two conceptions be placed in opposition and used to characterize
the processes in the living organism. The conception
of the metabolism of stimulation must always stand in relation
to that of an equilibrium of metabolism characterized by a constantly
existing metabolism of rest, just as the conception of
stimulus can likewise only be defined relatively to that of vital
conditions.
Nevertheless, the conception of the equilibrium of metabolism
requires a somewhat more accurate definition before we can feel
justified in using this term. Definitions are always trite, nevertheless
they are the basis of all our thinking and a definite understanding
is impossible unless we first clearly fix their contents.
The history of theology and philosophy even to the most recent
times furnishes a long line of instances in which the most eminent
minds, for the want of fixed definitions of the conceptions which
they made use of, failed to find a mutual basis for their ideas.
Without a sharp definition every conception is a mere word,
which each individual, according to his personal experiences and
views, endows with a different meaning. To such conceptions
we may apply Mephisto’s ironical comment to his pupil:
“Mit Worten lässt sich trefflich streiten,
Mit Worten ein System bereiten.”
The natural sciences, if they are to retain their reputation for
exactness and precision, require the strictest and clearest definitions
of all conceptions. If we seek to penetrate more deeply
into the varied happenings in concrete conditions, we must reconcile71
ourselves to dry pedantic definitions. In the case of that
of the equilibrium of metabolism indeed we have before us one
of the most important conceptions in physiology.
The justification to speak of an equilibrium of metabolism
arises from investigations of metabolism in mammals. The classical
experiments of the previous century, as is well known, have
shown that in the adult mammal receiving a necessary quantity
of nourishment and in a state of rest, the intake and outgo of
the constituent elements are the same. The carbon, hydrogen,
nitrogen, oxygen, sulphur, phosphorus, etc., taken in during
a lengthened period in the form of food and respired air,
appear again in equal quantity, in other combinations, in the
products of excretion of the organisms. Calorimetric experiments
likewise show an equilibrium of the consumption and elimination
of energy. If there thus exists an equilibrium of metabolism for
the whole cell community, it is clear that the same must also apply
to the individual cell, that is, for all living substance. The quantitative
relations of the foodstuffs taken in, and the excreted metabolic
products given off, are, however, merely a standard of the
metabolism. We know that the former are used to build up new
living substance and that the latter represent the result of disintegration
of that previously existing living substance; for we
find, as in the case of the plant, complicated protein combinations,
which are built up from comparatively simple constituents of the
food and are again broken down into comparatively simple substances.
And so the building up and breaking down processes
form the two great processes of metabolism, which with Hering41
we can briefly call “assimilation” and “dissimilation.” In the
terms assimilation and dissimilation are comprised the sum of all
processes of construction and disintegration in the living organism.
It is apparent that equilibrium of metabolism occurs when
assimilation and dissimilation are equal. The formula A : D, that
is, the relation of the sum of all assimilation to the sum of that of
all dissimilative processes, is a factor of fundamental importance
in the study of the course of the vital processes, for upon its72
value depends individual vital manifestation, and, in fact, the continuation
of life. I have, therefore, designated the formula
A = D “Biotonus.” The equilibrium of metabolism would then
be characterized by the biotonus42 of a living organism being
equal to one. This would be the metabolism of rest of a system,
whilst its metabolism of stimulation would consist in an alteration
of the biotonus. But is this state of living substance strictly
speaking ever realized?
In considering the nature of the equilibrium of metabolism
one factor has been disregarded which must be taken into account
at every point; this is growth. Growth changes, although
varying more or less, are never absent during the life of the
organism. An equilibrium of metabolism never exists in a
strictly mathematical sense, and here again we are working with
a conception which is faulty, because it is an abstraction, originating
from experience with rather too restricted boundaries.
But an error of which one is aware is not dangerous. In mathematics
we also consciously reckon with errors, without the result
being altered. In the before mentioned cases the equilibrium of
metabolism was maintained, because the investigations involved
only a short time in an adult mammal. In the adult mammal the
growth processes occur very slowly, so that alterations within
a relatively short time are not demonstrated.
If it were possible to subject the adult mammal to metabolic
or calorimetric experiments, extending for years, it would be
found that the intake would be qualitatively and quantitatively
different at the end of the investigation and that the same would
apply to the outgo. In the growing egg cell this takes place with
much more rapidity. In the organism which rapidly grows, it
can be seen at once that the quantity of the outgo of the products
of disintegration cannot be equal to that of the intake of foodstuffs.
If biotonus were equal to one, the organism could not
grow. Equilibrium of metabolism can only be understood when
we take into consideration a period of time in which the alterations
in growth take place with such imperceptible slowness that73
the resultant error is inconsiderably minute. This period of time
is of greatly varying length in different living organisms and this
fact must be taken into account in every living form. Only with
this restriction can we justify the use of the term “equilibrium
of metabolism.” Then, however, its use is of great value.
The metabolism of stimulation is then a disturbance of the
metabolism of rest, that is, a disturbance of the equilibrium of
metabolism through the effect of stimuli.
The question here follows: Is there a constancy of this interruption
of the equilibrium of rest produced by the stimulus which
can be formulated into a general law? To begin with, the number
of possible responses are greater than the variety of forms of
living substance, for every living organism with its specific properties
can undergo alteration in its metabolism in various directions.
Thereby results an infinite number of manifold reactions
to stimuli. However, in answer to the question, in which direction
the change in the specific metabolism of rest in response to a
stimulus takes place, we find a comparatively simple scheme of
general reaction. All phenomena can change in their rapidity as
well as in their nature. That is quantitatively and qualitatively.
In this way the specific vital process of an organism can be altered
by the stimulus, on the one hand, in its rapidity; on the other, in
the manner of its action.
The majority of all temporary responses to stimuli consist in
alterations of rapidity of the vital process, and form either a
quickening or retardation of its course. The former is manifested
in a strengthening or an increase, the latter in a decrease
or repression of the specific action of the living organism. The
stimuli have the same effect as in the case of the catalysers in
chemical processes. According to Ostwald’s43 well-known definition
of catalysis a catalyser is a substance which, without
appearing in the final product of a chemical reaction, alters its
rapidity. This group of reactions can, therefore, be referred to
as “catalytic stimulation and response.” When the response
consists in increase, we speak, in a physiological sense, of an74
excitation, and when there is decrease in the vital processes, we
speak of a depression.
The conception of excitation and depression are purely empirical.
They are terms for real things, referring, in fact, simply to
alterations in rapidity of life process, which can be as readily
observed as the process itself. I wish to lay particular stress on
this fact, for the reason that Cremer44 has recently made the
extraordinary statement that I have introduced hypothetical processes
into the definition of the conception of excitation. I have
always considered excitation as merely an increase or change of
intensity of the specific actions of a living system, and as such is
an established process without a trace of the hypothetical element.45
If, however, the excitation process is to be regarded as something
absolute, as a mysterious state sui generis, which is entirely independent
and totally unlike the metabolism of rest, then, of course,
it would appear utterly incomprehensible and would be without
purpose. As an absolute process excitation is merely a meaningless
word. Excitation and depression are relative conceptions
and can only acquire meaning when the process which is excitated
or depressed is more closely defined. This is the specific
vital process of a given organism, and the two conceptions only
have meaning in relation to it. The conception of the vital process,
however, is one directly gained from experience. However
complex or difficult to analyze the process may be, it still is as
little hypothetical as that of the combustion of carbon into carbon
dioxide, or the revolving of the earth around the sun. It can be
looked upon as something positive and real. Quite another
question is the manner in which we are to consider the mechanism
of the vital process. In analyzing this mechanism we cannot, at
least in the present state of our knowledge, entirely dispense with
hypothesis. But these hypotheses are in no way involved in the
definition of the process of excitation. If we look upon every75
excitation or depression produced by a stimulus as an alteration
in rapidity in the specific vital process of a given organism, we
are thereby expressing the same fact which Johannes Müller has
termed “specific energy.” We give, however, the doctrine of
specific energy a more general application in so far as it comprehends
not only the increase but likewise the decrease of activity
in response to stimuli. Johannes Müller’s doctrine of specific
energy of the living substance at all times has been the subject
of most animated discussion. When I refer here to the specific
energy of living substance, it is with the knowledge that Johannes
Müller did not use this expression of “living substance” in this
connection. He was already acquainted, however, as we have
seen, with the fact of the existence of the specific energy of all
living structures. For appertaining to the muscle he says: “This
is universal in all organic reaction.” The reason why the doctrine
of sense energy has become of importance in the discussion
of the specific energy of the living substance, is in consequence of
the theoretical interest, resulting from its connection with the
nature of the specific energy of our sense substances. The controversies
on this subject are still far from settled.46 Indeed,
according to the special philosophical standpoint taken by an
observer, the existence of a specific energy of the senses is
acknowledged or disputed. For any one acquainted with the
general physiological reaction to stimuli, such a discussion is
wholly without purport. The sense substances have as a matter
of course in common with all living substances their specific
energy, that is, the influence of stimuli can produce an increase
or decrease of their specific vital processes. “Specific energy” of
“sense substance” in this sense is like that of all other living substances,
a fact. In that the psychical capability of these sense
substances, in which we include not only the peripheral, but also
the central portion, are dependent upon their specific vital processes,
it must be self-evident that the excitation and the suppression
of sense sensation can be brought about by adequate and76
inadequate stimuli, no matter what one may think of the relations
between physical and psychical phenomena.
The only debatable question is that concerning the limits of the
validity of the doctrine of the specific energy of living substances.
This question will involve our attention when we have analyzed
somewhat more closely the happenings in the living substance
taking place under the influence of stimuli. We will, therefore,
return later on to a more detailed consideration of the last question.
Nevertheless, we will here refer to a fact which, upon a
superficial observation, seems to restrict the validity of the conception
of the specific energy of living substance.
In contrast to those reactions to stimuli, which consist merely
in the changes of a rapidity of the specific vital process, are
another group of reactions in which the influence of stimuli leads
to qualitative alterations in the specific vital process. In these
instances, the influence of the stimulus directs the metabolism of
rest into new channels, so that chemical processes occur in the
cell, which under ordinary circumstances do not take place. This
group of reactions, which I wish to term “metamorphic stimulation
and response,” are chiefly observed where weak stimuli act
continuously upon the living substance. These are essentially
weak chemical stimuli, which last for a prolonged period or frequently
reoccur in the life of the cell community. Examples of
this are found in the continual ingestion of alcohol and other
poisons by the human being, or in the formation of metabolic
products of bacteria, etc. The majority of chronic diseases
belong to this group of reactions; disease being simply response
to stimulation. Disease is life under altered vital conditions and
altered vital conditions are stimuli. This simple and self-evident
fact shows the immense importance which the knowledge of the
general laws of the physiology of stimulation has for pathology.
The pathologist, who does not wish to confine his observations
to a purely superficial symptomatology or a merely histological
morphology, must seek above all to penetrate as deeply as possible
into the nature of the general reactions to stimulation in the living
organism. It is the essential point which meets him everywhere.
In spite of their great interest for pathology, however, it is just77
these qualitative alterations of the normal vital process produced
by continuous stimulation which have up to now been least analyzed.
In this field we expect much from pathological investigation
which alone has the immense amount of material at its
command. This will take place only when pathology adds to the
almost exclusively histological direction of investigation, that also
of experimental physiology. It is true that the problems of the
qualitative alterations of a vital process by chronic stimulation
are much more complicated than those of the rapid responses to
temporary stimuli, consisting simply in mere alterations of
rapidity of the specific vital process. An understanding of the
nature of the former can only be expected when a deeper knowledge
of the latter is gained, for, as will be seen presently, there is
the closest relation between the two groups.
The reactions to catalytic stimuli of short duration, which produce
merely an alteration of rapidity in the specific phenomena
of a living organism, show on a closer analysis the interesting
fact, that it is not always the entire metabolic processes of the
cell which are perceptibly quickened, but that only certain constituent
processes of the same are affected by the action of excitation.
This is the more noticeable, as, considering the close correlation
which all the individual links of the chain of metabolism
bear to each other, it is to be expected that the alteration in rapidity
of one would be followed at once by a corresponding change
in all the others. An example of the case in question, in which
a special constituent process may be predominately affected, is
that of the specific activity of a muscle which is repeatedly stimulated
by nervous impulses. Since the classical investigation of
Fick and Wislicenus47 on themselves, and of Voit48 on the dog, we
know that the nitrogen metabolism is practically unaltered by
the functional use of the muscle and there is a remarkable
increase only in the breaking down of the nitrogen-free groups78
of the living substance. Sufficient importance has not as yet been
attached to this knowledge. This fact not only has a particular
interest for the much-discussed question of the source of muscle
energy, but also affords a deeper insight into the metabolic activity
of the living substance. It shows us that we must not imagine
a purely linear linking of the individual constituent metabolic
processes, but rather, at least at certain points, a branching formation,
the individual members spreading in various directions. An
alteration in an individual member can occur without an immediate
change in the other branches. This would not be the case
if there were only a linear connection of the constituent processes,
for the breaking of a single member of the chain would be
followed by a change in all the following members.
It shows us, further, that certain branches are more labile than
others. In the case referred to here, the branches of this system,
which bring about the nitrogen metabolism, are relatively firm
and stable, the branches, which are disturbed by the stimulus producing
functional activity of the muscle, are particularly labile.
I should like in passing to call here your attention to the fact that
as is well known, Ehrlich,49 in another field involving other conditions
and other experiences and considerations, has arrived in
analogous manner at his “side chain theory.” In order to have
an expression for those stimuli which involve rapid alteration of
the labile constituent processes and which are connected with the
specific action of the particular organism, I have called them
“functional stimuli,” and contrasted with them the “cytoplastic
stimuli.” In the latter the alterations produced include all the
constituent processes extending even to the stable processes of
nitrogen changes, and sometimes extend to complete disintegration
and rebuilding of living substance.50 To the first group belong
all adequate stimuli within certain limits of duration and intensity,
and the greater part of inadequate stimuli of brief duration so79
long as they do not exceed a certain intensity. To the latter
group belong in general all the stronger adequate and inadequate
stimuli of prolonged duration; such as extreme temperature, the
stronger electric currents, constant alteration in the supply of
food, water, oxygen, the prolonged or stronger influence of
extraneous chemical matter, etc.
Considering the close correlation of the individual part processes
it would appear very strange, however, if a single one of
these could undergo an alteration of its rapidity without the
course of the rest of the processes being in the least influenced.
One cannot comprehend such absolute independence of a process
brought about by functional stimulation from all the other constituent
processes, particularly when this is of prolonged duration
and involves to a considerable extent the alterations in rapidity,
for the individual constituent processes are dependent in a
high degree upon the quantity of the particular chemical substances
of which the living system is composed. The cycle of
the individual constituent processes of this system is determined
in the most delicate manner in its rapidity and extent, by the
relative quantities of the individual substances. Associated with
an alteration in the rapidity of an individual constituent process,
there would also be a relative alteration quantitatively of the
substances. And with the increase in the quantity of the disintegration
products, and also the increase of the substances for
their replacement, there would result, during this time, an alteration
in the amount of interaction of the molecules of the other
constituent processes, so that these processes secondarily suffer
an alteration in rapidity which is perceptible after long continued
involvement of the functional part of metabolism.
In fact, in the previously mentioned case of the functional
stimulation of the muscle, the proof has been furnished that a
long-continued increase of the functional metabolism is followed,
although to a less extent, by an increase in the entire
cytoplastic metabolism. Argutinski showed this on himself in
1890 in Pflüger’s laboratory. He found, namely, that after the
exertion of a long walk in a hilly district, a considerable increase
of nitrogen excretion in the urine took place, which extended80
over the succeeding two or three days. This increase of the
nitrogen metabolism in its totality is not nearly as great as that
of the breaking down of nitrogen-free substances, but it is,
nevertheless, present and shows us that functional metabolism
cannot experience a lasting excitation without being followed by
secondary results in the entire cytoplastic metabolism. This
fact is even more strikingly illustrated in the alteration of the
entire volume of a living organism as produced by the lengthened
duration of functional stimulation. It has been long known, that
the muscle as the result of frequent functional excitation by
means of adequate nerve impulses, that is, prolonged activity, is
considerably increased in size, whereas in the absence of such
it loses more and more in volume. A hypertrophy of activity,
produced by functional stimuli, and the atrophy of inactivity, the
result of the discontinuance of the functional excitation, is universal
and can be observed in the various tissues of our body.
We see it, for example, in the glands; we see it in the skin and
we see it in the elements of the nervous system. Berger,51 for
instance, established the fact that the ganglion cells of the optic
lobe in the cerebrum of newborn dogs only reach their full
development when functionally excitated by adequate light stimuli
(Figure 9, B), coming from the eye, whereas they remain in
the embryonic state when these light stimuli are eliminated. (Figure
9, A.) The cytoplastic increase of volume of the neurons
under the influence of functional stimuli is a fact of fundamental
importance for the entire happenings of the nervous system and
forms the physiological basis for reinforcement of reflexes, which,
in its turn, is essential for all acts of memory and intelligence.
For the increase in volume of the ganglion cell body is, when
functionally activated, accompanied at the same time by an
increase of specific capabilities and the intensity of discharge.
Its excitation impulses can, therefore, be conducted through a
greater number of neurons, with which it is connected, than would
be the case if development of the volume of the ganglion cell
increased to a less extent.
81
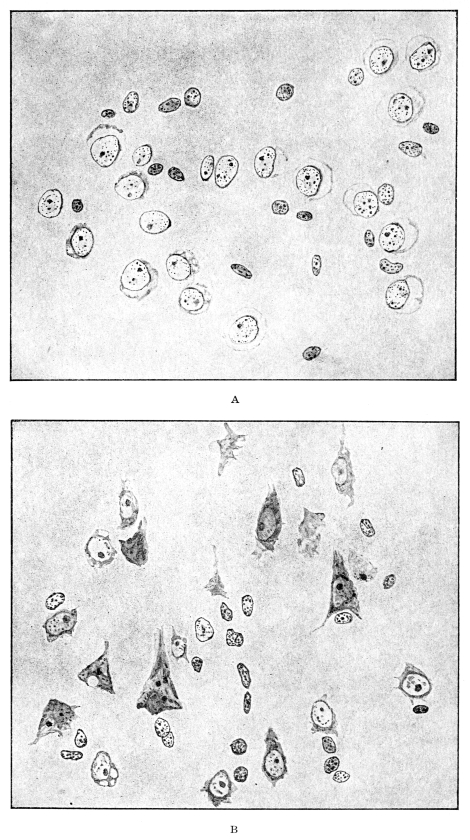
Fig. 9.
A—Undeveloped ganglia cells in the optic lobe of a dog, the eyes of which have been
sewn up immediately after birth. B—Fully developed ganglia cells in the same
region of a normal dog of the same age. (After Berger.)
82
The increase in volume under the influence of stimuli further
shows the relation between the group of those solely catalytic
effects of stimulation consisting in mere alterations of rapidity of
the specific vital process, and that of the metamorphotic effects
of stimulation, which manifest themselves in qualitative alterations
of the vital process. Simple observation shows us that a
qualitative change of individual constituent processes must necessarily
result from the increase of volume of a cell, and that considering
the close correlation of all the individual processes a
profound alteration of the entire metabolism must be produced.
I have already at another place52, 53 treated these conditions more in
detail and will, therefore, only briefly refer to them here. If we
study the growth of a ball-shaped cell, we find that the surface
then increases as a square, and the volume as the cube. It therefore
follows that, by progressive volume increase, the conditions
for the interchange of substance with the surrounding medium
must become more and more unfavorable for those cell portions
situated in the interior, whereas those at the exterior are at much
greater advantage. This must lead to a constantly increasing
difference of the rapidity of the metabolic processes between the
peripheral and central portions. Accordingly, the intricate interworkings
of the individual constituent processes, the rapidity of
action of all which is intimately connected, are, therefore, followed
by corresponding alterations in the entire metabolism.
Sooner or later a stage is reached in which the individual constituent
processes become so limited that certain metabolic products,
which previously were broken down as soon as formed, can be
no longer eliminated and remain in the cell acting as foreign
bodies. In this way the relative quantity of the individual cell
substances become more and more altered, and as the course of
chemical processes occurs in accordance with the law of mass
action, the whole metabolism is directed into another channel,
so that finally new constituent processes take place, which were
formerly not possible. These in their turn produce deep-seated83
alterations of the relations of the cell to its surrounding medium,
etc. Hence this mere increase of volume of the cell in growth
forms the source of an infinite mass of alterations in the activities
of cell metabolism, which we briefly term its “development,” and
which by constant progression, leads either to a process of cell
division, and with this to a correction of existing disorder, or
finally to irreparable disturbances ending in death. In this way
an inseparable relation exists between increase of volume and
the development of living substance. We have seen, however,
that the catalytic reactions of stimulation, which at first only
produce an alteration of rapidity of the individual constituent
processes, if of prolonged duration or of frequent recurrence,
secondarily effect a change of volume of the entire living organism.
One can, therefore, hardly reject the conclusion that seeing
the close interworkings of the individual part process of metabolism,
every change of rapidity of a single member, if of prolonged
duration or of frequent occurrence, must finally lead to
qualitative alterations of the entire metabolism. In consequence
there results an important dependence between catalytic stimulation
and metamorphic reaction. Indeed, it is not unlikely that
the metamorphic reactions, which are especially seen in the continued
effect of weak stimuli, result from alterations of rapidity,
which the individual members of the vital processes have
primarily undergone from this influence.
It is perhaps expedient to cite a concrete instance in illustration.
A simple example is furnished by asphyxiation. If oxygen is
withdrawn from any living organism, the result is a depression
of its oxydation processes. Here there is primarily only a change
in rapidity, especially a retardation of oxydation processes. The
metabolism, however, proceeds, the disintegration of living substance
continues, although at a slower rate, but produces an
accumulation of other products. Whereas formerly during the
existence of a sufficient supply of oxygen an oxydative disintegration
of nitrogen-free groups into carbon dioxide and water took
place, both of which could easily be eliminated from the cell, the
anaërobic disintegration furnishes only complex products, having
a higher carbon content, such as lactic acid, fatty acids, aceton,84
etc. These, being more difficult to excrete from the cell, accumulate.
These asphyxiation products have in their turn a depressing
effect and so on. In this way the whole metabolism is forced into
a wrong course. The accumulation of fat in those tissue-cells
with an insufficient blood supply, as we have seen in the case of
the fat metamorphosis, is doubtless brought about in the same
manner by relative oxygen insufficiency. The fatty acids accumulate
as products of an incomplete combustion and combine with
glycerine to form neutral fats. In like manner it may be that the
accumulation of amyloid substance in amyloid metamorphosis, of
lime salts in arteriosclerosis, etc., is produced by a primary
depression of the individual constituent processes of the particular
cells.
The relation here described, of the catalytic stimuli to the production
of the metamorphic processes, leads us to the distinctions
between primary and secondary effects of stimulation.
Should the general fact be established, which has up to now only
been pointed out in individual cases, that all the metamorphic
processes are merely secondary results of primary alterations in
rapidity of individual metabolic constituent processes, then the
primary reactions of every stimulus would consist purely in the
excitation or depression of the directly concerned constituent.
Whether or not, as may be assumed, this primary effect of stimulation
applies to all stimuli, is a question which only the future
can answer.
The metamorphic processes are not, however, the only secondary
effects of stimulation. The influence of long-continued
excitation of the functional constituent processes upon the entire
cytoplastic metabolism can be looked upon as a secondary response.
Therefore, they may be considered as a secondary effect
of stimulation which, in contrast to this primary excitation, may
be called the secondary excitation.
Further: While the secondary excitation and metamorphic
processes are generally produced by the continued existing effects
of weak stimulation, we also observe as the result of a stimulus
of short duration or frequently repeated at brief intervals, but
otherwise not exceeding the physiological limits of intensity, a85
secondary effect, which plays a very important part in the activity
of the organism. I refer to fatigue. Here a secondary depression
is developed in connection with the primary excitation, for
fatigue of a living organism must be characterized as a depression
of activity. This case shows that we have to distinguish between
a primary depression, as for example, produced by temperature
reduction, withdrawal of food, deficiency of oxygen, etc., which
occurs as a direct effect of stimulation, and secondary depression,
which as in fatigue is an indirect result of primary excitation.
After the cessation of a briefly catalytic stimulus, not exceeding
the physiological limit of intensity, another secondary result
is observed, which is of the greatest importance for the continued
existence of the living substance. The catalytic stimulus
brings about a disturbance of the equilibrium of metabolism,
which after cessation of the stimulus is reestablished by the living
substance. In other words: recovery takes place. This fundamental
principle has been known for a long time as the result
of observation. If a skeletal muscle of our body has been activated
for a prolonged period by nerve impulses, until it has
become completely fatigued and incapable of work, a recovery
takes place on the cessation of these impulses and the muscle is
again capable of action. Likewise, as the result of strong mental
activity during the day, we are mentally fatigued in the evening;
recovery, however, occurs during the night, which results from
the removal of the source of activity. The next morning finds
us refreshed. This restitution occurs in every cell, and the return
of its former capability of action, which had disappeared under
the influence of stimulation, shows that compensation has taken
place of the metabolism of rest, disturbed by the effects of the
stimulus. Hering54 has aptly termed this restitution as “the internal
self-regulation of metabolism.” All recovery after disease is
based on this self-regulation. The physician simply provides, by
means of therapy, for the possibility of its taking place. Healing
itself is brought about by the organism. “Natura sanat, medicus
curat.”
86
Finally, a third kind of secondary effect of stimulation claims
our interest. This is the secondary extension of the result of
stimulation from the part of a living organism directly and primarily
affected by the stimulus, to the surrounding structures.
All living substance has the capability of conducting an excitation,
which is produced locally through a catalytic stimulus, to a neighboring
part, not directly affected by the stimulus. It finds its
highest development in the nerve, but in no living structure is it
completely absent. This capability has been frequently termed
“conductivity of stimulation.” It is more precise, however, to
speak of conductivity of excitation, for it is not the primary
influencing external stimulus which is conducted in the living
substance, but the excitation which it has produced. I have intentionally
considered only the excitating effects of stimulation, and
not those of the depressing reactions, as only excitations, not
depressions, are conducted by the living substance. These questions,
however, demand a closer analysis. Here we were concerned
only with a survey of the general effects of stimulation.
If I, therefore, once more summarize the results which have been
gained, this is most clearly demonstrated by the following scheme:
Primary Effects of Stimulation
Excitation Depression
Functional Cytoplastic Functional
Secondary Effects of Stimulation
Secondary excitation Secondary depression
Conduction of excitation, Metamorphic processes, Self-regulation of metabolism
This, however, is simply a scheme, like all other schemes, having
for its purpose a superficial survey of the subject.
It brings to some extent order into the overwhelming mass of
manifold effects of stimulation but tells us nothing of the mechanism
and genesis. Our further task must, therefore, be a more
thorough analysis of this field.
87
CHAPTER V
THE ANALYSIS OF THE PROCESS OF EXCITATION
Contents: Indicators for the investigation of the process of excitation.
Latent period. The question of the existence of assimilatory excitations.
Dissimilatory excitations. Excitations of the partial components
of functional metabolism. Production of energy in the chemical
splitting up processes. Oxydative and anoxydative disintegration.
Theory of oxydative disintegration. Dependence of irritability on
oxygen. Experiments on unicellular organisms, nerve centers and
nerve fibers. Restitution after disintegration by metabolic self-regulation.
Organic reserve supplies of the cell. The question of a
reserve supply of oxygen of the cell. Metabolic self-regulation as a
form of the law of mass effect, and metabolic equilibrium as a condition
of chemical equilibrium. Functional hypertrophy.
If it is true that all primary effects of stimulation consist either
in an excitation or depression of the metabolism, and that all other
effects of stimulation secondarily follow this primary alteration
of the metabolism of rest, then every thorough analysis of the
mechanics of reaction must have its beginning in the investigation
of these primary processes. I desire to adopt this method
here and will analyze somewhat further the primary process of
excitation and its immediate and remote sequences. This will be
followed later by the analysis of the process of primary depression
and its results.
The investigation of the more obscure processes in the living
substance places us in a difficult position, for their details cannot
be observed by the unaided senses. That which we can perceive
is merely the grosser vital action, consisting of a complex combination
of the individual processes, the total result of a multitude
of different components. For this reason the conception of excitation
can only be established by observations based upon the
combined vital actions, which are produced by the effect of stimulation88
upon the complex system. In the beginning, the process
of excitation was studied exclusively on the muscle and nervous
system. A physical factor served as indicator, such as muscle
contraction or production of electricity. These showed, besides
the direct and primary effect of stimulation, the secondary process
of conductivity. Even graphic registration is merely an
expression of the phenomena composed of a great mass of individual
elements. The visible course of the phenomena, as shown,
for instance, by the latent period by the ascent and descent of the
curve of contraction, represents as it were a reflected picture of
the actual excitation processes similar to an object seen in a distorting
mirror; the first and the last parts of the process are not
even perceptible. Later, when organ physiology was extended
into a cell physiology the processes of excitation were studied in
numerous simple organisms, such as the plant cell, the rhizopoda,
the infusoria, etc. Later, in this way, by the use of comparative
methods many essential facts were discovered. However,
even the single cell, in spite of its minuteness, is, compared with
the size of a molecule, a gigantic system, and it would be a grave
error if we should consider this system even in its simplest aspect
as homogeneous. In order, therefore, to analyze the vital activities
in the cell, cell physiology must endeavor to penetrate into
molecular conditions. For this purpose the indicators employed
must be essentially of a chemical nature, capable of magnifying
the processes of molecular dimension to such a degree that we
are enabled to base conclusions upon these not otherwise directly
perceptible phenomena. To obtain a sufficient magnification we
must necessarily place somewhat larger quantities of living substance
under observation and apply a stimulus of such frequency
or length of duration that the chemical alterations as a result of
excitation are so increased as to be plainly perceptible with the
aid of our chemical indicators. Unfortunately, we do not possess
specific chemical indicators for every individual molecular
constituent process of the cell and so cannot dispose with the help
of indicators of the combined happenings in a greater quantity of
living substance. It remains for us to obtain data concerning the
cycle of excitation processes in the living substances by the aid89
of the combined employment of the most varied kinds of physical
as well as chemical indicators. If we use the most varied types of
living substance of widely differing properties, showing us the
greatest variety of vital manifestations, we may hope by the use
of comparative physiological methods, even though with difficulty,
to separate more and more the essential details of the general
processes of excitation. At present we are still at the very
beginning of this task and vast fields of unexplored regions are
yet before us. But it is the unknown which has a particular fascination,
especially if we succeed from time to time in making new
advances.
If we suppose a living system in a state of metabolism of rest
influenced by an instantaneously excitating stimulus, the entire
course of excitation extends from the first alteration produced
by the stimulation until the complete restitution of the metabolic
equilibrium, and we will, therefore, differentiate individually the
successive stages of this whole process.
The very beginning of the chain of alterations produced by the
excitating stimulus cannot be studied by any indicator. The
changes must first reach a certain dimension by conduction from
the point of stimulation before they influence even the most delicate
indicators. The application of the stimulus is, therefore,
followed at first by a measurable “latent period,” in which the
living substance remains apparently at rest. This latent period
has been particularly studied in muscle. After its discovery by
Helmholtz55 it was made the object of innumerable investigations
and met with an interest which can only be explained by the exactness
of the methods employed. Among others Tigerstedt56 has
made the most thorough study of the influence of various factors
on the duration of the latent period. These experiments have
established the fact that the duration of the latent period varies
according to the intensity of the stimulus, temperature, loading90
or fatigue. This is apparent when it is understood that the
amount of the alterations produced by the stimulus must ascend
from the value zero to a certain height before the changes are
perceptible, and that under various conditions this amount is, on
the one hand, attained in different lengths of time and, on the
other, must reach a varying amount before it is perceptible by
means of the indicator.
The facts concerning the whole latent period and its dependence
on various factors would be incomprehensible if it were
assumed that no alterations whatever take place during the latent
period although the stimulus is already operative. In reality, the
alterations following a stimulus occur with imperceptible rapidity
in the form of a molecular interchange, and the latent period is
simply an expression of the fact that the primary alterations,
being limited in nature, are not registered by our indicators.
The question first arises, In what do these first imperceptible
alterations consist? Nernst57 has evolved the theory for electric
stimulus, that the primary effect produced by the electric current
is an alteration in the ion concentration on the surface of the
living substance. In fact, we know that the surfaces of all protoplasm
possess the property of semi-permeable membranes and that
changes in the concentration of ions invariably occur when an
electric current flows through two electrolytes separated by a
semi-permeable membrane, in which the anions and cations have
a different rapidity of movement. It is apparent, therefore, that
such an alteration in the ion concentration must be followed by
further chemical processes in the living substance. According
to the theory of Nernst the first impetus for all further alterations,
which the electrical stimulus brings about in the metabolism
of rest, is the alteration in the concentration of the ions on both
sides of the semi-permeable membrane, which represents the surface
of the protoplasm. In view of the present findings of physical
chemistry, objections can hardly be made to this theory of
Nernst’s. It is a question, however, in how far this theory, especially
established for the electric stimuli, can be applied to other91
forms of stimuli and their action. It cannot be denied that the
degree of dissociation of an electrolyte can be altered by very
different factors, such as heat, light, chemical processes, etc.,
and in that the surfaces of the protoplasm, acting as semi-permeable
membranes, bring about a selective action on the passage
of the ions, there arises the opportunity for the development
of difference of electrical potential on both sides, and for further
chemical alterations in the protoplasm. These observations, however,
require further experimental investigations in many fields,
before we are justified in extending the Nernst theory of the
manner of action of the electric stimuli to a general explanation
of the primary alterations produced by all stimuli in the living
substance. For the present we must confine our observations to
those alterations which are known to be responses to an excitating
stimulus; these are the chemical alterations in the metabolism
of rest in the living substance.
If it is asked, which members of the entire metabolic chain
are increased primarily by the stimulating excitation of a vital
system, we should not be able to answer this question generally
for all living systems. To begin with, it appears highly
probable that the various forms of vital substances in this respect
act quite differently. It is to be regretted that, up to the present,
this question has not been treated from a comparative standpoint.
This inquiry should be extended to the greatest possible
number of organisms. Still there is enough material at hand,
obtained from the muscles, glands, ganglion cells, nerve fibers
and plants, to show that the complexity is by no means so great
as one might at first assume.
In considering the two stages of metabolism, assimilation and
dissimilation, in their entirety, it appears as a very remarkable
fact, that nearly all stimuli produce primarily a dissimilative
excitation. We are only acquainted with a primary assimilative
excitation, that is, an augmentation of the building up
processes, in short, the formation of living substance, occurring
as a primary result of stimulation, following increased introduction
of foodstuffs extending over a prolonged length of time.
With this exception it cannot be proved that any other stimuli,92
either especially those operative in the activity of the animal
organism or any of the physiological nerve impulses which regulate
the actions of the different organs and tissues, bring about
primarily an assimilative excitation, which leads to an increase
of new formation of living substance. The much-discussed
teaching of the existence of the trophic nerves has not given us
a single case in which there was positive proof that a nerve
impulse brought about a primarily assimilative excitation. I have
endeavored for nearly fifteen years to discover such a case.
My efforts have been, however, without avail. In the most recent
critical review by Jensen58 on the subject of the trophic nerves, the
same conclusion is reached although certain facts, as, for instance,
the excitation of assimilative processes in the green plant cell,
produced by light, seems at the first glance to clearly demonstrate
a primary excitation of the building up processes resulting from
a stimulation. Nevertheless closer observation invariably shows
that these conditions are much more complicated and that primarily
assimilative excitating reaction of the stimulus cannot be
conclusively shown. There remains, therefore, as a primary
assimilative excitating stimulus only the increased introduction
of nutrition in a living organism. This excitating effect on the
assimilative portion of metabolism is, as we shall see later, a
simple manifestation of the law of mass action.
As a result manifold effects of excitating stimulation, which
seemed possible at a first glance, are already considerably restricted.
The great mass of excitating stimuli produce an acceleration
of the dissimilative processes of the metabolic chain. But
here our former observations have already shown that certain
constituent processes are especially responsive and very readily
increase as a result of the most varied adequate and inadequate
stimuli. These are the “functional” members of metabolism.
These members are particularly labile, so that they are always
affected by every influence to which the system is subjected in
the form of a stimulus. The functional portion of metabolism
of the muscle, which is particularly labile and is always primarily93
affected by stimulation, consists as demonstrated in increase
of formation of carbon dioxide and water, and in the disintegration
of the nitrogen-free groups. The innumerable observations
on metabolism during the stage of the activity of the muscle, as
those of Hermann, v. Frey, Fletcher, Johannson, Thunberg,
and many others on the individual muscle, and those by Voit,
Fick and Wislicenus, Pflüger, Rubner, Zuntz, Lehmann and
Hagemann, Bernstein and Löwy and others on the muscle of the
entire organisms, have sufficiently proved this fact. However, we
should not apply in detail the conditions existing in the muscle
to all living substance. Comparative methods show us, rather,
that the functional portion of metabolism is very differently
involved in various forms of living substance. The formation of
carbon dioxide and water is constant in nearly all forms of living
substance. We must, however, exclude certain micro-organisms,
which have adapted themselves to unusual vital conditions.
Further, there appear in some forms manifold special constituent
processes consisting in a disintegration of living substance
which are in part converted into very complex combinations. In
the gland cells this type is represented in an especially high degree.
Here the functional disintegration leads to excretion of proteins,
glycoproteins, nucleoproteins, cholic acid, enzymes of various
kinds, all of which are complex and at the same time nitrogenous
organic combinations. This fact must not be lost sight of. The
origin of these special members, however, for the present is completely
unknown, while on the other hand, it is self-evident that
the general and constant constituents of the process of excitation
must claim a first place in our interest. It is just at this point,
therefore, that we must endeavor to penetrate somewhat more
deeply into the mechanism of the excitation process and analyze
in greater detail the acceleration of the functional constituent
parts of metabolism produced by the stimulus bringing about the
formation of carbon dioxide and water.
The question arises: By what means is the particular labile state
of just this constituent part of functional metabolism conditioned?
The lability of the functional portion of metabolism, excitated
by the stimulus, resembles the processes in the disintegration of94
explosive combinations. Iodide of nitrogen, for instance, in a
manner similar to the living substance in the state of the metabolism
of rest, constantly disintegrates even without the influence
of an impact. The disintegration is suddenly enormously increased
by the result of a jar. An explosion follows. In a like
manner the functional metabolism of rest is explosively excitated
by the stimulus, the transformation of the energy involved
likewise bears a similar relation.
In both instances the transformation of energy, constant in the
resting state, is by the impact of the stimulus suddenly increased.
The dynamic method of investigation of the excitation process
with its physical indicators, forms, therefore, in many respects
an excellent addition to the chemical analysis. A development,
that is, exothermic formation, of energy can only occur in a
chemical process when the chemical affinities which are to be
combined are stronger than those which have been separated.
When this process is brought about by a simple impact, the energy
value of which bears no relation to that of the quantity of energy
in the process itself and which occurs with explosive rapidity,
then it can be simply a question of a liberation process, that is,
a process by which the impact brought about a conversion of
latent chemical energy into that of kinetic energy. The comparison
of the functional excitation process with that of an explosion
does not, therefore, consist in a merely superficial analogy, but is
founded on the same dynamic principles.
When we study the chemical process which occurs in the explosive
transformation of potential into kinetic energy we find two
types of chemical processes. The first type includes the synthetic
processes. For this, the synthesis of water from explosive gas
may serve as a simple example. Here the weaker affinities in
comparatively simple molecules (H + H and O + O) are separated
and stronger affinities are combined in the formation of
more complicated molecules (H + O + H). The second type
represents the process of cleavage. As example for the latter, the
explosive disintegration of nitroglycerine may be quoted. Here
the atoms, held together in a complex molecule by weaker affinities,
are changed by transposition of nitroglycerine. For instance,95
the hydrogen atoms loosely combined with carbon enter
into strong combinations with oxygen and the oxygen loosely
combined with the nitrogen enters into strong combination with
carbon, so that water and carbon dioxide are formed and nitrogen
and oxygen set free.
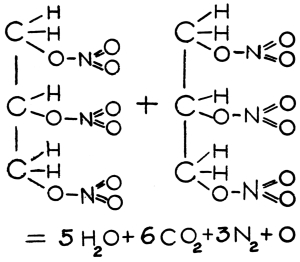
In the functional disintegration of living substance, the last
type is realized. Living substance contains loose complex combinations,
and we know that functional disintegration is accompanied
by the consumption of these organic combinations. In
the functional disintegration of muscle substance the nitrogen-free
groups are concerned, and we must, consequently, first consider
the carbohydrates. However, without further study we
should not generalize from that which is true in the case of
muscle. There are other forms of living substances which contain
different combinations, which disintegrate as a result of the
contact of a stimulus and yield carbon dioxide. A clue as to
which combinations in individual cases undergo disintegration
as a result of excitating stimulation, is furnished by the metabolism
of rest in the particular substance. Plants and micro-organisms
have been investigated more thoroughly in this connection
than animals. Plant physiology has demonstrated that96
the material employed for the CO2 formation and with it the
production of energy is carbohydrate, but that, on the other hand,
various plant organisms and protistæ also use a quantity of other
substances, such as fats and protein, indeed even such comparatively
simple organic combinations as alcohol, formic acid and
methane. It may be accepted that in all these various instances
of excitation of the functional metabolism as a result of stimulation,
the specific respiratory material of the substance concerned
is used in greater amount in the decomposition and likewise
invariably yields carbon dioxide.
The point of most essential interest for the analysis of the
excitation processes is, above all, the mechanism of the organic
combustion and the associated energy production. Here we may
base our observations on the disintegration of carbohydrates,
which is most extensive in the animal as well as in the vegetable
kingdom. We may now ask how dextrose, for instance, disintegrates
in the living system into carbon dioxide, for it is this,
or a sugar of similar chemical nature, which is generally concerned.
Plant physiology, which here, as in many other respects,
is in advance of animal physiology, has indicated two ways by
which this can be accomplished in the living substance. One is
oxydative, the other, anoxydative disintegration.
In the oxydative disintegration of dextrose, taking place in
aërobic organisms, if sufficient quantities of oxygen are present,
there occurs a splitting up of the carbohydrate molecule, as a
result of the introduction of oxygen, into simpler substances and
finally into carbon dioxide and water, just as the dextrose molecule,
when subjected to oxydative processes, is split up into simpler
molecules. In the living substance the oxydases play the
important rôle of oxygen carriers. It cannot be denied, however,
that up to now no carbohydrate splitting oxydases have been
obtained from living substance. This, of course, does not prove
its nonexistence. But this deserves consideration in connection
with an assumption very widely spread among plant physiologists
in regard to the aërobic disintegration of the carbohydrate molecule,
which I shall touch upon presently. If we suppose that
oxydases exist, which bring about primarily the oxydative disintegration97
of the dextrose molecule, its first point of attack must
obviously be sought in the aldehyde group. Here would be situated
the activator, as it were, for the whole carbon chain, from
which, as by a spark, the entire series of links would be ignited.
In an anoxydative disintegration of dextrose as observed in
anaërobic as well as in aërobic organisms, provided the latter have
an insufficient supply of oxygen, the dextrose molecule, by
enzymic action as a result of the splitting off of carbon dioxide,
is converted into substances having a comparatively large carbon
content. The best-known example of this anoxydative disintegration
is the formation of alcohol by fermentation in which
the dextrose molecule is split up by the yeast into alcohol and
carbon dioxide. (C6H12O6 = 2C2H5OH + 2CO2.) Instead of
the production of alcohol and CO2 we may have other enzymic
actions with the formation of other carbon-containing disintegration
products, such as lactic acid, fatty acids, hydrogen, etc. Of
course in such an anoxydative disintegration, which does not lead
to the formation of such simple combinations as carbon dioxide
and water, the quantity of energy set free is much less in amount
than in complete oxydative decomposition, the energy production
of the alcohol fermentation being only 11 per cent of the latter.
In order to produce the same amount of energy as in the former,
a much greater number of molecules is required. We find, therefore,
that the anoxydative type of disintegration develops either
only where the respiratory substances are present in sufficient
amounts, as for instance, in the case of yeast cells, existing in
nutritive solutions rich in sugar; or where the chemical and
energy transformations occur only to a limited extent, as, for
example, in the presence of low temperature. In this respect
Pütter59 has demonstrated in the leech that at a higher temperature,
the oxydative, at a lower, the anoxydative, decomposition
predominates. These are important facts in that they show us
the superiority of oxydative to that of the anoxydative disintegration
in the cell economy. This is of particular interest when
we consider those organisms in which great demands are made98
upon the capability of movement, above all, in homothermous
forms, the metabolism of which takes place on a continuously
high level. For this reason, in homothermous animals the respiration
of oxygen is the almost exclusive source of energy
production.
The previously mentioned facts make it clear that in one and
the same form of living substance both oxydative and anoxydative
decomposition processes are found, depending upon the conditions.
This does not apply merely to the individual organic
forms, such as the facultative anaërobic organisms, but generally
to all aërobic living substance. If oxygen is withdrawn from an
aërobic organism the disintegration does not cease in consequence.
In place of the oxydative we have anoxydative decomposition.
The various aërobic organisms are, however, adapted
in very different degrees to the possibility of an anaërobic existence.
While the facultative anaërobic organisms can continue to
exist without oxygen, the homothermous animals become asphyxiated
in a very short time in the absence of oxygen, in that they
are poisoned by the products of the anoxydative decomposition,
which are eliminated with much more difficulty than carbon
dioxide and water. The fact, however, that disintegration also
continues in an anoxydative form, if oxygen is withdrawn, has
given rise to the thought, which has been accepted especially by
plant physiologists with great readiness, that the decomposition
of organic respiratory substances of the aërobic organisms invariably
takes place in two stages; in that the dextrose molecule—to
again use this as an example—is split up first by an enzyme into
larger fragments, which then in the second stage of the process
undergo combustion to the formation of carbon dioxide and
water. Such a possibility cannot be repudiated. I wish, however,
to state that one should be very reluctant in generalization
of this assumption for all aërobic organisms. The types of metabolism
in the different organisms are so manifold and of such
immense variety that we should be very careful in our generalizations
before being in possession of material extending over a
great number of groups of organisms. Above all, it does not
seem justifiable to also accept this type for life existing at higher99
temperatures, and still less to apply it to those instances in which
the production of energy following stimulation is suddenly increased
to great amounts. Let us suppose that the disintegration
process occurs in two phases, the first of which after the type of
the fermentation of dextrose separates the molecule into larger
fragments, while in the second phase these fragments are split
up through oxydation into the formation of carbon dioxide and
water. We can then say with certainty that in the first stage only
a comparatively small amount of energy production occurs, for
energy production by enzymic processes of this kind is never
great; the second phase, on the other hand, must be associated
with a very considerable energy production, for by the addition
of oxygen and the formation of carbon dioxide and water the
strongest affinities possible are combined. With this assumption
in certain cases, as, for instance, in the sudden production of
energy in muscle contraction, which necessarily occurs in the
purely oxydative phase of the whole process, the view is forced
upon us, that, in these cases, the entrance of oxygen into the
molecule from the very beginning, even the first impact, produces
oxydative decomposition of the whole molecule. The view that,
in the reactions of warm-blooded animals, which occur with great
rapidity and considerable energy production, the oxygen primarily
explosively breaks up the whole carbon chain, certainly
presents no more difficulties than the supposition that the simpler
substances are attacked secondarily, provided sufficient oxygen
be present. This method would be obviously the simplest.
This is, however, mere speculation and a definite decision between
the two possibilities cannot be made at present. However,
whether the process takes place in two phases, an anoxydative
and an oxydative, or simply in an oxydative phase, in any case,
the sudden discharge of energy in the aërobic organism set free
by the stimulus, is brought about by the addition of oxygen.
This is a highly important fact and as such requires the most
thorough confirmation, and is best accomplished by the investigation
of the state of excitation of aërobic substances on the
withdrawal of oxygen. Experience gained by observation in this
respect on a great number of living substances shows that excitability101100
decreases upon the withdrawal of oxygen. In this connection
I should like to cite some particularly significant instances.
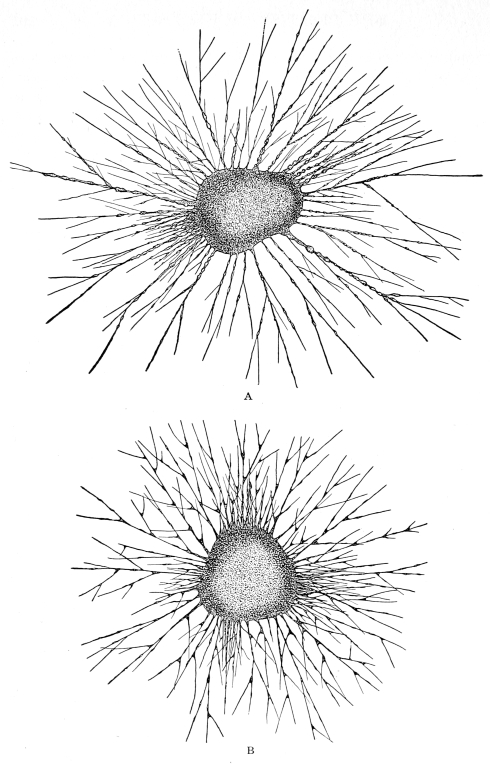
Fig. 10.
Rhizoplasma kaiseri. A—Under normal conditions.
B—In an atmosphere of pure hydrogen.
During a sojourn at the Red Sea in 1894–95 I was able to
establish this dependence in the single-celled organism, the Rhizoplasma
Kaiseri, a large naked orange-colored rhizopod. (Figure 10, A.)
Mechanical stimulation, which under normal vital conditions
of these organisms brings about contraction in the long-branched
pseudopods, becomes ineffective with a cessation of the
movement of protoplasm, when oxygen is removed and is replaced
by a stream of hydrogen. (Figure 10, B.) With renewed introduction
of oxygen there is a return of the protoplasmic movement
and entire recovery takes place.
This dependence of irritability upon oxygen is most clearly
demonstrated in the nerve centers. For this purpose I have
employed the spinal cord of the frog.60 A canula is introduced
and fixed into the aorta of the animal and the blood is replaced
by a current of oxygen-free saline solution. The centers of the
spinal cord are thereby wholly isolated from the supply of oxygen.
The indicator for the irritability here used is reflex excitation
from the skin to the gastrocnemius, or better, stimulation
of the central stump of the sciatic nerve with single induction
shocks, bringing about reflex response of the triceps. The reflex
may be considerably augmented by increasing the reflex excitability
of the spinal cord by poisoning the animal with strychnine.
On testing the reflex excitability at the beginning of the experiment
it will be found that the reaction to each individual stimulus
consists, in consequence of the strychnine poisoning, of a long-continued
maximal tetanus. The longer the deficiency of oxygen
continues, the briefer become the tetanic reflex contractions following
a single stimulus. Soon reflex tetanic responses are
merely short single contractions, which decrease more and more
with the continuance of oxygen deficiency. Finally, the same
stimuli which previously produced strong tetanic contractions
of long duration are altogether without effect. Although by102
increasing the intensity of stimulation brief contractions can
again be brought about, irritability decreases more and more,
until at last even the strongest stimuli remain without result.
If the oxygen-free saline solution is now replaced by one saturated
with oxygen, or blood of the ox, rendered arterial, the
excitability returns within a few minutes and soon reaches the
maximal height which it possessed under the influence of the
strychnine poison. Even the weakest single stimuli now again
produce tetanus. The same process reoccurs, if the fluid used
for transfusion containing oxygen is again replaced by an oxygen-free
saline solution. In this way, by repeated change of the perfusing
fluid, we can demonstrate in the most positive manner this
alteration in irritability, the result of the alternate presence and
removal of oxygen. This is perhaps the best example of the
close dependence of irritability on oxygen.
This same fact can be observed with equal clearness in the
nerve. At my suggestion H. v. Baeyer61 showed as the result of
investigations made in the Göttingen laboratory the dependence
of irritability of the nerve upon oxygen for the first time. By
employing as the method the ascertainment of the threshold of
stimulation I then made a closer study of the alterations in irritability
during asphyxiation. These observations were soon after
continued by Fröhlich.62 The method is as follows: the nerve of
a nerve-muscle preparation of the frog is drawn through a glass
chamber which is made completely air-tight and containing platinum
electrodes. The air in the chamber is then displaced by a
stream of pure nitrogen. (Figure 11.) On testing that part
of the nerve situated within the glass chamber with single break
induction shocks it can be observed that its irritability, measured
by the threshold of stimulation for muscle contraction, decreases
more and more, until after the lapse of some hours, the stimulation
required is so strong as to reach the region of the “Stromschleifengrenze.”
If in place of the stream of nitrogen, air or
pure oxygen is now allowed to flow through the chamber, the103
nerve recovers almost instantaneously. Within the space of a
minute its irritability has risen again to its full height and the
same experiment, with the same result, can be repeated. Finally,
as Fillié63 has shown, the like result is obtained when the nerve is
asphyxiated in a fluid medium.
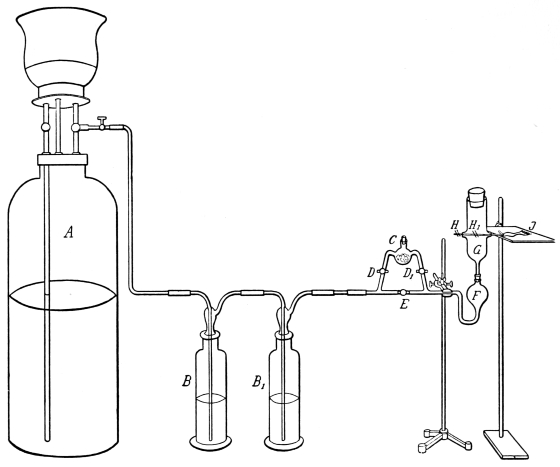
Fig. 11.
Arrangement for asphyxiating the nerve. A—Gasometer containing pure nitrogen. B and B1—Vessels for
washing the gas. C—Ether chamber for eventual experiments with narcosis. D, D1 and E—Glass
faucets. F—Moist chamber. G—Asphyxiation chamber. H and H1—Two pairs of electrodes over
which the nerve is laid. I—Nerve muscle preparation.
All these facts, the number of which indeed could be increased
greatly for other aërobic forms, suffice to establish the fundamental104
importance of oxygen to the maintenance of irritability of
living substance. Oxygen is of greatest importance for a high
degree of irritability in all aërobic organisms. All living systems
which are characterized by a great capability of activity and
evince strong responses under the influence of stimulation, such
as the vertebrates and insects, are necessarily aërobic, whereas
the living organisms of pronounced anaërobic character, as some
bacteria, yeast cells, parasitic organisms, etc., manifest on the
average much less capability of activity.
Finally, to briefly summarize the foregoing, the following
picture presents itself of disintegration produced by a momentarily
acting stimulus. It is immaterial how the stimulus produces
an excitating effect in the given case, whether through
changes in the ion concentration of the living system, by increase
of intramolecular atomic movement or in any other manner, it
invariably accelerates the disintegration of the complex molecules
concerned in functional metabolism, the nature of which
varies in the special cases. In the great majority of instances
nitrogen-free organic combinations serve as material for the
functional constituent members of metabolic processes. In the
anaërobic organisms this decomposition takes place anoxydatively
with the coöperation of enzymic processes, and as larger
fragments generally result from the disintegration of the complex
molecule, the production of energy is accordingly smaller.
The disintegration of aërobic organisms, on the other hand, occurs
in the form of an oxydative splitting up of the complex molecules
into carbon dioxide and water so that the production of
energy attains a high value. The details concerning the manner
in which the individual stages of this decomposition take place
and the interactions by which its end products are reached is
at present beyond our knowledge. It would be a mistake to generalize
in this connection from the behavior of certain groups of
organisms. The assumption that under certain conditions the
disintegration occurs in two phases, the splitting up resulting
from enzymic action of the complex molecule into larger fragments,
followed by an oxydative splitting up of these into carbon
dioxide and water, can in no case as yet be justifiably applied to105
all conditions and all aërobic organisms. This is more or less the
impression which we derive of the functional excitation process
as seen today.
Under normal conditions the functional excitation is at once
followed by a succession of secondary processes, the “self-regulation
of metabolism.” Self-regulation after a functional
excitation is a fact demonstrated by experience. But in what
manner does it take place in detail?
As the functional constituent members of metabolism involve
a disintegration of the nitrogen-free atom groups, the functional
self-regulation must necessarily furnish in sufficient quantity and
in proper form the carbon, hydrogen and oxygen atoms, which
have been removed in the production of carbon dioxide and water.
This is accomplished, as is well known, by the food and the intake
of oxygen. It is of importance to the maintenance of living substance
that after every functional activity it is as soon as possible
again capable of reaction. Therefore, it is absolutely necessary
that this material is in the proper place, where building up is
essential, and is at the same time constantly in proper form.
Indeed, the restitution of the original state follows under favorable
conditions with lightning rapidity, although varying in different
forms of living substance. This occurs in the nerve in
an extremely short time. From this it might be supposed that
the living system by accumulating a store of the necessary compensation
substances in suitable form, had made itself independent
to a certain degree of the frequently varying supply of
material obtained from the medium.
This may be held as the proper view, first with regard to compensation
substances. The fact that living organisms can under
some conditions remain for a lengthened period in a state of
starvation, without losing their capability of activity, can only
be explained by the presence of a great quantity of reserve supplies
of compensation substances. In the course of work in the
laboratory every physiologist has become acquainted with the
fact that frogs which have been kept without food for a year,
although much reduced in weight, are still capable of some
muscular activity.
106
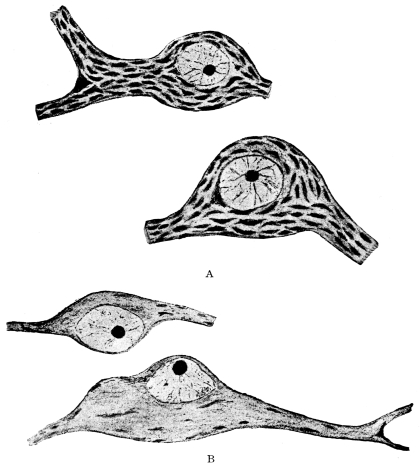
Fig. 12.
Motor ganglia cells from the spinal cord of the frog. A—In normal state.
B—After an asphyxiation lasting 8 to 9 hours. (After Gordon Holmes.)
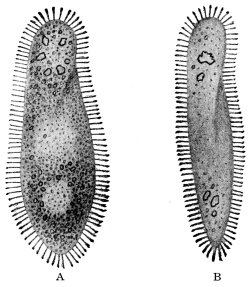
Fig. 13
Paramecium aurelia. A—In normal state. B—In a state of starvation.
Organs and tissue, which are cut off from all food supply
through the blood and lymph, may remain active for many hours.
H. v. Baeyer64 has shown that the ganglion cells in the frog, in
which saline solution was transfused at room temperature and
containing no trace of organic substances and where irritability
has been increased to the maximal by means of strychnine, were
capable of strenuous work for nine or ten hours before losing107
responsivity. The nerves and muscles of the animal retain their
excitability for even a longer period under the same conditions.
Indeed, we have histological evidence of the existence of organic
reserve material in the various cells in the form of embedded
bodies in the protoplasm. As for instance the disappearance of
the Nissl granules in the ganglion cells following great activity,65
(Figure 12), or that of the granules in infusoria cells during
starvation.66 (Figure 13.) We assume that a certain amount of
organic foodstuffs in a state properly prepared is present in the
cell. As the amount of these prepared substances is consumed,
new quantities of stores, having undergone various preparatory
processes, among which the enzymic actions may be considered
to play a chief rôle, are brought into that form in which they
appear suited to fill the gap produced by disintegration. Plant
physiologists in particular have here again furnished us with some108
essential data for the assumption of the existence of such processes
which regulate the transformation of reserve substances
as well as its extent. Pfeffer67 has found in several fungi and
bacteria that there exists a compensation between the diastatic
breaking down of the carbohydrates stored as reserve material
and the quantity of dextrose introduced. He further found that
the more the reserve substance is split up into dextrose the less
of the latter is introduced from without and vice versa. De Bary68
some time ago also observed in the bacillus amylobacter an analogous
relation between the enzymatic cellular digestion and the
quantity of dextrose introduced with the food. An equilibrium,
therefore, exists between the required amount of dextrose and
the extent of enzymic splitting up processes of the reserve material.
A great number of similar processes have been observed.
Even though the details of the whole preparatory assimilative
processes are beyond our knowledge we can still say with certainty
that, on the one hand, everywhere great quantities of organic
reserve substances are always present in the cell, and on the other,
that these substances are subjected to a transformation into suitable
material for building-up processes, the extent of which is
controlled according to need, by the processes of self-regulation.
Entirely different is the question if the cell also possesses a
reserve store of oxygen. In this respect views have widely
differed, and even today no conformity of opinions has been
arrived at. The fact that many purely aërobic organisms and tissues
can exist under complete exclusion of oxygen for a longer
or shorter period, retaining their excitability and producing carbon
dioxide, has for a long time led a great number of investigators,
such as Liebig, Matteucci, Engelmann, Pettenkofer and
Voit, Claude Bernard, Verworn, H. v. Baeyer and others, to the
supposition that a reserve store of oxygen must exist in the living
substance which maintains its excitability for a time. More
recent information, however, of the transition of the oxydative
to the anoxydative disintegration under a deficiency of oxygen,109
as can be observed in plants and certain invertebrate animals,
indicates that here also there is the possibility of another explanation
of these facts. Various attempts have been made to solve
the problem if reserve oxygen is present in the cell or not. The
experiments of Rosenthal,69 carried out with his respiration calorimeter,
seemed to point directly to an oxygen reserve in the
organism of the mammal. He observed that during respiration in
an atmosphere rich in oxygen the respiratory quotient (CO2 : O2)
became lower than in ordinary air, that is, that oxygen, and that
indeed in considerable quantity, must be retained in the organism.
Nevertheless Falloise70 found that when rabbits, which had been
kept in an atmosphere containing 80 per cent of oxygen, were
asphyxiated, the time necessary to produce death was no longer
than in animals which had been kept previously in ordinary air.
The correctness of the observations of Rosenthal have been disputed
by Durig.71 Winterstein72 also, employing the microrespiration
methods of Thunberg upon the spinal cord of the frog,
believed that he had found proof that an oxygen reserve cannot
take place. He reasoned thus: If the cells of the spinal cord
contain reserve oxygen, which is used up when pure nitrogen only
is breathed, then it necessarily follows that after reintroduction
of oxygen, following asphyxiation, a definite quantity must be
stored up again as reserve. In consequence, the respiratory
quotient following the intake of oxygen after asphyxiation should
be smaller than when the animal is in air. He found, however,
that the respiratory quotient does not essentially change and concluded
from this that storage of oxygen does not take place.
In these experiments, however, there exists no certain indicator
as to the state of the spinal cord during asphyxiation and recovery
in the given case. The spinal cord may be severely injured and110
even undergo degeneration during asphyxiation, and the recovery
following the reintroduction of oxygen may be either incomplete
or nil, without there being a method for its determination. Apart
from this, Lesser73 has already emphasized, in opposition to these
experiments, that the respiratory quotient in recovery is no criterion
to guide us. It is immaterial whether during asphyxiation
oxygen respiration occurs following a reserve supply, or that
an anoxydative formation of carbon dioxide has taken place,
for in both instances the respiratory quotient would be less after
asphyxiation when there is again an oxygen supply. It is, therefore,
quite impossible to decide the question by the employment
of this method. For this reason Lesser has attempted to solve
the problem by means of quite another method, and was convinced
that he had refuted finally the belief in the existence of
reserve oxygen. His method consists in the employment of the
Bunsen ice calorimeter, by which he determines the heat production
of frogs, kept first in air, then in nitrogen, and at the
end of each experiment ascertaining the amount of output of
carbon dioxide, respectively in air and nitrogen. He found that
the quantity of heat, calculated in terms of 100 grms. body
weight per hour, produced in nitrogen was considerably less than
that under corresponding conditions in air, but that the production
of carbon dioxide, on the other hand, during the first hours in
nitrogen was doubled in amount, as compared to that in air.
From this he concludes that the carbon dioxide formation in
nitrogen must be different from that in air, as it is associated
with a reduced heat production. In other words, carbon dioxide
formation, while the animal is in a nitrogen atmosphere, does not
have its origin in oxydative processes at the cost of stored up
oxygen. I regret that I am unable to accept these arguments as
conclusive evidence against the assumption of an oxygen reserve,
as this question cannot be decided by the use of such
methods. Lesser does not measure the amount of carbon dioxide
until the end of his experiments, that is, he learns merely the111
entire carbon dioxide production during a period of many hours.
No conclusions can be drawn from this as to the conditions
existing in the first period of time, directly after the animals
have been subjected to an atmosphere of nitrogen. It is quite
possible that subsequent to the change to nitrogen an oxydative
carbon dioxide formation may have continued in decreasing
degree, without this being shown in the final result. The problem
of the existence of a reserve supply of oxygen is in no way
solved by these experiments.
In assuming the presence of a reserve supply of oxygen in the
cell we must above all entertain no false conception as to its
amount. This must be, as I have often had occasion to emphasize,
exceedingly small and in no way comparable with the great
masses of organic reserve substances contained in the cell. The
assumption, especially for the nerve centers of the frog, that the
excitability remains after complete exclusion of oxygen must be
looked upon as demonstrating a reserve supply of oxygen, would
oblige one to suppose the presence of such a small store of oxygen
that it would be completely exhausted by continued activity in
room temperature within ten to twenty-five minutes. Strychninized
frogs, in which the blood has been replaced by an oxygen-free
saline solution, lose, as I have shown,74 their excitability completely
within ten to twenty-five minutes after the blood has
been displaced. Nevertheless the assumption of the existence of
a small oxygen supply in the cell can hardly be evaded. It must
not be imagined that the moment the blood of the frog has been
replaced with an oxygen-free solution, there is not a trace of
oxygen left in the organism. Were such the case, the irritability,
if measured by the extent of the response, would sink momentarily
to a very low level, for the anoxydative disintegration processes
are associated with an incomparably smaller production of
energy than those of oxydative disintegration. We see, however,
that the irritability in the muscles, nerves and nerve centers of the
frog even after the complete withdrawal of oxygen at first remains
practically at the former height and only very gradually112
decreases. Above all it would seem to me to be in the interest
of the preservation of the organism and especially of those parts
in which there is a high energy production and particularly those
substances in which energy production predominates, that the
material necessary for its formation is always at its disposal in
sufficient quantity. Otherwise the capability of action of the
organism would be impaired at every moment or at least suffer
great fluctuations.
In accordance with this we must suppose that under physiological
conditions all those substances required to replace the disintegrated
molecules are always present in the cell in sufficient
quantity and suitable form to replace at once those lost by excitation.
Further, without doubt, in the organism which is always
aërobic, oxygen must be present in certain quantities to assure at
any moment oxygen replacement following oxydative disintegration,
to guarantee sufficient amount for succeeding stimulation.
A further question arises: How is it that the material lost
in disintegration is always replaced in just sufficient quantity
to establish the metabolic equilibrium? In short, how are we
to understand in a mechanical sense the self-regulation of
metabolism?
In the preservation of metabolic equilibrium, we have a process
before us, the principle of which is nowadays restricted to
living substance. In my “Biogen hypothesis,”75 I have associated
the self-regulation of metabolism with the chemical equilibrium
in interreacting masses. I have considered the metabolic self-regulation
as the expression of the formation of a mass equilibrium
between the quantity of foodstuffs and the quantity of a
hypothetical combination of living substance, the biogen, which
continuously disintegrates and builds up again of its own accord.
In fact, however, we have in the chemical equilibrium of reacting
mixtures in the non-living world, a principle which is completely
analogous to the self-regulation in living substance. The chemical
facts are, indeed, well known. If we take the classical example
of the formation of ethylacetat from acetic acid and alcohol,113
we have a case of an inanimate system, in which the amounts of
the reacting substances are in constant equilibrium. The reaction
following the mixture of equal amounts of alcohol and acetic acid
is as follows:
1⁄3 Mol. C2H5OH + 1⁄3 Mol. CH3COOH
= 2⁄3 Mol. CH3COOC2H5 + 2⁄3 Mol. H2O.
In this reaction there is an alteration only in the absolute
quantity of the individual constituents but never in the relative
amount. In the living system we have a completely analogous
instance, which apart from its course differs from the inanimate
example merely in the following points: In the first place, certain
quantities of substances reacting on each other are continually
introduced into and certain reaction products continually removed
from the living system. Secondly, the reacting mixture
of the living substance is not homogeneous, and at the same
time is more complicated than that of the inanimate example.
Thirdly, the sum total of the reaction is not reversible in its entirety.
The question arises, should any essential difference
between metabolic self-regulation and the maintenance of chemical
equilibrium be assumed upon this statement? I must confess
that this does not appear to me to be the case. The fact
that organisms exist in a stream of substances by which their
nutrition is introduced and the metabolic products removed,
cannot have any influence on the state of equilibrium so long as
the conditions are again and again replaced in the same manner.
The equilibrium can only be influenced when the introduction of
foodstuffs or the output of metabolic products is changed in value.
Then they occur as the inanimate example, when various amounts
of material are brought together. A new equilibrium takes place,
having a higher or a lower mass level. This is also true in the
living substance, in growth and in atrophy. The equilibrium is
disturbed as happens in the inanimate reacting mixture, where
different quantities of reacting substances are brought together.
In both instances we have in principle a conformity of behavior
of the inanimate and the living system. Secondly, as far as the
greater complexity and inhomogeneity of the living reacting mixture114
is concerned, it is self-evident that this likewise does not
constitute an essential difference, for we are acquainted with conditions
of equilibrium in chemical reactions possessing a number
of members and in inhomogeneous mixtures. Finally, the fact
that the reaction in the living system is not totally reversible,
forms no barrier to the assumption in principle of metabolic self-regulation
as a chemical equilibrium. It is quite possible to conceive
of a chemical equilibrium in a reacting mixture, of which
only certain constituent processes are reversible, without the
totality of the reactions as a whole being necessarily so. Let us
assume, by way of example, that the assimilative processes of
the metabolic chain are reversible, then under constant quantitative
relations of foodstuffs, following every disintegration of
assimilative products with removal of the decomposition products,
the same amount of assimilatory processes is required for building
up. And this is just that which we observe in metabolic
equilibrium. Accordingly, we may look upon the metabolic equilibrium
as a special, although a very highly complicated, instance
of chemical equilibrium, and we may explain the metabolic self-regulation
following a dissimilative excitation of the same, by
those principles on which the rebuilding of chemical equilibrium
is founded. It is true that the special details of this process can
be differentiated in only that degree in which it is possible to
penetrate at all into the details of metabolism of the given cell
form. In this, as is well known, the advance is extremely slow.
The rebuilding process following decomposition of living substance
in response to an excitating stimulus consists not merely
in compensation for the decomposed atom groups but also in the
removal of disintegration products. This removal can be accomplished,
in so far as simple chemical substances such as carbon
dioxide and water are concerned, by diffusion. Observations
have shown that the semi-permeable protoplasm surface is pervious
to water and carbon dioxide. The latter can, therefore,
depending upon the amount of concentration, be eliminated from
the living substance. Output of water likewise takes place in so
far as the specific water content of the living substance is exceeded
and which is osmotically regulated by its amount of salt115
content. When, finally, osmotic pressure within the living cell
and in the surrounding medium is equal, the interchange of water
ceases. All these processes are explained by diffusion. Self-regulation
takes place in this regard simply by osmotic means.
The conditions in respect to those decomposition products consisting
in more complicated organic combinations, such as lactic
acid, fatty acids and nitrogen derivatives of protein disintegration,
are somewhat different in that the protoplasm surface possesses
the property of hindering the passage of these substances into the
medium. These are, as is well known, first transformed by secondary
chemical processes into transfusable substances. In this
transference the oxydative decomposition with the formation of
simpler substances plays the most important rôle, so that the substances
thereby formed, namely, carbon dioxide, water and
ammonia, are osmotically eliminated as the result of the selective
permeability of the surface of the protoplasm. In this way the
living cell rids itself of the useless products of metabolism.
Finally, the question remains, is the original state, as it existed
before the influence of the stimulus, really completely recovered
by metabolic self-regulation, or does even individual excitation of
brief duration produce a continued change in the protoplasm? It
is quite impossible to prove that such an effect follows the momentarily
acting single stimulus, if stimulation has not exceeded the
physiological limits of intensity. Should it exist, it must be
imperceptible. Nevertheless, it ought to be possible by frequently
repeated application of the stimulus to increase this which is
imperceptible to an extent in which it is perceptible. This is,
indeed, the case and is manifested as we have already seen in
the increase of the volume of living substance by frequently
recurring functional excitation. We can, therefore, assume with
great probability that even the momentarily acting individual
stimulus produces, although not perceptible per se, lasting effect
in the cell. The functional excitation must be followed secondarily
by an increase of the assimilative phase of the entire
cytoplastic metabolism. Otherwise the taking place of the increase
of volume of the living system following frequent excitation
of the functional constituent members of metabolism, is116
unintelligible. But how are we to interpret these secondary
results from a physical standpoint? First of all, it must be stated
that we do not know of such hypertrophy following activity in
unicellular organisms, but only in the tissues and organs of multi-cellular
forms, in muscles, nerve cells, glands, etc. In the cell
community of the vertebrates, however, the studies on the relations
between activity and the blood supply of the particular
tissue or organ furnish a physical interpretation for the existence
of the functional hypertrophy. The active portions show
a dilation of the blood vessels, therefore an increased supply of
blood and consequently an increase in the circulation of lymph.
In other words: the supply of nourishment to the individual cell
and the removal of the metabolic products in a unit of time is
increased. The preceding discussion of the dependence of the
conditions of equilibrium upon the quantitative relations of the
reacting substances makes it clear that under these conditions a
metabolic equilibrium on a higher quantitative level must occur;
that is, the living substance must increase in amount just as in
the inanimate example the absolute amount of the æthylacetat
increases if more alcohol and acetic acid are introduced to an
equal degree. Some time ago76 I expressed the opinion that the
increase of the blood supply in a functionally active organ must
be based on a physical self-regulation, which takes place as a
result of the fact that metabolic products of the tissue cells
influence the cells of the vessel walls in that part, so that the
vessels dilate and more lymph is formed. In the meantime this
has been proved to be indeed the case. Schwarz und Lemberger77
and Ishikawa78 have shown that especially the weak acids, which
are produced in larger amount as a result of strong activity of the
cells, bring about vessels’ dilation. By the demonstration of this
highly important process of self-regulation the last link has been
added for the physical understanding of the hypertrophy of
activity of the tissue cells by continued functional excitation.117
Whether or not the same applies to the single living cell, if the
unicellular organism likewise undergoes a quantitative increase
by a continuous functional excitation, and if the single cell possesses
in itself a corresponding mechanism of self-regulation
similar to the cell community in the vertebrates, cannot be
answered, for concerning all these problems information is lacking
for the present.
118
CHAPTER VI
CONDUCTIVITY
Contents: Only processes of excitation are conducted, not processes of
depression. Conduction of excitation in its two extreme instances.
Conduction in undifferentiated pseudopod protoplasm of rhizopoda.
Conduction of excitation with decrement of intensity and rapidity.
Conduction of excitation in the nerve. Rapidity of conduction of
excitation without decrement. Relation between irritability and conductivity.
Conduction of excitation with decrement of the nerve after
artificial depression of irritability by narcosis. Theory of the decrementless
conduction of the normal nerve. Proof of the validity of
the “all or none law” in the medullated nerve. Theory of the process
of the conductivity of excitation. Theory of core model (Kernleiter).
Electrochemical theory of conduction based on the properties of semi-permeable
surfaces.
When the response to a stimulus is studied in a living system,
whether it be a single cell, a tissue, or a complex organism, the
indicator used, either that of movement, current of action, production
of certain substances, the development of light, of heat
or the alteration of form, is the result of two distinct processes.
The first of these is primary excitation, brought about by the
stimulus at a local point, and the second is an extension of the
excitation to the surrounding tissue. We are not in a position
to experimentally bring about a response to stimulation, in which
the primary excitation occurs and not the secondary process,
that of conductivity. All living substance contains this property,
although to a very different degree, as all living substance possesses
irritability, and this presents the condition not only for
the taking place of the process of excitation but also that of its
conduction.
If I here speak only specifically of the conduction of excitation
instead of the conductivity of response to stimulation this is not119
only primarily for the reason that we intend especially to analyze
the conductivity of excitation on this occasion, but also because
no other effects of stimulation except those of excitation can be
conducted from the part affected by the stimulus to the surroundings.
Although considered on theoretical grounds it appears more
or less improbable that depression is extended from the place of
its origin, it is very easy to convince one’s self experimentally of
the fact that depression following a stimulus is invariably localized
to that portion directly affected by the stimulus. The nerve
furnishes a very favorable object for this purpose. If a nerve
muscle preparation of the frog is made and introduced in the
glass chamber previously described containing platinum electrodes,
and another pair is applied to the nerve between the chamber
and the muscle, it is possible to subject the stretch of nerve in
the chamber to various agents, producing a paralyzing effect. In
this way it may be exposed to an atmosphere of pure nitrogen
for example, or to narcosis as by ether, chloroform, carbon dioxide
and other gases, to an increase in temperature or to other
agents, without these in any way affecting the irritability of the
nerve stretch situated over the electrode between the chamber
and the muscle. The contractions of the muscle, which are produced
by stimulation of the periphery region of the nerve with
stimuli of a definite strength, remain unaltered, even when the
asphyxiated stretch of nerve in the chamber is already completely
degenerated. The central depression of a ganglion cell of a
motory neuron is likewise wholly without influence on the degree
of excitability of its nerve fiber, as I was able to demonstrate79
in the reflex inhibition of the motor neurons of the spinal cord
of the dog. (Figure 14.) That which is conducted by the
nerves is solely the process of excitation.
It is our task to analyze in detail the conditions involved in the
conduction of excitation in order to obtain a deeper insight into
the physics of this process. A comparative survey of a series of
various types of living substance shows us that they differ in120
respect to the conduction of excitation in the following points:
In regard to the rapidity with which the excitation is conducted,
the extent of the area over which it spreads, and the intensity
with which it extends. These conditions may be best illustrated
by citing two extreme examples. The one is formed by the
rhizopods, the other by the nerve fibers. Between these two
extremes we have manifold gradations in the conditions of conductivity.
Not all cell forms are suitable objects for the study of
conductivity. There are forms of rhizopods which are as favorable
to investigation as the nerve; this is due to the fact that,121
although they are often of microscopic dimensions, they possess
elongated fingerlike or threadlike pseudopods.
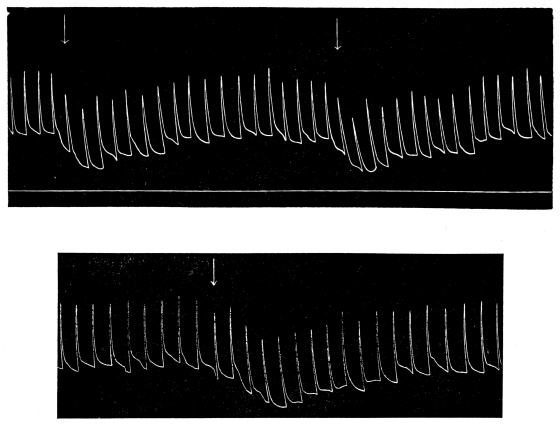
Fig. 14.
Contractions of the musculus extensor digitorum communis longus of the dog, brought about by rhythmic
stimulation of the nervus peroneus. The muscle is in the condition of tonic excitation which
proceeds from the center. The arrows indicate the point where reflex inhibition of the central tonus
is produced. The height of the single contraction undergoes no diminution.
Indeed, a rhizopod cell, with its straight, elongated pseudopods,
is preëminently fitted as an object of comparison with a neuron.
Although the difference in respect to the individual points is so
far-reaching, still, based on their outward morphological similarity
various physiological parallels in both are forced on our observation.
A comparison of the rhizopod cell with the neuron can
consequently guard us from many erroneous generalizations
which we might be inclined to deduce from a one-sided investigation
of the nerve. This is especially the case in regard to the
conductivity of excitation, which was formerly studied almost
exclusively on the nerve and only occasionally on the muscle,
which offers similar conditions. The nerve, in which the function
of the conductivity of excitation is particularly highly developed,
was considered at the same time as the type in which this
process could be most readily analyzed, and from which it was
believed general information of the process of the conductivity of
excitation could first be gained. This view has led to serious
errors, as the nerve, resulting from the high development of its
conductive capability, shows quite one-sided specialized conditions,
which can by no means be transferred to other forms of living
substance.
A very suitable object among rhizopods for the study of conductivity,
and which is everywhere easily procured, is Difflugia.
This species living in small pools has a delicate urn-shaped, pear-shaped
or flask-shaped capsule built up of sand grains, diatomes
or material produced by the organism itself. From the opening
the protoplasm extends often to a considerable length its finger-shaped
hyaline pseudopods. When Difflugia is placed in a flat
dish in water and observed under the microscope, it is frequently
seen to extend from the opening long pseudopods in exactly opposite
directions, which reach for a considerable distance on the
bottom. These offer particularly favorable conditions for the
study of the conduction of excitation. When this animal is
placed under a microscope, the pseudopods are very readily stimulated
at any position to a desired extent by means of a sharp122
needle, to which fat has been previously applied and subsequently
the excess removed. The extension of the response
from one point toward the other can then be followed with
great ease. The pseudopod of the rhizopod has the great advantage
over the nerve that its excitation can be directly observed.
The excitation following weaker stimulation is manifested by
a wrinkling of the previously completely smooth surface; stronger
stimulation produces differentiation of the hyaline protoplasm to
a strongly refractive strand in the axis and a turbid myelinlike
mass at the periphery, the pseudopod at the same time retracting
toward the central cell body. In spite of all these occurrences
being of microscopic dimensions, still with some practice it is
quite possible to experiment on them under the microscope. In
this way I found it comparatively simple to study the fundamental
principles of conductivity.80
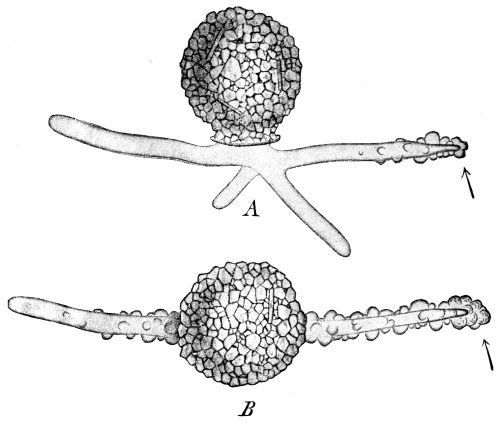
Fig. 15.
Difflugia urceolata. A—Weak local stimulation at the end of a long extended pseudopod.
B—Stronger local stimulation applied to the end of a long pseudopod.
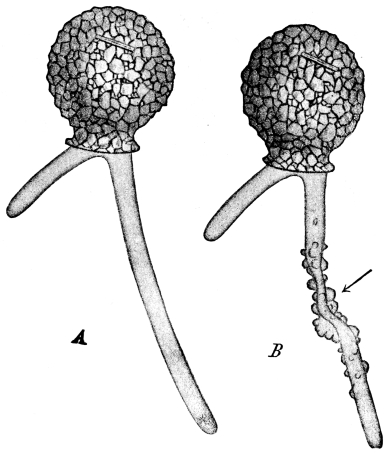
Fig. 16.
Difflugia urceolata. A—In non-stimulated condition. B—The same individual
locally stimulated in the middle of a long extended pseudopod. The excitation
spreads in both directions, centripetal as well as centrifugal.
All these factors, the intensity with which the excitation
extends from the point of stimulation, the rapidity of the extension,
and finally the area over which conduction takes place, are
manifestations of the intensity of stimulus, and as such alter
with these in corresponding manner. If the end of a pseudopod
is barely touched and thereby weakly stimulated, the response is
restricted to a slight wrinkling of the surface, which slowly extends
to the immediate neighborhood, whilst the more distant
parts of the pseudopod are not affected at all by the excitation.
(Figure 15, A.) On a stronger stimulation of the pseudopod by
slight pressure, the response is likewise stronger, and the characteristic
differentiation of the protoplasm, consisting in the
strongly refractive strand in the axis and the turbid myelinlike
outer mass, appears at the point of stimulation. From here a
peculiar alteration spreads gradually further over the pseudopod,
in that first upon its smooth surface a few myelinlike droplets
are seen, which become larger and with the development of the
strand in the axis, dissolve into a wrinkled mass on the surface.
The further this process extends from the point of stimulation,
the weaker it becomes and the more slowly it proceeds, until at123
last there is complete disappearance. (Figure 15, B.) The
pseudopod has at the same time retracted to a considerable
degree. If a still stronger stimulus is applied by firm pressure
at the end of the pseudopod the process takes place with much
greater violence. The differentiation of the protoplasm spreads
centripetally from the point of stimulation over the whole
pseudopod with great rapidity, and produces a quick retraction
in the same, then involves the oppositely directed pseudopod,
in which it then extends more and more slowly, until, proceeding
in a centrifugal direction, it is at last gradually completely obliterated.
When strong stimulation is applied, the process occurs
with such rapidity that the contraction of the pseudopod is
almost twitchlike. As the rapidity of the conduction alters124
within a wide limit according to the strength of the stimulus
and the distance from the point of stimulation, it is self-evident
that no constant figure can be stated. To give a general idea of
the rapidity, they might be estimated according to observations
I have made with second watch and ocular-micrometer as from
within a slight fraction to that of a millimeter in the second.
When a very long extended pseudopod is locally stimulated in
the middle, the response spreads from the point affected in both
directions diminishing in intensity and rapidity. The excitation
extends equally in all directions. (Figure 16.) These facts125
show very clearly that in Difflugia the excitation following a
localized stimulus is dependent on the intensity of the stimulus,
and that according to the degree of this, the wave progresses
in either stronger, more rapid and extended, or weaker,
slower and more limited manner. With the greater distance
from the point of stimulation the excitation undergoes an increasing
decrement of its intensity and rapidity of conduction. Different
species of Difflugia which I have investigated, Difflugia
lobostoma, urceolata, pyriformis, have shown a complete conformity126
in this respect. A great number of other fresh water and
marine rhizopods, the pseudopods of which I have used for analogous
experiments, although differing in the manner of reaction
in regard to the extent and rapidity of the course of excitation,
manifest exactly the same fundamental principles. A very
favorable form is, for instance, the much smaller Cyphoderia
margaritacea, which is distinguished by a somewhat higher degree
of excitability and rapidity of reaction.81 The long straightly
extended pseudopods are thinner and more threadlike than those127
of Difflugia and show upon stimulation as a result of their local
excitation a simple contraction into clumps of the stimulated
protoplasm without the characteristic differentiation of that of
Difflugia. (Figure 17.) In the case of the marine rhizopods,
Orbitolites (Figure 19), Amphistegina, etc., which I investigated
at the Red Sea, the conduction of excitation takes place also as
in Difflugia with a decrement of intensity and rapidity becoming
larger with the distance from the point of stimulation until the
wave of excitation is obliterated.
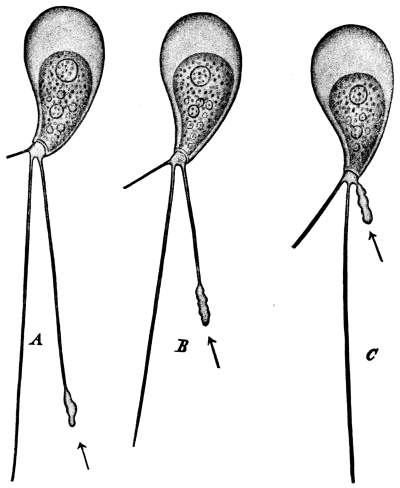
Fig. 17.
Cyphoderia margaritacea. Result of localized mechanical stimulation at the end
of a long extended pseudopod. A, B, C—three successive stages.
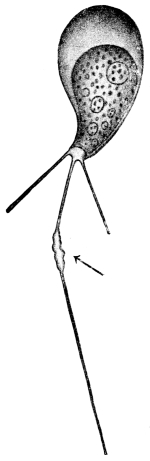
Fig. 18.
Cyphoderia margaritacea. Result of localized mechanical
stimulation in the middle of a long extended pseudopod.
Fig. 19.
A pseudopod of Orbitolites complanatus (cf. Fig. 7). a—In normal condition.
b—Severed by a cross section near the end. b-f—Five successive stages
of the effect. b-d—The pseudopod retracts by centripetal flowing of the
protoplasm contracted in the shape of microscopic balls and spindles. e
and f—The pseudopod begins to extend again. The centripetal flowing
balls and spindles begin to disappear.
128
A sharp contrast to this type is formed by the other extreme
as represented by that of the medullated nerve. As an indicator
of the course of excitation we will take the action current in
an isolated nerve of the frog. If this is stimulated at one end,
we can test the intensity of the conducted excitation by leading
off the action current from two points at varying distances from
the one influenced by the stimulus. Since the classical discovery
of Du Bois-Reymond of the action current of the nerve, we know
that in the fresh medullated nerve, if observed under favorable
experimental conditions, no decrement of intensity of excitation
during its course from the point of stimulation along the length
of the nerve can be demonstrated.82 If unpolarizable electrodes
are applied to a nerve in such a position that they are equidistant
from the cross section and are connected with apparatus for
testing the current, it will be found that there exists an “unwirksame
Ableitung” in the sense of Du Bois-Reymond, that is, in
which there is no demarcation current. When a tetanizing current
is applied to one end of the nerve, no difference of potential
between the two nonpolarizable electrodes is observed, which
indeed would be the case if excitation with its current of
action would have a decrement on its way from one to the other
point of leading off the current. This fact, which has been repeatedly
confirmed, shows us that the medullated nerve, under normal
conditions, conducts excitation without a perceptible decrement
of the intensity.
This specific property of a medullated nerve is in conformity
with the conditions in connection with the rapidity of conductivity.
Since Helmholtz83 has devised the method for measuring
the rapidity of conduction in the nerve, this investigator himself
and numerous others have studied the rate in different nerves.84
Helmholtz found the rate for motor nerves of the frog to be
27 meters per second, for the sensory nerves of man 60 meters,129
and the motor nerves of man 34 meters. Other investigators
have obtained quite different results; Hirsch, for the sensory
nerves of man, 34 meters; Schelske, for the same, 25–33 meters;
De Jaager, 26 meters; v. Wittich, 34–44 meters, and Kohlrausch,
56–225 meters; v. Wittich for the motor nerves of man, 30 meters;
Piper85 finally in the most recent investigations about 120 meters
in the second.
These differences may be explained in a large measure by the
variety of the methods used, in part also by the difference in the
structures. The methods employed for the study of the velocity
have also been used to solve the question, whether the velocity of
the excitation wave in its course over the nerve meets with a
decrement as it moves further and further away from the point
of stimulation. Here the endeavor was made to study the difference
in time of the latent period, which is observed by the indicator,
when the nerve is stimulated at two points at different distances
from the muscle, used as an indicator, or from the wires
leading the current to the indicator. The more recent investigators,
as René Du Bois-Reymond,86
Engelmann,87
G. Weiss,88 have
arrived at the same conclusion, that the rate of conductivity in
the medullated nerve under normal conditions is the same at all
distances from the point of stimulation. (Figure 20.)
The medullated nerve shows, therefore, under normal conditions
neither a decrement of its conductivity, nor of its irritability,
as the distance of the wave of excitation increases from that of
the position of stimulation; this means, in other words, that excitation
is conducted with the same intensity with which it is started,
and with a constant rate throughout the entire course of the
nerve.
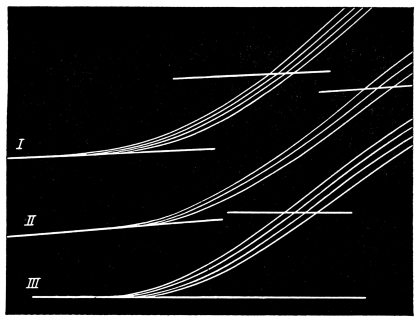
Fig. 20.
Curves of muscle contraction obtained by stimulation of 3 and 4 points situated
at equal distances from each other on the sciatic nerve of the frog. The
increase of length of the nerve stretch corresponds with an equal increase of
the latent period of contraction. From this follows, that the rapidity of the
wave of excitation is the same at all points of the entire length of the nerve.
(After Engelmann.)
There is, nevertheless, a third point of considerable difference130
between the types of conduction of excitation in the rhizopods
and in the nerve. Whereas in the rhizopods the rapidity of conduction
is dependent upon the intensity of the stimulus, it has
been long known as the result of investigation by Rosenthal,
Brücke and Lautenbach and at a more recent date by Gotch89 and
Piper,90 that in the nerve of the frog, as well as in man, the velocity
is not dependent upon the intensity of stimulation. (Figure 21.)
Contrary results have been obtained by a few early observers
wherein the latent period was shorter when the stimulation was
strong. Nicolai91 explains this shortening of the latent period,131
resulting from the application of strong electrical stimuli, to a
spreading out of the “Stromschleifen” from the point of application
and consequently there is a shortening of the stretch of
nerve between the point of stimulation and the indicator.
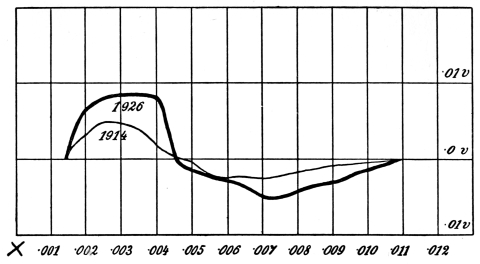
Fig. 21.
Course of the action current of the nerve. The thin line indicates the action current
produced by a weak, the thick line the action current produced by a strong stimulus.
The duration of the action current is the same in both cases. (After Gotch.)
This conspicuous difference in the conduction of the two
extreme types of living substance, which we have already observed,
arouses the question as to what properties of living substance
bring about these differences. In order to answer this
question, it is necessary, first of all, to make some general statements
concerning the processes of conductivity.
As already emphasized, all living substance possesses the capability
of conducting excitations to a definite degree. We may,
therefore, assume that the same fundamental property of conductivity
exists in all substances. A fact to be considered in the
conduction of excitation, is that the primary breaking down of
the complex molecules at the position of stimulation act in turn
as exciting stimuli upon the neighboring portion of the living
substance, which in turn undergoes a similar decomposition. And
so this process continues. This fact is evident from the observations
on the process of excitation. But the nature of the stimulus132
which produces the breaking down of the complex molecules
upon the surrounding molecules is a problem which can only be
studied later. Here only one point will be mentioned in advance
concerning the intensity of the stimulus. It is apparent from the
experiments on the rhizopods, that the greater the intensity of the
stimulus the more extensive must be the breaking down of the
living substance. A stronger primary stimulation must also
secondarily produce a stronger stimulus in the neighborhood. In
other words: the conduction of excitation is a function of irritability.
The greater the irritability, that is, the greater the number
of molecules broken down in a unit of time and space by a stimulus
of a certain intensity, the greater also is the conductivity of
the living system, that is, the stronger, the more rapidly and the
further excitation is extended. Conductivity of excitation is,
therefore, unthinkable without irritability. Both are inseparably
connected. The conclusion forced upon us by this chain of
reasoning admits of no argument. Nevertheless the endeavor
has been made, because of certain evidence at hand, to show that
the property of conductivity could exist without irritability. A
number of authors, such as Schiff,92 Erb,93 Grünhagen,94 Effron,95
Hirschberg96 and G. Weiss,97 have observed the fact that in spite of
a more or less strong decrease of excitability of a stretch of nerve,
stimuli applied above this stretch can still produce a conduction
of excitation through the affected part. They have concluded
from this that it is possible to separate the conductivity from
irritability. Erb and G. Weiss have even gone so far as to directly
express the opinion that capability of conduction and irritability
involve two different histological elements. In contrast to this,133
other investigators, such as Hermann,98 Szpilmann and Luchsinger,99
Gad,100 Piotrowski101 and Wedenski,102 have more or less
decidedly taken the stand that an actual separation of irritability
and of conductivity does not exist. The apparently contradictory
evidence as well as the conflicting theoretical views have been
cleared up by Werigo,103 Dendrinos,104 Noll105 and Fröhlich.106 These
investigators have shown that the length of the narcotized stretch
of the nerve plays an important rôle in the obliteration of conductivity.
It has been found by the application of a stimulus
above the narcotized stretch of nerve, that the longer this stretch
is, the less is the reduction of irritability which obliterates the
excitation wave reaching this area. Further: The shorter
the stretch, the greater must be the reduction in irritability
before this result is brought about. (Figure 22.) In other
words, the conductivity in the narcotized nerve is dependent
upon the length and the irritability of the narcotized stretch.
From this observation the important fact is evolved, that
the wave of excitation meets with a decrement of its intensity
in the narcotized area. This decrement becomes larger as
the wave progresses through the involved stretch. Further it
is progressively increased as the amount of the irritability is
reduced. Finally, when the stretch is long enough, the wave134
of excitation is obliterated. This important fact has been further
established by the experiments of Boruttau and Fröhlich,107
in which they studied the intensity of the current of action,
produced by a wave of excitation, from two points in the narcotized
stretch. The wave of negative variation, brought about by135
the excitation, gradually decreases in the narcotized stretch as
the electrode is further removed from the point of entrance.
Beside a decrement of intensity, as the investigations of Fröhlich108
prove, the wave of excitation shows a decrement of the velocity
in the narcotized stretch. And it is probable that the wave of
excitation extends with progressive reduction in the velocity,
corresponding to the decrement of intensity. The work of Koike109
under the direction of Garten, in which the conclusion arrived at
is that the reduction in the velocity is the same throughout the
narcotized area, should not be accepted as conclusive in spite
of the delicate method employed. These investigations are extremely
difficult, being in the field of the most delicate of present-day
methods. The decrement, which the wave of excitation
meets with in its progress in the narcotized stretch, makes the
conflicting testimony concerning the apparent separation of irritability
and conductivity intelligible. It depends entirely upon
the length of the narcotized area, and the amount of reduction
in irritability on the one hand, and the strength of the stimulus
used for testing the irritability on the other, whether the conductivity
will disappear before the irritability or vice versa. If
I test the irritability in the narcotized stretch with a weak stimulus,
just slightly above the threshold, then by slight reduction in
the irritability complete absence of response occurs, when the
same stimulus is applied. This occurs at a time when excitation
reaches the narcotized area from above and meets with a
decrement so slight that it can pass through the whole narcotized
stretch, that is, when the narcotized stretch is short enough.
If I test the irritability of the narcotized area with a strong
stimulus, far above that of the threshold, irritability will be
found to be present at a time when the conductivity for the excitation,
coming from above, is already obliterated. This is due
to the fact that the decrement in the narcotized area is already
great enough to bring about the complete disappearance of the136
wave of excitation coming from above. This, of course, only
occurs provided the length of the narcotized stretch is great
enough. The separation of conductivity and irritability is, therefore,
only an apparent one. In reality, the facts obtained from
experimentation indicate that with the reduction of irritability the
decrement of the wave of excitation increases, whilst the shorter
the stretch, the smaller is the decrement. This shows that conductivity
is a manifestation of irritability.
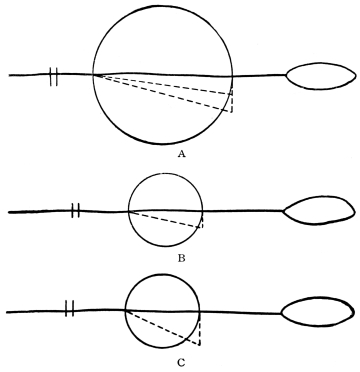
Fig. 22.
Scheme of the decrement of the excitation wave in the narcotized stretch of a
nerve. A—The narcotized stretch (indicated by the cross section of the
chamber) is 30 mm. long. The ordinates of the dotted lines indicate the
amount of the decrement. If the decrement is slight (upper dotted line), the
excitation wave passes the narcotized stretch and increases again on entering
the normal stretch. If the decrement is great (lower dotted line), the excitation
wave is obliterated towards the end of the narcotized stretch and the
muscle remains at rest. B—The narcotized stretch is 15 mm. long. The
decrement is slight. The excitation wave can therefore pass into the normal
stretch and here increase again. C—The narcotized stretch is 15 mm. long.
The decrement is great. The excitation wave is obliterated, therefore, in the
narcotized stretch, and the muscle remains at rest.
The facts just mentioned have, however, a much deeper meaning.
They show us that it is possible by means of narcosis to
convert an extreme type of a living system, with decrementless
conductivity, into another extreme type of living substance, in
which excitation in its progress meets with a strong decrement,
like that seen in the rhizopods. The same results may also be
obtained by asphyxiation and other forms of temporary and permanent
injury of the nerve. We are, therefore, in the fortunate
position in the case of the medullated nerve of having a substance
to study, which, depending upon conditions which are
under our control, may become a type in which conductivity
occurs with or without the presence of a decrement. We can
likewise reduce the irritability to various degrees, producing all
intermediate gradations between the two extremes. This latter
is particularly valuable in that it permits us to study the conditions
in one and the same substance necessary to bring about the
various peculiarities of conductivity. The great differences in
the conductivity of excitation are conditioned by variations in
the degree of irritability. If the irritability of the nerve is at the
normal level the wave of excitation progresses to the end of the
nerve without manifesting a decrement of its intensity or
rapidity.
If the level of irritability of the intact nerve is artificially
reduced, the wave of excitation meets with a greater decrement
and reduces in velocity, and in fact disappears the more quickly
in the stretch of nerve, as the reduction in irritability is increased.
These three factors, irritability, intensity and velocity of the
progress of the wave of excitation, are inseparable. All living
substances may be grouped according to their capability of conducting137
excitation into a long series, starting with those possessing
the least irritability, as we found in the rhizopods, then those
having greater irritability, as the smooth muscle and ganglion
cells, then those with still greater irritability, as the striped muscle,
and finally those having the greatest degree of irritability, as the
medullated nerves of the warm-blooded animal. Should the
processes of excitation, as we saw, result from the energy production
following the disintegration of the labile molecules of
the living substance, then the degree of irritability is determined
by the chemical constitution of the disintegrating molecules, by
the number of molecules which are broken down in a definite
space and a given time, and by the nature of the disintegration
itself. All of these individual components, if we observe the
problem from the physical standpoint, are manifested by the
quantity of energy production. The higher the irritability of a
living system, the greater is the amount of energy production in
a given time and space which the stimulus produces. This has
particular interest from the standpoint of the extreme cases of
medullated nerves of the vertebrates with their most highly developed
conductivity, and which will be analyzed in somewhat greater
detail. How are we to explain their decrementless conductivity?
When we study the decrement of the excitation wave in the
series of living substances, before alluded to, we see that this
reduces with a progressive increase of irritability. Consequently
the extreme irritability of the nerve is a manifestation of its
decrementless conductivity. If we study the course of a process
of excitation and its conduction in its molecular details, the fact
of the decrementless conduction indicates that in excitation, produced
by a stimulus, the same number of specific molecules
capable of disintegration are broken down in the same manner
at every following cross section, as at the point of stimulation;
or in other words: an equal amount of energy is set free at every
cross section, which, in its turn, acts as stimulus to the next,
etc. Such a condition presupposes, however, in an elementary
fiber of the nerve, that by the conduction of the wave of excitation
from cross section to cross section, all those molecules
capable of disintegration are broken down. If it is assumed that138
the entire number of molecules capable of disintegration do not
break down, but only a certain per cent. of the same, then it would
not be possible to conceive of a molecular structure of the nerve
in which this would take place without decrement of the wave of
excitation. With the assumption of a generally homogeneous
molecular structure (Figure 23, a) of the elementary fibers it
would be entirely incomprehensible how, with the decrementless
extension of the excitation, individual molecules capable of breaking
down could escape disintegration. If, on the contrary, the
molecular structure is not homogeneous it only is possible to
explain a conduction, on each cross section of which an equal
per cent. of irritable molecules break down, by the hypothesis
that the irritable molecules are in their turn ordered in fiber-shaped
series (Figure 23, b) within the elementary fiber and are
thus protected to a certain degree from one another and from
transverse conduction of excitation. This hypothesis would,
therefore, only mean that the elementary fiber is not such in139
reality and would thus transfer the difficulty to the ultimate
fiber unit, for which a homogeneous molecular structure would
have to be presumed. In short, whatever may be the assumption
on which molecular structure of elementary fibers is based, the
fact of the decrementless conduction peremptorily demands, from
the physical standpoint, that from cross section to cross section
the entire number of irritable molecules are broken down. This
conclusion is highly important, for it indicates very clearly that
the “all or none law” is applicable to the nerve.

This gives us occasion to return to the discussion of the question,
if living systems really exist which respond in accordance
with the “all or none law.” The medullated nerve forms an
object particularly suited to serve as a starting point for the
treatment of this especially important problem. The question
arises in this connection, if the validity of this law for the nerve
can be tested by other means.
At first it would seem as if the application of the “all or none
law” to the nerve were in contradiction to the well-known fact
that a weak stimulation of the nerve produces a weak, a strong
stimulation, a strong response. In this connection Gotch110 has
pointed out, as the result of experimental studies of the wave
of activity of the nerve, that the difference in response, following
the application of stimuli of varying strengths, is understandable
from the fact that threshold stimuli stimulate only a
few of the fibers of the nerve trunk, whereas progressively increasing
the intensity of the current involves more and more
fibers. There can be no doubt that this factor explains the difference
in the strength of the response. Therefore, in reality we
do not find here a contradiction of the “all or none law.” On
the other hand, the fact that the nerve, in contradistinction to
many other forms of living substance, the ganglion cell, for
example, upon a weak stimulation does not show the phenomena
of summation, even when the stimuli follow each other in a
rapid succession, indicates very strongly that the weakest operable
stimulus produces maximal excitation, so that the response140
cannot be further increased. But above all, there is a series of
facts, which have been gained in the Göttingen laboratory, which
demonstrate apparently without doubt the validity of the “all
or none law” for the medullated nerve. These observations I
wish now to consider in greater detail.
If a nerve of a nerve muscle preparation is drawn through a
specially devised glass chamber so that the middle portion can
be narcotized or asphyxiated and the nerve so arranged that it
rests upon a pair of electrodes in the chamber and upon a second
pair without the chamber and centrally located, then the nerve
can be narcotized or asphyxiated and thereby the alterations in
the irritability as well as the conductivity can be followed. In
order to obtain as distinct a picture of this alteration as possible,
I tested continuously the threshold of stimulation, which just produced
minimal contraction in the muscle, and Fröhlich111 continued
these observations. As a result the following very remarkable
conditions were observed. During the increase of the depth
of narcosis or asphyxia the irritability sinks more and more with
regularity. The conductivity remains unaltered for a long time,
as the strength of the threshold stimulus is not changed until
irritability has fallen to a definite point. When this is reached,
conductivity disappears. (Figure 24.) The most important point
in this connection, however, is, that the conductivity disappears
simultaneously and practically momentarily for the excitation
produced by both weak and strong stimuli. When the stimulation
at the electrode placed centrally to the chamber does not
bring about response for threshold stimuli, maximal stimuli at
the same time also become inoperative. This is a very interesting
point, the importance of which has not until now been recognized.
This fact is not in harmony with the view held until now, that in
the nerve fiber different strengths of stimuli bring about excitation
of different intensity, and are then conducted. Let us now
clearly comprehend this problem.
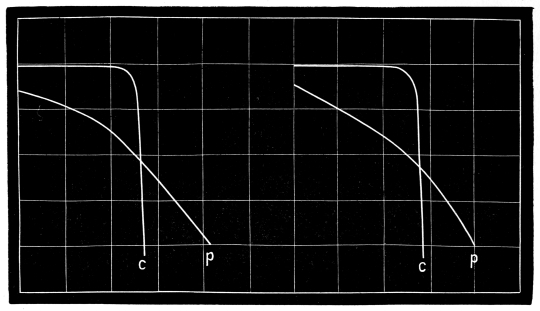
Fig. 24.
Curves of the changes in irritability (p) and conductivity (c) of a nerve under the influence of
narcosis or asphyxiation. (After Fröhlich.)
We have already seen that the wave of excitation meets with
a decrement of its intensity in the narcotized stretch, which141
increases in strength as the irritability diminishes. If the value
of the threshold is learned by stimulating the nerve at the electrodes
centrally placed to the chamber with minimal stimuli, it
would necessarily follow that this weak stimulus would bring
about a corresponding weak excitation of the individual fibers
and the wave of excitation already in the beginning of narcosis
would be obliterated, for it would meet with a decrement, the
result of the reduction in the irritability. A wave of excitation
of minimal strength could under these conditions no longer
reach the muscle, even in the beginning of narcosis. In spite
of this the excitation, even when produced with threshold
stimuli, passes through for a long time, even when the irritability
in the chamber is greatly reduced, as shown by testing
with the electrodes within the chamber. This is not consistent
with the assumption that a threshold stimulus brings about the
minimal excitation, even in the individual nerve fiber. But
further: with a definite decrease of irritability of the narcotized142
stretch the capability of conductivity disappears, and indeed
simultaneously for the weakest as well as the strongest stimuli.
If it is assumed that weak stimuli bring about weak excitations
in the nerve fiber, it must most certainly be expected that on the
cessation of the response, weak stimuli applied at the central
nerve end would still, by slight increase of the intensity of stimulation,
be followed anew by reaction in the muscle. This is all
the more to be expected, because the irritability of the narcotized
stretch, as shown by stimulation with the electrodes inside the
chamber, very gradually decreases, so that within the chamber
stimuli of moderate strength are still effective. Instead the capability
of conduction is completely obliterated, and even the strongest
stimuli, applied to the end of the nerve, produce no response
in the muscle. This in turn does not agree with the assumption
that the intensity of excitation varies with the strength of the
stimulus in the individual nerve fiber. The facts here alluded to
are, therefore, either not correct, or the intensity of excitation
in the individual nerve fibers is independent of the strength of the
stimulus, and the view which we have entertained up to the
present in this respect is incorrect.
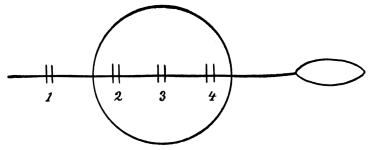
In order to examine these facts once more on an extensive
scale, and at the same time obtain an understanding of the development
of the decrement in the narcotized stretch, I have requested
Dr. Lodholz to register as many accurate curves as possible
in which the positions of the secondary coil of an inductorium
are the ordinates indicating the threshold of stimulation at four
points of a nerve stretch. Of these points three are situated at
prescribed distances from each other in the narcotized or asphyxiated
stretch; the fourth is centrally placed. (Figure 24.) As143
might be expected the result was the same as in former investigations.
They show however even more strikingly the abruptness
of the disappearance of conductivity simultaneously for the weakest
and the strongest stimuli. The curve produced by the centrally
placed electrode remains at the same height for a considerable
period, then suddenly abruptly declines. Those of the electrodes
within the chamber likewise sink, at first slowly, then with increasing
rapidity in successive order corresponding to the distance
which they are situated from the point of exit of the
nerve, so that the curve of the most distant electrode reaches the
abscissa first, that of the electrode nearest the muscle in the
chamber, last. The experiments demonstrate with complete
clearness that in contrast to all those points within the affected
stretch, where the conductivity, though already obliterated for
weaker stimuli, still exists for stronger, that with stimulation
of a point towards the center above the affected stretch, conduction
ceases simultaneously for all different strengths of
stimuli. This last state at the points within the affected stretch
might be ascribed to the diminution of the excitability of this
stretch, and the idea entertained that the weak stimuli no longer
produce excitation capable of further conduction.
This assumption is contradicted, however, by the fact that
subsequently to the disappearance of the response at a point situated
at the greatest distance from the place of exit, an effect of
stimulation can be obtained at the nearest point to the exit with
the same or even less strength of the current. As the irritability
in the affected stretch is reduced at all points in equal measure,
the fact of a weaker stimulus becoming inoperative whilst a
stronger remains effective can only be attributed to the circumstance
that the wave of excitation set free at some point of the
influenced stretch by a weaker stimulus is sooner obliterated on
its way to the muscle than that produced at the same point by a
stronger stimulus. These experiments, in which the manifestations
of the nerves in response to stimuli applied centrally above
the chamber in the normal stretch are compared to those in
response to a stimulus acting on the affected stretch, clearly
demonstrate the totally different effect in both cases. In stimulation144
of the centrally situated normal stretch, the wave of excitation,
which enters from here into the influenced stretch, is obliterated
at the same point simultaneously for the weakest as well as
for the strongest stimulus; stimulation of the affected stretch, the
wave of excitation which is set free at one point by a weak stimulus,
is obliterated sooner and after passing through a shorter
stretch than that which is produced by a stronger stimulus. It is
self-evident that in the first instance, in which the stimulus acts on
the centrally situated normal stretch, the wave of excitation,
thereby set free, must in passing through the affected stretch
undergo a decrement of its intensity. If, therefore, the wave
of excitation, coming from above, is obliterated exactly at the
same point, whether brought about by weak or strong stimuli, the
inevitable conclusion must be drawn that, whether either a weak
or a strong stimulus is operative, the wave of excitation must
have entered into the influenced stretch from the normal stretch
with exactly the same intensity. In other words: the weakest
as well as the strongest stimuli produce excitations of equal
intensity in the normal nerve, that is, the “all or none law” is
valid for the nerve.
This information can no longer be doubted in the presence of
such evidence as was presented above. This indeed is a fact of
far-reaching importance in the understanding of the functional
activity of our nervous system, for it is evident that the difference
of intensity in the conduction of excitation is not, as has
been assumed until now, the result of the conduction of varying
strengths of a single excitation in the same elementary
fibers, but rather the involvement of a various number of fibers,
and that a series of processes which we have to the present
attributed to the varying intensities are now to be explained
by difference in the duration and form of excitation. This gives
us an entirely different but nevertheless a more definite picture
of the physiological character of the processes in the nervous
system. Still, this question belongs to another chapter of physiology.
Here we are interested in the fact that we have in
the nerve a form of living substance, in which irritability has
reached a high degree, and every stimulus which is at all operative145
brings about disintegration of all the material involved
in excitation, and consequently the property of conductivity
in the nerve reaches the state of highest development of all
living structures, in that the medullated nerve conducts with
the greatest rapidity on the one hand, and on the other, there is
no decrement of the velocity and intensity of excitation. All
these characteristics: the existence of the “all or none law,” the
rapidity of the conduction of excitation, the absence of a decrement
in the velocity, the absence of a decrement of the intensity
of the excitation wave, the want of the capability of summation
of excitation, are all dependent upon one another, for they are
the combined expression of one and the same factor, that of the
high state of irritability. When the irritability is artificially reduced,
then the nerve approaches more and more, depending upon
the amount of reduction, to the series of living substances in
which we found the protoplasm of the rhizopoda to occupy the
other extreme. Between the normal medullary nerve with its
maximal, and the pseudopods of the rhizopods with their minimal
capability of reaction, we find innumerable gradations in groups
of living substances. Whether or not other forms of living substances
follow the type of the nerve, whether for example the
“all or none law” can be applied to the skeletal muscle as the
investigations of Keith Lucas112 seem to show, requires further
investigation.
Finally, there arises the important question as to the finer
mechanism of conductivity. The progression of excitation from
cross section to cross section in a living system is brought about
by the decomposition of the molecules in one region acting as a
stimulus and producing a disintegration of the molecules in
another region, etc. We have already seen that the intensity is
dependent upon the amount of energy produced by the disintegration
of the molecules following the stimulus, that is, upon the
amount liberated in a definite space in a definite time. The question
which now arises is this: What form of energy is produced146
by the stimulus at the point of stimulation, which acts upon the
neighboring molecules? The conduction of excitation is a property
of all living substance, and we may presume that this
occurs in all living systems in the same manner. If one examines
the forms of energy which are produced in a living substance by
the breaking down of the molecules, we find that chiefly three
forms of energy may be taken in consideration in the problem
of conductivity: heat, electricity and osmotic energy. Light
cannot be looked upon as a form of energy which is produced
by all living substance, and the other forms of energy, as the
chemical energy and surface tension, remain local. At a first
glance one is inclined to assume that heat is the form of energy
which is liberated by the breaking down of the stimulated molecule
and which spreads to the neighboring molecules and brings
about their decomposition. For we know that heat facilitates
dissociation, and the analogy between living substance and explosive
material is very close. In both instances the decomposition,
which extends over a great mass of molecules, is accomplished by
the heat produced in the breaking down of a few molecules. In
fact, the conduction of excitation of a nerve can in many respects
be compared with the burning of a fuse.113 Nevertheless, it must
not be forgotten that this analogy, which on first glance seems so
apt, upon closer observation presents serious difficulties. It can
be experimentally shown that an increase in the temperature in
the living substance follows stimulation, but it is also known that
in momentary excitation following a single stimulus, as in the
muscle after the application of an induction shock, the heat production
is extremely small. This difficulty becomes particularly
apparent if we endeavor to gain an approximate idea of the
numerical proportions of the irritable, that is the disintegrating
molecules to the remaining mass of a living system. The
water content above all represents an enormous proportion.
If we calculate this to be for the nerve, for instance, roughly
about 75 per cent., which is a low estimate, only 25 per cent.147
of dry substances remain. Even of this 25 per cent. by far
the largest part is apportioned to connective tissue, for which 15
per cent. is certainly not too high a figure. Neither can the remaining
10 per cent. of dry substances be regarded as consisting
entirely of molecules capable of decomposition. For in this is
also included the organic reserve material of the axis cylinder protoplasm,
which is doubtless of very considerable amount. Further,
the salts and products of disintegration, for which the estimate
for the sum total would probably not be too low if we assume the
amount to be equal to that of the group specially concerned in
the process of excitation. As, however, a constant metabolism
of rest takes place, these last molecules or atom groups are certainly
not at the moment of entrance of the stimulus in their
entirety in a condition capable of decomposition. It is quite certain,
therefore, that we are still overestimating the amount of the
molecules capable of disintegration, if we put them down as 5 per
cent. of the entire nerve substance. If we now suppose that this
5 per cent. of irritable molecules are broken down as a result of
stimulation, 95 per cent. of nonirritable substance, separating
these irritable molecules, must become heated to such a degree
by the disintegration of the latter that the amount of heat suffices
to bring about decomposition of the nearest surrounding molecules
or atom groups, for otherwise conduction of disintegration
could not take place in this manner. This condition presents a
serious difficulty for the assumption that heat is the form of
energy responsible for the conduction of disintegration. It is
true that we cannot reject this view at once as being completely
incorrect, as the possibility of conduction does not depend upon
the absolute amount of heat which reaches the next molecule
capable of decomposition, but upon the relative amount of heat
in regard to the degree of lability of the irritable molecules,
of which we cannot even approximately make an estimate. However,
by a comparison with other highly explosive substances,
such as iodide of nitrogen, we find that a slight trace of water
applied to the iodide of nitrogen suffices to prevent the extension
of the disintegration process, and with this the explosion of the
whole mass. Nor does the view of Pflüger remove this difficulty,148
which assumes that the atom groups capable of breaking down
are joined together by a chemical linking of atoms to long fiber-shaped
giant molecules through the whole nerve fiber, for this
assumption of a firm structure can hardly be reconciled with
the principles concerned of metabolism.
In consideration of this difficulty it seems easier to assign
the rôle of mediator of disintegration not to heat but to electricity.
Production of electricity is likewise a property of all
living substance. Differences of electrical potential between two
points may be equalized in the stretch by conduction through
the intervening space. Electricity would then fulfil the important
conditions, which must be demanded for the form of energy,
acting as mediator for the conduction of disintegration from
cross section to cross section.
Physiologists even at an early date, misled by the apparent likeness
in the conduction of excitation, especially in the nerve, to
that of electricity in a metal wire, regarded both processes as
identical. When, however, Helmholtz first demonstrated experimentally
the rapidity of the conduction in the nerve, the thought
that electrical conduction was concerned, such as takes place in
a metal wire, had to be abandoned, as the velocity shows too
great a difference in the two cases.
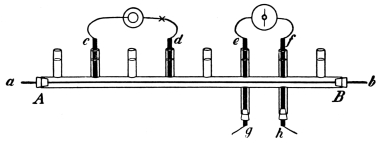
Fig. 25.
Model of a “Kernleiter.” A, B—Glass tube, with a number of side tubes
filled with saline solution, through which a wire is passed. c and d—Side
tubes with electrodes for stimulation. e and f—Tubes for connection
with a galvanometer. (After Hermann.)
The observations, on the other hand, on the conductivity in the
so-called “core model,” seemed to offer another possibility of
attributing the conduction of excitation in the nerve to electric149
processes. Matteucci, later Hermann and finally Boruttau114 have
endeavored to apply the results obtained when electricity is introduced
in a wire covered with a moist envelope (saline solution),
to the explanation of conductivity in the nerve. (Figure 25.)
The fact has been shown, that in such a model the application of
electricity to a point, as a result of polarization between the moist
envelope and the metal, produces a weak local current, which in
turn disturbs the electrical potential in the next cross section and
consequently a new local current is produced and so on through
the whole length of the wire. (Figure 26.) This fact, in connection
with the apparent similarity in the differentiation of the
axial fibers and peripheral envelope in the nerve, has led Boruttau
to apply the principles of conductivity in the “core model”
to that of the nerve. Then, however, Nernst and Zeyneck brought
forward their theory, according to which the galvanic current is
operative as a stimulus in that it brings about an alteration in
the concentration of the ions at the junction of two different
electrolites which, in turn, produce local currents. Boruttau then
dropped the assumption of the existence of a simple physical
polarization between the wire and the envelope and replaced it
by the assumption of an alteration in the concentration of the
ions at this position. Thereby the “core model explanation” was
already altered in principle, in that only the differentiation of a
central fibrilla and a peripheral enveloping substance was appropriated.
It seems to me that this factor can likewise be considered150
as completely dispensable and may, therefore, be omitted;
thus nothing remains of the “core model explanation” of the
conduction of excitation in the nerve.
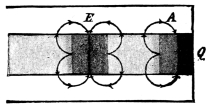
Fig. 26.
Scheme of the conduction by local electric currents
in a “Kernleiter.” (After Hermann.)
The results of continually increasing numbers of investigation
in recent times make it appear almost as a certainty that the elementary
fibrillæ in the axis cylinder are nothing else but skeletal
substances. Wolff,115 Verworn116 and others have first expressed
the view that the neurofibrillæ must be looked upon as skeletal
fibers for the soft neuroplasm, and more recently Lenhossek117 and
especially Goldschmidt118 have confirmed this assumption in detail.
Goldschmidt has shown by extensive comparative studies of cell
mechanism the rôle played by the neurofibrillæ in a physical connection
as internal skeletal formations, and has proved at the
same time, in complete unanimity with other investigators, their
continuity with other undoubted skeletal fibrillæ. By this the
numerous combinations and speculations of Apathy and Bethe
concerning the part taken by the neurofibrillæ have been rendered
untenable. In no case is there the slightest justification to regard
the apparent “Kernleiterstructur” of the nerve as the principal
condition for the process of conductivity, for should we dispense
completely with this point for the theory of the conduction of
the nerve, we can obtain, solely by the aid of the facts known
today in physical chemistry, the foundations for a theory of the
conductions of excitation which not merely renders the specific
case of the conduction of the nerve intelligible, but contains at the
same time the principles of the process of the conduction of excitation
for all living substance.
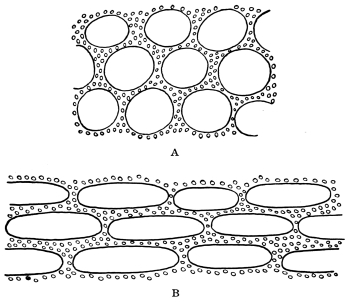
Fig. 27.
Scheme of the foam structure of living substance. A—In
undifferentiated protoplasm. B—In fibrillae protoplasm.
On the basis of investigation in the physical chemistry on the
properties of semi-permeable membranes, we know that such
membranes produce an elective effect on the diffusion of dissolved151
substances. This is in the way that the two different solutions,
separated by a semi-permeable surface, do not follow the known
laws of diffusion, but are altered in that certain substances in
contrast to their rapidity of diffusion pass through the membrane
or are prevented from entering by the latter. This applies likewise
to the two kinds of ions, which are dissociated in diluted
substances. If the surface exercises a selection in the way,
for instance, that the positive kations are allowed to pass through,
whilst the negative anions are held back, a difference of potential
must exist between the two. In this manner, wherever two different
solutions are separated from each other by a semi-permeable
surface, an opportunity occurs for the taking place of galvanic
currents. As we know, living protoplasm by reason of its colloidal
components possesses, in common with all colloidal substances,
on its surface the properties of semi-permeable membranes.
Between the cell and the medium, therefore, there is
always the opportunity for the occurrence of differences of electric
potential. But more. We likewise know that protoplasm itself153
represents a mixture of colloid substances and actual solutions.
Frequently, if not always, living structure presents a morphological
differentiation of two types, when seen under the microscope,
in the form of a foam structure described by Bütschli.
(Figures 27 and 28.) If we suppose that with the disintegration
of complex molecules, which we must assume as taking place
in the material of the walls of the protoplasm network, substances
are formed which are subjected to electrolytic dissociation,
the anions and kations hereby liberated must be diffused from
the place of their separation into the surroundings. Their diffusion,
however, is restricted by the protoplasmic network. The
positive ions may pass through, but the negative ions may not.
As a result: the reticulated substance is the seat of electric discharge,
which in turn gives the impact to the breaking down of
new molecules and with this to the occurrence of new potential
differences, and so on, consequently the disintegration is extended
further and further through the connected masses of the protoplasmic
framework.
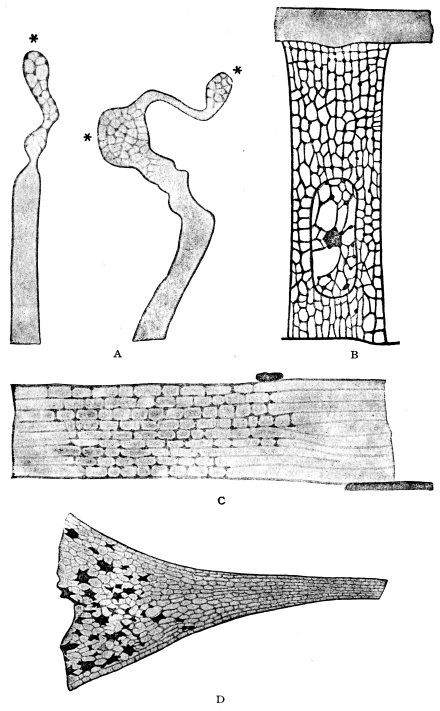
Fig. 28.
Protoplasm of different cells, showing foam structures. A—Pseudopod of a marine
rhizopod. The protoplasm only shows foam structure at the point of stimulation.
B—Epidermic cell of lumbricus. C—Nerve fiber. D—Part of the cell
body of a ganglia cell. (A-C after Bütschli, D after Held.)
This theory, founded on facts gained entirely from investigation,
would involve those forms of energy which play the rôle of
activator in the extension of the breaking down of the molecule
from cross section to cross section, namely, the osmotic and the
electrical energy. Based on the general properties of physical
chemistry and those of morphology of the living substances, they
would be applicable to all vital systems. It would be premature
to attempt to extend this assumption and further develop its
specific details, above all to make it responsible for the specific
differences in the process of the conduction of excitation in
various forms of living substance. For this our knowledge of
the properties of living substance is still far too incomplete.
Nevertheless, it furnishes us even now with various points of
view for the further analysis of a series of vital manifestations,
as, for instance, the facts concerning the production of electricity,
of galvanotaxis, chemotaxis and so on. This, however, exceeds
the limits of the task we have here mapped out. We are concerned
here solely with the general principle on which the conductivity
of excitation in the living substance is founded.
154
CHAPTER VII
THE REFRACTORY PERIOD AND FATIGUE
Contents: Conception of specific irritability. Alteration of specific irritability
during and after excitation. Refractory period in various
forms of living substance. Absolute and relative refractory period.
Curve of irritability during refractory period. Dependence of the
duration of the refractory period on the rapidity of the course of the
metabolic processes in the living substance. Dependence on temperature.
Dependence on supply of oxygen. Theory of refractory period.
Refractory period as basis of fatigue. Fatigue as a form of asphyxiation.
Alterations of irritability and the course of excitation in fatigue.
Recovery from fatigue. The rôle played by oxygen in recovery.
Fatigue as an expression of the prolongation of the refractory period
conditioned by the relative want of oxygen. Fatigue of the nerve.
Every living system possesses, as we know, a peculiar and characteristic
manner of reacting to stimulation. The muscle responds
with a contraction, the salivary cell with production of saliva, the
luminous cell with the emission of light. This is the specific
energy in the sense of Johannes Müller. Every living system is
likewise characterized by a certain degree of irritability, which
can be expressed by the threshold value of the stimulus at which
the specific reaction is just perceptible. This degree of irritability,
by which the system concerned is distinguished, may be
termed its specific irritability.
From the standpoint of the conditional method of investigation
it is at once apparent that specific energy, as well as specific irritability,
must be solely determined by the specific conditions existing
in the particular system. It follows from this that every alteration
in the conditions of the system, that is, every change of its
state, likewise entails a corresponding alteration of its specific
energy and its specific irritability. It is, therefore, self-evident
that the alteration of the state, which is undergone by the living155
system in the process of excitation, brings about an alteration of
its specific irritability. Likewise as the original state of the system
is restored by the metabolic self-regulation after the course of an
excitation, the specific irritability of the system must be reestablished.
The specific irritability is, therefore, a property of
the living system, which, like the metabolic equilibrium, undergoes
restitution by the process of self-regulation after variation
produced by a stimulus of any kind. It is scarcely necessary to
repeat each time that this is only applicable within the physiological
variations and for a limited period, during which the alterations
in development need not be considered.
These alterations of the specific irritability following an excitation
and their compensation through the metabolic self-regulation
will now claim our attention.
That the specific irritability of a living system undergoes a
diminution as the result of a stimulus of long duration has been
long known through the study of fatigue. This is especially so
with frequently recurring excitating stimuli. It is only within the
last decade, however, that the observation has been made in a few
instances that a single momentary excitation is likewise followed
by such a reduction of the specific irritability. But that this is a
fact of general physiological fundamental importance for the
whole field of response to stimulation in the living substance has
only been recognized within the last few years.
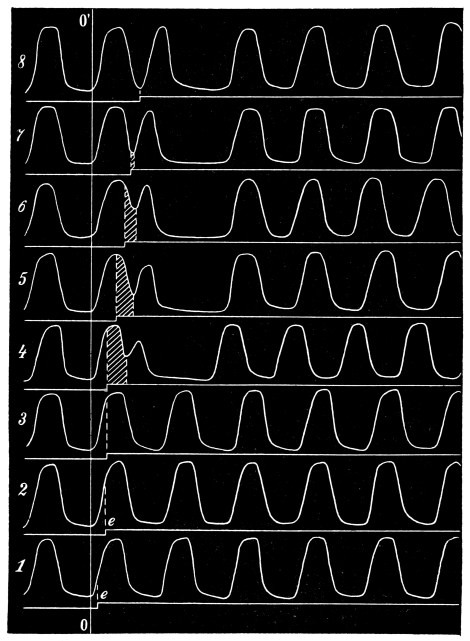
Fig. 29.
Eight series of heart contractions. The dotted lines e show the moment of an artificial
stimulus. The artificial stimulus is ineffective if it is applied before the height of a
systole. The artificial stimulus becomes the more effective in producing an extra
systole, followed by a compensatory pause, the later it is applied after the height of
the systolic contraction. (After Marey.)
In 1876 Marey119 found that the irritability of the heart in response
to artificial stimulation was greatly reduced during the
systole, and that recovery took place during the following diastole.
(Figure 29.) This fact was already apparent from the
observations made by Bowditch120 and Kronecker,121 that by stimulation
of the isolated frog’s heart with single induction shocks, an157
artificial systole can only be produced with certainty when the
stimuli succeed each other at certain intervals, which must be the
longer as the strength of the stimulation is weaker. Marey calls
this period of reduced irritability “phase réfractaire” of the heart.
The refractory period of the heart has been made the subject of
a great number of investigations, especially by Engelmann and
his pupils. It was Engelmann122 especially who determined more
exactly the duration of the course of the refractory period. He
found, namely, that irritability disappears immediately before
each systole and reappears shortly before the beginning of the
diastole, and again reaches its original height at the end of the
diastole. For a long time, however, this refractory period was
looked upon as a special peculiarity of the heart. It was not until
Broca and Richet,123 twenty years after Marey’s investigations, discovered
an analogous refractory period for the motor centers of
the cerebral cortex of the dog. They first made this observation
on a dog affected with chorea, in which the choreic movements
rhythmically occurred in intervals of one second. They found that
after each movement electrical stimulation of the cortex remained
without result for about .5 seconds. During the next .25 seconds
stimulation was followed by a weak response and it was not until
the last .25 seconds before the next movement that a strong
effect was produced. They also found in the normal dog a refractory
period after every artificial stimulation equal to .1
second, so that the number of contractions brought about by
rhythmical electrical stimulation were only ten per second. Following
this, numerous other investigations of the refractory period
have been made on the central nervous system. Zwaardemaker124
and Lans have observed a refractory period in the eyelid reflex
of the human being which, on stimulation of the optic nerve,158
amounts to about .5–1 second; on the stimulation of the trigeminus
produced by blowing on the cornea on the other hand,
it is somewhat shorter, less than .25 seconds. Zwaardemaker125
also was able to demonstrate an analogous refractory period for
the swallowing reflex of the cat. Further a refractory period
was found and closely analyzed by Verworn126 for the reflexes in
the spinal cord of the strychninized frog. Dodge127 found a refractory
period in the knee jerk reflex of man. Gotch and Burch128
showed, by two induction shocks following each other in quick
succession, a refractory period of the nerve, which is characterized
by its extremely brief duration. They found, depending
upon the temperature, a period of nonirritability of .001–.008
seconds after every stimulus. The investigations of Miss
Buchanan129 lead us to conclude that there is a refractory period
for the cross striated skeletal muscle. Miss Buchanan stimulated
the muscle at times through the nerve, at other times directly
after elimination of the nervous element, with very frequent
electrical stimuli (about 1000 in the second) and found by means
of the capillary electrometer a rhythmical reaction of the muscle
of about 50–100 excitation shocks per second. Likewise the
Ritter tetanus produced by the breaking of an increasing current
proved to be a rhythmical reaction of an analogous nature. In
a more direct manner Keith Lucas130 has determined the refractory
stage for the musculus sartorius of the frog. He allowed two
induction shocks to act successively on the muscle at intervals159
of varied duration and then registered the action currents by
means of the capillary electrometer. He then found that the
second stimulus was ineffective for about .005 seconds after the
application of the first stimulus. If the second stimulus follows
somewhat later, it produces a contraction which is weaker and
has a longer latent period the nearer the second stimulus approaches
the first in point of time. (Figure 30.) Massart131 and
Jennings132 likewise observed the existence of a refractory period
for the myoids of unicellular organisms brought about by mechanical
stimuli. Massart attributes this cessation of reaction to
stimuli following each other at certain intervals, to fatigue, an
explanation which has been disputed by Jennings as the result of
his investigations made on Stentor and Vorticella. Jennings looks
upon the behavior of the infusoria rather as an “adaptation” to
the stimulus. Pütter was the first to see in this the existence of
a refractory period. His experiments on Spirostomun ambiguum
in 1900 showed a refractory period in the reaction to rhythmical
mechanical stimuli. I wish to state, however, that these observations
of Pütter have not as yet been published. Thus the existence
of a refractory period has even today been proved for a
whole series of very different kinds of substances.
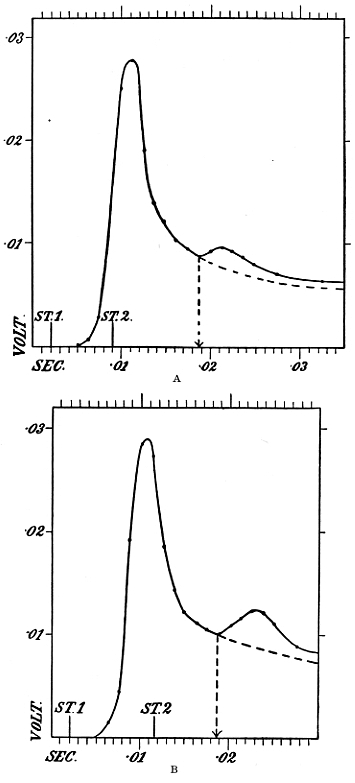
Fig. 30.
Curve of action current of the musculus sartorius excitated by two successive
stimuli (St. 1 and St. 2). The effect of the second stimulus
is the less and the latent period is the longer the more quickly the
first stimulus is followed by the second. (Keith Lucas.)
We will now examine the alterations of irritability which are
perceptible during the refractory period to complete restitution of
the specific irritability of the particular system, and endeavor by
the analysis of their special conditions to render them comprehensible
from a physical standpoint of view.
The first fact to take into consideration is, that, as is shown in
the heart, the refractory period begins at the moment of the
appearance of the systolic excitation. The irritability of the
heart is absent and remains so until the excitation has reached
its highest point, that is, shortly before the beginning of the diastole.
From this point the restitution of irritability begins, which
does not reach the maximum until the end of the diastole. In
other words: irritability undergoes the greatest reduction by disintegration162161
produced by the stimulus and is restored by the metabolic
self-regulation following the decomposition.
This point of view enables us to interpret this state from a
physical standpoint. In this discussion on the relations between
irritability and the extension of excitation, I have taken the
amount of energy which is produced during the time unit and
space unit in a living system as the general standard for the
degree of irritability, at the same time duly regarding the individual
components involved. This amount of energy is determined
in a given system by the quantity of substance broken down
by a stimulus of a given intensity. It is, therefore, clear that
during the time in which an increased disintegration produced
by a stimulus takes place, the irritability in response to a second
stimulus must be reduced, as during this period the second stimulus
has less of necessary decomposable substances at its disposal,
and at the same time there are more products of disintegration
in a given space. If a living organism is the subject of consideration,
to which the “all or none law” is applicable, as, for instance,
the heart at the moment of the beginning of excitation, irritability
is completely obliterated, as shown by the fact that the
second stimulus of any strength remains without response, for
during the excitation there is a complete breaking down of all
the substances capable of decomposition. If, on the contrary, a
system is the subject of observation, for which the “all or none
law” is not valid, then irritability is merely reduced but not wholly
obliterated during an excitation, and whether or not a response
is obtained to the stimulus depends upon its strength. To impress
the relations between the degree of irritability and the intensity
of the stimulus, I have, therefore, employed the term “relative
refractory period” in contrast to the “absolute refractory period,”
in which irritability is obliterated even for the strongest stimuli.
It is self-evident that irritability must again increase in the same
degree as the restitution of the living system by metabolic self-regulation
takes place, for the more molecules capable of disintegrating
are restored and the more products of disintegration
removed, the more molecules necessary for decomposition in the
unit of space are attacked and broken down by the stimulus. All163
these are self-evident facts which are in accordance with the conception
we have here developed of the course of the process of
excitation and its physical nature. But another important point
is evolved from the observations we have made of the nature of
the process of self-regulation. The process of self-regulation is
founded on the same principle as that which governs the taking
place of all chemical equilibrium, for metabolic equilibrium is
merely a special kind of a chemical equilibrium. The development
of a chemical equilibrium between reacting substances and
reaction products has, as known, a characteristic course in regard
to its duration. If the rapidity with which the equilibrium is
reached is expressed by a curve in which the abscissa represents
the time, while the ordinates signify the number of contacts of
the interacting molecules, the rapidity of reaction is altered with
the approach to the equilibrium in the form of a logarithmic
curve; that is, the approach to the state of equilibrium, which is
represented by ordinate value zero, takes place at first very rapidly,
then with more and more decreasing speed, for with the
decrease of the number of reacting molecules and the increase
of the amount of products of reaction, the contact of the interacting
molecules and with this the opportunity for the reaction
occurs less and less frequently. Although the self-regulation
of metabolic equilibrium is by no means such a simple process
as, for instance, that of the well-known example of the
forming of ethylester from acetic acid and æthyl alcohol, we have
still in every case to deal with the taking place of a chemical
mass equilibrium. Hence the progress to the metabolic equilibrium
must likewise correspond with a logarithmic curve, i.e., restitution
after a disturbance of the equilibrium must take place at
first rapidly, then at a constantly decreasing rate. For reasons
readily to be understood the special form of this restitution curve
has so far not been accurately ascertained for any kind of living
substance. Even in those cases where the restitution occurs
very slowly we meet with the difficulty that, when the tests are
applied which are necessary to determine the restitution at different
intervals, with each testing stimulus irritability is each
time reduced. Hence the construction of the restitution curve164
can only be achieved by indirect means, and we must content ourselves
with the ascertainment of a smaller number of its points
from which by interpolation its form can be constructed. Indeed
in this connection a certain number of results have already been
gained quite sufficient to experimentally confirm the correctness
of these types of curves, primarily obtained by purely theoretical
deductions. That irritability very gradually reaches its maximal
height has been already shown, as previously mentioned by
Bowditch133 in his investigations on the influence of rhythmical
induction shocks on the apex of the heart of the frog. He found
that in order to produce response, the weaker the stimuli the
longer must be the intervals between them. It follows from this,
that after a discharge the irritability in response to strong stimuli
reappears more rapidly than for weak, i.e., that they only gradually
regain their maximum. The exact periods of time for the
course of the return of irritability for the heart have unfortunately
not been so far ascertained. On the other hand, the investigations
of Ishikawa134 furnish the material for the construction of
the restitution curve for the centers of the spinal cord of the frog.
Ishikawa did not employ the threshold of stimulation as an indicator
for the course of restitution, but used instead the duration
of the reflex time following on a stimulus of a certain strength.
The reflex time is greatly prolonged after an excitation of extended
duration and only regains its normal value in the same
degree as restitution takes place. By a great number of painstaking
experiments Ishikawa ascertained the duration of the
reflex time at intervals of thirty seconds to one minute, and obtained
figures which show that restitution does actually take
place, at first rapidly and then with constantly decreasing
speed. The detailed study of the course of self-regulation of the
individual forms of living substance will doubtless be more exactly
determined in the near future. But even at the present
we are fully justified in describing the form of restitution
curve as a logarithmic in type. Therefore, a relative refractory165
period must be present in every metabolic self-regulation after
an excitation, during which stronger stimuli produce response,
while weaker are still without result. This is a fact which, as we
shall see later, is of fundamental importance for the comprehension
of the various kinds of interference responses to stimuli.
From the information here gained on the nature and origin
of the refractory period the conclusion must inevitably be drawn
that in all living substance there must exist, directly following an
excitation, a period of time in which its irritability is reduced,
that is, under proper conditions a refractory period can be
demonstrated for every living organism. Every living system
possessing irritability undergoes a period of reduced irritability
at the time of and subsequent to every excitation, for every
excitation momentarily decreases the amount of products capable
of disintegration and increases the disintegration products in the
unit of space. As restitution involves time, a stimulus occurring
in the phase preceding complete restitution cannot break down
the same quantity of molecules as would be the case after the
establishment of complete restitution, that is, the response is
weaker, the irritability is decreased. The refractory period during
and subsequent to excitation is as much a general property of the
living substance as irritability and metabolic self-regulation.
This conclusion appears so self-evident that it would seem
hardly to call for emphasis were it not that even at the present
time the view is still widely held that the refractory period is a
special characteristic of certain forms of living substance. This
assumption is explained on the one hand by the fact that our
information concerning the refractory period is still of comparatively
recent date and that few physiologists are in the habit of
connecting special observations with general physiological conceptions,
but also for the reason that some investigators have
vainly tried to find a refractory period in certain forms of living
substance. Langendorff and Winterstein,135 for instance, have not
succeeded in proving a refractory period for the spinal cord of
the frog. Langendorff stimulated the central sciatic stump166
with two stimuli in quick succession and used the contractions
of the triceps as indicator of the response. He found that when
the stimuli, if consisting in either single induction shocks or
faradic shocks, followed each other even at intervals of .004
seconds the second stimulus was still operative, this being perceptible
in an increase of the contraction or with greater intervals
of time in a summation of two contractions. Winterstein
concludes from this that the development of a refractory period
after a stimulation is not a general property of all nerve centers.
If the experiments of Langendorff failed to show the presence
of a refractory period it is not for the reason that this does not
take place in the centers of the spinal cord but rather results from
the fact that the conditions for the investigation were not suited
for its demonstration. In fact, Fröhlich136 and especially Vészi137
have incontestably proved the existence of relative refractory
periods in the normal spinal cord.
If the existence of the refractory period is based on the fact
that during the time of and subsequent to an excitation the quantity
of substances necessary for disintegration is decreased and
that of the breaking down products increased, and if it is limited
by the restitution of the substances required for decomposition
and the elimination of the disintegration products, its duration
must be dependent upon the length of these processes. All factors
which lessen the decomposition and hasten the metabolic
self-regulation must, therefore, shorten its duration. This is
completely confirmed by experimental investigations. As can be
understood, the factors of special interest for us are those which
influence the duration of the refractory period in the physiological
occurrences of the organism.
One of these factors is temperature. As we know, the rapidity
of chemical reactions increases with ascending and decreases with
falling temperature. As in the disintegration as well as in the
restitution, processes are chemical in nature, it is to be expected167
that the duration of the refractory period is influenced in like
manner by temperature. Indeed, Kronecker138 found some time ago
that in the isolated frog’s heart a much more frequent rhythm of
stimulation is effective at a higher than at a lower temperature.
When the heart is stimulated at a temperature of 11–12° C. with
twelve rhythmical induction shocks in the second, every stimulus
is operative and produces a systole. If a stimulus of the same
frequency is used at a temperature of 5° C., the heart responds
merely to every second stimulus. This shows that the refractory
period is of longer duration at a lower than at a higher temperature.
A factor of particular interest is the supply of oxygen, for we
know its fundamental importance in all aërobic organisms in the
breaking down of the living substance. The life of these organisms
is primarily dependent upon the supply of oxygen from
without. Organic reserve substances for restitution after disintegration
are contained in ample quantity in the reserve stores in
the living cell substance, whereas oxygen is present in very small
quantities in relation to the former. It is, therefore, self-evident
that the rapidity of the breaking down processes is very closely
dependent upon the amount of available oxygen at hand. Nevertheless
it is not the absolute quantity but the relative amount of
oxygen in relation to the momentary requirement which is of
importance. For instance, the quantity of oxygen present may
completely suffice for the oxydative disintegration in the metabolism
of rest or at lower temperature, whereas the same amount
would be much too small to meet the demand increased by excitation
or at higher temperature. In the latter case “a relative
deficiency of oxygen” occurs. I have introduced the term “relative
deficiency of oxygen”139 for I have found that a number
of authors by neglecting the relations of the available oxygen
to that which is required at the moment have been led to false
conclusions. There is no living object so preëminently fitted
to demonstrate in such a striking manner the dependence of169
the duration of the refractory period upon the supply of oxygen
as the spinal cord centers of the frog, when their irritability
has been increased to the maximum by strychnine.140
Various observers, such as Loven, Buchanan, H. von Baeyer and
others, investigated the action current by the capillary electrometer.
As a means of studying the number of impulses in the
strychnine tetanus, we can upon the basis of their figures roughly
assume the number of impulses to equal ten per second at room
temperature. In short, in the freshly strychninized frog the duration
of the refractory period is about .1 second. By means of the
method of artificial circulation already mentioned a deficiency of
oxygen can readily be brought about. It has been demonstrated
that the rhythmic in contrast to the continuous method of introduction
of circulatory fluid is superior in that the former reproduces
more closely the natural conditions of the circulation of
the blood and renders the smallest capillaries more permeable.
In consequence I have recently constructed a small appliance for
artificial circulation, which accomplishes this in a manner as
simple as it is complete. (Figure 31.)
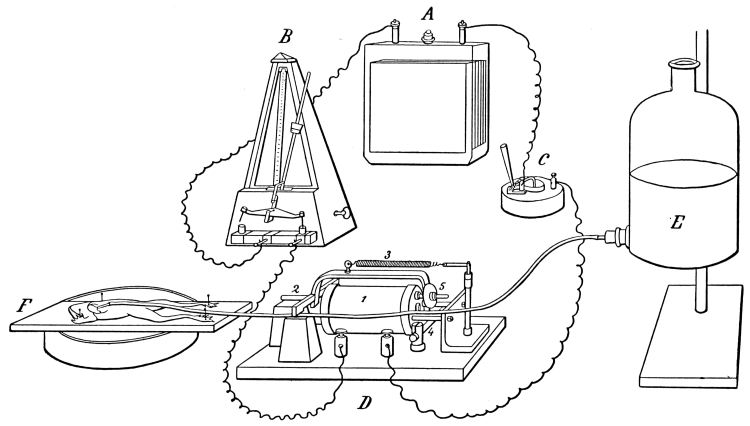
Fig. 31.
Arrangement for an artificial circulation in
the frog. A—Accumulator. B—Metronom. C—Mercury key.
D—Electromagnetic apparatus for compressing the rubber tube: 1,
wire spool with magnet; 2, anchor for the magnet; 3, spiral spring
which pulls back the anchor; 4, axis on which the anchor turns; 5,
plate for arresting the anchor. E—Vessel containing saline
solution. F—Slab of cork with frog.
The fluid flows from a vessel, E, provided with an outlet tube through a thin rubber tube
into a glass canula, which is introduced into the general aorta of
the frog, F. The tube is automatically occluded by the rhythmical
movement of the armative of an electromagnet, D, produced by a
metronome, B. The pressure of the circulating fluid can be readily
changed at will by varying the level of the vessel and the frequency
of the pulse by the rhythm of the metronome, which makes
and breaks the current to the electromagnet.141 In this way it is
possible to artificially replace the normal circulation with satisfactory
exactitude and substitute for the blood, circulating in the
vessels of the frog, any desired fluid. If the entire quantity of
blood of a frog is displaced by a continuous stream of oxygen-free
saline solution and a weak strychnine solution is injected
with a Pravaz syringe, a violent strychnine tetanus appears after170
the lapse of a few seconds. (Figure 32, A.) If the artificial circulation
with oxygen-free saline solution is now contained in the
rhythm of the natural heart beat, the further reactions can then
be readily observed. The first long-continued tetanic attack,
which can be produced by a slight touch of the skin, is followed
by a whole series of tetanic convulsions of prolonged duration,
which are repeatedly followed by periods of exhaustion. I wish
to emphasize this fact once more, as it appears to me as not without
interest for the understanding of the question of reserve substances.171
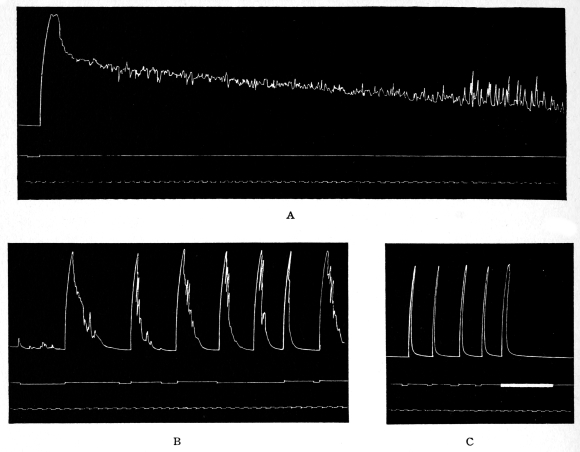
Fig. 32.
Muscle curve of strychnine tetanus in a frog with artificial oxygen-free circulation. Lower line indicates
seconds. Upper line indicates stimulation by induction shocks. A—A single shock produces a long
tetanic contraction. B—In a more advanced stage each shock produces a tetanus only of short
duration. C—In a still more advanced stage each shock brings about only a single contraction if the
stimuli do not succeed each other too rapidly. If they succeed more rapidly, as, for instance, in a
faradic current, only the first shock is effective.
If we assume that at the moment when the entire amount
of blood is removed from the vascular system, no oxygen remains
in the cells of the spinal cord and muscle, then disintegration of
the living substance could from this instant take place exclusively
anoxydatively, and there would be no further oxydative
breaking down into carbon dioxide and water. The energy production
compared in equal number of molecules, taking the
figures of Lesser for the fermentation of sugar, would approximately
amount to about 3.8 per cent. of that of the energy production
in the oxydative disintegration of dextrose into carbon
dioxide and water. In reality, however, the tetanic convulsions
are at first exactly as violent as in the frog with a normal circulation.
There simply remains the assumption, therefore, that
either the disintegration as soon as it becomes anoxydative involves
relatively greater number of molecules than would be the
case if it were oxydative in nature, or to suppose that even
after the complete displacement of the blood a certain, though
relatively small, amount of oxygen is present in the cells which
for a short time suffices for the taking place of oxydative disintegration
and with this an almost maximal production of energy
which naturally decreases as the oxygen is consumed. It seems
to me that the latter supposition contains more probability than
the first. To return, however, from this observation to a further
consideration of the animal we are studying, we see how the
complete tetanic convulsions in the refractory period which we
assumed to be .1 second are gradually transformed into incomplete
tetanus. After a time the tetanic convulsions become shorter
after each stimulus (Figure 32, B) and permit us to distinguish
their individual movements, even though the latter at first succeed
each other still very rapidly. Gradually this incomplete tetanic
convulsion assumes the form of a short series of individual contractions,
distinctly separated from each other and soon a stage is
reached in which each reaction to a peripheral stimulus consists
merely in a single contraction. (Figure 32, C.) The refractory
period is, however, even now less than a second. Nevertheless,
with a further continuation of the experiment, the refractory
period becomes more and more prolonged, so that stimuli succeeding172
each other at intervals of less than a second are without effect.
It is possible at this stage, as Tiedemann142 did, to graphically record
the reactions. He severed the sciatic nerve on one side and
stimulated its central stump, at the same time connecting the
triceps with a writing lever. It is then found that when the
single induction shocks follow each other at intervals of a second
or more every stimulus produces a contraction, but that on the
contrary only the first stimulus of a rhythmical series is operative
and all those succeeding ineffectual, if the stimuli follow each
other at shorter intervals. The refractory period becomes, however,
more and more prolonged. The rhythm of the stimulus
must become continually slower if each individual stimulus is
to remain effective. If the rhythm is even slightly too rapid
only the first few stimuli of a rhythmical series are effective
and this with decreasing response and later no contraction at all
is observed. With a further continuance of the experiment, the
stimuli are only effective when following each other at long
intervals. It is necessary that a period of recovery lasting
several seconds must take place before the following stimulus
can meet with response. (Figure 33.) The refractory period
can gradually be prolonged for the space of a minute or longer,173
until finally irritability does not reappear at all, and even the
strongest stimuli fail to produce the least contraction. The
continuous manner in which the refractory period is, in the
absence of oxygen, more and more prolonged until eventually a
prolonged state of nonirritability is developed, can be better
followed by observing the experiment than when described in
words. If at this stage instead of the oxygen-free saline solution,
defibered blood of the ox shaken in air or a saline solution saturated
with oxygen is circulated in the frog, restitution is often
within a few minutes so complete that tetanic attacks are once
more produced by a single stimulus, that is, the refractory period
has from being practically nil returned to the normal. This
experiment can be repeated several times on the same animal.
It is invariably found that the refractory period is prolonged
by the withdrawal of oxygen and shortened with a renewed
supply.
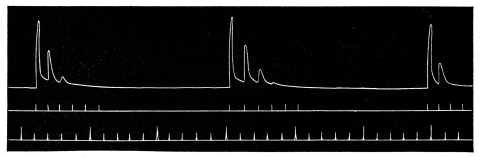
Fig. 33.
Development of the refractory period in the spinal cord of a strychninized frog. Lower
line indicates seconds; upper line stimuli. Of a series of stimuli only the first ones
are operative with decreasing effect.
I have described this experiment somewhat in detail as it contains
facts which are the key for the comprehension of a general
physiological process of paramount importance. I refer to
fatigue. The refractory period and fatigue are inseparably connected,
for fatigue is founded on the existence of the refractory
period and is an expression of prolongation of the former, brought
about by want of oxygen. This is shown at once by closer
analysis. It is here necessary to differentiate somewhat more in
detail the factors which bring about the prolongation of the
refractory period in deficiency of oxygen.
If we first turn our attention to the normal refractory period
which occurs in a system in metabolic equilibrium of rest in
direct connection with dissimilatory excitation, following a
momentary stimulus, we find that reduction of irritability or,
more exactly expressed, the lessening of the response is, as we
have seen, determined by the time involved in the metabolic
decomposition and recovery. Both these processes require time
and until their completion the quantity of substance demanded
for the oxydative disintegration is decreased in a given space,
and every stimulus must consequently be followed by a weaker
response. Our conceptions of the physical details of these processes174
depend essentially upon the question, if the oxydative disintegration
itself in the given living system occurs in one single
phase, in that the oxygen is the activator for the oxydative splitting
up of the carbon chain, or if this takes place in two periods,
in which the carbon chain is first anoxydatively split up into
larger fragments by the stimulus, which are then seized upon by
the oxygen to be split up into carbon dioxide and water. As we
have seen, this question must remain for the present undecided
as far as the metabolism of rest as well as the excitation produced
by a single momentary stimulus is concerned. It is highly
probable that a uniformity of the process for all living systems
does not exist. We are, therefore, not justified in assuming that
these special chemical processes resulting from single stimuli are
uniform throughout the refractory period.
On the contrary it is different in the case of oxygen deficiency.
Here we see with increasing want of oxygen a constantly increasing
duration of the refractory period, a prolongation which may
be attributed to the retardation of the oxydative disintegration.
It is necessary, however, that we now study more clearly these
alterations brought about by the deficiency of oxygen.
If we follow the course of the changes from that of the normal
state of equilibrium of metabolism, wherein oxygen is sufficient
to bring about complete disintegration of the molecules to the
formation of carbon dioxide and water, we must assume in spite
of the great explosive rapidity of this process on the basis of our
chemical knowledge, that first a series of intermediate products
are produced before finally the end products are formed. In
this way the oxydative disintegration produced by a stimulus
becomes more and more prolonged by an increasing want of
oxygen. If, as I have previously suggested, the amount of energy
which is liberated in a given space and time by an excitating stimulus
is taken as a standard of irritability, it is apparent that the
more the oxydative disintegration following a stimulus is retarded,
the greater must be the decrease in irritability. The less oxygen
there is at disposal and the more incomplete the oxydative breaking
down, the smaller is the degree of irritability, the weaker the
response and the slower the return of irritability after every175
stimulus. In other words, with the increasing deficiency of
oxygen, the response is not merely reduced for every stimulus,
but the duration of the refractory period is likewise progressively
prolonged until finally with an absolute want of oxygen, constant
and complete depression takes place. In the genesis of this process
another factor, however, has the same effect.
While with a sufficient supply of oxygen disintegration leads
to the formation of carbon dioxide and water, therefore to end
products, which can quickly and easily be removed by diffusion,
the want of oxygen produces complex products of incomplete
combustion and finally of anoxydative decomposition, such as
lactic acid, fatty acids and even more complex substances in constantly
increasing quantities. These products permeate the protoplasmic
surfaces with great difficulty, if at all, and as they cannot
subsequently be oxydatively split up, constantly accumulate.
These asphyxiation substances, as they may be briefly termed,
produce a depressing effect on further disintegration. This can
be experimentally demonstrated.
For this purpose I have modified the experiment previously
described in the way that after every introduction into the blood
of oxygen-free saline solution and after the injection of strychnine,
the artificial circulation was stopped so that stagnation of
the oxygen-free saline solution took place in the vascular system.
The processes then occurred in exactly the same manner with
the exception that the state of non-irritability appeared somewhat
earlier. If after the beginning of complete depression artificial
circulation with oxygen-free saline solution was again started, a
certain degree of recovery took place within one or more minutes.
The stimuli were once more effective and produced a number
of contractions. At times, several single contractions, following
each other in more or less quick succession, could be brought
about. But complete recovery or the appearance of even incomplete
tetanic convulsions was never again obtained, whereas by
the introduction of oxygen complete recovery could at once be
brought about. If, however, the circulation with oxygen-free
saline solution was continued, irritability gradually decreased.
The refractory periods after the individual stimuli became longer,176
and in spite of continuous artificial circulation irritability again
disappeared. The experiment shows that by the circulation of
oxygen-free solution irritability can simply be reduced up to a
certain degree. This partial restitution is produced by washing
out the depressing metabolic products. Being desirous to verify
the results of this investigation with greater exactitude I have
requested Dr. Lipschütz143 to repeat the experiments, taking the
utmost possible precaution in respect to the absolute exclusion of
oxygen. Lipschütz has tested the normal saline solution made
oxygen free with the sensitive Winkler method, in which the
slightest trace of oxygen is shown by the oxydation of manganous
chloride to manganic chloride in which the latter in a saline solution
sets free an amount of iodide from iodide of potassium corresponding
to that of the consumed oxygen. These experiments
of Lipschütz have shown that even with the absolute exclusion
of the slightest trace of oxygen a partial recovery can be brought
about by artificial circulation. There can be, therefore, no doubt
that recovery is actually founded on the removal of the depressing
asphyxiation substances by artificial circulation. Moreover
Fillié144 has previously succeeded in the laboratory at Göttingen in
obtaining by the same methods a corresponding result for the
nerve. In both cases the experiments are extremely complicated
and must be carried out with the most painstaking care.
The depressing influence of the asphyxiation products need not
be regarded as a specific effect of poisoning. It can be solely an
expression of mass relations, if we assume that the anoxydative
decomposition is controlled by a chemical equilibrium between
masses capable of disintegrating and products of the disintegration.
It is not possible to give any detailed account as to the part
taken by accumulating asphyxiation substances in the prolongation
of the refractory period. Indeed, we must for the present
relinquish the attempt to delimitate quantitatively the part taken
by the individual constituent processes in the symptoms of depression
resulting from the deficiency of oxygen. We can merely177
say, the individual alterations produced by the want of oxygen,
that is, the restriction and retardation of the oxydative disintegration,
the corresponding increase of the anoxydative decomposition
and the accumulation of the products of incomplete oxydation
and anoxydative breaking down have the same influence in
that they decrease the strength of the response and retard the
rapidity of the decomposition process. These are the general
effects perceptible in the refractory period by the deficiency of
oxygen.
The establishment of these facts of the dependence of the refractory
period upon oxygen are of the utmost importance for
the genesis of fatigue, for the state of fatigue in all aërobic organisms
is invariably brought about by deficiency of oxygen. In
other words: fatigue is invariably asphyxiation. A deficiency of
organic reserve substances never occurs in fatigue before the
effect of oxygen deficiency leads to complete depression, for the
quantity of organic reserve substances at the disposal of the cells
is greater comparatively than that of oxygen. This is shown by
transfusion experiments in which the time involved before complete
paralysis was brought about in the frog by the introduction
of an oxygen-free saline solution was ascertained and compared
with the period which elapsed before complete paralysis took
place, when the same solution saturated with oxygen was used.
Although the previously described experiments on the strychninized
frog show clearly the relations of fatigue to the refractory
period, I should, nevertheless, like to illustrate them somewhat
further.
The state of fatigue as it is developed in a living system by a
continuous functional activity is characterized by a series of
symptoms which can be best studied in the fatigue of the muscle,
the nervous centers, and the peripheral nerves.
If the muscle of the frog is isolated and rhythmically stimulated
with single induction shocks and the muscle contractions
graphically recorded, it will be found that the first perceptible
alteration during the course of stimulation is the increasing
height in the curve, which appears directly after the first
contraction and becomes more and more noticeable after every178
succeeding one. With the isolated apex preparation of the
frog’s heart an effect is produced which Bowditch145 has termed
the “Treppe” and Tiegel,146 Minot147 and others have obtained the
same result for the skeletal muscle. The Treppe has been often
regarded as an expression of increasing of capability of the
muscle following each succeeding stimulus in spite of the fact
that it is physiologically incomprehensible that an isolated muscle
can become more capable by increased demands. Fröhlich148 first
threw light on this seeming contradiction by showing that the
increase in height of the muscle contraction in the Treppe is
in reality the first indication of the beginning of fatigue, and
Fr. Lee149 arrived at the same result. The increase in height of the
contraction curve depends upon the retardation of the course of
contraction. As the contraction extends over the muscle substance
in the form of a wave, a longer stretch of the muscle will
be in a state of contraction when the wave is more extended than
when it is shorter, that is, the shortening of the muscle will be
greater, the contraction curve higher, when the wave is more
extended. With increasing fatigue the retardation in the course
of contraction, as Rollet150 already has shown, becomes continuously
greater. (Figure 34.) The consequence of this retardation
in the course of contraction is, therefore, perceptible in the
rhythmically activated muscle in the form of contracture. As
fatigue increases, the muscle requires an increasing length of time
to relax to its full extent and in consequence the period between
the two stimuli is very soon insufficient for this to occur. There180
remains a certain amount of shortening, when the next contraction
begins. This characteristic extension of the individual contraction
curve of the fatigued muscle is an expression of the retardation
of the oxydative disintegrating processes and of the Treppe. It
shows us that fatigue is perceptible to a slight degree even after
the first excitation. After every succeeding stimulus the oxydative
decomposition in the fatigued muscle is increasingly prolonged.
It is, therefore, self-evident that the capability of action
of the muscle likewise becomes less with increasing fatigue.
Every state of fatigue is, in fact, distinguished by the decrease of
response. This is perceptible in the later stages by the decline of
the height of contraction. Hence all symptoms of fatigue which
we observe form the expression of one single process; it is the
constantly increasing slowness of oxydative disintegration with
increasing fatigue.
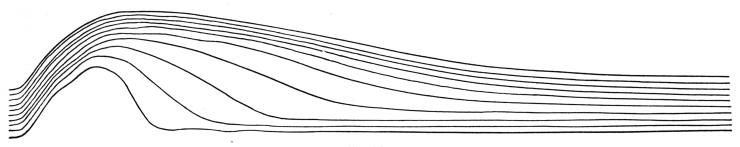
Fig. 34.
Series of muscle curves graphically recorded one over the other, showing the retardation in the course of contraction with increasing
fatigue. (After Rollet.)
Exactly similar conditions as those of the muscle are seen in
the central nervous system. The reflex contraction of the triceps
of the frog produced by stimulation of the central end of the
sciatic nerve with single induction shocks demonstrates clearly
as Ishikawa151 has proved in certain stages of fatigue, an increase
in height and a strong relaxation which does not depend upon the
fatigue of the muscle but on that of the centers. If the fatigue
is greater, the height of the contraction then decreases, whereas
the extension of the course of relaxation increases further. The
possibility of fatigue of the muscle during these experiments was,
of course, precluded by proper precautionary measures. Irritability
and the course of excitation in fatigue of the centers
show exactly the same alterations as developed in fatigue of the
muscle. The processes of oxydative breaking down are retarded
more and more with increasing fatigue, that is, fatigue is characterized
by exactly the same processes as is the prolongation of
the refractory period by the deficiency of oxygen, and likewise
in fatigue this retardation of the oxydative disintegration processes
is conditioned by the relative deficiency of oxygen. This
is shown by the rôle played by oxygen in recovery after fatigue.
181
It was found by Hermann152 in 1867 and confirmed by Mademoiselle
Joteyko153 in Richet’s laboratory, that the isolated muscle of
the frog, which was completely nonirritable as the result of
fatigue, does not regain irritability in an oxygen-free medium,
but does so when oxygen is introduced. The previously described
experiments of artificial circulation in the frog show
clearly how dependent the centers are upon the oxygen supply
for the restoration of irritability. In consequence of the strychnine
poisoning the irritability of the centers is so enormously
increased that the “all or none law” is applicable to the centers of
the spinal cord under these conditions.154 These are the best conditions
for the production of fatigue. One can readily demonstrate
the importance of the oxygen supply for the rapidity with which
irritability returns after fatigue if in the strychninized frog an
artificial circulation is used, at the same time varying on one hand
the amount of oxygen, on the other the activity of the centers. If
a saline solution containing merely a trace of oxygen is circulated,
the centers recover very slowly and incompletely after every
fatigue. Subsequent to every reaction produced by a stimulus,
an increasing length of time is required until irritability is so far
recovered that a new stimulus can meet with response. If, however,
a saline solution is circulated which has been saturated by
being shaken with oxygen and is continuously in a pure atmosphere
of oxygen, recovery takes place in comparison with far
greater rapidity and completeness. If the supply of oxygen is
ample and the stimuli act at longer intervals on the frog, irritability
always is quickly restored in the periods of rest between
the stimuli. With continuous stimulation of quickly succeeding
stimuli, irritability is soon completely obliterated, even though an
abundant oxygen supply be present, and it is not until a pause is
interpolated that oxygen is capable of bringing about a recovery.
By manifold variations of these experiments the connection
between fatigue and the refractory period can be more and more182
clearly recognized. Fatigue is simply the refractory period prolonged
by deficiency of oxygen. In both cases there is a diminution
of irritability. In both cases this diminution is conditioned
by a retardation of oxydative disintegration following every
stimulation. In both cases it is the relative deficiency of oxygen
which produces this delay. In both cases the oxydative decomposition
can be quickened and irritability restored, that is, the refractory
period lessened and fatigue removed by a sufficient supply of
oxygen. The amount of oxygen which suffices to constantly maintain
the specific irritability of a living system in an undisturbed
metabolism of rest is not sufficient if the system is continuously
functionally activated by stimulation. The refractory period
increases after excitation and merges, although very gradually,
finally into permanent nonirritability, that is, into complete
fatigue.
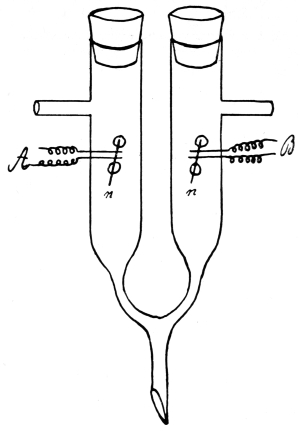
Fig. 35.
Double glass chamber for comparative experiments on
fatigue of the nerve (n n). A and B—Wires of
the electrodes. (After Thörner.)
The knowledge that fatigue represents a prolonged refractory
period resulting from relative deficiency of oxygen has enabled
me with the aid of my coworkers to demonstrate the existence
of fatigue and produce the typical symptoms experimentally for
a living tissue, which up to then was considered indefatigable:
I refer to the medullated nerve. After having found that the
condition necessary for the production of fatigue in the nervous
centers is a deficiency of oxygen, I arrived at the conclusion that
fatigue could only be obtained in the medullated nerve when subjected
to a deficiency of oxygen. Up to that time, however, no
consumption of oxygen was known for the nerve. It was, therefore,
necessary to first ascertain if the nerve possessed an oxydative
metabolism. At my request, H. von Baeyer investigated these
questions. After many vain attempts to obtain absolutely pure
nitrogen, we finally succeeded in finding a method by which it is
possible to gain nitrogen gas, which is, one might almost say, in
a mathematical sense absolutely pure. It was then possible for
H. von Baeyer155 to asphyxiate the nerve and subsequently to bring
about complete restoration by the introduction of oxygen. It
was shown that the nerve requires merely a minute quantity of183
oxygen and only completely asphyxiates when the last trace of
oxygen is removed, and further that recovery takes place within
a fraction of a minute if the oxygen is again supplied. These
experiments which have been carried further by Fröhlich156 were
afterwards confirmed in other laboratories,157 and form the basis
for proving the existence of fatigue of the medullated nerve.
Shortly after, Fröhlich158 was able to demonstrate symptoms of
fatigue in the medullated nerve. He found that the refractory
period of the nerve, which, as previously mentioned, Gotch and
Burch fixed at about .005 second duration, was prolonged by
oxygen deficiency to .1 second, so that stimuli following each
other oftener than ten times per minute produced merely single
initial contractions in the muscle concerned, that is, in a series
of stimuli of which the intervals are less than .1 per second, only
the first produces response, whereas the following occur in the
refractory period, brought about by those preceding, and are,
therefore, inoperative. The nerve is fatigued by the quick succession
of stimuli. The normal nerve on the contrary invariably
responds, as known, to an even more rapid succession of stimuli
with a rhythmical excitation corresponding to the number of
stimuli and which is manifest in the muscle by a tetanus. This
again confirmed the identity of fatigue with the prolonged refractory
period, conditioned by the relative want of oxygen. It
likewise explained the conditions of the analogous behavior that
Wedensky159 had observed in the narcotized nerve, but had neither184
recognized as manifestation of the prolonged refractory period
nor as fatigue. A further advance was made by the investigations
of Thörner. He placed two nerves of the same frog in a double
chamber under completely identical conditions with the exception
that one remained in a state of rest, whilst to the other
tetanic stimuli were applied. (Figure 35.) If this took place in
nitrogen, the irritability of the stimulated nerve invariably sank
with much greater velocity than that of the nonstimulated, whereas
after an introduction of oxygen, even when the stimulation was
continuous, both again recovered. In these experiments of185
Thörner160 the action current and not the muscle contraction served
as indicator. Here the fatigue of the medullated nerve brought
about by the deficiency of oxygen during prolonged stimulation
is demonstrated in the most obvious manner. (Figure 36.)
Thörner161 further succeeded by a continuous stimulation of the
nerve in obtaining even in atmospheric air the indications of primary
fatigue. The symptoms were exactly the same as those
characterizing fatigue of the muscle; the extension of the course
of excitation and, as a consequence of this, the appearance of a
summation of excitation produced by tetanic currents and a reduction
of irritability in response to single stimuli. The form of
the curve, resulting from alteration of irritability in fatigue and
recovery, likewise shows complete conformity with that of the
muscle. (Figure 37.) Finally Thörner162 proved that the nerve,
when fatigued by continuous tetanic stimulation in nitrogen,
could also partially recover in the latter if the stimulation was186
interrupted, whereas a complete recovery could not take place
unless a supply of oxygen was introduced. (Figure 38.) This
fact is in perfect accordance with the relations found by Verworn,
Lipschütz, in fatigue of the nervous centers. It is the
expression for the accumulation and removal of fatigue substances,
the depressing effect of which Ranke163 first established
for the fatigued muscle. The fact that the nerve could also partially
recover in an atmosphere of nitrogen would seem to likewise
contain the proof that among the fatigue substances products
in the form of gas must be present. It is probable that an escape
of carbon dioxide has taken place.
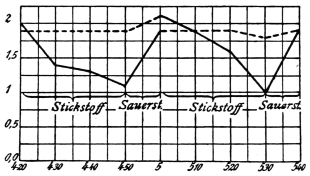
Fig. 36.
Curve of action current of two nerves, one of which is stimulated
(plain line) whilst the other remains at rest (dotted
line). After decrease of irritability of the stimulated nerve
in nitrogen, oxygen is introduced into the chamber and
irritability increases again. Then the previously resting
nerve is stimulated in nitrogen and the stimulated nerve
remains at rest. (After Thörner.)
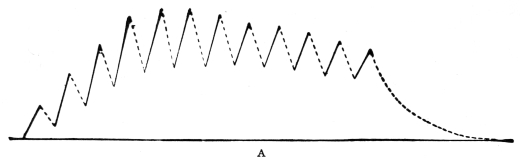
Scheme showing course of fatigue (plain line) and recovery (dotted line) of the nerve
as it is manifested on testing the irritability with tetanic stimuli, when fatigue and
recovery alternate at equal intervals. The curve shows at the beginning an apparent
increase of irritability corresponding to the “Treppe” of the muscle. (After Thörner.)
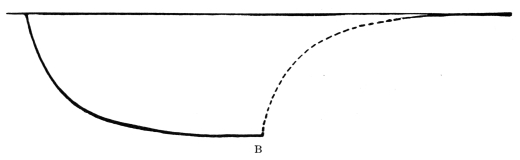
Fig. 37.
Scheme showing course of fatigue (plain line) and recovery (dotted line) on testing the
irritability of the nerve by single induction shocks. In fatigue irritability sinks at first
rapidly, then more and more slowly until a state of equilibrium is reached. Recovery
shows the same in reverse succession. (After Thörner.)
187
As a result of all these investigations, linked together in a systematic
series, the proof has now been obtained that the nerve
like all other living substances is fatigable. Its fatigue is solely
the manifestation of a prolonged refractory period and the extension
of the latter by continuous stimulation is, as in all aërobic
substances, a result of relative deficiency of oxygen.
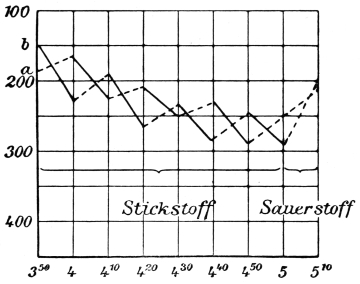
Fig. 38.
Curve of irritability as demonstrated by action current of two nerves
in nitrogen, which are alternatively stimulated (plain line) and at
rest (dotted line). Recovery in nitrogen is always merely partial
and relative. It only increases on introduction of oxygen. (After
Thörner.)
To briefly summarize in conclusion, I will repeat that just as
all living systems show a refractory period after an excitation, in
which irritability is reduced, all living systems are likewise capable
of fatigue. Both are most intimately connected and are based
fundamentally on the facts of metabolism.
An excitating stimulus disturbs the metabolic equilibrium of
rest by suddenly bringing about increased decomposition of certain
substances. During and directly after the breaking down,
irritability is reduced in the same degree as the amount of substances
required for disintegration in response to a succeeding
stimulus is decreased and the quantity of the decomposition188
products is increased. This is the refractory period. By the
metabolic self-regulation in accordance with the principle of
chemical equilibrium, the original metabolic equilibrium is
restored after every excitation. Irritability, therefore, increases
in the same measure as this occurs, that is, in the form of a
logarithmic curve, until it again reaches the specific degree of
irritability of the particular system. The refractory period
diminishes. If the processes of disintegration and self-regulation
are delayed, either by want of substance necessary for breaking
down or the accumulation of decomposition substances, the
refractory period is prolonged and the response to every further
stimulation decreased, that is, the system is fatigued. In all
aërobic organisms the retardation of the course of excitation and
self-regulation under a continuous influence of stimuli is the result
of the relative want of oxygen. The processes of oxydative disintegration
are prolonged and restricted by relative deficiency of
oxygen and merge more and more into anoxydative decomposition.
The products of incomplete oxydative and anoxydative
decomposition accumulate. Both factors decrease the strength
of the response after every stimulation. Thus the want of oxygen
leads to reduced activity. In the anaërobic organisms the refractory
period and symptoms of fatigue are, of course, produced by
the relative deficiency of other substances. Fatigue in the
anaërobic systems has, however, so far not been investigated. We
advance very slowly, step by step, in physiology, and, as in every
science, an acquirement of a new knowledge means a new problem.
In this lies the inexhaustible charm of our scientific
research.
189
CHAPTER VIII
INTERFERENCE OF EXCITATIONS
Contents: Examples of effects of interference of stimuli in unicellular
organisms. Interference of galvanic and thermic stimuli in Paramecia.
Interference of galvanic and thermic stimuli and narcotics.
Interference of galvanic and mechanical stimuli. Interference of galvanotaxis
and thigmotaxis in Paramecia and hypotrii infusoria. Real
or homotop interference, apparent or heterotop interference. The
two effects of homotop interference of excitations: Summation and
inhibition of excitations. Theory of the processes of inhibition.
Hering-Gaskell theory. Inhibition as an expression of the refractory
period. Individual possibilities of interference of two stimuli. Interference
of an excitating and a depressing stimulus. Interference of
two depressing stimuli. Interference of two excitating stimuli.
Analysis of the interference of two excitations. Interference of two
single stimuli. Conditions upon which the result of interference is
dependent. Heterobole and isobole living systems. Intensity of the
two stimuli. Interval between the stimuli. Specific irritability and
rapidity of reaction of the living system. Latent period. Interference
of single stimuli in a series. General scheme of the development of
the effect of interference. Summation and inhibition. Apparent
increase of irritability. Conditions of summation. Tonic excitations.
Conditions of inhibition. Various types of inhibition. Interference
of two series of stimuli. Relations in the nervous system. Peculiarities
of the nerve fibers. Conversion of the nerve by relative fatigue
from an isobolic into a heterobolic system.
Until now the mechanism of the single excitation has received
the major portion of our attention. It was not until we reached
the subject of the origin of fatigue that we became acquainted
with the effects of repeated stimulation. Here we found a case
of interference of individual excitations. But fatigue is simply
a special instance of such interference, for the subject of interference
action occupies a much greater field.
Every cell of the larger organisms, and more especially the
single celled organisms, is subjected to manifold stimuli. It is190
indeed, quite common that two stimuli interfere with each other
and manifold effects follow, depending upon the specific reaction
of the cell and the quality, intensity and duration of the interfering
stimuli. Sometimes the interference effect is readily
understandable from a knowledge of the specific effect of the
individual stimuli concerned. At other times, however, the
specific reaction seems entirely different in nature than would
be expected from a study of the effects of the individual stimuli.
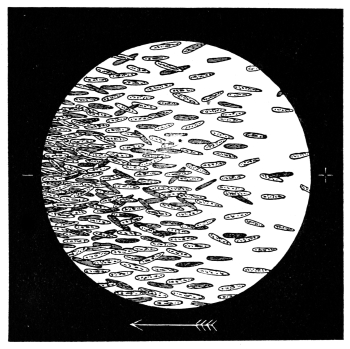
Fig. 39.
Galvanotaxis of Paramaecium aurelia.
When I place a drop of Paramecium culture on a slide having
on two sides parallel pieces of baked clay which serve as electrodes
and allow a constant current of about .2 milliampère to
flow through, it will be seen that the infusoria at room temperature
move toward the negative pole at a rate averaging 1–1.4 mm.
per second. (Figure 39.) If I increase the temperature, the rate
of movement is increased. Here the galvanic and the thermal
stimuli influence each other in such a manner that the reaction191
to the galvanic is increased by the thermal stimulation. This
summation of excitation is readily understood on the basis of the
laws concerning the effect of temperature upon the velocity of
chemical change established by van’t Hoff. If, however, the
Paramecia are in a 1 per cent. alcoholic solution, then, as was
shown by Nagai,164 the rapidity of movement following galvanic
stimulation is decidedly reduced. The interference effect between
the galvanic and chemical stimulation is, because of the depressing
effect of the latter, likewise readily understood.
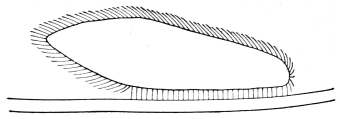
Fig. 40.
Thigmotaxis of Paramaecium aurelia. (After Jennings.)
Greater difficulty meets us, however, in the following instance.
The forward movements of the Paramecia follow in consequence
of the fact that the individual cilia of the body lash more powerfully
backward than forward. If now the Paramecia, while
moving forward, meet with a resisting body, they withdraw sideways
while executing a sudden strong forward ciliary stroke. The
strong mechanical stimulation brings about retraction of the
organism. Entirely different are the results when the impact is
weak. If Paramecia while slowly swimming touch a resisting
object with the anterior portion of the body, withdrawal does not
occur. The infusoria remain under proper conditions in contact
with the resistance, and the rhythmic activity of the cilia directly
against resistance, as well as those on the other side toward the
posterior portion of the body, are more or less inhibited. (Figure
40.) The degree of inhibition brought about by this weak
mechanical stimulation may vary considerably. At times the cilia192
of the whole body suddenly cease their movement. (Figure 41, A.)
At other times, this cessation is limited to the cilia in the
anterior portion of the body (Figure 41, B), while the movements
of those on the posterior portion of the body are of less amplitude
or are irregular and weak. In all cases the infusorium remains
quiescent in the water in contact with the resistance, and it is not
uncommon to find numerous individuals in apposition with
particles of ground, slimy detritus, plant fibers and so forth.
(Figure 41, C.) In short, the rhythmic activity of the cilia of
the Paramecia receiving their normal impulses of excitation from
the ectoplasm of the cell body interfere with strong mechanical
stimuli in such a manner that a negative thigmotaxis develops;
following weak mechanical stimuli a positive thigmotaxis results.
Here is an instance of the relation between the intensity of the
stimulus and the manner in which its effects interfere with an
already existing excitation.
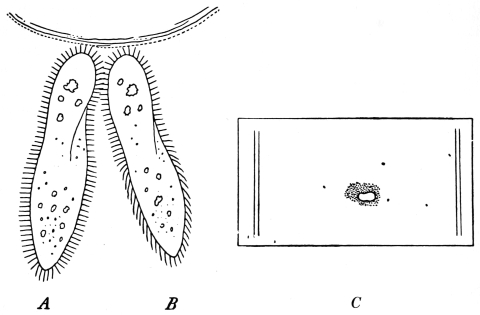
Fig. 41.
Thigmotaxis of Paramaecium aurelia.
However, the strength of the inhibitory effect of a weak contact
stimulus upon another excitation is best appreciated when193
positive thigmotaxis is interfered with by the effect of a thermal
or galvanic stimulus. Jennings165 and especially Pütter166 have, at
my request, more thoroughly investigated my original observations
and have given us a complete analysis of these interesting
interference effects. If the freely swimming Paramecia are subjected
to a constantly increasing temperature, the movements of
these infusoria become more and more active. At 30° C., the
rapidity is very violent and at about 37° C. they reach their maximal.
If now the same experiment is repeated with Paramecia
which have in consequence of thigmotaxis fixed themselves to
particles of slime, the temperature may be increased to 30° C.
without an observable effect. The infusoria remain throughout
in contact with the resistance. Only when the temperature is
37° C. do they release their contact and move violently through
the water. If a drop containing Paramecia is placed on a slide,
between parallel pieces of fired clay which serve as electrodes,
it will be seen that some freely swim about, whereas others
remain thigmotactically in contact with particles of slime. When
a constant current of about .2 of a milliampère is passed through,
it is observed that the freely swimming individuals hasten
towards the cathode. Those attached to objects, on the contrary,
do not respond in this manner to the electrical current.
(Figure 42.) The intensity of the current can be greatly increased
without bringing about detachment of the individuals
from their position of fixation. The typical influence of the
strong current upon the movement of the cilia of the thigmotactically
fixed individuals can be clearly seen. Nevertheless, the
inhibition, brought about by the contact stimulus, predominates
over that of the excitating effect of the current, so that a freeing
of the organisms from their position does not occur. Not until
the current becomes very strong is the excitation thereby produced
sufficient to bring about a separation of the infusoria,
whereupon they immediately swim toward the cathode. In this194
interference between the contact stimulus, on the one hand, and
the thermal or galvanic on the other, the inhibitory effect of the
former may overpower the strong excitation of the latter.
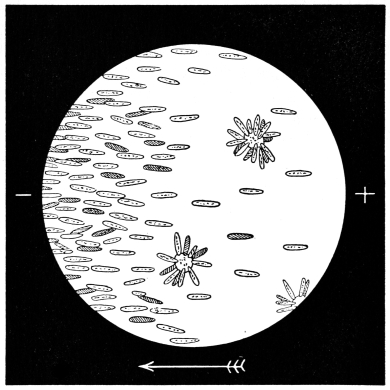
Fig. 42.
Interference of galvanotaxis and thigmotaxis in Paramaecium aurelia. The
individuals which are thigmotactically attached to slime particles
remain at rest while the freely swimming individuals move toward the
cathodic pole.
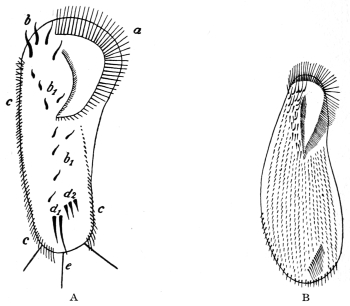
Fig. 43.
Hypotrichous infusoria. A—Stylonychia. B—Urostyla.
Still more complex and striking is finally the following case
of interference between thigmotaxis and galvanotaxis. The
hypotrichous infusoria as Stylonychia, Urostyla, Oxytricha, etc.,
have a marked functional and morphological differentiation of
their cilia. They possess a bow-like row of perioral cilia, which
sweep in the food; a number of cilia on the ventral surface used
for locomotion by which they move about upon objects in the
water; a row of border cilia on each side, which, during swimming,
contribute the propelling force. The perioral cilia also195
form the elements which bring about a screw-like movement on
the axis. They further possess several cilia, which permit a rebounding
of the organism, and finally certain forms have anal cilia,
which probably serve as breaks and to steer the organism.
(Figure 43.) Their usual mode of locomotion is that of creeping,
moving by means of the cilia on the ventral surface. These
movements depend upon the positive thigmotaxis of the cilia of
locomotion. At the same time there is inhibition of the cilia on
the sides. When the infusoria are excitated by a new stimulus,
the cilia used for rebounding become active, the body frees itself
from its position of attachment and begins to swim, wherein the
cilia on the sides, as well as the perioral cilia, act in the manner
mentioned above. I have made the striking observation that the
hypotrichous infusoria respond differently to the galvanic current,
depending on whether they are swimming or in a fixed
position. If one places a drop of water with numerous Urostyla
on a slide between parallel pieces of fired clay which serve as
electrodes, it will be seen, upon the closing of a current, that all196
of the individuals which are freely swimming and turning in
a screw-like manner around their axis, steer immediately toward
the cathode, exactly as in the case of the Paramecia. On the
other hand, those which are fixed to the bottom of the slide as
a result of thigmotaxis, upon closing of the current, make a short
turn and assume a position wherein the long axis is at right
angles to the direction of the current, and the perioral rim is
directed toward the cathode. In this position they move through
the field. (Figure 44.) When the current is broken the individuals
draw backwards, distribute themselves and creep and
swim in all directions in the water. If during the course of the
passage of the current, an individual which has been swimming
begins to creep, the axis immediately assumes the position above
described in the case of the organisms which are in contact with
the bottom and vice versa. The thigmotaxis, therefore, influences
galvanotactically swimming organisms in a most characteristic
manner. As a consequence of the interference of thigmotaxis
and galvanotaxis, the organisms move in a direction transversely
to the direction of the current. This most striking reaction has
been cleared up by Pütter,167 the explanation being based upon an197
accurate investigation of the mechanism of ciliary activity. The
galvanotactic swimming toward the cathode is explained by the
same principle as that applicable to all galvanotaxis.168 As a result
of the excitation produced by the anode, the cell body must
assume a position wherein the border cilia, which are of greatest
importance in swimming, are equally stimulated on both sides
of that part of the body directed toward the anode. It is only
in this position that forward swimming is possible, for as a result
of unsymmetrical excitation of the border cilia a turning must at
once occur, which automatically brings about a resumption of the
position of the long axis. The perioral cilia bring about the
screw-like movement around the axis during swimming. It follows
that the freely swimming individuals must necessarily move
towards the cathode. In the case of the thigmotactically moving
individuals the activity of the border cilia is inhibited. The
perioral and the locomotion cilia bring about the assumption of
the position of the axis, above described. The perioral cilia during
movement bring about a turning of the body on the vertical
axis toward the side opposite that of the orifice and it follows
that the body can occupy only that axial position wherein the
perioral cilia are least excitated. This is, however, only the
case when the long axis of the body is transverse to the direction
of the current, and the perioral cilia are directed toward the
cathode, for stimulation arises from the anode. The reason why
the infusoria do not turn toward the anode from this transverse
position of the axis is to be found in the fact that the anterior
locomotion cilia are stimulated to a greater extent by the turning
toward the anode, and bring about a movement in the contrary
direction. The transverse position of the axis is thus the result
of an antagonistic action between the perioral and the anterior
locomotion cilia. It therefore follows that the characteristic
position, which is necessarily assumed by the thigmotactically
creeping individuals, is brought about by an interference action
between tactile and galvanic stimulation.
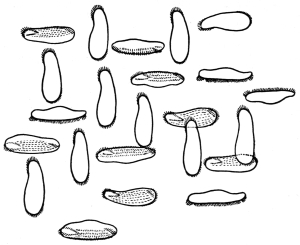
Fig. 44.
Urostyla grandis. Interference of galvanotaxis and thigmotaxis. The
freely swimming individuals move towards the cathode (left side).
The creeping individuals move in transverse direction.
These, then, are a few examples of the interference action of
various stimuli on the single cell. They show us in part fairly198
simple, and in part very complex states. It now behooves us to
obtain a general understanding of interference action, to learn
the fundamental laws in connection with these complex actions,
to shell out, as it were, the general factors involved in the special
conditions. In this connection the examples already referred to
furnish all of the data necessary for our first orientation. In the
simple instance in which the effect of galvanic stimulation was
augmented by increase of temperature and again in the case where
there was a diminution of excitation resulting from the alcohol,
the interference of the two stimuli is consequent upon the
fact that the location of attack is the same. The constant
current acts upon a portion of the infusorium, which also responds
to elevation of temperature. We have a real, or, as I
may term it, “homotopic interference,” for it is an interference
in which the general point of attack is the same for both stimuli.
In contradistinction to this case, we have the examples of the
interference of thigmotaxis and galvanotaxis in the hypotrichous
infusoria. Here the effect of interference, the characteristic
position of the axis of the cell body, is brought about by the fact
that the galvanic stimulus affects different elements than the
mechanical. The turning of a creeping Stylonychia or Urostyla,
when the current is closed, in which the anterior portion of the
body was previously directed towards the anode, results from
excitation of the perioral cilia from the anodic pole. The mechanical
stimulation, on the contrary, exerts its effect upon the
locomotion and border cilia. Only when there is a turning of
the anterior portion of the body towards the anode, would the
galvanic stimulus affect also the anterior locomotion cilia and
thereby counteract turning towards the anode. Therefore, we
have before us in this case of the assuming of a characteristic
position of the axis of the cell body the expression of an apparent,
or, as I prefer to express it, a “heterotopic interference,” in which
the two stimuli do not actually interfere in their action, but rather
influence the final result, in that the condition for the state of
the system in its totality is dependent upon its individual components.
This heterotopic interference is of particular importance
in the bringing about of the movements of the living system.199
The locomotion of the animal and especially the direction is
in part a manifestation of heterotopic interference of response.
At the same time, however, especially in the coördinated movements
of nervous origin, the homotopic interference also plays
an important rôle and, not rarely, is combined with heterotopic
interference.
Although the physical analysis of heterotopic interference is
extremely attractive, we must, however, temporarily set aside its
consideration, for at this point the question arises as to what
happens when there is interference of two stimuli at the same
point. In the heterotopic interference the effect of each stimulus
is the same as if it were applied singly. In the homotopic interference
the interfering effects of stimulation influence each other.
The above examples of homotopic interference introduce us to
the two principal types of these manifold kinds of interference
effects; the excitation brought about by galvanic stimulation is
summated by the excitation produced by temperature. The other
type consists of an inhibition of one effect of stimulation brought
about by another. The depression produced by alcohol on the
Paramecia weakens the excitation of the galvanic current. These
examples of the two principal types of interference effects are
quite simple; nevertheless, in other cases, the conditions are very
complex. This is especially true in the field of nervous inhibition,
so important in the functionation of the nervous system,
and which has presented the greatest difficulties to physiological
investigators until the last few years. That a stimulus bringing
about excitation in a ganglion cell can be inhibited by another
exciting stimulus, or that the development of excitation in a
ganglion cell may be prevented by another exciting stimulus cannot
be easily understood. The problem as to how two interfering
excitations can bring about inhibition is one that has received
many explanations. An interesting incident in the history of
physiology is that the first explanation of the principles of inhibitory
processes was close on the track of being a correct one,
but was subsequently abandoned by its originator. Schiff169
(1858) has endeavored to explain this inhibition as a manifestation200
of fatigue, and this idea he defended with the greatest
tenacity for a long time, until finally, twenty-five years after, in
a treatise which he called “Abschied von der Ershöpfungstheorie,”
he renounced the idea as untenable.
Among other investigations, which since this time have been
made to explain the mechanism of inhibition, those of Gaskell,170
Hering171 and Meltzer172 have received widest consideration. These
theories are built upon the existence of the two phases of metabolism,
and assume that inhibition, in contradistinction to dissimilatory
excitation processes, depends upon an increase of the
assimilative processes. The principal evidence which Gaskell
advances is that when the vagus nerve of the tortoise heart, a
typical inhibitory nerve, is stimulated, a positive variation of the
demarcation current of the heart muscle occurs, whereas when a
motor nerve of a skeleton muscle is stimulated the attached muscle
shows a negative variation of the demarcation current. I must
confess that this explanation of inhibitory processes, from the
standpoint of an interpretation of processes in the living substance,
seems very plausible, and I have accepted this even in my
address on excitation and depression before the Frankfurter
Naturforscher Versammlung.173 I have since then endeavored to
obtain experimental evidence to substantiate this theory, in that
I attempted to prove that increase of the assimilatory processes
brought about by stimulation would be associated with a reduction
of the specific irritability. For this purpose I have sought
for such cases in which a stimulus primarily and momentarily
increases assimilative processes in a system in a state of metabolic
equilibrium. I was disappointed, when, after years of investigation,
I could not find such cases. There is only one kind of
stimulus of which we can say with positiveness that it primarily
increases the assimilative processes, that is, increased supply of201
food. But here the increase in the processes of assimilation never
occurs momentarily, and indeed this increase is so extremely
slight that it can only be demonstrated over a long course of time.
These totally negative results of my investigation had awakened
strong doubts concerning the assimilation hypothesis of inhibition.
Above all, this explanation seemed to me to be impossible for the
nervous system. I searched, therefore, for another explanation
for the processes of inhibition in the nervous system. If the
increase of energy production resulting from the application of
a stimulus is dependent upon an excitation of a dissimilative
nature, then one is justified to look upon the reduction of functional
energy production as an expression of an antagonistic process
to that of dissimilatory excitation. In this respect the
Gaskell-Hering hypothesis of inhibition rests upon a firm foundation.
When, however, this hypothesis assumes an antagonism
between dissimilatory and assimilatory excitation, then it must
not be overlooked that a second antagonism is possible between
dissimilatory excitation and dissimilatory depression. The antagonism
need not involve the two types of metabolism, it may
depend upon variations of one type. When, therefore, the hypothesis
that inhibition is brought about by assimilatory excitation
meets with insuperable difficulties, the possibility should be
considered if it is not more likely dependent upon dissimilatory
depression. These reflections induced me to investigate if conditions
could not be produced experimentally wherein dissimilatory
depression could bring about inhibitory processes in the
nervous system. The most essential requirement was, that dissimilatory
depression should quickly develop and pass away with
like rapidity, for inhibition of the nervous system sets in momentarily
and disappears again momentarily. Another important
requisite is, that both interference stimuli are individually capable
of producing dissimilatory excitation, for the inhibitory processes
of the nervous type may be assumed to be the result of dissimilatory
excitation which produce by their interference inhibition,
for the nerve fibers, as already stated, are capable of conducting
only dissimilatory excitation to the responding organ. As I
studied the problem in this manner, it became clear to me that all202
the conditions necessary for the genesis of inhibition are realized
in the existence of the refractory period, and that I had already
produced inhibition by prolonging the refractory period, by
oxygen withdrawal, in the strychninized frog. If we take a
strychninized frog in which the refractory period has been somewhat
prolonged by oxygen withdrawal, so that the reaction is
simply a short reflex contraction, and rhythmically stimulate the
skin, a reaction is only obtained with the first few stimuli, which
reactions rapidly decrease until a stage is reached wherein the
succeeding stimuli are completely inoperative. (Figure 45.)174
This inhibition is demonstrated even more clearly by the following
experiment. Contractions of the triceps muscle of a strychninized
frog are recorded which reflexly follow from stimulation of the203
central end of the cut sciatic nerve. Oxygen is withdrawn in the
manner already referred to. At the proper stage of oxygen
deficiency, rhythmic induction shocks applied to the central end
of the nerve, the interval between the individual stimuli of which
being longer than the duration of the refractory period, elicit
reflex contractions of the muscles of the posterior extremity on
the opposite side following each individual stimulus. If, however,
in the same stage the central end of the nerve is stimulated
with induction shocks at intervals briefer than the duration of
the refractory period, a contraction is only observed during the
very beginning, being brought about by the first stimulus, whereas
the subsequent stimuli are ineffective, the muscles remaining at
rest during their entire application. (Figure 46.) Tiedemann175
at a later date continued these observations and analyzed them
more in detail. In all these experiments, therefore, there is an
interference of the frequent stimulus, because each succeeding
stimulus occurs in the refractory period of the proceeding. In204
consequence there is a strong reduction of irritability and reaction
is absent. That is, the centers during application of the
frequent current are inhibited. If cessation of stimulation by
frequent shocks takes place, stimulation by slowly succeeding
individual shocks becomes effective again in a few seconds. This
is the simplest example of the process of inhibition and by it I
was led to seek in the refractory period the key of the mechanisms
of the process of inhibition. This principle once recognized,
further material for the more detailed working out and extension
of the theory was gathered from the experiences already gained
during the course of the preceding years in the researches on
fatigue and the refractory period in the nerve. Here it became
apparent that the processes resembling inhibition discovered by
Schiff in the nerve preparation and which were studied anew at
a later date by Wedenski, F. B. Hofmann and Amaja and in part
attributed by Hofmann to fatigue of the nerve endings, by
Fröhlich to fatigue of the nerve itself, were in principle of the
same nature as the central inhibitions themselves. Fröhlich,176 by
his analysis of the observations of Richet, Luchsinger, Fick,
Biedermann and Piotrowski on inhibition in the claw of the crab,
then showed that inhibition can be influenced by the alteration of
the intensity of the stimulus as well as its frequency. In a series
of experimental researches he could then demonstrate that the
widely extended antagonistic inhibitions and other special processes
of inhibitions in the centers could on the basis of the same
principle be physiologically explained. Here the supposition was
confirmed that the development of a relative refractory period
plays a very important rôle in the inhibition of the nervous centers.
Thus, the relations of the processes of inhibition to the
refractory period, once established, their entire field, up to then205
shrouded in darkness, has gradually in the course of years been
completely elucidated.

Fig. 45.
Lower line indicates stimuli.
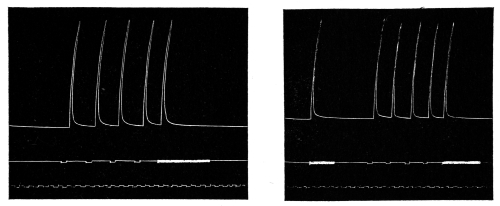
Fig. 46.
Reflex inhibition in the strychninized frog. Lower line indicates seconds, upper line stimuli.
When stimulation with single shocks at longer intervals is applied, each single stimulus
is effective. When faradic stimulation is used, only the first stimulus is operative,
and during the further continuance of stimulation inhibition takes place in the spinal
cord.
Before going back to the cases of inhibition and explaining
them by this general principle, it is necessary that we penetrate
more deeply into the details of the characteristic course of the
refractory period. By this means we will find the conditions
which universally determine the interference in the effects of
stimulation.
First of all, it is self-evident that the occurrence of interference
of stimulation in a living system can only take place when the
succeeding stimulus is applied before the effects of the previous
one have completely disappeared. Within the interval, however,
which is involved from the moment of the beginning of a stimulus
until its effect disappears through the self-regulation of metabolism,
there is the possibility of various interference results from
stimulation.
If we take into consideration the various instances which can
arise, perhaps we may best start with that type wherein the first
stimulation produces depression, whereas the second has an exciting
effect on disintegration. In this type the response to the
second stimulus is weaker than when the second stimulus alone
is applied. As a concrete example of this type, we may refer to
the interference of an induction shock in a nerve during the
relative want of oxygen. We arrange a nerve of a nerve muscle
preparation of a frog in a glass chamber, as already described, and
determine the threshold of stimulation of the stretch within the
chamber by the weakest induction shocks which produce response.
The oxygen is then removed and the effect on the threshold determined.
As shown by Baeyer it is found that with increasing
asphyxia the threshold of stimulation for induction shocks becomes
continually higher. The irritability is likewise decreased.
This occurs, as the investigations of Lodholz show, at first slowly,
then more and more rapidly. The curve of the decrease of irritability
has a logarithmic form. During the continuation of the
depressing stimulus, i.e., the want of oxygen, the exciting stimulus
has less and less effect. If oxygen is again brought in contact
with the nerve, irritability immediately returns to its original206
height. The cessation of the depressing stimulus has, therefore,
the effect that the exciting stimulus again brings about its original
response.
A second type of interference is produced when both stimuli
bring about depression. As an example, we may select the interference
of cold and deficiency of oxygen. If we assume, for
instance, that each of these stimuli of itself brings about only a
partial reduction of living processes and not a complete suppression,
then it would be possible to think of a summation of both
depressions. Nevertheless, the conditions for the summation of
depression have never been carefully analyzed. Quantitative
investigations upon the interference of depressing stimuli are
entirely lacking. One should not, however, in physiology presuppose
what may happen under certain given conditions without
first making the necessary experiments. The strength of
scientific investigation depends upon the fact that every deduction,
no matter how small, must be substantiated by experience
before further progress can be made. So, likewise, we must
await the results of thorough experimentation upon the interference
of depressing stimuli before we can establish a law. The
conditions are not as simple as they appear on first observation,
for the point of attack of the various kinds of the depressing
stimuli upon the chain of metabolic processes may be very different.
In such a case it is not at once possible to understand the
results of the interference.
There is a third type in which two dissimilatory excitations
interfere with each other. Fortunately there is a great amount
of experimental data at our command so that today we have a
clear understanding of the essential points of the conditions
necessary for the development of summation of excitation on the
one hand, and inhibition on the other. If we take an instance
of a momentary dissimilatory excitation operating upon an
aërobic system in metabolic equilibrium, it is necessary to recall
the two effects thereby produced. The stimulus brings about an
oxydative decomposition of the living substance. Likewise there
is a reduction of irritability. Both of these alterations are the
foundation of interference. Both processes have a specific time207
of occurrence. The disintegration, determined by energy production,
reaches a maximum suddenly, then diminishes, at first
rapidly, then more and more slowly until the zero point is reached.
In an analogous manner the irritability abruptly reaches a minimum,
then increases rapidly, then more slowly, until it again
reaches its previous value. When we represent these processes
by a curve, they assume the following form. (Figure 47.) In
this diagram the abscissa is the time, the ordinate value zero is
the level of the metabolism of rest and the specific irritability.
The points above the abscissa represent disintegration, that is,
energy production, those under the abscissa, the reduction of irritability.
A consideration of the latent period may be omitted.
At the end of the curve the effect of stimulation may be assumed
to have disappeared and the state of metabolic equilibrium reestablished.
If we base our further observations upon this curve
of excitation, we can study in them the factors upon which
responsivity is dependent when a second exciting stimulus is
operative during the course of the first.
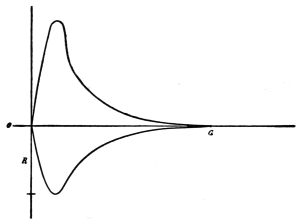
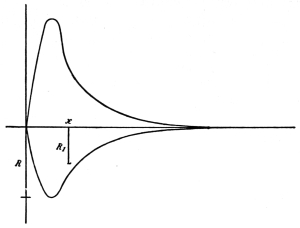
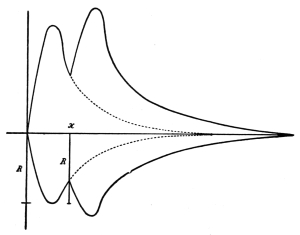
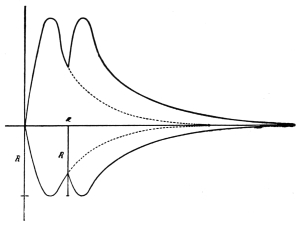
It is from the beginning apparent that the response to the
second stimulus is determined by the intensity of the second
stimulus in relation to the degree of irritability which exists at
the moment when this is effective. This relation is dependent
first upon the absolute intensity of the second stimulus. In the
following diagram the intensity of the existing threshold value is208
fixed for convenience as ordinates beneath the abscissa. If, for
example, at the time point x, a stimulus of weak intensity R1 acts,
this stimulus being under the existing threshold, produces no
perceptible effect. (Figure 48.) If now instead of a weak stimulus,
one of stronger intensity acts at the time point x, this stimulus
will produce an appreciable response. (Figure 49.) If the second
stimulus is of the same strength as the first, this second stimulus
will bring about relatively less disintegration, because the
system is then in a state in which irritability is still reduced.
But this lessened disintegration in that it summates the excitation
still existing as the result of the first stimulus can produce
an absolute increase of the height above that of the abscissa.209
Here then we see the possibility of an increase of response
resulting from summation. Accordingly the increase of disintegration
must occur simultaneously with a diminution of irritability
and this must fall below the level of the reduction of
irritability produced by the first stimulus. This augmentation of
the response through summation above the level of that produced
by the first stimulus acting upon an unexcitated system is,
however, connected with another condition. The above example
refers to systems in which weak stimuli bring about weak response
and strong stimuli strong response, that is, the response
is capable of increase. In systems in which the “all or none
law” is applicable, such an alteration in the absolute height of
excitation, as results in summation, is not possible. In order to
characterize these two types of living systems by a short expression
rather than by a long sentence, we will call the first a
“heterobolic system,” the latter in which the “all or none law” is
operative an “isobolic system.” The former term expresses
various degrees of discharge depending upon the intensity of the
stimulus, the latter term refers to the constancy of discharge
following stimuli of various intensities. Isobolic systems are in
contradistinction to the heterobolic systems not capable of summation.
The response to the second stimulus of equal intensity
cannot be greater than that of the first, it may be equal to the
first (Figure 50) or be less in extent, but it can never be greater210
than that resulting when a single stimulus is applied. These facts
have been known for a long time in the case of the heart muscle.
A word is necessary, however, concerning the effect of stimuli
beneath the threshold in heterobolic systems. We must here
distinguish between the “ideal” threshold, beneath which the
influence of a stimulus is nil, and the threshold of perceptible
effect, beneath which a stimulus apparently has no effect; nevertheless
a weak effect does occur, as is shown by succeeding reactions.
This effect is manifested by a sub-threshold disintegration
and a corresponding slight reduction of irritability.
(Figure 51.) The presence of such a sub-threshold effect is
recognized by various facts as, for example, the summation of the
sub-threshold stimuli to production of a perceptible result. Thus
stimulation of a sensory spinal cord root with a single sub-threshold
induction shock will not produce any evidence of a
reflex excitation, whereas, when induction shocks of the same
strength and of sufficient frequency are applied, a strong reflex
contraction results. The fact that sub-threshold stimuli can bring
about sub-threshold effects is also important in consideration of
the result of interference. The relation between the intensity of
the second stimulus and the degree of irritability of the system,
the intensity of the stimulus being absolutely constant, depends,211
secondly, upon the momentary amount of irritability which exists
just at the time when the second stimulus produces its effects.
It is, therefore, clear that the response produced by interference
must also alter with the momentary degree of irritability in a
manner analogous with variations of the intensity of the second
stimulus. One must, therefore, know the factors which control
the momentary degree of excitation.
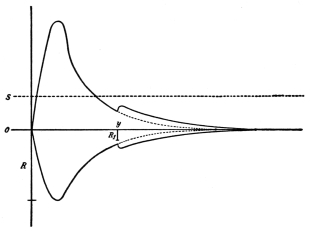
Fig. 51.
Effect of sub-threshold stimuli. o—Level of the ideal threshold.
s—Level of the threshold of perceptible effect.
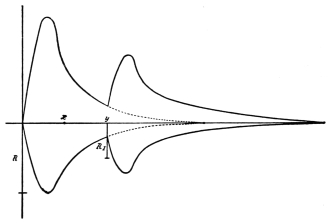
The first factor to be considered is the moment of time in which
the second stimulus is applied, that is, the interval between the
first and the second stimulus. If, for example, a weak second
stimulus follows very quickly after the first, the stimulus will
bring about no response, as the system at the time of its application
is in a relative refractory period. (Figure 48.) The
stimulus is, therefore, under the threshold. If, however, a stimulus
of the same strength is applied somewhat later, when the irritability
has already increased to a somewhat greater extent, then
at this moment the stimulus is above that of the threshold and
a response is obtained which, on account of the state of irritability
existing, is summated. (Figure 52.) But further, it is not a
question of the absolute interval between the stimuli, but rather
to the relative interval to the specific rapidity of the reaction of
the living substance under consideration. There are living substances,
as we have seen, in which the refractory period is unusually
short, as, for instance, the nerve. There are other substances212
wherein this period lasts a considerable time after stimulation,
that is, before the irritability returns to the original level,
as, for example, the smooth muscle. Indeed, depending upon
the specific properties of a system, a short or a long interval is
required before a stimulus of a given intensity is again operative.
Finally, in one and the same living system the duration of the
refractory period can be very different, depending upon the
momentary state of the system. Above all we know that the
refractory period is considerably prolonged in fatigue and likewise
after the influence of other agents, as narcotics, lowering of
the temperature, etc. In such states a second stimulus remains
inoperative when it follows at a definite interval from the first,
whereas under normal conditions the same stimulus applied at the
same interval would be operative.
Finally, there is another factor to be considered, namely, that
the latent period of the second stimulus is more and more prolonged
as the second stimulus approaches more closely to the
absolute refractory period of the first. In the above schemes the
latent period was not taken into consideration because practically
for all the intervals of stimulation considered at that time it could
be assumed to be the same. When, however, a decrease of the
intervals between the individual stimuli takes place, the prolongation
of the latent period can then not be overlooked, as it leads to
a retardation of response. (Figures 29, 30.) This fact was shown
in the classic investigations of Marey177 upon the refractory period
of the heart, and more recently has been the subject of study
by Samojloff,178 Keith Lucas179 and Gotch180 in the muscle and
nerve. These, then, are the essential factors which bring about
interference, and although there are special details which deserve213
more close analysis, nevertheless, we are in a position to attribute
to them the origins of summation and inhibitory processes, which
occur in all living systems, especially the nervous system.
For the analysis of summation and the inhibitory processes
which occur in the physiologically active organisms or which are
experimentally produced, a very important point should be observed,
that is, the fact that the stimuli which bring about these
phenomena are practically always a series of single stimuli. The
nerve impulses, for example, consist of a shorter or a longer
series of single discharges which follow each other in rapid
rhythmic sequence. Here, then, we have the conditions necessary
for the production of interference effects when these single
stimuli follow each other with sufficient frequency and also when
there is the combined action of two series.
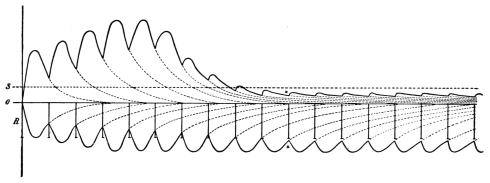
Fig. 53.
Curve showing the general development of the effect produced by interference of the
stimuli of the same series in an heterobolic system. The effect is first summation
and then inhibition. R indicates the intensity of the stimuli, S the level of the
threshold of perceptible effect.
We will first direct our attention to the simplest case brought
about by an interference between the individual effects of stimuli
in the same series. We will study the effect, which here occurs,
in the accompanying diagram, which shows the facts involved in
the interference of two stimuli of a series of stimuli. (Figure 53.)
The curve shows the development of summation and inhibition.
The single stimuli of equal intensity follow at the same
intervals, so that the succeeding stimuli meet with an incomplete214
recovery of excitation and accordingly a decreased state of irritability.
In spite of the diminution of the relative response to
each stimulus the summation of excitation brings about an
absolute increase of the same. At the same time the irritability
decreases more and more, for after each stimulation the oxydative
disintegration as well as restitution require a progressively greater
time and a relative fatigue must, therefore, necessarily develop.
The summation, consequently, reaches its limit very soon and
then decreases progressively, for, as a result of the increase of
fatigue, the oxydative decomposition which occurs at the instant
of every stimulation reduces and with this the energy production
becomes less and less. The system is relatively refractory for
the given intensity of stimulus. Accordingly the response to stimulation
falls below the threshold of perceptible response (dotted
line S) and finally an equilibrium between disintegration and
restitution occurs, wherein the small amount of material used at
each stimulation by oxydative decomposition is again replaced
before the next stimulus. In other words, the irritability is reduced
at each stimulation to an amount equal to that of the
recovery in the interval. If this all takes place beneath the threshold
of perceptible response, the system during the continuance
of the stimulation seems responseless, that is, inhibited.
The inhibition consists then of a reduction of irritability below
the perceptible threshold of response of the stimulus concerned.
It depends upon a continued lessening of dissimilative excitation
to a low level through the delay of the oxydative decomposition
processes. The inhibition is according to this a relative fatigue,
which is conditioned, as is true of every fatigue, by a lengthening
of the refractory period following a relative deficiency of oxygen.
The processes of inhibition are simply and solely an expression
of a refractory period persisting as a result of dissimilatory
excitating stimuli.
Accordingly the general conditions requisite for summation
on the one side and inhibition on the other may be formulated
as follows:
A summation may develop in a heterobolic system and by the
use of submaximal stimuli. It always develops when the following215
stimulus is applied before there is complete recovery of excitation
from the previous stimulus. The absolute increase of
excitation as a result of summation is, however, limited by the
diminution of irritability. By continuation of the series of stimuli
the state of equilibrium between the amount of excitation and
the irritability will be established on a higher or lower level.
There occurs then, depending on whether the feeble persistent
excitation remains above or below the level of perceptible effect,
either a tonus or an inhibition.
Summation can be transformed into inhibition by the continuance
of stimuli of constant intensity. The principles which underlie
both processes are in no way antagonistic and indeed are not
separated by distinct boundaries. The diagram here shown
(Figure 53) illustrates this development of summation and inhibition.
The time required for this development is in manifold
ways influenced by variations of the above-stated factors which
control the occurrence of interference. Thereby results an
immense number of special cases which differentiate themselves
in characteristic manner depending on whether an isobolic or
heterobolic system is involved, depending on whether the irritability
of the system, as measured by the threshold of stimulation,
is high or low, depending on whether fatigability is great or
small, depending upon the intensity and frequency of the stimuli,
etc. Analysis of every instance shows us different combinations
of the interaction of the individual factors. It is, therefore, self-evident
that we cannot here analyze a greater number of these
cases of summation and inhibition. I wish only to refer to a few
typical examples at this time.
It is known that summation of excitation in the normal nerve
does not occur. As already stated, the nerve is a system in which
the “all or none law” is operative. Such isobolic systems do not
summate, having no power of summation because each individual
stimulus brings about a maximum response. But we have seen
that the nerve, as a result of depressing factors, such as deficiency
of oxygen, narcosis, fatigue, etc., which decrease its irritability,
can be transformed from an isobolic into a heterobolic system.
In this state the nerve possesses the capability of summating216
excitations. Waller,181 Boruttau,182 Boruttau and Fröhlich,183
Thörner184 and others have shown that the action current of the
nerve during the application of tetanic stimulation becomes decidedly
greater during a certain stage of narcosis or asphyxiation,
so that the wave of negative variation is higher than when the
nerve is excitated by a single induction shock. Fröhlich185 first
threw light upon this subject in that he made the observation
that here a principle is involved which has far-reaching importance
in the phenomena occurring in the organism. He showed
that as a result of fatigue, cold and narcosis, etc., the course of
excitation brought about by the single stimulation undergoes
retardation. These conditions within certain limits become more
favorable for the production of summation, because each succeeding
stimulus meets with a more incomplete recovery of excitation
than the one previously applied. In consequence of this, the irritability
of the system in the beginning of fatigue, or narcosis, or
immediately after the application of cold, is apparently increased.
This “apparent excitation,” as it was called by Fröhlich, depends,
however, in reality upon a beginning depression which is evident
in that the course of the individual excitations are lengthened by
this means. The irritability is likewise also reduced. Reinecke186
later studied in further detail the retardation of excitation in the
muscle and attributed to this the characteristic property shown
in muscle in the so-called “reaction of degeneration.” Fatigue,
asphyxia, cold, degeneration, in fact all factors which retard the217
course of excitation, are favorable to the summation of excitation,
provided their influence does not exceed certain limits.
Although the nerve as an isobolic system can only be rendered
capable of exhibiting summation when artificially influenced,
there are other forms of living substance which normally are
systems with a slow course of excitation, in which excitation
may be summated, for this type possesses at the same time a
heterobolic character. For example, a single mechanical excitation
elicits a hardly perceptible response in Amœba, Actinosphærium,
Orbitolites. When it is perceptible at all, there occurs
a short interruption of the centrifugal movement of the protoplasm.
After a pause the movement of the protoplasm and the
stretching out of the pseudopods again return. But if the
organism is agitated one or more minutes by rhythmically shaking
the edge of the slide by a special device, as a result of the
summation of weak excitations there occurs a complete drawing
in of the pseudopods and the amœbæ become bell-shaped.187 The
ganglion cells also possess a great capability for summation. We
have already alluded to the fact that single induction shocks
below that of the threshold produce no evident effect, whereas
when rapidly repeated, summation occurs with reflex reaction.
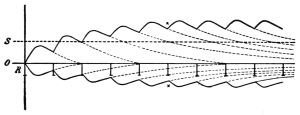
Fig. 54.
Development of tonus by interference of sub-threshold stimuli.
S—Level of the threshold of perceptible effect.
The summation of sub-threshold excitation to a certain height
offers very favorable conditions for the development of tonus.
(Figure 54.) This fact has been established for many kinds of
centers (cardio-inhibitory center, vasomotor center, etc.). During
the continuance of a series of stimuli, as we have already seen,
an equilibrium between disintegration and replacement soon takes218
place. The level of this state of equilibrium depends upon the
relative intensity of the stimuli. It is lower in the case of strong
and higher in that of weak stimuli. This fact becomes apparent
from the researches of Thörner188 on the fatigue of medullated
nerves in air. This investigator showed that during continued
tetanic stimulation of the nerve, the irritability fell to a certain
level, at which it remained so long as stimulation persisted. The
irritability decreased to a new level when the strength of the
stimulus was increased. These interesting experiments of Thörner
show that the level reached when stimulation is continued is
higher as the intensity is weaker. It is, therefore, clear that this
level in summation of stimulation beneath the threshold can be
above that of the threshold of perceptible response, that is, a perceptible
tonic excitation may result. In the genesis of tonus in
the muscle, there is another point to be taken into consideration.
Here we have a combination of a heterotopic interference with a
homotopic interference, for the total shortening of the muscle is
brought about in part by several contraction waves which occur
at various points at the same time and which follow each other,
therefore have a heterotopic sequence. If we consider a long
stretch of muscle, to one end of which a stimulus is applied, it will
be found that the contraction wave moves throughout the entire
length. If after a certain interval of time a second stimulus is
applied, the resultant wave moves along the muscle but does not
necessarily homotopically interfere with the first. In short, there
are two waves of contraction occurring coincidently in the muscle,
the muscle is now more strongly contracted. Fröhlich189 has made
the fact intelligible by this means that tetanic shortening of a
muscle is greater than that of maximal shortening which can be
produced by strong single stimulation. This heterotopic interference
dare not be overlooked in the genesis of muscle tonus.
If it is true, as appears from the investigations of Keith Lucas,190219
that the “all or none law” applies to striated muscle, then an
increase of the contraction from homotopic summation cannot
occur, because an isobolic system cannot show an increase of its
already maximal excitation by summation. Such being the case,
the tonic shortening of striated muscle can only be explained as
an expression of a heterotopic interference.
If we assume that the summation of sub-threshold stimulation,
by increasing excitation, brings about a state of equilibrium from
below, as it were, so also inhibition may be assumed to be the
reverse, the level of equilibrium being reached from above, as it
were, by decrease of the primary excitation from strong stimulation.
This is expressed in our general scheme of the development
of summation and inhibition resulting from the effect of a
series of stimuli. At the same time the first part of the curve to
the fall of irritation to the level of the sub-threshold equilibrium
can be shortened to a minimum by strong stimulation or greater
frequency of the same, and we have then the type of inhibition
with primary excitation. As example of this I wish to again recall
the strychninized frog which was used in the fundamental experiments
for understanding of the theory of inhibition. If we stimulate
a sensory nerve of a strychninized frog, in which the refractory
period is already lengthened, with rhythmic single induction
shocks of slow frequency, the muscle arranged to make a graphic
record will show reflex contraction following each stimulus. If,
on the other hand, we apply a series of stimuli, consisting of single
stimuli rapidly repeated, contraction is produced only by the first,
or the first few stimuli (Figures 45 and 46, pages 202, 203). For
the succeeding stimuli the centers remain inhibited, because each
succeeding stimulus occurs in the refractory period of the former.
The origin of this inhibition shows us with particular clearness
how excitation produced by each single stimulus depending upon
the frequency of the same, falls rapidly or slowly beneath the
threshold of perceptible response. In this case, the state of equilibrium
is reached which is maintained by the following stimuli.
That a single stimulus is not entirely without effect upon this
state of equilibrium follows from the fact that during the continuation
of the stimulus a recovery to the point of observable220
response does not occur, whereas such is the case immediately
upon the discontinuation of the stimulus. In inhibition, then, the
dissimilatory excitation produced by a single stimulus falls to a
low level as a result of the reduction of irritability and remains
at this level continuously. Inhibition as well as tonus is based
upon the development of a state of equilibrium between excitation
and recovery, or disintegration and restitution of the living
substance under the continuous effect of a rhythmic series of
stimuli. They differentiate themselves essentially by the height
of this equilibrium, which is dependent upon the intensity of the
stimulus.
We have to the present considered only the simplest conditions
existing as a result of the effect of a single series of stimuli and
also of the interference of its individual members. These elementary
conditions are at the basis of an understanding of complicated
interference effects which arise when two series of stimuli
interact. In that these processes can be readily explained by
the elementary processes previously described, I will, therefore,
dwell but briefly on this subject. From the standpoint already
taken it may be readily presumed what will happen when two
series of stimuli act upon the same system.
When there is interference of two series of stimuli, there are
two resultant possibilities. In one type the stimuli of the one are
active simultaneously with that of the other. In this instance
both stimuli would act as a single stimulus of greater intensity,
and we have essentially the same condition as exists when a single
series is operative. Nevertheless, such cases are practically hardly
realized in the physiological happenings of the organism. More
often a state exists wherein the single stimuli of one series occur
in the intervals of the stimuli of the other. In these cases there
is an increase in the frequency of the stimuli applied in a given
length of time. We have here, then, in principle the same conditions
as when a series of greater frequency is operative. (Figure 55.)
The effect of such alteration in the frequency consists in an
increase of the velocity of the development of summation or inhibition,
as the general scheme (Figure 55) has shown us. Depending
upon the special combination of the factors involved in interference,221
we may have a summation of the exciting effect of each
series of stimuli or an inhibition of one series by the exciting
effects of the other series. If the frequency of both series is
essentially different, we may have here the conditions for periodically
increasing and decreasing excitations. Nevertheless these
conditions have not been systematically analyzed and experimentally
studied.
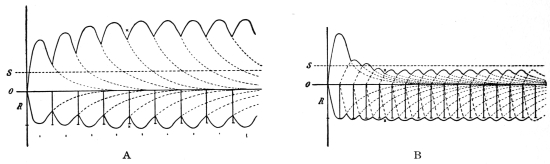
Fig. 55.
Interference of two series of stimuli. A—Effect of the one series alone. Development of tonus
by summation. The dots below the curve indicate the points of time at which the stimuli
of the second series will operate. B—Effect resulting from the interference of both series.
By the addition of the second series the frequency has been doubled. The result consists
in an inhibition.
The greatest number of instances of the interference of two
series of stimuli have been given to us by investigation of the
physiology of the nervous system. In the functionation of the
nervous system the fact that two series of stimuli from different
tracks affect the same ganglia plays a very important rôle. It
is this to which Sherrington191 has alluded as “the principle of
the common path.” Where two nervous excitations involve the
same paths, there arises an interference of the effect of the two
series of stimuli, for the impulses in the nervous system, as
already stated, possess a rhythmic character. This principle has
a broad application in the phenomena of association in the cerebral
cortex. The simpler and, therefore, the most easily understood
cases are, however, in the spinal cord. The motor neurons
of the anterior horns of the spinal cord are the junction of a222
great number of tracks, for example, the sensory neurons of the
spinal cord at different levels, the neurons of the cerebellum, the
pyramidal tracks from the motor areas of the cerebral cortex,
etc. On the contrary, for example, the sensory neurons of the
spinal cord are strictly “private paths” in the sense of Sherrington,
for excitation can enter by this means only from the special
paths of the spinal ganglia and, therefore, from the periphery.
The motor neurons of the anterior horns offer, therefore, excellent
opportunities for the experimental investigation of the interference
of two series of excitations which enter by different
paths. The spinal cord consequently has become a much-used
object of investigation for this purpose. In fact, we can observe
and produce all types of interference in the spinal cord. These
conditions have been quite thoroughly investigated by Sherrington192
and his coworkers on the dog, and Fröhlich,193 Vészi,194 Tiedemann195
and Satake196 on the frog.
A summation of two excitations was observed already by
Exner. This investigator connected the abductor pollicis of the
rabbit with an apparatus for making graphic records. He then
stimulated first the paw and then the motor areas of the cerebral
cortex with faradic shocks, the intensity of which was just sufficient
to bring about perceptible effect. If both stimuli were
simultaneously operative, an increase in the response was observed.
Even when the stimuli were sub-threshold in type, as a
result of summation there was a perceptible muscle contraction.
(Figure 56.) Exner had at that time referred to this increase of
the response as “Bahnung” (reinforcement). However, the word
“Bahnung” has more than one meaning, for processes of various
types are involved in this term. Thus writers have differentiated223
real and apparent “Bahnungen.” On account of this lack of clearness
in the meaning of the term “Bahnung,” I wish to discard its
use as it is not at all essential. We will speak simply of a summation
of excitation, for here it is simply a question of summation
of two excitations of the motor cells of the spinal cord.
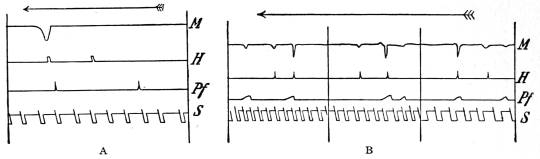
Fig. 56.
Summation of two excitations in the rabbit. The one proceeds from the paw, the other from
the motor sphere of the cerebral cortex. S—Time in seconds. Pf—Stimulation of the paw.
H—Stimulation of the motor sphere. M—Contractions of the abductor pollicis. (After
Exner.)
Fröhlich has shown that summation of two excitations upon a
motor cell of the anterior horn coming by way of different paths
is more readily obtained when the stimuli are somewhat strong,
or when the duration of the excitation processes in the ganglion
cells are somewhat prolonged by fatigue.
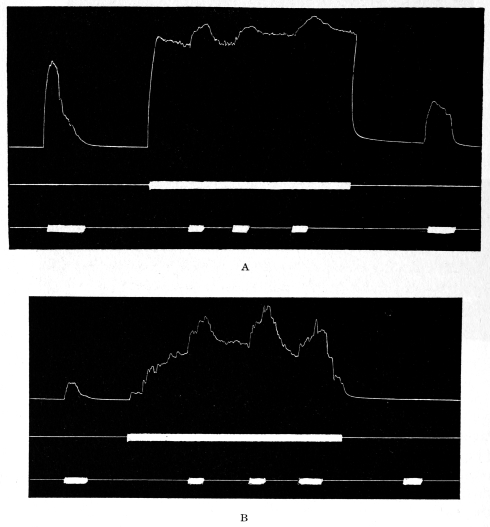
Fig. 57.
Summation of two excitations in the spinal cord produced by stimulation of the ninth and
tenth posterior root. Lower line indicates faradic stimulation of the tenth, upper line
of the ninth root.
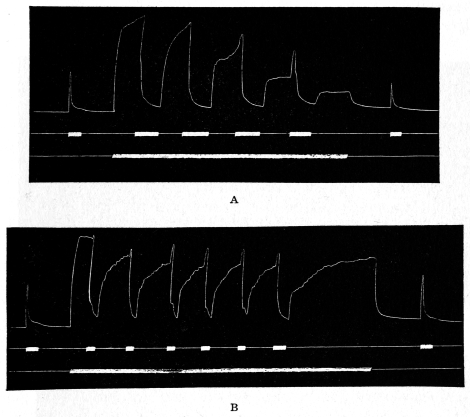
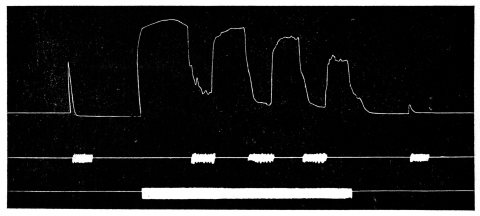
On the other hand, the conditions for the production of inhibition
are favored when the intensity of the series of stimuli is weak.
Here it is a question of the development of a relative refractory
period for the weak stimuli by increase in their frequency. A
relative fatigue of the motor ganglion cells for weak stimuli
rapidly occurs, and there develops a state of equilibrium beneath
that of the threshold of perceptible effect throughout the continuation
of stimulation. Vészi succeeded in isolating these types
of summation and inhibition in the spinal cord. His method consisted
in cutting the posterior roots of the spinal cord of the frog
and stimulating faradically the central ends, and at the same time
graphically recording the response of the gastrocnemius muscle.
Upon faradic stimulation of the ninth posterior root, one obtains
tetanic reflex contraction of this muscle. When the tenth posterior224
root is then stimulated, tetanus is also produced but of somewhat
shorter duration. If, while obtaining tetanus reflexly by
stimulation of the ninth root, a faradic current of short duration
and not too weak is applied to the tenth root, then a summation
of excitation occurs, an increase in the reflex contraction.
(Figure 57, A and B.) When, on the other hand, the tenth root225
is stimulated with weak shocks, one can obtain an increase of the
tetanus of short duration followed by inhibition. Here, as the
result of interference, we have an instance of inhibition with primary
tetanus. (Figure 58.) When the tenth root is stimulated
with very weak shocks, inhibition of the tetanus produced simultaneously
from the ninth root occurs without primary summation.
(Figure 59.) The fact that two series of stimuli, both of which
produce dissimilative excitation, bring about an inhibition by
their combined action, is sufficient to show the untenability of
the Gaskell-Hering hypothesis, that inhibitory processes result
from assimilatory excitation. It would be impossible to understand
how two dissimilatory exciting stimuli, by their simultaneous
action, could bring about assimilatory excitation. When226
the eighth or the seventh root is stimulated with stronger faradic
shocks during the time when tetanus is produced reflexly by
faradic stimulation of the ninth, an inhibition is practically always
obtained. Indeed, faradic currents that are so weak as to be far
below the threshold of perceptible response bring about when
applied to the seventh or eighth root a decided inhibition of the
tetanus, brought about by simultaneous stimulation of the ninth
root. The inhibitory effect of weak sub-threshold excitations
are here particularly apparent. This inhibition resulting from
excitation far below that of the threshold of perceptible response
is a common occurrence in the functional activities of the central
nervous system. In various parts of the nervous system, the
excitation in its conduction is weakened when passing through
intervening ganglion stations so that it has undergone a strong
decrement before reaching the responding structure, where an
inhibitory effect may be manifested. In this connection it is of
interest that the reciprocal “antagonistic reflexes” discovered by
Sherrington,197 who recognized their importance in the functional
processes of the nervous system, can be explained, as Fröhlich
showed, upon this principle of inhibition resulting from weakened
excitation. On the basis of numerous investigations in the227
Göttingen laboratory as well as that of Bonn198 we have come to
look upon the reflex arc in the spinal cord as consisting of the
following elements: a neurone in the spinal ganglion, a neurone
in the posterior horn and a motor neurone in the anterior horn.
This is the most direct route between the point of stimulation
and that of the responding organ of a unilateral reflex. (Figure 60.)
It is known that the excitation becomes weaker in passing
from the entrance of the excitation into the spinal cord to the
motor elements of a lower level on the same side or to those on
the opposite side. In order to obtain a response a stronger stimulus228
is necessary. Here the weakening of the excitation as well as
the prolongation of the reaction time is brought about by the
introduction of intercalated neurones. The reflex arc contains
more stations. (Figure 61.) If we accept the most plausible
assumption that the central connection of antagonistic muscles
possesses like relations, then the effects discovered by Sherrington
are self-explanatory. In this case stimulation of the sensory
path, which brings about a strong reflex excitation of the motor
neurons of the anterior horns controlling a muscle, at the same
time stimulates the antagonistic muscle with sub-threshold stimuli.
The result of this as shown by the experiments of Vészi is not a
motor response of the antagonists, but an inhibition if the motor
neurons of the antagonists are at the time in a state of excitation.
It is, therefore, understandable that reflex excitation of a229
muscle under normal conditions of irritability has an inhibitory
effect on its antagonist.
Fig. 60.
Scheme of the simplest unilateral reflex arc of the spinal cord.
Fig. 61.
Scheme of the simplest reflex arc from one to the other side, and
from a higher to a lower level.
Finally, I wish to conclude this discussion on the origin of
central inhibition and its dependence upon the strength of the
stimulus by referring to a point which apparently is contradictory.
We have already met with the fact that series of stimuli
by their interference in the nervous system may have different
effects depending upon their intensity; if this is strong, we obtain
summation of excitation, if weak an inhibition. The question
may be asked, how is it possible that a weak stimulus can have
a different effect when it is believed that the nerve as an isobolic
system responds to intensities of all gradations to the same extent,
namely, with maximum excitation? If the “all or none law” is
applicable, then the same intensity of excitation is always carried
to the centers and yet we see that various kinds of responses
follow various intensities of stimulation. Here, indeed, is a
difficulty which has not as yet been explained. Naturally between
the two facts there can be no contradiction. But the question
arises, how are we to bring them into harmony? Two entirely
different possibilities present themselves. If the various intensities
of stimulation always bring about excitation of the same
strength and we see in spite of this that various intensities of
stimulation produce various kinds of effects, then we must think
of the possibility that various intensities of stimulation bring
about some other effect than that of variations in intensity
in the course of the wave of excitation. In this connection
variations in the time involved must be taken into consideration.
One might think that strong stimuli may develop a longer wave
of excitation than such of weak intensity. Gotch199 tested these
questions experimentally with completely negative results. A single
strong stimulus does not result in an excitation differing in its
course from that of a weak stimulus. But there is another possibility
that requires testing. This was brought to light by the
investigation of Thörner200 on the fatigue of the nerve. His investigations230
showed that in a normal nerve in air the first typical
beginning of fatigue resulting from faradic stimulation can be
demonstrated in the characteristic summation of excitations.
This is shown by the nerve after fifteen minutes of stimulation
with faradic shocks applied for short intervals. The irritability,
when tested with single induction shocks, is at the same time
reduced. Thereby the amount of fatigue of the nerve, that is,
the amount of the reduction of irritability, is dependent upon the
strength and frequency of stimulation producing fatigue. When
the nerve is stimulated with weak faradic shocks of a slow rate
of frequency, there is a slight or a complete absence of the reduction
of irritability. On the other hand, if the nerve is fatigued
with strong faradic shocks of great frequency, the irritability
falls very considerably. This shows that when the nerve is stimulated
for a longer time, even under conditions favorable to the
supply of oxygen, a diminution of irritability occurs and with it
naturally an actual diminution of the wave of excitation, a diminution
the intensity of which becomes greater as the strength of
the stimulus increases. In other words, long-continued faradic
stimulation converts the nerve from a system isobolic in character
to that which is heterobolic in that the intensity of the excitation
which is conducted differs depending upon the intensity of the
stimulus. We have found other cases in the investigation of the
nervous system in which, as in fatigue, an isobolic is converted
into a heterobolic system. Vészi201 has shown that the centers of
the strychninized frog, which are isobolic in character, when
fatigued by weak faradic stimuli can be brought to react again
when the faradic stimulation is increased. According to this and
other experiments of a like nature, it is beyond doubt that an isobolic
system during the refractory period may assume a heterobolic
character, and only after completion of the refractory period
and entire recovery of the equilibrium of metabolism does the isobolic
character return. This permits us to understand the characteristic
properties of an isobolic system more accurately and precisely
than has thus far been possible. The “all or none law”231
with its associated properties, such as the conductivity without
decrement and the incapability of summating excitations, have
in a system of this character only relative validity. They are
realized only in the state of an equilibrium of metabolism. Only
when the stimuli follow each other at intervals greater than the
duration of the refractory period is there a response of equal
extent to stimuli of all intensities which are above the threshold.
During the refractory period and consequently in fatigue,
asphyxia, cooling and narcosis, etc., in short, in all states in which
the refractory period is prolonged this system loses its isobolic
properties and becomes heterobolic. In order that there may not
be a misunderstanding, we will consider more in detail the capability
in this state of summation of excitations. When we refer
to a summation of excitation of such a system under the influence
of one of these factors, we, of course, at no time mean an increase
of response beyond that of the degree of excitation which exists
in an isobolic system in a normal state consequent upon the application
of a single stimulus, for this degree of excitation is maximal.
We refer rather to a summation which has become reduced
as a result of fatigue.
On the basis of these facts it is readily understood when a level
of equilibrium of lower intensity has been reached that excitation
produced by weak faradic stimulation must have weaker
effects than when strong stimuli are applied, for when the system
assumes a heterobolic type as the result of relative fatigue weak
stimuli bring about weak, and strong, stronger excitation. Consequently,
during interference induced by a second series of excitations,
in the first case we have the conditions favorable for inhibition,
in the second for those of summation. If we also assume
that this characteristic alteration of the isobolic character of the
elementary nerve fibers which has been shown to occur in fatigue,
as seen when continued faradic stimulation is employed, develops
immediately after the beginning of stimulation then we can
readily understand the various kinds of effects produced by interference
observed in the reflex response following weak and strong
faradic stimulation to the different nerves in spite of the fact that
the nerve in the state of rest is a system isobolic in type. Experimental232
evidence, therefore, must be brought forward to show
that faradic stimulation of short duration produces the above-mentioned
alteration in the character of the system. Thörner in
his experiments on the nerve stimulated it faradically at least four
minutes and always found after this that excitation was reduced.
After shorter intervals of stimulation Thörner made no test of
the state of excitation. It is, however, highly probable that a
reduction of excitation is much more quickly reached. Indeed,
we are unavoidably compelled to accept the assumption that even
after the first single stimulus of the faradic current, alterations
of a slight degree are present which, after repeated stimulation,
become constantly greater and give to the system a heterobolic
character. As a result of fatigue, as we have already seen, the
refractory period becomes more and more prolonged. As the
individual shocks in faradic stimulation follow each other at
regular intervals, a necessary consequence is that the shocks are
operative before the refractory period has completely disappeared,
otherwise Thörner could not have obtained fatigue produced
by continued stimulation. The intervals of the individual
shocks must be somewhat shorter than the duration of the refractory
period, even in fatigue of a very slight degree. It is very
interesting in this connection that Thörner invariably obtained
positive evidences of fatigue by the application of stimuli at the
rate of 10–12 per second. When the number of stimuli per second
was less than this the above-mentioned result was not always
obtained. From this we can easily estimate the refractory period
of the nerve, which is present after reaching a state of equilibrium
under certain conditions. If we assume ten stimuli per
second to be the number required to produce slight fatigue when
stimulation is prolonged, we can conclude that the refractory
period in this state is somewhat longer than one tenth of a second.
Even though Gotch in his investigations already cited placed the
refractory period of the normal nerve at about .005 second, this
statement is in no way contradictory to the figure which we have
just given. Gotch measured simply the duration of the absolute
refractory period of the normal nerve, in other words, the duration
of the period in which no excitation at all could be brought233
about. On the contrary, my estimate, based upon the investigations
of Thörner, refers to the total refractory period of the
nerve, that is, to the point of complete recovery of the equilibrium
of metabolism and of the specific irritability. Experimental proof
of this assumption is already under way.
I have endeavored to show the elementary principles at the
basis of these extremely varied interference effects and to make
a few generalizations concerning the complicated conditions here
concerned. It has been shown that a great number of interference
effects possess characteristics in common if one takes into
consideration the process occurring in the course of a single
excitation. The altered state which exists in living substance
until the complete disappearance of excitation is the basis upon
which to explain the altered effects produced by a second stimulus.
This state alters during the whole course of the first stimulus
until the original equilibrium of the metabolism of rest is, by self-regulation,
again reached. It is, therefore, self-evident that the
second stimulus must have different effects depending upon the
momentary state of the living system at the time of its application.
The state of the system differs depending on the length
of the interval in which the second stimulation follows the first.
The most important factor is the phase of the excitation period
and the reduction of irritability. The second important factor
is the intensity of the second stimulus; the relation of the two
with each other determines the response. But the specific properties
of the given systems must also be taken into consideration.
It is important to know if the living system possesses isobolic
properties, that is, every intensity of stimulation produces a
maximal liberation of energy, or if it possesses a heterobolic
character, that is, stimuli of different strength bring about the
liberation of different amounts of energy. It is further important
to know the rapidity of reaction, whether the system rapidly
or slowly fatigues. In all cases it depends whether the second
stimulus produces a perceptible excitation or whether it occurs
in the refractory period and produces no perceptible effect. Upon
these factors depend the results of the interference of two
rhythmic series of stimuli, whether a summation or inhibition of234
excitation takes place. Here is the key to the understanding of
the great variety of interference effects. By determination of
these various factors in a given case and their sequence, we can
anticipate the nature of the interference which will follow. The
complex actions brought about by the various factors, which we
cannot at first clearly understand, can be at once interpreted as
soon as we convert them into their elements.
235
CHAPTER IX
THE PROCESSES OF DEPRESSION
Contents: Necessity of cellular physiological analysis of toxic depressions
by pharmacology. Apparent variety of processes of depression.
Depression of oxydative disintegration as the most extended principle
in the processes of depression. Asphyxiation, fatigue, heat depression,
as a consequence of restriction of oxydative disintegration.
Narcosis. Theories of narcosis. The alteration of specific irritability
and conductivity in narcosis. Depression of oxydative processes in
narcosis. Asphyxiation of living substance when oxygen is present
during narcosis. Persistence of anoxydative disintegration in narcosis.
Increase of the same by stimuli. Depression by narcosis as
a form of acute asphyxiation. Hypothesis on the mechanism of
depression of oxygen exchange by narcotics. Possibility of combining
the facts with the observations of Meyer and Overton.
The processes of excitation of all the effects of stimulation are
those which have invariably claimed place in the interest of physiologists.
The study of the processes of depression, on the other
hand, has remained more or less in the background. This is
readily understood when it is considered how much more apparent
the processes of excitation are than those of depression. Nevertheless,
these latter possess no less importance for the course
of vital phenomena than those of excitation. Without depression
no excitation can take place in the vital activity of the organism,
for, as we have seen, every excitation is secondarily followed
by a refractory period. To this must be added the great
number of primary depressions, directly brought about by the
most varied stimuli, such as cold, want of oxygen, poisons, etc.,
without the presence of a preceding excitation. Thus it is essential
that the processes of depression should be studied with no
less interest than those of excitation, and it is much to be desired
that the former should receive a more detailed analysis than has
up to now been the case. Even as it is, extensive material has236
been obtained for the analysis of this group of reactions. With
the closer study of the process of excitation the facts in connection
with the refractory period and fatigue make it necessary
that the processes of depression be taken into consideration.
Toxicology and pharmacology likewise furnish innumerable
effects of depression produced by poisons and drugs. Unfortunately
the investigation of these reactions has been in the main
purely superficial. This arises from the recency of the development
of these sciences. Even later than physiology they are
only now beginning to extend their investigations, directed up to
the present to the grosser organic reactions, to the cellular analysis
of the effects of poisons. How rarely we find instances in which
the effect of some drug is studied at the point of attack and systematically
followed to the specific cell form, and its primary
excitating or depressing effect on this or that constituent process
of the metabolic activities ascertained. And how great, on the
other hand, is the number of “medicines” making their appearance
each year in pharmacology of which nothing further is
known than a few secondary effects on the action of the heart,
the blood pressure, the secretion and excretion and on some other
outwardly perceptible organic actions! This deplorable condition
of present-day pharmacology must be ascribed to the regrettable
circumstances that pharmacological research is only in a very
small degree the result of careful investigations, carried out by
biologically and chemically trained pharmacologists, but is for
the most part undertaken at the instigation of chemical manufacturers.
This eager haste to obtain superficially practical
results has lessened in great degree the interest in the close and
painstaking theoretical analysis of reaction to poisons. Thus it
happens that, in spite of the numberless examples of the depressing
effects of poisons discovered by pharmacologists, it is only
in rare instances that the physical nature of these processes is
more closely studied. Therefore, investigation in pharmacology
and toxicology in so far as they are carried out in a purely
scientific spirit and not influenced by the desire for merely superficial
results, may find here a wide field of research work, rich
in future promise. It is from such investigation that we may237
expect an abundance of material for the closer analysis of the
processes of depression. For the present, however, we must
restrict ourselves to the consideration of some individual cases
which have been studied somewhat more in detail by physiologists.
Simple reflection shows the possibility that depression, that is,
the retardation of the normal vital processes, can be brought
about in various ways. As on the one hand the normal metabolism
of rest is composed of very numerous chemical constituent
processes, and on the other hand the closest interdependence
exists between these individual constituent processes, it follows
that every factor which increases or retards even one of these
must secondarily influence the course of the entire activity.
Hence a wide range of possibilities exists for the processes of
depression. As the complicated works of a clock can, by the
stopping of a single moving part, be brought to a standstill, so in
like manner the metabolic activity can be depressed by very
different constituent members. In spite of this we have every
reason to assume that the greater number of all processes of
depression result from the primary effect of one or a few constituent
members. A primary simultaneous depression of all or
at least of numerous constituent processes of the entire metabolism
may only be assumed as possible, resulting from decrease
of temperature within certain limits. But even in the case of
“cold depression” it is not probable, owing to the great effect of
every alteration in the relations of masses in the cell, that depression
is solely the manifestation of a uniform retardation of all
individual constituent metabolic processes. If, therefore, the
greater part of the processes of depression are brought about by
the primary effects of an individual constituent process, then the
possibility must be admitted that any component of the chain can
by the means of some specific external influence form the starting
point for a depression. The number of the various kinds of
processes of depression would be, therefore, enormous. The
knowledge obtained up to the present shows, however, that this
variety is not quite as great as the above facts might lead one to
expect. Even though future investigation will certainly not do238
away with the assumption of the existence of the most manifold
physical types of depression, the analysis of a few processes which
have been studied up to now demonstrates the singular fact that
a number of these which are brought about by quite different
external factors, are based on an absolute uniformity of their
mechanism. As we have previously seen, a certain constituent
of the metabolic chain can be excitated primarily by very different
kinds of stimuli. In like manner there exists in metabolic activity
a certain point of predilection for different kinds of stimuli, from
which their depressing effects proceed. Here the highly interesting
fact is shown that this point of predilection, which represents
that of the most frequent attack, is the same for excitating as for
depressing stimuli. These are the oxydative processes. As our
knowledge of the reactions to stimuli in anaërobic organisms is
still almost nil it is not possible at present to ascertain which
component in the metabolism of these organisms, adapted to life
without oxygen, plays an analogous rôle to that of the oxydative
in aërobic systems. Our investigations must, therefore, be restricted
to the world of aërobic organisms. Here we have seen
that the different stimuli which produce an excitating effect invariably
increase the oxydative disintegration of the living system
and we now find that these constituent processes of metabolism
likewise form a point from which depressing responses to stimuli
very readily proceed.
The prototype of this group of processes of depression in
which this is manifested in a most striking manner, is that of a
simple asphyxiation by the withdrawal of the oxygen supply
from the exterior. If the supply of oxygen is withheld from an
aërobic organism, oxydative disintegration is gradually found to
be more and more decreased and further breaking down takes
place anoxydatively, as oxydative decomposition forms the chief
source of energy production, and energy production consequently
undergoes a gradual decrease. Excitating stimuli, therefore, meet
with less response than when a sufficient supply of oxygen is
present, that is, irritability is diminished. As a result of this
decrease, a corresponding decrement in the extension of excitation
takes place, which, in turn, is likewise manifested by the239
restriction of the perceptible response to stimulation. In the
same degree in which oxydative disintegration becomes less,
anoxydative breaking down products are accumulated. The
accumulation of these products likewise plays a part in the production
of depression and increases the decrement in the conduction
of excitation. The decrease of energy production by
decline of the oxydative decomposition, as well as the accumulation
of anoxydative breaking down products, therefore, similarly
reduce irritability; that is, their effect is depressing. This
whole series of processes, which we have previously considered
in detail, takes place on the withdrawal of oxygen and leads to
the depression of asphyxiation. It can readily be observed in
the most varied kinds of aërobic organisms in rhizopods and
infusoria, in plant and ganglion cells, but finds its most complete
demonstration in the nerves. Here these processes can be easily
produced with any rapidity desired, accordingly as a relative or
absolute want of oxygen is brought about. These same typical
results are likewise shown in numerous processes in which the
external conditions are quite different in nature.
We have previously become acquainted with such a case and
studied it in detail. This is the state of fatigue. Fatigue is a
typical state of depression, that is, a state in which the vital process
is retarded and irritability in response to stimuli correspondingly
decreased. Fatigue is, however, as we have found,
the result of a relative deficiency of oxygen. The amount of
oxygen at disposal is not sufficient to allow of disintegration,
increased by constant functional activity oxydatively taking place,
to develop to its full extent. In consequence the previously cited
sequence of processes takes place. A “depression of activity” is
produced. Fatigue is true asphyxiation and it is here evident
that depression proceeds from the same constituent processes of
metabolism as excitation, brought about by a single stimulus.
Excitation produced by constant stimuli gradually merges into
depression as the amount of oxygen at disposal, even if augmented
in the intact organism by the increased blood supply, for instance,
is still insufficient to meet the demand made by the increased
oxygen consumption as a result of continuous functional activity.
240
A further very interesting example of depression produced
by oxygen deficiency is furnished by heat depression. It has
long been known that with increasing temperature the vital manifestations
of all poikilothermic organisms at first undergo a
heightening of their intensity. If, however, after a maximum
is reached, the temperature is still further increased a sudden
depression sets in. The increase in the rapidity of the vital process
as a result of increased temperature is readily understood
when based on the well-known law discovered by van’t Hoff.
Numerous investigations on the rapidity of the course of special
vital manifestations, as, for instance the growth of the eggs of
the frog and sea urchin, the assimilation of carbon dioxide in
green plant cells, the number of vacuole pulsations in the infusoria
cells, the frequency of the heart rate of the frog and of the
mammal, etc., have shown that their increase does in fact follow
the van’t Hoff law, being doubled or tripled in amount with every
increase of ten degrees of temperature. The genesis of depression
produced by heat, developed in different organisms at various
heights of temperature, requires a closer analysis. This depression
takes place at temperatures below that in which coagulation
of proteins occurs. Therefore, under certain conditions, with
which we shall presently become acquainted, it is capable of being
recovered from, whereas in higher temperatures, in which albumen
coagulates, vital activity is permanently obliterated. Depression
produced by heat is, therefore, in itself not a necrobiotic
process, which, as such, must necessarily lead to death. But
rather like fatigue it must be looked upon as an asphyxiation
process. Its relations to oxygen exchange have been chiefly
demonstrated by Winterstein202 by his investigations on the central
nervous system of frogs and on medusæ. He found that when
placed in a heated chamber in a temperature of 32–40° the
activity and reflex excitability of the frog are at first augmented.
Within the lapse of a short time this increase has become so
great that the slightest touch produces tetanic contractions, similar241
to those characteristic of strychnine poisoning. Very soon,
however, this state of high excitation is followed by one of depression,
in which no response to stimuli can be obtained. The
animal remains entirely motionless in any position in which it is
placed, in the same manner as a frog whose nerve centers have
been completely exhausted by strenuous activity. On the basis of
our knowledge of the rôle played by the deficiency of oxygen in
the bringing about of exhaustion the thought arose, if in this heat
depression exhaustion might not likewise be the result of oxygen
deficiency. This assumption has been most strikingly confirmed
by the investigations of Winterstein. It has been demonstrated
that recovery of the animal in a state of heat depression cannot
be obtained by mere cooling, but is only brought about when at
the same time a renewed oxygen supply is provided. For instance,
a frog is depressed in the warm chamber and even when a
strychnine injection has been introduced, does not show the
slightest reaction to stimuli. In the warm water bath artificial
circulation is now applied in the previously described manner with
an oxygen-free saline solution at 30° C., so that the blood is displaced
and thus the renewed oxygen supply to the nervous centers
prevented. The animal can now be cooled and the warm saline
solution be replaced by a cooled one without the least recovery
taking place. If, however, blood of the ox with contained oxygen
is substituted for the oxygen-free saline solution, the frog shows
signs of recovery within a few minutes and after ten or fifteen
minutes responds as a result of the strychnine to the merest touch
with tetanic contractions of the whole body. By modifying these
methods of investigation to a certain extent Bondy203 has confirmed
these results to the fullest extent. Later Winterstein by quantitative
determinations of oxygen consumption on medusæ showed
that at 30–35° C., at which temperature heat depression sets in,
the consumption of oxygen shows an increase of about three and
a half times compared to that in a temperature of 11–12° C.
These facts show that we have in heat depression a process which,
as far as its genesis is concerned, is completely analogous to that242
of fatigue. In fatigue, a relative want of oxygen is produced by
the increased consumption following functional activity, in heat
depression by the increase of the entire metabolism producing a
corresponding increase of oxygen requirement. In both instances
we have an excitation produced by external stimuli which result
in an increase in the amount of oxygen required, and in both
instances the oxygen at disposal is not sufficient to permanently
meet the augmented demand. In both types, therefore, decomposition
must become more and more anoxydative and the well-known
series of processes is developed, which find their expression
in depression.
In another direction likewise heat depression is of special
interest, that is, in regard to the theory of nature of the processes
in the living substance. According to the van’t Hoff law
we may assume that every individual constituent metabolic process,
if we imagine it as isolated and taking place in a test tube,
undergoes in more or less the same degree as all others an increased
rapidity of reaction as a result of increased temperature.
At the same time, in living substance we find on the contrary
that the van’t Hoff law is only within certain narrow limits more
or less applicable to the sum total of all metabolic processes.
Beyond certain degrees of temperature no further increase of
the vital process takes place, instead a retardation occurs. The
analysis of depression produced by heat shows us in the clearest
and simplest manner the reason for this apparent deviation from
the general law of van’t Hoff. This reasoning is based on the fact
that the rapidity of reaction of a chemical process is not merely
dependent upon the temperature, but likewise upon the mass
relations of the reacting substances. In spite of the effect of the
temperature in increasing the rapidity of reactions, the process
undergoes retardation which extends to a complete cessation if
the supply of material necessary to its existence does not keep
pace with the increase produced by temperature. In the present
instance the amount of reserve supplies for the building up of
the disintegrating molecules exists in abundance, and it is merely
the available oxygen which is in relatively a very small quantity.
As soon, however, as metabolism in its entirety, or even merely in243
those parts in which oxygen is directly required, is increased by
whatever means, the oxydative processes would be the first to fail
and it must be from this point that the disturbance of the harmony
in the interacting of the individual metabolic processes proceeds.
This principle which we here see manifested in its simplest form
in the effect of temperature on oxygen exchange in the form of
a disturbance in the correlations of the individual constituent
processes based on an alteration of the mass relation and the
rapidity of reactions of individual members is, however, not
merely restricted to effects of temperature and the results quickly
following on a relative oxygen deficiency. It has, indeed, a much
more general significance for all manner of constituent metabolic
processes, for it is applicable to all nutrition and to all growth,
and forms one of the most important factors which influence the
process of development, that is, the gradual “metachronic” alterations
in metabolism to which all living systems are subjected as
long as life endures.
A very extensive group of depression processes is produced
by the action of chemical stimuli. Among these the processes
to which we apply the collective term of “narcosis” must claim
our special interest. As is well known, an enormous number of
substances of very different chemical nature, such as carbon
dioxide, alcohol, ether, chloroform, chloral hydrate, etc., exist,
which, possessing the property of producing cessation of the
vital activities in all living systems, after withdrawal of their
application, if it has not been too prolonged or intense, permit
a complete restoration to normal vitality. These are the general
narcotics. Besides these there are a series of substances which
have a depressing effect only upon certain forms of living substance,
and which we may, therefore, term special narcotics. As,
however, the particular nature of depression following the
application of chemical substances has hitherto been closely
studied only in a very few instances, we are not, at present, in a
position to sharply define the limitations of the conception of
narcosis, a conception which originally had hardly any further
meaning than the production of unconsciousness by chemical
means. In the following discussion, therefore, we shall deal244
merely with narcosis produced by the well-known general narcotics,
such as carbon dioxide, alcohol, ether, chloroform, etc.
From the time of the introduction of ether narcosis into medical
practice by Jackson and Morton in the year 1848 up to the present
day, the theory of this process has awakened the liveliest interest.
Many attempts have since been made to explain the physical
nature of this interesting process without, however, any generally
acknowledged theory of narcosis being established. I will refrain
from entering into these former theories in detail as they have
been exhaustively treated by Overton204 in his studies on narcosis.
In connection with our present observations, however, I will
more closely analyze the process itself, following the results of
investigations extending over more than ten years carried out by
my coworkers and myself. In these investigations it has been
found that narcosis belongs to this group of depressing processes.
A satisfactory theory of narcosis, however, and this I must explain
from the first, can even today not be arrived at. Such a
theory would require the ascertainment of all primary and secondary
alterations produced by the narcotic in the course of normal
vital activity. For this, however, a number of minute details
are still lacking. Nevertheless, the careful and detailed investigations
during the last ten years have acquainted us with a large
number of alterations, which, acting as conditioning factors for
the process of narcosis, must be taken into consideration, and
which to a certain extent give us an idea of the mechanism of this
process. They are equally interesting from a theoretical as well
as from a practical point of view. The presentation will become
more detailed as more of such conditioning factors are established
by the deeper penetrating of future analysis. I will deal here with
the facts found up to the present and then proceed to the deductions
which these furnish for the theory of narcosis.
In the first place narcosis is stamped as a typical process of
depression, being characterized by a decrease of irritability with
a corresponding decrement of the extent of excitation. The chief
feature of all narcotized systems is, that in slight narcosis excitating245
stimuli produce a greatly weakened excitation, and that
in deep narcosis no perceptible response is obtained. This can
readily be ascertained in the various forms of living substance.
According to the previous observations on the inseparable relations
between conduction of excitation and irritability, it is self-evident
that with decrease of irritability there must be a corresponding
decrease in the capability of the conduction of excitation
from the point of stimulation. This decrease in conductivity
must, therefore, be the greater the more irritability is reduced;
that is, the deeper the narcosis, the greater must be the
decrement undergone by the wave of excitation in its extension
from the point of stimulation. These facts can be observed in
the highest perfection in the nerve, and have, as we have seen,
been demonstrated by the investigations of Werigo, Dendrinos,
Noll, Boruttau and Fröhlich.205 Upon deeper analysis of this process
of depression, the next task for the investigator must be the
ascertainment of the special components of the metabolic activity,
which are depressed as a result of the narcotic.
As a consequence of the result of my investigations on fatigue,
the idea occurred to me to test if possibly oxygen exchange likewise
undergoes depression during narcosis. The spinal cord
centers of the frog, which had served me in ascertaining the rôle
played by oxygen in the bringing about of the depression of
activity, appeared likewise a favorable object for this investigation.
Indeed, the question if consumption of oxygen takes
place during narcosis, could be experimentally determined in
direct connection with the investigations on fatigue. This was
based on the following consideration. If an oxygen-free saline
solution is introduced into the aorta of a frog and in order to
increase the activity of the spinal cord centers to the maximum
the animal is poisoned with strychnine, after a very short time
complete exhaustion takes place as a result of oxygen deficiency.
This exhaustion can only be removed by the introduction of
oxygen. In this condition the oxygen requirement of the centers246
is enormously increased. If the centers are narcotized by adding
a narcotic to the oxygen-free circulating fluid in amounts which,
as experience has found, would produce complete loss of reaction
in the normal animal, for example, about 5 per cent. of alcohol,
it can then be tested if, in this state of narcosis, the centers are
capable of oxygen consumption. It is merely necessary to replace
the oxygen-free saline solution containing alcohol by blood
rich in oxygen, containing alcohol in an amount sufficient to continue
the narcosis, but supplying an abundance of oxygen. If,
after this artificial circulation has lasted for a sufficient period,
the blood is then displaced by an oxygen-free saline solution
containing alcohol, and then this, in turn, is replaced by an
oxygen- and alcohol-free saline solution, so that cessation of the
narcosis is now produced, it can be ascertained by the responses
of the animal if consumption of the oxygen, when at the disposal
of the centers during narcosis, has taken place or not. If
the former is the case, then on the cessation of narcosis reflex
contraction must occur in the same manner as in every strychninized
frog totally exhausted by oxygen deficiency and into
which a saline solution containing oxygen is reintroduced. If
during narcosis, on the other hand, oxygen has not been consumed
by the centers, depression must continue to be present
after cessation of narcosis. Testing the recovery of the animal
on the introduction of blood, rich in oxygen, serves as an indicator
for the vital activity and capability of recovery of the
centers. A great number of experiments based on this scheme
of investigation were undertaken at my request by Winterstein.206
These were carried out with alcohol, ether, chloroform and also
carbon dioxide. His experiments have shown in the most
uniform manner that, in spite of the requirement of oxygen by
the centers being increased to its highest extent, and notwithstanding
the most ample oxygen supply during narcosis, after
cessation of the same and the introduction of an oxygen-free
saline solution no trace of recovery occurred, whereas after a
supply of oxygen was introduced tetanic contractions reappeared247
at once. During narcosis, therefore, the centers, in spite of their
great requirement of oxygen, lose their capability of oxydative
splitting up and consumption of oxygen.
After the methods for asphyxiation of the nerve had been
worked out and perfected the wish arose likewise to carry out
for these structures an analogous series of experiments to that
employed for the centers and based on the same chain of reasoning.
These investigations have the advantage of essentially
simpler conditions. After having convinced myself by experiments,
that the results on the nerve were in complete conformity
with those on the spinal cord, at my suggestion Fröhlich207 repeated
and continued these experiments on a more extended scale.
A nerve was asphyxiated by the previously described method.
This is accomplished in the simplest manner by the opening
or closing of stop cocks in the apparatus I have employed
which permit of pure nitrogen, or nitrogen with ether, and finally
also oxygen with ether or pure oxygen being conducted at will
through the glass chamber. If the nerve was so far depressed
in pure nitrogen that conductivity became obliterated for about
two cm. of the asphyxiated stretch, it was then narcotized in
nitrogen. Following this oxygen with ether was supplied for a
time. Then the oxygen-ether mixture was displaced by one of
nitrogen and ether and finally by pure nitrogen. Even after a
prolonged period, a recovery in pure nitrogen never took place.
On the other hand, the nerve recovered at once, as soon as oxygen
without ether was introduced. The results of these investigations
are, therefore, completely in harmony with those undertaken by
Winterstein on the nervous centers. They were later likewise
entirely confirmed by similar experiments of Heaton.208 All these
investigations furnished the proof that in narcosis, living substance,
notwithstanding even the greatest oxygen deficiency, is
not capable of producing oxydation, neither can consumption of
oxygen take place, with which, after cessation of the narcosis,
oxydative splitting up can be carried out.
248
Recently Warburg209 has likewise found an oxydative depression
during narcosis in the eggs of the sea urchin and in the red corpuscles
of geese, and the same fact has lately been also demonstrated
by Joannovics und Pick210 for the oxydative activity of the
liver cells of the dog.
This fundamental establishment of the fact that narcosis prevents
oxydations in living substance is at once followed by the
further problem, in what manner do the disintegration processes
undergo alterations during narcosis? That they must be altered,
and this in the form of a reduced energy production, is clearly
shown by the decrease of irritability and the increase of the
decrement of the conduction of excitation. Both become the
greater the deeper the narcosis. The observations just discussed
render these facts at once self-evident. They follow as a simple
and necessary result of the elimination of the oxydative processes.
If these are suppressed further breaking down, if not
influenced by addition of other factors, proceeds anoxydatively.
The previously observed series of processes is developed, which
invariably take place when oxygen deficiency occurs and which
produce in the clearest form the results of asphyxiation on the
withdrawal of oxygen supply. If, therefore, the disintegration
processes are not influenced in some other manner during narcosis,
they must then take place in the same way as in the withdrawal
of the oxygen supply. The question, if this is actually the
case, can be experimentally decided by comparing, on the one
hand, the development of the course of asphyxiation during narcosis,
and on the other, the withdrawal of the oxygen supply.
We have carried out this comparison for the spinal cord centers
as well as for the medullated nerve. A prolonged series of experiments
have been made by Bondy211 with the apparatus constructed249
for this purpose by Baglioni.212 Two frogs under uniform conditions
of temperature were submitted to artificial circulation, the
one merely with an oxygen-free fluid, the other with the same,
but with the addition of 5 per cent. of alcohol. In order to render
the least trace of irritability perceptible, responsivity was increased
in both animals by the employment of strychnine. It
then appeared that, on the average, irritability was obliterated in
the narcotized frog in about the same time as in the animal
simply asphyxiated. These experiments were controlled by introducing
at their conclusion a saline solution containing oxygen
into both frogs and by ascertaining the degree of recovery. In
like manner Fröhlich213 has established the same fact for the nerve.
The period of asphyxiation for the nerve in a nitrogen-ether
mixture is approximately the same as in pure nitrogen. Analogous
experiments have been carried out in amœbæ by Ishikawa.214
Here also it has been shown that living substance becomes
asphyxiated in narcosis and can finally recover only when oxygen
is supplied. In more than a hundred experiments Ishikawa has,
however, obtained the uniform result that amœbæ asphyxiate
rather sooner in narcosis than in pure nitrogen. The most striking
experiments are those which Heaton215 has carried out on the
nerve. Using both sciatic nerves of the same frog, he passed
each one through a separate glass chamber, as previously described,
and laid the central stumps projecting from the chamber
over a pair of platinum electrodes, while the stretch within was
likewise placed on platinum electrodes. The muscles served as
indicator of the capability of conduction and irritability. The
alterations thereof were tested by the ascertainment of the
threshold of stimulation. The nerve in the one chamber was
then subjected to a pure nitrogen current, that in the other merely
to one of pure air with ether. In order to test the degree of250
asphyxiation the air-ether current in the latter chamber was
replaced from time to time by an ether-nitrogen current, and
then by one of pure nitrogen, so that the narcosis was interrupted
without the entrance of oxygen being possible in the mean time.
During this suspension of the narcosis, the nerve recovered each
time in nitrogen, its irritability again increasing and its capability
of conduction returning with every test. However, recovery
showed itself as less and less complete. Finally irritability
had sunk so low that the capability of conduction disappeared
entirely. At the end of the experiment as control, nitrogen
was displaced by air in the two chambers and in both nerves
recovery took place.
In both cases recovery could only be brought about by an introduction
of oxygen. From the sum of all these experiments it
results that during narcosis in air the nerve, even when a sufficiency
of oxygen is present, gradually asphyxiates and loses
its capability of conduction, and this in about the same length
of time as the other nerve in pure nitrogen. These investigations
furnish two important facts for the theory of narcosis.
First, that in narcosis living substance becomes asphyxiated notwithstanding
the presence of an ample oxygen supply, and
secondly, that asphyxiation occurs in the same time, or somewhat
more rapidly, in pure nitrogen under otherwise similar
conditions than without narcosis. In other words, it is shown
that the breaking down processes of metabolism continue in narcosis
as anoxydative disintegration. In narcosis, therefore,
asphyxiation takes place with approximately the same or a somewhat
greater rapidity than that in an oxygen-free medium.
The fact here established explains in the simplest manner the
often described observation that in the human being and in mammals
during prolonged anæsthesia typical products of insufficient
combustion, such as fatty acids, lactic acid and above all aceton,
in not inconsiderable quantities are eliminated, as the case may
be, by the urine or the respiratory air.216 If, as has been shown by251
the foregoing experiments, the processes of disintegration can
continue to anoxydatively take place during narcosis, the problem
arises, if this anoxydative breaking down can be further
increased by excitating stimuli. This question has been answered
likewise by means of experiments on the nerve made by Heaton.217
The two sciatic nerves of the same frog were drawn through
a double glass chamber of the form previously described so that
each nerve lay on an electrode and with the central stump protruding
out of the chamber hanging likewise over an electrode.
As in the former instances the muscle contraction of the shank
again served as indicator. Both nerves were then subjected to
the same current of nitrogen-ether. When, as a result of the
narcosis, their irritability has sunk to the level of “stromschleifen”
the central stump of the one nerve was continuously stimulated
with faradic shocks during a prolonged period, while the other
nerve remained at rest. Finally, by displacement of the current
of nitrogen-ether with one of pure nitrogen, cessation of narcosis
was brought about. It was then seen that the irritability of the
continuously stimulated nerve showed a much greater decrease
than that of the nonstimulated. The control made by introduction
of air demonstrated that both nerves recovered in an oxygen
supply. There can, therefore, be no doubt, by comparative experiments
we find, that during narcosis anoxydative disintegration
can be still further increased by the action of stimuli.
In view of this knowledge of the influence of narcotics on
oxygen exchange it may be considered as a firmly established
fact, that a process of depression is developed during narcosis,
which can be classified with the large group of depressions, resulting
from deficiency of oxygen. This is followed by the important
problem, is it possible to attribute the whole series of alterations,
produced by the narcotic, solely to this one factor? In other
words, is narcosis the result of acute suppression of the oxydative
processes?
If the individual symptoms which characterize narcosis are
investigated from this point of view, one must indeed confess
that they are all readily understood when regarded as the results252
of suppression of the oxydative processes. Indeed, the disappearance
of the perceptible vital activities, the decrease of
irritability, the restriction of the conduction of excitation, the continuance
of an anoxydative breaking down, the recovery on cessation
of narcosis, provided oxygen is present, etc., in short, all
the characteristics of narcosis so far known must be expected
and demanded if a suppression of the oxydative processes exists
during narcosis.
There is only one point which at the first glance would not
seem to agree entirely with the assumption. This is the fact that
depression sets in with a relatively greater rapidity in narcosis
than when the supply of oxygen is completely withdrawn.
Depression of the centers in the spinal cord, which begins in
about five to ten minutes after artificial circulation of an oxygen-free,
alcohol-containing, saline solution, is not brought about for
more than an hour when the same saline solution but without
alcohol is introduced. This difference is still more strikingly
apparent in the nerve. The same degree of depression, which is
produced in the nerve in a nitrogen-ether mixture within about
five minutes, is not reached in pure nitrogen without ether until
after the lapse of from two to four hours. In order to investigate
this relation somewhat more closely I have questioned if it
is possible for a living system, which has been narcotized to a
certain extent, to regain its irritability in a completely oxygen-free
medium, if cessation of the narcosis takes place after a
period essentially shorter than the time of asphyxiation of the
system under equal conditions. If the depression of narcosis is
founded exclusively on asphyxiation, it would be expected that
no recovery could occur. Experiments which I have made on
the spinal cord centers as well as on the peripheral nerves have,
however, demonstrated exactly the contrary. If a frog is
subjected to an artificial circulation of an oxygen-free saline
solution containing 5 per cent. of alcohol until reaction is lost,
being certain of this by the injection of a weak dose of strychnine,
and if now a cessation of the narcosis is brought about by
the transfusion of oxygen-free saline solution, the centers of the
animal recover completely within ten to fifteen minutes, as shown253
by typical strychnine tetanus. If a nerve is placed in a gas chamber
through which a mixture of nitrogen and ether is allowed to
flow until irritability is greatly decreased, and is then displaced
by pure nitrogen, irritability increases more or less completely
according to the time which has passed from the beginning of
asphyxiation. This investigation proves that living substance,
even after the deepest narcotic depression, may recover on cessation
of the narcosis, although in an entirely oxygen-free medium.
Fröhlich, Bondy and Heaton, by the methods of their experiments
above described, have proved this fact in a great number
of instances. On the other hand, Ishikawa could not observe
a pronounced recovery in amœbæ from narcosis in pure nitrogen.
But it is possible that here the difference is perhaps merely
quantitative.
What position should be taken in the face of these facts? Does
recovery of a deeply narcotized tissue in an oxygen-free medium
really make it difficult to suppose that narcosis is the result of
an acute suppression of the processes of oxydation? On closer
view, it will be found that this difficulty is merely apparent. In
reality it is quite possible to bring these facts into harmony with
the assumption that narcosis consists in a suppression of these
processes. If one proceeds from the supposition that living substance
possesses a certain, even though merely a small supply of
oxygen in its interior, then it is at once evident that a more or
less complete recovery of irritability from narcosis depression is
possible, even in an oxygen-free medium. It can take place at
the cost of the oxygen still present in the living substance and
which during the narcosis, on account of the suppression of the
oxydation processes, could not be consumed. If the presence of
a certain oxygen reserve in living substance is entirely set aside
and a different explanation sought for the primary continuance
of irritability after a complete withdrawal of the oxygen supply
from without, the great difference of time in the setting in of the
depression in narcosis and that of the complete elimination of the
oxygen supply from without would make it necessary to assume
the processes occurring in narcosis are entirely different in nature.
The explanation that narcosis is the result of suppression of the254
oxydative processes would indeed be out of the question in such
a view.
The assumption, however, that in a living system at the same
moment when oxygen is removed from the neighborhood, let us
say by a stream of nitrogen, no oxygen would be present and
that in consequence every oxydative process must cease, contains
so little probability that I have rejected it on various occasions.218
The way in which irritability is lost in asphyxiation of the nerve
likewise very clearly demonstrates the untenability of this view.
The recent investigations of Lodholz219 have shown that decrease of
irritability takes place after a sudden displacement of all oxygen
from the surrounding medium uniformly and gradually in the
form of a logarithmic curve. If at the moment of oxygen withdrawal
from the outer medium, metabolism became entirely
anoxydative, the curve of irritability must under all circumstances
show a sudden steep decline at this point, and subsequent to this
a further slower decrease. For, as the oxydative processes constitute
by far the chief part in the energy production of living
substance, the production of energy, and with this irritability,
would undergo considerable loss at the same moment in which
oxydative was replaced by anoxydative disintegration. The curve
of decrease of irritability during the transition period from
oxygen supply to oxygen withdrawal shows, on the contrary, a
completely uniform course and it is not until later that a very
slow decline takes place, which only after a prolonged time assumes
increasing rapidity. But the assumption that at the
moment when the supply of oxygen ceases, anoxydative breaking
down could acquire such enormous dimensions that it furnishes
just exactly the same amount of energy as was before supplied
oxydatively, is a view which no one will seriously entertain. In
connection with this I wish to call attention to the experiments
of Fröhlich220 in which he compared the time required for asphyxiation
to take place in the nerves, when, on the one hand, the frogs
had been kept several days previous to the experiment in temperature255
of 14–40° C., and on the other, in one merely a few degrees
above zero. He found that the nerves of the cooled frogs required
on an average twice or three times as long for their irritability
to sink to the same degree as those of the heated frog,
although during the experiment the same temperature was present
in both. It was also shown that the asphyxiation period was prolonged
up to a certain limit, depending upon the length of time the
animals were kept at a low temperature. It would seem to me
that these facts admit of no other explanation than that in a low
temperature a greater amount of oxygen is stored in the nerve
than in high temperatures. From the standpoint that from the
moment of withdrawal of oxygen from without, disintegration
likewise takes place exclusively anoxydatively, these facts would
be completely incomprehensible. When, however, the assumption
is made, and this would appear to me as inevitable, that living
substance contains in itself a certain even though a very slight
quantity of oxygen, which in low temperature is greater, in a
high temperature less, the recovery from narcosis, when oxygen
is withheld, is not at all surprising. The comparatively rapid
setting in of depression in narcosis finds a simple explanation in
the violent manner in which the oxydative breaking down, notwithstanding
the presence of oxygen, is suddenly suppressed by
the flooding by the narcotic. Finally, this view receives unlooked-for
support by a group of facts which at the first glance would
appear to bear no relation whatever to the process of narcosis.
In a series of investigations on the mechanism of movement in
naked protoplasm,221 I have pointed out the rôle played by oxygen
in the genesis of the amœboid protoplasm movement. We can
distinguish two antagonistic phases in the movement of amœboid
cells, the expansion phase and the contraction phase. The first
consists in an increase, the latter in a diminution of the surface,
the mass remaining the same. The expansion phase is manifested256
in the stretching out of the pseudopods by a centrifugal outflowing
of the protoplasm into the surrounding medium, the contraction
phase by the indrawing of the pseudopods by the centripetal
inflowing of the protoplasm to the cell body. In total contraction,
such as occurs, for instance, in strong excitation following stimuli,
the cell body becomes ball shaped. In local contraction of the
long thread or net-shaped outstretched pseudopods of the sea
rhizopoda, the protoplasm of the retracting pseudopod forms balls
and spindles. Considered from a physical point of view the
expansion phase of amœboid movement is an expression of decrease,
the contraction phase an increase of the surface tension.
I have shown that the factor which under physiological conditions
decreases the surface pressure and thereby brings about the
expansion phase is the introduction of oxygen into the living
substance. With removal of oxygen the stretching out of the
pseudopods ceases. The cell gradually draws in all pseudopods
and assumes the shape of a ball. On the reintroduction of oxygen
the outflow of the pseudopods begins anew. This fact can be
observed in all amœboid cells. When, therefore, consumption of
oxygen and oxydative changes is suppressed during narcosis it is
to be expected that all naked protoplasm masses by being narcotized
lose their capability of assuming the expansion phase of
movement and contract into the shape of balls. Experimentation
confirms this deduction in the most striking manner. When
amœbæ are placed in a drop of water under the microscope in a
gas cell through which air and a little ether are allowed to flow,
the pseudopod formation of the amœbæ ceases within a few
minutes and they all assume the shape of a ball. (Figure 62.) In
asphyxiation in pure nitrogen, the changes in the amœbæ take
place in exactly the same manner with the exception that in this
case a longer period ensues according to the size and activity of
the animals. About 20 to 60 minutes elapse before depression
becomes complete. If larger sea rhizopoda are narcotized in the
same manner all pseudopods are more or less retracted and the
contained protoplasm flows centripetally and contracts in the
characteristic manner into balls and spindles. (Figure 63.) If
the narcosis is removed by displacing the ether by pure air, the257
stretching out of the pseudopods then begins anew, provided the
narcosis has not been too deep or too prolonged.
Fig. 62.
Amoeba limax. A—In normal state. B—Narcotized by ether.
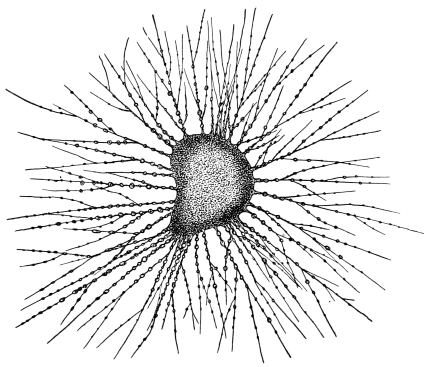
Fig. 63.
Rhizoplasma Kaiseri. Effect of chloroform.
In the face of all this evidence there can be indeed no further
barrier to the assumption that the symptoms in narcosis are a
result of a suppression of the oxydative processes. Nevertheless,
I would not at present venture to maintain that the entrance
of the narcotic into living substance produces no alterations whatever,
except just this oxydative suppression. For the present
it seems to me that the possibility is in no way precluded that258
the same process, which is expressed in the oxydative suppression,
is connected with other alterations in the living substance,
of which we are as yet ignorant. As far as the effects of larger
doses of narcotics are concerned, the assumption that other
alterations take place in the living substance can in any case
hardly be avoided. An application of larger quantities of narcotics
brings about destruction of the living system with great
rapidity. Here the alterations in the optical properties of the
cell are of such magnitude that the changes are directly perceptible
under the microscope. Binz222 has observed such alterations
in the nerve cell and looked upon them as coagulation. In unicellular
organisms these optical alterations can readily be followed.
If amœbæ, sea rhizopods or infusoria are narcotized
with stronger doses of ether or chloroform, the protoplasm becomes259
opaque and granulated, it appears darker than formerly
and in many cases displays a yellowish brown color in transmitted
light. Cells altered in this way no longer recover after
removal of the narcotic. These intense and rapidly appearing
alterations of protoplasm resulting from the application of
stronger doses of the narcotic can scarcely be explained as simply
the result of a mere decrease of the oxydative processes. They
would seem to consist rather, as suggested by Binz, as coagulation,
in an alteration of the state of certain components of living substance.
Whether these alterations are already present in a correspondingly
slight amount in those degrees of narcosis after which
complete recovery can take place and further whether in this
case they are in any way concerned in bringing about the individual
symptoms of the former, are questions the decision of
which must be left to future investigations. Höber223 indeed
makes such an alteration of the colloidal state of the lipoid the
basis of a theory of narcosis. But such assumptions are scarcely
more than speculations. This is one of the points in which our
present knowledge is lacking.
Even if we restrict ourselves to the actually established alterations
produced by the narcotic in living substance, new problems
present themselves, the investigation of which requires further
effort. Above all, the question arises as to the finer mechanism of
oxydative depression. In what manner does the narcotic molecule,
entering into the living substance, suppress the oxydative
processes? Here there are very different possibilities to be taken
into consideration and up to the present in our investigations of a
suppression of the oxydative processes resulting from narcosis,
we have stood on the firm ground of assured facts. However, the
discussion of the nature of this suppression leads us into the
domain of hypothesis. But without hypothesis there can be no
progress in knowledge. In all branches of scientific research,
working hypotheses are required for the obtainment of new facts.
On closer reflection, there are chiefly three possibilities, which,260
considered from the standpoint of our present knowledge of the
processes in living substance, offer an explanation of the oxydative
suppression as a result of narcosis.
One of these possibilities is, that the narcotic itself consumes
the oxygen which activates living substance and uses it for its
individual oxydation, so that the specific oxydable material of
living substance receives less oxygen from the oxygen carriers.
Based on a series of interesting experiments this view has been
recently maintained by Bürker.224 He observed that with the
electrolysis of acidulated water, to which a small per cent. of ether
was added, a much less amount of oxygen was at the anode than
in one used as means of control, containing acidulated water
without ether. The oxygen was replaced at the anode by
oxydation products of the ether, such as carbonic oxide, carbon
dioxide, acetate aldehyde and acetic acid. In experiments with
various narcotics he likewise found that the stronger the effect
produced by narcosis, the greater the oxygen amount required
for the oxydation taking place of electrolysis. Bürker applies
these results obtained for electrolysis to the processes in living
substance and takes the view that the narcotic seizes on the
active oxygen, and so withdraws it from the masses of living substance
possessing a great oxygen requirement. It cannot be
denied that this conception of the nature of certain narcotics
deserves careful investigation. It seems to me, however, that
before considering it in the light of a serious probability a grave
difficulty would first have to be removed. In living substance
the narcotic would occur under conditions essentially different
from those existing during the experiment in the voltameter.
In the former case there would be the struggle for oxygen
of the specific oxydable cell masses to be met with. Considering
the small amount of chemical activity of the greater number of
narcotics it would appear at least doubtful if in this battle for
supremacy the latter would achieve a victory. For some narcotics,
as, for instance, carbon dioxide, this method of a depression
of the oxydative processes would have no bearing whatever.261
This is rather to be looked for in the effects of oxydative suppression
of the aldehydes, which Warburg225 has recently observed and
investigated. Here, however, it is not a true narcosis which is
concerned.
A second possibility of a suppression of oxydation would be
the fixation of the molecules of the oxydable substances by chemical
or physical combinations in that they would lose their capability
of oxydative disintegration. Such a supposition would,
however, likewise contain but few elements of probability. As
has been shown, an anoxydative breaking down continues during
narcosis, which, and this we may assume with certainty, furnishes
very different products in great variety. These anoxydative disintegration
products, as recovery on the cessation of narcosis
shows, are removed during recovery by oxydation. If the effect
of the narcotic consisted in the prevention in spite of the presence
of oxygen of the oxydation by combination, it would be necessary
to assume that the narcotic was bound to a mass of completely
heterogeneous substances, a conclusion we should find
difficult to entertain.
If, however, depression of the oxydative processes is founded
neither on the seizure of oxygen by the narcotic nor the fixation
of oxydable substances by the former, there remains the possibility
that the narcotic suppresses the transmission of oxygen to
these points of consumption. We assume that the oxygen transmission
to those points where its consumption takes place is carried
out by special substances, the existence of which has been
established in the most varied vegetable and animal cell forms.
Unfortunately we only know these oxygen-carrying substances
by their effects. Of their chemical constitution we have no
knowledge, but we usually assume that the transmission of oxygen
occurs in the same manner as in catalytic processes. On
another occasion I have previously expressed the suggestion,226
that the narcotic suppresses oxydation by producing incapability
of the groups acting as oxygen carriers to carry out this function.262
If we assume that the substances possessing the character
of oxygen carriers, which activate the molecular oxygen and so
render it capable of attacking the oxydable substances, lose this
capability under the influence of narcotics, this supposition would
not only make all of the facts of suppression of oxygen exchange
in narcosis comprehensible, considered from one point, but likewise,
as careful investigation has shown, be in complete harmony
with all knowledge obtained up to the present of the process of
narcosis.
Here is the point where the interesting observations of Hans
Meyers227 and Overton228 on the relations of the depressing influence
of narcotics to their solubility of fat and water may be connected
with the facts of the suppression of oxydation. Meyer and Overton
have quite independently of each other made the same observation,
that the depressing effect of a narcotic is the greater, the
larger the coefficient of distribution between substances of a fatty
nature and water. Those narcotics produce the strongest effects
which are readily soluble in substances of a fatty nature, but not
easily so in water, that is, in which the coefficient distribution
between fat and water is very great. This law, which
has been demonstrated by Meyer and Overton for a large number
of narcotic processes, is in itself not a theory of narcosis, as
has been often erroneously assumed. It shows us, however,
an important condition, which must be considered in every
theory of narcosis. It demonstrates that it is the ease with
which transmission in the lipoid occurs which allows a substance
to develop narcotic effects. These facts would seem to indicate
that the lipoids of the cell are connected in some way or other
with the exchange of oxygen. If we assume that the oxygen
carriers, the chemical constitution of which is so far not known,
bear the character of lipoids and belong, say, to the generally
extended group of phosphatides, there results at once an apparent263
connection of the law established by Meyer and Overton with the
nature of narcosis.
The depressing effect of the narcotic would then consist in
producing incapability of the lipoids transmitting oxygen to act
as carriers of the same, and it is, therefore, self-evident that the
effect of the narcotic would be the stronger the more readily it
found entrance into the lipoids. It is perhaps not without interest
that in similar manner Mansfeld229 has attempted to establish a
connection between the facts which Meyer and Overton have
found and those ascertained by my coworkers and myself. He
expressed the view that the lipoids of the cells represent the
channels followed by the oxygen on its entrance, and that in consequence
of their accumulation in the lipoids, the narcotics bring
about asphyxiation by physically obstructing the transmission
of the oxygen from the outer medium through the surface layer
of the lipoid into the protoplasm. The divergence in our views
is not essential in their nature, and I attach the less importance
to them as we find ourselves here, as I must again emphasize,
on purely hypothetical ground.
In consideration of these observations we may perhaps establish
the following hypothesis of the effect of the oxydative suppression
of narcotics: The narcotics obstruct, either by absorption
or loose chemical combination the oxygen carriers of the cell and
render them incapable to activate the molecular oxygen. In consequence,
oxydation of the oxydable substances cannot take place
and disintegration occurs of an anoxydative form. The cell
asphyxiates.
In conclusion I wish to warn against erroneous assumption
that all oxydative depressions by chemical substances are narcosis
and that the mechanism is the same. It is true that a number
of chemical substances depress the processes of oxydation.
But the latter can be brought about in very varying ways. I
would like to mention the effect of oxydative depression of
aldehydes. To this Warburg230 has added hydrocyanic acid,264
arsenic acid, ammonia and substitution compounds of ammonia.
These substances do not follow the Meyer-Overton law of the
coefficient of distribution. We cannot consider them, therefore,
as narcotics. Future investigation will establish the existence of
a large number of substances belonging to this great group of
oxydation suppressing poisons, which are not narcotics. And
it is likewise certain that depressing substances will be found, the
depressing effects of which will not have their point of attack in
the oxygen exchange, but will be shown to exist in other constituents
of the metabolic chain. Our research in these fields, as
already said, is still in the first beginnings and its perspective
reaches into infinite space.
Return to transcriber’s notes
Spelling corrections:
possibilites → possibilities
deliminated → delimitated
equilibrum → equilibrium
fur → für
künstliche Immunisirungsprocesse → künstlichen Immunisierungsprozesse
methan → methane
aldehyd → aldehyde
Rüchenmarks → Rückenmarks
metronom → metronome
irrritability → irritability
tranverse → transverse
the the → the
Mittleilung → Mitteilung
whereever → wherever
oxdyative → oxydative
anoxdyative → anoxydative
Spelling inconsistencies:
ae/æ/e (inconsistent ligatures)
cannot/can not
cell-pathology/cell pathology (inconsistent hyphenation)
æthyl/ethyl
Return to transcriber’s notes
