
Title: Bacteria
Author: Sir George Newman
Release date: April 25, 2015 [eBook #48793]
Most recently updated: October 24, 2024
Language: English
Credits: Produced by Chris Curnow, Turgut Dincer and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive)
|
THE SCIENCE SERIES 1. The Study of Man.—By A. C. Haddon. Illustrated, 8o, $2.00. 2. The Groundwork of Science.—By St. George Mivart. 8o, $1.75. 3. Rivers of North America.—By Israel C. Russell. Illustrated, 8o, $2. 00. 4. Earth Sculpture.—By James Geikie. Illustrated, 8o, $2.00. 5. Volcanoes.—By T. G. Bonney. Illustrated, 8o, $2.00. 6. Bacteria.—By George Newman. Illustrated, 8o, $ ? G. P. PUTNAM'S SONS, New York and London |
EDITED BY |
BACTERIA
ESPECIALLY AS THEY ARE RELATED
TO THE ECONOMY OF NATURE
TO INDUSTRIAL PROCESSES
AND TO THE PUBLIC HEALTH
BY
GEORGE NEWMAN
M.D., F.R.S. (Edin.), D.P.H. (Camb.), etc.
DEMONSTRATOR OF BACTERIOLOGY IN KING'S COLLEGE, LONDON
ILLUSTRATED
NEW YORK
G. P. PUTNAM'S SONS
LONDON
JOHN MURRAY
1899
Copyright, 1899
BY
G. P. PUTNAM'S SONS
The Knickerbocker Press, New York
The present volume is not a record of original work, nor is it a text-book for the laboratory. Theoretical and practical text-books of Bacteriology plentifully exist both in England and America. There are two large works widely used, one by Professor Crookshank, entitled Bacteriology and Infective Diseases, the other by Dr. Sternberg, A Manual of Bacteriology. There are also, in English, a number of smaller works by Abbott, Ball, Hewlett, Klein, Macfarland, Muir and Ritchie, and Sims Woodhead. This book is of a less technical nature. It is an attempt, in response to the editor of the series, to set forth a popular scientific statement of our present knowledge of bacteria. Popular science is a somewhat dangerous quantity with which to deal. On the one hand it may become too popular, on the other too technical. It is difficult to escape the Scylla and Charybdis in such a voyage.
I am much indebted to Professor Crookshank, who, in reading the manuscript, has helped me by many valuable criticisms. My thanks are also due to Sir C. T. D. Acland, Bart., for many kind suggestions, and to Mr. E. J. Spitta, M.R.C.S., who has been good enough to take a number of excellent photo-micrographs for me. Some other illustrations have been derived from the Atlas of Bacteriology, brought out jointly by Messrs. Slater and Spitta. For these also I am glad to have an opportunity of expressing my thanks. It should be understood that the outline drawings are only of a diagrammatic nature.
GEORGE NEWMAN.
London, 1899.
| PAGE | |
Introduction | ix |
| CHAPTER I | |
The Biology of Bacteria | 1 |
| CHAPTER II | |
Bacteria in Water | 37 |
| CHAPTER III | |
Bacteria in the Air | 96 |
| CHAPTER IV | |
Bacteria and Fermentation | 111 |
| CHAPTER V | |
Bacteria in the Soil | 137 |
| CHAPTER VI | |
Bacteria in Milk, Milk Products, and Other Foods | 178 |
| CHAPTER VII | |
The Question of Immunity and Antitoxins | 240 |
| CHAPTER VIII | |
Bacteria and Disease | 264 |
| CHAPTER IX | |
Disinfection | 322 |
Appendix | 337 |
[Illustrations starred (*) are reproduced by permission of the Scientific Press from Drs. Spitta and Slater's Atlas of Bacteriology.]
| PAGE | |
Various Forms of Bacteria | 9 |
Sarcina | 10 |
Normal and Pleomorphic Forms of Tubercle | 13 |
Bacilli, Showing Flagella | 15 |
Various Forms of Spore Formation and Flagella | 18 |
Potato in a Roux Tube Prepared for Cultivation | 22 |
Staphylococcus Pyogenes Aureus Incubator | to face 22 |
Culture Media Ready for Inoculation | 23 |
Inoculating Needles | 24 |
Pasteur's Large Incubator for Cultivation at Room Temperature | to face 24 |
Method of Producing Hydrogen by Kipp's Apparatus for Cultivation of Anaërobes | 27 |
Anaërobic Culture | 28 |
Koch's Steam Steriliser | 31 |
Levelling Apparatus for Koch's Plate | 40 |
Moist Chamber in which Koch's Plates are Incubated | 41 |
Hot-Air Steriliser | 42 |
The Hanging Drop | 44 |
Drying Stage for Fixing Films | 45 |
Types of Liquefaction of Gelatine | 47 |
Wolfhügel's Counter | 49 |
Petri's Dish | 50 |
Berkefeld Filter | 52 |
Apparatus for Filtering Water to Facilitate its Bacteriological Examination | to face 52 |
Bacteria of Typhoid Fever | 56 |
Bacillus Coli Communis | 60 |
The Comma-Shaped Bacillus of Cholera | 66 |
*Bacillus Typhosus | to face 66 |
*Bacillus Typhosus | to"fac66 |
v*Bacillus Coli Communis | 66 |
*Bacillus Mycoides | 66 |
Pasteur-Chamberland Filter | 80 |
Proteus Vulgaris | 86 |
Bacillus Enteriditis Sporogenes | 86 |
A Plan of Septic Tank and Filter-Beds | 91 |
Filter-Beds | 94 |
Miquel's Flask | 97 |
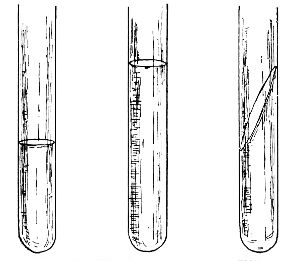
Sedgwick's Sugar-Tube | 99 |
Sedgwick's Tube | 100 |
Saccharomyces Cerevisiæ | 117 |
Ascospore Formation | 120 |
Gypsum Block | 121 |
Yeast | to face 122 |
Ascospore Formation in Yeast | to"fac122 |
Nitrogen-Fixing Bacteria from Rootlet Nodules | to"fac122 |
*Bacillus of Tetanus | to"fac122 |
Saccharomyces Ellipsoideus | 126 |
Saccharomyces Pastorianus | 126 |
Bacillus Acidi Lactici | 131 |
Bacillus Butyricus | 133 |
Kipp's Apparatus | 140 |
Fränkel's Tube | 141 |
Buchner's Tube | 141 |
A Method of Growing Cultivations in a Vacuum over Pyrogallic Solution | 143 |
Micrococcus from Soil | 151 |
Nitrous Organism | to face 158 |
Nitric Organism | to"face158 |
Nitrogen-Fixing Organism from Secretion of Root-Nodules | to"face158 |
Rootlet of Pea with Nodules | 163 |
Nitrogen-Fixing Bacteria in Situ in Nodule on Rootlet of a Pea | to face 164 |
Nitrogen-Fixing Bacteria in Situ in Rootlet-Nodule of a Pea | to"fac164 |
Nitrogen-Fixing Bacteria in Situ in Root-Nodule of a Pea | to"fac164 |
Bacillus of Tetanus | 170 |
Bacillus of Symptomatic Anthrax | 172 |
Bacillus of Malignant Œdema | 172 |
A Centrifuge | 228 |
Suspended Spinal Cord | 255 |
Flask Used in the Preparation of the Toxin of Diphtheria | 262 |
vi*Bacillus Tuberculosis | to face 280 |
*Bacillus Tuberculosis | 280 |
*Streptococcus Pyogenes | 280 |
*Bacillus Anthracis | 280 |
Flask Used in the Preparation of Tuberculin | 282 |
Bacillus of Diphtheria | 289 |
Types of Streptococcus | 298 |
Micrococcus Tetragonus | 299 |
Diplococcus of Neisser | 300 |
Bacillus of Anthrax and Blood Corpuscles | 302 |
Threads of Bacillus Anthracis, Showing Spores | 302 |
Bacillus of Plague | 306 |
*Bacillus of Plague | to face 310 |
*Bacillus of Leprosy | to"fac310 |
Streptothrix Actinomyces | to"fac310 |
Bacillus Mallei | to"fac310 |
Diplococcus of Pneumonia | 312 |
Bacillus of Influenza | 315 |
We live in a world that is teeming with life. From the earliest times of man that life has been studied and the observations recorded. Thus there has slowly come to be a considerable accumulation of knowledge concerning the various forms (morphology) and functions (physiology) of organised life. This we call the science of biology. It has for its object the study of organic beings, and for its end the knowledge of the laws of their organisation and activity. Slowly, too, in the midst of this gradual accumulation of facts, we begin to see incoherence becoming coherent, chaos becoming cosmos, chance and accident becoming law. Further, the contemplation and comprehension which built up the edifice of modern biology is assuming a new relationship to practical life. Biology can no longer be considered only as an academic occupation or as a theoretical pabulum upon which the leisured mind may ruminate. With rapid strides and determined face this giant of knowledge has marched into the arena of practical politics. The world is opening its eyes to a reality which it had mistaken for a vision.
This application of biology to life and its problems has in recent years been nowhere more marked than in the realm of bacteriology. This comparatively new science, associated with the great names of Pasteur, Koch, and Lister, furnishes indeed a stock illustration of the applicability of pure biology. Turn where we will, we shall find the work of the unseen hosts of bacteria daily claiming more and more attention from practical people. Thus biology, even when clothed in the form of microscopic cells, is coming to occupy a new place in the minds of men. "Its evolution," as Professorix Patrick Geddes declares, "forms part of the general social evolution." Certainly its recent rapid development forms a remarkable feature in the practical science of our time. Not only in the diagnosis and treatment of disease, nor even in the various applications of preventive medicine, but in ever-increasing degree and sphere, micro-organisms are recognised as agents of utility or otherwise no longer to be ignored. They occur in our drinking water, in our milk supply, in the air we breathe. They ripen cream, and flavour butter. They purify sewage, and remove waste organic products from the land. They are the active agents in a dozen industrial fermentations. They assist in the fixation of free nitrogen, and they build up assimilable compounds. Their activity assumes innumerable phases and occupies many spheres, more frequently proving themselves beneficial than injurious. They are both economic and industrious in the best biological sense of the terms.
Yet bacteriology has its limitations. It is well to recognise this, for the new science has in some measure suffered in the past from over-zealous friends. It cannot achieve everything demanded of it, nor can it furnish a cause for every disease. It is a science fuller of hope than proved and tested knowledge. We are as yet only upon the threshold of the matter. As in the neighbouring realm of chemistry, it is to be feared that bacteriology has not been without its alchemy. The interpretations and conclusions which have been drawn from time to time respecting bacteriological work have led to alarmist views which have not, by later investigation, been fully supported. Again, the science has had devotees who have fondly believed, like the alchemists, that the twin secret of transmuting the baser metals into gold and of indefinitely prolonging human life was at last to be known. But neither the worst fears of the alarmist nor the most sanguine hopes of the alchemist have been verified. Science, fortunately, does not progress at such speed, or with such kindly accomxmodation. It holds many things in its hands, but not finally life or death. It has not yet brought to light either "the philosopher's stone" or "the vital essence."
What has already been said affords ample reason for a wider dissemination of the elementary facts of bacteriological science. But there are other reasons of a more practical nature. Municipalities are expending public moneys in water analysis, in the examination of milk, in the inspection of cows and dairies, in the bacterial treatment of sewage, and in disinfection and other branches of public health administration. Again, the newly formed National Association for the Prevention of Tuberculosis, our increasing colonial possessions with their tropical diseases, even medical science itself, which is year by year becoming more preventive, make an increasing claim upon public opinion. The successful accomplishment and solution of these questions depend in a measure upon an educated public opinion respecting the elements of bacteriology. Recently it was urged that "the first elements of bacteriology should be shadowed forth in the primary school."1 This course was advised owing to such knowledge being of value to those engaged in dairying. As we shall point out at a later stage, many of the undesirable changes occurring in milk are due to bacteria, even as the success of the butter and cheese industries depends on the use and control of the fermentative processes due to their action. Much of the uncertainty attending the manufacture of dairy products can only be abolished by the careful application of some knowledge of the flora of milk. In Denmark and in Scandinavia the importance of such knowledge is realised and acted upon. America, too, has not been slow to respond to these needs; but in England comparatively little has been done in this direction.2
Whilst there can be no doubt as to the advantage of a wider dissemination of the ascertained facts concerning bacteria, it should be borne in mind that only patient, skilled observation and experimental research in well-equipped laboratories can advance this branch of science, or indeed train bacteriologists. The lives of Darwin and of Pasteur adequately illustrate this truth. Yet it is observable that States and public bodies are slow to act upon it, and frequently in the past the most useful and substantial support for the advancement of science has been forthcoming only from private sources. As the world learns its intimate relation to science and the interdependence between its life and scientific truth, it may be expected more heartily to support science.
BACTERIA
The first scientist who demonstrated the existence of micro-organisms was Antony von Leeuwenhoek. He was born at Delft, in Holland, in 1632, and enthusiastically pursued microscopy with primitive instruments. He corroborated Harvey's discovery of the circulation of the blood in the web of a frog's foot; he defined the red blood corpuscles of vertebrates, the fibres of the lens of the human eye, the scales of the skin, and the structure of hair. He was neither educated nor trained in science, but in the leisure time of his occupation as a linen-draper he learned the art of grinding lenses, in which he became so proficient that he was able to construct a microscope of greater power than had been previously manufactured. The compound microscope dates from 1590, and when Leeuwenhoek was about forty years old, Holland had already given to the world both microscope and telescope. Robert Hooke did for England what Hans Janssen had done for Holland, and established the same 2 conclusion that Leeuwenhoek arrived at independently, viz., that a simple globule of glass mounted between two metal plates and pierced with a minute aperture to allow rays of light to pass was a contrivance which would magnify more highly than the recognised microscopes of that day. It was with some such instrument as this that the first micro-organisms were observed in a drop of water. It was not until more than a hundred years later that these "animalcules," as they were termed, were thought to be anything more than accidental to any fluid or substance containing them. Plenciz, of Vienna, was one of the first to conceive the idea that decomposition could only take place in the presence of some of these "animalcules." This was in the middle of the eighteenth century. Just about a century later, by a series of important discoveries, it was established beyond dispute that these micro-organisms had an intimate causal relation to fermentation, putrefaction, and infectious diseases. Spallanzani, Pasteur, and Tyndall are the three who more than others contributed to this discovery. Spallanzani was an Italian, who studied at Bologna, and was in 1754 appointed to the chair of logic at Reggio. But his inclinations led him into the realm of natural history. Amongst other things, his attention was directed to the doctrine of spontaneous generation, which had been propounded by Needham a few years previously. In 1768 Spallanzani became Professor of Natural History at Pavia, and whilst there he demonstrated that if infusions of vegetable matter were placed in flasks and hermetically sealed, and then brought to the boiling point, no living organisms could thereafter be detected, nor did the vegetable matter decompose. When, however, the flasks were very slightly cracked, and air gained admittance, then invariably both organisms and decomposition appeared. Schwann, the founder of the cell-theory, and Schulze, both showed that if the air gaining access to the flask were either passed through highly heated3 tubes or drawn through strong acid the result was the same as if no air entered at all, viz., no organisms and no decomposition. The result of these investigations was that scientific men began to believe that no form of life arose de novo (abiogenesis), but had its source in previous life (biogenesis). It remained to Pasteur and Tyndall to demonstrate this beyond dispute, and to put to rout the fresh arguments for spontaneous generation which Pouchet had advanced as late as 1859. Pasteur collected the floating dust of the air, and found by means of the microscope many organised particles, which he sowed on suitable infusions, and thus obtained rich crops of "animalculæ." He also demonstrated that these organisms existed in different degrees in different atmospheres, few in the pure air of the Mer de Glace, more in the air of the plains, most in the air of towns. He further proved that it was not necessary to insist upon hermetic sealing or cotton filters to keep these living organisms in the air from gaining access to a flask of infusion. If the neck of the flask were drawn out into a long tube and turned downwards, and then a little upwards, even though the end be left open, no contamination gained access. Hence, if the infusion were boiled, no putrefaction would occur. The organisms which fell into the open end of the tube were arrested in the condensation water in the angle of the tube; but even if that were not so, the force of gravity acting upon them prevented them from passing up the long arm of the tube into the neck of the flask. A few years after Pasteur's first work on this subject Tyndall conceived a precise method of determining the absence or presence of dust particles in the air by passing a beam of sunlight through a glass box before and after its walls had been coated with glycerine. Into the floor of the box were fixed the mouths of flasks of infusion. These were boiled, after which they were allowed to cool, and might then be kept for weeks or months without putrefying or revealing the presence of germ life. Here all the con4ditions of the infusions were natural, except that in the air above them there was no dust.
The sum-total of result arising from all these investigations was to the effect that no spontaneous generation was possible, that the atmosphere contained unseen germs of life, that the smallest of organisms responded to the law of gravitation and adhered to moist surfaces, and that micro-organisms were in some way or other the cause of putrefaction.
The final refutation of the hypothesis of spontaneous generation was followed by an awakened interest in the unseen world of micro-organic life. Investigations into fermentation and putrefaction followed each other rapidly, and in 1863 Davaine claimed that Pollender's bacillus of anthrax, which was found in the blood and body tissues of animals dead of anthrax, was the cause of that disease. From that time to this in every department of biology bacteria have been increasingly found to play an important part. They cause changes in milk, and flavour butter; they decompose animal matter, yet build up the broken-down elements into compounds suitable for use in nature's economy; they assist in the fixation of free nitrogen; they purify sewage; in certain well-established cases they are the cause of specific disease, and in many other cases they are the likely cause. No doubt the disposal of spontaneous generation did much to arouse interest in this branch of science. Yet it must not be forgotten that the advance of the microscope and bacteriological method and technique have played a large share in this development. The sterilisation of culture fluids by heat, the use of aniline dyes as staining agents, the introduction of solid culture media (like gelatine and agar), and Koch's "plate" method have all contributed not a little to the enormous strides of bacteriology. Owing to its relation to disease, physicians have entered keenly into the arena of bacteriological research. Hence, from a variety of causes, it has come about that the advance has been phenomenal.
We shall now take up a number of points in the biology of bacteria which call for early attention, and which are mostly the outcome of comparatively recent work on the subject.
The Place of Bacteria in Nature. As we have seen, for a considerable period of time after their first detection these unicellular organisms were considered to be members of the animal kingdom. As late as 1838, when Ehrenberg and Dujardin drew up their classification, bacteria were placed among the Infusorians. This was in part due to the powers of motion which these observers detected in bacteria. It is now, of course, recognised that animals have no monopoly of motion. But what, after all, are the differences between animals and vegetables so low down in the scale of life? Chiefly two: there is a difference in life-history (in structure and development), and there is a difference in diet. A plant secures its nourishment from much simpler elements than is the case with animals; for example, it obtains its carbon from the carbonic acid gas in air and water. This it is able to do, as regards the carbon, by means of the green colouring matter known as chlorophyll, by the aid of which, with sunlight, carbonic acid is decomposed in the chlorophyll corpuscles, the oxygen passing back into the atmosphere, the carbon being stored in the plant in the form of starch or other organic compound. The supply of carbon in the chlorophyll-free plants, among which are the bacteria, is obtained by breaking up different forms of carbohydrates. Besides albumen and peptone, they use sugar and similar carbohydrates and glycerine as a source of carbon. Many of them also have the capacity of using organic matters of complex constitution by converting such into water, carbonic acid gas, and ammonia. Their hydrogen comes from water, their nitrogen from the soil, chiefly in the form of nitrates. From the soil, too, they obtain other necessary salts. Now all these substances are in an elementary condition, and as6 such plants can absorb them. Animals, on the other hand, are only able to utilise compound food products which have been, so to speak, prepared for them; for example, albuminoids and proteids. They cannot directly feed upon the elementary substances forming the diet of vegetables. This distinction, however, did not at once clear up the difficult matter of the classification of bacteria. It is true, they possess motion, are free from chlorophyll, and even feed occasionally upon products of decomposition—three physiological characters which would ally them to the animal kingdom. Yet by their structure and capsule of cellulose and by their life-history and mode of growth they unmistakably proclaim themselves to be of the vegetable kingdom. In 1853 Cohn arrived at a conclusion to this effect, and since that date they have become more and more limited in classification and restricted in definition.
Even yet, however, we are far from a scientific classification for bacteria. Nor is this matter for surprise. The development in this branch of biology has been so rapid that it has been impossible to assimilate the facts collected. The facts themselves by their remarkable variety have not aided classification. Names which a few years ago were applied to individual species, like Bacillus subtilis, or Bacterium termo, or Bacillus coli, are now representative, not of individuals, but of families and groups of species. Again, isolated characteristics of certain microbes, such as motility, power of liquefying gelatine, size, colour, and so forth, which at first sight might appear as likely to form a basis for classification, are found to vary not only between similar germs, but in the same germ. Different physical conditions have so powerful an influence upon these microscopic cells that their individual characters are constantly undergoing change. For example, bacteria in old cultures assume a different size, and often a different shape, from younger members of precisely the same species; Bacillus7 pyocyaneus produces a green to olive colour on gelatine, but a brown colour on potato; the bacillus of Tetanus is virulently pathogenic, and yet may not act thus unless in company with certain other micro-organisms. Hence it will at once appear to the student of bacteriology that, though there is great need for classification amongst the six or seven hundred species of microbes, our present knowledge of their life-history is not yet advanced enough to form more than a provisional arrangement.
We know that bacteria are allied to moulds on the one hand and yeasts on the other, and that they have no differentiation into root, stem, or leaf; we know that they are fungi (having no chlorophyll), in which no sexual reproduction occurs, and that their mode of multiplication is by division. From such facts as these we may build up a classification as follows:—
| Vegetable Kingdom. | |||||||
| │ | |||||||
| ┌─────────────────┬─────────────┬───────────┐ | |||||||
| Thallophyta. [= The lowest forms of vegetable life. No differentiation into root, stem, or leaf.] │ Protophyta. [= No sexual reproduction.] |
Muscineæ | Pteridophyta. | Phanerogamia. | ||||
| │ | |||||||
| ┌──────────┐ | |||||||
| Algæ. [= Chlorophyll present.] |
Fungi. [= No chlorophyll.] │ |
||||||
| ┌─┬─┬─┬─┬─┬─┬─┬─┬─┐ | |||||||
| │ Schizomycetes [= multiplication by cell division or by spores] or Bacteria |
 |
(1) Coccaceæ4—round cells. (2) Bacteriaceæ—rods and threads. |
|||||
| (3) Leptotricheæ. (4) Cladotricheæ. |
 |
Higher Bacteria | |||||
Structure and Form. Having now located micro-organisms in the economy of nature, we may proceed to describe their subdivisions and form. For practical convenience rather than academic accuracy, we may accept the simple division of the family of bacteria into three chief forms, viz.:—
| Lower Bacteria |  |
(1) Round cell form—coccus. (2) Rod form—bacillus. (3) Thread form—spirillum. |
Higher Bacteria—Leptothrix, Streptothrix, Cladothrix, etc.
A classification dependent as this is upon the form alone is not by any means ideal, for it ignores all the higher and complicated functions of bacteria, but it is, as we have said, practically convenient.

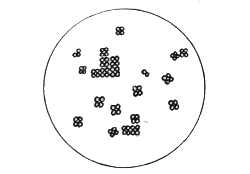
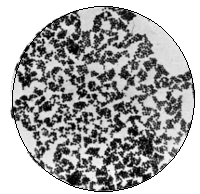
Various Forms of Bacteria
| 1. Micrococcus | 2. Diplococcus | 3. Streptococcus |
| 4. Staphylococcus | 5. Leuconostoc, showing Arthrospores | |
| 6. Merismopedia | 7. Sarcina | 8. Bacilli |
| 9. Spirillum | ||
1. The Coccus. This is the group of round cells. They vary in size as regards species, and as regards the conditions, artificial or natural, under which they have been grown. Some are less than 1/25000 of an inch in diameter; others are half as large again, if the word large may be used to describe such minute objects. No regular standard can be laid down as reliable with regard to their size. Hence the subdivisions of the cocci are dependent not upon the individual elements so much as upon the relation of those elements to each other. A simple round cell of approximately the size already named is termed a micrococcus (μικρος, small). Certain species of micrococci always or almost always occur in pairs, and such a combination is termed a diplococcus. Some diplococci are united by a thin capsule, which may be made apparent by special methods of staining; of others no limiting or uniting membrane can be seen with the ordinary high powers of the microscope.5 Again, one frequently finds a species which is exactly described by saying that two micrococci are in contact with each other, and move and act as one individual, but otherwise show no alteration; whilst others are seen 9which show a flattening of the side of each micrococcus which is in relation to its partner. Perhaps the diplococci in an even greater degree than the micrococci respond to external conditions both as regards size and shape. It must further be borne in mind that a dividing micrococcus assumes the exact appearance of a diplococcus during the transition stage of the fission. Hence, with the exception of several well-marked species of diplococci, this form is somewhat arbitrary. The third kind of micrococcus is that formed by a number of elements in a twisted chain, named streptococcus (στρεπτος, twisted). This form is produced by cells dividing in one axis, and remaining in contact with each other. It occurs in a number of different species, or what are supposed10 by many authorities to be different species, owing to their different effects. Morphologically all the streptococci are similar, though a somewhat abortive attempt was once made to divide them into two groups, according to whether they were long chains or short. As a matter of fact, the length of streptococci depends in some cases upon biological properties, in others upon external treatment or the medium of cultivation which has been used. Sometimes they occur as straight chains of only half a dozen elements; at other times they may contain thirty to forty elements, and twist in various ways, even forming rosaries. The elements, too, differ not only in size, but in shape, appearing occasionally as oval cells united to each other at their sides. The fourth form is constituted by the micrococci being arranged in masses like grapes, the staphylococcus (σταφυλις, a bunch of grapes). The elements are often smaller than in the streptococcus, and the name itself describes the arrangement. There is no matrix and no capsule. This is the commonest organism found in abscesses, etc. The sarcina is best classified amongst the cocci, for it is composed of them, in packets of four or multiples of four, produced by division vertically in two planes. If the division occurs in one plane, we have as a result small squares of round cells known as11 merismopedia. In both these conditions it frequently happens that the contiguous sides of the elements of packets become faceted or straightened against each other. It may happen, too, particularly in the sarcinæ, that segmentation is not complete, and that the elements are larger than in any other class of cocci. They stain very readily. Nearly all the cocci are non-motile, though Brownian movement may readily be observed.
Sarcina
2. The Bacilli. These consist of rods, having parallel sides and being longer than they are broad. They differ in every other respect according to species, but these two characteristics remain to distinguish them. Many of them are motile, others not. The ends or poles of a bacillus may be pointed, round, or almost exactly square and blocked. They all, or nearly all, possess a capsule. Individuals of the same species may differ greatly, according to whether they have been naturally or artificially grown, and pleomorphic forms are abundant.
3. The Spirilla. This wavy thread group is divisible into a number of different forms, to which authorities have given special names. It is sufficient, however, to state that the two common forms are the non-septate spiral thread (like the Spirillum Obermeier of relapsing fever), which takes no other form but a lengthened spirillum; and the spirillum which breaks up into elements or units, each of which appears comma-shaped (like the cholera bacillus). The degree of curvature in the spirilla, of course, varies. They are the least important of the lower bacteria.
The Higher Bacteria group includes more highly organised members of the Schizomycetes. They possess filaments, which may be branched, and almost always have septa and a sheath. Perhaps the most marked difference from the lower bacteria is in their reproduction. In the higher bacteria we have what is in fact a flower—terminal fructification by conidia. In this group of vegetables we have the12 Beggiatoa, Leptothrix, Cladothrix, and, at the top, the Streptothrix. It has been demonstrated that Streptothrix actinomycotica and Streptothrix maduræ are the organismal cause, respectively, of Actinomycosis and Madura-foot, two diseases which have hitherto been obscure.
Pleomorphism. This term designates an irregular development of a species. Different media and external conditions bring about in protoplasm as susceptible as mycoprotein a variety of morphological phases. These may occur in succession, and represent different stages in the life-history of a bacterium, or they may be involution forms resulting from a change of environment, and occurring as "faults" in the species. In the Bacillus coli, B. typhosus, bacillus of Plague, and B. tuberculosis pleomorphism undoubtedly occurs, and is manifest in the change of shape. This is particularly marked in old cultures of the last named. The ordinary well-known bacillus may grow out into threads, with bulbous endings, granular filaments, drumsticks, and diplococcal forms. Speaking generally, the older the culture, the more marked is the variation.
Polymorphism is a term used to define the theory which held that bacteria were one of the intermediate shapes or forms between something lower and something higher in the vegetable kingdom. Neither pleomorphism nor polymorphism is fully understood, and many bacteriologists find shelter from both in the term involution form. What we do know is that the species already named, for example, take on divers forms when placed under different conditions.
Composition. From what we have seen of the diet of micro-organisms, we shall conclude that in some form or other they contain the elements nitrogen, carbon, and hydrogen. All three substances are combined in the mycoprotein or protoplasm of which the body of the microbe consists. This is generally homogeneous, and there is no sign of a nucleus. It possesses a fortunate affinity for aniline dyes, and by this13 means organisms are stained for the microscope. Besides the variable quantity of nitrogen present, mycoprotein may also contain various mineral salts. The uniformity of the cell protoplasm may be materially affected by disintegration and segmentation due to degenerative changes. Vacuoles also may appear from a like cause, which it is necessary to differentiate from spores. Two other signs of degeneration are the appearance of granules in the body of the cell protoplasm known as metachromatic granules, owing to their different staining propensities, and the polar bodies which are seen in some species of bacteria. Surrounding the mass of mycoprotein, we find in most organisms a capsule or membrane composed, in part at least, of cellulose. This sheath plays a protective part in several ways. During the adult stage of life it protects the mycoprotein, and holds it together. At the time of reproduction or degeneration it not infrequently swells up, and forms a viscous hilum or matrix, inside which are formed the new sheaths of the younger generation. It may be rigid, and so maintain the normal shape of the species, or, on the other hand, flexible, and so adapted to rapid movement of the individual.

Normal and Pleon-Forms of the Bacillus of Tubash.
Here, then, we have the major parts in the constitution of a bacillus—its body, mycoprotein; its capsule, cellulose. But, further than this, there are a number of additional dis14tinctive characteristics as regards the contents inside the capsule which call for mention. Sulphur occurs in the Beggiatoa which thrive in sulphur springs. Starch is commoner still. Iron as oxide or other combination is found in several species. Many are highly coloured, though these are generally the "innocent" bacteria, in contradistinction to the disease-producing. A pigment has been found which is designated bacterio-purpurin. According to Zopf, the colouring agents of bacteria are the same as, or closely allied to, the colouring matters occurring widely in nature. Migula holds that most of the bacterial pigments are non-nitrogenous bodies. There are a very large number of chromogenic bacteria, some of which produce exceedingly brilliant colours. Among some of the commoner forms possessing this character are Bacillus et micrococcus violaceus (violet); B. et M. aurantiacus (orange); B. et M. luteus (yellow); M. roseus (pink); many of the Sarcinæ; B. aureus (golden-yellow); B. fluorescens liquefaciens et non-liquefaciens (green); B. pyocyaneus (green); B. prodigiosus (blood-red).
Motility. When a drop of water containing bacteria is placed upon a slide, a clean cover glass superimposed, and the specimen examined under an oil immersion lens, various rapid movements will generally be observed. These are of four kinds: (1) A dancing stationary motion known as Brownian movement. This is molecular, and depends in some degree upon heat and the medium of the moving particles. It is non-progressive, and is well known in gamboge particles. (2) An undulatory serpentine movement, with apparently little advance being made. (3) A rotatory movement, which in some water bacilli is very marked, and consists of spinning round, with sometimes considerable velocity, and maintained for some seconds or even minutes. (4) A progressive darting movement, by which the bacillus passes over some considerable distance.
The conditions affecting the motion of bacteria are but15 partly understood. Heating the slide or medium accelerates all movement. A fresh supply of oxygen, or indeed the addition of some nutrient substance, like broth, will have the same effect. There are also the somewhat mysterious powers by which cells possess inherent attraction or repulsion for other cells, known as positive and negative chemiotaxis. These powers have been observed in bacteria by Pfeiffer and Ali-Cohen.
Bacilli, Showing Flagella
The essential condition in the motile bacilli is the presence of flagella.6 These cilia, or hairy processes, project from the sides or from the ends of the rod, and are freely motile and elastic. Sometimes only one or two terminal flagella are present; in other cases, like the bacillus of typhoid fever, five to twenty may occur all round the body of the bacillus, varying in length and size, sometimes being of greater length even than the bacillus itself. It is not yet established as to whether these vibratile cilia are prolongations of capsule only, or whether they contain something of 16 the body protoplasm. Migula holds the former view, and states that the position of flagella is constant enough for diagnostic purposes. They are but rarely recognisable except by means of special staining methods. Micrococcus agilis (Ali-Cohen) is the only coccus which has flagella and active motion.
Modes of Reproduction. Budding, division, and spore formation are the three chief ways in which Schizomycetes and Saccharomycetes (yeasts) reproduce their kind. Budding occurs in some kinds of yeast, and would be classified by some authorities under spore formation, but in practice it is so obviously a "budding" that it may be so classified. The capsule of a large or mother cell shows a slight protrusion outwards which is gradually enlarged into a daughter yeast and later on becomes constricted at the neck. Eventually it separates as an individual. The protoplasm of spores of yeasts differs, as Hansen has pointed out, according to their conditions of culture.
Division, or fission, is the commonest method of reproduction. It occurs transversely. A small indentation occurs in the capsule, which appears to make its way slowly through the whole body of the bacillus or micrococcus until the two parts are separate, and each contained in its own capsule. It has been pointed out already that in the incomplete division of micrococci we observe a stage precisely similar to a diplococcus. So also in the division of bacilli an appearance occurs described as a diplobacillus.
Simple fission requires but a short period of time to be complete. Hence multiplication is very rapid, for within half an hour a new adult individual can be produced. It has been estimated that at this rate one bacillus will in twenty-four hours produce 17,000,000 similar individuals; or, expressed in another way, Cohn calculated that in three days, under favourable circumstances, this rate of increase would form a mass of living organisms weighing 7300 tons,17 and numbering about 4772 billions. Favourable conditions do not occur, fortunately, to allow of such increase, which, of course, can only be roughly estimated. But the above figures illustrate the enormous fertility of micro-organic life. When we remember that in some species it requires 10,000 or 15,000 fully grown bacilli placed end to end to stretch the length of an inch, we see also how exceedingly small are the individuals composing these unseen hosts.
Spore formation may result in the production of germinating cells inside the capsule of the bacillus, endospores, or of modified individuals, arthrospores. The body of a bacillus, in which sporulation is about to occur, loses its homogeneous character and becomes granular, owing to the appearance of globules in the protoplasm. In the course of three or four hours the globule enlarges to fill the diameter of the rod, and assumes a more concentrated condition than the parent cell. At its maturity, and before its rupture of the bacillary capsule, a spore is observed to be bright and shining, oval and regular in shape, with concentrated contents, and frequently causing a local expansion of the bacillus. In a number of rods lying endwise, these local swellings produce a beaded or varicose appearance, even simulating a streptococcus. In the meantime the rod itself has become slightly broader and pale. Eventually it breaks down by segmentation or by swelling up into a gelatinous mass. The spore now escapes and commences its individual existence. Under favourable circumstances it will germinate. The tough capsule gives way at one point, generally at one of the poles, and the spore sprouts like a seed. In the space of about one hour's time the oval refractile cell has become a new bacillus. One spore produces by germination one bacillus. Spores never multiply by fission, nor reproduce themselves.
Hueppe has stated that there are certain organisms (like leuconostoc, and some streptococci) which reproduce by the method of arthrospores. Defined shortly, this is simply an18 enlargement of one or more cell elements in the chain which thus takes on the function of maternity. On either side of the large coccus may be seen the smaller ones, which it is supposed have contributed of their protoplasm to form a mother cell. An arthrospore is said to be larger, more refractile, and more resistant than an ordinary endospore. Many bacteriologists of repute have declined hitherto to definitely accept arthrospore formation as a proved fact.

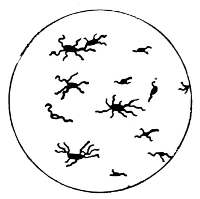
Various Forms of Spore Formation and Flagella
A. Stages in formation of spore and its after development. B. Spirillum with terminal flagella.
It is important to note that spore formation in bacteria
must not be considered as a method of multiplication. The
general rule is undoubtedly that one bacillus produces one
spore, and one spore germinates into one bacillus. It is a reproduction,
not a multiplication. Indeed, the whole process
is of the nature of a resting stage, and is due (a) to the
arrival of the adult bacillus at its biological zenith, or (b) to
the conditions in which it finds itself being unfavourable19
to its highest vegetative growth, and so it endeavours to
perpetuate its species. Most authorities are probably of the
latter opinion, though there is not a little evidence for the
former. Exactly what conditions are favourable to sporulation
is not known. Nutriment has probably an intimate
effect upon it. The temperature must not be below 16° C.,
nor much above 40° C. Oxygen, as we have seen, is favourable,
if not necessary, to many species, which will in cultivation
in broth rise to the surface and lodge in the pellicle to
form their seeds. Moisture, too, is considered a necessity.
The position and size of the spore are of considerable use in differential diagnosis. The terminal spore of Bacillus tetani is well known. It is rarely seen at both ends of the bacillus, and hence when poised only at one end causes the "drumstick" appearance. In the bacillus of Quarter Evil the spore is generally towards one end of the rod rather than in the middle; in Malignant Œdema the bacillus in the blood grows out into long threads, and when such a thread sporulates the spore is also near one end. The latter further illustrates the fact that in some species the spore is of greater diameter than the mother cell, and hence dilates the bacillary capsule. The spores of anthrax are typical oval endospores. When free in the field of the microscope, spores must be distinguished from fat cells, micrococci, starch cells, some kinds of ova, yeast cells, and other like objects. Spores are detected frequently by their resistance to ordinary stains and the necessity of colouring them by special staining methods. When, however, a spore has taken on the desired colour, it retains it with tenacity. In addition to their shape, size, thickened capsule, and staining characteristics, spores also resist desiccation and heat in a much higher degree than bacilli not bearing spores. Roux and some other eminent bacteriologists suggest that bacteria should be classified according to their method of spore formation.
Nutritive Medium. In the very earliest days of the study of micro-organisms it was observed that they mostly congregate where there is pabulum for their nourishment. The reason why fluids such as milk, and dead animal matter such as a carcass, and living tissues such as a man's body contain so many microbes is because each of these three media is favourable to their growth. Milk affords almost an ideal food and environment for microbes. Its temperature and constitution frequently meet their requirements. Dead animal matter, too, yields a rich diet for some species (saprophytes). In the living tissues bacteria obtain not only nutriment, but a favourable temperature and moisture. Outside the human body it has been the endeavour of bacteriologists to provide media as like the above as possible, and containing many of the same elements of food. Thus the life-history may be carried on outside the body and under observation. By means of cover-glass preparations for the microscope we are able to study the form, size, motility, flagella, spore formation, and peculiarities of staining, all of which characters aid us in determining to what species the organism under examination belongs. By means of artificial nutrient media we may further learn the characters of the organism in "pure culture,"7 its favourable temperature, its power or otherwise of liquefaction, the curdling milk, or of gas production, its behaviour towards oxygen, its power of producing indol, pigment, and chemical bodies, as well as its thermal death point and resistance to light and disinfectants. It is well known that under artificial cultivation an organism may be greatly modified in its morphology and physiology, and yet its conformity to type 21remains much more marked than any degeneration which may occur.
The basis of many of these artificial media is broth. This
is made from good lean beef, free from fat and gristle, which
is finely minced up and extracted in sterilised water (one
pound of lean beef to every 1000 cc. of water). It is then
filtered and sterilised. It will be understood that such an
extract is acid. To provide peptone beef-broth, ten grains of
peptone and five grains of common salt are added to every
litre of acid beef-broth. It is rendered slightly alkaline by the
addition of sodium carbonate, and is filtered and sterilised.
Glycerine-broth indicates that 6 to 8 per cent. of glycerine has
been added after filtration, glucose-broth 1 or 2 per cent. of
grape-sugar. This latter is used for anaërobic organisms.
The use of broth as a culture medium is of great value. It
is undoubtedly our best fluid medium, and in it may not
only be kept pure cultures of bacteria which it is desired to
retain for a length of time, but in it also emulsions and mixtures
may be placed preparatory to further operations.
Gelatine is broth solidified by the addition of 100 grams of
best French gelatine to the litre. Its advantage is twofold:
it is transparent, and it allows manifestation of the power of
liquefaction. When we speak of a liquefying organism we
mean a germ having the power of producing a peptonising
ferment which can at the temperature of the room break
down solid gelatine into a liquid. Grape-sugar gelatine is
made like grape-sugar broth. Agar was introduced as a
medium which would not melt at 25° C., like gelatine, but
remain solid at blood-heat (37·5° C.; 98·5° F.). It is a seaweed
generally obtained in dried strips from the Japanese
market. Ten to fifteen grams are added to every litre of
peptone-broth. Filtration is slow and often difficult, and
the result not as transparent as desirable. The former difficulty
is avoided by filtering in the Koch's steamer or with
a hot-water filter, the latter by the addition of the white of22
an egg. Glycerine and grape-sugar may be added as elsewhere.
Blood agar is ordinary agar with fresh sterile blood
smeared over its surface. Blood serum is drawn from a jar
of coagulated horse-blood, in which the serum has risen to
 Potato in a
Roux Tube
Prepared for
Cultivation
Glycerine is
placed in the
bulb of the tube
the top. This is collected in sterilised tubes and
coagulated in a special apparatus (the serum inspissator).
Potato is prepared by scraping ordinary
potatoes, washing in corrosive sublimate, and
sterilising. They may then be cut into various
shapes convenient for cultivation. Upon any of
these forms of solid media the characteristic
growth of the organism can be observed. Of the
nutrient elements required, nitrogen is obtained
from albumens and proteids, carbon from milk-sugar,
cane-sugar, or the splitting up of proteids;
salts (particularly phosphates and salts of potassium)
are readily obtainable from those incorporated
in the media; and the water which is required is
obtainable from the moisture of the media.
Potato in a
Roux Tube
Prepared for
Cultivation
Glycerine is
placed in the
bulb of the tube
the top. This is collected in sterilised tubes and
coagulated in a special apparatus (the serum inspissator).
Potato is prepared by scraping ordinary
potatoes, washing in corrosive sublimate, and
sterilising. They may then be cut into various
shapes convenient for cultivation. Upon any of
these forms of solid media the characteristic
growth of the organism can be observed. Of the
nutrient elements required, nitrogen is obtained
from albumens and proteids, carbon from milk-sugar,
cane-sugar, or the splitting up of proteids;
salts (particularly phosphates and salts of potassium)
are readily obtainable from those incorporated
in the media; and the water which is required is
obtainable from the moisture of the media.
There are two common forms of test-tube culture, viz.: on the surface and in the depth of the medium. In the former the medium is sloped, and the inoculating needle is drawn along its surface; in the latter the needle is thrust vertically downwards into the depth of the solid medium. Plate cultures and anaërobic cultures will be described at a later stage. When the medium has been inoculated the culture is placed at a temperature which will be favourable. Two standards of temperature are in use in bacteriological laboratories. The one is called room temperature, and varies from 18° C.-20° C.; the other is blood-heat, and varies from 35° C.-38° C. It is true, some species will grow below 18° C., and others above 38° C. The pathogenic (disease-producing) bacteria thrive best at 37° C., and the non-pathogenic at the ordinary temperature of the room. The different degrees of temperature are regulated by means of incubators. For 23the low temperatures gelatine is chosen; as a medium for the higher temperatures agar.
Staphylococcus Pyogenes Aureus
× 1000
Incubator
(Temperature of blood-heat, registered by thermometer, and regulated
by thermo-regulator)
Moisture has been shown to have a favourable effect upon the growth of microbes. Drying will of itself kill many species (e. g., the spirillum of cholera), and, other things being equal, the moister a medium is, the better will be the growth upon it. Thus it is that the growth in broth is always more luxuriant than that on solid media. Yet the growth of Bacillus subtilis and other species is an exception to this rule, for they prefer a dry medium.
Culture Media Ready for Inoculation
Temperature. Most bacteria grow well at room temperature, but they will grow more luxuriantly and speedily at blood-heat. The optimum temperature is generally that of the natural habitat of the organism. In exceptional cases growth will occur as low as 5° C. or as high as 70° C. Indeed, some have been cooled to-20° C. and-30° C., and yet retained their vitality,8 whereas some few can grow at 2460–70° C. These latter are termed thermophilic bacteria. The average thermal death-point is at or about 50° C.
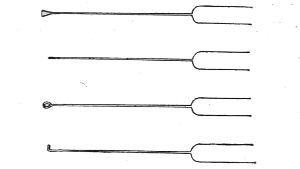
Inoculating Needles
Plantinum wire fused into glass handles
Light acts as an inhibitory or even germicidal agent. This fact was first established by Downes and Blunt in a memoir to the Royal Society in 1877. They found by exposing cultures to different degrees of sunlight that thus the growth of the culture was partially or entirely prevented, being most damaged by the direct rays of the sun, although diffuse daylight acted prejudicially. Further, these same investigators proved that of the rays of the spectrum which acted inimically the blue and violet rays acted most bactericidally, next to the blue being the red and orange-red rays. The action of light, they explain, is due to the gradual oxidation which is induced by the sun's rays in the presence of oxygen. Duclaux, who worked at this question at a later date, concluded that the degree of resistance to the bactericidal influence of light which some bacteria possess might be due to difference in species, difference in culture media, and difference in the degrees of intensity of light. Tyndall tested the growth of organisms in flasks exposed to air and light on the Alps, and found that sunlight inhibited the growth temporarily. A large number of experimenters in Europe and England have worked at this fascinating subject since 251877, and though many of their results appear contradictory, we may be satisfied to adopt the following conclusions respecting the matter:
(1) Sunlight has a deleterious effect upon bacteria, and to a less extent on their spores.
(2) This inimical effect can be produced by light irrespective of rise in temperature.
(3) The ultra-violet rays are the most bactericidal, and the infra-red the least so, which indicates that the phenomenon is due to chemical action.
(4) The presence of oxygen and moisture greatly increases this action.
(5) The sunlight acts prejudicially upon the culture medium, and thereby complicates the investigation and after-growth.
(6) The time occupied in the bactericidal action depends upon the heat of the sun and the intrinsic vitality of the organism.
(7) With regard to the action of light upon pathogenic organisms, some results have recently been obtained with Bacillus typhosus. Janowski maintains that direct sunlight exerts a distinctly depressing effect on typhoid bacilli. At present more cannot be said than that sunlight and fresh air are two of the most powerful agents we possess with which to combat pathogenic germs.

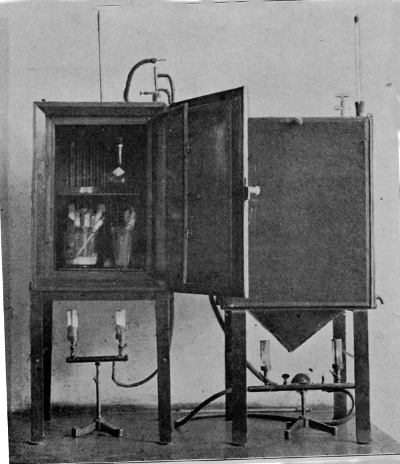
Pasteur's Large Incubator for Cultivation at Room Temperature
A very simple method of demonstrating the influence of light is to grow a pure culture in a favourable medium, either in a test-tube or upon a glass plate, and then cover the whole with black paper or cloth. A little window may then be cut in the protective covering, and the whole exposed to the light. Where it reaches in direct rays it will be found that little or no growth has occurred; where, on the other hand, the culture has been in the dark, abundant growth occurs. In diffuse light the growth is merely somewhat inhibited. It has been found that the electric light has but26 little action upon bacteria, though that which it has is similar to sunlight. Recent experiments with the Röntgen rays have given negative results.
In 1890 Koch stated that tubercle bacilli were killed after an exposure to direct sunlight of from a few minutes to several hours. The influence of diffuse light would obviously be much less. Professor Marshall Ward has experimented with the resistant spores of Bacillus anthracis by growing these on agar plates and exposing to sunlight. From two to six hours' exposure had a germicidal effect.
It should be remembered that several species of sea-water bacteria themselves possess powers of phosphorescence. Pflüger was the first to point out that it was such organisms which provided the phosphorescence upon decomposing wood or decaying fish. To what this light is due, whether capsule, or protoplasm, or chemical product, is not yet known. The only facts at present established are to the effect that certain kinds of media and pabulum favour or deter phosphorescence.
Desiccation. A later opportunity will occur for consideration of the effect of drying upon bacteria. Here it is only necessary to say that, other things being equal, drying diminishes virulence and lessens growth.
Oxygen. Pasteur was the first to lay emphasis upon the effect which free air had upon micro-organisms. He classified them according to whether they grew in air, aërobic, or whether they flourished most without it, anaërobic. Some have the faculty of growing with or without the presence of oxygen, and are designated as facultative aërobes or anaërobes. As regards the cultivation of anaërobic germs, it is only necessary to say here that hydrogen, nitrogen, or carbonic acid gas may be used in place of oxygen, or they may be grown in a medium containing some substance which will absorb the oxygen.
Modes of Bacterial Action. In considering the specific 27 action of micro-organisms, it is desirable, in the first place, to remember the two great functional divisions of saprophyte and parasite. A saprophyte is an organism that obtains its nutrition from dead organic matter. Its services, of whatever nature, lie outside the tissues of living animals. Its life is spent apart from a "host." A parasite, on the other hand, lives always at the expense of some other organism which is its host, in which it lives and upon which it lives. There is a third or intermediate group, known as "facultative," owing to their ability to act as parasites or saprophytes, as the exigencies of their life-history may demand.
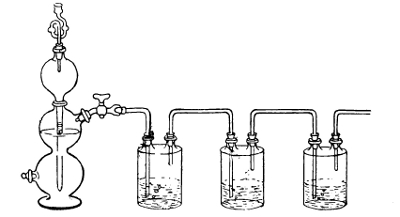
Method of Producing Hydrogen by Kipp's Apparatus for Cultivation of Anaërobes (See page 139)
The saprophytic organisms are, generally speaking, those
which contribute most to the benefit of man, and the parasitic
the reverse, though this statement is only approximately true.
In their relation to the processes of fermentation, decomposition,
nitrification, etc., we shall see how great and invaluable
is the work which saprophytic microbes perform. Their result
depends, in nearly all cases, upon the organic chemical
constitution of the substances upon which they are exerting28
their action, as well as upon the varieties of bacteria themselves.
Nor must it be understood that the action of saprophytes
is wholly that of breaking down and decomposition.
As a matter of fact, some of their work is, as we shall see,
of a constructive nature; but, of whichever kind it is, the
result depends upon the organism and its environment.
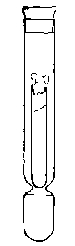 Anaërobic
Culture
Anaërobic
Culture
(Buckner's Tube)
with Pyrogallic
Solution in Bulb.
This, too, may be said of the pathogenic species, all of
which are in a greater or less degree parasitic. It is well
known how various are the constitutions of man,
how the bodies of some persons are more resistant
than those of others, and how the invading microbe
will find different receptions according to the constitution
and idiosyncrasy of the body which it
attacks. Indeed, even after invasion the infectivity
of the special disease, whatever it happens to be,
will be materially modified by the tissues. When
we come to turn to the micro-organisms which
are pathogenic parasites we shall further have to
keep clear in our minds that their action is double
and complex, and not single or simple. In the
first place, we have an infection of the body due
to the bacteria themselves. It may be a general
and widespread infection, as in anthrax, where the
bacilli pass, in the blood or lymph current, to
each and every part of the body; or it may be a
comparatively local one, as in diphtheria, where the invader
remains localised at the site of entrance. But, be
that as it may, the micro-organisms themselves, by their
own bodily presence, set up changes and perform functions
which may have far-reaching effects. It is obvious that
the wider the distribution the wider is the area of tissue
change, and vice versâ. Yet there is something of far
greater importance than the mere presence of bacteria in
human or animal tissues; for the secondary action of disease-producing
germs—and possibly it is present in all bacteria29—is
due to their poisonous products, or toxins, as they
have been termed. These may be of the nature of ferments,
and they become diffused throughout the body,
whether the bacteria themselves occur locally or generally.
They may bring about very slight and even imperceptible
changes during the course of the disease, or they may kill
the patient in a few hours. Latterly bacteriologists have
come to understand that it is not so much the presence
of organisms which is injurious to man and other animals,
as it is their products which cause the mischief; and the
amount of toxic product bears no known proportion to the
degree of invasion by the bacteria. The various and widely
differing modes of action in bacteria are therefore dependent
upon these three elements: the tissues or medium, the bacteria,
and the products of the bacteria; and in all organismal
processes these three elements act and react upon each other.
A word may be said here respecting the much-discussed question of species in bacteria. A species may be defined as "a group of individuals which, however many characters they share with other individuals, agree in presenting one or more characters of a peculiar and hereditary kind with some certain degree of distinctness."9 Now, as regards bacteria, there is no doubt that separate species occur and tend to remain as separate species. It is true, there are many variations, due in large measure to the medium in which the organisms are growing,—variations of age, adaptation, nutrition, etc.,—yet the different species tend to remain distinct. Involution forms occur frequently, and degeneration invariably modifies the normal appearance. But because of the occurrence of these morphological and even pathological differences it must not be argued that the demarcation of species is wholly arbitrary.
Means of Sterilisation. As this term occurs frequently in even a book of this untechnical nature, and as it is expressive 30of an idea which must always be present to the mind of the bacteriologist, it may be desirable to make some passing allusion to it.
Chemical substances, perfect filtration, and heat are the three means at our command in order to secure germ-free conditions of apparatus or medium. The first two, though theoretically admissible, are practically seldom used, the former of the two because the addition of chemical substances annuls or modifies the operation, the latter of the two on account of the great practical difficulties in securing perfection. Hence in the investigation involved in bacteriological research heat is the common sterilising agent. A temperature of 70° C. (158° F.) will kill all bacilli; even 58° C. will kill most kinds. Boiling at 100° C. (212° F.) for three minutes will kill anthrax spores, and boiling for thirty to sixty minutes will kill all bacilli and all spores. This difference in the thermal death-point between bacilli and their spores enables the operator to obtain what are called "pure cultures" of a desired bacillus from its spores which may be present. For example, if a culture contains spores of anthrax and is contaminated with micrococci, heating to 70° C. (158° F.) will kill all the micrococci, but will not affect the spores of anthrax, which can then grow into a pure culture of anthrax bacilli. Fractional or discontinuous sterilisation depends on the principle of heating to the sterilising point for bacilli (say 70°C.) on one day, which will kill the bacilli, but leave the spores uninjured. But by the following day the spores will have germinated into bacilli, and a second heating to 70°C. will kill them before they in their turn have had time to sporulate. Thus the whole will be sterilised, though at a temperature below boiling.
Successful sterilisation, therefore, depends upon killing
both bacteria and their spores, and nothing short of that
can be considered as sterilisation. The following methods
are those generally used in the laboratory. For dry heat31
(which is never so injurious to organisms as moist heat)10:
(a) the Bunsen burner, in the flame of which platinum
needles, etc., are sterilised; (b) hot-air chamber, in which
flasks and test-tubes are heated to a temperature of 150–170°
 Koch's Steam Steriliser
C. for half an hour. For moist
heat: (c) boiling, for knives and instruments;
(d) Koch's steam steriliser,
by means of which a crate is
slung in a metal cylinder, at the
bottom of which the water is
boiled; (e) the autoclave, which is
the most rapid and effective of all
the methods. This is in reality a
Koch steriliser, but with apparatus
for obtaining high pressure. The
last two (d, e) are used for sterilising
the nutriment media upon
which bacteria are cultivated outside
the body. Blood serum
would, however, coagulate at a
temperature over 60° C. (124° F.),
and hence a special steriliser has
been designed to carry out fractional
sterilisation daily for a week
at about 55° C.-58° C.
Koch's Steam Steriliser
C. for half an hour. For moist
heat: (c) boiling, for knives and instruments;
(d) Koch's steam steriliser,
by means of which a crate is
slung in a metal cylinder, at the
bottom of which the water is
boiled; (e) the autoclave, which is
the most rapid and effective of all
the methods. This is in reality a
Koch steriliser, but with apparatus
for obtaining high pressure. The
last two (d, e) are used for sterilising
the nutriment media upon
which bacteria are cultivated outside
the body. Blood serum
would, however, coagulate at a
temperature over 60° C. (124° F.),
and hence a special steriliser has
been designed to carry out fractional
sterilisation daily for a week
at about 55° C.-58° C.
The Association of Organisms. At a later stage we shall have an opportunity of discussing symbiosis and allied conditions. Here it is only necessary to draw attention to a fact that is rapidly 32becoming of the first importance in bacteriology. When species were first isolated in pure culture it was found that they behaved somewhat differently under differing circumstances. This modification in function has been attributed to differences of environment and physical conditions. Whilst it is true that such external conditions must have a marked effect upon such sensitive units of protoplasm as bacteria, it has recently been proved that one great reason why modification occurs in pure artificial cultures is that the species has been isolated from amongst its colleagues and doomed to a separate existence. One of the most abstruse problems in the immediate future of the science of bacteriology is to learn what intrinsic characters there are in species or individuals which act as a basis for the association of organisms for a specific purpose. Some bacteria appear to be unable to perform their regular function without the aid of others. An example of such association is well illustrated in the case of tetanus, for it has been shown that if the bacilli and spores of tetanus alone obtain entrance to a wound the disease may not follow the same course as when with the specific organism the lactic-acid bacillus or the common organisms of suppuration or putrefaction also gain entrance. There is here evidently something gained by association. Again, the virulence of other bacteria is also increased by means of association. The Bacillus coli is an example, for, in conjunction with other organisms, this bacillus, although normally present in health in the alimentary canal, is able to set up acute intestinal irritation, and 33various changes in the body of an inflammatory nature. It is not yet possible to say in what way or to what degree the association of bacteria influences their rôle. That is a problem for the future. But whilst we have examples of this association in streptococcus and the bacillus of diphtheria, B. coli and yeasts, tetanus and putrefactive bacteria, Diplococcus pneumoniæ and streptococcus, and association amongst the various suppurative organisms, we cannot doubt that there is an explanation to be found here of many hitherto unsolved results of bacterial action. This is the place in which mention should also be made of higher organisms associated for a specific purpose with bacteria. There is some evidence to support the belief that some of the Leptotricheæ (Crenothrix, Beggiatoa, Leptothrix, etc.) and the Cladotricheæ (Cladothrix) perform a preliminary disintegration of organic matter before the decomposing bacteria commence their labours. This occurs apparently in the self-purification of rivers, as well as in polluted soils.
Antagonism of Bacteria. Study of the life-history of many of the water bacteria will reveal the fact that they can live and multiply under conditions which would at once prove fatal to other species. Some of these water organisms can indeed increase and multiply in distilled water, whereas it is known that other species cannot even live in distilled water, owing to the lack of pabulum. Thus we see that what is favourable for one species may be the reverse for another.
Further, we shall have opportunity of observing, when considering the bacteriology of water and sewage, that there is in these media in nature a keen struggle for the survival of the fittest bacteria for each special medium. In a carcass it is the same. If saprophytic bacteria are present with pathogenic, there is a struggle for the survival of the latter. Now whilst this is in part due to a competition owing to a limited food supply and an unlimited population, as occurs34 in other spheres, it is also due in part to the inimical influence of the chemical products of the one species upon the life of the bacteria of the other species. Moreover, in one culture medium, as Cast has pointed out, two species will often not grow. When Pasteur found that exposure to air attenuated his cultures, he pointed out that it was not the air per se that hindered his growth, but it was the introduction of other species which competed with the original. The growth of the spirillum of cholera is opposed by Bacillus pyogenes fœtidus. B. anthracis is, in the body, opposed by either B. pyocyaneus or Streptococcus erysipelatis, and yet it is aided in its growth by B. prodigiosus. B. aceti is, under certain circumstances, antagonistic to B. coli communis.
In several of the most recent of the admirable reports of Sir Richard Thorne issued from the Medical Department of the Local Government Board, we have the record of a series of experiments performed by Dr. Klein into this question of the antagonism of microbes. From this work it is clearly demonstrated that whatever opposition one species affords to another it is able to exercise by means of its poisonous properties. These are of two kinds. There is, as is now widely known, the poisonous product named the toxin, into which we shall have to inquire more in detail at a later stage. There is also in many species, as Dr. Klein has pointed out, a poisonous constituent or constituents included in the body protoplasm of the bacillus, and which he therefore terms the intracellular poison. Now, whilst the former is different in every species, the latter may be a property common to several species. Hence those having a similar intracellular poison are antagonistic to each other, each member of such a group being unable to live in an environment of its own intracellular poison. Further, it has been suggested that there are organisms possessing only one poisonous property, namely, their toxin—for example, the bacilli of tetanus and diphtheria—whilst there are other species, as above, possess35ing a double poisonous property, an intracellular poison and a toxin. In this latter class would be included the bacilli of Anthrax and Tubercle.
Reference has been made to the associated work of higher vegetable life and bacteria. The converse is also true. Just as we have bacterial diseases affecting man and animals, so also plant life has its bacterial diseases. Wakker, Prillieux, Erwin Smith, and others have investigated the pathogenic conditions of plants due to bacteria, and though this branch of the science is in its very early stages, many facts have been learned. Hyacinth disease is due to a flagellated bacillus. The wilt of cucumbers and pumpkins is a common disease in some districts of the world, and may cause widespread injury. It is caused by a white microbe which fills the water-ducts. Wilting vines are full of the same sticky germs. Desiccation and sunlight have a strongly prejudicial effect upon these organisms. Bacterial brown-rot of potatoes and tomatoes is another plant disease probably due to a bacillus. The bacillus passes down the interior of the stem into the tubers, and brown-rots them from within. There is another form of brown-rot which affects cabbages. It blackens the veins of the leaves, and a woody ring which is formed in the stem causes the leaves to fall off. This also is due to a micro-organism, which gains entrance through the water-pores of the leaf, and subsequently passes into the vessels of the plants. It multiplies by simple fission, and possesses a flagellum.
There can be no doubt that these complex biological properties of association and antagonism, as well as the parasitic growth of bacteria upon higher vegetables, are as yet little understood, and we may be glad that any light is being shed upon them. In the biological study of soil bacteria in particular may we expect in the future to find examples of association, even as already there are signs that this is so in certain pathogenic conditions. In the alimentary canal, on36 the other hand, and in conditions where organic matter is greatly predominating, we may expect to see further light on the subject of antagonism.
Attenuation of Virulence or Function. It was pointed out by some of the pioneer bacteriologists that the function of bacteria suffered under certain circumstances a marked diminution in power. Later workers found that such a change might be artificially produced. Pasteur introduced the first method, which was the simple one of allowing cultures to grow old before sub-culturing. Obviously a pure culture cannot last for ever. To maintain the species in characteristic condition it is necessary frequently to sub-culture upon fresh media. If this simple operation be postponed as long as possible consistent with vitality, and then performed, it will be found that the sub-culture is attenuated, i. e., weakened. Another mode is to raise the pure culture to a temperature approaching its thermal death point. A third way of securing the same end is to place it under disadvantageous external circumstances, for example a too alkaline or too acid medium. A fourth, but rarely necessary, method is to pass it through the tissues of an insusceptible animal. Thus we see that, whilst the favourable conditions which we have considered afford full scope for the growth and performance of functions of bacteria, we are able by a partial withdrawal of these, short of that ending fatally, to modify the character and strength of bacteria. In future chapters we shall have opportunity of observing what can be done in this direction.
In entering upon a consideration of such a common article of use as water, we shall do well to describe in some detail the process by which we systematically investigate the bacteriology of a water, or, indeed, of any similar fluid suspected of bacterial pollution.
The collection of samples, though it appears simple enough, is sometimes a difficult and responsible undertaking. Complicated apparatus is rarely necessary, and fallacies will generally be avoided by observing two directions. In the first place, the sample should be chosen as representative as possible of the real substance or conditions we wish to examine. Some authorities advise that it is necessary to allow the tap to run for some minutes previous to collecting the sample; but if we desire to examine for lead chemically or for micro-organisms in the pipes biologically, then such a proceeding would be injudicious.11 Hence we must use common sense in the selection and obtaining of a sample, following this one guide, namely, to collect as nearly as possible a sample of the exact water the quality of which it is desired to learn. In the second place, we must observe strict bacteriological cleanliness in all our manipulations. This means that we must use only sterilised vessels or flasks for collecting the sample, and in the manipulation required we must be extremely careful 38to avoid any pollution of air or any addition to the organisms of the water from unsterilised apparatus. A flask polluted in only the most infinitesimal degree will entirely vitiate all results.
Accompanying the sample should be a more or less full statement of its source. There can be no doubt that, in addition to a chemical and bacteriological report of a water, there should also be made a careful examination of its source. This may appear to take the bacteriologist far afield, and in point of fact, as regards distance, this may be so. But until he has seen for himself what "the gathering-ground" is like, and from what sources come the feeding streams, he cannot judge the water as fairly as he should be able to do. The configuration of the gathering-ground, its subsoil, its geology, its rainfall, its relation to the slopes which it drains, the nature of its surface, the course of its feeders, and the absence or presence of cultivated areas, of roads, of houses, of farms, of human traffic, of cattle and sheep—all these points must be noted, and their influence, direct or indirect, upon the water carefully borne in mind.
When the sample has been duly collected, sealed, and a label affixed bearing the date, time, and conditions of collection and full address, it should be transmitted with the least possible delay to the laboratory. Frequently it is desirable to pack the bottles in a small ice case for transit. On receipt of such a sample of water the examination must be immediately proceeded with, in order to avoid, as far as possible, the fallacies arising from the rapid multiplication of germs. Even in almost pure water, at the ordinary temperature of a room, Frankland found organisms multiplied as follows:—
| Hours. | No. of Germs per cc. |
| 0 | 1,073 |
| 6 | 6,028 |
| 24 | 7,262 |
| 48 | 48,100 |
Another series of observations revealed the same sort of rapid increase of bacteria. On the date of collection the micro-organisms per cc. in a deep-well water (in April) were seven. After one day's standing at room temperature the number had reached twenty-one per cc. After three days under the same conditions it was 495,000 per cc. At blood-heat the increase would, of course, be much greater, as a higher temperature is more favourable to multiplication. But this would depend upon the degree of impurity in the water, a pure water decreasing in number on account of the exhaustion of the pabulum, whereas, for the first few days at all events, an organically polluted water would show an enormous increase in bacteria.
Furthermore, it is desirable to remember that organisms, in an ordinary water, do not continue to increase indefinitely. There is a limit to all things, even to numbers in bacteriology. Cramer, of Zurich, examined the water of the Lake after it had been standing for different periods, with the following results:—
| Hours and Days of Examination. |
No. of Micro-organisms per cc.12 |
|
| 0 | hours | 143 |
| 24 | " | 12,457 |
| 3 | days | 328,543 |
| 8 | " | 233,452 |
| 17 | " | 17,436 |
| 70 | " | 2,500 |
The writer's own experience is entirely in agreement with this cessation of multiplication at or about the end of a week, and the later decline.
Method of Examination. At the outset of a systematic study of a water it is well to observe its physical characters. The colour, if any, should be noted. Suspended matter and 40deposit may indicate organic or inorganic pollution. If abundant or conspicuous, a microscopic examination of the sediment may be made. The reaction, whether acid, neutral, or alkaline, must be tested, and the exact temperature taken. Any and every fact will help us, perhaps not so much to determine the contents of the water as to interpret rightly the facts we deduce from the entire examination.
Levelling Apparatus for Koch's Plate
At the beginning of the bacteriological work the water should be examined by means of the gelatine plate method. This consists in drawing up into a fine sterilised pipette a small quantity of the water and introducing it thereby into a test-tube of melted gelatine at a temperature below 40° C.13 It will depend upon the apparent quality of the water as to the exact quantity introduced into the gelatine; about .5 or .1 of a cubic centimetre is a common figure. The stopper is then quickly replaced in the test-tube, and the contents gently mixed more or less equally to distribute the one-tenth cubic centimetre throughout the melted gelatine. A sterilised sheet of glass (4 inches by 3) designated a Koch's plate is now taken and placed upon the stage of a levelling apparatus, which holds iced water in a glass jar under the stage. The gelatine is now poured out over the glass plate, and by 41means of a sterilised rod stroked into a thin, even film all over the glass. It is then covered with a bell-jar and left at rest to set. The level stage prevents the gelatine running over the edge of the plate; the iced water under the stage expedites the setting of the gelatine into a fixed film. When it is thus set the plate is placed upon a small stand in a moist chamber, and the whole apparatus removed to the room temperature incubator. A moist chamber is a glass dish, in which some filter paper, soaked with corrosive sublimate, is inserted, and the dish covered with a bell-jar. By this means the risks of pollution are minimized, and moisture maintained. In all cases at least two plates must be prepared of the same sample of water, and it is often advisable to make several. They may be made with different media for different purposes, and with different quantities of water, though the same method of procedure is adopted. In a highly polluted water extremely small quantities would be taken, and, vice versâ, in pure water a large quantity.
Moist Chamber in which Koch's Plates are Incubated
When we come to discuss the relation of disease organisms to water, particularly those causing typhoid fever, we shall learn that they are both scarce and intermittent. This point has been dwelt upon frequently by Dr. Klein, and it is clear that such a state of things greatly enhances the difficulties in detecting such bacteria, and he has proposed a simple procedure by which the difficulty of finding the Bacillus typhosus in a large body of water may be met.

Hot Air Steriliser
For the Sterilization of Glass Apparatus, etc.
One or two thousand cubic centimetres of the water under examination are passed through a sterilised Berkefeld filter by means of siphon action or an air-pump. The candle of the filter retains on its outer surface all, or nearly all, the particulate matter contained in the water. The matter thus retained on this outer surface is brushed by means of a sterile brush into 10 or 20 cc. of sterilised water. Thus we have all the organisms contained in two litres of the water reduced into 10 cc. of water. From this, so to speak, con43centrated emulsion of the bacteria of the original water, phenol-gelatine plates or Eisner plates (both acid media) may be readily made. In this way we not only catch many bacteria which would evade us if we were content with the examination merely of a few drops of the water, but we eliminate by means of the acid those common water bacteria, like Bacillus fluorescens liquefaciens, which so greatly confuse the issue.

In the course of two or three days the film of gelatine on the plate becomes covered with colonies of germs, and the next step is to examine these quantitatively and qualitatively. We may here insert a simple scheme by which this may be most fully and easily accomplished:—
1. Naked-Eye Observation of the Colonies. By this means44 at the very outset certain facts may be obtained, viz., the size, elevation, configuration, margin, colour, grouping, number, and kinds of colonies, all of which facts are of importance, and assist in final diagnosis. Moreover, in the case of gelatine plates (it is otherwise in agar) one is able to observe whether or not there is present what is termed liquefaction of the gelatine. Some organisms produce in their development a peptonizing ferment which breaks down gelatine into a fluid condition. Many have not this power, and hence the characteristic is used as a diagnostic feature.
2. Microscopic Examination of Colonies, which confirms or corrects that which has been observed by the naked eye. Fortunately some micro-organisms when growing in colonies produce cultivation features which are peculiar to themselves (especially is this so when growing in test-tube cultures), and in the early stages of such growths a low power of the microscope or magnifying glass facilitates observation.
3. Make cover-glass preparations: (a) unstained—"the hanging drop"; (b) stained—single stains, like gentian-violet, methyl blue, fuchsin, carbol fuchsin, etc.; double stains—Gram's method, Ziehl-Neelsen's method, etc.
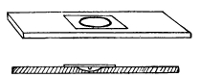
The Hanging Drop
This third part of the investigation is obviously to prepare specimens for the microscope. "The hanging drop" is a simple plan for securing the organisms for microscopic examination in a more or less natural condition. A hollow ground slide, which is a slide with a shallow depression in it, is taken, and a small ring of vaseline placed round the edge45 of the depression. Upon the under side of a clean cover-glass is placed a drop of pure water, and this is inoculated with the smallest possible particle taken from one of the colonies of the gelatine plate on the end of a sterilised platinum wire. The cover-glass is then placed upon the ring of vaseline, and the drop hangs into the space of the depression. Thus is obtained a view of the organisms in a freely moving condition, if they happen to be motile bacteria. As a matter of practice the hollow slide may be dispensed with, and an ordinary slide used.
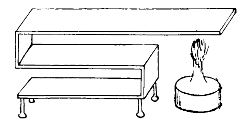
Drying Stage for Fixing Films
With regard to staining, it will be undesirable here to dwell at length upon the large number of methods which have been adopted. The "single stain" may be shortly mentioned. It is as follows: A clean cover-glass is taken (cleaned with nitric acid and alcohol, or bichromate of potash and alcohol), and a drop of pure sterilised water placed upon it. This is inoculated with the particle of a colony on the end of a platinum needle, and a scum is produced. The film is now "fixed" by slowly drying it over a flame. When the scum is thus dried, a drop of the selected stain (say gentian-violet) is placed over the scum and allowed to remain for varying periods: sarcinæ about thirty seconds; for many of the bacilli three or four minutes. It is then washed off with clean water, dried, and mounted in Canada balsam. The organisms will now appear under the microscope as violet in colour, and will thus be clearly seen.
The "double staining" is adopted when we desire to46 stain the organisms one colour and the tissue in which they are situated a contrast colour. Some of the details of these methods are mentioned in the Appendix.
4. Sub-culture. The plate method was really introduced by Koch in order to facilitate isolation of species. In a flask it is impossible to isolate individual species, but when the growth is spread over a comparatively large area, like a plate, it is possible to separate the colonies, and this being done by means of a platinum wire, the colonies may be replanted in fresh media; that is to say, a sub-culture may be made, each organism cultivated on its favourite soil, and its manner of life closely watched. We have already mentioned the chief media which are used in the laboratory, and in an investigation many of these would be used, and thus pure cultures would be obtained. Let us suppose that a water contains six kinds of bacteria. On the plate these six kinds would show themselves by their own peculiar growth. Each would then be isolated and placed in a separate tube, on a favourite medium, and at a suitable temperature. Thus each would be a pure culture; i. e., one and only one, species would be present in each of the six tubes. By this simple means an organism can be, we say, cultivated, in the same sort of way as in floriculture. From day to day we can observe the habits of each of our six species, and probably at an early stage of their separated existences we should be able to diagnose what species of bacteria we had found in the water. If not, further microscopic examination could be made, and, if necessary, secondary or tertiary sub-cultures.
5. Inoculation of Animals. It may be necessary to observe the action of supposed pathogenic organisms upon animals. This is obviously a last resource, and any abuse of such a process is strictly limited by law. As a matter of fact, an immense amount of bacteriological investigation can be carried on without inoculating animals; but, strictly47 speaking, as regards many of the pathogenic bacteria, this test is the most reliable of all. Nor would any responsible bacteriologist be justified in certifying a water as healthy for consumption by a large community if he was in doubt as to the disease-producing action of certain contained organisms.
Types of Liquefaction of Gelatine
By working through some such scheme as the above we are able to detect what quantity and species of organisms, saprophytic or parasitic, a water or similar fluid contains. For, observe what information we have gained. We have learned the form (whether bacillus, micrococcus, or spirillum), size, consistence, motility, method of grouping, and staining reactions of each micro-organism; the characters of its culture, colour, composition, presence or absence of liquefication or gas formation, its rate of growth, smell, or reaction; and lastly, when necessary, the effect that it has upon living tissues. Here, then, are ample data for arriving at a satisfactory conclusion respecting the qualitative estimation of the suspected water.
As to to the quantitative examination, that is fulfilled by counting the number of colonies which appear, say by the third and fourth day, upon the gelatine plates. Each colony has arisen, it is assumed, from one individual, so that if we count the colonies, though we do not thereby know how many organisms we have upon our plate, we do know approximately how many organisms there were when the plate was first poured out, which are the figures we require, and which can at once be multiplied and returned as so many organisms per cubic centimetre. There is, unfortunately, at present no exact standard to which all bacteriologists may refer.
Miquel and Crookshank have suggested standards which allow "very pure water" to contain up to 100 micro-organisms per cc. Pure water must not contain more than 1000, and water containing up to 100,000 bacteria per cc. is contaminated with surface water or sewage. Macé gives the following table:
| Very pure water | 0- | 10 bacteria per cc. |
| Very good water | 20- | 100bac"eria per"eri |
| Good water | 100- | 200bac"eria per"eri |
| Passable (mediocre) water | 200- | 500bac"eria per"eri |
| Bad water | 500- | 1,000bac"eria per"eri |
| Very bad water | 1000- | 10,000 and overeri"eri |
Koch first laid emphasis on the quantity of bacteria present as an index of pollution, and whilst different authorities have all agreed that there is a necessary quantitative limit, it has been so far impossible to arrive at one settled standard of permissible impurity.
Besson adopts the standard suggested by Miquel, and, on the whole, French bacteriologists follow suit. They also agree with him, generally speaking, in not placing much emphasis upon the numerical estimation of bacteria in water. In Germany and England it is the custom to adopt a stricter limit. Koch in 1893 fixed 100 bacteria per cc. as the maximum number of bacteria which should be present in a49 properly filtered water. Hence the following has been recognised more or less as the standard:
| 0- | 100 bacteria per cc. = | a good potable water, |
| 100- | 500bact"erieri"teria= | a suspicious water. |
| 500- | 1000 or moreeri"teria= | a water which should have further filtration before being used for drinking purposes. |
The personal view of the writer after some experience of water examination would favour a standard of "under 500" being a potable water, if the 500 were of a nature indicating neither sewage pollution nor disease. Miquel holds that not more than ten different species of bacteria should be present in a drinking water, and such is a useful standard. The presence of rapidly liquefying bacteria associated with sewage or surface pollution would, even though present in fewer numbers than a standard, condemn a water. Thus it will be seen that it is impossible to judge alone by the numbers unless they are obviously enormously high.
Wolfhügel's Counter
When we are counting colonies upon a Koch's plate, Wolfhügel's counter may be used. This is a thin plate of glass a size larger than Koch's plates, and upon it are scratched squares, each square being divided into nine smaller squares. The Wolfhügel plate is superimposed50 upon the Koch's plate, and the colonies counted in one little square or set of squares and multiplied.

Petri's Dish
By using flat, shallow, circular glass dishes, generally known as Petri's dishes, instead of Koch's plates, much manipulation and time is saved, and, on the whole, less risk of pollution occurs. Moreover, these are easily carried about and transferred from place to place. When counting colonies in a Petri's dish it is sufficient to divide the circle into eight equal divisions, and counting the colonies in the average divisions, multiply and reduce to the common denominator of one cc. For example, if the colonies of the plate appear to be distributed fairly uniformly we count those in one of the divisions. They reach, we will suppose, the figure of 60; 60 × 8=480 micro-organisms in the amount taken from the suspected water and added to the melted gelatine from which the plate was made. This amount was .25 cc. Therefore we estimate the number of micro-organisms in the suspected water as 60 × 8=480 × 4= 1920 m.-o. per cc., which is over standard by about 1500. A water might then be condemned upon its quantitative examination alone or qualitative alone, or both. If the quantity were even that of an artesian well, say 4–10 m.-o. per cc., but those four or ten were all Bacillus typhosus, it would clearly be condemned on its quality, though quantitatively it was an almost pure water. If, on the contrary, the water contained 10,000 m.-o. per cc., and none of them disease-producing, it would still be condemned on the ground that so large a number of organisms indicated some kind of organic pollution to supply pabulum for so many organisms to live in one cc. of the water. It is not the number per se which condemns. The large51 number condemns because it indicates probable pollution with surface water or sewage in order to supply pabulum for so many bacteria per cc.
It should always be remembered that a chemical report and a bacteriological report should assist each other. The former is able to tell us the quantity of salts and condition of the organic matter present; the latter the number and quality of micro-organisms. Neither can take the place of the other and, generally speaking, both are more or less useless until we can learn, by inspection and investigation of the source of the water, the origin of the organic matter or contamination. Hence a water report should contain not only a record of physical characters, of chemical constituents, and of the presence or absence of micro-organisms, injurious and otherwise, but it should also contain information obtained by personal investigation of the source. Only thus can a reasonable opinion be expected. Moreover, it is generally only possible to form an accurate judgment of a water from watching its history, that is, not from one examination only, but from a series of observations. A water yielding a steady standard of bacterial contents is a much more satisfactory water, from every point of view, than one which is unstable, one month possessing 500 bacteria per cc. and another month 5000. It is obvious that rainfall and drought, soil and trade effluents, will have their influence in materially affecting the bacterial condition of a water.
It is perhaps scarcely necessary to add that we have not
in the above account of the examination of water included
all, or nearly all, the various methods adopted for acquiring
a knowledge of the bacterial contents of the water. Many of
these are of too detailed and technical a nature to enter into
here. Three points, however, we may touch upon. In the
first place, as we have said, the particulate matter out of a
large body of water should be concentrated in a small
quantity. Accordingly it has become the custom to pass52
2000 or 3000 cc. of the suspected water through a Berkefeld
filter. When this has been accomplished, by means of a
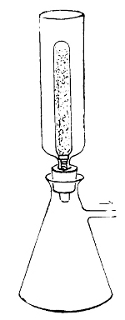 Berkefeld Filter
Berkefeld Filter
In Position for Filtration
of Water to
be Examined.
sterile brush the particulate matter on the
candle of the filter is brushed off into 10 or
15 cc. of sterilised water. This simple arrangement
is analogous to the use of gravity
or centrifugal methods of securing the solid
matter in milk. The smaller quantity of
water is then readily examined, and scanty
germs more readily detected. A second
point elaborating the scheme of water examination
is the choice of media for sub-culturing.
Mere examination on gelatine is not
sufficient. Even in making the primary
plate cultivations it is well to vary the media—agar,
carbol-gelatine, Elsner, etc. But
when colonies have appeared upon these
plates it is important to sub-culture with
accuracy and good judgment upon all or any
media—gelatine, agar, broth, potato, milk,
blood serum, glucose agar, glycerine agar,
etc.—that will reveal the real characters of
the bacteria present. A method proposed
by Professor Sheridan Delépine is to place
some of the suspected water in sterilised
test-tubes without further treatment, and incubate at 37°
C. for twelve or eighteen hours, and then plate out and
estimate the number of bacteria as in the ordinary course.
"In polluted water, containing an excess of organic matter,"
he says, "an extremely rapid multiplication of bacteria is
observed. In unpolluted water, containing only water bacteria
and a very small amount of organic matter, very little or no
multiplication takes place, and the growth of the water bacteria
liquefying gelatine is checked to a remarkable extent."
Thirdly, by none of these methods should we be able to
53isolate anaërobic bacteria, and to furnish a complete report
these also must receive careful attention.

Apparatus for Filtering Water to Facilitate its Bacteriological Examination
The Bacteriology of Water. In many natural waters there will be found varied contents even in regard to flora alone: algæ, diatoms, spirogyræ, desmids, and all sorts of vegetable detritus. Many of these organisms are held responsible for divers disagreeable tastes and odours. The colour of a water may also be due to similar causes. Dr. Garrett, of Cheltenham, has recorded the occurrence of redness of water owing to a growth of Crenothrix polyspora, and many other similar cases make it evident that not unfrequently great changes may be produced in water by contained microscopic vegetation.
With the exception of water from springs and deep wells,
all unfiltered natural waters contain numbers of bacteria.
The actual number roughly depends upon the amount of organic
pabulum present, and upon certain physical conditions
of the water. As we have already seen, bacteria multiply with
enormous rapidity. In some species multiplication does not
appear to depend on the presence of much organic matter,
and, indeed, some can live and multiply in sterilised water:
Micrococcus aquatilis and Bacillus erythrosporus. Again,
others depend not upon the quantity of organic matter, but
upon its quality. And frequently in a water of a high degree
of organic pollution it will be found that bacteria have
been restrained in their development by the competition of
other species monopolising the pabulum. Probably at least
one hundred different species of non-pathogenic organisms
have been isolated from water. Some species are constantly
occurring, and are present in almost all natural waters.
Amongst such are B. liquefaciens, B. fluorescens liq., B. fluorescens
non-liquefaciens, B. termo, B. aquatilis, B. ubiquitus,
and not a few micrococci, etc. Percy Frankland14 collected
water from various quarters at various times and seasons,
and some of his results may here be added:
RIVER THAMES WATER COLLECTED AT HAMPTON
Number of Micro-organisms Obtained from 1 cc. of Water.
| Month. | 1886. | 1887. | 1888. |
| January | 45,000 | 30,800 | 92,000 |
| February | 15,800 | 6,700 | 40,000 |
| March | 11,415 | 30,900 | 66,000 |
| April | 12,250 | 52,100 | 13,000 |
| May | 4,800 | 2,100 | 1,900 |
| June | 8,300 | 2,200 | 3,500 |
| July | 3,000 | 2,500 | 1,070 |
| August | 6,100 | 7,200 | 3,000 |
| September | 8,400 | 16,700 | 1,740 |
| October | 8,600 | 6,700 | 1,130 |
| November | 56,000 | 81,000 | 11,700 |
| December | 63,000 | 19,000 | 10,600 |
Again, another example:
RIVER LEA WATER COLLECTED AT CHINGFORD
Number of Micro-organisms Obtained from 1 cc. of Water.
| Month. | 1886. | 1887. | 1888. |
| January | 39,300 | 37,700 | 31,000 |
| February | 20,600 | 7,900 | 26,000 |
| March | 9,025 | 24,000 | 63,000 |
| April | 7,300 | 1,330 | 84,000 |
| May | 2,950 | 2,200 | 1,124 |
| June | 4,700 | 12,200 | 7,000 |
| July | 5,400 | 12,300 | 2,190 |
| August | 4,300 | 5,300 | 2,000 |
| September | 3,700 | 9,200 | 1,670 |
| October | 6,400 | 7,600 | 2,310 |
| November | 12,700 | 27,000 | 57,500 |
| December | 121,000 | 11,000 | 4,400 |
"During the summer months these waters are purest as
regards micro-organisms, this being due to the fact that
during dry weather these rivers are mainly composed of
spring water, whilst at other seasons they receive the washings
of much cultivated land."—Frankland.
Prausnitz has shown that water differs, as would be expected, according to the locality in the stream at which examination is made. His investigations were made from the river Isar before and after it receives the drainage of Munich:
| No. of Colonies per cc. |
|
| Above Munich | 531 |
| Near entrance of principal sewer | 227,369 |
| 13 kilometres from Munich | 9,111 |
| 22kilom"metres"muni" | 4,796 |
| 33kilom"metres"muni" | 2,378 |
Professor Percy Frankland also points out how the river Dee affords another example, even more perfect, of pollution and restoration repeated several times until the river becomes almost bacterially pure.
We cannot here enter more fully into the many conditions of a water which affect its bacterial content than to say that it varies considerably with its source, at different seasons, and under different climatic conditions. An enormous increase will occur if the sediment is disturbed, and conversely sedimentation and subsidence during storage will greatly diminish the numbers of bacteria. Sand filtration, plus a "nitrifying layer," will remove more than 90 per cent. of the bacteria. Sea-water contains comparatively few bacteria, and the deeper the water and the farther it is from shore so much less will be the bacterial pollution.
We will now consider several of the more important disease-producing bacteria found in water.
Bacillus Typhosus (Eberth-Gaffky). In 1880–81 Eberth announced the discovery of this bacillus in cases of clinical enteric fever. In 1884 it was first cultivated outside the body by Gaffky. Since then other organisms have been held56 responsible for the causation of enteric (or typhoid) fever. In 1885 the B. coli communis was recognised, and it has been a matter of great debate amongst bacteriologists as to how far these two organisms are the same species, and the typhoid germ merely a higher evolution of the B. coli. The differentiating signs between them will be referred to shortly. Bacteriologists generally regard the Eberth-Gaffky bacillus as the specific cause of the disease, though complete proof is still wanting.

Bacteria of Typhoid Fever
Microscopic Characters (in pure culture). Rods, 2–4 µ long, .5 µ broad, having round ends. Sometimes threads are observable, being 10 µ in length. In the field of the microscope the bacilli differ in length from each other, but are all the same thickness approximately. Round and oval cells constantly occur even in pure culture, and many of these shorter forms of typhoid are identical in morphology with some of the many forms of Bacillus coli. There are no spores. Motility is marked; indeed, in young culture it is the most active pathogenic germ we know. The small forms dart about with extreme rapidity; the longer forms move in a vermicular manner. Its powers of movement are due to some five to twenty flagella of varying length, some of them being much longer than the bacillus itself, though, owing to the swelling of the bacillus under flagellum-staining methods,57 it is difficult to gauge this exactly. The flagella are terminal and lateral, and are elastic and wavy. Their active contraction produces an evident current in the field of the microscope.
Cultures. This organism may be isolated from ulcerated Peyer's patches in the intestine, from the liver, the spleen, and the mesenteric glands. Owing to the mixture of bacteria found elsewhere, it is generally best to isolate it from the spleen. The whole spleen is removed, and a portion of its capsule seared with a hot iron to destroy superficial organisms. With a sterilised knife a small cut is made into the substance of the organ, and by means of a sterilised platinum wire a little of the pulp is removed and traced over the surface of agar. Agar reveals a growth in about twenty-four hours at 37° C., which is the favourite temperature. A greyish, moist, irregular growth appears, but it is invariably attached to the track of the inoculating needle. On gelatine the growth is much the same, but its irregular edge is, if anything, more apparent. There is no liquefaction and no gas formation. On plates of gelatine the colonies appear large and spreading, with jagged edges. The whole colony appears raised and almost limpet-shaped, with delicate lines passing over its surface. There is an appearance under a low power of transparent iridescence. The growth on potato is termed "invisible," and is of the nature of a potato-coloured pellicle, which looks moist, and may at a late stage become a light brown in colour, particularly if the potato is alkaline. Milk is a favourable medium, and is rendered slightly acid. No coagulation takes place. Broth is rendered turbid.
Micro-pathology. Typhoid fever is an infiltration and coagulation, necrosis, and ulceration of the Peyer's patches in the small intestine of man. The mesenteric glands show the same features, except that there is no ulceration. The spleen is enlarged, and contains the germs of the disease in almost a pure culture. The bacillus is present in the intestinal con58tents and excreta, particularly so when the Peyer's glands have commenced ulceration. In the blood of the general circulation the bacillus is not demonstrable, except in very rare instances. Typhoid fever is not, like anthrax, a blood disease.
COMPARATIVE FEATURES OF BACILLUS TYPHOSUS AND B. COLI
| B. TYPHOSUS | B. COLI |
Morphology: Cylindrical bacillus 2.4 µ, unequal lengths; some filaments. |
Shorter, thicker; filaments rare. |
Flagella: Long, wavy, spiral, and very numerous; movement very active. |
Shorter, stiffer, fewer; movement less active. |
On Gelatine and Agar: Angular, irregular, raised colonies; slow growth; translucent; medium remains clear. |
Even edge, homogeneous; much larger, quicker growth, and less translucent than B. typhosus; medium becomes turbid or coloured. |
In Gelatine: In ordinary gelatine and in sugar gelatine no gas is produced. |
Under the same circumstances abundant gas is produced. |
Milk: Not curdled by the bacillus. |
Milk is coagulated (within three days). |
Indol: The production of indol in ordinary broth is nil. |
Indol is present. |
Potato: The "invisible growth," if potato is acid. |
Thick, yellow growth. |
Lactose: Fermentation very slight. |
Fermentation marked. |
25 per cent. Gelatine at 37° C.: Strongly and uniformly turbid (Klein). |
Gelatine remains limpid and clear, but possesses thick pellicle. |
Elsner's Iodised Potato Gelatine: Slow growth; small, very transparent colonies. |
Very fast growth; larger, brown, less transparent colonies. |
Widal's Test: Bacilli become motionless and clumped together when suspended in a drop of blood serum from a typhoid patient. |
Bacilli remain actively motile. |
59Broth containing 0.3 per cent. Phenol or Formalin (1:7000): No growth. |
Grows well. |
Thermal Death Point: 62° C. for five minutes (Klein). |
66° C. for five minutes (Klein). |
Vitality in Water and Sewage: Typhoid bacillus soon ceases to multiply and readily dies (Klein). |
The B. coli retains for a much longer time its vitality and power of self-multiplication (Klein). |
The two species, Bacillus typhosus and B. coli, agree in possessing the following characters: no spores, no liquefaction of gelatine; both grow well on phenolated gelatine, and in Parietti's broth; both act similarly upon animals, though typhoid fever is not a specific disease of animals.
The Bacillus typhosus, though a somewhat susceptible bacillus, can when dried retain its vitality for weeks. In sewage it is very difficult indeed to detect, and is soon crowded out. Dr. Andrews and Mr. Parry Laws, in their bacterial researches into sewage for the London County Council,15 found that when they examined specially infected typhoid sewage it was only with extreme difficulty they isolated Eberth's bacillus. In ordinary sewage it is clear such difficulty would be greatly enhanced.

B. Coli Communis
We have pointed out elsewhere the relation between soil and typhoid. In water, even though we know it is a vehicle of the disease, the Bacillus typhosus has been only very rarely detected. The difficulties in separating the bacillus from waters (like that at Maidstone, for example), which appear definitely to have been the vehicle of the disease, are manifold. To begin with, the enormous dilution must be borne in mind, a comparatively small amount of contamination being introduced into large quantities of water. Secondly, the huge group of the B. coli species considerably complicates the issues, for it copiously accompanies the typhoid, and is always able to outgrow it. Further, we must bear in 60mind a point that is systematically neglected, namely, that the bacteriological examination of a water which is suspected of having conveyed the disease is from a variety of circumstances conducted too late to detect the causal bacteria. The incubation period of typhoid we may take at fourteen days. Let us suppose a town water supply is polluted with some typhoid excreta on the 1st of January. Until the 14th of January there may be no knowledge whatever of the state of affairs. Two or three days are required for notification of cases. Several more days elapse generally before bacteriological evidence is demanded. Hence arises the anomalous position of the bacteriologist who sets to work to examine a water suspected of typhoid pollution three weeks previously. There can be no doubt that these difficulties are very real ones. The solution to the problem will be found in Dr. Klein's dictum that "a water in which sewage organisms have been detected in large numbers should be regarded with suspicion"16 as the vehicle of typhoid, even though no typhoid bacilli were discoverable. The chief of these sewage bacteria are believed to be Proteus vulgaris, B. coli, P. zenkeri, and B. enteritidis, and they are all nearly related to B. typhosus. The presence of the B. coli in limited numbers is not suffi61cient to indicate sewage pollution, seeing that it is so widely distributed. But in large numbers, and in company with the other named species, it is almost certain evidence of sewage-polluted water.
It may occur to the general reader that, as the typhoid bacillus is not extremely rare, drinking water may frequently act as a vehicle to carry the disease to man. But, to appreciate the position, it is desirable to bear in mind the following facts: the typhoid bacillus is only found in the human excrement of patients suffering from the disease; it is short-lived; in ordinary waters there exist organisms which can exert an influence in diminishing its vitality; exposure to direct sunlight destroys it; and it has a tendency to be carried down-stream, or in still waters settle at the bottom by subsidence. Even when all the conditions are fulfilled, it must not be forgotten that a certain definite dose of the bacillus is required to be taken, and that by a susceptible person. Into these latter questions of how bacteria produce disease we shall have an opportunity of inquiring at a later stage.
We must now mention several of the special media and tests used in the separation of Bacillus typhosus and B. coli.
1. The Indol Reaction. Indol and skatol are amongst the final products of digestion in the lower intestine. They are formed by the growth, or fermentation set up by the growth, of certain organisms. Indol may be recognised on account of the fact that with nitrous acid it produces a dull red colour. The method of testing is as follows. The suspected organism is grown in pure culture in broth, and incubated for forty-eight hours at 37° C. Two cc. of a 4 per cent. solution of potassium nitrite are added to 100 cc. of distilled water, and about 1 cc. of this is added to the test-tube of broth culture. Now a few drops of concentrated sulphuric acid (unless quite pure, hydrochloric should be used) are run down the side of the tube. A pale pink to dull red colour62 appears almost at once, and may be accentuated by placing the culture in the blood-heat incubator for half an hour. Much dextrose (derived from the meat of the broth) inhibits the reaction. Bacillus typhosus does not produce indol, and therefore does not react to the test; B. coli and the bacillus of Asiatic cholera do produce indol, and react accordingly. It should be pointed out, however, that the bacillus of cholera also produces nitrites. Hence the addition of acid only to a peptone culture of cholera yields the "red reaction" of indol.
2. Carbolised Gelatine. To ordinary gelatine .05 per cent. of phenol is added. This inhibits many common water bacteria.
3. "_Shake Cultures._" To 10 cc. of melted gelatine a small quantity of the suspected organism is added. The test-tube is then shaken and incubated at 22° C. If the organism is Bacillus coli, the next day reveals a large number of gas-bubbles.
4. Elsner's Medium. This special potassium-iodide-potato-gelatine medium is used for the examination of typhoid excreta. It is made as follows: 500 grams of potato gratings are added to 1000 cc. of water; stand in cool place for twelve hours, and filter through muslin; add 150 grams of gelatine; sterilise and add enough deci-normal caustic soda until only faintly acid; add white of egg; sterilise and filter. Before use add half a gram of potassium iodide to every 50 cc. Upon this acid medium common water bacteria will not grow, but Bacillus typhosus and B. coli flourish.
5. Parietti's Formula consists of—phenol, five grams; hydrochloric acid, four grams; distilled water, 100 cc. To 10 cc. of broth 0.1–0.3 cc. of this solution is added. The tube is then incubated in order to see if it is sterile. If that is so, a few drops of the suspected water are added, and the tube reincubated at 37° C. for twenty-four hours. If the water contains the B. typhosus or B. coli, the tube will show a turbid growth.
6. Widal's Reaction. Mix a loopful of blood from a patient suspected of typhoid fever with a loopful of young typhoid broth culture in a hanging drop on a hollow ground slide. Cover with a cover glass and examine under 1/6-inch objective. If the patient is really suffering from typhoid, there will appear in the hanging drop two marked characteristics, viz., agglutination and immotility. This aggregation, together with loss of motility, is believed to be due to the inhibitory action of certain bacillary products in the blood of patients suffering from the disease. The test may be applied in various ways, and its successful issue depends upon one or two small points in technique into which we cannot enter here, but which the reader will find dealt with in the appendix.
7. Flagella-staining. Special methods must be adopted for staining the flagella of Bacillus typhosus and B. coli. The cover glasses should be absolutely clean, the cultures young (say eighteen hours old), and a diluted emulsion with distilled water must be made in a watch-glass in order to get bacilli discrete and isolated enough. Van Ermengem's Method is as follows:—Place a loopful of the emulsion on a clean cover glass and dry it in the air, fixing it lastly by passing it once or twice through the flame of a Bunsen burner. Place films for thirty minutes in a solution of one part boric acid (2 per cent.) and two parts of tannin (15.25 per cent.), which also contains four or five drops of glacial acetic acid to every 100 cc. of the mixture. Wash in distilled water and alcohol. Then place for five to ten seconds in a 25.5 per cent. solution of silver nitrate. Immediately thereafter, and without washing, treat the cover glass to the following solution for two or three seconds: gallic acid, five grams; tannin, three grams; fused potassium acetate, ten grams; distilled water, 350 cc. After this place in a fresh capsule of silver nitrate until the film begins to turn black. Wash in distilled water, dry, and mount. The process contracts the bacilli somewhat, but the flagella stain well.
The Bacillus coli communis occupies such an important place in all bacteriological investigation that a few words descriptive of it are necessary in this place. The "colon bacillus," as it is termed, appears to be almost ubiquitous in distribution. The idea once held that it belonged exclusively to the alimentary canal or sewage is now discarded. It is one of the most widely distributed organisms in nature, though, as its name implies, its habitat is in the intestinal tract of man and animals. It is an aërobic, non-sporulating, non-liquefying bacillus, about .4 µ in thickness, and twice that measurement in length; hence it often appears oval or egg-shaped. Its motility is in varying degree, occasionally being as active as B. typhosus, but generally much less so. It possesses lateral flagella. On gelatine plates at 20° C. B. coli produces non-liquefying, greyish-white, round colonies; in a stroke culture on the same medium, a luxuriant greyish band, much broader and less restricted to the track of the needle than B. typhosus. In depth of medium or "shake" cultures there is an abundant formation of bubbles of gas (methane or carbon dioxide) in the medium. On potato it produces a light yellow, greasy growth, which must be distinguished from the growth of B. fluorescens liquefaciens, B. pyocyaneus, and several other species on the same medium. If the potato is old or alkaline, the yellow colour may not appear. Milk is curdled solid in from twenty-four to forty-eight hours, and a large amount of lactic acid produced. In broth it produces a uniform turbidity, with later on some sediment and a slight pellicle. It gives the reaction to indol.
It is now the practice to speak of the family of Bacillus coli rather than the individual. The family is a very large one, and shows throughout but few common characters. The morphology readily changes in response to medium, temperature, age, etc. Fermentation of sugar, coagulation of milk, or indeed the indol reaction cannot always be used65 as final tests as to whether or not the organism is B. coli, for unfortunately some members of the family do not show each of these three features. Most varieties, however, appear to show some motility, a small number of flagella, a typical growth on potato, and develop more rapidly on all media than B. typhosus. These characters, plus one or more of the three features above named, are diagnostic data upon which reliance may be placed.
Cholera. This word is used to cover more a group of diseases rather than one specific well-restricted disease. In recent years it has become customary to speak of Asiatic cholera and British cholera, as if indeed they were two quite different diseases. But, as a matter of fact, we know too little as yet concerning either form to dogmatise on the matter. Until 1884 practically nothing was known about the etiology of cholera. In that year, however, Koch greatly added to our knowledge by isolating a spirillum from the intestine and in the dejecta of persons suffering from the disease.
Cholera has its home in the delta of the Ganges. From this endemic area it spreads in epidemics to various parts of the world, often following lines of communication. It is a disease which is characterised by acute intestinal irritation, manifesting itself by profuse diarrhœa and general systemic collapse, with cramps, cardiac depression, and subnormal temperature. The incubation period varies from only a few hours to several days. In the intestine, and setting up its pathological condition, are the specific bacteria; in the general circulation their toxic products, bringing about the systemic changes. Cholera is generally conveyed by means of water.
The spirillum of Asiatic cholera (Koch, 1884) generally appears, in the body and in artificial culture, broken into elements known as "commas." These are curved rods with round ends, showing an almost equal diameter throughout, and sometimes united in pairs or even a chain (spirillum).66 The latter rarely occur in the intestine, but may be seen in fluid cultures. The common site for Koch's comma is in the intestinal wall, crowding the lumina of the intestinal glands, situated between the epithelium and the basement membrane, abundant in the detached flakes of mucous membrane, and free in the contents of the intestine. They do not occur in the blood, nor are they distributed in the organs of the body.
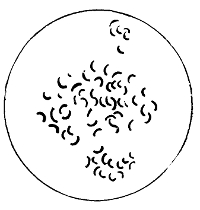
The Comma-Shaped Bacilli of Cholera
The bacilli are actively motile, and possess at least one terminal flagellum. The organism is aërobic, and liquefies gelatine. It stains readily with the ordinary aniline dyes. It does not produce spores, though certain refractile bodies inside the protoplasm of the bacillus in old cultures have been regarded as such. The virulence of the bacillus is readily attenuated, and both the virulence and morphology appear to show in different localities and under different conditions of artificial cultivation a large variety of what are termed involution forms. Unless the organism is constantly being sub-cultured, it will die. Acid, even the .2 per cent. present in the gastric juice, readily kills it. Desiccation, 55° C. for ten minutes, and weak chemicals have the same effect. The bacilli, however, have comparatively high powers of resistance to cold. Unless examined by the microscope 67in a fresh and young stage, it is difficult to differentiate Koch's comma from many other curved bacilli.
 |
 |
| Bacillus Typhosus (Showing Flagella) × 1000 |
Bacillus Typhosus (Widal Reaction) (Agglutination by serum from typhoid patient) × 400 |
 |
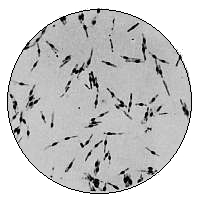 |
| Bacillus Coli Communis (From agar culture, 48 hours growth) × 1000 By permission of the Scientific Press, Limited |
Bacillus Mycoides (Spore formation. From agar culture) × 1000 |
Its cultivation characters are not always distinctive. Microscopically the young colonies in gelatine appear as cream-coloured, irregularly round, and granular. Liquefaction sets in on the second day, producing a somewhat marked "pitting" of the medium, which soon becomes reduced to fluid. In the depth of gelatine the growth is very characteristic. An abundant, white, thick growth exactly follows the track of the needle, here and there often showing a break in continuity. Liquefaction sets in on the second day, and produces a distinctive "bubble" at the surface. The liquefied gelatine does not fall from the sides of the tube, as in the Finkler-Prior comma of cholera nostras, but occurs inside the border where the gelatine joins the glass. In the course of a week or two all the gelatine may be reduced to fluid. On agar Koch's comma produces with rapidity a thick, greyish, irregular growth. On potato, especially if slightly alkaline, an abundant brownish layer is formed. Broth and peptone water are excellent media. In milk it rapidly multiplies, curdling the medium, with production of acid. Unlike Bacillus coli, it does not form gas, but, like B. coli, it produces large quantities of indol and a reduction of nitrates to nitrites. Hence the indol test may be applied by simply adding to the peptone culture several drops of strong sulphuric acid, when in the course of several hours, if not at once, there will be produced a pink colour, the "cholera red reaction." Although it readily loses virulence, and its resistance is little, the comma bacillus retains its vitality for considerable periods in moist cultures, upon moist linen, or in moist soil. In cholera stools kept at ordinary room temperature the cholera bacillus will soon be outgrown by the putrefactive bacteria. The same is true of sewage water.
The lower animals do not suffer from any disease at all68 similar to Asiatic cholera, and hence it is impossible to fulfil the postulate of Koch dealing with animal inoculation. In this respect it is like typhoid. It is, however, provisionally accepted that Koch's bacillus is the cause of the disease. The four or five other bacteria which have from time to time been put forward as the cause of cholera have comparatively little evidence in their support. It is less from these, and more from several spirilla occurring in natural waters, that difficulties of diagnosis arise.
Some hold that, however many comma bacilli be introduced into the alimentary canal, they will not produce the disease unless there is some injury or disease of the wall of the intestine. It need hardly be added that cholera acts, like other pathogenic bacteria, by the production of toxins. Brieger separated cadaverin and putrescin and other bodies from cholera cultures, and other workers have separated a tox-albumen.
Methods of Diagnosis of Cholera:
1. The nature of the evacuations and the appearance of the mucous membrane of the intestine afford striking evidence in favour of a positive diagnosis. Nevertheless it is upon a minute examination of the flakes and pieces of detached epithelium that reliance must be placed. In these flakes will be found in cholera abundance of bacilli having the size, shape, and distribution of the specific comma of cholera. The size and shape have been already touched upon. The distribution is frequently in parallel lines, giving an appearance which Koch described as the "fish-in-stream arrangement." This distribution of comma bacilli in the flakes of watery stools is, when present, so characteristic of Asiatic cholera that it alone is sufficient for a definite diagnosis. But unfortunately it is not always present, and then search for other characters must be made.
2. The appearance of cultivation on gelatine, to which reference has been made, is of diagnostic value.
3. The "cholera red reaction." It is necessary that the culture be pure for successful reaction.
4. Isolation from water is, according to Dr. Klein, best accomplished as follows: A large volume of water (100–500 cc.) is placed in a sterile flask, and to it is added so much of a sterile stock fluid containing 10 per cent. peptone, 5 per cent. sodium chloride, as will make the total water in the flask contain 1 per cent. peptone and .5 per cent. salt. Then the flask is incubated at 37° C. If there have been cholera vibrios in the water, however few, it will be found after twenty-four hours' incubation that the top layer contains actively motile vibrios, which can now be isolated readily by gelatine-plate culture.
5. To demonstrate in a rapid manner the presence of cholera bacilli in evacuations, even when present in small numbers, a small quantity must be taken up by means of a platinum wire and placed in a solution containing 1 per cent. of pure peptone and .5 per cent. sodium chloride (Dunham). This is incubated as in the case of the water, and in twelve hours is filled with a turbid growth, which when examined by means of the hanging drop shows characteristic bacilli.
We have already noticed that rivers purify themselves as they proceed. There are many excellent examples of this self-purification. The Seine as it runs through Paris becomes highly polluted with every sort of filthy contamination. But twenty or thirty miles below the city it is found to be even purer than above the city before it received the city sewage. In small rivers it is the same, provided the pollution is less in amount. Whilst authorities differ with regard to the mode of self-purification, all agree that in some way rivers receiving crude sewage are able in a marvellous degree to become pure again.
The conditions influencing this phenomenon are as follows:
(a) The Movement of the Water. It is probable, however, that any beneficial result accruing from this cause is due, not to any mechanical factor in the movement, but to the extra surface of water available for oxidation processes.
(b) The Pressure of the Water. It is believed that the volume of water pressing down upon any given area beneath it weakens the vitality of certain microbes. In support of this theory, it is urged that the number of bacteria capable of developing is less the greater the depth from the surface. Yet it must be remembered that mud at the bottom of a river, or at the bottom of the sea, is teeming with living organisms.
(c) Light. We have seen how prejudicial is light to the growth of organisms in culture media. This is so, though to a less extent, in water. Arloing held that sunlight could not pierce a layer of water an inch in thickness and still act inimically on micro-organisms. But Buchner found that the sun's rays could pass through fifteen or twenty inches and yet be bactericidal. This evidence appears contradictory. On the whole, however, authorities agree that the influence of the sun's rays upon water is distinctly bactericidal and causes a marked diminution in the quantity of organisms after acting for some hours. Especially will this be so when the water is spread over a wide area and is therefore shallow and stationary, or moving but slowly.
(d) Vegetation in Water. Pettenkofer, in his observations upon the Iser below Munich, has shown how algæ bring about a marked reduction in the organic matters present in water.
(e) Dilution. There can be no doubt in anyone's mind that the pollutions passing into a flowing river are very soon diluted with the large quantities of comparatively pure water always forthcoming. But this, whilst it would lower the percentage of impurity, cannot remove impurities.
(f) Sedimentation. Whilst Pettenkofer attributes self-purification to oxygenation and vegetation, most authorities are now agreed that it is largely brought about by the subsidence of impure matters, and by their subsequent disintegration at the bottom of the river. Sedimentation obviously is greatest in still waters. Hence lake water contains as a rule very few bacteria. "The improvement in water during subsidence is the more rapid and pronounced the greater the amount of suspended matter initially present" (Frankland). Tils has pointed out that the number of micro-organisms was invariably smaller in the water collected from the reservoir than in that taken from the source supplying the latter. Percy Frankland has demonstrated the same effect of sedimentation by storage as follows:
| No. of Colonies in 1 cc. of water. |
|
| 1. Intake from Thames, June 25, 1892 | 1,991 |
| 2. First small storage reservoir | 1,703 |
| 3. Second all st"orage re" | 1,156 |
| 4. Large storage reservoir | 464 |
The large reservoir would of course necessitate a prolonged subsidence, and hence a greater diminution than in the small reservoirs. Many like examples might be cited, but a typical one such as the above will suffice.
(g) Oxidation. Many experiments and observations have been made to prove that large quantities of oxygen are used up daily in oxygenising the Thames water. Oxygenated water will come up with the tide and down with the fresh water from above London. There will also be oxygen absorption going on upon the surface of the water, and from these three sources enough oxygen is obtained to oxidise impurities and produce what is really an effluent. In many smaller streams the opportunity for oxidation is afforded by weirs and falls.
Probably all these factors play a part in the self-purifica72tion of rivers, but we may take it that oxidation, dilution, and sedimentation are three of the principal agencies.
We may here digress to refer in passing to the facts obtainable from Sir Edward Frankland's report on Metropolitan water supply in 1894, as they will afford a connecting link between self purification and artificial purification. Judged by the relatively low proportion of carbon to nitrogen, the organic matter present in the water was, as usual, found to be chiefly, if not entirely, of vegetable origin. An immense destruction of bacteria was found to be effected by storage in subsidence reservoirs. The bacterial quality of the water might differ widely from its chemical qualities. These three facts are of primary importance in the interpretation of water reports, and it will be well to bear them in mind. Sir E. Frankland also refers to the physical conditions affecting microbial life in river waters. The importance of changes of temperature, the effect of sunlight, and rate of flow had been referred to in previous reports. Respecting the relative proportion of these factors, he adds:
"The number of microbes in Thames water is determined mainly by the flow of the river, or, in other words, by the rainfall, and but slightly, if at all, by either the presence or absence of sunshine, or a high or low temperature. With regard to the effect of sunshine, the interesting researches of Dr. Marshall Ward leave no doubt that this agent is a powerful germicide, but it is probable that the germicidal effect is greatly diminished, if not entirely prevented, when the solar rays have to pass through a comparatively thin stratum of water before they reach the living organisms."
From which it is clear that evidence favours the effect of sedimentation and dilution. These two factors in conjunction with filtration are, practically speaking, the methods of artificial water purification, with which we are now in a position to deal.
Sedimentation and Precipitation. Naturally, we see this factor in action in lakes or reservoirs. For example, the water supply of Glasgow is the untreated overflow from Loch Katrine. Purification has been brought about by means of subsidence of impurities. Nothing further is needed. Artificially, we find it is this factor which is the mechancial purifier of biological impurity in such methods as Clark's process. By this mode "temporary hardness," or that due to soluble bicarbonate of lime, is converted into insoluble normal carbonate of lime by the addition of a suitable quantity of lime-water. Carbonates of lime and magnesia are soluble in water containing free carbonic acid, but when fresh lime is added to such water it combines with the free CO2 to form the insoluble carbonate, which falls as a sediment:
CaCO3 + CO2 + CaH2O2 (lime-water) = 2 CaCO3 + H2O.
As the carbonate falls to the bottom of the tank it carries down with it the organic particles. Hence sedimentation is brought about by means of chemical precipitation. It is obviously a mechanical process as regards its action upon bacteria. Nevertheless its action is well-nigh perfect, and 300 or 400 m.-o. per cc. are reduced to 4 or 5 per cc. We shall refer to this same action when we come to speak of bacterial purification of sewage. Alum has been frequently used to purify waters which contain much suspended matter. Five or six grains of alum are added to each gallon of water, with some calcium carbonate by preference. Precipitation occurs, and with it sedimentation of the bacteria, as before. But, as Babes has pointed out, alum itself acts inimically on germs; in such treatment, therefore, we get sedimentation and germicidal action combined.
As a matter of actual practice, however, sedimentation74 alone is rarely sufficient to purify water. It is true that the collection of water in large reservoirs permits subsidence of suspended matters, and affords time for the action of light and the competitive suicidal behaviour of the common water bacteria. Yet, after all, filtration is the most important and most reliable method.
Sand Filtration, as a means of purifying water, has been practised since the early part of the present century. But it was not till 1885 that Percy Frankland first demonstrated the great difference in bacterial content between a water unfiltered and a water which had passed through a sand filter. Previous to this time the criterion of efficiency in water purification had been a chemical one only, and the presence or absence of bacteria in any appreciable quantity was described, not in mathematical terms, but in indefinite descriptive words, like "turbid," "cloudy," etc. It is needless to say that this difference in estimation was due to the introduction by Koch of the gelatine-plate method of examination. As a result of Percy Frankland's work, he formulated the following conclusions as regards the chief factors influencing the number of microbes passing through the filter.
It depends upon:
(1) The Storage Capacity for Unfiltered Water. This, of course, has reference to the advantages, which we have noticed above, of securing a large collection of water previous to filtration for subsidence, etc.
(2) The Thickness of Fine Sand through which Filtration is Carried on. An argument needing no further support, for it is clear, other things being equal, the more sand water passes through the greater the opportunity of leaving its impurities behind.
(3) Rate of Filtration. The slower filtration will be generally the more complete in its results.
(4) Renewal of Filter-Beds. After a certain time the filter-bed becomes worn out and inefficient; at such times renewal75 is necessary. Not only may the age of the filter act prejudicially, but the extra pressure required will tend to force through it bacteria which ought to have remained in the filter.
In 1893, Koch brought out his monograph upon Water Filtration and Cholera, and his work had a deservedly great influence upon the whole question. He shows how the careful filtration of water supplied to Altona from the Elbe saved the town from the epidemic of cholera which came upon Hamburg as a result of drinking unfiltered water, although Altona is situated several miles below Hamburg, and its drinking water is taken from the river after it has received the sewage of Hamburg. Now, from his experience of water filtration, Koch arrived at several important conclusions. In the first place, he maintained that the portion of the filter-bed which really removed micro-organisms effectively was the slimy organic layer upon the surface. This layer is produced by a deposit from the still unpurified water lying immediately above it. The most vital part of the filter-bed is this organic layer, which, after formation, should not be disturbed until it requires removal owing to its impermeability. A filter-bed, as is well known, consists of say three feet of sand and one foot of coarse gravel. The water to be filtered is collected into large reservoirs, where subsidence by gravitation occurs. Thence it is led by suitable channels to the surface of the filter-bed. Having passed through the three or four feet of the bed, it is collected in a storage reservoir and awaits distribution. The action of the whole process is both mechanical and chemical. Mechanically by subsidence, much suspended matter is left behind in the reservoir. Again, mechanically, much of that which remained suspended in the water when it reached the filter-bed is waylaid in the substance of the sand and gravel of the filter-bed. Chemically also the action is twofold. Oxidation of the organic matter occurs to some extent as the water passes through the sand. Until recently this chemical action76 and the double mechanical action were believed to be the complete process, and its efficiency was tested by chemical oxidation and alteration, and absence of the suspended matter.
Now, however, it is recognised that the second portion of the chemical action is vastly the more important, indeed, the only vital, part of the process. This is the chemical effect of the layer of scum and mud on the surface of the sand at the top of the filter-bed. The mechanical part of this layer is, of course, the holding back of the particulate matter which has not subsided in the reservoir; the vital action consists in what is termed nitrification of unoxidised substance, which is accomplished in this layer of organic matter. We shall deal at some length with the principles of nitrification when we come to speak of soil. But we may say here that by nitrification is understood a process of oxidation of elementary compounds of nitrogen, by which these latter are built up into stable bodies which can do little harm in drinking water. From what has been said it will be seen that the action of a filter-bed is of a complicated nature. There is (1) subsidence of the grosser particles of impurity in the water; (2) mechanical obstruction to impurities in the interstices of the scum, sand, and gravel in the filter; (3) oxidation of organic matter by the oxygen held in the pores of the sand and gravel; (4) nitrification in the vital scum layer, which is accomplished by micro-organisms themselves. This latter is now considered to be incomparably the most important part of the filter. That being so, its removal, except when absolutely necessary, is to be avoided as detrimental to the efficiency of the filter. New filters have obviously but little of this action. Hence it is wise to allow a new filter-bed to act for a short period (say twenty-four to forty-eight hours) before the filtered water is used for domestic purposes, in order to allow the organic layer to be formed. This must also be borne in mind after a filter-bed has been cleaned.
To maintain this nitrifying action of a filter in efficiency, Koch suggested, in the second place, that the rate of filtration must not exceed four inches per hour. At the Altona water-works this rate of filtration was maintained, and the number of organisms always remained below 100 per cc., which, as we have seen, is the standard. Thirdly, it is important that periodic bacteriological examinations should be made. Koch's emphasis upon this point is well known, and the cumulative experience of bacteriologists only further supports such a course being taken. If it be true that efficient sand filtration is a safeguard against pathogenic germs like typhoid and cholera, then there can be but one criterion of efficiency, viz., their absence in the filtered water, which can only be ascertained by regular examination. But it is not alone for pathogenic germs that filtration is proposed. Filtered water containing more than 100 micro-organisms of any kind per cc. is below the standard in purity, and should on no account be distributed for drinking purposes. In this country chemical analysis, with a more or less cursory microscopic examination, has been almost invariably accepted as reliable indication of the condition of the water. But such an examination is not really any more a fair test of the working of the filter than it is of the actual condition of the water. It is true, the quantity of organic matter can be estimated and the condition in which it exists in combination obtained; but it cannot tell us what a bacteriological examination can tell us, viz., the quantity and quality of living micro-organisms present in the water. Upon this fact, after all, an accurate conclusion depends. There is abundant evidence to show that no valuable opinion can be passed upon a water except by both a chemical and a bacteriological examination, and further by a personal investigation, outside the laboratory, of the origin of the water and its liabilities to pollution.
So convinced was Koch of the efficiency of sand filtration78 as protection against disease-producing germs that he advocated an adaptation of this plan in places where it was found that a well yielded infected water. Such pollution in a well may be due to various causes; surface-polluted water oozing into the well is probably the commonest, but decaying animal or vegetable matter might also raise the number of micro-organisms present almost indefinitely. Koch's proposal for such a polluted well was to fill it up with gravel to its highest water level, and above that, up to the surface of the ground, with fine sand. Before the well is filled up in this manner it must, of course, be fitted with a pipe passing to the bottom and connected with a pump. This simple procedure of filling up a well with gravel and sand interposes an effectual filter-bed between the subsoil water and any foul surface water percolating downwards. Such an arrangement yields as good, if not better, results than an ordinary filter-bed, on account of there being practically no disturbance of the bed nor injury done to it by frost.
The effect of the remedies we have been discussing upon the number of bacteria is demonstrated in the results which Sir Edward Frankland arrived at in his investigation of London waters.17
Mean of Monthly Examinations for the Year
| Name of Company. | Source of Supply |
M.-o. per cc. | Average per cent. of Micro-organisms Removed by Filtration. | ||||
| At Source. | After Storage. | After Filtration. | |||||
| The Chelsea Co. | Thames at Hampton | 16,138 | 1067 | 34 | 98.96 | ||
| West Middlesex Co. | " | 16,138 | 1788 | 58 | 99.40 | ||
| Southwark & Vauxhall Co. | " | 16,138 | .... | 80 | 97.72 | ||
| Grand Junction Co. | " | 16,138 | 2500 | |
623 100 96 |
 |
98.46 |
| Lambeth Co. | " | 16,138 | 7820 | 75 | 99.50 | ||
The teaching of these figures could, with great ease, be reproduced again and again if such was necessary; but these will suffice to show that sand filtration, when carefully carried out, offers a more or less absolute barrier to the passage of bacteria, whether non-pathogenic or pathogenic.
Domestic Purification of Water. Something may here be said, from a bacteriological point of view, relative to what is called domestic purification. There is but one perfectly reliable method of sterilising water for household use, viz., boiling. As we have seen, moist heat at the boiling point maintained for five minutes will kill all bacteria and their spores. The only disadvantages to this process are the labour entailed and the "flat" taste of the water. Nevertheless in epidemics due to bad water it is desirable to revert to this simple and effectual purification.
There are a large number of filters on the market with, in
many cases, but little modification from each other. The
materials out of which they are made are chiefly the following:
carbon and charcoal, iron (spongy iron or magnetic
oxide), asbestos, porcelain and other clays, natural porous
stone, and compressed siliceous and diatomaceous earths.
From an extended research in 1894 by Dr. Sims Woodhead
and Dr. Cartwright Wood our knowledge of the quality of
these substances as protectives against bacteria has been
largely increased. They concluded that a filter failed to act
in one of two ways. It was either pervious to micro-organisms,
or its power of filtering became modified owing to (a)
structural alteration of its composition, or to (b) the growing
through of the micro-organisms. The conditions which
chiefly influence the growth of bacteria through a filter appear
to be the temperature, the intermittent use of the filter,
and the species of bacteria. The higher the temperature
and the longer the organisms are retained in the filter
the more likely is it that they will grow through, and in
the next usage of the filter appear in the filtrate. As to the80
species, those multiplying rapidly and possessing the power
of free motility will naturally appear earlier in a filtrate
than others. Woodhead and Wood, from their searching
and most able investigation, concluded that the Pasteur-Chamberland
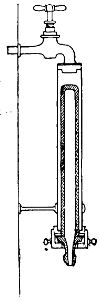 Pasteur-Chamberland
Filter
Pasteur-Chamberland
Filter
Attached to Water Supply
candle filters (composed of porcelain
formed by a mixture of kaolin and other
clays) were the only filters out of the substances
named above which were reliable and protective
against bacteria. They tested over three dozen
of the Pasteur filters, and "in every case these
gave a sterile filtrate." Pure cholera bacillus
in suspension (5000 bacilli to every cc.) and
typhoid bacillus in suspension (8000 per cc.)
were passed through these filters, and not a
single bacillus was detectable in the filtrate.
The Berkefeld filter (siliceous earth) came
second on the list as an effective filter, and had
but the fault of not being a "continuous"
steriliser. A certain Parisian filter ("Porcelaine
d'Amiante"), made of unglazed porcelain,
rendered water absolutely free from
bacteria. Its action was, however, very slow.
Setting aside these three efficient filters, we
are face to face with the fact that most filters
do not produce germ-free filtrates, even though
they are nominally guaranteed to do so. It is professed
for animal charcoal, which is widely used, that it absorbs
oxygen, and so fully oxidises whatever passes through it.
This may be so at first, but after a little use it probably
does more harm than good. It appears to add nitrogen
and phosphates to water, which are both nutritive substances
on which bacteria grow. Moreover it readily absorbs
impurities from the air. As a matter of experiment
and practice, it has been found by Frankland, Woodhead,
and others, that charcoal actually adds to the number of
germs after it has been in use for some days.
Diseases Conveyed by Water. There are a few preliminary features to be noticed before we enter in detail upon the characteristics of several of the chief pathogenic bacteria in water.
In sterilised water, and in very highly polluted water or sewage, pathogenic bacteria do not flourish. In the former case they die of starvation, although there are some experiments on record which appear not to support this view; in the latter case they are killed by the enormous competition of common bacteria. Even in ordinary water there is a wide divergence of behaviour. Some bacteria are destroyed in a few hours; others appear to flourish for weeks. In all cases the spores are able to resist whatever injurious properties the water may have much more persistently than the bacilli themselves. These changes in the vitality of bacteria in water, partly due to the water and partly to the other micro-organisms, bring about two characteristics which it is important to remember, viz., that pathogenic germs in water are, as a rule, scanty and intermittent. It is these features in conjunction with the enormous quantities of common water bacteria which make the search for the bacillus of typhoid what Klein has called "searching for a needle in a rick of hay." Not that it cannot be detected, but its detection is one of the most difficult of investigations. We shall refer to this matter again when Bacillus typhosus is under consideration.
In artificial cultivation water bacteria respond very readily to external conditions. Increase of alkalinity (.01 grams of sodium carbonate added to 10 cc. of ordinary gelatine) causes the number of colonies to be five or six times greater than that revealed by using ordinary gelatine; on the other hand, very slightly increasing the acidity of a medium as markedly diminishes the number of bacteria. Advantage is taken of this in culturing the bacillus of typhoid, which does not object to an acid medium.
Water may become polluted in a variety of ways, and it82 is helpful to classify these as pollutions at the source, in the course, and at the periphery. Gathering-grounds are frequently the locality of the pollution. The recent Maidstone epidemic is an example. Here some of the springs supplying the town with water were contaminated by several typhoid patients. Frequently on the gathering-ground one may find a number of houses the waste and refuse of which will furnish ample surface pollution, which in its turn may readily pass into a collecting reservoir or a well. Only recently the writer investigated the cause of typhoid in a large country house, and traced it to pollution of the private well by surface washings from the stable quarters. Leakage of house-drains into wells is not an infrequent source of contamination. The same cause is generally operative in cases of pollution of a water supply in its course from the source to the cisterns or taps at the periphery, viz., a sewer or drain leaking into the water supply.
Water companies and those responsible for water supply appear to hold the opinion that so long as there is sand filtration or subsidence reservoirs it is unnecessary to consider the gathering-ground or transit. But, as we have seen, a frost may completely dislocate the efficient action of a filter, and times of flood may prevent proper sedimentation; then our dependence for pure water is wholly upon the gathering-ground and source. Hence we find water contaminated at its source by polluted wells, by sewage-infected rivers and streams, by drainage of manured fields, by innumerable excremental pollutions over the areas of the gathering-grounds, and in transit by careless laying, poor construction, bad jointing, and close proximity of water-and drain-pipes. In the third place, we may get a water infected at the periphery, in the house itself. Such cases are generally due to one of two causes: filthy cisterns or suction. Cisterns per se are more or less indispensable where a constant service does not exist, but they should be inspected from time to83 time and maintained in a cleanly condition. Suction into the tap has been recently emphasised by Dr. Vivian Poore as a cause of pollution. It is liable to occur whenever a tap is left turned on, and a vacuum is produced in the supply-pipe by intermission of the water supply, so that foul gas or liquid is sucked back into the house-pipe.
One more point requires our attention. It has relation to bacterially polluted water when it has gained entrance to the body. It has been known for some time past that not all waters polluted with disease germs produce disease. As we have before said, this may depend upon the infective agent, its quantity and quality; the body being able in many cases to resist a small dose of poison. It is, however, necessary to infection, especially in water-borne disease, that the tissues shall be in some degree disordered. The perverted action of the stomach influences the acid secretion of the gastric juice, through which bacilli might then pass uninjured. Particularly must this be so in the bacillus of cholera, which is readily killed by the normal acid reaction of the stomach. Hence, in this disease at least, it is the opinion of bacteriologists that the condition of the mucous membrane of the stomach is of primary importance. Metschnikoff has indeed demonstrated the presence of the bacillus of cholera in the intestinal excretion of apparently healthy persons, which shows that they were protected by the resistance of their tissues to the bacilli. Further light has been thrown on this question by the researches of MacFadyen, who has pointed out that suspensions of cholera bacilli in water passed through the stomach untouched, and were thus able to exert their evil influence in other parts of the alimentary canal. When, however, cholera bacilli were suspended in milk, none appeared to escape the germicidal action of the gastric juice. The explanation of this is probably the simple one that the stomach reacted with its secretion of gastric juice only to food (milk), but simply passed84 the water on into the lower and more absorptive parts of the alimentary canal. Such a condition of affairs clearly increases the danger of water-borne germs.
It will not be needful to insist upon the obvious fact that bacteria abound in sewage. Such a large quantity of organic matter, in which decomposition is constantly taking place, will afford an almost ideal nidus for micro-organic life. There is indeed but one reason why such a medium is not absolutely ideal from the microbe's point of view, and that reason is, that in sewage the vast number of bacteria present make the struggle for existence exceptionally keen. Not only are the numbers incredibly large, but we also find a very extensive representation of species, including both saprophytes and parasites, non-pathogenic and pathogenic. Not infrequently it is from pollution by sewage that drinking water is contaminated with disease. A patient, we will say, suffers from typhoid fever. The specific organism has its habitat largely, though not exclusively, in the alimentary canal. It passes out in the excreta, and though sometimes partially disinfected, may escape without hindrance into the drains, and thus to the sewer or cesspool. How often, by means of direct connection or by percolation, sewage, from sewers or cesspools, gains access to drinking water, the history of typhoid outbreaks in this country only too fully records.
It is impossible to lay down any exact standard of the chemical and bacteriological quality of sewage. The quality will differ according to the size of the community, the inclusion or otherwise of trade-waste effluents, the addition of rain-water, and other like physical conditions. Moreover, sewage itself when, so to speak, fully formed is liable to undergo rapid changes owing to fermentation and the com85petition of micro-organisms. It is clear that these latter are the chief agents in bringing about the change, because, if sewage be placed in hermetically sealed flasks and sterilised by heat, it is found that no change occurs. From facts such as the above it will be apparent that no exact standard of chemical or bacterial contents is possible. Respecting the chemical condition we may shortly say that the chief characteristic of sewage is its enormous amount of contained organic matter in suspension or solution; respecting the bacterial content we may say that the chief species of the very numerous organisms are those commonly concerned in fermentative putrefaction. London crude sewage contains on an average about four millions of micro-organisms per cc. Many of these are "liquefying" bacteria; that is to say, they have the power of liquefying gelatine, which is generally one of the features of putrefactive species. In considering the quality of the bacteria present in sewage, a still wider field of research opens before us. For though we can say that, roughly, all sewage will contain probably between four and eight millions of bacteria, we cannot even lay down a rough standard respecting the kinds of bacteria present more than we have done already in stating that a very large number indeed out of the total will belong to putrefactive species.
We may, however, make a provisional list of normal sewage bacteria18 as follows:
1. Bacillus coli communis and all its varieties and allies.
2. Proteus vulgaris and the various protean species.
3. B. enteritidis sporogenes (Klein).
4. Liquefying bacteria, e. g., Bacillus fluorescens liquefaciens, B. subtilis, B. mesentericus.
5. Non-liquefying bacteria.
6. Sarcinæ, yeasts, and moulds.
 |
 |
Proteus Vulgaris |
B. Enteritidis Sporogenes |
We have not included in the above inventory any pathogenic bacteria. Doubtless such species (e. g., typhoid19) not infrequently find their way into sewage. But they are not normal habitants, and though they struggle for survival, the keenness of competition among the dense crowds of saprophytes makes existence almost impossible for them. Nor can they expect much sympathy from us in the difficulties of life which fortunately confront them in sewage.
Of those we have named as normally present it is unnecessary to speak in detail, with the exception of the newly 87discovered anaërobe, Bacillus enteritidis sporogenes of Klein.20 This bacillus is credited to be a causal agent in diarrhœa, and has been isolated by Dr. Klein from the intestinal contents of children suffering from severe diarrhœa, and from adults having cholera nostras. It has been readily detected in sewage from various localities, and also in sewage effluents, after sedimentation, precipitation, and filtration. Its biological characters are shortly as follows: It is in thickness somewhat like the bacillus of symptomatic anthrax, thicker and shorter than the bacillus of malignant œdema, and standing therefore between the latter and anthrax itself. It is motile and possesses flagella, but has no threads. It readily forms spores, which develop as a rule near the ends of the rods and are thicker than the bacilli. It is stained by Gram's method. In various media (particularly milk) it produces gas rapidly. It is an anaërobe, and is cultivated in Buchner's tubes. A recent epidemic of diarrhœa affecting 144 patients in St. Bartholomew's Hospital was traced to milk in which B. enteritidis was present.
Sewer Air. Though not of material importance as regards bacterial treatment of sewage, this subject calls for some remark. For long it has been known that air polluted by sewage emanations is capable of giving rise to various degrees of ill-health. These chiefly affect two parts of the body; one is the throat, and the other is the alimentary canal. Irritation and inflammation may be set up in both by sewer air. Such conditions are in all probability produced by a lowering of the resistance and vitality of the tissues, and not by either a conveyance of bacteria in sewer air or any stimulating effect upon bacteria exercised by sewer air. What evidence we have is against such factors. (See p. 105.)
Several series of investigations have been made into the 88bacteriology of sewer air, amongst others by Uffelmann, Haldane, Laws, and Andrewes. From their labours we may formulate four simple conclusions:
1. The air of sewers contains very few micro-organisms indeed, sometimes not more than two organisms per litre (Haldane), and generally fewer than the outside air (Laws and Andrewes).
2. There is no relationship between the microbes contained in sewer air and those contained in sewage. Indeed, there is a marked difference which forms a contrast as striking as it is at first sight unexpected. The organisms isolated from sewer air are those commonly present in the open air. Micrococci and moulds predominate, whereas in sewage bacilli are most numerous. Liquefying bacteria, too, which are common in sewage, are extremely rare in sewer air. Bacillus coli communis, which occurs in sewage from 20,000 to 200,000 per cc., is altogether absent from sewer air.
3. Pathogenic organisms and those nearly allied to them are found in sewage, but absent in sewer air. Uffelmann isolated the Staphylococcus pyogenes aureus (one of the organisms of suppuration), but such a species is exceptional in sewer air. Hence, though sewer air is popularly held responsible for conveying diphtheria and all sorts of other virulent bacteria, there is up to the present no evidence of a substantial nature in support of such views. Sewer air neither conducts pathogenic organisms nor stimulates the virulence of such.
4. Lastly, only when there is splashing in the sewage, or when bubbles are bursting (Frankland), is it possible for sewage to part with its contained bacteria to the air of the sewer.
Whilst we cannot here enter more fully into an account of the bacteria found in sewage or of their functions, it is necessary to remark upon one distinguishing feature. A very large number of sewage bacteria are decomposing and denitri89fying, that is to say, breakers down, by means of putrefaction, of organic compounds. The knowledge of this fact has recently been applied, in conjunction with oxidation, to the biological treatment of sewage. As this illustrates in a marked degree some of the facts we have dwelt upon in considering the bacteriology of soil, and as it is likely that the future will witness a still wider application of these same facts, it will be necessary to refer in some detail to the matter.
Hitherto there has been adopted one of four methods of treatment of sewage. In the first place, in towns situated on the coast the sewage has, by means of a conduit, been carried out to sea. It is clear that such a course, which is in itself open to criticism, is applicable to but few towns. In the second place, methods of chemical treatment have been practised. This has generally been of the nature of a "precipitation" process. Six to twelve grains of quicklime have been added to each gallon of sewage. The process is simple and cheap, but it does not remove the organic matter in solution. On the one hand, it does not produce a valuable manure; on the other, it fails to purify the effluent. A dozen other methods have been tried, but all based on the addition of chemical substances to precipitate or change the organic matter of the sewage. Electrolysis, too, has been proposed. The third mode adopted in the past has been that known as intermittent downward filtration. This may be defined as "the concentration of sewage at short intervals on an area of specially chosen porous ground, as small as will absorb and cleanse it, not excluding vegetation, but making the product of secondary importance" (Metropolitan Sewage Commission). The action is mechanical and biological, that is to say, due in part to nitrification by bacteria in the upper layers of soil. The fourth plan is that of irrigation, or "the distribution of sewage over a large surface of ordinary agricultural ground, having in view a maximum90 growth of vegetation (consistently with due purification) for the amount of sewage supplied." Like the former, there is biological influence at work here, though in a less degree. About one acre is required for every hundred persons in the population. These two latter modes are much to be preferred to chemical treatment, yet on account of space and management, as well as on account of the non-removal of the "sludge," their success has not been all that could be desired. Until comparatively recent times the above methods of treating sewage were the only ones available.
In 1881 it appears that M. Louis Mouras, of Vesoul (Haute Saône), published an account of a hermetically sealed, inodorous, and automatically discharging cesspool, in which sewage was anaërobically broken down by "the mysterious agents of fermentation." This is the first record we have of the newly applied treatment of sewage by simply allowing Nature to fulfil her function by means of bacteria. We shall most easily arrive at an appreciation of the recent developments of the process in England by describing the so called septic tank and cultivation beds.
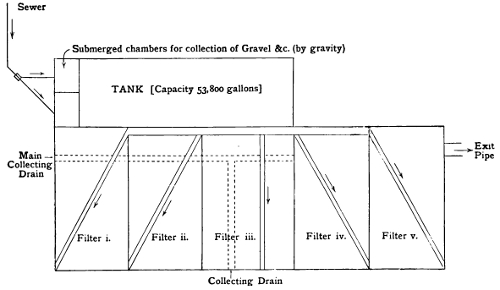
A Plan of Septic Tank and Filter-Beds
As Used at Exeter
The septic tank is a large underground vault of cemented
brick, having a capacity of thousands of gallons, according
to the population. That at Exeter has a capacity of 53,800
gallons, and takes the average sewage of 1500 inhabitants in
twenty-four hours. Near the entrance is a submerged wall,
seven feet from the entrance and twelve inches below the
surface of the liquid in the full tank. Within this are caught,
by gravity, gravel and such-like deposits. The remaining
solid matter of the sewage becomes deposited in the tank itself.
Both in the sediment at the bottom of the tank and in
the thick scum on the surface the organic compounds are
broken down and made soluble. In the former position this is
accomplished by anaërobic bacteria, in the latter on the surface
by aërobic bacteria. It need hardly be added that these
are denitrifying and putrefactive bacteria, and that those at
91
92the bottom of the tank perform greater service than those
at the top. When the liquid sewage passes out of the tank
it differs from the crude sewage which enters the tank in the
following particulars: (a) The gravel and particulate débris
have been removed; (b) the organic solids in suspension are
so greatly diminished that they are almost absent; (c) there
is an increase of organic matter in solution; (d) the sewage
is darker in colour and more opalescent; (e) compounds like
albuminoid ammonia, urea, etc., have been more or less
completely broken down, and reappear in elementary conditions,
like ammonia, methane, carbon dioxide, and sulphuretted
hydrogen. These latter bodies may be in solution
or may have escaped as gas.
The cultivation beds are four or five filters, to which the sewage from the tank flows in such a manner as to produce a weir. By an automatic arrangement the fluid is distributed to each filter in turn. When the second filter is full the first is discharged, and remains empty during the time that the third and fourth are being filled. Each filter is thus full, say, about six hours, and has from ten to twelve hours' rest. These filter-beds (at Exeter) have an area of eighty square yards and a depth of five feet; collecting drains are laid on the bottom of the filters, joining main collectors, the latter terminating in discharging wells. The filtrant is broken furnace clinker or broken coke.
The changes occurring in these filters are of the nature of oxidation, with the result that the proportion of the oxidised nitrogen increases (as nitrites and nitrates), the ammonia becomes less, and the total solids and organic nitrogen almost disappear. It will thus be seen that the work of these filters is not merely a straining action. It is true that particulate matter in the effluent from the tank is caught on the surface by the film (resulting from previous effluents), but the real work of the bed is nitrification, an oxidation of ammonia into nitrites and nitrates. This change obviously93 begins when the tank effluent flows over the "weir" on to the filter-beds, and the oxygen thus obtained by the effluent is carried down in solution into the coke-breeze. Upon the surface of the filtrant are oxidising bacteria. When the effluent is on the bed they oxidise its contained products; when the bed is empty and "resting" they oxidise carbon. An advantage arising from the periodical emptying and filling of the filter is that the products of decomposition which would eventually inhibit the action of the aërobic bacteria are washed away, and pass into the nearest stream, where they become absolutely innocuous.
The "filter" is more correctly termed a cultivation bed, for its purpose is to furnish a very large surface upon which the nitrifying organisms present, as we have seen, in all soils, may flourish, and thus feeding upon the organic matter of the sewage, may perform their function of oxidation.
It is not possible to lay down exact limits as to where denitrification ends and oxidation begins. To a certain extent, and in varying degree, they overlap each other. But roughly we may say that in the tank there is a breaking down (denitrification and decomposition) and in the filter-beds a building up (nitrification). The case is precisely parallel to similar changes occurring in soil, and which we have dealt with elsewhere. The advantage indeed of this biological treatment of sewage is that it exactly follows the processes of nature, in contradistinction to the mechanical and chemical methods hitherto adopted.
At Sutton and some other places the same principles are applied,—that is to say, bacterial filtration,—but there is no tank. A metal screen in some measure takes its place, and holds back solid matter from being carried on to the beds. The filtrant is burnt clay, and it is forked over occasionally to let in oxygen. The crude sewage is run over the top of the burnt ballast, where it is left for two or three hours. It is then slowly run off on to a finer filter, where it also94 stays two hours. Thence the effluent is run into the stream.
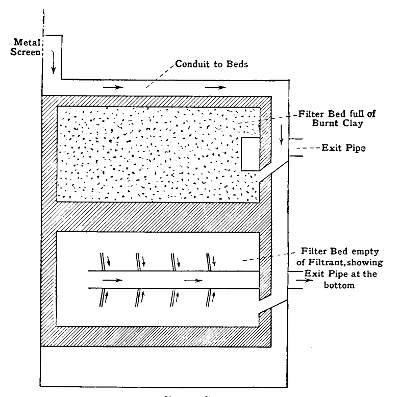
Filter-Beds
As Used at Sutton
It must be admitted that the bacterial treatment of sewage, though exhibiting such excellent results where it has been given a fair trial, is still in a probationary stage. It appears to stand on reason. The sludge of previous methods is avoided. The sewage is entirely broken down, and the effluent is a comparatively pure one, yet taking back nitrogen, as nitrate, to the soil. The whole change, indeed, in the opinion of Dr. Dupré, is more effective and radical than in chemical treatment. Further, it has been tested as regards95 its action upon the pathogenic bacilli—those of tubercle and typhoid—with the result that these infective bacteria have been completely destroyed. It appears that such destruction of infective germs occurs in the tank, and depends in degree upon the rapidity with which sewage is passed through the tank. The cultivation beds also have an inimical effect upon infective bacteria. Hence the final effluent is practically germ-free as regards pathogenic organisms.
The basis of the usual methods in practice is to pass air over or through some nutrient medium. By this means the contained organisms are waylaid, and finding themselves under favourable conditions of pabulum, temperature, and moisture, commence active growth, and thus reveal themselves in characteristic colonies. These are examined, as directed on page 43, by the microscope and sub-culture. Quantitative estimation is not generally made, as a fixed standard is even less a possibility than in milk and soil. Returns of the number of bacteria in the sample taken may be made for the sake of information, but little or no conclusion of value can be drawn from such data. The standard recognised in Europe is the cubic metre, and one may speak, for example, of the air of a room containing 500, 1000, or 3000 germs per cubic metre.
The following are the chief methods:
1. Pouchet's Aëroscope. This apparatus was in use some time ago in France, and by its means all the solid matter of a given quantity of air was drawn through an air-tight glass tube by aspiration and made to impinge upon a small plate of glycerine. The air escaped to the aspirator at the sides, leaving upon the glycerine plate only its particulate matter. This remnant could then be examined.
2. Koch adopted the simplest of all the culture methods, viz., exposing a plate of gelatine or agar for a longer or shorter time to the air of which examination is desired. By gravity the suspended bacteria fall on the plate and start growth. As a matter of quantitative exactitude, this method is not to be recommended, but it frequently proves an excellent method for qualitative estimation.
3. The Method of Miquel. Pasteur was the first to analyse air by the culture method, and he adopted a plan which in principle is washing the air in some fluid culture medium which will retain all the particulate matter, which may then be cultured directly or sub-cultured into any favourable medium.
Miquel has contrived a simple piece of apparatus for the
carrying out of this principle. It consists of a flask with a
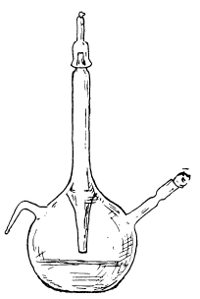 Miquel's Flask
central tube through its own neck
for the entrance of the air. On
one side of the flask is a tube to
be connected with the aspirator,
on the other side of the flask a
tube through which to pour off
the contained fluid at the end of
the process. In the flask are
placed 30 cc. of sterilised water
(or, indeed, if it be preferred,
sterilised broth). The entrance
tube is now unplugged, and the
aspirator draws through a fair
sample of the air in the room (say
ten litres). This air perforce
passes through the water and by
the exit tube to the aspirator, and
is thereby washed, leaving behind
in the water all its bacteria. The
aspiration is then stopped, and the98
entrance tube closed. The water (plus bacteria) is now
poured out into test-tubes of media or plated out on Petri's
dishes. Provided the apparatus has been absolutely sterilised,
and that the water was also sterile, any colonies
developing upon the Petri dish are composed of micro-organisms
from the air examined.
Miquel's Flask
central tube through its own neck
for the entrance of the air. On
one side of the flask is a tube to
be connected with the aspirator,
on the other side of the flask a
tube through which to pour off
the contained fluid at the end of
the process. In the flask are
placed 30 cc. of sterilised water
(or, indeed, if it be preferred,
sterilised broth). The entrance
tube is now unplugged, and the
aspirator draws through a fair
sample of the air in the room (say
ten litres). This air perforce
passes through the water and by
the exit tube to the aspirator, and
is thereby washed, leaving behind
in the water all its bacteria. The
aspiration is then stopped, and the98
entrance tube closed. The water (plus bacteria) is now
poured out into test-tubes of media or plated out on Petri's
dishes. Provided the apparatus has been absolutely sterilised,
and that the water was also sterile, any colonies
developing upon the Petri dish are composed of micro-organisms
from the air examined.
4. The Method of Hesse. This method is somewhat akin to Pouchet's aëroscope, but is in addition a culture method. Hesse's tube is about 2 feet long and 1-1/2 inches bore throughout. At one end is an india-rubber stopper bored for a glass tube to the aspirator. The other end is open. Before using, the tube is sterilised, and 40 or 50 cc. of sterilised gelatine replaced in it. The tube is now rapidly rotated in a groove on a block of ice or under a cold-water tap, and by this simple means the gelatine becomes fixed and forms a layer inside the tube throughout. We have therefore, so to speak, a tube of glass with a tube of gelatine inside it. The apparatus is now ready for use. It is fixed on the tripod, and fifteen litres of air are drawn through, and the tube is properly plugged and incubated at room temperature. In a day or two days the colonies appear upon the gelatine. They are most numerous generally in the first part of the tube, and might be roughly estimated as follows:
15 litres of air, 6 colonies.
⁂ 6/15 × 10,000 = 4000 aërobic bacteria in the cubic metre.
The disadvantages of this process are that dried gelatine does not catch germs like the broth cultures of Pasteur or Miquel, and that many organisms are able to go straight through the tube, and failing to be deposited, pass out at the aspirator exit, and thus are neither caught nor counted. The Hesse tube is generally used in practice with a pump consisting of two flasks and a double-way india-rubber tube. The flasks have a capacity for one litre of water. By a simple adaptation it is possible to secure siphon action, and99 hence measure with considerable exactitude the amount of air passing through the tube.
5. Methods of Filtration. To-day most of the above methods have been discarded, with the exception, perhaps, of Miquel's and modifications thereof.
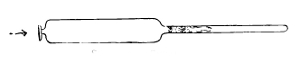
Sedgwick's Sugar-Tube
Frankland, Petri, Pasteur, Sedgwick, and others have suggested the adoption of methods of filtration. These depend upon catching the organisms contained in the air by filtering them through sterilised sand or sugar, and then examining these media in the ordinary way. Many different kinds of apparatus have been invented. Petri aspirates through a glass tube containing sterilised sand, which after use is distributed in Petri dishes and covered with gelatine. The principal objection to this method is the presence of the opaque particles of sand in and under the gelatine. Probably it was this which suggested the use of soluble filters like sugar. Pasteur introduced the principle, and Frankland and others have followed it out. The apparatus most largely used is that known as Sedgwick's Tube. This consists of a comparatively small glass tube, about a foot long. Half of it has a bore of 2.5 cm., and the other half a bore of .5 cm. It is sterilised at 150° C., after which the dry, finely granulated cane-sugar is inserted in such a way as to occupy an inch or more of the narrow part of the tube next the wide part. Next to it is placed a wool plug, and the whole is again sterilised at 130° C. for two hours, care being taken that the sugar does not melt. After sterilisation an india-rubber tube is fixed to the end of the narrow portion, and thus it is attached to the aspirator. The measured quantity (5–20 litres) of air is drawn through, and any particulate100 matter is caught in the sugar. Warm, nutrient gelatine (10–15 cc.) is now poured into the broad end of the tube, and by means of a sterilised stilette the sugar is pushed down into the gelatine, where it quickly dissolves. We have now in the gelatine all the micro-organisms in the air which has been drawn through the tube. After plugging with wool at both ends, the tube is rolled on ice or under a cold-water tap in order to fix the gelatine all round the inner wall of the tube, which is incubated at room tempera101ture. In a day or two the colonies appear, and may be examined.

Sedgwick's Tube
Fixed upon Tripod for Air Examination
Micro-organisms in the Air. Schwann was one of the first to point out that when a decoction of meat is effectually screened from the air, or supplied solely with calcined air, putrefaction does not set in. Helmholtz and Pasteur confirmed this, but it may be said with some truth that Schwann originated the germ theory, and Lister applied it in the treatment of wounds. Lister believed that if he could surround wounds with filtered air the results would be as good as if they were shut off from the air altogether.
It was Tyndall21 who first laid down the general principles upon which our knowledge of organisms in the air is based. That the dust in the air was mainly organic matter, living or dead, was a comparatively new truth; that epidemic disease was not due to "bad air" and "foul drains," but to germs conveyed in the air, was a prophecy as daring as it was correct. From these and other like investigations it came to be recognised that putrefaction begins as soon as bacteria gain an entrance to the putrefiable substance, that it progresses in direct proportion to the multiplication of bacteria, and that it is retarded when they diminish or lose vitality.
Tyndall made it clear that both as regards quantity and quality of micro-organisms in the air there neither is nor can be any uniformity. They may be conducted on particles of dust—"the raft theory"—but being themselves endowed with a power of flotation commensurate with their extreme smallness and the specific lightness of their composition, dust as a vehicle is not really requisite. Nevertheless the estimation of the amount of dust present in a sample of air is a very good index of danger. It is to Dr. Aitken that we are indebted for devising a method by which we can measure dust particles in the air, even though they 102be invisible. His ingenious experiments, reported in the Transactions of the Royal Society of Edinburgh (vol. xxxv.), have demonstrated that by supersaturation of air the invisible dust particles may become visible. As is now well known, Dr. Aitken has been able to prove that fogs, mists, and the like do not occur in dust-free air, and are due to condensation of moisture upon dust particles. But it should be remembered that, though dust forms a vehicle for bacteria, dusty air is often comparatively free from bacteria. Hence, after all, the necessary conditions for dissemination of bacteria in air are two, namely, some degree of air-current and dry surfaces.
This latter condition is one of essential importance. Bacteria cannot leave a moist surface either under evaporation or by means of air-currents.22 Only when there is considerable molecular disturbance, such as splashing, can there possibly be microbes transmitted to the surrounding air. This fact, coupled with the influence of gravitation, is the reason why sewer gas and all air contained within moist perimeters is almost germ-free; whereas from dry surfaces the least air-current is able to raise countless numbers of organisms. Quite recently this principle has been admirably illustrated in two series of investigations made upon expired and inspired air. In a report to the Smithsonian Institution of Washington (1895) upon the composition of expired air, it is concluded that "in ordinary quiet respiration no bacteria, epithelial scabs, or particles of dead tissue are contained in the expired air. In the act of coughing or sneezing such organisms or particles may probably be thrown out." The interior of the cavity of the mouth and external respiratory tract is a moist perimeter, from the walls of which no organisms can rise except under molecular dis103turbance. The position is precisely analogous to the germ-free sewer air as established by Messrs. Laws and Andrewes for the London County Council. The popular idea that infection can be "given off by the breath" is contrary to the laws of organismal pollution of air. The required conditions are not fulfilled, and such breath infection must be of extremely rare occurrence. The air can only be infective when filled with organisms arising from dried surfaces.
The other series of investigations were conducted by Drs. Hewlett and St. Clair Thompson, and dealt with the fate of micro-organisms in inspired air and micro-organisms in the healthy nose. They estimated that from 1500 to 14,000 bacteria were inspired every hour. Yet, as we have pointed out, expired air contains practically none at all. It is clear, then, that the inspired bacteria are detained somewhere. Lister has pointed out, from observation on a pneumo-thorax caused by a wound of the lung by a fractured rib, that bacteria are arrested before they reach the air-cells of the lung; hence it is at some intermediate stage that they are detained. Hewlett and Thomson examined the mucus from the wall of the trachea, and found it germ-free. It was only when they reached the mucous membrane and moist vestibules and vibrissæ of the nose that they found bacteria. Here they were present in abundance. The ciliated epithelium, the moist mucus, and the bactericidal influence of the wandering or "phagocyte" cells probably all contribute to their final removal.23
There can be no doubt that the large number of bacteria present in the moist surfaces of the mouth is the cause of a variety of ailments, and under certain conditions of ill-health organisms may through this channel infect the whole 104body. Dental caries will occur to everyone's mind as a disease possibly due to bacteria. As a matter of fact, probably acids (due to acid secretion and acid fermentation) and micro-organisms are two of the chief causes of decay of teeth. Defects in the enamel, inherent or due to injury, retention of débris on and around the teeth, and certain pathological conditions of the secretion of the mouth are predisposing causes, which afford a suitable nidus for putrefactive bacteria. The large quantity of bacteria which a decayed tooth contains is easily demonstrated.
From the two series of experiments which we have now considered we may gather the following facts:
(a) That air may contain great numbers of bacteria which may be readily inspired.
(b) That in health those inspired do not pass beyond the moist surface of the nasal and buccal cavities.
(c) That here there are various influences of a bactericidal nature at work in defence of the individual.
(d) That expired air contains, as a rule, no bacteria whatever.
The practical application of these things is a simple one. To keep air free from bacteria, the surroundings must be moist. Strong acids and disinfectants are not required. Moisture alone will be effectual. Two or three examples at once occur to the mind.
Anthrax spores are conveyed from time to time from dried infected hides and skins to the hands or bodies of workers in warehouses in Bradford and other places. If the surroundings were moist, and the hides moist, anthrax spores and all other bacteria would not remain free in the air.
The bacilli or spores of tubercle present in sputum in great abundance cannot, by any chance whatever, infect the air until, and unless, the sputum dries. So long as the expectorated matter remains on the pavement or handkerchief105 wet, the surrounding air will contain no bacilli of tubercle. But when in the course of time the sputum dries, then the least current of air will at once infect itself with the dried spores and bacilli.
Typhoid Fever, too, occupies the same position. Only when the excrement dries can the contained bacteria infect the air. It is of course well known that the common channel of infection in typhoid fever is not the air, whereas the reverse holds true of tuberculosis. The writer recently obtained some virulent typhoid excrement, and placed it in a shallow glass vessel under a bell-jar, with similar vessels of sterilised milk and of water, all at blood-heat. So long as the excrement remained moist, even though it soon lost its more or less fluid consistence, the milk and water remained uninfected. But when the excrement was completely dried it required but a few hours to reveal typhoid bacilli in the more absorptive fluid, milk, and at a later stage the water also showed clear signs of pollution. This evidence points in the same direction as that which has gone before. If the excrement of patients suffering from typhoid dries, the air will become infected; if, on the other hand, it passes in a moist state into the sewer, even though untreated with disinfectants, all will be well as regards the surrounding air.
Before passing on to consider other matters concerning organisms in the air, we may draw attention to some interesting observations recorded by Mr. S. G. Shattock24 on the negative action of sewer air in raising the toxicity of lowly virulent bacilli of diphtheria. Some direct relationship, it has been surmised, exists between breathing sewer air and "catching" diphtheria. Clearly it cannot be that the sewer air contains the bacillus. But some have supposed that the sewer air has had a detrimental effect by increasing the virulent properties of bacilli already in the 106human tissues. Two cultivations of lowly virulent bacilli were therefore grown by Mr. Shattock in flasks upon a favourable medium over which was drawn sewer air. This was continued for two weeks or five weeks respectively. Yet no increased virulence was secured. Such experiments require ample confirmation, but even from this it will be seen that sewer air does not necessarily have a favouring influence upon the virulence of the bacilli of diphtheria.
It should be noted that the bacilli of diphtheria are capable of lengthened survival outside the body, and are readily disseminated by very feeble air-currents. The condition necessary for their existence outside the body for any period above two or three days is moisture. Dried diphtheria bacilli soon lose their vitality. It is probably owing to this fact that the disease is not as commonly conveyed by air as, for example, tubercle.25
The influence of gravity upon bacteria in the air may be observed in various ways, in addition to its action within a limited area like a sewer or a room. Miquel found in some investigations in Paris that, whereas on the Rue de Rivoli 750 germs were present in a cubic metre, yet at the summit of the Pantheon only 28 were found in the same quantity of air. At the tops of mountains air is germ-free, and bacteria increase in proportion to descent. As Tyndall has pointed out, even ultra-microscopic cells obey the law of gravitation. This is equally true in the limited areas of a laboratory or warehouse and in the open air.
The conditions which affect the number of bacteria in the air are various. After a fall of rain or snow they are very markedly diminished; during a dry wind they are increased. In open fields, free from habitations, they are fewer, as would be expected, than in the vicinity of manufactories, houses, or towns. A dry, sandy soil or a dry surface of any kind will obviously favour the presence of organisms in 107the air. Frankland found that fewer germs were present in the air in winter than in summer, and that when the earth was covered with snow the number was greatly reduced. Miquel and Freudenreich have declared that the number of atmospheric bacteria is greater in the morning and evening between the hours of six and eight than during the rest of the day. But we venture to express the hope that such coincidental facts may not be exalted into principles.
There is no numerical standard for bacteria in the air as there is in water. The open air possibly averages about 250 per cubic metre. On the seacoast this number would fall to less than half; in houses and towns it would rise according to circumstances, and frequently in dry weather reach thousands per cubic metre. When it is remembered that air possesses no pabulum for bacteria as do water and milk, it will be understood that bacteria do not live in the air. They are only driven by air-currents from one dry surface to another. Hence the quality and quantity of air organisms depend entirely upon environment and physical conditions. In some researches which the writer made into the air of workshops in Soho in 1896, it was instructive to observe that fewer bacteria were isolated by Sedgwick's sugar-tube in premises which appeared to the naked eye polluted in a large degree than in other premises apparently less contaminated. In the workroom of a certain skin-curer the air was densely impregnated with particles from the skin, yet scarcely a single bacterium was isolated. In the polishing-room of a well-known hat firm, in which the air appeared to the naked eye to be pure, and in which there was ample ventilation, there were found four or five species of saprophytic bacteria. Quite recently Mr. S. R. Trotman, public analyst for the city of Nottingham, estimated the bacterial quality of the air of the streets of that town during "the goose fair" held in the autumn. He used a modification of Hesse's apparatus in which the gelatine is replaced by108 glycerine. The air was slowly drawn through and measured in the usual way. Sterilised water was then added to bring the glycerine to a known volume, the liquid thoroughly mixed, and a series of gelatine and agar plates made with quantities varying from 0.1 to 2 cc. By this method a large number of bacteria were detected in this particular investigation, including Staphylococcus pyogenes aureus et albus, the common Bacillus subtilis, and B. coli communis.26
During a six years' investigation the air of the Montsouris Park yielded, according to Miquel, an average of 455 bacteria per cubic metre. In the middle of Paris the average per cubic metre was nearly 4000. Flügge accepts 100 bacteria per cubic metre as a fair average. From this fact he estimates that "a man during a lifetime of seventy years inspires about 25,000,000 bacteria, the same number contained in a quarter of a litre of fresh milk."27 Many authorities would place the average much below 100 per cubic metre, but even if we accept that figure it is at once clear how relatively small it is. This is due, as we have mentioned, to sunlight, rain, desiccation, dilution of air, moist surfaces, etc. So essentially does the bacterial content of air depend upon the facility with which certain bacteria withstand drying that Dr. Eduardo Germano28 has addressed himself first to drying various pathogenic species and then to mixing the dried residue with sterilised dust and observing to what degree the air becomes infected. Typhoid appears to withstand comparatively little dessication, without losing its virulence. Nevertheless, it is able to retain vitality in a semi-dried condition, and it is owing to this circumstance in all probability that it possesses such power of infection. Diphtheria, on the other hand, is, as we have pointed out, capable of lengthened survival outside the body, particularly 109when surrounded with dust. The question of their power of resisting long drying is an unsettled point. The power of surviving a drying process is, according to Germano, possessed by the streptococcus. This is not the case with cholera or plague. Dr. Germano classifies bacteria, as a result of his researches, into three groups: first, those like plague, typhoid, and cholera, which cannot survive drying for more than a few hours; second, those like the bacilli of diphtheria, and streptococci, which can withstand it for a longer period; thirdly, those like tubercle, which can very readily resist drying for months and yet retain their virulence. It will be obvious that from these data it is inferred that Groups 1 and 2 are rarely conveyed by the air, whereas Group 3 is frequently so conveyed. Miquel has recently demonstrated that soil bacteria or their spores can remain alive in hermetically sealed tubes for as long a time as sixteen years. Even at the end of that period the soil inoculated into a guinea-pig produced tetanus.29
The presence of pathogenic bacteria in the air is, of course, a much rarer contamination than the ordinary saprophytes. Tubercle has been not infrequently isolated from dry dust in consumption hospitals, and in exit ventilating shafts at Brompton the bacillus has been found. From dried sputum it has, of course, been many times isolated, even after months of desiccation. M. Lalesque failed to isolate it from the dry soil surrounding some garden seats in a locality frequented by phthisical patients. The writer also failed to isolate it from the same soil. But a very large mass of experimental evidence attests the fact that the air in proximity to dried tubercular sputum or discharges may contain the specific bacillus of the disease. Diphtheria in the same way, but in a lesser degree, may be isolated from the air, and from the nasal mucous membrane of nurses, attendants, and patients in a ward set apart for the treat110ment of the disease. Delalivesse, examining the air of wards at Lille, found that the contained bacteria varied more or less directly with the amount of floating matter, and depended also upon the vibration set up by persons passing through the ward and the heavy traffic in granite-paved streets adjoining. Bacillus coli, staphylococci, and streptococci, as well as B. tuberculosis, were isolated by this observer.
Some new light has been thrown upon the subject of pathogenic organisms in air by Neisser in his investigations concerning the amount and rate of air-currents necessary to convey certain species through the atmosphere. He states that the bacteria causing diphtheria, typhoid fever, plague, cholera, and pneumonia, and possibly the common Streptococcus pyogenes, are incapable of being carried by the molecules of atmospheric dust which the ordinary insensible currents of air can support, whilst Bacillus anthracis, B. pyocyaneus, and the bacillus of tubercle are capable of being aërially conveyed. This work will require further confirmation, but if its truth be established, it proves that attempted aërial disinfection of the first group of diseases is useless.
It was Pasteur who in 1857 first propounded the true cause and process of fermentation. The breaking down of sugar into alcohol and carbonic acid gas had been known, of course, for a long period. Since the time of Spallanzani (1776) the putrefactive changes in liquids and organic matter had been prevented by boiling and subsequently sealing the flask or vessel containing the fluid. Moreover, this successful preventive practice had been in some measure correctly interpreted as due to the exclusion of the atmosphere, but wrongly credited to the exclusion of the oxygen of the air. It was not until the beginning of the present century that authorities modified their view and declared in favour of yeast cells as the agents in the production of fermentation. That this process was due to oxygen per se was disproved by Schwann, who showed that so long as the oxygen admitted to the flask of fermentative fluid was sterilised no fermentation occurred. It was thus obvious that it was not the atmosphere or the oxygen of the atmosphere, but some fermenting agent borne into the flask by the admission of unsterilised air. It was but a step to further establish this hypothesis by adding unsterilised air plus some antiseptic substance which would destroy the fermenting agent. Arsenic was found by Schwann to have this germicidal faculty. Hence Schwann supported Latour's theory that fermentation was due to something borne in by the air, and112 that this something was yeast. Passing over a number of counter-experiments of Helmholtz and others, we come to the work of Liebig. He viewed the transformation of sugar into alcohol and carbonic acid gas simply and solely as a non-vital chemical process, depending upon the dead yeast communicating its own decomposition to surrounding elements in contact with it.
Liebig insisted that all albuminoid bodies were unstable, and if left to themselves would fall to pieces—i. e., ferment—without the aid of living organisms, or any initiative force greater than dead yeast cells. It was at this juncture that Pasteur intervened to dispel the obscurities and contradictory theories which had been propounded.
As in all the conclusions arrived at by Pasteur, so in those relating to fermentation, there were a number of different experiments which were performed by him to elucidate the same point. We will choose one of many in relation to fermentation. If a sugary solution of carbonate of lime is left to itself, after a time it begins to effervesce, carbonic acid is evolved, and lactic acid is formed; and this latter decomposes the carbonate of lime to form lactate of lime. This lactic acid is formed, so to speak, at the expense of the sugar, which little by little disappears. Pasteur demonstrated the cause of this transformation of sugar into lactic acid to be a thin layer of organic matter consisting of extremely small moving organisms. If these be withheld or destroyed in the fermenting fluid, fermentation will cease. If a trace of this grey material be introduced into sterile milk or sterile solution of sugar, the same process is set up, and lactic acid fermentation occurs.
Pasteur examined the elements of this organic layer by aid of the microscope, and found it to consist of small short rods of protoplasm quite distinct from the yeast cells which previous investigators had detected in alcoholic fermentation. One series of experiments was accomplished with113 yeast cells and these bacteria, a second series with living yeast cells only, a third series with bacteria only, and the conclusions which Pasteur arrived at as the result of these labours were as follows:
"As for the interpretation of the group of new facts which I have met with in the course of these researches, I am confident that whoever shall judge them with impartiality will recognise that the alcoholic fermentation is an act correlated to the life and to the organisation of these corpuscles, and not to their death or their putrefaction, any more than it will appear as a case of contact action in which the transformation of the sugar is accomplished in the presence of the ferment without the latter giving or taking anything from it."
Pasteur occupied six years (1857–1863) with further elucidation of his wonderful discovery of the potency of these hitherto unrecognised agents, and the establishment of the fact that "organic liquids do not alter until a living germ is introduced into them, and living germs exist everywhere."
It must not be supposed that to Pasteur is due the whole credit of the knowledge acquired respecting the cause of fermentation. He did not first discover these living organisms; he did not first study them and describe them; he was not even the first to suggest that they were the cause of the processes of fermentation or disease. But, nevertheless, it was Pasteur who "first placed the subject upon a firm foundation by proving with rigid experiment some of the suggestions made by others." Thus it has ever been in the times of new learning and discovery: many contributors have added their quota to the mass of knowledge, even though one man appearing at the right moment has drawn the conclusions and proved the theory to be fact.
In order that no confusion may arise in the mind of the reader, we may here say that, although fermentation is114 always due to a living agent, as proved by Pasteur, the process is conveniently divided into two kinds.30 (1) When the action is direct, and the chemical changes involved in the process occur only in the presence of the cell, the latter is spoken of as an organised ferment; (2) when the action is indirect, and the changes are the result of the presence of a soluble material secreted by the cell, acting apart from the cell, this soluble substance is termed an unorganised soluble ferment, or enzyme. The organised ferments are bacteria or vegetable cells allied to the bacteria; the unorganised ferments, or enzymes, are ferments found in the secretions of specialised cells of the higher plants and animals. With the former this book deals in an elementary fashion; with the latter we have little concern. It will be sufficient to illustrate the enzymes by a few of the more familiar examples. They form, for example, the digestive agents in human assimilation. This function is performed, in some cases, by the enzyme combining with the substance on which it is acting and then by decomposition yielding the new "digested" substance and regenerating the enzyme; in other cases, the enzyme, by its molecular movement, sets up molecular movement in the substance it is digesting, and thus changes its condition. These digestive enzymes are as follows: in the saliva, ptyalin, which changes starch into sugar; in the gastric juice of the stomach, pepsin, which digests the proteids of the food and changes them into absorptive peptones; the pancreatic ferments, amylopsin, trypsin, and steapsin, capable of attacking all three classes of food stuffs; and the intestinal ferments, which have not yet been separated in purer condition. In addition to these, there are ferments in bitter almonds, mustard, etc. Concerning these unorganised ferments we have nothing further to say. Perhaps the commonest of them all is diastase, which occurs in malt, and to which some reference will be made later.
Its function is to convert the starch which occurs in barley into sugar. These unorganised ferments act most rapidly at about 75° C. (167° F.).31
We may now return to the work of Pasteur and the question of organised ferments. Let us preface further remark with an axiom with which Professor Frankland sums up the vitalistic theory of fermentation, which was supported by the researches of Pasteur: "No fermentation without organisms, in every fermentation a particular organism." From these words we gather that there is no one particular organism or vegetable cell to be designated the micro-organism of fermentation, but that there are a number of fermentations each started by some specific form of agent. It is true that the chemical changes induced by organised ferments depend on the life processes of micro-organisms which feed upon the sugar or other substance in solution, and excrete the product of the fermentation. Fermentation nearly always consists of a process of breaking down of complex bodies, like sugar, into simpler ones, like alcohol and carbonic acid. Of such fermentation we may mention at least five: the alcoholic, by which alcohol is produced; the acetous, by which wine absorbs oxygen from the air and becomes vinegar; the lactic, which sours milk; the butyric, which out of various sugars and organic acids produces butyric acid; and ammoniacal, which is the putrefactive breaking down of compounds of nitrogen into ammonia. We have already referred at some length to this process when considering denitrifying organisms in the soil.
There are four chief conditions common to all these five kinds of organised fermentation. They are as follows:—
1. The presence of the special living agent or organism of the particular fermentation under consideration. This, as Pasteur pointed out, differs in each case.
2. A sufficiency of pabulum (nutriment) and moisture to favour the growth of the micro-organism.
3. A temperature at or about blood-heat (35–38° C., 98.5° F.).
4. The absence from the solution or substance of any obnoxious or inimical substances which would destroy or retard the action of the living organism and agent. Many of the products of fermentation are themselves antiseptics, as in the case of alcohol; hence alcoholic fermentation always arrests itself at a certain point.
We are now in a position to consider particular fermentations and their causal micro-organisms. These latter are of various kinds, belonging, according to botanical classification, to various different subdivisions of the non-flowering portion of the vegetable kingdom. A large part of fermentation is based upon the growth of a class of microscopic plants termed yeasts. These differ from the bacteria in but few particulars, mainly in their method of reproduction by budding (instead of dividing or sporulating, like the bacteria). Their chemical action is closely allied to that of the bacteria. Secondly, there are special fermentations and modifications of yeast fermentation due to bacteria. Thirdly, a group of somewhat more highly specialised vegetable cells, known as moulds, make a perceptible contribution in this direction. According to Hansen, these latter, so far as they are really alcoholic ferments, induce fermentation, not only in solutions of dextrose and invert sugar, but also in solutions of maltose. Mucor racemosus is the only member that is capable of inverting a cane-sugar solution; M. erectus is the most active fermenter, yielding eight per cent. by117 volume of alcohol in ordinary beer wort. Each of these will be referred to as they occur in considering the five important fermentations already mentioned.

Saccharomyces Cerevisiæ
The general microscopic appearance of yeast cells may be shortly stated as follows: they are round or oval cells, and by budding become daughter yeasts. Each consists of a membrane and clear homogeneous contents. As they perform their function of fermentation, vacuoles, fat-globules, and other granules make their appearance in the enclosed plasma. As in many vegetable cells a nucleus was detected by Schmitz by means of special methods of staining, Hansen has found the nucleus in old yeast cells from "films" without any special staining.
Cause, yeast; medium, sugar solutions; result, alcohol and carbonic acid.
It was Caignard-Latour who first demonstrated that yeast cells, by their growth and multiplication, set up a chemical change in sugar solutions which resulted in the transference of the oxygen from the hydrogen in the sugar compound to the carbon atoms, that is to say, in the evolution of carbonic acid gas and the production, as a result, of alcohol. If we were to express this in a chemical formula, it would read as follows:
C6H12O6 (plus the yeast) = 2 C2H6O + 2 CO2.
A natural sugar, like grape-sugar, present in the fruit of the vine, is thus fermented. The alcohol remains in the liquid; the carbonic acid escapes as bubbles of gas into the surrounding air. It is thus that brandy and wines are made. If we go a step further back, to cane-sugar (which possesses the same elements as grape-sugar, but in different proportions), dissolve it in water, and mix it with yeast, we get exactly the same result, except that the first stage of the fermentation would be the changing of the cane-sugar into grape-sugar, which is accomplished by a soluble ferment secreted by the yeast cells themselves. If now we go yet one step further back, to starch, the same sort of action occurs. When starch is boiled with a dilute acid it is changed into a gum-like substance named dextrin, and subsequently into a sugar named maltose, which latter, when mixed with these living yeast cells, is fermented, and results in the evolution of carbonic acid gas and the production of alcohol. In the manufacture of fermented drinks from cereal grains containing starch there is therefore a double chemical process: first the change of starch into sugar by means of conversion,32 and secondly the change of the sugar into alcohol and carbonic acid gas by the process of fermentation, an organic change brought about by the living yeast cells.
In all these three forms of alcoholic fermentation the principal features are the same, viz., the sugar disappears; the carbonic acid gas escapes into the air; the alcohol remains behind. Though it is true that the sugar disappears, it would be truer still to say that it reappears as alcohol. Sugar and alcohol are built up of precisely the same elements: carbon, hydrogen, and oxygen. They differ from 119each other in the proportion of these elements. It is obvious, therefore, that fermentation is really only a change of position, a breaking down of one compound into two simpler compounds. This redistribution of the molecules of the compound results in the production of some heat. Thus we must add heat to the results of the work of the yeasts.
When alcohol is pure and contains no water it is termed absolute alcohol. If, however, it is mixed with 16 per cent. of water, it is called rectified spirit, and when mixed with more than half its volume of water (56.8 per cent.) it is known as proof spirit.
We shall have to consider elsewhere a remarkable faculty which some bacteria possess of producing products inimical to their own growth. In some degree this is true of the yeasts, for when they have set up fermentation in a saccharine fluid there comes a time when the presence of the resulting alcohol is injurious to further action on their part. It has become indeed a poison, and, as we have already mentioned, a necessary condition for the action of a ferment is the absence of poisonous substances. This limit of fermentation is reached when the fermenting fluid contains 13 or 14 per cent. of alcohol.
Having discussed shortly the "medium" and the results, we may now turn to the bacteriology of the matter, and enumerate some of the chief forms of the yeast plant. Professor Crookshank33 gives more than a score of different members of this family of Saccharomycetes. Before dwelling upon some of the chief of these, it will be desirable to consider a number of properties common to the genus.
The yeast cell is a round or oval body of the nature of a fungus, composed of granular protoplasm surrounded by a definite envelope, or capsule. It reproduces itself by budding, or, as it is sometimes termed, gemmation. At one end 120of the cell a slight swelling or protuberance appears, which slowly enlarges. Ultimately there is a constriction, and the bud becomes partly and at last completely separated from the parent cell. In many cases the capsules of the daughter cell and the parent cell adhere, thus forming a chain of budding cells. The character of the cell and its method of reproduction do not depend merely upon the particular species alone, but are also dependent upon external circumstances. There are differences in the behaviour of species towards different media at various temperatures, towards the carbohydrates (especially maltose), and in the chemical changes which they bring about in nutrient liquids. In connection with this Professor Hansen has pointed out that, whilst some species can be made use of in fermentation industries, others cannot, and some even produce diseases in beer.34
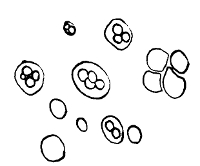
Ascospore Formation
One of the most remarkable evidences of the adaptability of the yeasts to their surroundings and a specific characteristic occurs in what is sometimes called ascospore formation. If a yeast cell finds itself lacking nourishment or in an unfavourable medium, it reproduces itself not by budding, but by forming spores out of its own intrinsic substance, and within its own capsule. To obtain this kind of spore formation Hansen used some gypsum blocks as medium on which 121to grow his yeast cells. Well-baked plaster of Paris is mixed with distilled water, and made into a liquid paste. Small moulds are made by pouring this paste into cardboard dishes, where it hardens again. The mould is sterilised by heat, and a small portion of yeast is placed on its upper surface, and then the whole is floated in a small vessel of water and covered with a bell-jar. Under these conditions of limited pabulum the cell undergoes the following changes: it increases in size, loses much of its granularity, and becomes homogeneous, and about thirty hours after being sown on the gypsum there appear several refractile cells inside the parent cell. These are the ascospores. In addition to the gypsum, it is necessary to have a plentiful supply of oxygen, some moisture (gained from the vessel of water in which the gypsum floats), a certain temperature, and a young condition of the protoplasm of the parent yeast cells. Hansen found that the lowest temperature at which these ascospores were produced was .5–3° C., and at the other extreme up to 37° C., which is blood-heat. The rapidity of formation also varies with the temperature, the favourable degree of warmth being about 22–25° C.
Gypsum Block
Hansen pointed out that it was possible by means of sporulation to differentiate species of yeasts. For it happens that different species show slight differences in spore formation, e. g.:
(a) The spores of Saccharomyces cerevisiæ expand during the first stage of germination, and produce partition walls, making a compound cell with several chambers. Budding can occur at any point on the surface of the swollen spores. To this group belong S. pastorianus and S. ellipsoideus.
(b) The spores of Saccharomyces Ludwigii fuse in the first stage, and afterwards grow out into a promycelium, which produces yeast cells.
(c) The spores of Saccharomyces anomalus are different in shape from the others in that they possess a projecting rim round the base.
Another point in the cultivation of yeasts has been elucidated by a number of workers, chief among whom perhaps is Hansen, namely, methods of obtaining pure cultures. We know, generally speaking, what this term means, and there is no difference in its meaning here to what is understood as its meaning with regard to bacteria. There is, however, some difference in the mode of securing it. It is only by starting with one individual cell that we can hope to secure a pure culture of yeasts. For the study of the morphology of yeasts under the microscope the problem was not a difficult one. It was comparatively easy to keep out foreign germs from a cover-glass preparation enough to perceive germination of spores and growth of mycelium. But when we require pure cultures for various physiological purposes, then a different standard and method are necessary.
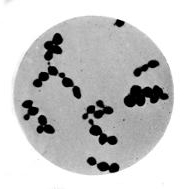 |
 |
| Yeast (Saccharomyces Cerevisiæ) × 1000 |
Ascospore Formation in Yeast (The capsule of the parent cell around the spores is invisible) × 1000 |
 |
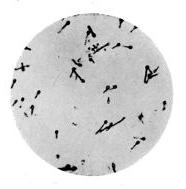 |
| Nitrogen Fixing Bacteria from Rootlet-Nodules (Subculture) × 1000 |
Bacillus of Tetanus (From broth culture, showing spore formation) × 1000 By permission of the Scientific Press, Limited |
Pasteur and Cohn adopted a practice based upon the fact
that when organisms find themselves in a favourable medium
they multiply to the exclusion of others to which the medium
is less favourable. Hence if an impure mixture be placed
under such circumstances there comes a time when those
organisms for which the circumstances are favourable multiply
to such an extent that they form an almost pure culture.
The method is open to fallacy, and will rarely result
in a really pure culture; and even if that be secured, it is
123quite possible that it will be to the exclusion of the desired
culture. Hansen has devised a much improved process for
securing a pure culture of yeast which depends upon dilution.
We believe Lister was one of the first who, in the
seventies, introduced some such plan as this. Hansen employed
dilution with water in the following manner:
Yeast is diluted with a certain amount of sterilised water. A drop is carefully examined under the microscope, a single cell of yeast is taken, and a cultivation made upon wort. When it has grown abundantly a quantity of sterilised water is added. From this, again, a single drop is taken and added, to, say 20 cc. of sterilised water in a fresh flask. This flask will contain we will suppose ten cells. It is now vigorously shaken, and the contents are divided into twenty portions of 1 cc. each, and added to twenty tubes of sterilised water. It is highly probable that half of those tubes have received one cell each. In the course of a few days it can be seen how far a culture is pure. If only one colony is present, the culture is a pure one, and as this grows we obtain an absolutely pure culture in necessary quantity. Even when the gelatine-plate method is used it is desirable to start with a single cell (Hansen). The advantage of Hansen's yeast method over Koch's bacterial-plate method is that it has a certain definite starting-point. This is obviously impossible when dealing with such microscopic particles as the bacteria proper.
A third matter in the differentiation of yeast species is the question of films. Hansen set to work, after having obtained pure cultures and ascospores, to examine films appearing on the surface of liquids undergoing fermentation. The object of this was to ascertain whether all yeasts produced the same mycelial growth on the surface of the fermenting fluid. To produce these films the process is as follows: Drop on to the surface of sterilised wort in a flask a very small quantity of a pure culture of yeast; secure the124 flask from movement, and protect it, not from air, which is necessary, but from falling particles in the air. In a short time small colonies appear, which coalesce and form patches, then a film or membrane which covers the liquid and attaches itself to the sides of the flask. By the differences in the films and the temperatures at which they form it is possible to obtain something of a basis for classification. The further advances in a yeast culture and in our knowledge of the agencies of fermentation have, however, tended to show that no strict dividing lines can be drawn. Hansen's researches have, notwithstanding, been of the greatest moment to the whole industry of fermentation. What has been found true in bacteriology has also been demonstrated in fermentation, namely, that though many yeasts differ but little in structure and behaviour, they may produce very different products and possess very different properties. Industrial cultivation of these finer differences in fermentative action has to a large extent revolutionised the brewing industry.
The formation of films is not a peculiarity of certain species, but must be regarded as a phenomenon occurring somewhat commonly amongst yeasts. The requisites are a suitable medium, a yeast cell, a free, still surface, direct access of air, and a favourable temperature. The wort loses colour, and becomes pale yellow. Microscopic differences soon appear between the sedimentary yeast and the film yeast of the same species, the latter growing out into long mycelial forms, the character of which depends in part upon the temperature. This often varies from 3° to 38° C.
A fourth point helpful in diagnosis is the temperature which proves to be the thermal death-point. Saccharomyces cerevisiæ is killed by an exposure to 54° C. for five minutes, and 62° C. kills the spores. As a rule, yeasts can resist a considerably higher temperature when in a dry state than in the presence of moisture.
Lastly, yeasts may be cultivated on solid media. Hansen125 employed wort-gelatine (5 per cent. gelatine), and found that at 25° C. in a fortnight the growths which develop show such microscopic differences as to aid materially in diagnosis. Saccharomyces ellipsoideus I. exhibits a characteristic network which readily distinguishes it.
There is one other point to which reference must be made. The process of fermentation may be set up by a "high" or a "low" yeast. These terms apply to the temperature at which the process commences. "High" yeasts rise to the surface as the action proceeds, accomplish their work rapidly, and at a comparatively high temperature, say about 16° C.; "low" yeasts, on the contrary, sink in the fermenting fluid, act slowly, and only at the low temperature of 4° or 5° C. This is maintainable by floating ice in the fluid. Formerly all beer was made by the "high" mode, but on the continent of Europe "low" yeast is mostly used, while the "high" is in vogue in England. This latter method is more conducive to the development of extraneous organisms, and therefore risky in all but well-ordered brewing establishments. Whether high and low yeasts consist of one or several species is not known.
Before proceeding to mention shortly some of the commoner forms of yeast we must again emphasise Hansen's method of analysis in separating a species. The shape, size, and appearance of cells are not sufficient for differentiation, because it is found that the same species when exposed to different external conditions can occur in very different forms. Hence Hansen established the analytical method of observing (1) the microscopic appearance, (2) the formation of ascospores, and (3) the formation of films. In addition, the temperature limits, cultivation on solid media, and behaviour towards carbohydrates, are characters which aid in the separation of yeasts. By basing differentiation of species upon these features, the following can be distinguished:
Saccharomyces Cerevisiæ. Oval or ellipsoidal cells; reproduction by budding; ascospores, rapidly at 30° C., slowly at 12° C., not formed at all at lower temperatures; film formation, seven to ten days at 22° C.; an active alcoholic ferment, producing in a fortnight in beer wort from 4 to 6 per cent. by volume of alcohol (Jörgensen). This species is a typical English "high" yeast, possessing the power of "inverting" cane-sugar previous to producing alcohol and carbonic acid. It is said to have no action on milk-sugar.
 |
 |
S. Ellipsoideus |
S. Pastorianus |
Saccharomyces Ellipsoideus I. Round, oval, or sausage-shaped cells, single or in chains; ascospores in twenty-four hours at 25° C. (not above 30° C., not below 4° C.). Grown on the surface of wort-gelatine, a network is produced by which they can be recognised (in eight to twelve days at 33° C.). At 13–15° C. a characteristic branching mass is produced. It is an alcoholic ferment as active as S. cerevisiæ. S. Ellipsoideus II. Round and oval, rarely elongated, a widely distributed yeast, causing "muddiness" in beer and a bitter taste. It is essentially a "low" yeast.
Saccharomyces Conglomeratus is a round cell, often united in clusters, and occurring in rotting grapes, and at the commencement of fermentation.
Saccharomyces Pastorianus I. Oval or club-shaped cells, occurring in after-fermentation of wine, etc., and producing a bitter taste, unpleasant odour, and turbidity. The spores frequently occur in the air of breweries.
S. Pastor. II. Elongated cells, possessing an invertose ferment. They do not, like S. pastor I., produce disease in beer.
S. Pastor. III. Oval or elongated cells, producing turbidity in beer. Grown on yeast-water gelatine, the colonies show after sixteen days crenated hairy edges.
Saccharomyces Apiculatus. Lemon-shaped cells. They give rise to a feeble alcoholic fermentation, and produce two kinds of spores—round and oval; they appear at the onset of vinous fermentation, but give way later on to S. cerevisiæ.
Saccharomyces Mycoderma. Oval or elliptical cells, often in branching chains. They form the so-called "mould" on fermented liquids, and develop on the surface without exciting fermentation. When forced to grow submerged they produce a little alcohol.
Saccharomyces Exiguus. Conical cells, appearing in the after-fermentation of beer.
Saccharomyces Pyriformis. Oval cells, converting sugary solutions containing ginger into ginger-beer.
Saccharomyces Illicis, Hansenii, et Aquifolii produce a small percentage of alcohol.
Cause, Mycoderma aceti; medium, wine and other alcoholic liquids; result, the formation of vinegar.
If alcohol be diluted with water, and the specific ferment mixed with it and exposed to the air at 22° C., it is rapidly converted into vinegar. The change is accompanied by the absorption of oxygen, one atom of which combines with two of hydrogen to form water, and a substance remains128 called aldehyde, further oxidation of which produces the acetic acid. We may express it chemically thus:
| Alcohol. | Aldehyde. Water. |
| C2H6O (+ oxygen and the ferment) = C2H4O + H2O. | |
The aldehyde becomes further oxidised:
C2H4O + O = C2H4O2 (acetic acid).
Now this method of simply oxidising alcohol to obtain acetic acid may be carried out chemically without any ferment. If slightly diluted alcohol be dropped upon platinum black, the oxygen condensed in that substance acts with energy upon the spirit, and union readily occurring, acetic acid results. Here the whole business of the platinum sponge is to persuade the oxygen of the air and the hydrogen of the alcohol to unite. In the ordinary manufacture this is accomplished by the vegetable cells of Mycoderma aceti.
There are two chief methods adopted in the commercial manufacture of vinegar, both of which depend upon the presence of the Mycoderma. The method in vogue at Orleans when Pasteur (about 1862) commenced his studies of the vinegar organism was to fill vats nearly to the brim with a weak mixture of vinegar and wine. Where the process is proceeding the surface is covered with a fragile pellicle, "the mother of vinegar," which is produced by and consists of certain micro-organisms whose function is to convey the oxygen of the air to the liquor in the vats, thus oxidising the alcohol into vinegar. This oxidation may be carried on even beyond the stage of acetic acid (when no more alcohol remains to be oxidised), resulting in carbonic acid gas, which escapes into the air. But as in the alcoholic, so in the acetic, fermentation, there comes a time when the presence of an excess of the acid inhibits the further growth of the organism. This point is approxim129ately when the acetic acid has reached a percentage as high as 14. But if the acid be removed, and fresh alcohol added, the process recommences.
The second method, sometimes called by the Germans the "quick vinegar process," is to pour the weakened alcohol through a tall cylinder filled with wood-shavings, having first added some warm vinegar to the shavings. After a number of hours the resulting fluid is charged with acetic acid. What has occurred? Liebig maintained that a chemical and mechanical change had brought about the change from the alcohol put into the cylinder and the vinegar drawn off at the exit tube. It was reserved for Pasteur to demonstrate by experiment that the addition of the warm vinegar to the shavings was in reality an addition of a living micro-organism, which, forming a film upon the shavings, became "the mother of vinegar," and oxidised the alcohol which passed over it, inducing it to become aldehyde and then acetic acid.
Mycoderma Aceti (described by Persoon 1822, Kützing 1837, and Pasteur 1864). It must be understood that this term is the name rather of a family than an individual. Pasteur believed it to be a specific individual, but Hansen pointed out that it was composed of two distinctly different species (Bacterium aceti and B. pasteurianum), and subsequently other investigators have added members to the acetic fermentation group of which M. aceti is the type.
This bacterium is made up of small, slightly elongated cells, with a transverse diameter of 2 or 3 µ, sometimes united in short chains of curved rods. They frequently show a central constriction, are motile, and produce in old cultures involution forms. The way in which the cells act and are made to perform their function is as follows: A small quantity, taken from a previous pellicle, is sown on the surface of an aqueous liquid, containing 2 per cent. of alcohol, 1 per cent. of vinegar, and traces of alkaline phos130phates. Very rapidly indeed the little isolated colonies spread, and, becoming confluent, form a membrane or pellicle over the whole area of fluid. When the surface is covered the alcohol acidifies to vinegar. After this it is necessary to add each day small quantities of alcohol. When the oxidation is completed the vinegar is drawn off, and the membrane is collected and washed, and is then again ready for use. It ought not to remain long out of fermenting liquid, nor ought it to be allowed to over-perform its function, for thus having oxidised all the alcohol it will commence oxidation of the vinegar.
In wort-gelatine Bacterium pasteurianum develops round colonies with a smooth or wavy border, whilst B. aceti has a tendency towards stellate arrangement. Spores have not been observed, and from a morphological point of view the two species behave alike. Neither produces any turbidity in the liquid containing them. In order to flourish, B. aceti requires a temperature of about 33° C. and a plentiful supply of oxygen. In a cool store or cellar there is, therefore, nothing to fear from B. aceti. Frankland has isolated a Bacillus ethaceticus, which is a fermentative organism producing ethyl-alcohol and acetic acid. By oxidation the ethyl-alcohol may be converted into acetic acid.
Cause, Bacillus acidi lactici; medium, milk-sugar, cane-sugar, glucose, dextrose, etc.; result, lactic acid.
The process set up by the lactic ferment is simply a decomposition, an exact division of one molecule of sugar into two molecules of lactic acid, there being neither oxidation nor hydration. The conditions under which the ferment acts are very similar to those we have already considered. There is frequently carbonic acid gas formed; there is a cessation of fermentation when the medium becomes too131 acid; there is the same method of starting the process by inoculation of sour milk or cheese or any substance containing the specific bacillus. It is probable that such inoculated matter will contain a mixture of micro-organisms, but if the lactic bacillus is present, it will grow so vigorously and abundantly that the fermentation will be readily set up.

B. Acidi Lactici
The Bacillus Acidi Lactici. Rods about 2 µ long and 4 µ wide, occurring singly or in chains and threads. It is non-motile. Spore formation is present, the spores appearing irregularly or at one end of the rod.
On the surface of gelatine a delicate growth appears along the track of the needle, with round colonies appearing at the edges of the growth. It does not liquefy gelatine. It grows best at blood-heat; but much above that it fails to produce its fermentation, and it ceases to grow under 10° C. It inverts milk-sugar and changes it to dextrose, from which it then produces lactic acid. Sugars do, however, differ considerably in the degrees to which they respond to the influence of the lactic ferment, and some which are readily changed by the alcoholic ferment are untouched by the Bacillus acidi lactici. It will be necessary to refer again to this micro-organism when we come to speak of milk and other dairy products.
Van Laer has described a saccharobacillus which produces lactic acid amongst other products, and brings about a characteristic disease in beer, named tourne. The liquid132 gradually loses its brightness and assumes a bad odour and disagreeable taste. The bacillus is a facultative anaërobe. A number of workers have separated organisms, having a lactic acid effect, which diverge considerably from the orthodox type of lactic acid bacillus. This is but further evidence of a fact to which reference has been made: that nomenclature restricted to one individual has now become adapted to a family.
Cause, Bacillus butyricus and B. amylobacter; medium, milk, butter, sugar and starch solutions, glycerine; result, butyric acid.
When sugars are broken down by the Bacillus acidi lactici the lactic acid resulting may, under the influence of the butyric ferment, become converted into butyric acid, carbonic acid, and hydrogen. Neither butyric acid nor lactic acid is as commonly used as alcohol or vinegar. Both, like vinegar, can be manufactured chemically, but this is rarely practised. Butyric acid is a common ingredient in old milk and butter, and its production by bacteria is historically one of the first bacterial fermentations understood. Moreover, in its investigation Pasteur first brought to light the fact that certain organisms acted only in the absence of oxygen. In studying a drop of butyric fermenting fluid, it was observed that the organisms at the edge of the drop were motionless and apparently dead, whilst in the central portion of the drop the bacilli were executing those active movements which are characteristic of their vitality. To Pasteur's mind this at once suggested what he was able later to demonstrate, namely, that these bacilli were paralysed by contact with oxygen. When he passed a stream of air through a flask containing a liquid in butyric fermentation, he observed the process slacken and eventually cease. So were discovered the anaërobic micro-organisms. The aërobic133 ferments give rise to oxidation of certain products of decomposition; the anaërobic organisms, on the other hand, only commence to grow when the aërobic have used up all the available oxygen. Thus in such fermentations certain bodies (carbohydrates, fatty acids, etc.) undergo decomposition, and by oxidation become carbonic acid gas, and the remainder is left as a "reduced" product of the whole process. Hence sometimes this is termed fermentation by reduction. The chemical formula of this butyric reaction may be expressed thus:—
| C6H12O6(by simple decomposition) = 2 C3H6O3 | |
| Glucose, | Lactic acid. |
which is followed by the fermentation of the lactic acid:—
| 2 C3H6O3 = C4H8O2 + 2 CO2 + 2 H2 | |||
| Lactic acid. | Butyric acid. | Carbonic acid gas. | Free hydrogen. |
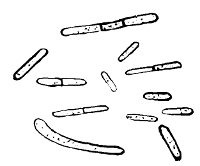
B. Butyricus
Bacillus Butyricus. Long and short rods, generally straight, with rounded ends, single or in chains, reproducing themselves both by fission and spores, and sometimes growing out into long threads, actively motile, anaërobic, and liquefying. The spores are widely distributed in nature, and grow readily on fleshy roots, old cheese, etc. The favourable temperature is blood-heat, and on liquid media they produce a pellicle. The resistant spores are134 irregularly placed in the rod, and may cause considerable variations in morphology. The culture gives off a strong butyric acid odour. It grows most readily at a temperature of about 40° C.
Although, according to Pasteur's researches, the butyric acid ferment performs its functions anaërobically, many butyric organisms can act in the presence of oxygen, and yield somewhat different products.
All of them, however, ferment most actively at a temperature at or about blood-heat, and the spores are able to withstand boiling for from three to twenty minutes (Fitz). It will be observed that as in lactic acid fermentation so in butyric, the results are not due to one species only.
Diseases in Beer. We have seen how a knowledge of fermentation has been compiled by a large number of workers. Spallanzani, Schwann, Pasteur, and Hansen all made epoch-making contributions. In the same way the investigations of diseases in beers and wines were carried out by many observers, and were closely connected with those relating to spontaneous generation and mixed cultures of bacteria in fermentation. These so-called "diseases" are analogous to the taints occurring in milk and due to fermentations. Turning (tourne), turbidity, ropiness, bitterness, acidity, mouldiness, are all terms used to describe these diseases. They are chiefly brought about by four agencies:—
1. Bacteria.
2. Mixed yeasts.
3. "Wild" yeasts.
4. Moulds.
To each species of wild yeast there belongs some taint-producing power in the fermentations for which it is responsible. Saccharomyces ellipsoideus II. and S. pastorianus I., III., are such yeasts; they only produce their diseases when introduced at the commencement of the fermentation.
Saccharomyces pastorianus I. is a low fermentative yeast in elongated cells, producing a bitter taste to beer and an unpleasant odour. It can also produce turbidity. S. pastorianus III. produces turbidity, and S. ellipsoideus II. has a similar effect.
In 1883 Hansen demonstrated that the much-dreaded turbidity and disagreeable tastes and smells in beer may be due to mixture of two yeasts, each of which by itself gives a faultless product.
Industrial Application of Bacterial Ferments. From what has been said we trust it has been made evident that bacteriology has a place of ever-increasing importance in regard to fermentative processes. Not only have the causal agents of various fermentations been isolated and studied, but from their study practical results follow. The question of pure cultures alone is one of practical importance; the recognition of the causes of "diseases" of beer is another.
We cannot enter into a full discussion of the rôle of bacteria in industrial processes, but several of the chief directions may be pointed out. Without exception, bacteria have a part in them on account of their powers of fermentation. In securing their food, bacteria break down material, and bring about chemical and physical change. The power which organisms have of chemically destroying compounds is in itself of little importance, but the products which arise as a result are of an importance in the world which has not hitherto been recognised. We have used bacteria abundantly in the past, but we have not perceived that we were doing so. The maceration industries may be mentioned as illustrative of this use without acknowledgment. The flax stem is made up of cellular substance, flax fibres, and wood fibres; the later are of no service in the making of linen, but the whole is bound together by a gummy, resinous substance. Now this connective element is got rid of in the process of retting. There is dew-retting and water-retting.136 The former is practised in Russia, and consists in spreading the flax on the grass and exposing it to the influence of dew, rain, air, and light. The result is a soft and silky fibre. Water-retting is accomplished by means of steeping the flax in bundles, roots downwards, in tanks or ponds. In ten to fourteen days, according to the weather, fermentation sets in, and breaks the "shore" or "shive" from the fibre, and the process is complete. This is always done by the aid of bacteria, which, under the favourable circumstances, multiply rapidly, and cause decomposition of the pectin resinous matter. The same operation occurs in jute and hemp. Sponges, too, are cleared in this manner by the rotting of the organic matter in their interstices. The preparation of indigo from the indigo plant is brought about by a special bacterium found on the leaves. If the leaves are sterilised, no fermentation occurs, and no indigo is formed. Tobacco-curing is also in part due to decomposition bacteria, and several bacteriologists have experimented independently in fermenting tobacco leaves by the action of pure cultures obtained from tobacco of the finest quality.
In all these applications we have advanced only the first stage of the journey. Nevertheless, here, as in nature on a big scale in the formation of fertile soils and coal-measures, we find bacteria silently at work, achieving great ends by co-operating in countless hordes.
Surface soils and those rich in organic matter supply a varied field for the bacteriologist. Indeed, it may be said that the introduction of the plate method of culture and the improved facilities for growing anaërobic micro-organisms have opened up possibilities of research into soil microbiology unknown to previous generations of workers.
From the nature of bacteria it will be readily understood that their presence is affected by geological and physical conditions of the soil, and in all soils only within a few feet of the surface. As we go down below two feet, bacteria become less, and below a depth of five or six feet we find only a few anaërobes. At a depth of ten feet, and in the "ground water region," bacteria are scarce or absent. This is held to be due to the porosity of the soil acting as a filtering medium. Regarding the numbers of micro-organisms present in soil, no very accurate standard can be obtained. Ordinary earth may yield anything from 10,000 to 5,000,000 per gram, whilst from polluted soil even 100,000,000 per gram have been estimated. These figures are obviously only approximate, nor is an exact standard of any great value. Nevertheless, Fränkel, Beumer, Miquel, and Maggiora have, as the result of experiments, arrived at a number of conclusions respecting bacteria in soil which are of much more practical use. From these results it appears that, in addition to the "ground water region" being free,138 or nearly so, virgin soils contain much fewer than cultivated lands, and these latter, again, fewer than made soils and inhabited localities. In cultivated lands the number of organisms augments with the activity of cultivation and the strength of the fertilisers used. In all soils the maximum occurs in July and August.
But the condition which more than all others controls the quantity and quality of the contained bacteria is the degree and quality of the organic matter in the soil. The quantity of organic matter present in soil having a direct effect upon bacteria will be materially increased by placing in soil the bodies of men and animals after death. Dr. Buchanan Young two or three years ago performed some experiments to discover to what degree the soil bacteria were affected by these means. "The number of micro-organisms present in soil which has been used for burial purposes," he concludes, "exceeds that present in undisturbed soil at similar level, and this excess, though apparent at all depths, is most marked in the lower reaches of the soil."35 The numbers were as follows:—
Virgin soil, 4 ft. 6 in. = 53,436 m.o. per gram of soil.
Burial soil (8 years), 4 ft. 6 in. = 363,411 m.o. per gram of soil.
Burial"lsoil(3 " ), 6 ft. 6 in. = 722,751m.o. pe"per gra"
Methods of Examination of Soil. Two simple methods are generally adopted. The first is to obtain a qualitative estimation of the organisms contained in the soil. It consists simply in adding to test-tubes of liquefied gelatine or broth a small quantity of the sample, finely broken up with a sterile rod. The test-tubes are now incubated at 37° C. and 22° C., and the growth of the contained bacteria observed in the test-tube, or after a plate culture has been made. The second plan is adopted in order to secure more accurate quantitative results. One gram or half-gram of the sample 139is weighed on the balance, and then added to 1000 cc. of distilled sterilised water in a sterilised flask, in which it is thoroughly mixed and washed. From either of these two different sources it is now possible to make sub-cultures and plate cultures. The procedure is, of course, that described under the examination of water (p. 41 et seq.), and Petri's dishes, Koch's plates, or Esmarch's roll cultures are used. Many of the commoner bacteria in soil will thus be detected and cultivated. But it is obvious that this by no means covers the required ground. It will be necessary for us here to consider the methods generally adopted for growing anaërobic bacteria, that is to say those species which will not grow in the presence of oxygen. This anaërobic difficulty may be overcome in a variety of ways.
1. The air contained in the culture tube may be removed by ebullition and rapid cooling. And whilst this may accurately produce a vacuum, it is far from easy to introduce the virus without also reintroducing oxygen.
2. The oxygen may be displaced by some other gas, and though coal-gas, nitrogen, and carbon dioxide may all be used for this purpose, it has become the almost universal practice to grow anaërobes in hydrogen. The production of the hydrogen is readily obtained by Kipp's or some other suitable apparatus for the generation of hydrogen from zinc and sulphuric acid. The free gas is passed through various wash-bottles to purify it of any contaminations. Lead acetate (1–10 per cent. water) removes any traces of sulphuretted hydrogen, silver nitrate (1–10) doing the same for arseniated hydrogen; whilst a flask of pyrogallic acid will remove any oxygen. It is not always necessary to have these three purifiers if the zinc used in the Kipp's apparatus is pure. Occasionally a fourth flask is added of distilled water, and this or a dry cotton wool pledget in the exit tube will ensure germ-free gas. From the further end of the exit tube of the Kipp's apparatus an india-rubber tube will carry140 the hydrogen to its desired destination. With some it is the custom to place anaërobic cultures in test-tubes, and the test-tubes in a large flask having a two-way tube for entrance and exit of the hydrogen; others prefer to pass the hydrogen immediately into a large test-tube containing the culture (Fränkel's method). Either method ends practically the same, and the growth of the culture in hydrogen is readily observed. Yet another plan is to use a yeast flask, and after having passed the hydrogen through for about half an hour, the lateral exit tube is dipped into a small flask containing mercury. The entrance tube is now sealed, and the whole apparatus placed in the incubator. The interior containing the culture is filled with an atmosphere of hydrogen. No141 oxygen can obtain entrance through the sealed entrance tube, or through the exit tube immersed in mercury. Yet through this latter channel any gases produced by the culture could escape if able to produce sufficient pressure.

Kipp's Apparatus
For the Production of Hydrogen
3. The Absorption Method. Instead of adding hydrogen
to the tube or flask containing the anaërobic culture, it is
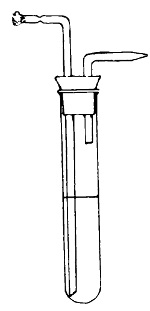 Fränkel's Tube
Fränkel's Tube
For Cultivation of Anaërobes
feasible to add to the medium
some substances, like glucose
 Buchner's
Tube
Buchner's
Tube
For Cultivation
of Anaërobes
or pyrogallic acid, which will
absorb the oxygen which is
present, and thus enable the
anaërobic requirement to be
fulfilled. To various media—gelatine,
agar, or broth (the
latter used for obtaining the
toxins of anaërobes)—2 per
cent. of glucose may be added.
Pyrogallic acid, or pyrogallic
acid one part and 20 per cent.
caustic potash one part, is also
readily used for absorptive purposes.
A large glass tube of
25 cc. height, named a Buchner's
cylinder, having a constriction
near the bottom, is taken; and about
two drachms of the pyrogallic solution are placed
in the bottom of it. A test-tube containing the
culture is now lodged in the upper part above the
constriction. The apparatus is now placed in
the incubator at the desired temperature, and the
contained culture grows under anaërobic conditions.
As the pyrogallic solution absorbs the
oxygen it assumes a darker tint.
4. Mechanical Methods. These include various ingenious tricks for preventing an admittance of oxygen to142 the culture. An old-fashioned one was to plate out the culture and protect it from the air by covering it with a plate of mica. A more serviceable mode is to inoculate, say, a tube of agar with the anaërobic organism, and then pour over the culture a small quantity of melted agar, which will readily set, and so protect the culture itself from the air. Oil may be used instead of melted agar. Another mechanical method is to make a deep inoculation and then melt the top of the medium over a bunsen burner, and thus close the entrance puncture and seal it from the air.
5. Absorption of Oxygen by an Aërobic Culture. This method takes advantage of the power of absorption of certain aërobic bacteria, which are planted over the culture of the anaërobic species. It is not practically satisfactory, though occasionally good results have been obtained.
6. Lastly, there is the Air-pump Method. By this means it is obviously intended to extract air from the culture and seal of it in vacuo. The culture tubes are connected with the air-pump, and exhausted as much as possible.
Of these various methods it is on the whole best to choose either the hydrogen method, the vacuum, or the plan of absorption by grape-sugar or pyrogallic. In anaërobic plate cultures grape-sugar agar plus 0.5 per cent. of formate of soda may be used. The poured inoculated plate should be placed over pyrogallic solution under a sealed bell-glass and incubated at 37° C. Pasteur, Roux, Joubert, Chamberland, Esmarch, Kitasato, and others have introduced special apparatus to facilitate anaërobic cultivation, but the principles adopted are those which have been mentioned.
We may now turn to consider the species of bacteria found in the interstices of soil. They may be classified in five main groups. The division is somewhat artificial, but convenient:
1. The Denitrifying Bacteria. A group whose function has been elucidated in recent years (largely by the investigations of Professor Warington) are held responsible for the breaking down of nitrates. With these may be associated the Decomposition or Putrefactive Bacteria, which break down complex organic products other than nitrates into simpler bodies.

A Method of Growing Cultivations in a Vacuum over Pyrogallic Solution
2. The Organisms of Nitrification. To this group belong144 the two chief types of nitrifying bacteria, viz., those which oxidise ammonia into nitrites, and those which change nitrites into nitrates.
3. The Nitrogen-fixing Bacteria, found mainly in the nodules on the rootlets of certain plants.
4. The Common Saprophytic Bacteria, whose function is at present but imperfectly known. Many are putrefactive germs.
5. The Pathogenic Bacteria. This division includes the three types, tetanus, malignant œdema, and quarter evil. Under this heading we shall also have to consider in some detail the intimate relation between the soil and such important bacterial diseases as tubercle and typhoid.
To enable us to appreciate the work which the "economic bacteria" perform, it will be necessary to consider shortly the place they occupy in the economy of nature. This may be perhaps most readily accomplished by studying the accompanying table (p. 145).
A SCHEME SHOWING THE PLACE AND FUNCTION OF THE ECONOMIC MICRO-ORGANISMS FOUND IN SOIL
| Water | Chemical Substances [Nitrates, etc.] |
Gases [CO2, H, N, O] |
|||
| ⬊ | ⬇ | ⬋ | |||
| Plant Life | |||||
| │ | |||||
| ┌────────┬──────┬──────┬──────────┬───────────┐ | |||||
Carbohydrates [albumoses, sugar, starch, etc.] |
Fats |
Proteids [bodies containing Nitrogen] |
Vegetable Acids |
Mineral Salts |
Water |
| Animal Life | |||||
| │ | |||||
| ┌───────┬──────────┬──────────────────┐ | |||||
Gases [CO2, etc.] |
Water |
Urea, Albuminoids, Ammonia compounds, etc. |
Nitrogen in many forms locked up in the body |
||
 |
|||||
| PUTREFACTIVE AND DENITRIFYING BACTERIA │ |
|||||
| ┌───────┬──────┬──────────┬───────────┐ | |||||
Free Nitrogen │ │ │ │ │ │ │ │ |
Gases [CO2] |
Water |
Ammonia and other elements of broken-down complex bodies. |
[Nitrites] |
|
 |
|||||
| NITRIFYING BACTERIA | |||||
| │ | |||||
| Nitrites[=Nitrous organism │ (Nitrosomans)] |
|||||
| NITROGEN-FIXING BACTERIA |
Nitrates[=Nitric organism (Nitrosomans)] |
||||
| [In soil and in the nodules on the rootlets of Leguminosæ] | [In soil and available for plant life] | ||||
The threefold function of plant life is nutrition, assimilation,
and reproduction: the food of plants, the digestive and
storage power of plants, and the various means they adopt
for multiplying and increasing their species. With the two
latter we have little concern in this place. Respecting the
nutrition of plant life, it is obvious that, like animals, they
must feed and breathe to maintain life. Plant food is of
three kinds, viz., water, chemical substances, and gas. Water
is an actual necessity to the plant not only as a direct food
and food-solvent, but as the vehicle of important inorganic
materials. The hydrogen, too, of the organic compounds
is obtained from the decomposition of the water which permeates
every part of the plant, and is derived by it from the
soil and from the aqueous vapour in the atmosphere. The
chief chemical substances of which vegetable protoplasm
is constituted are six, viz, potassium, magnesium, calcium,
iron, phosphorous, and sulphur. These inorganic
145
146elements do not enter the plant as such, but combined with
other substances or dissolved in water. Potassium occurs in
salt form combined with various organic acids (tartaric,
oxalic, etc.), calcium and magnesium as salts of lime and
magnesia in combination both with organic and inorganic
acids. Iron contributes largely to the formation of the
green colouring matter of plants, and is also derived from
the soil. Phosphorus, one of the chief constituents of seeds,
generally occurs as phosphate of lime. Sulphur, which is
an important constituent of albumen, is derived from the
sulphates of the soil. In addition to the above, there are
other elements, sometimes described as non-essential constituents
of plants. Amongst these are silica (to give stiffness),
sodium, chlorine, iodine, bromine, etc. All these
elements contribute to the formation or quality of the
protoplasm of plants.
The gases essential to plants are four: Carbon dioxide (carbonic acid), Hydrogen, Oxygen, and Nitrogen. By the aid of the green chlorophyll corpuscles, and under the influence of sunlight, we know that leaves absorb the carbon dioxide of the atmosphere, and effect certain changes in it. The hydrogen, as we have seen, is obtained from the water. Oxygen is absorbed through the root from the interstices of the soil. Each of these contributes vitally to the existence of the plant. The fourth gas, nitrogen, which constitutes more than two thirds of the air we breathe (79 per cent. of the total volume and 77 per cent. of the total weight of the atmosphere), is, perhaps, the most important food required by plants. Yet, although this is so, the plant cannot absorb or obtain its nitrogen in the same manner in which it acquires its carbon—viz., by absorption through the leaves—nor can the plant take nitrogen into its own substance by any means as nitrogen, with the exception of the flesh-feeding plants (insectivorus). Hence, although this gas is present in the atmosphere surrounding the plant, the147 plant will perish if nitrogen does not exist in some combined form in the soil. Nitrates and compounds of ammonia are widely distributed in nature, and it is from these bodies that the plant obtains, by means of its roots, the necessary nitrogen.
Until comparatively recently it was held that plant life could not be maintained in a soil devoid of nitrogen or compounds thereof. But it has been found that certain classes of plants (the Leguminosæ, for example), when they are grown in a soil which is practically free from nitrogen at the commencement, do take up this gas into their tissues. One explanation of this fact is that free nitrogen becomes converted into nitrogen compounds in the soil through the influence of micro-organisms present there. Another explanation attributes this fixation of free nitrogen to micro-organisms existing in the rootlets of the plant. These two classes of organisms, known as the nitrogen-fixing organisms, will require our consideration at a later stage. Here we merely desire to make it clear that the main supply of this gas, absolutely necessary to the existence of vegetable life upon the earth, is drawn not from the nitrogen of the atmosphere, but from that contained in nitrogen compounds in the soil. The most important of these are the nitrates. Here then we have the necessary food of plants expressed in a sentence: water, gases, salts, the most important and essential gas and some of the salts being combined in nitrates.
Plant life seizes upon its required constituents, and by means of the energy furnished by the sun's rays builds these materials up into its own complex forms. Its many and varied forms fulfil a place in beautifying the world. But their contribution to the economy of nature is, by means of their products, to supply food for animal life. The products of plant life are chiefly sugar, starch, fat, and proteids. Animal life is not capable of extracting its nutriment from soil, but it must take the more complex foods which have148 already been built up by vegetable life. Again, the complementary functions of animal and vegetable life are seen in the absorption by plants of one of the waste materials of animals, viz., carbonic acid gas. Plants abstract from this gas carbon for their own use, and return the oxygen to the air, which in its turn is of service to animal life.
By animal activity some of these foods supplied by the vegetable kingdom are at once decomposed into carbonic acid gas and water, which goes back to nature. Much, however, is built up still further into higher and higher compounds. The proteids are converted by digestion into albumoses and peptones, ultimately entirely into peptones; these in their turn are reconverted into proteids, and become assimilated as part of the living organism. In time they become further changed into carbonic acid, sulphuric acid, water, and certain not fully oxidised products,36 which contain the nitrogen of the original proteid. In the table these bodies have been represented by one of their chief members, viz., urea.
It is clear that there is in all animal life a double process continually going on; there is a building up (anabolism, assimilation), and there is a breaking down (katabolism, dissimilation). These processes will not balance each other throughout the whole period of animal life. We have, as possibilities, elaboration, balance, degeneration; and the products of animal life will differ in degree and in substance according to which period is in the predominance. These products we may subdivide simply into excretions during life and final materials of dissolution after death, both of which may be used more or less immediately by other forms of animal or vegetable life, or mediately after having passed to the soil. We may shortly summarise the final products of animal life as carbonic acid, water, and nitrogenous remnants. These latter will occur as urea, new albumens, com149pounds of ammonia, and nitrogen compounds of great complexity stored up in the tissues and body of the animal. The carbonic acid, water, and other simple substances like them will return to nature and be of immediate use to vegetable life. But otherwise the cycle cannot be completed, for the more complex bodies are of no service as such to plants or animals.
1. In order that this complex material should be of service in the economy of nature, and its constituents not lost, it is necessary that it should be broken down again into simpler conditions. This prodigious task is accomplished by the agency of two groups of organisms, the decomposition and denitrifying37 bacteria. The organisms associated with decomposition processes are numerous; some denitrify as well as break down organic compounds. This group will be referred to under "Saprophytic Bacteria." The reduction by the denitrifying bacteria may be simply from nitrate to nitrite, or from nitrate to nitric or nitrous oxide gas, or indeed to nitrogen itself. In all these processes of reduction the rule is that a loss of nitrogen is involved. How that free nitrogen is brought back again and made subservient to plants and animals we shall understand at a later stage.
Professor Warington has again recently set forth the chief facts known of this decomposition process.38 That the action in question only occurs in the presence of living organisms was first established by Mensel in 1875 in natural waters, and by Macquenne in 1882 in soils. If all living organisms are destroyed by sterilisation of the soil, denitrification cannot take place, nor can vegetable life exist. "Bacteria reduce nitrates," says Professor Warington, "by bringing about the combustion of organic matter by the oxygen of the nitrate, the temperature distinctly rising 150during the operation." The reduction to a nitrite is a common property of bacteria. But only a few species have the power of reducing a nitrate to gas. These few species are, however, widely distributed. In 1886 Gayon and Dupetit first isolated the bacteria capable of reducing nitrates to the simplest element, nitrogen. They obtained their species from sewage, but ten years later denitrifying bacteria were isolated from manure. That soil contains a number of these reducing organisms is known by introducing a particle of surface soil into some broth, to which has been added one per cent. of nitre. During incubation of such a tube gas is produced, and the nitrate entirely disappears.
Whenever decomposition occurs in organic substances there is a reduction of compound bodies, and in such cases the putrefying substances obtain their decomposing and denitrifying bacteria from the air. The chief conditions requisite for bringing about a loss of nitrogen by denitrification are enumerated by Professor Warington as follows: (1) the specific micro-organism; (2) the presence of a nitrate and suitable organic matter; (3) such a condition as to aëration that the supply of atmospheric oxygen shall not be in excess relatively to the supply of organic matter; (4) the usual essential conditions of bacterial growth. "Of these," he says, "the supply of organic matter is by far the most important in determining the extent to which denitrification will take place." The necessarily somewhat unstable condition facilitates its being split up by means of bacteria. The bacteria in their turn are ready to seize upon any products of animal life which will serve as their food. Thus, by reducing complex bodies to simple ones, these denitrifying organisms act as the necessary link to connect again the excretions of the animal body, or after death the animal body itself, with the soil.
In a book of this nature it has been deemed advisable not to enter into minute description of all the species of bacteria151 mentioned. Some of the chief are described more or less fully. We cannot, however, do more than name several of the chief organisms concerned in reducing and breaking down compounds. As we shall find in the bacteria of nitrification, so also here, the entire process is rarely, if ever, performed by one species. There is indeed a remarkable division of labour, not only between decomposition bacteria and denitrification bacteria, but between different species of the same group. Bacillus fluorescens non-liquefaciens, Mycoderma ureæ, and some of the staphylococci break down nitrates (denitrification), and also decompose other compound bodies. Amongst the group of putrefactive bacteria found in soil may be named B. coli, B. mycoides, B. mesentericus, B. liquidus, B. prodigiosus, B. ramosus, B. vermicularis, B. liquefaciens, and many members in the great family of Proteus. Some perform their function in soil, others in water, and others, again, in dead animal bodies. Dr. Buchanan Young, to whose researches in soil we have referred, has pointed out that in the upper reaches of burial soil, where these bacteria are most largely present, there is as a result no excess of organic carbon and nitrogen. Even in the lower layers of such soil it is rapidly broken down.

Micrococcus from Soil
It will be observed, from a glance at the table, that the chief results of decomposition and denitrification are as fol152lows: free nitrogen, carbonic acid, gas and water, ammonia bodies, and sometimes nitrites. The nitrogen passes into the atmosphere, and is "lost"; the carbonic acid and water return to nature and are at once used by vegetation. The ammonia and nitrites await further changes. These further changes become necessary on account of the fact, already discussed, that plants require their nitrogen to be in the form of nitrates in order to use it. Nitrates obviously contain a considerable amount of oxygen, but ammonia contains no oxygen, and nitrites very much less than nitrates. Hence a process of oxidation is required to change the ammonia into nitrites and the nitrites into nitrates.
2. This oxidation is performed by the nitrifying micro-organisms, and the process is known as nitrification. It should be clearly understood that the process of nitrifaction may, so to speak, dovetail with the process of denitrification. No exact dividing line can be drawn between the two, although they are definite and different processes. In a carcass, for example, both processes may be going on concomitantly; so also in manure. There is no hard and fast line to be drawn in the present state of our knowledge. Other organisms beside the true nitrification bacteria may be playing a part, and it is impossible exactly to measure the action of the latter, where they began and where the preliminary attack upon the nitrogenous compounds terminated. In all cases, however, according to Professor Warington, the formation of ammonia has been found to precede the formation of nitrous or nitric acid.
It was Pasteur who (in 1862) first suggested that the production of nitric acid in soil might be due to the agency of germs, and it is to Schlösing and Müntz that the credit belongs for first demonstrating (in 1877) that the true nature of nitrification depended upon the activity of a living microorganism. Partly by Schlösing and Müntz and partly by Warington (who was then engaged in similar work at Roth153amsted), it was later established (1) that the power of nitrification could be communicated to substances which did not hitherto nitrify by simply seeding them with a nitrified substance, and (2) that the process of nitrification in garden soil was entirely suspended by the vapour of chloroform or carbon disulphide. The conditions for nitrification, the limit of temperature, and the necessity of plant food, have furnished additional proof that the process is due to a living organism. These conditions are briefly as follows:
1. Food (of which phosphates are essential constituents). "The nitrifying organism can apparently feed upon organic matter, but it can also, apparently with equal ease, develop and exercise all its functions with purely inorganic food" (Warington).
Winogradsky prepared vessels and solutions carefully purified from organic matter, and these solutions he sowed with the nitrifying organism, and found that they flourished. Professor Warington has employed the acid carbonates of sodium and calcium with distinct success as ingredients of an ammoniacal solution undergoing nitrification.
2. The next condition of nitrification is the presence of oxygen. Without it the reverse process, denitrification, occurs, and instead of a building up we get a breaking down, with an evolution of nitrogen gas. The amount of oxygen present has an intimate proportion to the amount of nitrification, and with 16 to 21 per cent. of oxygen present the nitrates are more than four times as much as when the smallest quantity of oxygen is supplied. The use of tillage in promoting nitrification is doubtless in part due to the aëration of the soil thus obtained.
3. A third condition is the presence of a base with which nitric acid when formed may combine. Nitrification can take place only in a feebly alkaline medium, but an excess of alkalinity will retard the process.
4. The last essential requirement is a favourable temper154ature. The nitrifying organism can act at a temperature as low as 37° or 39° F. (3–4° C.), but at a higher temperature it becomes much more active. According to Schlösing and Müntz, at 54° F. (12° C.) nitrification becomes really active, and it increases as the temperature rises to 99° F. (37° C.), after which it falls. A high temperature or a strong light are prejudicial to the process.
We are now in a position to consider shortly some of the characters of these nitrification bacteria. They may readily be divided into two chief groups, not in consideration of their form or biological characteristics, but on account of the duties which they perform. Just as we observed that there were few denitrifying organisms which could break down ammonia compounds to nitrogen gas, so is it also true that there are few nitrifying bacteria which can build up from ammonia to the nitrates. Nature has provided that this shall be accomplished in two stages, viz., a first stage from ammonia bodies to nitrites, and a second stage from nitrites to nitrates. The agent of the former is termed the nitrous organism, the latter the nitric organism. Both are contributing to the final production of nitrates which can be used by plant life.39
The Nitrous Organism (Nitrosomonas). Prior to Koch's gelatine method the isolation of this bacterium proved an exceedingly difficult task. But even the adoption of this isolating method seemed to give no better results, and for an excellent reason: the nitrifying organisms will not grow on gelatine. To Winogradsky and Percy Frankland belongs the credit of separately isolating the nitrous organism on the surface of gelatinous silica containing the necessary inorganic 155food. Professor Warington, in his lectures under the Lawes Agricultural Trust, has described this important germ as follows:
"The organism as found in suspension in a freshly nitrified solution consists largely of nearly spherical corpuscles, varying extremely in size. The largest of these corpuscles barely reaches a diameter of one-thousandth of a millimetre, and some are so minute as to be hardly discernible in photographs. The larger ones are frequently not strictly circular, and are sometimes seen in the act of dividing.
"Besides the form just described, there is another, not universally present in solutions, in which the length is considerably greater than its breadth. The shape varies, being occasionally a regular oval, but sometimes largest at one end, and sometimes with the ends truncated. The circular organisms are probably the youngest.
"This organism grows in broth, diluted milk, and other solutions without producing turbidity. When acting on ammonia it produces only nitrites. It is without action on potassium nitrite. It is, in fact, the nitrous organism which, as we have previously seen, may be separated from soil by successive cultivations in ammonium carbonate solution."
The elongated forms appear to be a sign of arrested growth. Normally the organ is about 1.8 µ long, or nearly three times as long as the nitric organism. It possesses a gelatinous capsule. "The motile cells, stained by Löffler's method, are seen to have a flagellum in the form of a spiral." When grown on silica the nitrous organism appears in the same two forms—zooglea and free cells—as when cultivated in a fluid. It commences to show growth in about four days, and is at its maximum on about the tenth day. Winogradsky found that there were considerable differences in the morphology of the organism according to the soil from which it was taken. One of the Java soils he investigated contained a nitrous organism having a spiral156 flagellum of thirty micromillimetres; but its movement was slow.
As we have already seen, the most astonishing property of this organism is its ability to grow and perform its specific function in solutions absolutely devoid of organic matter. Some authorities hold that it acquires its necessary carbon from carbonic acid. The mode of culturing it is as follows:
To sterilised flasks add 100 cc. of a solution made of one gram of ammonium sulphate, one gram of potassium sulphate, and 1000 cc. of pure water. To this add one gram of basic magnesium carbonate which has been previously sterilised by boiling. Now inoculate the flask with a small portion of the soil under investigation, and after four or five days sub-culture on the same medium in fresh flasks, and let this be repeated half a dozen times. Now, as this inorganic medium is unfavourable to ordinary bacteria of soil, it is clear that after several sub-cultures the nitrous organism will be isolated in pure culture.
Winogradsky employs for culturing upon solid media a mineral gelatine. A solution of from 3 to 4 per cent. of silicic acid in distilled water is placed in flasks. By the addition of the following salts to such a solution gelatinisation occurs:
| (a) | |
Ammonium sulphate | 0.4 gram |
| Magnesium sulphate | 0.05 " | ||
| Calcium chloride | A trace | ||
| (b) | |
Potassium phosphate | 0.1 gram |
| Sodium carbonate | 0.6, 0.9 " | ||
| Distilled water | 100 cc. |
The sulphates and chloride are mixed in 50 cc. of distilled water, and the latter substance in the remaining 50 cc. in separate flasks. After sterilisation and cooling these are all mixed and added in small quantities to the silicic acid.
Upon this medium it is possible to sub-culture a pure growth from the film at the bottom of the flasks in which the nitrous organism is first isolated.
The Nitric Organism. It was soon learned that the nitrous organism, even when obtainable in large quantities and in pure culture, was not able entirely to complete the nitrifying process. As early as 1881 Professor Warington had observed that some of his cultures, though capable of changing nitrites into nitrates, had no power of oxidising ammonia. These he had obtained from advanced sub-cultures of the nitrous organism, and somewhat later Winogradsky isolated and described this companion of the nitrous organism. It develops freely in solutions to which no organic matter has been added; indeed, much organic matter will prevent its growing. He isolated it from soils from various parts of the world on the following media:
| Water | 1000.0 |
| Potassium phosphate | 1.0 |
| Magnesium sulphate | 0.5 |
| Calcium chloride | A trace |
| Sodium chloride | 2.0 |
About 20 cc. of this solution is placed in a flat-bottom flask, and a little freshly washed magnesium carbonate is added. The flask is closed with cotton wool, and the whole is sterilised. To each flask 2 cc. of a 2 per cent. solution of ammonium sulphate is subsequently added. The temperature for incubation is 30° C. Winogradsky concluded that the oxidation of nitrites to nitrates was brought about by a specific organism independently of the nitrous organism. He successfully isolated it in silica jelly. He believes the organism, like its companion, derives its nutriment solely from inorganic matter, but this is not finally established.
The form of the nitric organism (or nitromonas, as it158 was once termed) is allied to the nitrous organism. The cells are elongated, rarely oval, but sometimes pear-shaped. They are more than half a micromillimetre in length, and somewhat less in thickness. The cells have a gelatinous membrane. Like the other nitrifying bacteria, its development and action are favoured by the presence of the acid carbonates of calcium and sodium. Of the latter, six grams per litre or even a smaller quantity gives good results. The sulphate of calcium can be used, but the organism prefers the carbonates. The differences between these two bacteria are small, with the exception of their chemical action. The nitric organism has no action upon ammonia, and the presence of any considerable amount of ammonium carbonate hinders its development and prevents its action on a nitrite.40
We may here summarise the general facts respecting nitrification. Winogradsky proposes to term the group nitro-bacteria, and to classify thus:
| Nitrous organisms | = | |
Nitrosomonas, containing at least two species, viz., the European and the Java. |
| Nitrosococcus. | |||
| Nitric organism | = | Nitrobacter. |
Nitrification occurs in two stages, each stage performed by a distinct organism. By one (nitrosomonas) ammonia is converted into nitrite; by the other (nitrobacter) the nitrite is converted into nitrate.41 Both organisms are widely and 159abundantly distributed in the superficial soils. They act together and in conjunction, and for one common purpose. They are separable by employing favourable media.
"If we employ a suitable inorganic solution containing potassium nitrite, but no ammonia, we shall presently obtain the nitric organism alone, the nitrous organism feeding on ammonia being excluded. If, on the other hand, we employ an ammonium carbonate solution of sufficient strength, we have selected conditions very unfavourable to the growth of the nitric organism, and a few cultivations leave the nitrous organism alone in possession of the field" (Warington).
 Nitrous Organism |
 Nitric Organism |
 |
 |
| Nitrogen Fixing Organism from Secretion of Root-Nodules × 1000 |
|
A word upon the natural distribution of these nitrifying bacteria before we leave them. They belong to the soil, river water, and sewage. They are also said to be frequently present in well water. From some experiments at Rothamsted it appears that the organisms occur mostly in the first twelve inches, and in subsoils of clay down to three or four feet. In sandy soils nitrification may probably occur at a greater depth. These facts should be borne in mind when arranging for the purification of sewage by intermittent filtration.
We have now given some consideration to the chief events in the life-cycle of nature depicted in the table. There is but one further process in which bacteria play a part, and which requires some mention. It will have been noticed that at certain stages in the cycle there is more or less appreciable "loss" of free nitrogen. In the process of decomposition brought about by the denitrifying bacteria, a very considerable portion of the nitrogen is dissipated 160into the air in the form of a free gas. This is the last stage of all proteid decomposition, so that wherever putrefaction is going on there is a continual "loss" of an element essential to life. Thus it would appear at first sight that the sum-total of nitrogen food must be diminishing.
But there are other ways also in which nitrogen is being set free. In the ordinary processes of vegetation there is a gradual draining of the soil and a passing of nitrogen into the sea; the products of decomposition pass from the soil by this drainage, and are "lost" as far as the soil is concerned. Many of the methods of sewage disposal are in reality depriving the land of the return of nitrogen which is its necessity. Again, nitrogen is freed in explosions of gunpowder, nitroglycerine, and dynamite, for whatever purpose they are used. Hence the great putrefactive "loss" of nitrogen, with its subsidiary losses, contributes to reduce this essential element of all life, and if there were no method of bringing it back again to the soil, it would seem that plant life, and therefore animal life, would speedily terminate.
It is at this juncture, and to perform this vital function, that the nitrogen-fixing bacteria play their wonderful part: they bring back the free nitrogen and fix it in the soil. Excepting a small quantity of combined nitrogen coming down in rain and in minor aqueous deposits from the atmosphere, the great source of the nitrogen of vegetation is the store in the soil and subsoil, whether derived from previous accumulations or from recent supplies by manure.
Sir William Crookes has recently42 pointed out the vast importance of using all the available nitrogen in the service of wheat production. The distillation of coal in the process of gas-making yields a certain amount of its nitrogen in the form of sulphate of ammonia, and this, like other nitrogenous manures, might be used to give back to the soil 161some of the nitrogen drained from it. But such manuring cannot keep pace, according to Sir W. Crookes, with the present loss of fixed nitrogen from the soil. We have already referred to several ways in which "loss" of nitrogen occurs. To these may well be added the enormous loss occurring in the waste of sewage when it is passed into the sea. As the President of the British Association pointed out,43 the more widely this wasteful system is extended, recklessly returning to the sea what we have taken from the land, the more surely and quickly will the finite stocks of nitrogen, locked up in the soils of the world, become exhausted. Let us remember that the plant creates nothing in this direction; there is nothing in wheat which is not absorbed from the soil, and unless the abstracted nitrogen is returned to the soil, its fertility must be ultimately exhausted. When we apply to the land sodium nitrate, sulphate of ammonia, guano, and similar manurial substances, we are drawing on the earth's capital, and our drafts will not be perpetually responded to.44 We know that a virgin soil cropped for several years loses its productive powers, and without artificial aid becomes unfertile. For example, through this exhaustion forty bushels of wheat per acre have dwindled to seven. Rotation of crops is an attempt to meet the problem, and the four-course rotation of turnips, barley, clover, and wheat witnesses to the fact that practice has been ahead of science in this matter.
The store of nitrogen in the atmosphere is practically unlimited, but it is fixed and rendered assimilable only by cosmic processes of extreme slowness. We may shortly glance at these, for it is upon these processes, plus a return 162to the soil of sewage, that we must depend in the future for storing nitrogen as nitrates.
1. Some combined nitrogen is absorbed by the soil or plant from the air, for example, fungi, lichens, and some algæ, and the absorption is in the form of ammonia and nitric acid. This is admittedly a small quantity.
2. Some free nitrogen is fixed within the soil by the agency of porous and alkaline bodies.
3. Some, again, may be assimilated by the higher chlorophyllous plants themselves, independently of bacteria (Frank).
4. Electricity fixes, and may in the future be made to fix more, nitrogen. If a strong inductive current be passed between terminals, the nitrogen from the air enters into combination with the oxygen, producing nitrous and nitric acids.
5. Abundant evidence has now been produced in support of the fact that there is considerable fixation by means of bacteria.
Bacterial life in several ways is able to reclaim from the
atmosphere this free nitrogen, which would otherwise be
lost. The first method to which reference may be made is
that involving symbiosis. This term signifies "a living
together" of two different forms of life, generally for a
specific purpose. It may be to mutual advantage, a living
for one another, or it may be, by means of an interchange
of metabolism or products, finally to produce or obtain some
remote chemical result. It is convenient to restrict the term
symbiosis to complementary partnerships such as exist between
algoid and fungoid elements in lichens, or between
unicellular algæ and Radiolarians,45 or between bacteria and
higher plants. The partnerships between hermit crabs and
sea-anemones and the like are sometimes defined by the term
commensalism (joint diet). Symbiosis and commensalism
must be distinguished from parasitism, which indicates that
163all the advantage is on the side of the parasite, and nothing
but loss on the side of the host. The distinction between
symbiosis and commensalism cannot be rigid, but between
these conditions which are advantageous to the partners and
parasitism, there is an obvious and radical difference. Association
of organisms together for increase of virulence and
function should be distinguished from symbiosis, and mere
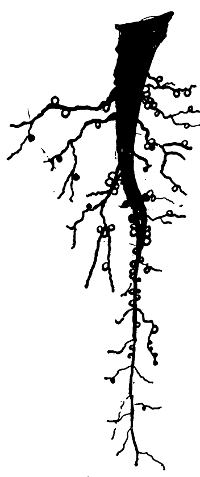 Rootlet of Pea with Nodules
existence of two or more species
of bacteria in one medium is
not, of course, symbiosis. Most
frequently such a condition
would result in injury and the
subsequent death of the weaker
partner, an effect precisely opposite
to that defined by this
term.
Rootlet of Pea with Nodules
existence of two or more species
of bacteria in one medium is
not, of course, symbiosis. Most
frequently such a condition
would result in injury and the
subsequent death of the weaker
partner, an effect precisely opposite
to that defined by this
term.
The example of bacteriological symbiosis with which we are concerned here is that partnership between bacteria and some of the higher plants (Leguminosæ) for the purpose of fixing nitrogen in the plant and in the surrounding soil.
The Nitrogen-fixing Bacteria, the third group of micro-organisms connected with the soil, exist in groups and colonies situated inside the nodules appearing, under certain circumstances, on the rootlets of the pea, bean, and other Leguminosæ. It was Hellriegel and Wilfarth who first pointed out that, although the higher chlorophyllous plants could not directly obtain or utilise free164 nitrogen, some of them at any rate could acquire nitrogen brought into combination under the influence of bacteria. Hellriegel found that the gramineous, polygonaceous, cruciferous, and other orders depended upon combined nitrogen supplied within the soil, but that the Leguminosæ did not depend entirely upon such supplies.
It was observed that in a series of pots of peas to which no nitrogen was added most of the plants were apparently limited in their growth by the amount of nitrogen locked up in the seed. Here and there, however, a plant, under apparently the same circumstances, grew luxuriantly and possessed on its rootlets abundant nodules. The experiments of Sir John Lawes and Sir Henry Gilbert at Rothamsted46 demonstrated further that under the influence of suitable microbe-seeding of the soil in which Leguminosæ were planted there is nodule formation on the roots, and coincidentally increased growth and gain of nitrogen beyond that supplied either in the soil or in the seed as combined nitrogen. Presumably this is due to the fixation, in some way, of free nitrogen. Nobbe proved the gain of nitrogen by non-leguminous plants (Elœagnus, etc.) when these grow root nodules containing bacteria, but to all appearances, bacteria differing morphologically from the Bacillus radicicola of the leguminous plants.
 |
—Cellular sheath of Rootlet forming capsule of nodule. —Colonies of bacteria in situ. |
| Nitrogen Fixing Bacteria in Situ in Nodule on Rootlet of Pea × 400 |
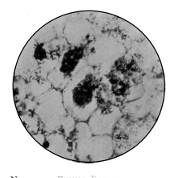 |
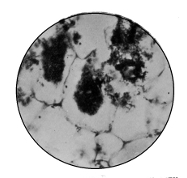 |
Nitrogen Fixing Bacteria in Situ |
Nitrogen Fixing Bacteria in Situ |
These facts being established, the question naturally arises, How is the fixation of nitrogen to be explained, and by what species of bacteria is it performed? In the first place, these matters are simplified by the fact that there is very little fixation indeed by bacteria in the soil apart from symbiosis with higher plants. Hence we have to deal mainly with the work of bacteria in the higher plant. Sir Henry Gilbert concludes47 that the alternative explanations 165of the fixation of free nitrogen in the growth of Leguminosæ seem to be:
"1. That under the conditions of symbiosis the plant is enabled to fix the free nitrogen of the atmosphere by its leaves;
"2. That the nodule organisms become distributed within the soil and there fix free nitrogen, the resulting nitrogenous compounds becoming available as a source of nitrogen to the roots of the higher plant;
"3. That free nitrogen is fixed in the course of the development of the organisms within the nodules, and that the resulting nitrogenous compounds are absorbed and utilized by the host." "Certainly," he adds, "the balance of evidence at present at command is much in favour of the third mode of explanation."
If this is finally proved to be the case, it will furnish another excellent example of the power existing in bacteria of assimilating an elementary substance.
Most authorities would agree that all absorption of free nitrogen, if by means of bacteria, must be through the roots. As a matter of fact, legumes, especially when young, use nitrogen, like all other plants, derived from the soil. It has been pointed out that, unless the soil is somewhat poor in nitrogen, there appears to be but little assimilation of free nitrogen and but a poor development of root nodules.48 The free nitrogen made use of by the micro-organism is in the air contained in the interstices of the soil. For in all soils, but especially in well-drained and light soils, there is a large quantity of air. Although it is not known how the micro-organisms in legumes utilise free nitrogen and convert it into organic compounds in the tissues of the rootlet or plant, it is known that such nitrogen compounds migrate into the stem and leaves, and so make the roots really poorer 166in nitrogen than the foliage. But the ratio is a fluctuating one, depending chiefly on the stage of growth or maturity of the plant.
If the nodules from the rootlets of Leguminosæ be examined, the nitrogen-fixing bacteria can be readily seen. The writer has isolated these and grown them in pure culture as follows: The nodules are removed, if possible at an early stage in their growth, and placed for a few minutes in a steam steriliser. This is advisable in order to remove the various extraneous organisms attached to the outer covering of the nodule. They may then be washed in antiseptic solution, and their capsules softened by soaking. When opened with a sterile knife, thick creamy matter exudes. On microscopic examination this is found to be densely crowded with small round-ended bacilli or oval bodies, known as bacteroids. By a simple process of hardening and using the microtome, excellent sections of the nodules can be obtained which show these bacteria in situ. In the central parts of the section may be seen densely crowded colonies of the bacteria, which in some cases invade the cellular capsule of the nodule derived from the rootlet. Aërobic and anaërobic pure cultures of these bacteria were made. In some cases these cultures very closely resembled the feathery growth of the bacillus of anthrax.
4. The Saprophytic Bacteria in Soil. This group of micro-organisms is by far the most abundant as regards number. They live on the dead organic matter of the soil, and their function appears to be to break it down into simpler constitution. Specialisation is probably progressing among them, for their name is legion, and the struggle for existence keen. After we have eliminated the economic bacteria, most of which are obviously saprophytes, the group is greatly reduced. It is also needless to add that of the remnant little beyond morphology is known, for as their function is learned they are classified otherwise. It is prob167able, as suggested, that many of the species of common saprophytes normally existent in the soil act as auxiliary agents to denitrification and putrefaction. At present we fear they are disregarded in equal measure, and for the same reasons, as the common water bacteria. An excess of either, in soil or water, is not of itself injurious as far as we know; indeed, it is probably just the reverse. It is, however, frequently an index of value as to the amount and sometimes condition of the contained organic matter. The remarks made when considering water bacteria apply here also, viz., that an excess of saprophytes acts not only as index of increase of organic matter, but as at first auxiliary, and then detrimental, to pathogenic organisms. It will require accurate knowledge of soil bacteria generally to be able to say which saprophytic germs, if any, have no definite function beyond their own existence. It may be doubted whether the stern behests of nature permit of such organisms. However that may be, we may feel confident, though at present there are many common bacteria in soil, as also in water, the life object of which is not ascertained, that as knowledge increases and becomes more accurate this special provisional group will become gradually absorbed into other groups having a part in the economy of nature, or in the production of disease. At present the decomposition, denitrifying, nitrifying,49 and nitrogen-fixing organisms are the only saprophytes which have been rescued from the oblivion of ages, and brought more or less into daylight. It is but our lack of knowledge which requires the present division of saprophytes whose business and place in the world is unknown.
5. The Pathogenic Organisms found in Soil. In addition to these saprophytes and the economic bacteria, there are, as is now well known, some disease-producing bacteria find168ing their nidus in ordinary soil. The three chief members of this group are the bacillus of Tetanus (lockjaw), the bacillus of Quarter Evil, and the bacillus of Malignant Œdema.
Tetanus. The pathology of this terrible disease has during recent years been considerably elucidated. It was the custom to look upon it as "spontaneous," and arising no one knew how; now, however, after the experiments of Sternberg and Nicolaier, the disease is known to be due to a micro-organism common in the soil of certain localities, existing there either as a bacillus or in a resting stage of spores. Fortunately Tetanus is comparatively rare, and one of the peculiar biological characteristics of the bacillus is that it grows only in the absence of oxygen. This fact contributed not a little to the difficulties which were met with in securing its isolation.
Tetanus occurs in man and horses most commonly, though it may affect other animals. There is usually a wound, often an insignificant one, which may occur in any part of the body. The popular idea that a severe cut between the thumb and the index finger leads to tetanus is without scientific foundation. As a matter of fact, the wound is nearly always on one or other of the limbs, and is infected simply because they come more into contact with soil and dust than does the trunk. It is not the locality of the wound nor its size that affects the disease. A cut with a dirty knife, a gash in the foot from the prong of a gardener's fork, the bite of an insect, or even the prick of a thorn have before now set up tetanus. Wounds which are jagged, and occurring in absorptive tissues, are those most fitted to allow the entrance of the bacillus. The wound forms a local manufactory, so to speak, of the bacillus and its secreted poisons; the bacillus always remains in the wound, but the toxins may pass throughout the body, and are especially absorbed by the cells of the central nervous system, and thus give rise169 to the spasms which characterise the disease. Suppuration generally occurs in the wound, and in the pus thus produced may be found a great variety of bacteria, as well as the specific agent itself. After a few days or, it may be, as much as a fortnight, when the primary wound may be almost forgotten, general symptoms occur. Their appearance is often the first sign of the disease. Stiffness of the neck and facial muscles, including the muscles of the jaw, is the most prominent sign. This is rapidly followed by spasms and local convulsions, which, when affecting the respiratory or alimentary tract, may cause a fatal result. Fever and increased rate of pulse and respiration are further signs of the disease becoming general. After death, which results in the majority of cases, there is very little to show the cause of fatality. The wound is observable, and patches of congestion may be found on different parts of the nervous system, particularly the medulla (grey matter), pons, and even cerebellum. Evidence has recently been forthcoming at the Pasteur Institute to support the theory that tetanus is a nervous disease, more or less allied to rabies, and is best treated by intra-cerebral injection of antitoxin, which then has an opportunity of opposing the toxins at their favourite site. (Roux and Borrel.)
In the wound the bacillus is present in large numbers, but mixed up with a great variety of suppurative bacteria and extraneous organisms. It is in the form of a straight short rod with rounded ends, occurring singly or in pairs of threads, and slightly motile. It has been pointed out that by special methods of staining, flagella may be demonstrated.50 These are both lateral and terminal, thin and thick, and are shed previously to sporulation. Branching also has been described. Indeed, it would appear that, like the bacillus of tubercle, this organism has various pleomorphic forms. Next to the ordinary bacillus, filamentous 170forms predominate, particularly so in old cultures. Clubbed forms, not unlike the bacillus of diphtheria, may often be seen from agar cultures. Without doubt the most peculiar characteristic of this bacillus is its sporulation. The well-formed round spores occur readily at incubation temperature. They occupy a position at one or other pole of the bacillus, and have a diameter considerably greater than the rod. Thus the well-known "drumstick" form is produced. In practice the spores occur freely in the medium and in microscopical preparation. Like other spores, they are extremely resistant to heat, desiccation, and antiseptics. They can resist boiling for several minutes.

Bacillus of Tetanus
As we have seen, this bacillus is a strict anaërobe, growing only in the absence of oxygen. The favourable temperature is 37° C., and it will only grow very slowly at or below room temperature.
An excellent culture is generally obtainable in glucose gelatine. The growth occurs, of course, only in the depth of the medium, and appears as fine threads passing horizontally outwards from the track of the needle. At the top and bottom of the growth these fibrils are shorter than at the middle or somewhat below the middle. For extraction of the soluble products of the bacillus glucose broth may be used.
In some countries, and in certain localities, the bacillus of tetanus is a very common habitant of the soil, and when one thinks how frequently wounds must be more or less contaminated with such soil, the question naturally arises, How is it that the disease is, fortunately, so rare? Probably we must look to the advance of bacteriological science to answer this and similar questions at all adequately. Much has recently been done in Paris and elsewhere to emphasise the relation which other organisms have to such bacteria as those of typhoid and tetanus. When considering typhoid, we saw that in addition to the presence of the specific germ other conditions were requisite before the disease actually occurred. So in tetanus, Kitasato and others have pointed out that the presence of certain other bacteria, or of some foreign body, is necessary to the production of the disease. The common organisms of suppuration are particularly accused of increasing the virulence of the bacillus of tetanus. How these auxiliary organisms perform this function has not been fully elucidated. Probably, however, it is by damaging the tissues and weakening their resistance to such a degree as to afford a favourable multiplying ground for the tetanus. It is right to state that some authorities hold that they act by using up the surrounding oxygen, and so favouring the growth of tetanus.
Quarter Evil (or symptomatic anthrax) is a disease of animals, produced in a manner analogous to tetanus. It is characterised by a rapidly increasing swelling of the upper parts of the thigh, sacrum, etc., which, beginning locally, may attain to extraordinary size and extent. It assumes a dark colour, and crackles on being touched. There is high temperature, and secondary motor and functional disturbances. The disease ends fatally in two or three days.
Slight injuries to the surface of the skin or mucous membrane are sufficient for the introduction of the causal bacillus. This organism is, like tetanus, an anaërobe, existing172 in the superficial layers of the soil. From its habitat it readily gains entrance to animal tissues. It has spores, but though they are of greater diameter than the bacillus itself they are not absolutely terminal. Hence they merely swell out the capsule of the bacillus, and produce a club-shaped rod. They form gas while growing in the tissues and in artificial culture. External physical conditions have little effect upon this bacillus, and the dried and even buried flesh retains infection for a very long period of time.
 |
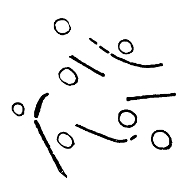 |
B. of Symptomatic Anthrax |
B. of Malignant Œdema |
The third disease-producing microbe found naturally in soil is that which produces the disease known as Malignant Œdema. Pasteur called this gangrenous septicœmia. Unlike quarter evil, malignant œdema may occur in man in cases where wounds have become septic. Animals become inoculated with this bacillus from the surface of soil, straw-dust, upper layers of garden-earth, or decomposing animal and vegetable matter.
The bacillus occurs in the blood and tissues as a long thread, composed of slender segments of irregular length. It is motile and anaërobic. The spores are larger than the diameter of the bacillus, and the organism produces gas; so much is this the case in artificial culture, that the medium itself is frequently split up.
Both malignant œdema and symptomatic anthrax are similar in some respects to anthrax itself. There are, however, a number of points for differential diagnosis. The enlargement of the spleen, the non-motility of the bacillus, the enormous numbers of bacilli throughout the body, the square ends, equal inter-bacillary spaces, aërobic growth, and characteristic staining afford ample evidence of anthrax.
The Relation of Soil generally to certain Bacterial Diseases. Recent investigations have, in effect, considerably added to our knowledge of pathogenic germs in soil; and whilst the three species enumerated above are still considered as types normally present in soil, it must not be forgotten that other virulent disease producers either live in the soil or are greatly influenced by its conditions.
Fränkel and Pasteur have both demonstrated the possible presence of anthrax. Fränkel maintained that it could not live there long, and at ten feet below the surface no growth occurred. This may have been due to the low temperature of such a depth. Pasteur held that earthworms are responsible for conveying the spores of anthrax from buried carcasses to the surface, and thus bringing about reinfection. Cholera, too, has been successfully grown in soil, except during winter. The presence of common saprophytes in the soil is prejudicial to the development of the cholera spirillum, and under ordinary circumstances it succumbs in the struggle for existence. From experiments recently conducted for the Local Government Board by Dr. Sidney Martin, evidence is forthcoming in support of the view that the bacillus of typhoid can live in certain soils. Samples of soil polluted with organic matter formed a favourable environment for living bacilli of typhoid for 456 days, whereas in sterilised soil, without organic matter, these organisms lived only twenty-three days. Tubercle also has been kept alive for several weeks in soil.
In passing, a single remark may be made in relation to174 the long periods during which bacteria can retain vitality in soil. Farm soils have, as is well known, been contaminated with anthrax in the late summer or autumn, and have retained the infectious virus till the following spring, and it has even then cropped up again in the hay of the next season. In 1881 Miquel took some samples of soil at a depth of ten inches, containing six and a half million bacteria per gram. After drying for two days at 30° C., the dust was placed in hermetically sealed tubes, which were put aside in a dark corner of the laboratory for sixteen years. Upon re-examination it is reported that more than three million germs per gram were still found, amongst them the specific bacillus of tetanus. Whether or not there is any fallacy in these actual figures, there is abundant evidence in support of the fact that bacteria, non-pathogenic and pathogenic, can and do retain their vitality, and sometimes even their virulence, for almost incredibly long periods of time.
It is now some years since Sir George Buchanan, for the English Local Government Board, and Dr. Bowditch, for the United States, formulated the view that there is an intimate relationship between dampness of soil and the bacterial disease of Consumption (tuberculosis of the lungs). The matter was left at that time sub judice, but the conclusion has been drawn, and surely a legitimate one, that the dampness of the soil acted injuriously in one of two ways. It either lowered the vitality of the tissues of the individual, and so increased his susceptibility to the disease, or in some way unknown favoured the life and virulence of the bacillus. That is one fact. Secondly, Pettenkofer traced a definite relationship between the rise and fall of the ground water with pollution of the soil and enteric (typhoid) fever.51 A 175third series of investigations concluded in the same direction, viz., the researches of Dr. Ballard respecting summer diarrhœa. This, it is generally held, is a bacterial disease, although no single specific germ has been isolated as its cause. Ballard demonstrated that the summer rise of diarrhœa mortality does not commence until the mean temperature of the soil, recorded by the four-foot thermometer, has attained 56.4° F., and the decline of such diarrhœa coincides more or less precisely with the fall in soil temperature. This temperature (56.4° F.) is, therefore, considered as the "critical" four-foot earth temperature, that is to say, the temperature at which certain changes (putrefactive, bacterial, etc.) take place in the pores of the earth, with the consequent development of the diarrhœal poison.
After a very elaborate and prolonged investigation on behalf of the Local Government Board, Dr. Ballard formulates the causes of diarrhœa in the following conclusions:52
(a) cause of diarrhœa resides ordinarily in the superficial layers of the earth, where it is intimately associated with the life processes of some micro-organism not yet detected or isolated.
(b) That the vital manifestations of such organism are dependent, among other things, perhaps principally upon conditions of season and the presence of dead organic matter, which is its pabulum.
(c) That on occasion such micro-organism is capable of getting abroad from its primary habitat, the earth, and having become air-borne, obtains opportunity for fastening on non-living organic material, and of using such organic 176matter both as nidus and as pabulum in undergoing various phases of its life history.
(d) That from food, as also from contained organic matter of particular soils, such micro-organism can manufacture, by the chemical changes wrought therein through certain of its life processes, a substance which is a virulent chemical poison.
Here, then, we have a large mass of evidence from the data collected by Buchanan, Bowditch, Pettenkofer, and Ballard. But much of this work was done anterior to the time of the application of bacteriology to soil constitution. Recently the matter has received increased attention from various workers abroad, and in England from Dr. Sidney Martin, Professor Hunter Stewart, Dr. Robertson, and others. The greater part of this work we cannot here consider. But some reference must be made to Dr. Robertson's admirable researches into the growth of the bacillus of typhoid in soil. By experimental inoculation of soil with broth cultures, he was able to isolate the bacillus twelve months after, alive and virulent. He concludes that the typhoid organism is capable of growing very rapidly in certain soils, and under certain circumstances can survive from one summer to another. The rains of spring and autumn or the frosts and snows of winter do not kill them off so long as there is sufficient organic pabulum. Sunlight, the bactericidal power of which is well known, had, as would be expected, no effect except upon the bacteria directly exposed to its rays. The bacillus typhosus quickly dies out in the soil of grass-covered areas. Dr. Robertson holds that the chief channel of infection between typhoid-infected soil and man is dust. As in tubercle and anthrax, so in typhoid, dried dust or excreta containing the bacillus is the vehicle of disease.
Hitherto we have addressed ourselves to those diseases the known causal organisms of which reside, normally or177 abnormally, in the soil. But closely allied to these matters connected with the rôle of pathogenic bacteria in soil is the question of what has been termed the miasmatic influence of soil. The term "miasm" has had an extensive and somewhat diffuse application in medical science. It may happen in the future that typhoid will be classified strictly as a miasmatic disease. But at present, in the transition state of the science, it would hardly be justifiable to classify typhoid with a typically miasmatic disease like malaria. Yet it is clear that mention should here be made of a group of diseases of which malaria is the type, and of which the tropics generally are the native land. The bacterial etiology of the group is by no means worked out. The cause of malaria alone is not yet a closed subject. However the details of the etiology of this group finally arrange themselves, there is little doubt of two facts, viz., the diseases are probably produced by bacteria or allied protozoa, and soil plays an important part in their production.
From what has been said, it will be seen that though a considerable amount of knowledge has been obtained respecting bacteria in the soil, it may be conjectured that actually there is still a great deal to ascertain before the micro-biology of soil is in any measure complete or even intelligent. The mere mention of tetanus and typhoid in the soil, and their habits, nutriment, and products therein, not to mention the work of the economic bacteria, is to open up to the scientific mind a vast realm of possibility. It is scarcely too much to say that a fuller knowledge of the part which soil plays in the culture and propagation of bacteria may suffice to revolutionise the practice of preventive medicine. Truly, our knowledge at the moment is rather a heterogeneous collection of isolated facts and theories, some of which, at all events, require ample confirmation; still, there is a basis for the future which promises much constructive work.
Injurious micro-organisms in foods are, fortunately for the consumers, usually killed by cooking. Vast numbers are, as far as we know, of no harm whatever. Alarming reports of the large numbers of bacteria which are contained in this or that food are generally as irrelevant as they are incorrect. Bacteria, as we have seen, are ubiquitous. In food we have abundance of the chief thing necessary to their life and multiplication—favourable nutriment. Hence we should expect to find in uncooked or stale food an ample supply of saprophytic bacteria. There was much wholesome truth in the assertions made some two years ago by the late Professor Kanthack, to the effect that good food as well as bad frequently contained large numbers of bacteria, and often of the same species. It is well that we should become familiarised with this idea, for its accuracy cannot be doubted, and its usefulness at the present time may not be without its beneficial effect.
Nevertheless, it is well we should know the bacterial flora of good and bad foods for at least two reasons. First, there is no doubt whatever that a considerable number of cases of poisoning can be traced every year to food containing harmful bacteria or their products. To several of the more notorious cases we shall have occasion to refer in passing. Secondly, we may approach the study of the bacteriology of foods with some hope that therein light will be found179 upon some important habits and effects of microbes. There can be little doubt that food-bacteria afford an example of association and antagonism of organisms to which reference has already been made. Any information that can be gleaned to illumine these abstruse questions would be very welcome at the present time. But there is a still further, and possibly an equally important, point to bear in mind, namely, the economic value of microbes in food. In a short account like the present it will be impossible to enter into hypotheses of pathology, but we shall at least be able to consider some of these interesting experiments which have been conducted in the sphere of beneficial bacteria.
The injurious effects of organisms contained in foods has been elucidated by the excellent work of the late Dr. Ballard. From the careful study of a number of epidemics due to food poisoning, this patient observer was able, without the aid of modern bacteriology, to arrive at a simple principle which must not be forgotten. Food poisoning is due either to bacteria themselves or to their products, which are contained in the substance of the food. In cases of the first kind, bacteria gaining entrance to the human alimentary canal, set up their specific changes and produce their toxins, and by so doing in course of time bring about a diseased condition, with its consequent symptoms. On the other hand, if the products, sometimes called ptomaines, are ingested as such, the symptoms set up by their action in the body tissues appear earlier. From these facts Dr. Ballard deduced the simple principle that if there is no incubation period or, at all events, a comparatively short space of time between eating the poisoned food and the advent of disease, the agents of the disease are products of bacteria. If, on the other hand, there is an incubation period, the agents are probably bacteria.
It is necessary to mention two other facts. Dr. Cautley53 180has recently been engaged in isolating from poisoned foods the different species of bacteria present. It would appear that these are limited, as a rule, to two or three kinds. As regards disease, the organisms of suppuration are the most common. Liquefying or fermentative bacteria are frequently present, the Proteus family being well represented. In addition there are, according to circumstances, a number of common saprophytes. Now, as we have pointed out, these organisms may act injuriously by some kind of cooperation, or they may by themselves be harmless, and pathological conditions be due to the occasional introduction of pathogenic species.
The other fact, requiring recognition from anyone who proposes to study the bacteriology of foods, is that a certain appreciable amount of the responsibility for food poisoning rests with the tissues of the individual ingesting the food. There is ample evidence in support of the fact that not all the persons partaking of infected food suffer equally, and occasionally some escape altogether. We know little or nothing of the causes of such modification in the effect produced. It may be due to other organisms, or chemical substances already in the alimentary canal of the individual, or it may be due to some insusceptibility or resistance of the tissues. Be that as it may, it is a matter which must not be neglected in estimating the effects of food contaminated with bacteria or their products.
Milk. There are few liquids in general use which contain such enormous numbers of germs as milk. To begin with, milk is in every physical way admirably adapted to be a favourable medium for bacteria. It is constituted of all the chief elements of the food upon which bacteria live. It is frequently at a temperature favourable to their growth. It is par excellence an absorptive fluid. A dish of ordinary water and a dish of newly drawn milk laid side by side, and under similar conditions of temperature, will rapidly demon181strate the difference in degree of absorptivity between the two fluids. Yet, whilst this general fact is true, we must emphasise at the outset the possibility and practicability of securing absolutely pure sterile milk. Recently some milking was carried out under strict antiseptic precautions, with the above sterile result. The udder was thoroughly cleansed, the hands of the milker washed with corrosive sublimate and then pure water, the vessels which were to receive the milk had been carefully sterilised, and the whole process was carried out in strict cleanliness. The result was that the sample of milk remained sweet and good and contained no germs. It should be stated that the first flow of milk, washing out the milk-ducts of the udder, was rejected. This fact of the sterility of cleanly drawn milk is not a new one, and has been established by many bacteriologists. Milk, then, is normally a sterile secretion. How does it gain its enormously rich flora of bacteria?
Sources of Pollution of Milk. These are various, and depend upon many minor circumstances and conditions. For all practical purposes there are three chief opportunities between the cow and the consumer when milk may become contaminated with bacteria:
1. At the time of milking.
2. During transit to the town, or dairy, or consumer.
3. After its arrival.
Pollution at the Time of Milking arises from the animal, the milker, or unclean methods of milking. It is now well known that in tuberculosis of the cow affecting the udder the milk itself shows the presence of the bacillus of tubercle. In a precisely similar manner all bacterial diseases of the cow which affect the milk-secreting apparatus must inevitably add their quota of bacteria to the milk. To this matter we shall have occasion to refer again. There is a further contamination from the animal when it is kept unclean, for it happens that the unclean coat of a cow will more materi182ally influence the number of micro-organisms in the milk than the popularly supposed fermenting food which the animal may eat. It is from this external source rather than from the diet that organisms occur in the milk. The hairy coat offers many facilities for harbouring dust and dirt. The mud and filth of every kind that may be habitually seen on the hinder quarters of cattle all contribute largely to polluted milk. Nor is this surprising. Such filth at or near the temperature of the blood is an almost perfect environment for many of the putrefactive bacteria.
The milker is also a source of risk. His hands, as well as the clothes he is wearing, can and do readily convey both innocent and pathogenic germs to the milk. Clothed in dust-laden garments, and frequently characterised by dirty hands, the milker may easily act as an excellent purveyor of germs. Not a few cases are also on record where it appears that milkers have conveyed germs of disease from some case of infectious disease, such as scarlet fever, in their homes. But under the more efficient registration of such disease which has recently characterised many dairy companies, the danger of infection from this source has been reduced to a minimum. The habit of moistening the hands with a few drops of milk previous to milking is one to be strongly deprecated.
Professor Russell recounts a simple experiment which clearly demonstrates these simple but effective sources of pollution:
"A cow that had been pastured in a meadow was taken for the experiment, and the milking done out of doors, to eliminate as much as possible the influence of germs in the barn air. Without any special precaution being taken the cow was partially milked, and during the operation a covered glass dish, containing a thin layer of sterile gelatine, was exposed for sixty seconds underneath the belly of the cow in close proximity to the milk-pail. The udder, flank, and legs of the cow were then thor183oughly cleaned with water, and all of the precautions referred to before were carried out, and the milking then resumed. A second plate was then exposed in the same place for an equal length of time, a control also being exposed at the same time at a distance of ten feet from the animal and six feet from the ground to ascertain the germ contents of the surrounding air. From this experiment the following instructive data were gathered. Where the animal was milked without any special precautions being taken there were 3250 bacterial germs per minute deposited on an area equal to the exposed top of a ten-inch milk-pail. Where the cow received the precautionary treatment as suggested above, there were only 115 germs per minute deposited on the same area. In the plate that was exposed to the surrounding air at some distance from the cow there were 65 bacteria. This indicates that a large number of organisms from the dry coat of the animal can be kept out of milk if such simple precautions as these are carried out."54
The influence of the barn air, and the cleanliness or otherwise of the barn, is obviously great in this matter. As we have seen, moist surfaces retain any bacteria lodged upon them; but in a dry barn, where molecular disturbance is the rule rather than the exception, it is not surprising that the air is heavily laden with microbic life. Here again many improvements have been made by sanitary cleanliness in various well-known dairies. Still there is much more to be done in this direction to ensure that the drawn milk is not polluted by a microbe-impregnated atmosphere.
The risks in transit differ according to many circumstances. Probably the commonest source of contamination is in the use of unclean utensils and milk-cans. Any unnecessary delay in transit affords increased opportunity for multiplication; particularly is this the case in the summer months, for at such times all the conditions are favourable to an enormous increase of any extraneous germs which may have 184gained admittance at the time of milking. Thus we have (1) the milk itself affording an excellent medium and supplying ideal pabulum for bacteria, (2) a more or less lengthened railway journey or period of transit giving ample time for multiplication, (3) the favourable temperature of summer heat. We shall refer again to the rate of multiplication of germs in milk.
Lastly, many are the advantages given to bacteria when milk has reached its commercial destination. In milk-shops and in the home there are not a few risks to be added on to the already imposing category. Water is occasionally, if not frequently, added to milk to increase its volume. Such water of itself will make its own contribution to the flora of the milk, unless indeed, which is unlikely, the water has been recently and thoroughly boiled before addition to the milk. Again, it is impossible to suppose that in small homes—perhaps of only one room—where the milk stands for several hours, pollution is avoidable. From a hundred different sources such milk runs the risk of being polluted.
Before proceeding, a word must be said respecting the first milk which flows from the udder in the process of milking, and which is known as the fore-milk. This portion of the milk is always rich in bacterial life on account of the fact that it has remained in the milk-ducts since the last milking. However thorough the manipulation, there will always be a residue remaining in the ducts, which will, and does, afford a suitable nidus and incubator for organisms. The latter obtain their entrance through the imperfectly closed teat of the udder, and pass readily into the milk-duct, sometimes even reaching the udder itself and setting up inflammation (mastitis). Professor Russell states that he has found 2800 germs in the fore-milk in a sample of which the average was only 330 per cc. Schultz found 83,000 micro-organisms per cc. in the fore-milk, and only 9000 in185 the mid-milk. As a matter of fact, most of this large number belong to the lactic-acid fermentation group, and the fore-milk rarely contains more than two or three species, and still more rarely any disease-producing bacteria. Still, they occur in such enormous numbers that their addition to the ordinary milk very materially alters its quality. Bolley and Hall, of North Dakota, report sixteen species of bacteria in the fore-milk, twelve of which produced an acid reaction. Dr. Veranus Moore, of the United States Department of Agriculture,55 concludes from a large mass of data that freshly drawn fore-milk contains a variable but generally enormous number of bacteria, but only several species, the last milk containing, as compared with the fore-milk, very few micro-organisms. The bacteria which become localised in the milk-ducts, and which are necessarily carried into the milk, are for the greater part rapidly acid-producing organisms, i. e., they ferment milk-sugar, forming acids. They do not produce gas. Still their presence renders it necessary to "pasteurise" as soon as possible. Dr. Moore holds that much of the intestinal trouble occurring in infants fed with ordinarily "pasteurised" milk arises from acids produced by these bacteria between the drawing of the milk and the pasteurisation.
The Number of Bacteria in Milk. From all that has been said respecting the sources of pollution and the favourable nidus which milk affords for bacteria, it is not surprising that a very large number of germs are almost always present in milk. The quantitative estimation of milk appears more alarming than the qualitative. It is true some diseases are conveyed by bacteria in milk, but on the whole most of the species are non-pathogenic. Nor need the numbers, though serious, too greatly alarm us, for, as we shall see at a later stage, disease is a complicated condition, and due to other agencies and conditions than merely the bacteria, which may 186be the vera causa. In addition to the fact that the high numbers have but a limited significance, we must also remember that there is no uniformity whatever in these numbers. The conditions which chiefly control them are (1) temperature, (2) time.
The Influence of Temperature. We have already noticed, when considering the general conditions affecting bacteria, how potent an agent in their growth is the surrounding temperature. Generally speaking, temperature at or about blood-heat favours bacterial growth. Freudenreich has drawn up the following table which graphically sets forth the effect of temperature upon bacteria in milk:
| 3 hours. | 6 hours. | 9 hours. | 24 hours. | |
| 59° F. | 1 + | 2.5 | 5 | 163 |
| 77° F. | 2 | 18.5 | 107 | 62,100 |
| 95° F. | 4 | 1,290 | 3,800 | 5,370 |
This instructive table claims some observations. It will be noticed that at 59° F. there is very little multiplication. That may be accepted as a rule. At 77° F. the multiplication, though not particularly rapid at the outset, results finally, at the end of the twenty-four hours, in the maximum quantity. These were probably common species of saprophytic bacteria, which increase readily at a comparatively low temperature. During the subsequent hours, after the twenty-four, we should expect a decline rather than an increase in 62,000, owing to the keen competition consequent upon the limitation of the pabulum. From a consideration of these figures we conclude that a warm temperature, somewhat below blood-heat, is most favourable to multiplication of bacteria in milk; that the common saprophytic organisms multiply the most rapidly; that, in the course of time, competition kills off a large number.
Let us take another example, from Professor Conn:
| 77° F. | 95° F. | ||||
| 2 | hours | after | milking | (liquefied the plate of gelatine) |
1,275,000 |
| 7 | " | " | " | 14,620,000 | 45,900,000 |
| 9 | " | " | " | 36,550,000 | 57,800,000 |
| 24 | " | " | " | 13,702,000,000 | 13,812,500,000 |
| [Bacteria per cub. inch.] | |||||
These almost incredibly large figures illustrate much the same points, particularly the rapid multiplication at blood-heat, and the later rise at 77° C.
The Influence of Time is not less marked than that of temperature, as the following table will show:
| Milk drawn at 59° C. | = | 153,000 | m.o. | per | cub. | in. | ||
| After | 1 | hour | = | 616,000 | " | " | " | " |
| " | 2 | hours | = | 539,000 | " | " | " | " |
| " | 4 | " | = | 680,000 | " | " | " | " |
| " | 7 | " | = | 1,020,000 | " | " | " | " |
| " | 9 | " | = | 2,040,000 | " | " | " | " |
| " | 24 | " | = | 85,000,000 | " | " | " | " |
| (Conn.) | ||||||||
Freudenreich gives another example, as follows:
| Milk drawn at 15.5° C. | = | 27,000 | m.o. | per | cc. | ||
| After | 4 | hours | = | 34,000 | " | " | " |
| " | 9 | " | = | 100,000 | " | " | " |
| " | 24 | " | = | 4,000,000 | " | " | " |
Concerning these figures little comment is necessary. But here again, we may remember that this rapid multiplication continues only up to a certain point, after which competition brings about a marked reduction.
The effect of temperature and time has been illustrated by Dr. Buchanan Young's recent researches, laid before the188 Royal Society of Edinburgh. He estimated that in the Edinburgh milk supply three hours after milking there were 24,000 micro-organisms per cc. in winter; 44,000 in spring; 173,000 in late summer and autumn. Again, he found that five hours after milking there were 41,000 micro-organisms per cc. in country milk, and more than 350,000 micro-organisms per cc. in town milk. Many London milks would exceed 500,000 per cc.56
There is no standard or uniformity in the numerical estimation of bacteria in milk. A host of observers have recorded widely varying returns due to the widely varying circumstances under which the milk has been collected, removed, stored, and examined. Nor is it possible to establish any standard which may be accepted as a normal or healthy number of bacteria, as is done in water examination. Bitter has suggested 50,000 micro-organisms per cc. as a maximum limit for milk intended for human consumption.
But owing to differences of nomenclature and classification, in addition to differences in mode of examination at present existing in various countries, it is impossible to state even approximately how many bacteria and how many species of bacteria have been isolated from milk. Until some common international standard is established mathematical computations are practically worthless. They are needlessly alarming and sensational. And it should be remembered that great reliance cannot be placed upon these numerical estimations. They vary from day to day, and even hour to hour. Furthermore, vast numbers of bacteria are economic in the best sense of the term, and the bacteria of milk are chiefly those of a fermentative kind, and not disease-producers.
Kinds of Bacteria in Milk. It is clear from the foregoing that the only valuable estimation of bacteria in milk is a 189qualitative one. The kinds commonly found may be classified thus:
1. Non-pathogenic; fermenting and various unclassified micro-organisms.
2. Pathogenic; tuberculosis, typhoid, cholera, scarlet fever, diphtheria, and suppurative diseases have all been spread by the agency of milk.
1. The Fermentation Bacteria
At the most we can make a merely provisional classification of these processes. Many of them are intimately related. Of others, again, our knowledge is at present very limited. It may be advisable, before proceeding, to consider shortly what are the constituents of milk upon which living ferments of various kinds exert their action. A tabulation of the chief constituents would be as follows:
| Ordinary fresh milk = 100 per cent. |
 |
(1) Water | 87.5 | per | cent. |
| (2) Milk-sugar | 4.9 | " | " | ||
| (3) Fat | 3.6 | " | " | ||
| (4) Proteids (casein, etc.) | 3.3 | " | " | ||
| (5) Mineral matter | 0.7 | " | " | ||
| —— 100.0 |
|||||
Another mode of expressing average milk constitution would be thus:
| Fat | 4.1 | per | cent. |
| Solids not fat | 8.8 | " | " |
| —— | |||
| 12.9 | " | " |
It is probably too obvious to need remark that milks vary in standard, but the above figures may be taken as authentic averages.
Milk-sugar, or Lactose (C12H24O12). This is an important and constant constituent of milk. It forms the chief substance in solution in whey or serum. Milk-sugar approximates to dextrose in its action on polarised light. By boiling with sulphuric acid it is converted into dextrose and galactose.
Fat occurs in milk as suspended globules, and by churning may be made into butter.
The Proteids include casein, albumen, lactoprotein, and a small quantity of globulin. These are the nitrogenous bodies.
Mineral Matter. The ash of milk, obtained by careful ignition of the solids, contains calcium, magnesium, potassium, sodium, phosphoric acid, sulphuric acid, chlorine, and iron, phosphoric acid and lime being present in the largest amounts.
(1) Lactic Acid Fermentation. If milk is left undisturbed, it is well known that eventually it becomes sour. The casein is coagulated, and falls to the bottom of the vessel; the whey or serum rises to the top. In fact, a coagulation analogous to the clotting of blood has taken place. In addition to this, the whole has acquired an acid taste. Now, this double change is not due to any one of the constituents we have named above. It is, in short, a fermentation set up by a living ferment introduced from without. The constituent most affected by the ferment is the milk-sugar, which is broken down into lactic acid, carbonic acid gas, and other products.
For many years it has been known that sour milk contained bacteria. Pasteur first described the Bacillus acidi lactici, which Lister isolated and obtained in pure culture. Hueppe contributed still further to what was known of this bacillus, and pointed out that there were a large number of varieties, rather than one species, to be included under the term B. acidi lactici. We have already seen that these191 bacilli do not as a rule liquefy gelatine, form spores, are non-motile, and are easily killed by heat.
When a certain quantity of lactic acid has been formed the fermentation ceases. It will recommence if the liquid be neutralised with carbonate of lime, or pepsine added. Since Pasteur's discovery of a causal bacillus for this fermentation, other investigators have added a number of bacteria to the lactic acid family. Some of these in pure culture have been used in dairy industry to add to the butter a pure sour taste, a more or less aromatic odour, and a higher degree of preservation.
(2) Butyric Acid Fermentation. This form of fermentation is also one which we have previously considered.
Both in lactic and butyric fermentation we must recognise that in the decomposition of milk-sugar there are almost always a number of minor products occurring. Some of the chief of these are gases. Hydrogen, carbonic acid, nitrogen, and methane occur, and cause a characteristic effect which is frequently deleterious to the flavour of the milk and its products. Most of the gas-producing ferments are members of the lactic acid group, and are sometimes classified in a group by themselves. In cheese-making the gases create the pin-holes and air-spaces occasionally seen.
(3) Curdling Fermentations without Acid Production. Of these there are several, caused by different bacteria. What happens is that the milk coagulates, as we have described, but no acid is produced, the whey being sweet to the taste rather than otherwise. Digestion of casein may or may not take place.
We must now mention several fermentations about which little is known. They are designated by terms denoting the outward condition of the milk, without giving any information respecting the real physiological alteration which has occurred.
(4) Bitter Fermentation. Some bitter conditions of milk192 are due to irregularity of diet in the cow. Similar changes occur in conjunction with some of the acid fermentations. Weigmann and Conn have, however, shown that there is a specific bitterness in milk due to bacteria which appear to produce no other change. Hueppe suggests that it may be due in part to a proteid decomposition resulting in bitter peptones. There seems to be some evidence for supposing that the bitter bacteria produce very resistant spores, which enable them to withstand treatment under which the lactic acid succumbs.
(5) Slimy Fermentation. This graphic but inelegant word is used to denote an increased viscosity in milk, and its tendency when being poured to become ropy and fall in strings. Such a condition deprives the milk of its use in the making of certain cheeses, whilst in other cases it favours the process. In Holland, for example, in the manufacture of Edam cheese, this "slimy" fermentation is desired. Tættemœlk, a popular beverage in Norway, is made from milk that has been infected with the leaves of the common butter wort, Pinguicula vulgaris, from which Weigmann separated a bacillus possessing the power of setting up slimy fermentation. There are, perhaps, as many as a dozen species of bacteria which have in a greater or less degree the power of setting up this kind of fermentation. In 1882 Schmidt isolated the Micrococcus viscosus, which occurs in chains and rosaries, affecting the milk-sugar. It grows at blood-heat, and is not easily destroyed by cold. Its effect on various sugars is the same. The M. Freudenreichii, the specific micro-organism of "ropiness" in milk, is a large, non-motile, liquefying coccus, which can produce its result in milk within five hours. On account of its resistance to drying, it is difficult to eradicate when once it makes its appearance in a dairy. The organism used in making Edam cheese is the Streptococcus Hollandicus, and in hot milk it can produce ropiness in one day. A number of bacilli have193 been detected by several observers and classified as slime fermentation bacteria. The Bacillus lactis pituitosi, a slightly curved, non-liquefying rod, which is said to produce a characteristic odour, in addition to causing ropiness, brings about some acidity. B. lactis viscosus is slow in starting its fermentation, but maintains its action for as long as a month. Many of the above organisms, with others, produce "slimy" fermentation in alcoholic beverages as well as in milk.
(6) Soapy Milk. This is still another form of fermentation, the etiology of which has been elucidated by Weigmann. The Bacillus saponacei imparts to milk a peculiar soapy flavour. It was detected in the straw of the bedding and hay of the fodder, and from such sources may infect the milk. There is little or no coagulation.
(7) Chromogenic Changes. We have already remarked that colour is the natural and apparently only product of many of the innocent bacteria. They put out their strength, so to speak, in the production of bright colours. The chief colours produced by germs in milk are as follows:
Red Milk. Bacillus prodigiosus, in the presence of oxygen, causes a redness, particularly on the surface of milk. It was the work of this bacillus that caused "the bleeding host," which was one of the superstitions of the Middle Ages. B. lactis erythrogenes produces a red colour only in the dark, and in milk that is not strongly acid in reaction. When grown in the light this organism produces a yellow colour. There is a red sarcina (Sarcina rosea) which also has the faculty of producing red pigment. One of the yeasts is another example.
It must not be forgotten that redness in milk may actually be due to the presence of blood from the udder of the cow. In such a case the blood and milk will be inextricably mixed together, and not in patches or a pellicle.
Blue Milk is due to the growth of Bacillus cyanogenus.194 This is an actively motile rod, the presence of which does not materially affect the milk, but causes the milk products to be of poor quality.
Yellow Milk. Bacillus synxanthus is held responsible for curdling the milk, and then at a later stage, in redissolving the curd, produces a yellow pigment.
Violet and Green Pigments in milk are also the work of various bacteria.
2. Various Unclassified Bacteria
In milk this is a comparatively small group, for it happens that those bacteria in milk which cannot be classified as fermentative or pathogenic are few. The almost ubiquitous Bacillus coli communis occurs here as elsewhere, and might be grouped with the gaseous fermentative organisms on account of its extraordinary power of producing gas and breaking up the medium (whether agar or cheese) in which it is growing. What its exact rôle is in milk it would be difficult to say. It may act, as it frequently does elsewhere, by association in various fermentations. Some authorities hold that its presence in excessive numbers may cause epidemic diarrhœa in infants.
Several years ago a commission was appointed by the British Medical Journal to inquire into the quality of the milk sold in some of the poorer districts of London. Every sample was found to contain Bacillus coli, and it was declared that this particular microbe constituted 90 per cent. of all the organisms found in the milk.57 We record this statement, but accept it with some misgiving. The diagnosis of B. coli four or five years ago was not such a strict matter as to-day. Still, undoubtedly, this particular organism is not uncommonly found in milk, and its source is unclean dairying. In the same investigation Proteus
vulgaris, B. fluorescens, and many liquefying bacteria were frequently found. Their presence in milk means contamination with putrefying matter, surface water, or a foul atmosphere.
A number of water bacteria find their way into milk in the practice of adulteration, and foul byres afford ample opportunity for aërial pollution.
Another unclassified group occasionally present in milk is represented by moulds, particularly Oidium lactis, the mould which causes a white fur, possessing a sour odour. It is allied to the Mycoderma albicans (O. albicans), which also occurs in milk, and causes the whitish-grey patches on the mucous membrane of the mouths of infants (thrush). These and many more are occasionally present in milk.
3. The Disease-Producing Power of Milk
The general use of milk as an article of diet, especially by the younger and least resistant portion of mankind, very much increases the importance of the question as to how far it acts as a vehicle of disease. Recently considerable attention has been drawn to the matter, though it is now a number of years since milk was proved to be a channel for the conveyance of infectious diseases. During the last twenty years particular and conclusive evidence has been deduced to show that milch cows may themselves afford a large measure of infection. The recent extensive work in tuberculosis by the Royal Commission has done much to obtain new light on the conveyance of that disease by milk and meat. The enormous strides in the knowledge of diphtheria and other germ diseases have also placed us in a better position respecting their conveyance by milk. Generally speaking, for reasons already given, milk affords an ideal medium for bacteria, and its adaptibility therefore for conveying disease is undoubted. We may now suitably196 turn to speak shortly of the outstanding facts of the chief diseases carried by milk.
Tuberculosis. It is well known that this disease is not a rare one amongst cattle. The problem of infective milk is, however, simplified at the outset by recognising the now well-established fact that the milk of tuberculous cows is only certainly able to produce tuberculosis in the consumers when the tuberculous disease affects the udder. This is not necessarily a condition of advanced tuberculosis. The udder may become affected at a comparatively early stage. But to make the milk infective the udder must be tubercular, and milk from such an udder possesses a most extraordinary degree of virulence. When the udder itself is thus the seat of disease, not only the derived milk, but the skimmed milk, butter-milk, and even butter, all contain tuberculous material actively injurious if consumed. Furthermore, tubercular disease of the udder spreads in extent and degree with extreme rapidity. From these facts it will be obvious that it is of first-rate importance to be able to diagnose udder disease. This is not always possible in the early stage. The signs upon which most reliance may be placed are the enlargement of the lymph-glands lying above the posterior region of the udder; the serous, yellowish milk which later on discharges small coagula; the partial or total lack of milk from one quarter of the udder (following upon excessive secretion); the hard, diffuse nodular swelling and induration of a part or the whole wall of the udder; and the detection in the milk of tubercle bacilli. The whole organ may increase in weight as well as size, and on post-mortem examination show an increase of connective tissue, a number of large nodules of tubercle, and a scattering of small granular bodies, known as "miliary" tubercles. Tuberculin may be used as an additional test. The udder is affected in about two per cent. of tuberculous cows.
There are a variety of causes in addition to the vera causa,197 the presence of the bacillus of tubercle, which make the disease common amongst cattle. Constitution, temperament, age, work, food, and prolonged lactation are the individual features which act as predisposing conditions; they may act by favouring the propagation of the bacillus or by weakening the resistance of the tissues. To this category must further be added conditions of environment. Bad stabling, dark, ill-ventilated stalls, high temperature, prolonged and close contact with other cows, all tend in the same direction.
Though there can be no doubt as to the virulence of tuberculous milk, it may be remembered with satisfaction that only about two per cent. of tuberculous cows have unmistakably tubercular milk. Even of this tubercular milk, unless it is very rich in bacilli and is ingested in large quantities, the risks are practically small or even absent. Practically the danger from drinking raw milk exists only for persons who use it as their sole or principal food, that is to say, young children and certain invalids. With adults in normal health the danger is greatly minimised, as the healthy digestive tract is relatively insusceptible. Moreover, dairy milk is almost invariably mixed milk; that is to say, if there is a tubercular cow in a herd yielding tubercle bacilli in her milk, the addition of the milk of the rest of the herd so effectually dilutes the whole as to render it almost innocuous.
But if for practical purposes we look upon all milk derived from tubercular udders as highly infective, we may adopt a comparatively simple and efficient remedy. To avoid all danger it is sufficient to bring the milk to a boil for a few minutes before it is consumed; in fact, the temperature of 85° C. (160° F.) prolonged for five minutes kills all bacilli. The common idea that boiled milk is indigestible, and that the boiling causes it to lose much of its nutritive value, is largely groundless.
Milk may become tubercular through the carelessness or198 dirty habits of the milker. Such a common practice as moistening the hands with saliva previously to milking may, in cases of tubercular milkers, effectually contaminate the milk. Again, it may become polluted by dried tubercular excreta getting into it. Such conveyances must be of rare occurrence, yet their possibility should not be forgotten.
An infant suckled by a tuberculous mother would run similarly serious risks of becoming infected with the disease.
In Liverpool, Dr. E. W. Hope, the Medical Officer of
Health, has organised an admirable system of examination
by skilled bacteriologists to find to what degree the Liverpool
milk supply is contaminated with tubercle. The final
result of this pioneer work, which ought really to be undertaken
by every great corporation responsible to the citizens
for a pure water and pure milk supply, is to the effect that
in Liverpool 5.2 per cent. of the samples of milk taken
from the city shippons contains tubercle bacilli. As regards
the milk sent in from the country, the return is that 13.4
per cent. is contaminated with the bacillus of tubercle.
| Town Shippons. | Country Shippons. | Total. | ||||
| Total. 228 |
Infected. 12 |
Per cent. 5.2 |
Total. 67 |
Infected. 9 |
Per cent. 13.4 |
295 |
Such results are very significant, and indicate the importance
of all large corporations obtaining the service of systematic
and periodic bacteriological examination of the milk
supply. Nor are the results surprising, for when we remember
the habits of the tubercle bacillus we cannot conceive
a more favourable nurture ground than the typical byre.
"Nothing worse than the insanitary conditions of the life
of the average dairy cow," says Sir George Brown, late of
the Board of Agriculture, "can be imagined." It will be
obvious that the above facts make it incumbent upon re199sponsible
authorities to see that not a stone is left unturned
to enforce cleanliness in all dairy work, isolation of diseased
cows, and strict treatment of all infected milk.
Typhoid Fever. Jaccoud in France and Hart in England have shown that enteric fever (typhoid) is not infrequently spread by milk. An epidemic affecting 386 persons in Stamford, Conn., U.S.A., was traced to milk, 97 per cent. of the cases coming from one single milk supply. Dr. McNail recently recorded an outbreak of twenty-two cases of enteric, due to a polluted milk supply.
Within the last twelve months much attention has been drawn to a milk source of typhoid infection by the epidemic of typhoid at Bristol. Dr. D. S. Davies has pointed out that a brook received the sewage of thirty-seven houses, the overflow of a cesspool serving twenty-two more, the washings from fields over which the drainage of several others was distributed, and the direct sewage from at least one other, and then flowed directly through a certain farm. The water of this stream supplied the farm pump, and the water itself, it is scarcely necessary to add, was highly charged with putrescent organic matter and micro-organisms. This water was used for washing the milk-cans from this particular farm, otherwise the dairy arrangements were efficient. Part of the milk was distributed to fifty-seven houses in Clifton; in forty-one of them cases of typhoid occurred. Another part of the milk was sold over the counter; twenty households so obtaining it were attacked with typhoid fever, and a number of further infections and complications arose. This evidence would appear to support the fact that milk may act in the same way, though not in such a high degree, as water in the conveyance of typhoid fever.
It may be pointed out that specific typhoid is not a disease of animals; consequently no danger need be apprehended from milk if it is properly cared for after it comes from the cow. Typhoid milk is almost invariably due to200 the addition of typhoid-infected water, either by way of adulteration or in the process of washing out the milk-cans. Cases have, however, been recorded in which there has been direct transmission to the milk from a person convalescing from the disease, and also indirect transmission by a milker serving also in the capacity of nurse to a patient in his own family.
Though the typhoid bacillus appears not to have the power of multiplying in milk, it has the faculty of existing and thriving in milk, even when it has curdled or soured, for a considerable time, and may thus infect milk products like butter and cheese. But infection by milk products may be eliminated as of too rare occurrence to deserve attention. The bacillus does not coagulate the milk like its ally the Bacillus coli communis, which is a much more frequent and less injurious inhabitant of milk.
Cholera. The cholera bacillus, as we have already pointed out, is unable to live in an acid medium. Hence its life in milk is a limited one, and generally depends on some alkaline change in the milk. Heim found that cholera bacilli would live in raw milk from one to four days, depending upon the temperature. D. D. Cunningham, from the results of a large number of investigations in India, concludes that the rapidly developing acid fermentations normally or usually setting in, connected with the rapid multiplication of other common bacteria and moulds, tend to arrest the multiplication of cholera bacilli, and eventually to destroy their vitality. Boiling milk appears, on the contrary, to increase the suitability of milk as a nidus for cholera bacilli, partly by its germicidal effect upon the acid-producing microbes, and partly because it removes from the milk the enormous numbers of common bacteria, which in raw milk cause such keen competition that the cholera bacillus finds existence impossible.
Professor W. J. Simpson, lately the Medical Officer of201 Health for Calcutta, has placed on record an interesting series of cholera cases on board the Ardenclutha, in the port of Calcutta, which arose from drinking milk which had been polluted with one quarter of its volume of cholera-infected water. This water came from a tank into which some cholera dejecta had passed. Of the ten men who drank the milk four died, five were severely ill, and one, who drank but very little of the milk, was only slightly ill. There was no illness whatever amongst those who did not drink the milk.
Diphtheria. Recent observations on the infectivity of diphtheria in milk by Schottelius have established the fact that milk is a good medium for the bacillus of diphtheria, but that it rarely acts as a vehicle for transmitting the disease. Klein has emphasised the possibility of this means of infection. In the first place, it is obvious that the milk may become infected from a human source—from pollution with diphtheritic discharges or dried "fomites." Secondly, from a variety of different quarters evidence has been forthcoming to throw some suspicion upon the cow itself as the agent. Klein states that "a new eruptive disease on the teats and udder of the cow," consisting of papules, vesicles, and induration, may be set up by the subcutaneous inoculation of a pure culture of the Bacillus diphtheriæ. In these eruptions a bacillus similar to the B. diphtheriæ was demonstrated. On a priori grounds this evidence substantiates a belief that diphtheria, in some form or other, may be a disease of cows. Other observers have not been able to confirm these observations, and the whole matter of cow diphtheria must remain for the present sub judice.
As long ago as 1879 W. H. Power traced an epidemic of diphtheria in North London to the milk supply. In 1887 the same authority studied another outbreak, and other observers have produced further evidence in favour of the conveyance of this disease by milk. Air infection of milk by the Bacillus diphtheriæ probably occurs only very rarely,202 on account of the fact that the organism is readily killed by desiccation, and yet such is necessary before it can be airborne. The most frequent mode of infection of milk with this disease is from the throats, hands, bodies, or clothing of dairy workers suffering from a mild or acute form of the disease.
The specific and proved cases in which milk has acted as the vehicle of diphtheria are, it is true, comparatively few. Yet, nevertheless, the possibility of milk infection in this disease is not one which we can afford to neglect.
Scarlet Fever. Here again the evidence is not complete, chiefly owing to the fact that no specific organism of scarlet fever has yet been discovered. Many cases have, however, illustrated the undeniable conveyance of the disease by milk. Even before 1881 a number of milk epidemics of scarlet fever had been traced out. In 1882 these were further added to by Mr. W. H. Power's report concerning a series of cases in Central London. That report was remarkable for the introduction of a new feature, viz., the evidence produced in favour of the infection of milk from some disease of the cow. The Medical Department of the Local Government Board from that time took up a position of suspended judgment concerning the belief hitherto credited that milk could only be infected by human scarlet fever. In 1886 there was a remarkable epidemic in Marylebone, and the theory was suggested by Dr. Klein and Mr. Power that the cow from which the milk was derived suffered from scarlet fever.
Into the extensive controversy which raged round "the Hendon disease," as it was called, affecting the cows supplying the Marylebone milk, we cannot here enter. It will be sufficient to say that a long discussion took place as to whether or not this Hendon disease was or was not scarlet fever. The difficulty of course largely arose from the fact before mentioned that we do not at present know the specific203 micro-organism of scarlet fever. The Agricultural Department supported the view of Professor Crookshank that the cow disease at Hendon was cowpox, and Professor Axe further pointed out that there was evidence of the Hendon milk having been contaminated with human scarlet fever. Whichever conclusion was adopted, all were agreed upon one point, viz., that the disease had been conveyed from Hendon to persons in Marylebone by means of the milk.
Mr. Ernest Hart in 1897 published a very large number of records of scarlatinal milk infection from all parts of the country, and though the cause of the disease is obscure, there is now no doubt that it may be and is conveyed by means of milk.
Other Diseases Conveyed by Milk. In addition to the above, there are other diseases spread by means of polluted milk. From time to time exceptional cases have occurred in which a disease like anthrax has been spread by this means. But it is not to such rare cases that we refer. There are two very common diseases in which milk has been proved to play a not inconsiderable part, viz., thrush and diarrhœa.
The mould which gives rise to the curd-like patches in the throats of children, and which is known as Oidium albicans, frequently occurs in milk. Soft white specks are seen on the tongue and mucous membrane of the cheeks and lips, looking not unlike particles of milk curd. If a scraping be placed upon a glass slide with a drop of glycerine and examined by means of the microscope, the spores and mycelial threads of this mould will be seen. The spores are oval, and possess a definite capsule. The threads are branched and jointed at somewhat long intervals. Milk affords an excellent medium for the growth of this parasite. Thus undoubtedly we must hold milk partly responsible for spreading this complaint. Penicillium, Aspergillus, and Mucor are also frequent moulds in milk.
Professor MacFadyen58 has given a full account of the ways in which milk becomes pathogenic, and his views have received further support from Professor Sheridan Delépine, who has examined more than one hundred samples of milk from Liverpool and Manchester. The result of this investigation has been that milk must be held to be one of the most potent causes of the summer diarrhœa of children. Indeed, a bacillus has been isolated identical with one which was apparently the cause of this complaint, which carries off such a large number of infants every summer. It resembles closely the Bacillus coli communis, which is an almost constant inhabitant of the alimentary canal, and is held by many bacteriologists to play, especially in conjunction with yeasts and other saprophytic organisms, an active rôle in the intestine of man.
In a recent official report59 Dr. Hope, of Liverpool, states that "the method of feeding plays a most important part in the causation of diarrhœa; when artificial feeding becomes necessary, the most scrupulous attention should be paid to feeding-bottles." Careless feeding, in conjunction with a warm, dry summer, invariably results in a high death-rate from this cause. These two causes interact upon each other. A warm temperature is a favourable temperature for the growth of the poisonous micro-organism; a dry season affords ample opportunity for its conveyance through the air. Unclean feeding-bottles are obviously an admirable nidus for these injurious bacteria, for in such a resting-place the three main conditions necessary for bacterial life are well fulfilled, viz., heat, moisture, and pabulum. The heat is supplied by the warm temperature, the moisture and food by the dregs of milk left in the bottle; and the dry air assists in transit.
Before passing on to other matters, reference must be made to poisonous products other than bacteria which occur in milk and set up ill-health. Vaughan, of Michigan, pointed out at the London Congress of Hygiene in 1891 that he had separated a poisonous alkaloid, which he called tyrotoxicon. This, as its name denotes, was a toxic or poisonous substance, probably produced by some form of microbe. It may be taken as a type of the organic chemical substances frequently occurring in milk.
From the somewhat extensive category of diseases which may be milk-borne, it will be suitable now to speak of some of the means at our disposal for obtaining and preserving good, pure milk.
We considered at the commencement of this chapter the most frequent channels of contamination. If these be avoided or prevented, and if the milk be derived from cows in good health and well kept, the risk of infection is reduced to a minimum. But we have seen that much, if not most, of the pollution of milk arises after the milking process and during transit and storage preparatory to use. Bacteria are so ubiquitous that to prevent the entrance of any at all is almost beyond hope. Can anything be done to prevent their multiplication or to kill them in the milk? Fortunately the answer is in the affirmative.
There are two means at hand to secure these results. First, we may add to the milk various chemical or physical preservatives. Borax or boric acid, formaldehyde, salicylic acid, and other chemical bodies are used for this purpose. The commonest of these is that named first. The Food and Drugs Act (Section VI., 1875) permits the addition of an ingredient not injurious to health if the same is required for protection or preparation of the article in question. It206 is, however, a difficult matter to determine what amount of boric acid is injurious to health, for this differs widely in different persons. It has been laid down by one authority that even so small an amount as one-tenth per cent. might have inconvenient results, owing to its cumulative effect. Formaldehyde is without doubt an excellent antiseptic, and the more its efficacy becomes known so much the more probably will it be used. The salicylates, which are mild antiseptics, have long been used as preservatives. These substances, then, can be added to milk in quantities not recognisable to the taste (salicylic acid about .75 grain, and boracic acid .4 grain, to the litre of milk). They will materially increase the time that milk will remain sweet, they will prevent a number of micro-organisms living in the milk, and will inhibit multiplication of others.60 Secondly, it is possible very perceptibly to remove the infectivity of milk by filtration and temperature variations.
Filtration has been practised for some time by the Copenhagen Dairy Company and by Bolle, of Berlin. The filters used consist of large cylindrical vessels divided by horizontal perforated diaphragms into five superposed compartments, of which the middle three are filled with fine sand of three sizes. At the bottom is the coarsest sand, and at the top the finest. The milk enters the lowest compartment by a pipe under gravitation pressure, and is forced upwards, and finally is run off into an iced cooler, and from that into the distribution cans. By this means the number of bacteria is reduced to one-third. The difficulty of drying and sterilising enough sand to admit a large turnover of milk is a serious one. This, in conjunction with the belief that 207filtration removes some of the essential nutritive elements of milk, has caused the process to be but little adopted. Dr. Seibert states that if milk be filtered through half an inch of compressed absorbent cotton, seven-eighths of the contained bacteria will be removed, and a second filtration will further reduce the number to one-twentieth. One quart of milk may thus be filtered in fifteen minutes.
The common methods now in vogue for the protection of milk are based upon germicidal temperatures. Low temperatures, it is true, do not easily destroy life, but they have a most beneficial effect upon the keeping quality of milk. At the outset of the process of cooling, strong currents of air are started in the milk-can, which act mechanically as deodorisers. But if the temperature be lowered sufficiently, the contained bacteria become inactive and torpid, and eventually are unable to multiply or produce their characteristic fermentations. At about 50° F. (10° C.) the activity ceases, and at temperatures of 45° F. (7° C.) and 39° F. (4° C.) organisms are deprived of their injurious powers. If it happens that the milk is to be conveyed long distances, then even a lower temperature is desirable. The most important point with regard to the cooling of milk is that it should take place quickly. Various kinds of apparatus are effective in accomplishing this. Perhaps those best known are Lawrence's cooler and Pfeiffer's cooler, the advantage of the latter being that during the process the milk is not exposed to the air. It must not be forgotten that cooling processes are not sterilising processes. They do not necessarily kill bacteria; they only inhibit activity, and under favourable circumstances the torpid bacteria may again acquire their injurious faculties. Hence during the cooling of milk greater care must be taken to prevent aërial contamination than is necessary during the process of sterilising milk. No cooling whatever should be attempted in the stable; but, on the other hand, there should be no delay. Climate208 makes little or no difference to the practical desirability of cooling milk, yet it is obvious that less cooling will be required in the cold season.
We now come to the protective processes known as sterilisation and pasteurisation. As we have already seen, sterilisation indicates a complete and final destruction of bacteria and their spores. As applied to methods of preserving milk, sterilisation means the use of heat at, or above, boiling-point, or boiling under pressure. This may be applied in one application of one to two hours at 250° F., or it may be applied at stated intervals at a lower temperature. The milk is sterilised—that is to say, contains no living germs—is altered in chemical composition, and is also boiled or "cooked," and hence possesses a flavour which to many people is unpalatable.
Now, such a radical alteration is not necessary in order to secure non-infectious milk. The bacteria causing the diseases conveyable by milk succumb at much lower temperatures than the boiling-point. Advantage is taken of this in the process known as "pasteurisation." By this method the milk is heated to 167–185° F. (75–85° C.). Such a temperature kills harmful microbes, because 75° C. is decidedly above their average thermal death-point, and yet the physical changes in the milk are practically nil, because 85° C. does not relatively approach the boiling-point. There is no fixed standard for pasteurisation, except that it must be above the thermal death-point of pathogenic bacteria, and yet below the boiling-point. As a matter of fact, 158° F. (70° C.) will kill all souring bacteria as well as disease-producing organisms found in milk. If the milk is kept at that temperature for ten or fifteen minutes, we say it has been "pasteurised." If it has been boiled, with or without pressure, for half an hour, we say it has been "sterilised." The only practical difference in the result is that sterilised milks have a better keeping quality than209 pasteurised, for the simple reason that in the latter some living germs have been unaffected.
Sterilisation may of course be carried out in a variety of modifications of the two chief ways above named. When the process is to be completed in one event an autoclave is used, in order to obtain increased pressure and a higher temperature. Milk so treated is physically changed in greater degree than in the slower process. The slow or intermittent method is, of course, based on Tyndall's discovery that actively growing bacteria are more easily killed than their spores. The first sterilisation kills the bacteria, but leaves their spores. By the time of the second application the spores have developed into bacteria, which in turn are killed before they can sporulate.
The methods of pasteurisation are continually being modified and improved, especially in Germany and America. Most of the variations in apparatus may be classed under two headings. There are, first, those in which a sheet of milk is allowed to flow over a surface heated by steam or hot water. This may be a flat, corrugated surface or a revolving cylinder. The milk is then passed into coolers. Secondly, milk is pasteurised by being placed in reservoirs surrounded by an external shell containing hot water or steam. Dr. A. L. Russell61 has described one apparatus consisting of a pasteuriser, a water-cooler, and an ice-cooler. The pasteuriser is heated by hot water in the outside casement. To equalise rapidly the temperature of the water and milk a series of agitators must be used. These are suspended on movable rods, and hang vertically in the milk and water chambers. By this ingenious arrangement the heat is diffused rapidly throughout the whole mass, and as the temperature of the milk reaches the proper point the steam is shut off, and the heat of the whole body of water and milk will remain constant for the proper length of time.
The somewhat difficult problem of drawing off the pasteurised milk from the vat without reinfecting it by contact with the air is solved by placing a valve inside the chamber, and by means of a pipe leading the pasteurised milk directly and rapidly into the coolers. These are of two kinds, which may be used separately or conjointly. In one set of cylinders there is cold circulating water, in the other finely crushed ice.
Domestic pasteurisation can be accomplished readily by heating the milk in vessels in a water-bath raised to the required temperature for half an hour.
Without entering into a long discussion upon the various methods adopted, we may summarise some of the chief essential conditions. It need scarcely be said that the operation must be efficiently conducted, and in such a way as to maintain absolute control over the time and temperature. The apparatus should be simple enough to be easily cleansed, sterilised, and economical in use. Arrangements must always be made to protect the milk from reinfection during and after the process. The entire preparation of the milk for market may be summed up in four items:
1. Pasteurisation in heat reservoir.
2. Rapid cooling in water-or ice-coolers.
3. All cans, pails, bottles, and other utensils to be thoroughly sterilised in steam.
4. The prepared milk must be placed in sterilised bottles and sealed up.
The quality of the milk to be pasteurised is an important point. All milks are not equally suited for this purpose, and those containing a large quantity of contamination, especially of spores, are distinctly unsuitable. Such milks, to be purified, must be sterilised. Dr. Russell has laid down a standard test for the degree of contamination which may be corrected by pasteurisation by estimating the degree of acidity, a low acidity (e. g., 0.2 per cent.) usually in211dicating a smaller number of spore-bearing germs than that which contains a high percentage of acid.
Lastly, while the heating process is of course the essential feature of efficient pasteurisation, it must not be forgotten that rapid and thorough cooling is almost equally important. As we have seen, pasteurisation differs from complete sterilisation in that it leaves behind a certain number of microbes or their spores. Cooling inhibits the germination and growth of this organismal residue. If after the heating process the milk is cooled and kept in a refrigerator, it will probably keep sweet from three to six days, and may do so for three weeks.
Before leaving this subject we may glance for a moment at the bacterial results of pasteurisation and sterilisation. The chief two of these are the enhanced keeping quality and the removal of disease-producing germs. The former is due in part to the latter, and also to the removal of the lactic acid and other fermentative bacteria. As a general rule these bacteria do not produce spores, and hence they are easily annihilated by pasteurisation. True, a number of indifferent bacteria are untouched, and also some of the peptonising species. The cooling itself contributes to the increased keeping power of the milk, especially in transit to the consumer.
Pasteurised milks have the following three economical and commercial advantages over sterilised milks, namely, they are more digestible, the flavour is not altered, and the fat and lact-albumen are unchanged. Professor Hunter Stewart, of Edinburgh, about two years ago, compiled from a number of experiments the following instructive and comprehensive table (page 212).
It will be admitted that this table exhibits much in favour
of pasteurisation; yet the crucial test must ever be the effect
upon pathogenic bacteria. Flügge has conducted a series
of experiments upon the destruction of bacteria in milk, and212
he states that a temperature of 158° F. (70° C.) maintained
for thirty minutes will kill the specific organisms of tubercle,
diphtheria, typhoid, and cholera. MacFadyen and Hewlett
have demonstrated,62 by sudden alternate heating and cooling,
that 70° C. maintained for half a minute is generally
sufficient to kill suppurative organisms and such virulent
types of pathogenic bacteria as Bacillus diphtheriæ, B.
typhosus, and B. tuberculosis.
| No. of Experiments. |
Average No. of Microbes spe cc. in Milk before Treatment. | Temperature and Duration of Pasteurisation in Minutes. |
No. of Microbes per cc. in Pasteurised Milk after 24 Hours. | Soluble Albumen in Fresh Milk, per cent. | Soluble Albumen in Pasteurised Milk, per cent. | Taste of Pasteurised Milk |
| 5 | 136,262 | 10' 60° C. | 1722 average | 0.423 | 0.418 | Unaffected |
| 4 | 53,656 | 30' 60° C. | 1 sterile 3 averaged 955 |
0.435 | 0.427 | " |
| 12 | 78,562 | 10' 65° C. | 6 sterile 3 averaged 686 |
0.395 | 0.362 | Not appreciably affected |
| 12 | 132,833 | 30' 65° C. | 9 sterile 3 averaged 233 |
0.395 | 0.362 | " |
| 13 | 49,867 | 10' 70° C. | sterile | 0.422 | 0.269 | Slightly boiled |
| 9 | 38,320 | 30' 70° C. | " | 0.421 | 0.253 | " |
| 2 | 77,062 | 10' 75° C. | " | 0.380 | 0.070 | Boiled |
| 3 | 48,250 | 30' 75° C. | " | 0.380 | 0.050 | " |
| 1 | 1,107,000 | 10' 80° C. | " | 0.375 | 0.000 | " |
| 1 | 1,107,000 | 30' 80° C. | " | 0.375 | 0.000 | " |
Respecting the numerical diminution of microbes brought
about by pasteurisation and sterilisation, respectively, we
may take the following two sets of experiments. Dr. N.
L. Russell63 tabulates the immediate results of pasteurisation
as follows:
| Unpasteurised. | Pasteurised. | |||||
| Minimum. | Maximum. | Average. | Minimum. | Maximum. | Average. | |
| Full cream milk. | 25,300 | 18,827,000 | 3,674,000 | 0 | 37,500 | 6,140 |
| Cream, 25%. | 425,000 | 32,800,000 | 8,700,000 | 0 | 57,000 | 24,250 |
As regards the later effect of the process, he states that in fifteen samples of pasteurised milk examined from November to December nine of them revealed no organisms, or so few that they might almost be regarded as sterile; in those samples examined after January the lowest number was 100 germs per cc., while the average was nearly 5,000. With the pasteurised cream a similar condition was to be observed.
Dr. Hewlett64 defines pasteurisation briefly as heating the milk to 68° C. for twenty or thirty minutes, and this treatment he quotes as destroying 99.75 per cent. of the total number of organisms. Bitter's table of results at 158° F. bears out the same:
| Before Pasteurisation. No. of Bacteria in 10 Drops. |
After Pasteurisation. No. of Bacteria in 10 Drops. |
|
| 1. | 102,600 | 2—3 |
| 2. | 251,600 | 30—40 |
| 3. | 25,000 | 3—5 |
| 4. | 37,500 | 2—5 |
| 5. | 94,000 | 2 |
Cream is generally richer in bacteria than milk. Set cream contains more bacteria than separated cream, but germs are abundant in both. Yet whilst it is true that cream contains a large number of bacteria, it must be pointed out that the butter fat in cream is a less suitable food for organisms than is the case with milk. Hence the fermentative changes set up in cream are of less degree than in milk, particularly so if separated from the milk. Butter-milk and whey vary much in their bacterial content. Butter necessarily follows the standard of the cream. But as the butter fat is not well adapted for bacterial food, the number of bacteria in 214butter is usually less than in cream.65 Moreover, they are soon reduced both in quality and quantity. Butter examined after it is several months old is often found to be almost free from germs; yet in the intervening period a variety of conditions are set up directly or indirectly through bacterial action.
Rancid butter is partly due to organisms. Putrid butter is caused, according to Jensen, by various putrefactive bacteria, one form of which is named Bacillus fœtidus lactis. This organism is killed at a comparatively low temperature, and is therefore completely removed by pasteurisation. Ill-flavoured butter may be due to germs or an unsuitable diet of the cow and a retention of the bad quality of the resulting milk. Lardy and oily butters have been investigated by Storch and Jensen and traced to bacteria. Lastly, bitter butter occasionally occurs, and is due to fermentative changes in the milk. Butter may also contain pathogenic bacteria, like tubercle. The B. coli can live for one month in butter.
Cheese suffers from very much the same kind of "diseases" as butter, except that chromogenic conditions occur more frequently. The latter are, under certain circumstances, more the result of chemical than bacterial action. Most of the troubles in cheese originate in the milk.
Method of Examination of Butter. Several grams of the butter should be placed in a large test-tube, which is then two-thirds filled with sterilised water and placed in a water-bath at about 45° C. until the butter is completely melted. A small quantity may then be added to gelatine or agar and plated out on Petri dishes or in flat-bottomed flasks in the usual way. After which the tube may be well shaken and returned to the bath inverted. In the space of twenty or thirty minutes the butter has separated from the water with 215which it has been emulsified. It is then placed in the cold to set. The water may be now either centrifugalised or placed in sedimentation flasks, and the deposit examined for bacteria.
The Uses of Bacteria in Dairy Produce. In considering the relation of bacteria to milk we found that many of the species present were injurious rather than otherwise, and when we come to consider bacteria in dairy products, like butter and cheese, we find that the dairyman possesses in them very powerful allies. Within recent years almost a new industry has arisen owing to the scientific application of bacteriology to dairy work.
As a preliminary to butter-making the general custom in most countries is to subject the cream to a process of "ripening." As we have seen, cream in ordinary dairies and creameries invariably contains some bacteria, a large number of which are in no sense injurious. Indeed, it is to these bacteria that the ripening and flavouring processes are due. They are perfectly consistent with the production of the best quality of butter. The aroma of butter, as we know, controls in a large measure its price in the market. This aroma is due to the decomposing effect upon the constituents of the butter of the bacteria contained in the cream. In the months of May and June the variety and number of these types of bacteria are decidedly greater than in the winter months, and this explains in part the better quality of the butter at these seasons. As a result of these ripening bacteria the milk becomes changed and soured, and slightly curdled. Thus it is rendered more fit for butter-making, and acquires its pleasant taste and aroma. It is then churned, after which bacterial action is reduced to a minimum or is absent altogether. Sweet-cream butter lacks the flavour of ripened or sour-cream butter. The process is really a fermentation, the ripening bacteria acting on each and all of the constituents of the milk, resulting in the216 production of various bye-products. This fermentation is a decomposition, and just as we found when discussing fermentation, so here also the action is beneficial only if it is stopped at the right moment. If, for example, instead of being stopped on the second day, it is allowed to continue for a week, the cream will degenerate and become offensive, and the pleasant ripening aroma will be changed to the contrary.
Bacteriologists have demonstrated that butters possessing different flavours have been ripened by different species of bacteria. Occasionally one comes across a dairy which seems to be impregnated with bacteria that improve cream and flavour well. In other cases the contrary happens, and a dairy becomes impregnated with a species having deleterious effects upon its butter. This species may arise from unclean utensils and dairying, from disease of the cow, or from a change in the cow's diet. Thus it comes about that the butter-maker is not always able to depend upon good ripening for his cream. At other times he gets ripening to occur, but the flavour is an evil one, and the results correspond. It may be bitter or tainted, and just as certainly as these flavours develop in the cream, so is it certain that the butter will suffer. Fortunately the bacterial content of the cream is generally either favourable or indifferent in its action. Thus it comes about that the custom is to allow the cream simply to ripen, so to speak, of its own accord, in a vat exposed to the influence of any bacteria which may happen to be around. This generally proves satisfactory, but it has the great disadvantage of being indefinite and uncertain. Occasionally it turns out wholly unsatisfactory, and results in financial loss.
There are various means at our command for improving the ripening process. Perfect cleanliness in the entire manipulation necessary in milking and dairying, combined with freedom from disease in the milch cows, will carry us a217 long way on the road towards a good cream-ripening. Recently, however, a new method has been introduced, largely through the work and influence of Professor Storch in Denmark, which is based upon our new knowledge respecting bacterial action in cream-ripening. We refer to the artificial processes of ripening set up by the addition of pure cultures of favourable germs.66 If a culture of organisms possessing the faculty of producing in cream a good flavour be added to the sweet cream, it is clear that advantage will accrue. This simple plan of starting any special or desired flavour by introducing the specific micro-organism of that flavour may be adopted in two or three different ways. If cream be inoculated with a large, pure culture of some particular kind of bacteria, this species will frequently grow so well and so rapidly that it will check the growth of the other bacteria which were present in the cream at the commencement and before the starter was added. That is, perhaps, the simplest method of adding an artificial culture. But secondly, it will be apparent to those who have followed us thus far, that if the cream is previously pasteurised at 70° C. these competing bacteria will have been mostly or entirely destroyed, and the pure culture, or starter, will have the field to itself. There is a third modification, which is sometimes termed ripening by natural starters. A natural starter is a certain small quantity of cream taken from a favourable ripening—from a clean dairy or a good herd—and placed aside to sour for two days until it is heavily impregnated with the specific organism which was present in the whole favourable stock of which the natural starter is but a part. It is then added to the new cream the favourable ripening of which is desired. Of the species which produce good flavours in butter the majority are found to 218be members of the acid-producing class; but probably the flavour is not dependent upon the acid. Moreover, the aroma of good ripening is also probably independent of the acid production.
Of all the methods of ripening—natural ripening, the addition of natural starters, the addition of pure cultures with or without pasteurisation—there can be no doubt that pure culture after pasteurisation is the most accurate and dependable. The use of natural starters is a method in the right direction; yet it is, after all, a mixed culture, and therefore not uniform in action. In order to obtain the best results with the addition of pure cultures, Professor Russell has made the following recommendations:
1. The dry powder of the pure culture must be added to a small amount of milk that has been first pasteurised, in order to develop an active growth from the dried material.
2. The cream to be ripened must first be pasteurised, in order to destroy the developing organisms already in it, and thus be prepared for the addition of the pure culture.
3. The addition of the developing starter to the pasteurised cream and the holding of the cream at such a temperature as will readily induce the best development of flavour.
4. The propagation of the starter from day to day. A fresh lot of pasteurised milk should be inoculated daily with some of the pure culture of the previous day, not the ripening cream containing the culture. In this way the purity of the starter is maintained for a considerable length of time. Those starters are best which grow rapidly at a comparatively low temperature (60–75° F.), which produce a good flavour, and which increase the keeping qualities of the butter. Now, whilst it is true that the practice of using pure cultures in this way is becoming more general, very few species have been isolated which fulfil all the desirable qualities above mentioned. In America starters are pre219ferred which yield a "high" flavour, whereas in Danish butter a mild aroma is commoner. In England as yet very little has been done, and that on an experimental scale rather than a commercial one.67 In 1891 it appears that only 4 per cent. of the butter exhibited at the Danish butter exhibitions was made from pasteurised cream plus a culture starter; but in 1895, 86 per cent. of the butter was so made. Moreover, such butter obtained the prizes awarded for first-class butter with preferable flavour. Different cultures will, of course, yield different flavoured butter. If we desire, say, a Danish butter, then some species like "Hansen's Danish Starter" would be added; if we desire an American butter, we should use a species like that known as "Conn's Bacillus, No. 41." But whilst these are two common types, they are not the only suitable and effective starters. On certain farms in England there are equally good cultures, which, placed under favourable temperatures in new cream, would immediately commence active ripening.
Professor H. W. Conn, who, with Professor Russell, has done so much in America for the advancement of dairy bacteriology, reports68 a year's experience with the bacillus to which reference has been made, and which is termed No. 41. It was originally obtained from a specimen of milk from Uruguay, South America, which was exhibited at the World's Fair in Chicago, and proved the most successful flavouring and ripening agent among a number of cultures that were tried. The conclusions arrived at after a considerable period of testing and experimentation appear to be on the whole satisfactory. A frequent method of testing 220has been to divide a certain quantity of cream into two parts, one part inoculated with the culture and the other part left uninoculated. Both have then been ripened under similar conditions, and churned in the same way; the differences have then been noted. It is interesting to know that, as a result of the year's experience, creameries have been able to command a price varying from half a cent to two cents a pound more for the "culture" butters than for the uninoculated butters. The method advised in using this pure culture is to pasteurise (by heating at 155° F.) six quarts of cream, and after cooling to dissolve in this cream the pellet containing bacillus No. 41. The cream is then set in a warm place (70° F.), and the bacillus is allowed to grow for two days, and is then inoculated into twenty-five gallons of ordinary cream. This is allowed to ripen as usual, and is then used as an infecting culture, or "starter," in the large cream vats in the proportion of one gallon of infecting culture to twenty-five gallons of cream, and the whole is ripened at a temperature of about 68° F. for one day. The cream ripened by this organism needs to be churned at a little lower temperature (say 52°-54° F.) but to be ripened at a little higher temperature than ordinary cream to produce the best results. Cream ripened with No. 41 has its keeping power much increased, and the body or grain of the butter is not affected. More than two hundred creameries in America used this culture during 1895, and Professor Conn reports that this has proved that its use for the production of flavour in butter is feasible in ordinary creameries and in the hands of ordinary butter-makers provided they will use proper methods and proper discretion.
Bacteria in Cheese-making. The cases where it has been possible to trace bacterial disease to the consumption of butter and cheese have been rare. Notwithstanding this fact, it must not be supposed that therefore cheese contains few or no bacteria. On the contrary, for the making of221 cheese bacteria are not only favourable, but actually essential, for in its manufacture the casein of the milk has to be separated from the other products by the use of rennet, and is then collected in large masses and pressed, forming the fresh cheese. In the course of time this undergoes ripening, which develops the peculiar flavours characteristic of cheese, and upon which its whole value depends.
We have said that the casein is separated by the addition of rennet, which has the power of coagulating the casein. But this precipitation may also be accomplished by allowing acid to develop in the milk until the casein is precipitated, as in some sour-milk or cottage cheeses. The former method is of course the usual one in practice. It has been suggested that the bacteria contained in the rennet exert a considerable influence on the cheese, but this, although rennet contains bacteria, is hardly established. It is not here, however, that bacteria really play their rôle. After this physical separation, when the cheese is pressed and set aside, is the period for the commencement of the ripening process.
That bacteria perform the major part of this ripening process, and are essential to it, is proved by the fact that when they are either removed or opposed the curing changes immediately cease. If the milk be first sterilised, or if antiseptics, like thymol, be added, the results are negative. It is not yet known whether this peptonising process is due to the influence of a single organism or not. The probability, however, is that it is to be ascribed to the action of that group of bacteria known as the lactic-acid organisms. Nor is it yet known whether the peptonisation of the casein and the production of the flavour are the results of one or more species. Freudenreich believes them to be due to two different forms.
However that may be, we meet with at least four common groups of bacteria more or less constantly present in222 cheese-ripening, either in the early or late stages. First, there are the lactic-acid bacteria, by far the largest group, and the one common feature of which is the production by fermentation of lactic acid; secondly, there are the casein-digesting bacteria, present in relatively small numbers; thirdly, the gas-producing bacteria, which give to cheese its honeycombed appearance; lastly, an indifferent or miscellaneous group of extraneous bacteria, which were in the milk at the outset of cheese-making, or are intruders from the air or rennet. All these four groups may bring about a variety of changes, beneficial and otherwise, in the cheese-making.
In order that the relation of bacteria to cheese may be more fully understood, we may draw attention to some experiments conducted by Professor H. L. Russell as to the numbers of bacteria present during different stages of the ripening, excluding those already referred to as present in the rennet. It appears that there is always at first a marked increase in the number of micro-organisms, which is soon followed by a more gradual decline. While the casein-digesting and gas-producing classes suffer a general and more or less rapid decline, the lactic-acid bacteria develop to an enormous extent, from which fact it would appear that cheese offers ideal conditions for the development of the latter. In some most interesting records Professor Russell has divided the ripening process into three divisions:
1. Period of Initial Bacterial Decline in Cheese. Where the green cheeses were examined immediately after removing from the press, it was usually found that a diminution in numbers of bacteria had taken place. This period of decline lasts but a short time, not beyond the second day. Lower temperature and expulsion of the whey would account for this general decline in all species of bacteria.
2. Period of Bacterial Increase. Soon after the cheese is removed from the press a most noteworthy change takes place223 in green cheese. A very rapid increase of bacteria occurs, confined almost exclusively to the lactic-acid group. This commences in green cheese about the eighth day, and continues more or less for twenty days. In Cheddar cheese it commences about the fifth day, reaches its maximum about the twentieth day, declines rapidly to the thirtieth day, and gradually for a hundred following days. During the first forty days of this period the casein-digesting and gas-producing organisms are present, and at first increasing, but relatively to only a very slight degree. With this rapid increase in organisms the curd begins to lose its elastic texture, and before the maximum number of bacteria is reached the curing is far advanced. Freudenreich has shown that acid inhibits the growth of the casein-digesting microbes and vice versâ.
3. Period of Final Bacterial Decline. The cause of this decline can only be conjectured, but it is highly probable that it is due to a general principle to which reference has frequently been made, viz., that after a certain time the further growth of any species of bacteria is prevented by its own products. We may observe that the gas-producing bacteria in Cheddar cheese last much longer than the peptonising organisms, for they are still present up to eighty days. Professor Russell aptly compares the bacterial vegetation of cheese with its analogue in a freshly seeded field. "At first multitudes of weeds appear with the grass. These are the casein-digesting organisms, while the grass is comparable to the more native lactic-acid flora. In course of time, however, grass, which is the natural covering of soil, 'drives out' the weeds, and in cheese a similar condition occurs." In milk the lactic-acid bacteria and peptonising organisms grow together; in ripening cheese the former eliminate the latter.
We have seen that the conclusion generally held respecting these lactic-acid bacteria is that they are the main agents224 in curing the cheese. Upon this basis a system of pure starters has been adopted, the characteristics of which must be as follows: (a) The organism shall be a pure lactic-acid-producing germ, incapable of producing gaseous products; (b) it should be free from any undesirable aroma; (c) it should be especially adapted for vigorous development in milk. The starter may be propagated in pasteurised or sterilised milk from a pure culture from the laboratory. The advantages accruing from the uses of this lactic-acid culture, as compared with cheese made without a culture, are that with sweet milk it saves time in the process of manufacture; that with tainted milk, in which acid develops imperfectly, it is an aid to the development of a proper amount of acid for a typical Cheddar cheese; and that the flavour and quality of such cheese is preferable to cheese which has not been thus produced. Professor Russell is of opinion that the lactic-acid organisms are to be credited with greater ripening powers than the casein-digesting organisms, but it must not be forgotten that these two great families of bacteria are still more or less on trial, and it is not yet possible finally to dispose of either of them. Mr. F. J. Lloyd holds that though "the greater the number of lactic-acid bacilli in the milk the greater the chance of a good curd," still "this organism alone will not produce that nutty flavour which is so sought after as being the essential characteristic of an excellent Cheddar cheese."69
There are several difficulties to be encountered by dairymen starting a ripening by the addition of a pure culture. To begin with, there is the initial difficulty of not being able to pasteurise milk intended for cheese, as rennet will not coagulate pasteurised milk (Lloyd). Hence it is impossible 225to avoid some contamination of the milk previous to the addition of the culture. The continual uncontaminated supply of pure culture is by no means an easy matter. The maintenance of a low temperature to prevent the rapid multiplication of extraneous bacteria will, in some localities, be a serious difficulty. These difficulties have, however, not proved insurmountable, and by various workers in various localities and countries culture-ripening is being carried on.
Abnormal Ripening. Unfortunately, from one cause or another, faulty fermentations and changes are not infrequently set up. Many of these may be prevented, being due to lack of cleanliness in the process or in the milking; others are due to the gas-producing bacteria being present in abnormally large numbers. When this occurs we obtain what is known as "gassy" cheese, on account of its substance being split up by innumerable cavities and holes containing carbonic acid gas, or sometimes ammonia or free nitrogen. Some twenty-five species of micro-organisms have been shown by Adamety to cause this abnormal swelling. In severe cases of this gaseous fermentation the product is rendered worthless, and even when less marked the flavour and value are much impaired. Winter cheese contains more of this species of bacteria than summer. Acid and salt are both used to inhibit the action of these gas-producing bacteria and yeasts, and with excellent results.
We may remark that the character of the gas holes in cheese is not of import in the differentiation of species. If a few gas bacteria are present, the holes will be large and less frequent; if many, the holes will be small, but numerous. (Swiss cheese having this characteristic is known as Nissler cheese.)
Many of these gas germs belong to the lactic-acid group, and are susceptible to heat. A temperature of 140° F. maintained for fifteen minutes is fatal to most of them, largely because they do not form spores. The sources of226 the extensive list of bacteria found in cheese are of course varied, more varied indeed than is the case with milk. For there are, in addition to the organisms contained in the milk brought to the cheese factory, the following prolific sources, viz., the vats and additional apparatus; the rennet (which itself contains a great number); the water that is used in the manufacture.
In addition to the abnormalities due to gas, there are also other faulty types. The following chromogenic conditions occur: red cheese, due to a micrococcus; blue cheese, produced, according to Vries, by a bacillus; and black cheese, caused by a copious growth of low fungi. Bitter cheese is the result of the Micrococcus casei amari of Freudenreich, a closely allied form of Conn's micrococcus of bitter milk. Sometimes cheese undergoes a putrefactive decomposition, and becomes more or less putrid. These latter conditions, like the gassy cheeses, are due to the intrusion of bacteria from without, or from udder disease of the cow. Healthy cows, clean milking, and the introduction of pure cultures are the methods to be adopted for avoiding "diseases" of cheese and obtaining a well-flavoured article which will keep.
Finally, we may quote five conclusions from the prolonged researches of Mr. Lloyd70 which cannot but prove helpful to the Cheddar cheese industry in England:
1. To make Cheddar cheese of excellent quality, the Bacillus acidi lactici alone is necessary; other germs will tend to make the work more rather than less difficult. Hence scrupulous cleanliness should be a primary consideration of the cheese-maker.
2. No matter what system of manufacture be adopted, two things are necessary. One is that the whey be separated from the curd, so that when the curd is ground it shall contain not less than 40 per cent. of water, and not more than 22743 per cent.; the other point is that the whey left in the curd shall contain, developed in it before the curd is put in the press, at least 1 per cent. of lactic acid if the cheese is required for sale within four months, and not less than 8 per cent. of lactic acid if the cheese is to be kept ripening for a longer period.
3. The quality of the cheeses will vary with the quality of the milk from which they have been made, and proportionately to the amount of fat present in that milk.
4. "Spongy curd" is produced by at least five organisms, and one of these is responsible for a disagreeable taint found in curd. They occur in water. Hence the desirability of securing clean water for all manipulative purposes, and also for the drinking purposes of the milch cow.
5. The fact that certain bacteria are found in certain localities and dairies is due more to local conditions than to climatic causes.
It is needless to remark that these conclusions once more emphasise the fact that strict and continual cleanliness is the one desideratum for bacteriologically good dairying. That being secured in the cow at the milking, in the transit, and at the dairy, it is a comparatively simple step, by means of pasteurisation and the use of good pure cultures of flavouring bacteria, to the successful application of bacteriology to dairy produce.
Methods of Examination of Milk:
1. Preparation of Microscopic Slides. This course might at once occur to the mind as the first to adopt in searching for bacteria in milk. Devices have accordingly been proposed for saponification previous to staining. Some recommend the addition of a few drops of a solution of sodium carbonate; others use methylene blue and chloroform. But, whatever plan of staining is adopted, this method of examination in its simplest form is in no degree a criterion of the bacterial content of a large quantity of milk.
Hence it has come to be recognised that one of two
manipulations must precede such microscopic examination.
These simple processes are known by the terms of sedimentation
and centrifugalisation. Sedimentation means merely
 A Centrifuge
A Centrifuge
Used in the Examination
of Milk
placing the milk in conical glasses in a cool
place for twenty-four hours. The introduction
of improved forms of the centrifuge
has brought the second method of
securing a sediment into preference. Five
cubic centimetres of the milk are introduced
into the graduated bottle, which is
then placed in the centrifuge, and whirled
for one or two minutes. Thus a deposit
of particulate matter is ensured. Cover-glass
specimens of the sediment or deposit
are then prepared and stained in the ordinary
way.
In testing for tubercle something more is generally necessary. To the 50 cc. of the milk set aside for sedimentation 10 cc. of liquefied, colourless carbolic acid are added. The mixture is shaken and poured into the conical glass. After standing for twenty-four hours a little of the sediment is taken by means of a pipette and examined by ordinary methods, though after "fixing" the films with heat they are some times passed through equal parts of alcohol and ether. The stain is of course that usually adopted in tubercle, namely, the Ziehl-Neelsen. Scheurlen suggested a method for demonstrating the tubercle bacillus in milk by steeping the cover glasses first in alcohol and then ether, after which they were stained with Ziehl-Neelsen.
2. Plate Culture. The milk is to be diluted a thousand or229 more times with sterile water, and ordinary plate cultures made in Petri dishes or flat-bottomed conical flasks. The colonies should be counted as late as possible; but even then the isolation of pathogenic germs is uncertain. As regards further procedure, the ordinary methods of sub-culturing adopted in water examination must be strictly followed, and the special tests for Bacillus typhosus and B. coli applied. As we have already seen, the quantitative estimation of organisms in milk is not of the same value as in water.
3. Inoculation. To test the capacity of the milk for causing disease, before or after centrifugalisation, preferably the latter, a certain quantity of the sediment may be inoculated into guinea-pigs. In suspected tubercle 2 cc. may be taken; in diphtheria a little less will suffice. The inoculation should be either intraperitoneal or subcutaneous. Many authorities hold that this test is the only safe one to protect the public from milk containing germs of disease.
Shell-fish have recently claimed the attention of bacteriologists, owing to the outbreak of typhoid and other epidemics apparently traceable to oysters.
It is four or five years since Professor Conn startled the medical world by tracing an epidemic of typhoid fever to the consumption of some uncooked oysters.71 Almost at the same time Sir William Broadbent published in the British Medical Journal a series of cases occurring in his practice which illustrated the same channel of infection. Since then a number of similar items of evidence to the same effect have cropped up. Hence there is little wonder that a number of investigators concentrated their attention upon this matter. Professors Herdman and Boyce, of Liverpool, Dr. Cartwright Wood, Dr. Klein, and Dr. Timbrell Bulstrode 230are some of the chief contributors to the elucidation of this problem.
The mode of infection of oysters by pathogenic bacteria is briefly as follows: The sewage of certain coast towns is passed untreated out to sea. At or near the outfall, oyster-beds are laid down for the purpose of fattening oysters. Thus they become contaminated with saprophytic and pathogenic germs contained in the sewage. It will be at once apparent that several preliminary questions require attention before any deductions can be drawn as to whether or not oysters convey virulent disease to consumers. To the solution of these Dr. Cartwright Wood was one of the first to address himself.
The precise conditions which render one locality more favourable than another in respect to oyster culture are not fully known. But it has been observed that they do not flourish in water containing less than three per cent. of salt. Hence they are absent from the Baltic Sea, which, owing to the fresh water flowing into it in rivers, contains a smaller percentage of salt than three. Oysters appear in addition, to favour a locality where they find their chosen food of small animalculæ and particles of organic matter. Such a favourable locality is the mouth of a river, where tides and currents also assist in bringing food to the oyster. Unfortunately, however, in a crowded country like England such localities round her coasts are frequently contaminated by sewage from outfalls. Thus the oysters and the sewage come into intimate relation with each other.
Professor Giaxa carried out some experiments in 1889 at Naples which appeared to show that the bacilli of cholera and typhoid rapidly disappeared in ordinary sea-water. Other observers at about the same time, notably Foster and Freitag, arrived at an opposite conclusion. In 1894 Professor Percy Frankland, in a report to the Royal Society, declared "that common salt, whilst enormously stimulating231 the multiplication of many forms of water bacteria, exerts a directly and highly prejudicial effect on the typhoid bacilli, causing their rapid disappearance from the water, whether water bacteria are present or not." It was at this time, when the matter was admittedly in an unsatisfactory stage, that Dr. Cartwright Wood made his experiments.72 We have not space here to enter into this work. But his conclusions seem to have been amply established, and were to the effect that typhoid and cholera bacilli could, as a matter of fact, exist over very lengthened periods in ordinary sea-water. The next step was to demonstrate the length of time the bacilli of cholera remained alive in the pallial cavity and body of the oyster. Dr. Wood found they did so for eighteen days after infection, though in greatly diminished numbers. This diminution was due to one or all of three reasons: (a) the effect of the sea-water already referred to as finally prejudicial to bacilli of typhoid; (b) the vital action of the body-cells of the oyster; (c) the washing away of bacilli by the water circulating through the pallial cavity.
It will have been noticed that up to the present we have learned that typhoid bacilli can and do live in sea-water, and also inside oysters up to eighteen days, but in ever-diminishing quantities. The question now arises: What is the influence of the oyster upon the contained bacilli? Under certain conditions of temperature organisms may multiply with great rapidity inside the shell of the oyster. Yet, on the other hand, the amœboid cells of the oyster, the acid secretion of its digestive glands, or the water circulating through its pallial cavity, may act inimically on the germs. Proof can be produced in favour of the third and last-named mode by which an oyster can cleanse itself of germs. So far, then, we have met with no facts which make it impossible for oysters to contain for a lengthened period the specific bacteria of disease. Let us now turn to 232their opportunity for acquiring such disease germs. It is afforded them during the process of what is termed "fattening." By this process the body of the oyster acquires a plumpness and weight which enhances its commercial value. This desired condition is obtained by growing the oyster in "brackish" water, for thus it becomes filled out and mechanically distended with water. But if this water contains germs of disease, what better opportunity could such germs have for multiplication than within the body-cavity of an oyster? "The contamination of sea-water, therefore, in the neighbourhood of oyster-beds may undoubtedly lead to the molluscs becoming infected with pathogenic organisms" (Wood). Yet we have seen that, apart altogether from the individual susceptibilities or otherwise of the consumer, there are in the series of events necessary to infection many occasions when circumstances would practically free the oysters from infection.
The sources of pollution of oysters are not the fattening beds alone. The native beds also may afford opportunity for contamination. Thirdly, in packing and transit, and fourthly, in storage in shops and warehouses, there is frequently abundant facility for putrefactive bacteria to gain entrance to the shells of oysters.
Dr. Klein's researches73 into this question have been wholly confirmatory of the facts elicited by Dr. Cartwright Wood. Despite the tendency of the bacilli of cholera and typhoid to die out quickly in crude sewage, the sewage is sufficiently altered or diluted at the outfall for these organisms to exist there in a virulent state. We may give Dr. Klein's conclusions:
1. That the cholera and typhoid bacilli are difficult of demonstration in sewage known to have received them.
2. Both organisms may persist in sea-water tanks for two 233 or three weeks, the typhoid bacillus retaining its characteristics unimpaired, the cholera bacillus tending to lose them.
3. Oysters from sources free of sewage contained no bacteria of sewage.
4. Oysters from sources exposed to risk of sewage contamination did contain colon bacilli and other sewage bacteria.
5. In one case Eberth's typhoid bacillus was found in the mingled body and liquor of the oyster.
Nor do typhoid bacilli lose activity or virulence by passing through an oyster.
These researches once and for all established the fact that oysters ordinarily grown on oyster-beds contaminated with bacteria may, and do on occasion, contain the virulent specific bacillus of typhoid, which can live both in sea-water and within the shell of the oyster. This being so, it will probably appear to the reader that the risk of infection of typhoid by oysters is very serious indeed. Yet in actual practice many conditions have to be fulfilled. For, in addition to the fact that the oysters must be consumed, as is usual, uncooked, the following conditions must also be present.
(a) Each infective oyster must contain infected sewage, which presupposes that typhoid excreta from patients suffering from the disease have passed into that particular sewage untreated and not disinfected.
(b) The infective oyster must be fed upon infected sewage, and still contain the virus in its substance.
(c) It has to be eaten by a susceptible person.
(d) There must have been no period of natural cleansing after "fattening."
Even to this formidable list of conditions we must add the further remark that, owing to the vitality of the body-cells of the oyster, or to the lessened vitality of the bacilli of cholera and typhoid, it is generally the case that the tend234ency of these organisms is rather to decrease and die out than live and multiply.
We shall probably maintain a satisfactory balance of truth if we place alongside these facts the summary of the Local Government Board Report.
"There can be no doubt," wrote Sir Richard Thorne, "that oysters which have been brought into sustained relation with the typhoid bacillus are liable to exhibit that microbe within the shell contents and to retain it for a while under circumstances not only permitting its rapid multiplication when transferred again to appropriate media, but conserving at the same time its ability to manifest its hurtful properties."
From what has been said the preventive treatment is obvious. All oyster-layings and shell-fish beds round the coast should be superintended and inspected by the sanitary authority of the Government. The importation of foreign oysters, grown on uncontrolled beds, should, if possible, be restricted or supervised. Further, as a protective measure of the first importance, oysters should be cleansed, after fattening on a contaminated bed, by being deposited for several weeks at some point along the coast which is washed by pure sea-water. Retention in dirty water-tanks, in uncleanly shops and warehouses, is also to be greatly deprecated.
In order to examine oysters bacteriologically, it is necessary to pay particular attention to the water in the pallial cavity, the contents of the alimentary canal, and the washings of the shell itself. Ordinary media may be used for obtaining a growth of the contained organisms.
Other shell-fish than oysters do, from time to time, cause epidemics or individual cases of gastro-intestinal irritation, and probably contain various germs. These they acquire in all probability from their food, which by their own choice is frequently of a doubtful character.
Meat. Parasites are occasionally found in meat, but bacteria are comparatively rare. Not that they do not235 occur in the bodies of animals used for human consumption, for in the glands, mesenteries, and other organs they are common. But in those portions of the carcass which are used by man, namely the muscles, bacteria are rare. The reasons alleged for this are the acid reaction (sarcolactic acid) and the more or less constant movement during life. A bacterial disease which, perhaps more than any other, might be expected to be conveyed by meat is tubercle. Yet the recent Royal Commission on Tuberculosis has again emphasised the absence of bacilli in the meat substance:
"In tissues which go to form the butcher's joint, the material of tubercle is not often found even where the organs (lungs, liver, spleen, membranes, etc.) exhibit very advanced or generalised tuberculosis; indeed, in muscle and muscle juice it is very seldom that tubercle bacilli are to be met with; perhaps they are somewhat more often to be discovered in bone, or in some small lymphatic gland embedded in intermuscular fat."74
The only way in which such meat substance becomes infected with tubercle appears to be through carelessness in the butcher, who perchance smears the meat substance with a knife that has been used in cutting the organs, and so has become contaminated with infected material. Very instructive also are the results at which Dr. Sims Woodhead arrived in compiling evidence for the same Commission on the effect of cooking upon tuberculous meat:
"Ordinary cooking, such as boiling and more especially roasting, though quite sufficient to sterilise the surface, and even the substance for a short distance from the surface of a joint, cannot be relied upon to sterilise tubercular material included in the centre of rolls of meat, especially when these are more than three pounds or four pounds weight. The least reliable method of cooking for this purpose is roasting before a fire; next comes roasting in an oven, and then boiling."75
From this statement it will be understood that rolled meat may be a source of infection to a greater degree than the ordinary joint.
Notwithstanding this negative evidence, more than twenty species of bacteria have been isolated from canned meats and hams, and a considerable number of poisoning cases have occurred from meat contaminated with bacteria or their products. The general symptoms of such meat poisoning are vomiting, diarrhœa, fever, and more or less prostration. Ballard and Klein isolated a specific microbe from samples of bacon which appear to have caused an epidemic of infectious pneumonia at Middlesborough. In 1880 occurred the well-known "Welbeck disease" epidemic. A public luncheon was followed by severe and even fatal illness. Seventy-two persons were affected, and four died. A specific bacillus was isolated by Klein. In 1881 much the same thing happened at Nottingham, in which fifteen persons were attacked, and one died. The same bacillus was isolated from the pernicious pork. Again in 1889 an outbreak of diarrhœa at Carlisle was traced to bacterially diseased pork. But taking these and similar cases at their worst, there can be no doubt that under no circumstances is meat as infective as milk.
Ice-cream. In 1894 Dr. Klein had occasion to bacteriologically examine ice-creams sold in the streets of London. In all six samples were analysed, and in each sample the conclusions resulting were of a nature sufficiently serious to support the view that the bacterial flora was not inferior to ordinary sewage. The water in which the ice-cream glasses were washed was also examined, and found to contain large numbers of bacteria.
Since that date many investigations have been made into ice-creams. It appears that they are often made under extremely foul circumstances, and with anything but sterilised appliances. Little wonder, then, that the numbers of bac237teria present run into millions. In nearly all recorded cases the quality of the germs as well as the quantity has been of a nature to cause some concern. Bacillus coli communis, which, though not now considered absolutely indicative of alimentary pollution, is looked upon as a highly unsatisfactory inhabitant of water, has been found in considerable abundance. The Proteus family, which also possesses a putrefactive function, is common in ice-creams. The common water bacteria are nearly always present.
Bacillus typhosus itself, it is said, has been isolated from some ice-cream which was held responsible for an outbreak of enteric fever. The material had become infected during process of manufacture in the house of a person suffering from unnotified typhoid fever.
Now, whilst reports of the above nature appear very alarming, the fact is that hundreds of weakly children devour ice-cream with apparent impunity, and when evil follows it is not infrequently due to other than bacterial conditions. The cold mass itself may inhibit the resistance of the gastric tissues. Tyrotoxicon, the alkaloid separated from cheese and cream by Vaughan, may be responsible for some alimentary irritation. On the whole, the practical effect upon the community is not in proportion to the bacterial content of the ice-cream. Yet, nevertheless, we ought to be much more watchful than in the past to preserve ice-cream from pollution with harmful bacteria.
The two chief constituents which contribute their quota of germ life to ice-cream are ice and cream. In addition, the uncleanly methods of manufacture render the material likely to contain the six or seven millions of micro-organisms per cc. which have been on several occasions estimated. To cleanly methods of dairying we have already fully referred; to the bacterial content of milk and cream we have also paid some attention; but we have not had an opportunity of saying anything of germs in ice.
Ice contains bacteria in varying quantities from 20 per cc. to 10,000 or more. Nor is variation in number affected alone by the condition of the water, for samples collected from one and the same place differ widely. The quality follows in large measure the standard of the water.
Water bacteria, Bacillus coli, putrefactive bacteria, and even pathogenic have been found in ice. Many of the latter can live without much difficulty and are most numerous in ice containing air-bubbles.
Dr. Prudden, of New York, performed a series of experiments in 1887 to show the relative behaviour of bacteria in ice. Taking half a dozen species, he inoculated sterilised water and reduced it to a very low temperature for a hundred and three days, with the following results:—Bacillus prodigiosus diminished from 6,300 per cc. to 3,000 within the first four days, to 22 in thirty-seven days, and vanished altogether in fifty-one days; a liquefying water bacillus, numbering 800,000 per cc. at the commencement, had disappeared in four days; Staphylococcus pyogenes aureus and B. fluorescens showed large numbers present at the end of sixty-six and seventy-seven days respectively; B. typhosus, which was present 1,000,000 per cc. after eleven days, fell to 72,000 after 77 days, and 7,000 at the end of 103 days. Anthrax bacilli are susceptible to freezing, but their spores are practically unaffected (Frankland).
From these facts it will be seen that bacteria live, but do not multiply, in ice.
In making a bacterial investigation into the flora of ice-cream, it is necessary to remember that considerable dilution with sterilised water is required. The usual methods of examining water and milk are adopted.
Bread forms an excellent medium for moulds, but unless specially exposed the bacteria in it are few. Waldo and Walsh have, however, demonstrated that baking does not sterilise the interior of bread. These observers cultivated239 numerous bacteria from the centre of newly baked London loaves.76 The writer has recently made a series of examinations of the air of several underground bakehouses in Central London; but, though the air was highly impregnated with flour-dust, few bacteria were present.
Other foods and beverages may be, and are, from time to time contaminated in some small degree with bacteria or their spores. Such contaminations are generally due to uncleanly manufacture or unprotected storage. The principles of examination or of the prevention of pollution are similar to those already described.
THE term natural immunity is used to denote natural resistance to some particular specific disease. It may refer to race, or age, or individual idiosyncrasies. We not infrequently meet with examples of this freedom from disease. Certain races of men do not, as a rule, take certain diseases. For example, plague and leprosy, though endemic in some countries, fail to get a footing in England. This, of course, is due in great measure to the sanitary organisation and cleanly customs of the English people. Still, it is also due to the fact that the English appear in some degree to be immune. Some authorities hold that the immunity against leprosy is due to the fact that the disease has exhausted itself in the English race. However that may be, we know that immunity, entire or partial, exists. Children, again, are susceptible to certain diseases and insusceptible to certain others to which older people are susceptible. We know, too, that some individuals have a marked protection against some diseases. Some people coming into the way of infection at once fall victims to the disease, whilst others appear to be proof against it. It is only in recent times that any very intelligent explanations have been offered to account for this phenomenon. The most recent of these, and that which appears to have most to substantiate it, is known as immunity due to antitoxins.
The products of bacteria are chiefly six:
1. Pigment. We have already seen how many organisms exhibit their energy in the formation of many coloured pigments. They are, as a rule, "innocent" microbes. Oxygen is required for some, darkness for others, and they all vary according to the medium upon which they are growing. Red milk, yellow milk, and green pus afford examples of pigment produced by bacteria.
2. Gas. Quite a number of the common bacteria, like Bacillus coli, produce gas in their growth; hydrogen (H), carbonic acid (CO2), methane (CH4), and even nitrogen (N) being formed by different bacteria. Many gases produced during fermentative processes are the result, not directly of the growth of the bacillus causing the fermentation, but indirectly owing to the splitting up of the fermenting fluids.
3. Acids. Lactic, acetic, butyric, etc., are common types of acids resulting from the growth of bacteria.
4. Liquefying Ferment. As we have seen, bacteria may be classified with regard to their behaviour in gelatine medium, whether or not they produce a peptonising ferment which liquefies the gelatine.
5. Phosphorescence. Some species of bacteria in sea-water possess the power of producing light.
6. Organic Chemical Products. When a pathogenic bacillus grows either in the body or in a test-tube, it produces as a result of its metabolism certain poisonous substances called toxins. These may occur in the blood as a direct result of the life of the bacillus, or they may occur as the result of a ferment produced by the bacillus. They are of various kinds according to the various diseases, and by their effect upon the blood and body tissues they cause the symptoms of the disease in question. We know, for instance, that a characteristic symptom common to many diseases is fever. Now, fever is produced by the action of the albumoses242 (bodies allied to the proteids) upon the heat-regulating centres in the brain. Whenever we get a bacillus growing in the body which has the power of producing a toxin albumose, we get fever as a result of that product acting upon the brain. Albumoses, as a matter of fact, cause a number of symptoms and poisonous effects, but the mention of one as an illustration will suffice. Toxins act, roughly speaking, in two ways:
(1) They have a local action, as, for example, in the formation of an abscess. The presence of the causal bacteria in the tissue brings about very marked changes. There is a multiplication of connective-tissue corpuscles, an emigration of leucocytic cells, a congestion of blood corpuscles. All these elements assist in creating a swelling and redness, and pain by the subsequent pressure upon the delicate nerve endings. These, as we all know, are the symptoms of a "gathering" or abscess. It is a "gathering" in a strict pathological sense—a gathering of cells to oust the intruder or build around it a wall or capsule as a protective measure. Now the toxin will commence its local action. The oldest cells in the mass of congestion will be caused to break down into liquid; what is called a necrosis, or death, will rapidly set in; and we shall have the connective-tissue cells, leucocytes, blood corpuscles, etc., losing their form and function, and "coming to a point" as matter, or pus. The local breaking down of these gatherings of cells into fluid matter is believed to be the work, not of the bacteria themselves, but of their toxins.
(2) Toxins may be absorbed and distributed generally throughout the body. They produce degenerative changes in muscles, in organs, and in the blood itself. Let us take diphtheria as an example. The bacillus occurs in a false membrane in the throat and occasionally other parts. It causes first the inflammatory condition giving rise to the membrane, and then it breaks it down. In the body of the243 membrane the bacillus appears to secrete a ferment which by its action and interaction with the body cells and proteids, chiefly those of the spleen, produces albumoses and an organic acid. These latter bodies are the toxins. They are absorbed, and pass throughout the body. There are albumoses, therefore we get the frequent pulse and high temperature of fever; the toxins irritate the mucous membrane of the intestine, and cause various fermentative changes in the contents of the intestine, therefore we get the symptoms of diarrhœa; they penetrate the liver, spleen, and kidney, therefore we get fatty degeneration and its results in these organs; they finally affect many of the motor and sensory nerves, breaking up their axis cylinders into globules, and therefore we get the characteristic paralysis. Loss of weight naturally follows many of these degenerative or wasting changes. Here, then, we have some of the chief changes set up by the toxins, and these changes constitute the leading symptoms in the disease as it is known clinically.
In addition to the presence of the specific bacillus in the membrane, we also have a number of other organisms, like the Bacillus coli, Coccus Brissou, Streptococcus pyogenes, and various staphylococci, diplococci, etc. Each of these produces or endeavours in the midst of keen competition and strife to produce, its own specific effect. Thus we obtain the complications of diphtheria, for example various suppurative and septic conditions. The whole of this compound process we may tabulate roughly as follows77:
| Bacillus coli. Coccus Brissou. Staphylococci. Diplococci. Streptococci. xxxxxx│ xxxxxx│ Toxins. xxxxxx│ xxxxxx│ Suppurative glands, septic poisoning, etc. |
Bacillus of Diphtheria xxxxxxxxx│ |
= primary infective agent | |
| Inflammatory changes and fibrinous exudation. xxxxxxxxx│ |
|||
| Ferment in Membrane xxxxxxxxx│ |
= secondary infective agent | ||
| Passes through body, and by digestion of proteids produces |
 |
ALBUMOSES; AN ORGANIC ACID |
|
| xxxxxx│ xxxxxx│ |
|||
| 1. Fever. 2. Diarrhœa. 3. Loss of body weight. 4. Fatty degeneration. 5. Degeneration of peripheral |
|||
Such is the general effect of toxins in diphtheria. The same principles apply with equal force in tetanus, typhoid, etc., the only differences being in degree of virulence, mode of onset, and portions of the body chiefly affected.
Sidney Martin has recently78 elaborated the views announced by him in 1892, and it is right that reference should be made to his new classification of bacterial poisons. This may be represented as follows:—
1. The poisons secreted by the bacterium itself = (ferment? toxin?) |
 |
= Extracellular bacterial poisons. |
2. Products of digestive action of bacterium = albumoses: |
||
3. Final non-proteid products = animal alkaloid; |
||
4. Poisons present in the bodies of the bacillus |
= Intracellular bacterial poisons. |
|
The poisons of bacteria are, according to Sidney Martin, of a kind which cannot be fully expressed chemically, but only pathologically. They may be of a ferment nature in diphtheria and tetanus. The arguments in support of that view are—(1) that they act in infinitesimal doses, (2) that they may act slowly and produce death after many days by 245profoundly affecting the general nutrition, and (3) that they are sensitive to the action of heat in a way that no chemical poisons are known to be. If they are considered as ferments, they must be substances which have a peculiar affinity for certain tissues of the body on which they produce their special toxic effect. As for the products of digestion, they are formed either by the bacillus ingesting the proteid and discharging it as albumose, or the digestion occurs by means of a ferment secreted by the bacillus in the body of an individual or animal suffering from the disease.
Sidney Martin suggests that anthrax produces albumoses and an alkaloidal substance, the former producing fever, the latter stupor. In tetanus the bacillus produces a secretion of the bacillus which causes the convulsions. The albumoses present in this disease are probably due to the secretory toxin. In diphtheria, too, we have a secretory poison in the membrane and in the tissues, and an albumose which is possibly the result of the secretion. It will be seen that these views differ in some particulars from those to which we have already referred.
However the details of the modus operandi of the formation of toxins are finally settled, we know that there comes a time when the disease symptoms vanish, the disease declines, and the patient recovers. Many of the older schools of medicine explained this satisfactory phenomenon by saying that this disease exhausted itself after having "gone through" the body. In a sense that idea is probably true; but recently a large number of investigators have applied themselves to this problem, and with some promising results.
Various protective inoculations against anthrax were practised as early as 1881, and the protected animals remained healthy. In 1887 Wooldridge succeeded in protecting rabbits from anthrax by a new method, by which he showed that the growth of the anthrax bacillus in special culture fluids gave rise to a substance which, when inoculated, con246ferred immunity. In 1889 and 1890 Hankin and Ogata worked at the subject, and announced the discovery in the blood of animals which had died of anthrax of some substances which appeared to have an antagonistic and neutralising effect upon the toxins of anthrax and upon the anthrax bacilli themselves. These substances, they afterwards found, were products of the anthrax bacillus. Behring and Kitasato arrived at much the same results for tetanus and diphtheria. The next step was to isolate these substances, and, separating them from the blood, investigate still further their constitution. A number of workers were soon occupied at this task, and since 1891 Buchner, Hankin, the Klemperers, Roux, Sidney Martin, and others have added to our knowledge respecting these toxin-opposing bodies known as antitoxins. In diphtheria, as we have seen, the toxins turned out to be soluble bodies allied to the proteids, albumoses, and an organic acid. Then arose the question of the source of antitoxins. Some believed they were a kind of ultratoxin—bodies of which an early form was a toxin; others held that, as the toxins were products of the bacteria invading the tissues, the antitoxins were of the nature of ferments produced by the resisting tissues. Finally, they came to be looked upon as protective substances produced in the body cells as a result of toxin action, and held in solution in the blood, and there and elsewhere exerting their influence in opposition to the toxins.79 The progress of disease is therefore a struggle between the toxins 247and the antitoxins: when the toxins are in the ascendency we get an increase of the disease; when the antitoxins are in the ascendency we get a diminution of disease. If the toxins triumph, the result is death; if the antitoxins and resistance of the tissues triumph, the result is recovery.
We may now consider shortly how these new facts were received and what theories of explanation were put forward to explain continued insusceptibility to disease. It had of course been known for a long time past that one attack of small-pox, for example, in some degree protected the individual from a subsequent attack of the same disease. To that experience it was now necessary to add a large mass of experimental evidence with regard to toxins and antitoxins. The theories of immunity were as follows:
1. The Exhaustion Theory. The supporters of this idea argued that bacteria of disease circulating in the body exhausted the body of the supply of some substance or condition necessary for the growth and development of their own species.
2. The Retention Theory. It was surmised that there were certain products of micro-organisms of disease retained in the body after an attack which acted antagonistically to the further growth in the body of that same species.
3. The Acquired Tolerance Theory. Some have advanced the theory that, after a certain time, the human tissues acquired such a degree of tolerance to the specific bacteria or their specific products that no result followed their action in the body. The tissues become acclimatised to the disease.
4. The Phagocyte Theory. This theory, which gained so many adherents when first promulgated by Metschnikoff, attributes to certain cells in the tissues the powers of "scavenging," overtaking germs of disease, and absorbing them into their own protoplasm. This, indeed, may be actually witnessed, and had been observed before the time of Metschnikoff. But it was he who applied it to disease.248 He came to the conclusion that the successful resistance which an animal offered to bacteria depended upon the activity of these scavenging cells, or phagocytes. These cells are derived from various cellular elements normally present in the body: leucocytes, endothelial cells, connective-tissue corpuscles, and any and all cells in the body which possess the power of ingesting bacteria. If they are present in large numbers and active, the animal is insusceptible to certain diseases; if they are few and inactive, the animal is susceptible.
It appears that the bacteria or other foreign bodies in the blood which are attacked by the phagocyte become assimilated until they are a part of the phagocyte itself. Metschnikoff explained also how it comes to pass that the phagocyte is able to encounter bacteria when both are circulating through the blood. It is guided in this attack upon the organisms by a power termed chemiotaxis. The bacteria elaborate a chemical substance which attracts the phagocyte, and this is termed "positive chemiotaxis."80 But it may occur that the chemical substance produced by the bacteria may have an opposite, or repellent, effect upon the leucocytes, in which case we have "negative chemiotaxis."81 It is not to be wondered at that such a theory of immunity based upon microscopical observations, should at first have been widely accepted, and there can be no doubt that Metschnikoff has collected a considerable mass of evidence in support of a theory of phagocytosis. But when it came to be known that blood serum, from which all leucocytes (phagocytes) had been removed, possessed the same immunising effect as before, it was clear that such effect was a property of the serum per se, and not only or wholly due to the scavenging power of certain cells in it. Even the 249phagocyte theory depends largely for its validity upon chemiotaxis, which latter was a property of the products of the bacteria contained in the blood serum.
5. The Antitoxin Theory. We have gathered, then, that whenever bacteria, introduced into the blood and tissues, fail to multiply or produce infection (as in saprophytic bacteria, or in immunity of a particular animal from a specific microbe), this inability to perform their rôle is brought about by some property in the living and normal blood serum which opposes their life and action; and further we have learned that this protective property is exhaustible according to the number of bacteria, and differs with various species of bacteria, and in different animals. Buchner designates these protective bodies, held in solution in the blood, alexines, and regards them as belonging to the albuminous bodies of the lymph and plasma. Where the blood and tissues do not possess this power, the animal is susceptible. Now, as we have already seen from the experiments of Ogata, Kitasato, and others, the blood of an animal dead of anthrax is protective against anthrax, from which and the foregoing it appears that microbes produce by their growth in the tissues poisonous substances we term toxins, which have the power of producing in the blood and body cells substances inimical to themselves, named antitoxins, and so long as these latter substances remain in the tissues the body remains insusceptible to further attacks of the same disease. Alexines are naturally produced antitoxins; antitoxins are acquired alexines. Hence we have the well-known terms "natural" and "acquired immunity." Of the former we have already spoken. Acquired immunity is a protection not belonging to the tissues of individuals naturally and as part of their constitution, but it is acquired during their lives as a further accomplishment, so to speak, of their tissues. This may happen in one or both of two ways. Either it may be an involuntary acquired immunity,250 or a voluntary acquired immunity. For example, the former is at once illustrated by an attack of the disease.
Small-pox, typhoid fever, even scarlet fever, are diseases which very rarely attack the same individual twice. That is because each of these diseases leaves behind it, on its first appearance, its antitoxic influence. Hence the individual has involuntarily acquired immunity against these diseases. An example of voluntary acquired immunity is also at hand in the old method of preventive inoculation for small-pox, or variolation. This was clearly an inoculation setting up an artificial and mild attack of small-pox, by which the antitoxins of that disease were produced, and protected the individual against further infection of small-pox; that is to say, it was a voluntary acquired immunity. This form of artificial production of protection is generally called artificial immunity. Let us now marshal together these various terms in a table as follows:
| Immunity in man = a condition of protection of insusceptibility to certain diseases. | ||
| 1. Natural immunity = constitutional protection produced by alexines. | ||
| 2. Acquired immunity | ||
| = | |
Acquired naturally (involuntarily) produced by antitoxins formed by an attack of the disease. |
| Acquired artificially (voluntary)= | ||
(a) Active immunity, produced by direct inoculation of the weakened bacteria or weakened toxins of the disease, e. g., vaccination, or Pasteur's treatment of rabies, or Haffkine's inoculation for cholera. |
||
(b) Passive immunity, produced by inoculation, not of the disease of an animal suffering from the specific disease. |
||
It is hoped that previous remarks will have explained the meaning of the terms used in the above table, with the exception of the last two phrases of active and passive immunity. We propose now to consider in some detail the four illustrations quoted under these two headings, viz.,251 vaccination, Pasteur's treatment of rabies, anti-cholera inoculation, and antitoxin inoculation. From all accounts, it is to be feared that these four phases of artificial immunity are hopelessly confused in the educated public mind. Nor is this to be wondered at when we reflect upon the rapid growth of the whole science of immunity, and upon the ever-varying forms of nomenclature through which it has passed.
Vaccination for Small-pox. In 1717 Lady Mary Wortley Montagu82 described the inoculation of small-pox as she had seen it practised in Constantinople. So greatly was she impressed with the efficacy of this process that she had her own son inoculated there, and in 1721 Mr. Maitland, a surgeon, inoculated her daughter in London. This was the first time inoculation was openly practised in England.83 For one hundred and twenty years small-pox inoculation (or variolation, as it is more correctly termed) was practised in England, until by Act of Parliament in 1840 it was prohibited.
There were different ways of performing variolation, but the most approved method was similar to the modern system of arm-to-arm vaccination, the arm being inoculated with a lancet in one or more places with small-pox lymph instead of, as now, with vaccine lymph. As a rule, only local results or a mild attack of small-pox followed, which prevented an attack of natural small-pox. Its disadvantage is apparent on the surface. It was a means of breeding small-pox, for the inoculated cases were liable to create fresh centres of infection. In 1796 Edward Jenner, who was a country practitioner in Gloucestershire, observed that those persons 252affected with cow-pox, contracted in the discharge of their duty as milkers, did not contract small-pox, even when placed in risk of infection. Hence he inferred that inoculation of this mild and non-infectious disease would be preferable to the process of variolation then so widely adopted in England. Jenner therefore suggested the substitution of cow-pox lymph (vaccine) in place of small-pox lymph, as in ordinary variolation.
It should not be forgotten that variolation was thus the first work done in this country in producing artificial immunity, and was followed by vaccination, which was only partly understood. Even to-day there is probably much to learn respecting it. Both variolation and vaccination may be described as active immunisation by means of an attenuated form of the specific virus causing the disease. The nature of the specific virus of both small-pox and cow-pox awaits discovery. Burdon Sanderson, Crookshank, Klein, and Copeman have all demonstrated bacteria in cow-pox or vaccine lymph, and in 1898 Copeman announced that he had isolated a specific bacillus and grown it upon artificial media.84 Numerous statements have been made to the effect that a specific bacillus has been found in small-pox also. But neither in small-pox nor cow-pox is the nature of the contagion really known.85
These facts, however, do not remove the suspicion which has hitherto rested upon vaccine lymph as a vehicle for bacteria of other diseases which by its inoculation may thus be contracted. A few remarks are therefore called for at this juncture upon the recent work of Dr. Monckton Copeman and Dr. Frank Blaxall in respect to what is known as glycerinated calf lymph. Evidence has been forthcoming to sub253stantiate in some measure the distrust which many of the public have from time to time felt in the vaccine commonly used in vaccination, hence the new form as above designated. This retains the toxic qualities required for immunity, but is so produced that it possesses in addition three very important advantages; namely, it is entirely free from extraneous organisms, it is available for a large number of vaccinations, and it retains full activity for eight months. It is prepared as follows:
A calf, aged three to six months, is kept in quarantine for a week. If then found upon examination to be quite healthy, it is removed to the vaccination station, and the lower part of its abdomen antiseptically cleaned. The animal is now vaccinated upon this sterilised area with glycerinated calf lymph. After five days the part is again thoroughly washed, and the contents of the vesicle, which have of course appeared in the interval, are removed with a sterilised sharp spoon, and transferred to a sterilised bottle. This is now removed to the laboratory, and the exact weight of the material ascertained. A calf thus vaccinated will yield from 18 to 24 grams of vaccine material. This is now thoroughly triturated and mixed with six times its weight of a sterilised solution of 50 per cent. chemically pure glycerine in distilled water. The resulting emulsion is aseptically stored in sealed tubes in a cool place. For four weeks it is carefully examined bacteriologically until the glycerine has absolutely killed any possible germ that may have obtained entrance. When by agar plates it is demonstrably sterile it is ready for distribution.
Pasteur's Treatment of Rabies. Rabies is a disease affecting dogs (in Western Europe) and wolves (in Russia), and can be transmitted to other animals and man, infection being carried by the bite of a rabid animal. It takes two chief forms: (1) furious rabies and (2) paralytic rabies. The former is more common in dogs. The animal becomes rest254less, has a high-toned bark, and snaps at various objects. Sometimes it exhibits depraved appetite; spasms of the throat follow, and these soon develop into convulsions, which are followed by coma and death. In man the incubation period is fortunately a very long one, averaging about forty days. Nervous irritability is the first sign; spasms occur in the respiratory and masticatory muscles, and the termination is similar to rabies in the dog. The symptom of fear of water is a herald of coming fatality.
Although a number of the workers at the Pasteur Institute and elsewhere have addressed themselves to the detection of a specific microbe, none has as yet been found, although, in the opinion of Pasteur, such an agent may be suspected as the cause.
Pathologically rabies and tetanus (see page 168) are closely allied diseases, and the recent remarkable additions to our knowledge of the latter disease only make the similarity more evident. There are in rabies three chief sets of post-mortem signs. First, and by far the most important, are the changes in the nervous system. Here we find patches of congestion in the brain, and breaking down of the axis cylinders of the nerves. The stomach, in the second place, exhibits hæmorrhagic changes, not unlike acute arsenical poisoning. Thirdly, the salivary glands show a degenerative change in a breaking down of their secreting cells. Roux has pointed out that in life the saliva of a mad dog becomes virulent three days before the appearance of the symptoms of disease.
We may now turn to the method of treatment which was introduced by Pasteur. Before his time cauterisation of the wound was the only means adopted. If more than half an hour has elapsed since the bite, cauterisation is of little or no avail. The basis for Pasteur's treatment was the difference in virulence obtainable in spinal cords infected with rabies. Pasteur found that drying the cord led to a lessen255ing of its virulence, just as certain other conditions increased its virulence. Next he established the fact that subcutaneous injection of a weak virus, followed up with doses of ever-increasingly virulent cords, immunised dogs against infection or inoculation of fully virulent material. From this he reasoned that if he could establish a standard of weakened virulence he would have at hand the necessary "vaccine" for the treatment of the disease.
Subsequent research and skilled technique resulted in a
method of securing this standard, which he found to be a
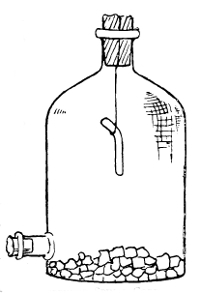 Suspended Spinal Cord
Suspended Spinal Cord
In drying jar containing Calcium Chloride
spinal cord dried for fourteen
days. The exact details are as
follows: The spinal cords of two
rabbits dead of rabies are removed
from the spinal canal in their entirety
by means of snipping the
transverse processes of the vertebrae.
Each cord is divided into
three more or less equal pieces,
and each piece, being snared by a
thread of sterilised silk, is carefully
suspended in a sterilised glass
jar. At the bottom of the jar
is a layer, about half an inch
deep, of sterilised calcium
chloride. The jars are then
removed to a dark chamber,
where they are placed at a
temperature of 20–22° C. in
wooden cases. Here they
are left to dry. Above each case is a tube of broth, to
which has been added a small piece of the corresponding
cord, in order to test for any organismal element that may
by chance be included. In case of the slightest turbidity
in the broth, the cord is rejected. Fourteen series of cords256
are thus suspended on fourteen consecutive days. The
first, second, and third are found to be of practically equal
virulence, but from the third to the fourteenth the virulence
proportionately decreases, and on the fifteenth day the cord
would be practically innocuous and non-virulent. When
treatment is to be commenced, obviously the weakest—that
is, the fourteenth day—cord is used to make the "vaccine,"
and so on in steadily increasing doses (as regards virulence)
up to and including a third-day cord. The fourteenth-day
cord is therefore taken, and a small piece cut off and macerated
in 10 cc. of sterile broth, which are placed in a conical
glass and covered with two layers of thick filter-paper, the
glass with its covering having been previously sterilised by
dry heat. When the patient bitten by the rabid animal is
prepared, 3 cc. of this broth emulsion of spinal cord are inoculated
by means of a hypodermic needle into the flanks
or abdominal wall. On the following day the patient returns
for an inoculation of a cord of the thirteenth day, and so on
until a rabid cord emulsion of the first three days has been
inoculated. As a matter of practice, the dosage depends
upon the three recognised classes of bites, viz. (1) bites
through clothing (least severe); (2) bites on the bare skin
of the hand; (3) bites upon the face or head, most severe
owing to the vascularity of these parts. An example of
each, which the writer was permitted to take in the Pasteur
Institute, may be here added to make quite clear the entire
practice. (See page 258.)
It may be well to add the returns of inoculation made at the Pasteur Institute, Rue Dutot, Paris, as above described. They are as follows:
| Year. | No. of Persons inoculated. |
No. of Deaths. |
Rate of Mortality. |
| 1886 | 2,671 | 25 | 0.94 |
| 1887 | 1,770 | 14 | 0.79 |
| 1888 | 1,622 | 9 | 0.55 |
| 2571889 | 1,830 | 7 | 0.38 |
| 1890 | 1,540 | 5 | 0.32 |
| 1891 | 1,559 | 4 | 0.25 |
| 1892 | 1,790 | 4 | 0.22 |
| 1893 | 1,648 | 6 | 0.36 |
| 1894 | 1,387 | 7 | 0.50 |
| 1895 | 1,520 | 2 | 0.13 |
| 1896 | 1,308 | 4 | 0.30 |
| 1897 | 1,521 | 6 | 0.39 |
Pasteur's treatment of rabies by inoculation of emulsions of dried spinal cord is, therefore, a "vaccination" of attenuated virus, resulting in antitoxin formation, to the further protection of the individual against rabies.
One further example of the modern application of the principles of active acquired immunity may be shortly mentioned. We refer to the cholera and plague vaccinations. The vaccination in small-pox is an inoculation of the virus of the disease; the rabies inoculation is a transmission of the vital products of the disease attenuated; the plague and cholera vaccinations are inoculations of pure cultures of living virus from outside the body. Inoculating cholera virus against cholera has been made illegal, as variolation was in 1840. But Haffkine has prepared two vaccines. The weak one is made from pure cultures of Koch's spirillum of Asiatic cholera, attenuated by growth to several generations on agar or broth at 39°C. The strong one is from similar culture the virulence of which has been increased. One cubic centimetre of the first vaccine is injected hypodermically into the flank, and the second vaccine three or four days afterwards. The immunisation is prophylactic, not remedial, and its action takes effect five or six days after the second vaccine has been injected.
INOCULATION TREATMENT FOR PERSONS AFFECTED WITH RABIES
| 1. For those Bitten through Clothes | 2. For those Bitten on Uncovered Skin of Hands, Etc. | 3. For those Bitten on Face or Head. | ||||
| Days of Treatment. | Doses of Emulsion per cc. | Dates of Cord Drying. | Doses of Emulsion per cc. | Dates of Cord Drying. | Doses of Emulsion per cc. | Dates of Cord Drying. |
| 1 at 11 A.M. | 3 | 14 | 3 | 14 | 3 | 14 |
| 1 at"11ii"A | 3 | 13 | 3 | 13 | 3 | 13 |
| 1 at 3 P.M. | .. | .. | .. | .. | 3 | 12 |
| 1 at"11ii"A | .. | .. | .. | .. | 3 | 11 |
| 2 at 11 A.M. | 3 | 12 | 3 | 12 | 3 | 10 |
| 2 at"11ii"A | 3 | 11 | 3 | 11 | 3 | 9 |
| 2 " 3 P.M. | .. | .. | .. | .. | 3 | 8 |
| 2 at"11ii"A | .. | .. | .. | .. | 3 | 7 |
| 3 at 11 A.M. | 3 | 10 | 3 | 10 | 3 | 6 |
| 3 at"11ii"A | 3 | 9 | 3 | 9 | 3 | 6 |
| 4 at 11 A.M. | 3 | 8 | 3 | 8 | 3 | 5 |
| 4 at"11ii"A | 3 | 7 | 3 | 7 | .. | .. |
| 5 at"11ii"A | 3 | 6 | 3 | 6 | 3 | 5 |
| 5 at"11ii"A | 3 | 6 | 3 | 6 | .. | .. |
| 6 at"11ii"A | 3 | 5 | 3 | 5 | 3 | 4 |
| 7 at"11ii"A | 3 | 5 | 3 | 5 | 2 | 3 |
| 8 at"11ii"A | 3 | 4 | 3 | 4 | 3 | 4 |
| 9 at"11ii"A | 2 | 3 | 2 | 3 | 3 | 3 |
| 10 at"11ii"A | 3 | 5 | 3 | 5 | 3 | 5 |
| 11 at"11ii"A | 3 | 5 | 3 | 5 | 3 | 5 |
| 12 at"11ii"A | 3 | 4 | 3 | 4 | 3 | 4 |
| 13 at"11ii"A | 3 | 4 | 3 | 4 | 3 | 4 |
| 14 at"11ii"A | 3 | 3 | 3 | 3 | 3 | 3 |
| 15 at"11ii"A | 3 | 3 | 3 | 3 | 3 | 3 |
| 16 at"11ii"A | ... | .. | 3 | 5 | 3 | 5 |
| 17 at"11ii"A | ... | .. | 3 | 4 | 3 | 4 |
| 18 at"11ii"A | ... | .. | 3 | 3 | 3 | 3 |
| 19 at"11ii"A | .. | .. | .. | .. | 3 | 5 |
| 20 at"11ii"A | .. | .. | .. | .. | 3 | 4 |
| 21 at"11ii"A | .. | .. | .. | .. | 3 | 3 |
In plague the same plan has been followed. Luxurious
crops of Kitasato's plague bacillus are grown on ordinary
nutritive media plus large quantities of fat. The fat of
milk, as clarified butter, is that generally used. Under the
globules of fat flakes of culture grow like stalactites, hanging
down into the clear broth. These are in time shaken to the
bottom, and a second crop grows on the under-surface of the
fat. In the course of a month perhaps half a dozen such
crops are obtained and shaken down into the fluid, until the
latter assumes an opaque milky appearance. This is now,
unlike the cholera vaccine, exposed to a temperature of 70°
C., by which the microbes are killed. The culture contains
all the toxins, and the dose is 3 cc. This preparation has
the advantage of being easily prepared, obtainable in large
quantities, and requires no animals in its preparation.
When inoculated it produces local pain and swelling at the
site of inoculation, and general reactive symptoms such as
fever. From a careful analysis of the results of this inoculation,
it is shown that the efficacy of the prophylactic depends
upon the virulence of the bacillus culture from which
the vaccine is prepared, and upon its dose and ability to
produce a well-marked febrile reaction. It appears to be
more effective in the prevention of deaths than of attacks.
The anti-typhoid vaccination is another example of inoculation to secure active immunity. It is needless, perhaps, to point out that all these vaccinations, except rabies, are prophylactic, and not curative.
Passive Immunity; Preparation of Antitoxins. We must now consider the question of passive immunity. This, it will be remembered, may be defined as a protection (against a bacterial disease) produced by inoculation, not of the disease itself, as in small-pox inoculation, nor yet of its weakened toxins, as in rabies, but of the antitoxins produced in the body of an animal suffering from that particular disease. Examples of this treatment are increasing every year, and the term "antitoxin" has now become almost a household260 word. The chief examples are to be found in diphtheria, tetanus, streptococcus, and pneumococcus.
To be of value, antitoxins must be used as early as possible, before tissue change has occurred and before the toxins have, so to speak, got the upper hand. When the toxins are in the ascendency the patient suffers more and more acutely, and may succumb before there has been time for the formation in his own body of the antitoxins. If he can be tided over the "crisis," theoretically all will be well, because then his own antitoxin will eventually gain the upper hand. But in the meantime, before that condition of affairs, the only way is to inject antitoxins prepared in some animal's tissues whose disease began at an earlier date, and thus add antitoxins to the blood of our patient, early in the disease, and the earlier the better, for, however soon this is done, it is obvious that the toxins begin their work earlier still. It should not be necessary to add that general treatment must also be continued, and indeed local germicidal treatment, e. g., of the throat in diphtheria and the poisoned wound in tetanus. Further, in a mixed infection, as in glandular abscesses with diphtheria, it must be borne in mind that the antitoxin is specific and may therefore probably fail in such mixed cases.
After these preliminary remarks we will now consider shortly some of the methods employed for the production of antitoxins. An animal is required from whose body a considerable quantity of blood can be drawn without injurious effect. Moreover, it must be an animal that can stand an attack of such diseases as diphtheria and tetanus. Such an animal is the horse. Now, by injecting into the horse (a) living organisms of the specific disease, but in non-fatal doses, or (b) dead cultures, or (c) filtered cultures containing no bacteria and only the toxins, we are able to produce in the blood of the horse first the toxins and then the antitoxins of the disease in question. The non-poisonous doses of261 living organisms can be weakened, or, as we say, attenuated, by various means. Dead cultures have not been much used to produce immunity except by Pfeiffer. In actual practice the third method is much the most general, viz., filtering a fluid culture free from the bacteria, and then inoculating this in ever-increasing doses. The preparation of diphtheria antitoxin may be taken as an example, but what follows would be equally applicable to other diseases, such as tetanus.
1. To Obtain the Toxin. First grow a pure culture of the
Klebs-Löffler bacillus of diphtheria in large flasks containing
"Löffler's medium," or a solution made by mixing
three parts of blood serum with one of beef broth and adding
one per cent. of common salt (Na Cl) and one per cent.
of peptone. An alkaline medium is preferable. The flask
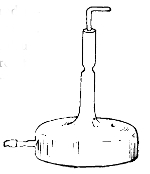 Flask used for the Preparation of
the Toxin of Diphtheria
was thoroughly sterilised before use, and is
now plugged with sterile cotton-wool and incubated
at 77° C. for three or four weeks.
Pure air may be passed over the culture
periodically, thereby aiding the growth.
After the lapse of about a month a scum of
diphtheria growth will have appeared
over the surface of the fluid. This is
now filtered into sterilised flasks, and
some favourable antiseptic added to
ensure that nothing foreign to the
toxin shall flourish, and the flasks are
kept in the dark. Here, then, we
have the product, the toxin, ready for injection into the horse.
Flask used for the Preparation of
the Toxin of Diphtheria
was thoroughly sterilised before use, and is
now plugged with sterile cotton-wool and incubated
at 77° C. for three or four weeks.
Pure air may be passed over the culture
periodically, thereby aiding the growth.
After the lapse of about a month a scum of
diphtheria growth will have appeared
over the surface of the fluid. This is
now filtered into sterilised flasks, and
some favourable antiseptic added to
ensure that nothing foreign to the
toxin shall flourish, and the flasks are
kept in the dark. Here, then, we
have the product, the toxin, ready for injection into the horse.
2. Immunisation of the Horse. It is evident that only healthy horses are of service in providing healthy antitoxin, even as healthy children are necessary in arm-to-arm vaccination. To provide against any serious taint the horse is tested for glanders (with mallein) and for tuberculosis (with tuberculin). The dose of the injection of toxin is at the commencement about 1/10 cc., or a little more. The site of262 the inoculation is the apex of the shoulder, which has been antiseptically cleaned. A mere prick is the whole operation. After the first injection there is generally a definite febrile reaction and a slight local swelling. From 1/10 or 1/2 cc. the dose is steadily increased, until at the end of two or three months86 perhaps as much as 300 cc. (or even half a litre) may be injected without causing the reaction which the initial injection of 1/10 cc. caused at the outset. This shows an acquired tolerance of the tissues of the horse to the toxic material. After injecting 500 cc. into the horse without bad effect, the animal has a rest of four or five days.
3. To Obtain the Antitoxin. During this period of rest the interaction between the living body cells of the animal and the toxins results in the production in the blood of an antitoxin. By means of a small sterilised cannula, five, or eight, or even ten litres of blood are drawn from the jugular vein of the horse into sterilised flasks or jars. The top of the jar is closed by two paper coverings before it is sterilised. Then it is again covered with a further loose one. Before use the loose one is removed and replaced by a metal (zinc) lid, which has been separately sterilised. This metal lid contains an aperture large enough for the tube which conveys the blood from the cannula to pass through. The tube, therefore, passes through the metal lid and two paper covers, which it was made to pierce. When enough blood has passed into the vessel the tube is withdrawn, and the metal lid slightly turned. Thus the contained blood is protected from the air.87
The jar containing the blood (which contains the antitoxin) is next placed in a dark, cool cellar, where it stands for two or three days. During this time the blood naturally coagulates, the corpuscles falling as a dense clot to the bottom, and the faintly yellow serum rising to the top. The serum, or liquor sanguinis, averages about 50 per cent. of the total blood taken. Sometimes antiseptics are added with a view to preservation. It is generally filtered before bottling for therapeutic use, and sometimes examined bacteriologically as a test of purity.
4. The Use of Antitoxins. The antitoxins are now ready for injection into the patient who has contracted diphtheria, and in whose blood toxins are in the ascendency and under which the individual may succumb. They are injected in varying doses, as we have already pointed out.88 The general result is that mortality has been greatly lessened, and that in fatal cases there has been a considerable lengthening of the period of life. Moreover, the whole clinical course of the disease has been greatly modified, and suffering lessened.89
Probably the most universally known fact respecting bacteria is that they are related in some way to the production of disease. Yet we have seen that it was not as disease-producing agents that they were first studied. Indeed, it is only within comparatively the latest period of the two centuries during which they have been more or less under observation that our knowledge of them as causes of disease has assumed any exactitude or general recognition. Nor is this surprising, for although an intimate relationship between fermentation and disease had been hinted at in the middle of the seventeenth century, it was not till the time of Pasteur that the bacterial cause of fermentation was experimentally and finally established.
In the middle of the seventeenth century men learned, through the eyes of Leeuwenhoek, that drops of water contained "moving animalcules." A hundred years later Spallanzani demonstrated the fact that putrefaction and fermentation were set up in boiled vegetable infusions when outside air was admitted, but when it was withheld from these boiled infusions no such change occurred. Almost a hundred years more passed before the epoch-making work of Tyndall and Pasteur, who separated these putrefactive germs from the air. Quickly following in their footsteps came Davaine and Pollender, who found in the blood of animals suffering from anthrax the now well-known specific265 and causal bacillus of that disease. Improvements in the microscope and in methods of cultivation (Koch's plate method in particular) soon brought an army of zealous investigators into the field, and during the last twenty years first this disease and then that have been traced to a bacterial origin. We may summarise the vast mass of historical, physiological, and pathological research extending from 1650 to 1898 in three great periods: the period of detection of living, moving cells (Leeuwenhoek and others in the seventeenth century); the period of the discovery of their close relationship to fermentation and putrefaction (Spallanzani, Schulze, Schwann, in the eighteenth century); and, thirdly, the period of appreciation of the rôle of bacteria in the economy of nature and in the production of disease (Tyndall, Pasteur, Lister, Koch, in the nineteenth).
But we must look less cursorily at the growth of the idea of bacteria causing disease. More than two hundred years ago Robert Boyle (1627–91), the philosopher, who did so much towards the foundation of the present Royal Society, wrote a learned treatise on The Pathological Part of Physic. He was one of the earliest scientists to declare that a relationship existed between fermentation and disease. When more accurate knowledge was attained respecting fermentation, great advance was consequently made in the etiology of disease. The preliminary discoveries of Fuchs and others between 1840 and 1850 had relation to the existence in diseased tissues of a large number of bacteria. But this was no proof that such germs caused such diseases. It was not till Davaine had inoculated healthy animals with bacilli from the blood of an anthrax carcass, and had thus produced the disease, that reliance could be placed upon that bacillus as the vera causa of anthrax. Too much emphasis cannot be laid upon this idea, that unless a certain organism produces in healthy tissues the disease in question, it cannot be considered as proven that the particular organism is related to266 the disease as cause to effect. In order to secure a standard by which all investigators should test their results, Koch introduced four postulates. Until each of the four has been fulfilled, the final conclusion respecting the causal agent must be considered sub judice. The postulates are as follows:
(a) The organism must be demonstrated in the circulation or tissues of the diseased animal.
(b) The organism thus demonstrated must be cultivated in artificial media outside the body, and successive generations of a pure culture of that organism must be obtained.
(c) Such pure cultures must, when introduced into a healthy and susceptible animal, produce the specific disease.
(d) The organism must be found and isolated from the circulation or tissues of the inoculated animal.
It is evident that there are some diseases—for example, cholera, leprosy, and typhoid—which are not communicable to lower animals, and therefore their virus cannot be made to fulfil postulate (c). In such cases there is no choice. They cannot be classified along with tubercle and anthrax. Bacteriologists have little doubt that Hansen's bacillus of leprosy is the cause of that disease, yet it has not fulfilled postulates (b) and (c). Nor has the generally accepted bacillus of typhoid fulfilled postulate (c), yet by the majority it is provisionally accepted as the agent in producing typhoid. Hence it will be seen that, though there is an academical classification of causal pathogenic bacteria according as they respond to Koch's postulates, yet nevertheless, there are a number of pathogenic bacteria which are looked upon as causes of disease provisionally. Anthrax and tubercle, with perhaps the organisms of suppuration, tetanus, plague, and actinomycosis, stand in the first order of pathogenic germs. Then comes a group awaiting further confirmation. It includes the organisms related to typhoid, cholera, malaria, leprosy, diarrhœa,and pneumonia.267 Then comes in a third category, a long list of diseases, such as scarlet fever, small-pox, rabies, and others too numerous to mention, in which the nature of the causal agent is still unknown. Hence it must not be supposed that every disease has its germ, and without a germ there is no disease. Such universal assertions, though not uncommonly heard, are devoid of accuracy.
In the production of bacterial disease there are two factors. First, there is the body tissue of the individual; secondly, there is the specific organism.
Whatever may be said hereinafter with regard to the power of micro-organisms to cause disease, we must understand one cardinal point, namely, that bacteria are never more than causes, for the nature of disease depends upon the behaviour of the organs or tissues with which the bacteria or their products meet (Virchow). Fortunately for a clear conception of what "organs and tissues" mean, these have been reduced to a common denominator, the cell. Every living organism, of whatever size or kind, and every organ and tissue in that living organism, contains and consists of cells. Further, these cells are composed of organic chemical substances which are not themselves alive, but the mechanical arrangement of which determines the direction and power of their organic activity and of their resistance to the specific agents of disease. With these facts clearly before us, we may hope to gain some insight into the reasons for departure from health.
The normal living tissues have an inimical effect upon bacteria. Saprophytic bacteria of various kinds are normally present on exposed surfaces of skin or mucous membrane. Tissues also which are dead or depressed in vitality from injury or previous disease, but which are still in contact with the tissues, afford an excellent nidus for the growth of bacteria. Still these have not the power, unless specific, to thrive in the normal living tissue. It has been definitely268 shown that the blood fluids of the body have in their fresh state the germicidal power (alexines) which prevents bacteria from flourishing in them. Such action does undoubtedly depend in measure upon the number of germs as well as their quality, for the killing power of blood and lymph must be limited. Buchner has pointed out that the antagonistic action of these fluids depends in part possibly upon phagocytosis, but largely upon a chemical condition of the serum. The blood, then, is no friend to intruding bacteria. Its efforts are to a certain extent seconded by the lymphoid tissue throughout the body. Rings of lymphoid tissue surround the oral openings of the trachea (windpipe) and œsophagus (gullet); the tonsils are masses of lymphoid tissue. Composed as it is of cells having a germicidal influence when in health, the lymphoid tissue may afford formidable obstruction to intruding germs.
All the foregoing points in one direction, namely, that if the tissues are maintained in sound health, they form a very resistant barrier against bacteria. But we know from experience that a full measure of health is not often the happy condition of human tissues; we have, in short, a variety of circumstances which, as we say, predispose the individual to disease. One of the commonest forms of predisposition is that due to heredity. Probably it is true that what are known as hereditary diseases are due far more to a hereditary predisposition than to any transmission of the virus itself in any form. Antecedent disease predisposes the tissues to form a nidus for bacteria; conditions of environment or personal habits frequently act in the same way. Damp soils must be held responsible for many disasters to health, not directly, but indirectly, by predisposition; dusty trades and injurious occupations have a similar effect. Any one of these three different influences may in a variety of ways affect the tissues and increase their susceptibility to disease. Not infrequently we may get them combined. For example, the269 following is not an unlikely series of events terminating in consumption (tuberculosis of the lungs):—(a) The individual is predisposed by inheritance to tuberculosis; (b) an ordinary chronic catarrh, which lowers the resisting power of the lungs, may be contracted; (c) the epithelial collections in the air vesicles of the lung—i. e., dead matter attached to the body—afford an excellent nidus for bacteria; (d) owing to occupation, or personal habits, or surroundings, the patient comes within a range of tubercular infection, and the specific bacilli of tubercle gain access to the lungs. The result, it is needless to state, will be a case of consumption more or less acute according to environment and treatment.
The channels of infection by which organisms gain the vantage-ground afforded by the depressed tissues are various, and next to the maintenance of resistant tissues they call for most attention from the physician and surgeon. It is in this field of preventive medicine—that is to say, preventing infective matter from ever entering the tissues at all—that science has triumphed in recent years. It is, in short, applied bacteriology, and therefore claims consideration in this place.
1. Pure Heredity. By this term may be understood the actual transmission from the mother to the unborn child of the specific virus of the disease. That such a conveyance may occur is generally admitted by pathologists, but it is impossible to enter fully into the matter in such a book as the present. Summarily we may say that, though this sort of transmission is possible, it is not frequent, nor is disease appreciably spread through such a channel. Sixty per cent. of consumptives, it has been estimated, have tuberculous progenitors, and this is the highest figure. Many would be justified from experience in placing it at half that number.
2. Inoculation, or inserting virus through a broken surface of skin, is itself a sufficiently obvious mode of infection to call for little comment. Yet it is under this heading that270 a word must be said of that remarkable application of preventive medicine known as the antiseptic treatment of wounds. When Lord Lister was Professor of Surgery in Glasgow, he was impressed with the greatness of the evil of putrefaction in wounds, which was caused, not by the oxygen of the air, as Liebig had declared, but by the entrance into the wound of fermentative organisms from the air. This was demonstrated by Pasteur, who pointed out that they could not arise de novo in the wound. Hence it appeared to Lister that these fermentative bacteria which produce putrefaction in wounds must either be kept out of the wound altogether, or killed, or their action prevented, in the wound. To keep air away from wounds is an almost impossible task, and thus it came about that wounds were dressed with a solution of carbolic acid.
From time to time examples occur of bacterial disease being directly inoculated in wounds made with polluted instruments, or in cuts made by contaminated broken glass, or in gunshot wounds. Tetanus is, of course, one of the most marked examples.
3. Contagion is a term which has suffered from the many ways in which it has been used. Defined shortly and most simply, we should say a disease is contagious when it can be "caught" by contact, through the unbroken surfaces, between diseased and healthy persons. Ringworm is an example, and there are many others.
4. The Alimentary Canal: Food. The recent Royal Commission on Tuberculosis has collected a large mass of evidence in support of the view that tubercle may be spread by articles of food. Milk and meat from tuberculous animals naturally come in for the largest amount of condemnation. To these matters we refer elsewhere.
5. The Respiratory Tract: Air. The air may become infected with germs of disease from dusty trades, dried sputum, etc. If such infected air be inhaled, pathogenic271 results will follow, especially if the bacteria are present in sufficient numbers, or meet with devitalised, and therefore non-resisting, tissues.
These, then, are the five possible ways in which germs gain access to the body tissues. The question now arises, How do bacteria, having obtained entrance, set up the process of disease? For a long time pathologists looked upon the action of these microscopic parasites in the body as similar to, if not identical with, the larger parasites sometimes infesting the human body. Their work was viewed as a devouring of the tissues of the body. Now, it is well known that, however much or little of this may be done, the specific action of pathogenic bacteria is of a different nature. It is twofold. We have the action of the bacteria themselves, and also of their products or toxins. In particular diseases, now one and now the other property comes to the front. In bacterial diseases affecting or being transmitted mostly by the blood, it is the toxins which act chiefly. The convenient term infection is applied to those conditions in which there has been a multiplication of living organisms after they have entered the body, the word intoxication indicating a condition of poisoning brought about by their products. It will be apparent at once that we may have both these conditions present, the former before the latter, and the latter following as a direct effect of the former. Until intoxication occurs there may be few or no symptoms, but directly enough bacteria are present to produce in the body certain poisons in sufficient amount to result in more or less marked tissue change, then the symptoms of that tissue change appear. This period of latency between infection and the appearance of the disease is known as the incubation period. Take typhoid, for example. A man drinks a typhoid-polluted water. For about fourteen days the bacilli are making headway in his body without his being aware of it. But at the end of that incubation272 period the signs of the disease assert themselves. Professor Watson Cheyne and others have maintained that there is some exact proportion between the number of bacteria gaining entrance and the length of the incubation period.
Speaking generally, we may note that pathogenic bacteria divide themselves into two groups: those which, on entering the body, pass at once, by the lymph or blood stream, to all parts of the body, and become more and more diffused throughout the blood and tissues, although in some cases they settle down in some spot remote from the point of entrance, and produce their chief lesions there. Tubercle and anthrax would be types of this group. On the other hand, there is a second group, which remain almost absolutely local, producing only little reaction around them, never passing through the body generally, and yet influencing the whole body eventually by means of their ferments or toxins. Of such the best representatives are tetanus and diphtheria. The local site of the bacteria is, in this case, the local manufactory of the disease.
Whilst the mere bodily presence of bacteria may have mechanical influence injurious to the tissues (as in the small peripheral capillaries in anthrax), or may in some way act as a foreign body and be a focus of inflammation (as in tubercle), the real disease-producing action of pathogenic bacteria depends upon the chemical poisons (toxins) formed directly or indirectly by them. Though within recent years a great deal of knowledge has been acquired about the formation of these bodies, their exact nature is not known. They are allied to albuminous bodies and proteoses, and are frequently described as tox-albumens. It may be found, after all, that they are not of a proteid nature. Sidney Martin has pointed out that there is much that is analogous between the production of toxins and the production of the bodies of digestion. Just as ferments are necessary in the intestine to bring about a change in the food by which273 the non-soluble albumens shall be made into soluble peptones and thus become absorbed through the intestinal wall, so also a ferment may be necessary to the production of toxins. Such ferments have not as yet been isolated, but their existence in diphtheria and tetanus is, as we have seen, extremely likely. However that may be, it is now more or less established that there are two kinds of toxic bodies, differing from each other in their resistance to heat. It may be that the one most easily destroyed by heat is a ferment and possibly an originator of the other. A second division which has been suggested for toxic bodies, and to which reference has been made, is intracellular and extracellular, according to whether or not the poison exists within or without the body of the bacillus.
Lastly, we may turn to consider the action of the toxins on the individual in whose body-fluids they are formed. It is hardly necessary to say that any action which bacteria or toxins may have will depend upon their virulence, in some measure upon their number, and not a little upon the channel of infection by which they have gained entrance. It could not be otherwise. If the virulence is attenuated, or if the invasion is very limited in numbers, it stands to reason that the pathogenic effects will be correspondingly small or absent. The influence of the toxins is twofold. In the first place (i.) they act locally upon the tissues at the site of their formation, or at distant points by absorption. There is inflammation with marked cell-proliferation, and this is, more or less rapidly, followed by a specific cell-poisoning. The former change may be accompanied by exudation, and simulate the early stages of abscess formation; the latter is the specific effect, and results, as in leprosy and tubercle, in infective nodules. The site in some diseases, like typhoid (intestinal ulceration, splenic and mesenteric change) or diphtheria (membrane in the throat), may be definite and always the same. But, on the other hand, the site may274 depend upon the point of entrance, as in tetanus. The distant effects of the toxin are due to absorption, but what controls its action it is impossible to say. We only know that we do find pathological conditions in certain organs at a distance and without the presence of bacteria. We have a parallel in the action of drugs; for example, a drug may be given by the mouth and yet produce a rash in some distant part of the body. In the second place (ii.) toxins produce toxic symptoms. Fever and many of the nervous conditions resulting from bacterial action must thus be classified. We have, it is true, the chemical symptoms of the pathological tissue change, for example, the large spleen of anthrax or the obstruction from diphtheritic membrane. But, in addition to these, we have general symptoms, as fever, in which after death no tissue change can be formed.
We may now consider briefly some of the more important types of disease produced by bacteria:
1. Tuberculosis.90 As far back as 1794 Baillie drew attention to the grey miliary nodules occurring in tuberculous tissue which gave rise to the term "tubercles." This preliminary matter was confirmed by Bayle in 1810.
In 1834 Laennec described all caseous deposits as "tubercles," insisting upon four varieties:
(1) Miliary, which were about the size of millet seeds, and in groups;
(2) Crude, miliary tubercles in yellow masses;
(3) Granular, similar to the last, but scattered;
(4) Encysted, a hard mass of crude tubercle with a fibrous or semi-cartilaginous capsule.
The tubercle possesses in many cases a special structure, and certain cell-forms frequently occur in it and give it a characteristic appearance. The central part of the tubercle 275usually contains giant cells with numerous nuclei. The uninuclear cells are partly lymphoid, partly large epithelial or endothelial cells; these are called epithelioid cells.
It was not till 1865 that the specific nature of tuberculosis was asserted by Villemin. Burdon Sanderson (1868–69) in England confirmed his work, and it was extended by Connheim, who a few years later laid down the principle that all is tubercular which by transference to properly constituted animals is capable of inducing tuberculosis, and nothing is tubercular unless it has this capability.
Klebs (1877) and Max Schiller (1880) described masses of living cells or micrococci in many tuberculous nodules in the diseased synovial membrane and in lupus skin. In 1881 Toussaint declared he had cultivated from the blood of tubercular animals and from tubercles an organism which was evidently a micrococcus, and in the same year Aufrecht stated that the centre of a tubercle contained small micrococci, diplococci, and some rods. But it was not till the following year, 1882, that Koch discovered and demonstrated beyond question the specific Bacillus tuberculosis.
It is now held to be absolutely proved that the introduction of the bacillus, or its spores or products, is the one and only essential agent in the production of tuberculosis. Its recognised manifestations are as follows:
| Tuberculosis | in the lungs = acute or chronic phthisis; |
| " | in the skin = lupus91; |
| " | in the mesenteric glands = Tabes mesenterica; |
| " | in the brain = hydrocephalus; |
| " | in lymphatic glands = Scrofula.91 |
The disease may occur generally throughout the body or locally in the suprarenal capsules, prostate, intestine, larynx, membranes of the heart, bones, ovaries, pleura, kidneys, spleen, testicles, Fallopian tubes, uterus, etc.
We may summarise the history of the pathology of tubercle thus:
1794. Baillie drew attention to grey miliary nodules occurring in tuberculosis, and called them "tubercles."
1834. Laennec described four varieties: miliary; crude; granular; encysted.
1843. Klencke produced tuberculosis by intravenous injection of tubercular giant cells.
1865. Villemin demonstrated infectivity of tubercular matter by inoculation of discharges; Connheim, Armanni, Burdon Sanderson, Wilson Fox, and others showed that nothing but tubercular matter could produce tuberculosis.
1877. Living cells were found in tubercles, "micrococci" (Klebs, Toussaint, Schiller).
1882. Koch isolated and described the specific bacillus, and obtained pure cultivations (1884).
The Bacillus of Koch, 1882. Delicate cylindrical rods, measuring 1.5–4 micromillimetres in length and about .2 µ in breadth; non-motile. Many are straight with rounded ends; others are slightly curved. They are usually solitary, but may occur in pairs, lying side by side or in small masses. They are chiefly found in fresh tubercles, more sparingly in older ones. Some lie within the giant cells; others lie outside; shorter in tissue sections of bovine tuberculosis, but longer in the milk (Crookshank).
When stained they appear to be composed of irregular cubical or spherical granules within a faintly stained sheath. In recent lesions the protoplasm appears more homogeneous, and takes on the segmented or beaded character only in old lesions, pus, or sputum.
Morphological differences are found under different circumstances, and within limits variation occurs according to the environment.
Cultivation on Various Media. Koch inoculated solid blood serum with tubercular matter from an infected lymphatic gland of a guinea-pig, and noticed the first signs of growth in ten or twelve days in the form of whitish, scaly patches. These enlarged and coalesced with neighbouring patches, forming white, roughened, irregular masses. Nocard and Roux showed that by adding 5/8 per cent. of glycerine to the media commonly used in the laboratory, such as nutrient agar or broth, the best growth is obtained.
On glycerine broth or glycerine agar abundant growth appears at the end of seven or eight days. By continuous sub-culture on glycerine agar the virulence of the bacillus is diminished. But in fifteen days after inoculation of the medium the culture equals in extent a culture of several weeks' age on blood serum.
Sub-cultures from glycerined media will grow in ordinary broth without glycerine (Nocard, Roux, Crookshank).
In alkaline broth to which a piece of boiled white of egg was added Klein obtained copious growths, and found that continued sub-culturing upon this medium also lessens the virulence.
Description of Cultivations:—On glycerine agar minute white colonies appear in about six days, raised and isolated, and coalescing as time advances, forming a white lichenous growth, fully developed in about two months.
On glycerine broth a copious film appears on the surface of the liquid, which if disturbed falls to the bottom of the flask as a deposit.
Spore Formation. In very old cultivations spore-like bodies can be observed both in stained and unstained preparations, but neither the irregular granules within the capsule nor the unstained spaces between the granules are278 spores (Babes and Crookshank). That the bacilli possess spores is believed on account of the following facts:
1. That tubercular sputum, when thoroughly dried, maintains its virulent character (Koch, Schill, Fischer, etc.). No sporeless bacillus is known which can survive through drying.
2. That tubercular matter and cultures survive temperature up to 100° C. Non-spore-bearing bacilli and micrococci are killed by being exposed for five minutes to a temperature of 65–70° C., whereas spores of other bacilli withstand much higher temperatures.
3. Tubercular sputum distributed in salt solution does not lose its virulence by being kept at 100° C. for one or two minutes; sporeless bacilli certainly would (Klein).
4. A solution of per-chloride of mercury does not kill the tubercle bacilli, as it does sporeless bacilli (Lingard and Klein).
Koch and many bacteriologists have declared the bacillus to be a "true parasite." Koch based this view upon the belief which he entertained that the bacillus can grow only between 30° C. and 41° C., and therefore in temperate zones is limited to the animal body and can originate only in an animal organism. "They are," he said, "true parasites, which cannot live without their hosts. They pass through the whole cycle of their existence in the body." But at length Koch and others overcame the difficulties and grew the bacillus as a saprophyte.
Schottelius92 has observed that tubercle bacilli taken from the lung of phthisical persons buried for years still retains its virulence and capability of producing tuberculosis upon inoculation. He further shows that tubercular lung kept in soil (enclosed in a box) shows a marked rise in temperature. Klein quotes these experiments as indications that "tubercle bacilli are not true parasites, but belong to the ecto279genic microbes which can live and thrive independent of a living host."
It has now been abundantly proved that the bacillus of tuberculosis is capable of accommodating itself to circumstances much less favourable than had been supposed, especially as regards temperature.
Temperature of Growth of Bacillus. 30–41° C. have been laid down by Koch as the limits of temperature at which the bacillus will grow in culture medium outside the body. The generally accepted temperatures as most favourable to the growth of the bacillus are between 36° C. and 38° C.
Sir Hugh Beevor, however, was able to grow the bacillus upon glycerine agar at 28° C. (82° F.), obtaining an ample culture which developed somewhat more slowly than on blood serum, and to a less extent than at 37° C. In both Beevor succeeded in growing the bacillus at a lower temperature even than on agar, viz., at a temperature rarely above 60° F. Sheridan Delépine and others have also been successful in obtaining growths at room temperature both in summer and winter.
Although, speaking generally, there is an actual cessation of growth at low temperature, the bacillus may be exposed to very low temperatures for a considerable time without losing its power of again becoming active when returned to a favourable environment (Woodhead).
The Relation of the Bacillus to the Disease. All four of Koch's postulates have been fulfilled in the case of Bacillus tuberculosis. Hence we are dealing with the specific cause of the disease. Yet, whilst this is so, we may usefully ask ourselves: How does the bacillus set up the changes in normal tissues which result in tubercular nodules? In arriving at a solution of this problem we are materially aided if we bear in mind the fact that such an organism in healthy tissues has a double effect. First, there is an ordinary inflammatory irritation, and secondly, there is a specific change280 set up by the toxins of the bacillus. Directly the invading bacilli find themselves in a favourable nidus they commence multiplication. In three or four days this acts as an irritant upon the surrounding connective-tissue cells, which proliferate, and become changed into large cells known as epithelioid cells. At the periphery of this collection of epithelioid cells we have a congested area. This change has been accomplished by the presence of the bacilli themselves. The production of their specific poisons changes the epithelioid cells in the centre of the nodule, some of which become fused together, whilst others expand and undergo division of nucleus. By this means we obtain a series of large multi-nucleated cells named giant cells. If the disease is very active, these soon caseate and break down in the centre. In a limb we get a discharge; in a lung we get an expectoration. Both discharge and expectoration arise from a breaking down of the new cell formation. Previously to breaking down we have in a fully developed nodule healthy tissue, inflammatory zone, epithelioid cells, giant cells, containing nuclei and bacilli. The sputum or the discharge will, during the acute stage of the disease at all events, contain countless numbers of the bacilli, which may thus be readily detected, and their presence used as evidence of the disease. It is obvious that if the centre of the nodule degenerates and comes away as discharge a cavity will be left behind. By degrees this small cavity may become a very large one, as is frequently the case in the lung, which particularly lends itself to such a condition. Hence, though at the outset a tubercular lung is solid, at the end it is hollow.
 |
 |
Bacillus Tuberculosis |
Bacillus Tuberculosis |
 |
 |
Streptococcus Pyogenes |
Bacillus Anthracis |
The exact period of giant-cell formation depends on the rapidity of the formative processes. Thus different conditions occur. Inside the giant cells the bacilli are arranged in relation to the nuclei in one of three ways: (a) polar, (b) zonal, or (c) mixed. The breaking down of the nodule is partly due to the cell-poisons, and partly because the nodule is non-vascular, owing to the fact that new capillaries cannot grow into the dense nodule, and the old ones are all occluded by the growth of the nodule.
From the local foci of disease the tubercle process spreads chiefly by three channels:
(a) By the lymphatics, affecting particularly the glands. Thus we get tuberculosis set up in the bronchial, tracheal, mediastinal, and mesenteric glands, and it is so frequently present as to be a characteristic of the disease. This is the common method of dissemination in the body.
(b) By the blood-vessels, by means of which bacilli may be carried to distant organs.
(c) By continuity of tissues, infective giant-cell systems encroaching upon neighbouring tissues, or discharge from lungs or bronchial glands obtaining entrance to the gullet and thus setting up intestinal disease also.
It has been abundantly proved that the respiratory and digestive systems are principally affected by Koch's bacillus. Wherever the bacilli are arrested, they excite formation of granulations or miliary tubercular nodules, which increase and eventually coalesce. The lymphatic glands which collect the lymph from the affected region are the earliest affected, always the nearest first, and then the disease appears to be appreciably stopped on its invading march. Each lymphatic gland acts as a temporary barrier to progress until the disease has broken its structure down. It remains local, in spite of increase in number and importance of the foci of disease, as long as the bacilli have not gained access to the blood stream.
Toxins and Tuberculin. Koch, Crookshank, and Herroun, Hunter, and others have isolated products from pure cultures of the tubercle bacillus. These have comprised chiefly albumoses, alkaloids, and various extractives. Koch's observations led him to suppose that in pure cultures of tubercle a substance appeared having healing action on tuberculosis,282 and an extract of this in glycerine he termed "tuberculin." It was made as follows: A veal broth containing peptone and glycerine was inoculated with a pure culture of the bacillus and incubated at 38° C. for six or eight weeks. An abundant growth with copious film formation appeared. The culture was then concentrated by evaporation over a water-bath until reduced to about one-tenth of its volume.
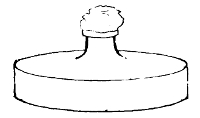
Flask used in the Preparation of Tuberculin
The announcements in 1890 and 1891 to the effect that a "cure" had been discovered for consumption will be remembered. The hopes thus raised were unfortunately not to be realised. Koch advocated injections of this tuberculin in cases of skin tubercle (lupus) and consumptive cases. In many of these benefit was apparently derived, but its general application was not founded upon any substantial basis. Dead tissue, full of bacilli, could not thus be got rid of; nor could the career of the isolated bacilli distributed through the body be thus checked.
Tuberculin has, however, found a remarkable sphere of usefulness in causing reaction in animals suffering from tuberculosis. Indeed, tuberculin is the most valuable means of diagnosis that we possess (MacFadyen). When injected (dose, 30–40 centigrammes) it causes a rise of one and a half to three degrees. The fever begins between the twelfth and fifteenth hour after injection, and lasts several hours. The duration and intensity of the reaction have no relation to the number and gravity of the lesions, but the same dose injected into healthy cattle causes no appreciable febrile reaction. The tuberculous calf reacts just as well as the283 adult, but the dose is generally 10–20 centigrammes. Injections of tuberculin have no troublesome effect on the quantity or quality of the milk of cows or on the progress of gestation.
Tuberculosis of Animals. Cattle come first amongst animals liable to tubercle. Horses may be infected, but it is comparatively rare, and among small ruminants the disease is rarer still. Dogs, cats, and kittens may be easily infected. Amongst birds, fowls, pigeons, turkeys, and pheasants, the disease assumes almost an epidemic character. Especially do animals in confinement die of tubercle, as is illustrated in zoölogical gardens. Respecting the lesions of bovine tuberculosis, it will be sufficient to say that nothing is more variable than the localisation or form of its attacks. The lungs and lymphatic glands come first in order of frequency, next the serous membranes, then the liver and intestines, and lastly the spleen, joints, and udder (Nocard).
The anatomical changes in bovine tubercle are mostly found in the lungs and their membranes, the pleuræ. It also affects the internal membrane lining, the abdomen and its chief organs, the peritoneum, and the lymphatic glands. In both these localities a characteristic condition is set up by small grey nodules appearing, which increase in size, giving an appearance of "grapes." Hence the condition is called grape disease, or Perlsucht. The organs, as we have said, are equally affected, and when we add the lymphatic glands we have a fairly complete summary of the form of the disease as it occurs in cattle. As has been clearly pointed out by Martin, Woodhead, and others in their evidence before the Royal Commission, the organs, glands, and membranes are the sites for tubercle, not the muscles (or "meat"). This latter is most liable to convey infection when the butcher smears it with the knife which he has used to remove tubercular organs.
As regards the udder in its relation to milk infection, it284 may be desirable to state that the initial lesion, according to Nocard and Bang, takes the form of a progressive sclerosis. The interlobular connective tissue, normally scanty, becomes thickened, fibrous, and infiltrated by minute miliary granulations. The granular tissue is thus "smothered by the hypertrophy and fibrous transformation of the interstitial connective tissue" (Nocard). The walls of the ducts are thickened and infiltrated, the lumen frequently dilated by masses of yellow caseous material. On the whole it may be said that tubercle of the udder is rare. Usually only one quarter is attacked, and by preference the posterior. For some time the milk remains normal, but gradually it becomes serous and yellow, and contains coagula holding numbers of bacilli. Lastly, it becomes purulent and dries up altogether. While the milk is undergoing these changes the lesion of the udder is becoming more marked, the tissue becomes less supple, and the toughness increases almost to a wooden hardness.
The general anatomical characteristics of the disease are similar to those occurring in man.
The percentage of cattle suffering from tubercle varies. In Germany it appears to vary from 2 to 8 per cent. of all cattle, in Saxony 17 to 30 per cent., in England 22 per cent. approximately (in London 40 per cent.), in France 25 per cent. Lowland breeds are much more infected than mountain breeds, which possess stronger constitutions.
Tuberculosis of the pig is less common than that of cattle, but not so rare as that of the calf (Nocard). In nine out of ten cases the pig is infected by ingestion, particularly when fed on the refuse from dairies and cheese factories. The disease follows the same course as in cattle. The finding of the bacillus is difficult, and the only safe test is inoculation (Woodhead).
Sheep are very rarely tuberculous by nature, though there is evidence to believe that very long cohabitation with285 tuberculous cattle would succeed in transmitting tuberculosis to some sheep.
Tuberculosis in the horse is relatively very rare. It attacks the organs of the abdominal cavity, especially the glands; it affects the lung secondarily as a rule. The cases are generally isolated ones, even though the animal belongs to a stud. Nocard holds that the bacillus obtained from the pulmonary variety is like the human type, whilst the abdominal variety is more like the avian bacillus.
Nocard says93:
"If the dog can become tuberculous from contact with man, the converse is equally true. Infection is at any rate possible when a house-dog scatters on the floor, carpet, or bed, during its fit of coughing, virulent material, which is rendered extremely dangerous by drying, especially for children, its habitual playmates. The most elementary prudence would recommend the banishment from a room of every dog which coughs frequently, even though it only seems to be suffering from some common affection of the bronchi or lung."
Tuberculosis is a common disease among the birds of the poultry-yard: poultry, pigeons, turkeys, pea-fowl, guinea-fowl, etc. They are infected almost exclusively through the digestive tract, generally by devouring infected secretions of previous tubercular fowls. Whatever the position or form of avian tuberculosis, the bacilli are present in enormous numbers, and are often much shorter and sometimes much longer than those met with in tuberculous mammalia, and grow outside the body at a higher temperature (43° C.). They are also said to be more resistant and of quicker growth. The species is probably identical with Koch's bacillus, though there are differences. In the nodule, which is larger than in human tuberculosis, there are few or no giant cells, and it does not so readily break down.
Nocard and others have demonstrated the fact that the Bacillus tuberculosis of Koch is the common denominator in all tubercular disease, whatever and wherever its manifestations, in all animals. The bacillus, they hold, may, however, experience profound modifications by means of successive passages through the bodies of divers species of animals. But if the modifications which it undergoes as a result of transmissions through birds, for example, are profound enough to make the bacillus of avian tubercle a peculiar variety of Koch's bacillus, they are not enough, it is generally believed, to make these bacilli two distinct species.
We may, therefore, take it for granted that tuberculosis is one and the same disease, with various manifestations, common to man and animals, intercommunicable, and having but one vera causa: the Bacillus tuberculosis of Koch.
The Prevention of Tuberculosis. At the present time much attention is being directed to the administrative personal control of tuberculosis. How greatly this is needed in so preventable a disease is evident from a perusal of the following quotation from the Registrar-General's reports. (See opposite page.)
These figures show a marked decline in the three worst forms of the disease. But this decline is apparently less marked in tabes than in phthisis or tubercular meningitis, i. e., less in the kind of tubercle due to the ingestion of infected milk. Fortunately the State is beginning to realise its duty in regard to preventive measures. The abolition of private slaughter-houses, the protection of meat and milk supplies, the seizure of tuberculous milch cows, and such like measures fall obviously within the jurisdiction of the State rather than the individual, and claim the earnest and urgent attention of the public health departments of states.94
ENGLISH DEATH-RATES FROM ALL TUBERCULAR DISEASES
THE FOLLOWING IS A TABLE OF DEATH-RATES TO A MILLION LIVING (ENGLAND), 1877–1897 (Reg.-Gen. Annual Reports):—
| 1877 | 1878 | 1879 | 1880 | 1881 | 1882 | 1883 | 1884 | 1885 | 1886 | 1887 | |
| Tabes Mesenterica | 316 | 348 | 300 | 370 | 284 | 313 | 289 | 310 | 251 | 300 | 253 |
| Tubercular Meningitis | 319 | 338 | 322 | 330 | 276 | 264 | 262 | 264 | 253 | 257 | 236 |
| Phthisis | 2079 | 2111 | 2021 | 1869 | 1825 | 1850 | 1880 | 1827 | 1770 | 1739 | 1615 |
| Other Forms | 126 | 124 | 116 | 129 | 145 | 153 | 160 | 170 | 157 | 177 | 179 |
| Total | 2840 | 2921 | 2759 | 2698 | 2530 | 2580 | 2591 | 2571 | 2431 | 2473 | 2283 |
| 1888 | 1889 | 1890 | 1891 | 1892 | 1893 | 1894 | 1895 | 1896 | 1897 | |
| Tabes Mesenterica | 240 | 269 | 265 | 251 | 242 | 265 | 192 | 243 | 196 | 201 |
| Tubercular Meningitis | 239 | 234 | 240 | 247 | 227 | 226 | 211 | 222 | 210 | 213 |
| Phthisis | 1568 | 1573 | 1682 | 1599 | 1468 | 1468 | 1385 | 1398 | 1307 | 1341 |
| Other Forms | 174 | 183 | 189 | 203 | 199 | 186 | 185 | 200 | 179 | 175 |
| Total | 2221 | 2259 | 2376 | 2300 | 2136 | 2145 | 1973 | 2063 | 1892 | 1930 |
Tabes mesenterica is tuberculosis of the alimentary canal and mesenteric lymph glands.
Tubercular meningitis is the name of the same disease as it affects the membranes of the brain (acute hydrocephalus).
Phthisis is the term applied to "consumption," or tubercle in the lungs.
But personal hygiene and the prevention of the transmission of the disease depend very largely indeed upon the mass of the population. Hence we hail with satisfaction the recent endeavours to educate public opinion. In order to make this matter very simple indeed, we have placed in a footnote a series of statements embodying some of the chief facts which every individual in our crowded communities should know.95
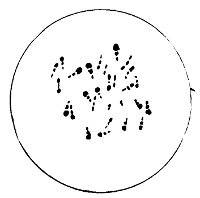
Bacillus of Diphtheria
Diphtheria (Klebs-Löffler Bacillus, 1882–1884). Diphtheria is an infective disease characterised by a variety of clinical symptoms, but commonly by a severe inflammation followed by a fibrous infiltration (constituting a membrane) of certain parts. The membrane ultimately breaks down. 290 The parts affected are the mucous membrane of the fauces, larynx, pharynx, trachea, and sometimes wounds and the inner wall of the stomach. The common sign of the disease is the membrane in the throat; but muscle weakness, syncope, albuminuria, post-diphtheritic paralysis, convulsions, and many other symptoms guide the physician in diagnosis and the course of the disease.
The Bacillus diphtheriæ was isolated from the many bacteria found in the membrane by Löffler. Klebs had previously identified the bacillus as the cause of the disease. 291 It is a slender rod, straight or slightly curved, and remarkable for its beaded appearance; there are also irregular and club-shaped forms. It differs in size according to its culture medium, but is generally 3 or 4 µ in length. In the membrane which is its strictly local habitat in the body—indeed, the bacillus is found nowhere else in the body—it almost invariably shows parallel grouping, lying between the fibrin of the membrane, and most largely in its deeper parts. Here it is mixed with other bacilli, micrococci, staphylococci, and streptococci, all of which are present and performing 292 their part in complicating the disease. The bacillus possesses five negative characters; namely, it has no spores, threads, or power of mobility, and does not produce liquefaction or gas. It stains with Löffler's methylene blue, and shows metachromatic granules and polar staining. Its favourable temperature is blood-heat, though it will grow at room temperature. It is aërobic, and, indeed, prefers a current of air. Löffler contrived a medium for cultivation which has proved most successful. It is made by mixing three parts of ox-blood serum with one part of broth containing 1 per cent. of glucose, 1 per cent. of peptone, and 1/2 per cent. of common salt; the whole is coagulated. Upon this medium the Klebs-Löffler bacillus grows rapidly in eighteen or twenty hours, producing scattered "nucleated" round white colonies, becoming yellowish. It grows well in broth, but without producing either a pellicle or turbidity; it can grow on the ordinary media, though its growth on 293 potato is not visible; on the white of egg it flourishes extremely well.
It retains its vitality in cultures and sometimes in the throat for months. Three or four weeks is the average length of time for its existence in the membrane, but, owing to the difficulty of killing it in situ, it may live on for as long as a year. All the conditions in the throat—mucous membrane, blood-heat, moisture, air—are extremely favourable to the bacillus; but it is very materially modified in virulence. It is secured for diagnostic purposes by one of two methods: (a) Either a piece of the membrane is detached, and after washing carefully examined by culture as well as the microscope; or (b) a "swab" is made from the infected throat and cultured on serum, and incubated at 37° C. for eighteen hours and then microscopically examined. Both methods—and there is no further choice—present some difficulties owing to the large number of bacteria found in the throat. Hence a negative result must be accepted with reserve.
We have already referred at some length to the question of toxins in diphtheria, and need not dwell further upon that matter. Still a word or two may be said here summarising the general action of the bacillus. Locally it produces inflammatory change with fibrinous exudation and some cellular necrosis. In the membrane a ferment is probably produced which, unlike the localised bacilli, passes throughout the body and by digestion of the proteids produces albumoses and an organic acid which have the toxic influence. The toxins act on the blood-vessels, and nerves, and muscle fibres of the heart, and many of the more highly specialised cells of the body. Thus we get degenerative changes in the kidney, in cells of the central nervous system, in the peripheral nerves (post-diphtheritic paralysis), and elsewhere, these pathological conditions setting up, in addition to the membrane, the signs of the disease. The bacil294lus is pathogenic for the horse, ox, rabbit, guinea-pig, cat, and some birds. Cases are on record of supposed infection of children by cats suffering from the disease. The horse, it will be remembered, yields the antitoxin which has saved so many lives (Metropolitan Asylums Board Report, 1896).
The influence of drainage, milk, and schools must not be forgotten by sanitary authorities any more than the essential importance of adequate isolation hospital accommodation. Mr. Shattock's experiments on the effect of sewer air upon attenuated Klebs-Löffler bacilli have been mentioned (see p. 105). Nevertheless there can be no doubt that emanations from defective drains have a materially predisposing effect, not, it is true, upon the bacilli, but upon the tissues. Sore throats thus acquired are par excellence the site for the development of diphtheria.
The influence of school attendance has claimed the recent attention of the Medical Officer of the London School Board and the Medical Officer of the administrative County of London. In London since 1881 there has been a marked increase of diphtheria, which has occurred, though in a much less degree, throughout England and Wales.
The Registrar-General has only classified diphtheria as a separate disease since 1855, when the death-rate per 1,000,000 in England and Wales was stated as 20. The following are the figures for four decades up to 1895:
AVERAGE DEATH-RATE PER MILLION OF THE POPULATION FROM DIPHTHERIA IN ENGLAND AND WALES AND IN LONDON (IN DECADES 1856–95)
| England and Wales | London. | |
| 1856–65 | 246.9 | 225.4 |
| 1865–75 | 124.8 | 123.5 |
| 1875–85 | 129.0 | 176.7 |
| 1885–95 | 210.6 | 421.4 |
From these figures the extraordinary increase during the last few years is clearly demonstrated.
Sir Richard Thorne Thorne, in 1891, drew attention to the influence of damp soils and schools upon diphtheria. In 1894 Mr. Shirley Murphy, Medical Officer to the London County Council, reported that there had been an increase in diphtheria mortality in London at school ages (three to ten) as compared with other ages since the Elementary Education Act became operative in 1871; that the increased mortality from diphtheria in populous districts, as compared with rural districts, since 1871, might be due to the greater effect of the Education Act in the former; and that there was a diminution of diphtheria in London during the summer holidays at the schools in 1893, but that 1892 did not show any marked changes for August.
In 1896 Professor W. R. Smith, the Medical Officer to the London School Board, furnished a report96 on this same subject of school influence, in which he produces evidence to show that the recrudescence of the disease in 1881–90 was greatest in England and Wales at the age of two to three years, and in London at the age of one to two years, in both cases before school age; that age as an absolute factor in the incidence of the disease is enormously more active than any school influence, and that personal contact is another important source of infection.
Although it is said that "statistics can be made to prove anything," there can be little doubt that both of these reports contain a great deal of truth; nor are these truths incompatible with each other. They both emphasise age as a great factor in the incidence of the disease, and whatever affects the health of the child population, like schools, must play, directly or indirectly, a not unimportant part in the transmission of the disease.
The Pseudo-diphtheria Bacillus.97 Löffler and Hoffman described a bacillus having the same morphological characters as the true Bacillus diphtheriæ, except that it had no virulence. Roux believes this is merely an attenuated diphtheria bacillus. It is frequently found in healthy throats. The chief differences between the real and the pseudo-bacillus are:
1. The pseudo-bacillus is thicker in the middle than at the poles, and not so variable as the Bacillus diphtheriæ. Polar staining is absent.
2. Its growth on potato reveals cream-coloured colonies visible in a couple of days; the real bacillus is invisible.
3. The pseudo-bacillus will not grow at all anaërobically in hydrogen, but the Bacillus diphtheriæ is able to do so.
4. There is the great difference in virulence.
Suppuration. This term is used to designate that general breaking down of cells which follows acute inflammation. An "abscess" or "gathering" is a collection, greater or smaller, of the products of suppuration. The word pus is generally used to describe this matter. We may have such an advanced inflammatory condition in any locality of the body, and it will assume different characters according to its site. Hence there are connected with suppuration, as causal agents, a variety of bacteria. Pus is not matter containing a pure culture of any specific species, but, on the contrary, is generally filled with a large number of different species. The most important are as follows:
1. Staphylococcus pyogenes aureus. These are micrococci arranged in groups, which have been likened to bunches of grapes. They are the common organisms found in pus, and were with other auxiliary bacteria first distinguished as such by Professor Ogston, of Aberdeen. There are several forms of the same species, differing from each other in colour.
Thus we have the S. pyogenes aureus (golden yellow), albus (white), citreus (lemon), and others. They occur commonly in nature, in air, soil, water, on the surface of the skin, and in all suppurative conditions. The aureus is the only one credited with much virulence. It occurs in the blood in blood-poisoning (septicæmia, pyæmia), and is present in all ulcerative conditions, including ulcerative disease of the valves of the heart.
The Staphylococcus cereus albus and S. cereus flavus are slightly modified forms of the S. pyogenes aureus, and are differentiated from it by being non-liquefying. They produce a wax-like growth on gelatine.
Staphylococcus pyogenes aureus, the type of the family, is grown in all ordinary media at room temperature, though more rapidly at 37° C. Liquefaction sets in at a comparatively early date, and subsequently we have in the gelatine test-tube cultures a flocculent deposit of a bright yellow amorphous mass, and in gelatine plates small depressions of liquefaction with a yellow deposit. It renders all media acid, and coagulates milk. Its thermal death-point in gelatine is 58° C. for ten minutes, but when dry considerably higher. It is a non-motile and a facultative anaërobe; but the presence of oxygen is necessary for a bright colour. Its virulence readily declines.
2. Streptococcus pyogenes. In this species of micrococcus the elements are arranged in chains. Most of the streptococci in pus, from different sources, are one species, having approximately the same morphological and biological characters. Their different effects are due to different degrees of toxic virulence; they are always more virulent when associated with other bacteria, for example, the Proteus family.
The chains vary in length, consisting of more elements when cultured in fluid media. They multiply by direct division of the individual elements, and in old cultures it has been observed that the cocci vary in form and size.298 This latter fact gave support to the theory that streptococcus reproduced itself by arthrospores, or "mother-cells."
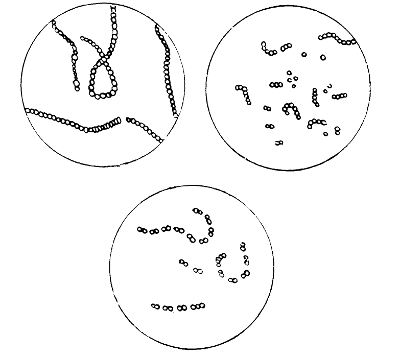
Types of Streptococcus
In culture upon the ordinary media streptococcus is comparatively slow-growing, producing minute white colonies on or about the sixth day. It does not liquefy gelatine, and remains strictly localised to the track of the inoculating needle. Like the staphylococcus, it readily loses virulence. The thermal death-point is, however, lower: 54° C. for ten minutes. Marmorek has devised a method by which the virulence may be greatly increased, and he holds that it depends upon the degree of virulence possessed by any particular streptococcus as to what effects it will produce. By the299 adoption of Marmorek's methods attempts have been made to prepare an antitoxin.
Streptococcus pyogenes has been isolated from the membrane of diphtheria, and from small-pox, scarlet fever, vaccinia, and other diseases. In such cases it is not the causal agent, but merely associated with the complications of these diseases. Suppuration and erysipelas are the only pathological conditions in which the causal agency of streptococcus has been sufficiently established.
3. The Bacillus pyocyaneus occurs in green pus, and is the cause of that colouration. Gessard was the first to prove its significance, and he describes two varieties.

Micrococcus Tetragonus
It is a minute, actively motile, non-sporulating bacillus, which occasionally complicates suppuration and produces green pus. Oxygen is necessary for pigmentation, which is due to two substances: pyocyanin, a greenish-blue product extracted with chloroform, and pyoxanthose, a brown substance derived from the oxidation of the former pigment. Both these colours are produced in cultivation outside the body. On gelatine the colour is green, passing on to olive. There is liquefaction. On potato we generally obtain a brown growth (compare Bacillus coli, B. mallei, and others). The organism grows rapidly on all the ordinary media, which it has a tendency to colour throughout.
It will be remembered that when speaking of the antagonism of organisms, we referred to the inimical action of Bacillus pyocyaneus upon anthrax.
4. Micrococcus Tetragonus. This species occurs in phthisi300cal cavities and in certain suppurations in the region of the face. It is a micrococcus usually in the form of small tetrads. A capsule is always present and sometimes discernible.
5. Bacillus coli communis and many putrefactive germs commonly occur in suppurative conditions, but they are not restricted to such disorders (see p. 64).
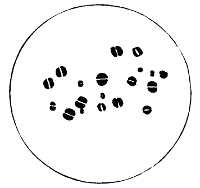
Diplococcus of Neisser
6. Micrococcus gonorrhœæ (Neisser, 1879). This organism is more frequently spoken of as a diplococcus. It occurs at the acute stage of the disease, but is not readily differentiated from other similar diplococci except by technical laboratory methods. Each element presents a straight or concave surface to its fellow. A very marked concavity indicates commencing fission. The position which these diplococci take up in pus is intracellular, and arranged more or less definitely around the nucleus. Difficulty has often been found in cultivating this organism in artificial media outside the body. Wertheim and others have suggested special formulæ for the preparation of suitable media, but it is a very simple matter to secure cultures on agar plates smeared with human blood from a pricked finger. The plate is incubated at 37° C. At the end of twenty-four hours small raised grey colonies appear, which at the end of301 about four days show adult growth. The margin is uneven, and the centre more opaque than the rest of the colony. This diplococcus is readily killed, and sub-cultures must be frequently made to retain vitality and virulence. Light, desiccation, and a temperature of 55° C. all act germicidally. The organism stains readily in Löffler's blue, but is decolourised by Gram's method. It is more or less strictly parasitic to man. Its shape, size, character of growth, and staining properties assist in differentiating it from various similar diplococci.98
Anthrax. This disease was one of the first in which the causal agency of bacteria was proved. In 1849 Pollender found an innumerable number of small rods in the blood of animals suffering from anthrax. In 1863 Davaine described these, and attributed the disease to them. But it was not till 1876 that Koch finally settled the matter by isolating the bacilli in pure culture and describing their biological characters.
It is owing in part to its interesting bacterial history, which opened up so much new ground in this comparatively new science, that anthrax has assumed such an important place in pathology. But for other reasons, too, it claims attention. It appears to have been known in the time of Moses, and was perhaps the disease described by Homer in the First Book of the Iliad. Rome was visited by it in 740 B.C.
Anthrax is an acute disease, affecting sheep, cattle, horses, goats, deer, and man. Cats, white rats, and Algerian sheep are immune. Swine become infected by feeding on the offal of diseased cattle (Crookshank).
The post-mortem signs are mainly three: The spleen is greatly enlarged and congested, is friable to the touch, and contains enormous numbers of bacilli; the skin may show exudations forming dark gelatinous tumours; and the blood 302 remains fluid for some time after death, is black, tar-like, contains bubbles of air, and shows other degenerative changes in the red corpuscles, whilst the small blood-vessels contain such vast quantities of bacilli that they may be ruptured by them. Particularly is this true in the peripheral arteries. Many of the organs of the body show marked congestion.
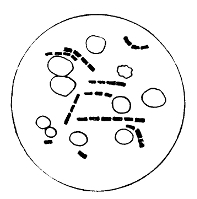 |
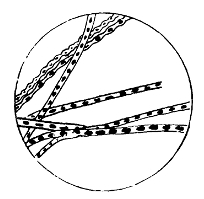 |
Bacillus of Anthrax and |
Threads of Bacillus |
Clinically there is rise of temperature and rapid loss of muscular power. The bacilli of anthrax are square-ended rods 1 µ broad and 4–5 µ long. In the tissues of the body they follow the lines of the capillaries, and are irregularly situated. In places they are so densely packed as to form obstructions to the onward flow of blood. In cultures they are in chains end to end, having as a rule equal interbacillary spaces. In cultures long filaments and threads occur. The exact shape of the bacillus depends, however, upon two things: the staining and spore formation. Both these factors may very materially modify the normal shape. The spores of anthrax are oval endospores, produced only in the presence of free oxygen, and at any temperature between 18 and 41° C. On account of requiring free oxygen, they are formed only outside the body. The homogeneous pro303toplasm of the bacillus becomes granular; the granules coalesce, and we have spores. Each spore possesses a thick capsule, which enables it to resist many physical conditions which kill the bacillus. When the spore is ripe or has exhausted the parent bacillus, it may take on a resting stage, or under favourable circumstances commence germination, very much after the manner of a seed. The spores may infect a farm for many months; indeed, cases are on record which appear to prove that the disease on a farm in the autumn may by means of the spores be carried on by the hay of the following summer into a second winter. Thus, by means of the spores, the infection of anthrax may cling to the land for very long periods, even for years. Spores of anthrax can withstand 5 per cent. carbolic acid or 1–1000 corrosive sublimate for more than an hour; even boiling does not kill them at once, whilst the bacilli without their spores are killed at 54° C. in ten minutes. When the spores are dry they are much more resistant than when moist. Hence the persistence of the anthrax bacillus is due to its spores.
The bacillus is aërobic, non-motile, and liquefying. Broth cultures become turbid in thirty-six hours, with nebulous masses of threads matted together. The pellicle which forms on the surface affords an ideal place for spore formation.
Cultures in the depth of gelatine show a most characteristic growth. From the line of inoculation delicate threads and fibrillæ extend outwards horizontally into the medium. Liquefaction commences at the top, and eventually extends throughout the tube. On gelatine plates small colonies appear in thirty-six hours, and on the second or third day they look, under a low power of the microscope, like matted hair. The colonies after a time sink in the gelatine, owing to liquefaction. On potato, agar, and blood serum anthrax grows well.
Channels of Infection. 1. The Alimentary Canal. This is the usual mode of infection in animals grazing on infected pasture land. A soil suitable for the propagation of anthrax is one containing abundance of air and proteid material. Feeding on bacilli alone would probably not produce the disease, owing to the germicidal effect of the gastric juice. But spores can readily pass uninjured through the stomach and produce anthrax in the blood. Infected water as well as fodder may convey the disease. Water becomes infected by bodies of animals dead of anthrax, or, as was the case once at least in the south-west of England, by a stream passing through the washing-yard of an infected tannery. Manure on fields, litter in stalls, and infected earth may all contribute to the transmission of the disease. Darwin pointed out the services which are performed in superficial soils by earthworms bringing up casts; Pasteur was of opinion that in this way earthworms were responsible for continually bringing up the spores of anthrax from buried corpses to the surface, where they would reinfect cattle. Koch disputed this, but more recently Bollinger has demonstrated the correctness of Pasteur's views by isolating anthrax contagium from five per cent. of the worms sent him from an anthrax pasture. Bollinger also maintains that flies and other insects may convey the disease from discharges or carcasses round which they congregate.
Alimentary infection in man is a rare form, and it reveals itself in a primary diseased state known as mycosis intestinalis, an inflamed condition of the intestine and mesenteric lymph glands.
2. Through the Skin. Cutaneous anthrax goes by the name of malignant pustule, and is caused by infective anthrax matter gaining entrance through abrasions or ulcers in the skin. This local form is obviously most contracted by those whose occupation leads them to handle hides or other anthrax material (butchers and cleaners of hides).305 Two or three days after inoculation a red pimple appears, which rapidly passes through a vesicular stage until it is a pustule. Concomitantly we have glandular enlargement, general malaise, and a high temperature. Thus from a local sore a general infection may result. Unless this does occur, the issue will not be fatal, and the bacilli will never gain entrance into the blood or be anything but local.
3. Respiratory Tract. In man this is the commonest form of all, and is well known under the term "wool-sorters' disease," or pulmonary anthrax. This mode of infection occurs when dried spores are inhaled in processes of skin-cleaning. It frequently commences as a local lesion affecting the mucous membrane of the trachea or bronchi, but it rapidly spreads, affecting the neighbouring glands, which become greatly enlarged, and extending to the pleura and lung itself. Such cases, as a rule, rapidly end fatally.
From what has been said, it will be clear that anthrax carcasses are better not opened and exposed to free oxygen. An extended post-mortem examination is not necessary. Burning the entire carcass in a crematorium would be the ideal treatment. As such is not generally feasible the next best thing is to bury the carcass deeply with lime below and above it, and rail in the area to prevent other animals grazing off it.
A very small prick will extract enough blood to examine for the anthrax bacilli which are driven by the force of the blood-current to the small surface capillaries. This occurs, of course, only when the disease has become quite general, for in the early stage the healthy blood limits the bacilli to the internal organs. In such cases examination of the blood of the spleen is necessary.
Anthrax covers a wide geographical area all over the world, and no country seems altogether exempt. In Germany as many as 3700 animals have been lost in a single year. About 900 animals were attacked in 1897 in Great Britain.
Plague. This disease, like anthrax and leprosy, has a long historical record behind it. As the Black Death it decimated the population of England in the fourteenth century, and visited the country again in epidemic form in the middle of the seventeenth century, when it was called the Great Plague. Now, it is highly probable that these two scourges and the recent epidemic in the East are all forms of one and the same disease. As a matter of fact, it is difficult to be sure what was the exact pathology of a number of the grievous ailments which troubled our country in the Middle Ages, but from all accounts bubonic plague and true leprosy were amongst them. The former came and went spasmodically, as is its habit; the latter dragged through the length of several centuries.

Bacillus of Plague
The distribution of plague at the present time is fortunately a somewhat limited one, namely, a definite area in Asia known as the "Plague Belt." From Mesopotamia, as a sort of focus, the disease spreads northwards to the Caspian Sea, westwards to the Red Sea, southwards as far as Central India, and eastwards as far as the China Sea. This constitutes the "belt," but the disease may take an epidemic form, and is readily, though very slowly, conveyed by infection or contagion. It appears to be infectious by means of infective dust, and contagious by prolonged and intimate contact with the plague-stricken. Rats have been accused of conveying the disease from port to port, and307 even infecting man. It is clear that rats are not the only agency acting in this way. Nevertheless it is true that rats contract the disease more readily than any other animals, and that when suffering from it they may spread the infection. How it is thus spread it is not known. Drs. Cantlie and Yersin have pointed out that previously to an epidemic of plague rats die in enormous numbers.
The bacteriology of plague is almost the latest addition to the science. Kitasato, of Tokio, demonstrated the cause of plague to be a bacillus during the Hong Kong epidemic in 1894. This was immediately confirmed by Yersin, and further proved by the isolation in artificial media of a pure culture of a bacillus able to cause the specific disease of bubonic plague.
The bacillus was first detected in the blood of patients suffering from the disease. It takes the form of a small, round-ended, oval cell, with marked polar staining, and hence having an appearance not unlike a diplococcus. In the middle there is a clear interspace, and the whole is surrounded with a thick capsule, stained only with difficulty. The organisms are often linked together in pairs or even chains, and exhibit involution forms. In culture the bacillus is even more coccal in form than in the body.
The plague bacillus grows readily on the ordinary media at blood-heat, producing circular cream-coloured colonies with a wavy outline, which eventually coalesce to form a greyish film. The following negative characters help to distinguish it: No growth occurs on potato, milk is not coagulated, and gelatine is not liquefied; Gram's method does not stain the bacillus, and there are no spores; the bacillus is readily killed by heat and by desiccation over sulphuric acid at 30° C. Both in cultures and outside the body the bacillus loses virulence. To this may be attributed possibly the variety of forms of the plague bacillus which differ in virulence. On gaining entrance to the human body308 the bacillus affects in particular two organs, the spleen and the lymph glands. The latter become inflamed in groups, commencing frequently with those in the armpit (axillary) or groin (inguinal). The spleen suffers from inflammatory swelling, which may affect other organs also. In both places the bacilli occur in enormous numbers. Kitasato considers that the bacillus may enter the body by the three channels adopted by anthrax, namely, the skin, alimentary canal, and respiratory tract.
Haffkine has prepared a vaccine to be used as a prophylactic. He grows a pure culture of Kitasato's bacillus in broth upon the surface of which some globules of fat ("ghee") have been placed. The bacillus grows upon this fat in copious stalactite form. From time to time this growth is shaken down, until after five or six weeks the shaken broth appears milky. The contained bacilli are killed by heating the fluid to 70° C. for one hour. The resultant is the vaccine, of which the dose is 3 cc. Haffkine believes that inoculated persons in India have suffered twenty times less than non-inoculated living under the same conditions.
Plague is essentially a "filth disease," and it is frequently preceded by famine. Temperature and overcrowding exert an influence upon it. The areas affected by the disease in the Middle Ages, in the seventeenth century, and in 1894–96 are alike in being characterised by filth and overcrowding. There is little fear, speaking generally, of the plague ever flourishing under Western civilisation, where the conditions are such that even when it appears there is little to encourage or favour its development.
Leprosy. This ancient disease is said to have existed in Egypt 3500 B.C., and was comparatively common in India, China, and even in parts of Europe 500 B.C. We know it has existed in many parts of the world in the past, in which regions it is now extinct. Some of the earliest notices we309 have of it in this country come from Ireland, and date back to the fifth and sixth centuries. Even at that period of time also various classical descriptions of the disease had been written and various decrees made by the Church councils to protect lepers and prevent the spread of the disease, which was often looked upon as a divine visitation. In the tenth century leprosy was prevalent in England; it reached its zenith in the thirteenth century, or possibly a little earlier, and declined from that date to its extinction in the sixteenth. But even two hundred years later leprosy was endemic in the Shetlands, and it is recorded that in 1742 there was held a public thanksgiving in Shetland on account of the disappearance of leprosy.
At one time or another there were as many as two hundred institutions in the British Isles for the more or less exclusive use of lepers. Many of these establishments were of an ecclesiastical or municipal character, and probably the exact diagnosis of diseases was a somewhat lax matter. Bury St. Edmunds, Bristol, Canterbury, London, Lynn, Norwich, Thetford, and York were centres for lepers. Burton Lazars and Sherburn, in Durham, were two of the more famous leper institutions.
At the present time the distribution of the disease is mostly Asiatic. Norway contains about 1200 lepers, Spain approximately the same number. Scattered through Europe are perhaps another 600 or 700, in India 100,000, and a large number in Japan. The Cape possesses a famous leper hospital on Robben Island, with a number of patients. The disease is also endemic in the Sandwich Islands.
Descriptions of the pathological varieties of leprosy have been very diverse. The classification now generally adopted includes three forms: the tuberculated, the anæsthetic (or maculo-anæsthetic), and the mixed. Lepra tuberculosa is that form of the disease affecting chiefly the skin, and resulting in nodular tuberculated growth or a diffuse infil310tration. It causes great disfigurement. The anæsthetic form causes a destruction of the nerve fibres, and so produces anæsthesia, paralysis, and what are called "trophic" changes. Not infrequently patches occur on the skin, which appear like parchment, owing to this trophic change. Bullæ may arise. When the tissue change is radical or far advanced, considerable distortion may result. The mixed variety of leprosy, as its name implies, is a mixture of the two other forms.
The Bacillus lepræ was discovered by Hansen in 1874. He found it in the lepra cells in the skin, lymph glands, liver, spleen, and thickened parts of the nerves. It is common in the discharges from the wounds of lepers. It is conveyed in the body by the lymph stream, and has rarely been isolated from the blood (Köbner).
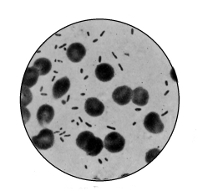 |
 |
Bacillus of Plague |
Bacillus of Leprosy (Hansen) |
 |
 |
Streptothrix Actinomyces |
Bacillus Mallei (Glanders) |
The bacillus is present in enormous numbers in the skin and tissues, and has a form very similar indeed to Bacillus tuberculosis. It is a straight rod, and showing with some staining methods marked beading, but with others no beading at all. It measures 4 µ long and 1 µ broad. Young leprosy bacilli are said to be motile, but old ones are not. Neisser has maintained that the bacillus possesses a capsule and spores. The latter have not been seen, but Neisser holds that this is the form in which the bacillus gains entrance to the body. There is a characteristic which fortunately aids us in the diagnosis of this disease in the tissues, and that is the arrangement of the bacilli, which are rarely scattered or isolated, but gathered together in clumps and colonies. Bordoni-Uffreduzzi and Campania claim to have isolated the bacillus and grown it on artificial media, the former aërobically on peptone-glycerine-blood-serum, at 37° C., the latter anaërobically. But no other worker has been able to do this. Hence we are not able to study the bacteriology of leprosy at all completely, nor have inoculation experiments proved successful. Nevertheless there is 311 little doubt that leprosy is a bacterial disease produced by the bacillus of Hansen. Bordoni-Uffreduzzi maintains that the parasitic existence of the Bacillus lepræ may alternate with a saprophytic stage. This may be of importance in the spread of the disease. There is evidence in support of the non-communicability of the disease by heredity or contagion. Segregation does not appear always to result in a decline of the disease, as we should expect if it were purely contagious. Ehlers, of Copenhagen, has, however, as recently as 1897, reaffirmed his belief in the contagiousness of leprosy; Virchow, on the other hand, has declared that it is not highly contagious. There is evidence to show that persons far advanced in the disease may live in a healthy community and yet not infect their immediate neighbours. Indeed, the transmission of the disease is still an unsolved problem. Mr. Hutchinson suggests diet, particularly uncooked or putrid fish, as a likely channel; on the other hand, leprosy appears in districts where no fish is eaten. Deficiency of salt, telluric and climatic conditions, racial tendencies, social status, poverty, insanitation, drinking water, even vaccination, have all secured support from various seekers after the true channel by which the bacillus gains entrance to the human body. The real mode of transmission is, however, still unknown. The decline and final extinction of leprosy in the British Islands was probably due in part to the natural tendency of the disease, under favourable hygienic circumstances, to die out, and in part to a general and extensive social improvement in the life of the people, to a complete change in the poor and insufficient diet, and to agricultural advancement, improved sanitation, and land drainage.
At the Leprosy Congress held in Berlin in 1897, Hansen again emphasised his belief that segregation was the cause of the decline of leprosy wherever it had occurred. But there appears to be some evidence to show that leprosy has312 declined where there has been no segregation whatever, and therefore, however favourable to decline such isolation may be, it would seem not to be actually necessary to the decline. At the same Congress Besnier declared in favour of the infective virus being widely propagated by means of the nasal secretion. Sticker states that the nasal secretion contains millions of lepra bacilli, especially in the acute stage of the disease, and Besnier and Sticker have pointed out how frequently and severely the septum nasi and skin over the nose are affected in leprosy. Several leprologists in India have recorded similar observations. These facts appear to support Besnier's contention that the disease is spread by nasal secretion.
We may fitly add here the conclusions arrived at by the English Leprosy Commission99 in India:
"1. Leprosy is a disease sui generis; it is not a form of syphilis or tuberculosis, but has striking etiological analogies with the latter.
"2. Leprosy is not diffused by hereditary transmission, and, for this reason and the established amount of sterility among lepers, the disease has a natural tendency to die out.
"3. Though in a scientific classification of diseases leprosy must be regarded as contagious, and also inoculable, yet the extent to which it is propagated by these means is exceedingly small.
"4. Leprosy is not directly originated by the use of any particular article of food, nor by any climatic or telluric conditions, nor by insanitary surroundings, neither does it peculiarly affect any race or caste.
"5. Leprosy is indirectly influenced by insanitary surroundings, such as poverty, bad food, or deficient drainage or ventilation, for these by causing a predisposition increase the susceptibility of the individual to the disease.
313"6. Leprosy, in the great majority of cases, originates de novo, that is, from a sequence or concurrence of causes and conditions dealt with in the Report, and which are related to each other in ways at present imperfectly known."
The practical suggestions of the Commission for preventive treatment included voluntary isolation, prohibition of the sale of articles of food by lepers, leper farms, orphanages, and "improved sanitation and good dietetic conditions" generally. Serum-therapy has been attempted on behalf of the French Academy of Medicine, but without success. Many forms of treatment ameliorate the miserable condition of the leper, but up to the present no curative agent has been found.
Diplococcus of Pneumonia
Pneumonia. Some of the difficulty which has surrounded the bacteriology of inflammation of the lungs is due to the confusion arising from supposing that attacks of the disease differed only in degree. Pneumonia, however, has various forms, arising now from one cause, now from another. The specific or croupous pneumonia is associated with two organisms: Fraenkel's diplococcus and Friedländer's pneumo-bacillus. Several other bacteria have from time to time been held responsible for pneumonia, a streptococcus re314ceiving, at one time, some support. But whilst opinion is divided on the rôle of various extraneous and concomitant bacteria in lung disease, importance is attached to Fraenkel's and Friedländer's organisms.
The diplococcus of Fraenkel is a small, oval diplococcus found in the "rusty" sputum of croupous pneumonia. It is non-motile, non-liquefying, and aërobic. When examined from cultures the diplococci are frequently seen in chains, not unlike a streptococcus, and there is some reason to suppose that this form gave rise to the belief that it was another species; when examined from the tissues it possesses a capsule, but in culture this is lost. It is difficult to cultivate, but grows on glycerine agar and blood serum at blood-heat. On ordinary gelatine at room temperature it does not grow, or, if so, very slightly. The ideal fluid is a slightly alkaline liquid medium, and in twenty-four hours a powdery growth will occur in such broth. On potato there is apparently no growth. It rapidly loses its virulence on solid media, and is said to be non-virulent after three or four sub-culturings. A temperature of 54–58° C. for a few minutes kills the bacteria, but not the toxin. This, however, is removed by filtration, and is therefore probably intracellular. It is attenuated by heating to 70° C.
Fraenkel's diplococcus occurs, then, in the acute stage of pneumonia, in company with streptococci and staphylococci. It also occurs in the blood in certain suppurative conditions, in pleurisy and inflammation of the pericardium, and sometimes in diphtheria, and therefore it is not peculiar to pneumonia.
There is one other point to which attention should be drawn. Fraenkel's organism is said to be frequently present in the saliva of healthy persons. Pneumonia depresses the resistant vitality of the tissues, and thus affords to the diplococcus present in the saliva an excellent nidus for its growth.
Friedländer's Pneumo-bacillus is a capsulated oval coccus, assuming the form of a small bacillus. It is inconstant in pneumonia, unequally distributed, and scarce; it is aërobic, and facultatively anaërobic; it occasionally occurs in long forms and filaments; it is non-motile, non-liquefying, and has no spores; it does not stain by Gram's method, which stain is therefore used for differential diagnosis; it will grow fairly well in ordinary gelatine at 20° C.; and it is a denitrifying organism, and also an actively fermentative one, even fermenting glycerine. It is not unlike Bacillus coli communis, and to distinguish it from that organism we may remember that the B. coli is motile, never has a capsule, produces indol, and does not ferment glycerine.

Bacillus of Influenza
Influenza. In 1892, during the pandemic of influenza, Pfeiffer discovered a bacillus in the bronchial mucus of patients suffering from the disease. It is one of the smallest bacilli known, and frequently occurs in chains not unlike a streptococcus. Carron obtained the same organism from the blood. In the bronchial expectoration it can retain its virulence for as long as a fortnight, but it is quickly destroyed by drying. The bacillus is aërobic, non-motile, and up to the present spores have not been found. It grows somewhat feebly in artificial media, and readily dies out. Blood serum, glycerine agar, broth, and gelatine have all been used at blood-heat. It does not grow at room tem316perature. Pfeiffer's bacillus appears most abundantly at the height of the disease, and disappears with convalescence. It is said not to appear in any other disease.
Yellow Fever. Sternberg and Havelburg have both isolated bacilli from cases of yellow fever; but the organism discovered by Sanarelli, the Bacillus icteroides, is now accepted as the causal agent of the disease. It is a small, short rod, with round ends, and generally united in pairs; it has various pleomorphic forms; it grows well on all the ordinary media; it is killed in sea-water at 60° C., and also by direct sunlight in a few hours.
Diarrhœa of Infants. From time to time different organisms have been isolated in this diseased condition. Bacillus coli and B. enteriditis sporogenes (Klein) have been held responsible for it. W. D. Booker, of Johns Hopkins University, sums up an extended research into the question as follows:
"No single micro-organism is found to be the specific exciter of the summer diarrhœa of infants, but the affection is generally to be attributed to the result of the activity of a number of varieties of bacteria, some of which belong to well-known species, and are of ordinary occurrence and wide distribution, the most important being the streptococcus and Proteus vulgaris.
"The first step in the pathological process is probably an injury to the epithelium from abnormal or excessive fermentation, from toxic products of bacteria, or from other factors.
"Bacteria exert a direct injury upon the tissues in some instances, whereas in others the damage is brought about indirectly through the production of soluble poisons."
Actinomycosis. This disease affects both animals and man. As Professor Crookshank points out, it has long been known in this country,100 but its various manifestations have 317been mistaken for other diseases or have received popular names.
Here we can only mention the most outstanding facts concerning the disease. It is caused by the "ray fungus," or Streptothrix actinomyces, which, growing on certain cereals, often gains entrance to the tissues of man and beast by lacerations of the mucous membrane of the mouth, by wounds, or by decayed teeth. Barley has been the cereal in question in some cases. The result of the introduction of the parasite is what is termed an "infective granuloma." This is, generally speaking, of the nature of an inflammatory tumour composed of round cells, epithelioid cells, giant cells, and fibrous tissue, forming nodules of varying sizes. In some cases they develop to large tumours, in others they soon break down. Actinomycosis in some ways closely resembles tuberculosis in its tissue characters.
In the discharge or pus from human cases of the disease small sulphur-yellow bodies may be detected, and these are tufts of "_clubs_" which are the broken-down rays of the parasite; for in the tissues which are affected the parasite arranges itself in a radiate manner, growing and extending at its outer margin and degenerating behind. In cattle the centre of the old ray becomes caseated, like cheese, or even calcified, like a stone. In the human disease abundant "_threads_" are formed as a tangled mass in the middle of the colony. As clubs characterise the bovine actinomycosis, so threads are a feature of the human form of the disease. But in both there is a third element, namely, small round cells, called by some spores, by others simply cocci. Authorities are not yet agreed as to the precise significance and rôle of these round cells. The life-history of the micro-organism may be summed up thus:
"The spores sprout into excessively fine, straight or sinuous, and sometimes distinctly spirilliform threads, which branch irreg318ularly and sometimes dichotomously. The extremities of the branches develop the club-shaped bodies. The clubs are closely packed together, so that a more or less globular body is formed, with a central core composed of a dense mass of threads" (Crookshank).
Possibly these clubs represent organs of fructification, and produce the spores. These latter are, it is believed, set free in the vicinity of the ray, and create fresh centres of disease.
In man the disease manifests itself in various parts according to the locality of entrance. When occurring in the mouth it attacks the lower jaw most frequently. In one recorded case the disease was localised to the bronchi, and did not even extend into the lungs. It was probably contracted by inhalation of the parasite. The disease may spread to distant parts by means of the blood stream, and frequently the abscesses are apt to burrow in various directions.
In the ox the disease remains much more localised, and frequently occurs in the lower jaw, palate, or tongue. In the last site it is known as "wooden tongue," owing to the hardness resulting. The skin and subcutaneous tissues are also a favourite seat of the disease, producing the so-called wens or clyers so commonly seen in the fen country (Crookshank). Actinomycosis in cattle is specially prevalent in river valleys, marshes, and on land reclaimed from the sea. The disease occurs at all seasons, but perhaps more commonly in autumn and winter. It is more frequently met with in young animals. The disease is probably not hereditary nor readily communicated from animal to animal.
Actinomyces may be cultivated, like other parasitic diseases, outside the body. Gelatine, blood serum, agar, glycerine agar, and potato have been used for this purpose. After a few days on glycerine agar at the temperature of the blood little white shining colonies appear, which in319crease and coalesce. In about ten days' time the culture often turns a bright yellow, though it may remain white or even take on a brown or olive tint. The entire mass of growth is raised dry and crinkled, and composed almost exclusively of threads. In its early stage small bacillary forms occur, and in its later stage coccal forms. True clubs never occur in pure cultures, although the threads may occasionally show bulbous endings.
Glanders in the horse and ass, and sometimes by communication in man also, is caused by a short, non-motile, aërobic bacillus, named, after the old Roman nomenclature (malleus), Bacillus mallei. It was discovered in 1882 by Löffler and Schütz. It is found in the nasal discharge of glandered animals. In appearance the bacillus is not unlike B. tuberculosis, except that it is shorter and thicker. The beading of the bacillus of glanders, like that in tubercle, does not denote spores. B. mallei can be cultivated on the usual media, especially on glycerine agar and potato. On the latter medium it forms a very characteristic honey-like growth, which later becomes reddish-brown.
In the horse glanders particularly affects the nasal mucous membrane, forming nodules which degenerate and emit an offensive discharge. From the nose, or nasal septum, as a centre, the disease spreads to surrounding parts. It may also occur as nodules in and under the skin, when it is known as "farcy." Persons attending a glandered animal may contract the disease, often by direct inoculation.
Mallein is a substance analogous to tuberculin, and is made by growing a pure culture of Bacillus mallei in glycerine-veal broth in flat flasks, with free access of calcined air. After a month's growth the culture is sterilised, filtered, concentrated, and mixed with an equal volume of a .5 per cent. solution of carbolic. The dose is 1 cc., and it is used, like tuberculin, for diagnostic purposes. If the suspected animal reacts to the injection, it is suffering from glanders.320 Reaction is judged by three signs, namely, a rise of temperature 2–3° C., a large "soup-plate" swelling at the site of inoculation, and an enlargement of the lymphatic glands.
Swine fever, foot-and-mouth disease, chicken cholera, dysentery, rinderpest, and other diseases of animals have micro-organisms intimately related to them.
There is a group of diseases due to the presence in the blood or tissues of hæmatozoa, that is, protozoa which can live and perform their function in the blood. Amongst these are malaria, sleeping sickness, and other tropical diseases in man, and surra and various hæmatozoa in horses, fish, frogs, or rats.
Malaria. Although a Bacillus malariæ has been described as the cause of this disease, it is now almost universally supposed that the true cause is a protozoan parasite. In 1880 Laveran first described this organism, and the discovery was confirmed by Marchiafava, Celli, and others. Laveran claimed that it occurred in four different forms during the progress of its life-history:
(a) Spherical or Irregular Bodies attached to the blood corpuscle, or free in the blood plasma. They are a little smaller than the blood-cells, and may or may not contain pigment. They eventually invade the corpuscles, possess more pigment, and lose their amœboid movement. Within the red blood corpuscles they increase in size until they reach the adult stage.
(b) Segmentation Forms, often assuming a rosette shape, follow next. They are pigmented, are possibly a sporing stage, and are finally set free in the blood.
(c) The Crescents, or Semilunar Bodies, are free in the blood, but motionless. They are colourless, have a distinct membrane, and generally show a little pigment about the middle; they taper towards the poles. They appear in the blood after the fever has existed for some time, occurring321 chiefly, sometimes only, in the quotidian and malignant types of malaria.
(d) The Flagellated Bodies apparently occur only in the blood outside the body. They are extracorpuscular bodies, and possess several long flagella, and are therefore actively motile. They are derived from the crescents or irregular intracorpuscular bodies.
What is the precise significance of these various forms and modifications of them is not at present known. Possibly the semilune is a resting stage inside the body, and the flagellated body another similar stage outside. Attempts to cultivate the parasite outside the body have failed. There is a good deal of evidence to show that the mosquito is the host outside the human body. There may be different forms and varieties of parasite, if not actually different species, causing the diverse forms of clinical malaria.
The above account of diseases caused by bacteria does not profess to be in any sense exhaustive. It is merely illustrative. It reveals some of the disease-producing powers of micro-organisms. There are a large number of other diseases in which bacteria have been found. They are not the causes, but only accidentally present or associated with "secondary infection." Variola (small-pox), scarlet fever, and measles are excellent examples. It is possible that the danger at the present time is rather in the direction of supposing that every disease will readily yield its secret to the bacteriologist. Such, of course, is not the case. Nevertheless, as in the past, so in the future, constant research and patient investigation is the only hope we have for the elucidation of truth in respect to the causes of disease.
The object of modern bacteriology is not merely to accumulate tested facts of knowledge, nor only to learn the truth respecting the biology and life-history of bacteria. These are most important things from a scientific point of view. But they are also a means to an end; that end is the prevention of preventable diseases and the treatment of any departure from health. In a science not a quarter of a century old much has already been accomplished in this direction. The knowledge acquired of, and the secrets learned from, these tiny vegetable cells which have such potentiality for good or evil have been, in some degree, turned against them. When we know what favours their growth and vitality and virulence, we know something of the physical conditions which are inimical to their life; when we know how to grow them, we also know how to kill them.
We have previously made a cursory examination of the methods which are adopted for opposing bacteria and their products in the tissues and body fluids. We must now turn to consider shortly the modes which may be adopted in preventive medicine for opposing bacteria outside the body.
It will be clear at once that we may have varying degrees of opposition to bacteria. Some substances kill bacteria, and they are known as germicides; other substances prevent their development and resulting septic323 action, and these are termed antiseptics. The word disinfectant is used more or less indiscriminately to cover both these terms. A deodorant is, of course, a substance removing the odour of evil-smelling putrefactive processes. Here, then, we have the common designations of substances able to act injuriously on bacteria and their products outside, or upon the surface of, the body. But a moment's reflection will bring to our minds two facts not to be forgotten. In the first place, an antiseptic applied in very strong dose, or for an extended period, may act as a germicide; and, vice versâ, a germicide in too weak solution to act as such may perform only the function of an antiseptic. Moreover, the action of these disinfecting substances not only varies according to their own strength and mode of application, but it varies also according to the specific resistance of the protoplasm of the bacteria in question. Examples of the latter are abundant, and readers who have only assimilated the simple facts set forth in these pages are aware that between the bacillus of diphtheria and the spores of anthrax there is an enormous difference in power of resistance. In the second place, reflection will enable us to recall what has already been said, when discussing the requirements necessary for bacterial growth, respecting the physical conditions injurious to development. In a cold temperature, as a general rule, bacteria do not multiply with the same rapidity as at blood-heat. Within the limits of a moist perimeter the air is, to all intents and purposes, germ-free. Direct sunlight has a definitely germicidal effect in the course of time upon some of the most virulent bacteria we know. Here, then, are three examples of physical agents—low temperature, moist perimeter, sunlight—which, if strong enough in degree, or acting for a long enough period of time, become first antiseptics and then germicides. Yet for a limited period they have no injurious effect upon bacteria. These are simple points, and call for little comment, yet the pages324 of medical and sanitary journals reveal not a few keen controversies upon the injurious action of certain substances upon certain bacteria owing to the discrepancies, of necessity arising, between results of different skilled observers who have been carrying out different experiments with different solutions of the same substance upon different protoplasms of the same species of bacteria. We feel no doubt that in these pioneering researches much labour has been to some extent misspent, owing to the neglect of a common denominator. Only a more accurate knowledge of bacteria or a recognised standard for disinfecting experiments can ever supply such common denominator.
Species of bacteria for comparative observation-experiments upon the action of chemical or physical agents must be not only the same species, but cultured under the same conditions, and treated by the agent in the same manner, otherwise the results cannot be compared upon a common platform, or with any hope of arriving at exactly the same conclusions.
Sir George Buchanan laid down, in 1884, a very simple and suitable standard of what true disinfection meant, viz., the destruction of the most stable known infective matter. Such a test is high and difficult to attain unto; nevertheless, it is the only satisfactory one. Obviously many substances which are useful antiseptics in practical life would fall far short of such a standard, yet for true and complete disinfection such an ideal is the only adequate one.
Quite recently three or four workers at Leipzig101 have drawn up simple directions, the adoption of which would considerably assist in securing a common standard for disinfectant research. They are as follows:
1. In all comparative observations it is imperative that molecularly equivalent quantities of the reagents should be employed.
2. The bacteria serving as test objects should have equal power of resistance.
3. The numbers of bacteria used in comparative observation should be approximately equal.
4. The disinfecting solution must be always used at the same temperature in comparative experiments.
5. The bacteria must be brought into contact with the disinfectant with as little as possible of the nutrient material carried over. (This obviously will depend upon the object of the research.)
6. After having been exposed to the disinfectant for a fixed time, they should be freed from it as far as possible.
7. They should then be returned in equal numbers to the respective culture medium most favourable to the development of each, and kept at the same, preferably the optimum, temperature for their growth.
8. The number of surviving bacteria capable of giving rise to colonies in solid media must be estimated after the lapse of equal periods of time.
We may now turn from general principles to mention shortly some of the commoner methods and substances adopted to secure efficient disinfection. They are all divisible, according to Sir George Buchanan's standard, into two groups:
1. Heat in various forms;
2. Chemical bodies in various forms.
It should at the outset be understood that we desire in practical disinfection to inhibit or kill micro-organisms without injury to, or destruction of, the substance harbouring the germs for the time being. If this latter is of no moment, as in rags or carcasses, burning is the simplest and most thorough treatment. But with mattresses and beddings, bedclothes and garments, as well as with the human body, it is obvious that something short of burning is required.
1. From the earliest days of bacteriology heat has held a prominent place as a disinfector. But it is only in comparatively recent times that it has been fully established that moist heat is the only really efficient form of heat disinfection. Boiling at atmospheric pressure (100° C.) is the oldest form of moist heat disinfection, and because of the simplicity of its application it has gained a large degree of popularity. But it must not be forgotten that mere boiling (100° C.) may not effectually remove the spores of all bacilli. Besides, boiling is not applicable to furniture, mattresses, and such-like frequently infected objects. For many of these hot-air ovens were used in the early days. But it was found that such disinfection was no disinfection at all, for not only did it leave many organisms and spores untouched, but the degree of temperature was rarely, if ever, uniform throughout the substance being treated.
The failures following in the track of these methods were an indication of the need of some form of moist heat, viz., steam.
Here it will be necessary to digress for a moment into some of the characters of steam. When water is heated certain molecular changes take place, and at a certain temperature (100° C., 212° F.) the water becomes steam, or vapour, and on very little cooling will condense. But if the vapour is heated, it will become practically a gas, and will not condense until it has lost the whole of the heat, i. e., the heat of making water into vapour plus the heat of making vapour into gas. A gas proper is, then, the vapour of a liquid of which the boiling-point is substantially below its actual temperature. But we know that the temperature at which it boils depends upon the pressure to which it is subjected (Regnault's law). Hence in reality "steam at any temperature whatever may be a vapour proper, provided the pressure is such as prevents the liquid from boiling below that temperature." In such a condition327 of vapour it is termed saturated steam. But if it is at that same pressure further heated, it becomes practically a gas, and is called superheated steam. The former can condense without cooling; the latter cannot so condense at the same pressure. Saturated steam condenses immediately it meets the object to be disinfected, and gives out its latent heat; superheated steam acts by conduction, and not uniformly throughout the object. Its advantage is that it dries moistened objects. As a disinfecting power, superheated steam is much less than saturated steam. There is one further term which must be defined, namely, current steam. This is steam escaping from a disinfector as fast as it is admitted, and may be at atmospheric or higher temperatures. The disinfecting temperature which is now used as a standard is an exposure to saturated steam of 115° C. for fifteen minutes.
A number of different kinds of apparatus have been invented to facilitate disinfection to this standard on a large scale. Most sanitary authorities of importance are now supplied with some form of steam disinfector, though many are unable to go to the expense of high-pressure disinfectors. Professor Delépine has pointed out102 that a current of steam at low pressure may completely disinfect. Whilst such simple current-steam machines have thus been demonstrated as efficient bactericides, for all practical purposes it is important to have disinfectors capable of giving temperatures considerably above 100° C., of simple construction, having steam power of uniform temperature and rapid penetration, and containing, when in action, a minimum of superheated steam. In addition to these characters of a first-rate steam disinfector, two other important points should be borne in mind, namely, the air must be completely ejected from the disinfection chamber before the results due to steam are obtained, and some sort of automatic index giving a record of each disinfection is indispensable.
We may turn from these general principles to mention shortly some of the types of steam disinfectors most commonly in use. They are four, namely, the Washington Lyon, the Equifex (Defries), the Thresh, and the Reck.
Washington Lyon's apparatus consists of an elongated boiler having double walls, with a door at each end. The body of the apparatus is jacketed. The whole is large enough to admit of bedding and mattresses, and generally is so arranged that one end opens into one room, and the other end opens into another room. This convenient position admits of inserting infected articles from one room and receiving them disinfected into the other room. Possible reinfection is thereby prevented. Steam is admitted into the jacket at a pressure of between twenty and twenty-five pounds, and is generally twenty pounds in the interior of the cylinder. At the end of the operation a partial vacuum is created, by which means much of the moisture on the articles may be removed. In some cases a current of warm air is admitted before disinfection in order to diminish the extent of condensation.
The Equifex (Defries) contains no steam jacket, but coils of pipes are placed at the top and bottom of the apparatus, with the object of imparting to the steam as much heat as is lost by radiation through the walls of the disinfecting chamber, and at the same time of preventing undue condensation. The air is first removed by a preliminary current of steam, after which steam at a pressure of ten pounds is intermittently introduced and allowed to escape. The object of this proceeding is to remove air from the pores of the articles to be disinfected by the sudden expansion of the film of water previously condensed on their surface. The apparatus introduced by Dr. Thresh was constructed with a view of overcoming the objection to some of the other machines that bulky articles retained a large percentage of moisture, thus necessitating the use of some additional329 drying apparatus. A central chamber receives the articles to be disinfected, and is surrounded by a boiler containing a solution of calcium chloride at a temperature of 225° F. This is heated by a small furnace, and the steam given off (218–300° F.) is conducted into the central chamber. The steam is not confined under any pressure except that of the atmosphere. When the steam has passed for a sufficient length of time, it is readily diverted into the open air. Hot air is now introduced, and at the expiration of an hour the articles may be taken out disinfected and as dry as they were when inserted. The apparatus is comparatively inexpensive, and not of a complicated nature. The current steam is saturated, and at a temperature a few degrees above the boiling-point. Many experiments have been performed with this apparatus, and there is now a large amount of evidence in favour of it and current steam disinfection.
Reck's apparatus is another kind of saturated steam disinfector, which resembles the Equifex, but differs from it in employing steam as a current.
It is probable that many other forms of steam disinfector will be invented, and each will have its enthusiastic supporters. Even at the time of writing some excellent results are announced from America.
2. The effects of chemical substances as solutions, or in spray form, upon bacteria have been observed from the earliest days of bacteriology. To some decomposing matter or solution a disinfectant was added and sub-cultures made. If bacteria continued to develop, the disinfection had not been efficient; if, on the other hand, the sub-culture remained sterile, disinfection had been complete. From such rough-and-ready methods large deductions were drawn, and it is hardly too much to say that no branch of bacteriology contains such a vast mass of unassimilated and unassimilable statements as that relating to research into disinfectants. Most of the tabulated and recorded results are conspicuous330 in having no standard as regards bacterial growth. Yet without such a standard results are not comparable.
Silk threads, impregnated with anthrax spores, were placed in bottles containing carbolic acid of various strengths, and at stated periods threads were removed and placed in nutrient media, and development or otherwise observed. But, as Professor Crookshank103 has pointed out, this method is fallacious, the thread being still wet with the solution when transferred to the medium, and thus modified in culture, possibly even inhibited altogether. It is unnecessary for us here to discuss every mode adopted by investigators in similar researches. We may just mention that the most approved methods at the present time are based upon two simple plans of exposure. In one we use a known volume of recent broth culture of an organism grown under specified conditions. To this is added a measured quantity of the antiseptic. At stated periods loopfuls of the broth and antiseptic mixture are sub-cultured in fresh-sterilised broth, and resulting development or otherwise closely observed. The other method is practicable when we are dealing with volatile bodies. In such cases a standard culture is made of the organism in broth at a standard temperature. Into this are dipped small strips of sterilised linen. When thoroughly impregnated these are removed from the broth and subsequently dried over sulphuric acid in a vacuum at 38° C. These may now be exposed for a longer or shorter period to the fumes of the antiseptic in question, and broth cultures made at the end of the exposure. It is obvious that a very large number of modifications are possible of these two simple devices for testing the bactericidal power of chemical substances. It should be remembered that here, perhaps, more than anywhere else in bacteriological research, careful control experiments are absolutely necessary.
Mineral acids (nitric, hydrochloric, sulphuric), especially concentrated, are all germicides.
The halogens—chlorine, bromine, iodine, and fluorine—are, all four, disinfectants, but not used in practice. They are named in their order of power as such.
A number of separate bodies, such as chloroform and iodoform, have been much advocated as antiseptics. The cost of the former and odour of the latter have, however, greatly militated against their general adoption.
Chloride of lime is a powerful disinfectant. Professor Sheridan Delépine and Dr. Arthur Ransome have demonstrated its germicidal effect as a solution applied directly to the walls of rooms inhabited by tuberculous patients.104 It may also be used in solid form for dusting decomposing matter.
Mercuric chloride (corrosive sublimate) has been an accepted germicide for some time. But the experiments of Behring, Crookshank, and others have proved that the weaker solutions cannot be relied upon. This is, in part, due to the fact that it forms in albuminous liquids an albuminate of mercury which is inactive. Dilute solutions have the further disadvantage of being unstable. Various authorities recommend a solution of 1–500 as a germicide, and much weaker solutions are, of course, antiseptic. An ounce each of corrosive sublimate and hydrochloric acid in three gallons of water makes an efficient disinfectant.
Potassium permanganate is, of course, the chief substance in Condy's fluid, as zinc chloride is in Burnett's disinfecting fluid. A 5 per cent. of the former and a 2-1/2 per cent. of the latter are germicidal.
Boracic acid is one of the most useful antiseptics with which to wash sore eyes, or preserve tinned foods or milk. It is not a strong germicide, but an unirritating and effective wash. Many cases of its addition to milk have found their 332 way into the law courts, owing to cumulative poisoning, and it should only be used with the very greatest care as a food preservative.
Carbolic acid has come into prominence as an antiseptic since its adoption by Lister in antiseptic surgery. It is cheap, volatile, and effective. One part in 400 is antiseptic, and 1 in 20 germicidal. As a wash for the hands the former is used, and a weaker solution for the body generally. Carbolic soap and similar toilet combinations are now very common. At one time it appeared as if corrosive sublimate would oust carbolic from the first place as an antiseptic solution, but a large number of experiments have confirmed opinion in favour of carbolic. Professor Crookshank found that carbolic acid, 1 in 40, acting for only one minute is sufficient to destroy Streptococcus pyogenes, S. erysipelatis, and Staphylococcus pyogenes aureus, and in the strength of 1 in 20 carbolic acid completely sterilised tubercular sputum when shaken up with it for one minute.
Creosol, a member of the phenol series, is a good disinfectant, and the active element in lysol, Jeye's fluid, creoline, izal, and creosote.
Sulphurous acid is one of the commonest disinfectants employed for fumigation—the old orthodox method of disinfecting a room in which a case of infective disease has been nursed. It is evolved, of course, by burning sulphur. For each thousand cubic feet from one to five pounds of sulphur is used, and the walls may be washed with carbolic acid. Dr. Kenwood carried out some experiments in 1896105 which appear to support the disinfecting power of sulphur fumes. He found that the Bacillus diphtheriæ was not killed, though markedly inhibited, when the sulphurous gas (SO2) did not much exceed .25 per cent. But the bacillus was killed where the sulphur fumes exceeded .5 per cent. Both these results had reference to the
SO2 in the air in the centre of the room at a height of four feet, and after the lapse of four hours. There can be little doubt that fuming a sealed-up room with sulphur fumes in a moist atmosphere, and leaving it thus for twenty-four hours, is generally, if not always, efficient disinfection. It will kill the bacillus of diphtheria, though not always more resistant germs. Moreover, its simplicity of adoption is greatly in its favour. Anyone can readily apply it by purchasing a few pounds weight of ordinary roll sulphur and burning this in a saucer in the middle of a room which has had all its crevices and cracks in windows and walls blocked up with pasted paper. Nitrous fumes may also be used in this way.
Recently formalin has come much in favour as a room disinfectant. Formalin is a 40 per cent. solution of formaldehyde in water, a gas discovered by Hofmann in 1869. This gas is a product of imperfect oxidation of methyl alcohol, and may be obtained by passing vapour of methyl alcohol, mixed with air, over a glowing platinum wire or other heated metals, such as copper and silver. It is the simplest of a series of aldehydes, the highest of which is palmitic aldehyde. Its formula is CH2O, and it is a colourless gas with a pungent odour, and having penetrating and irritating properties, particularly affecting the nasal mucous membrane and the eyes of those working with it. It is readily soluble in water, and in the air oxidises into formic acid (CH2O2). This latter substance occurs in the stings of bees, wasps, nettles, and various poisonous animal secretions. Formalin is a strong bactericide even in dilute solutions, and, of course, volatile. A solution of 1 to 10,000 is said to be able to destroy the bacilli of typhoid, cholera, and anthrax. A teaspoonful to ten gallons of milk is said to retard souring. When formalin is evaporated down, a white residue is left known as paraform. In lozenge form this latter body is used by combustion of methylated334 spirit to produce the gas. Hence we have three common forms of the same thing—formalin, formic aldehyde, paraform—each of which yields formic acid, and thus disinfects. The vapour cannot in practice be generated from the formalin as readily as from the paraform.
By a variety of ingenious arrangements formic aldehyde has been tested by a large number of observers during the last two or three years. We may refer to three modes of application. 1. The sprayer (Equifex apparatus) produces a mixture of air and solution for spraying walls, ceilings, floors, and sometimes garments. 2. The autoclave (Trillat's apparatus). In this a mixture of a 30–40 per cent. watery solution of formaldehyde and calcium chloride (4–5 per cent.) is heated under a pressure of three or four atmospheres, and the almost pure, dry gas is conducted through a tube passing through the keyhole of the door into the sealed-up room. 3. The paraform lamp (the Alformant). The principle of this lamp is that the hot, moist products from the combustion of methylated spirit act upon the paraform tablets, converting them into gas. Most of the conclusions derived from experiments with these three different forms of apparatus are the same. It is agreed that the gas is harmless to colours and metal and polished wood. The vapour acts best in a warm atmosphere. As for its action on bacteria, it compares favourably with any other disinfectant. In 1 per cent. solution formalin destroys non-spore-bearing bacteria in thirty to sixty minutes.
Many observers have decried formaldehyde on account of its professed lack of penetrating power. Professor Delépine, however, states106 that it possesses "penetration powers probably greater than those of most other active gaseous disinfectants. Bacillus coli, B. tuberculosis, B. pyocyaneus, and Staph. pyogenes aureus were killed in dry or moist state, even when protected by three layers of filter paper." In
Professor Delépine's opinion, the vapours of phenol, izal, dry chlorine, and sulphurous acid have, under the same conditions, given inferior results.
We may now shortly summarise the foregoing facts respecting antiseptics and disinfection in the simplest terms possible to afford facility to the uninitiated in practical application:
To disinfect a room, seal up cracks and crevices, and burn at least one pound of roll sulphur for every 1,000 cubic feet of space.107 Many authorities recommend four or five pounds of sulphur to the same space. Let the room remain sealed up for twenty-four hours.
To disinfect walls, wash with chloride of lime solution (1–100) or carbolic acid (1–40). This latter solution may be used to wipe down furniture. Either or both may be used after sulphur fuming. Formic aldehyde may also be used by lamp or autoclave.
To disinfect bedding, etc., the steam sterilisation secured in a Thresh, Equifex, or Lyon apparatus is the best. Rags and infected clothing, unless valuable, should be burnt.
To disinfect garments and wearing apparel, they should be washed in a disinfectant solution, or fumed with formic aldehyde.
To disinfect excreta or putrefying solutions, enough disinfectant should be added to produce in the solution or matter being disinfected the percentage of disinfectant necessary to act as such. Adding a small quantity of antiseptic to a large volume of fluid or solid is as useless as pouring a small quantity of antiseptic down a sewer with the idea that such treatment will disinfect the sewage. The mixture of the disinfectant with the matter to be disinfected must contain the standard percentage for disinfection. Chloride of lime is a common substance for use in this way. Potassium 336permanganate (1–100) and carbolic (1–100), and many manufactured bodies containing them, are also widely used. Drs. Hill and Abram recommend108 that the excreta and disinfectant be thoroughly mixed and stand for at least half an hour. For various reasons they particularly advise chinosol as the most convenient disinfectant for this specific purpose.
Antiseptics for wounds. Carbolic acid (1–40) or corrosive sublimate (1–1,000) are commonly used in surgical practice. Boracic acid is one of the most unirritating antiseptics which are known. It may be used in saturated watery solution (1–30) or dusted on copiously as fine powder. It is especially applicable in open wounds, and as an eye-wash.
To disinfect hands and arms. Operating surgeons are those to whom it is a most urgent necessity to cleanse hands and arms antiseptically. Carbolic acid (1–20, or 1–40) is used for this purpose.
It is hardly necessary to add that in a case of infectious disease occurring in a household many of these modes of application, perhaps all of them, must be adopted. Formalin is probably the best gaseous disinfectant which we have, but its use does not, and should not, preclude the simultaneous adoption of other methods.
It is proposed to add one or two notes on certain technical points in bacteriological work, with a view to assisting those medical men not able to obtain the advantages of a well-equipped laboratory, and yet desirous of occasionally attempting some practical bacteriology.
1. General Examination. All fluids may be examined for bacteria in two chief ways:
(a) A small quantity may be placed on a cover-glass or slide, dried over a lamp or bunsen flame, and stained with aniline dyes for a few minutes. It is then ready for microscopic examination. It is obvious that the result will generally be a mixture of bacteria, for which differentiating stains may be used (Gram, Ziehl-Neelsen, etc.).
(b) A minute drop of the suspected fluid may be added to various fluid media (broth, liquefied gelatine, etc.) and then plated out upon small sterilised sheets of glass. In the course of two or three days the contained bacteria will reveal themselves in characteristic colonies, which may be examined, and if possible sub-cultured, and carefully studied.
Double-Staining Methods. These are various, and are used when it is desired to stain the bacteria themselves one colour, and the matrix or ground substance in which they are situated another colour. Three of the commoner methods are those of Ehrlich, Neelsen, and Gram. They are as follows:
Ehrlich's Method. "Five parts of aniline oil are shaken up with 100 parts distilled water, and the emulsion filtered through moistened filter paper. A saturated alcoholic solution of fuchsine, methyl-violet, or gentian-violet is added to the filtrate in a watch-glass,338 drop by drop, until precipitation commences. Cover-glass preparations are floated in this mixture for fifteen to thirty minutes, then washed for a few seconds in dilute nitric acid (one part nitric acid to two of water), and then rinsed in distilled water. The stain is removed from everything except the bacilli; but the ground substance can be after-stained brown if the bacilli are violet, or blue if they have been stained red" (Crookshank, Bacteriology and Infective Diseases, p. 89).
Gram's Method. The primary stain in this method is a solution of aniline gentian-violet (saturated alcoholic solution of gentian-violet 30 cc., aniline water 100 cc.), which stains both ground substance and bacteria in purple. The preparation is next immersed in the following solution for half a minute or a little more:
| Iodine | 1 part |
| Potassium iodide | 2 parts |
| Distilled water | 300 parts |
In this short space of time the iodine solution acts as a mordant of the purple colour in the bacteria, but not in the ground substance. Hence, if the preparation be now (when it has assumed a brown colour) washed in alcohol (methylated spirit), the ground substance slowly loses its colour and becomes clear. But the bacteria retain their colour, and thus stand out in a well-defined manner. Cover-glass preparations decolourise more quickly than sections of hardened tissue, and they should only be left in the methylated spirit until no more colour comes away. The preparation may now be washed in water, dried, and mounted for microscopic examination, or it may be double-stained, that is, immersed in some contrast colour which will lightly stain the ground substance. Eosin or Bismarck brown are commonly used for this purpose. The former is applied for a minute or two, the latter for five minutes, after which the specimen is passed through methylated spirit (and preferably xylol also) and mounted. The result is that the bacteria appear in a dark purple colour on a background of faint pink or brown. Carbol-thionine blue, picro-carmine, and other stains are occa339sionally used in place of the aniline gentian-violet, and there are other slight modifications of the method.
Ziehl-Neelsen Method. Here the primary stain is a solution of carbol-fuchsin:
| Fuchsin | 1 part |
| Absolute alcohol | 10 parts |
| 5 per cent. aqueous solution of carbolic acid | 100 parts |
It is best to heat the dye in a sand-bath, in order to distribute the heat evenly. The various stages in the staining process are as follows: (a) The cover-glass with the dried film upon it is immersed in the hot stain for one to three minutes. (b) Remove the cover-glass from the carbol-fuchsin, and place it in a capsule containing a 25 per cent. solution of sulphuric acid to decolourise it. Here its redness is changed into a slate-grey colour. (c) Wash in water, and alternately in the acid and water, until it is of a faint pink colour. (d) Now place the cover-glass for a minute or two in a saturated aqueous solution of methylene-blue, which will counter-stain the decolourised ground substance blue. (e) Wash in water. (f) Dehydrate by rinsing in methylated spirit, dry, and mount. A pure culture of bacteria will not necessarily require the counter-stain (methylene-blue). Sections of tissue may require twenty to thirty minutes in the primary stain (carbol-fuchsin). This stain is used for tubercle and leprosy. With a little practice the staining of the bacillus of tubercle when present in pus or sputum becomes a very simple and accurate method of diagnosis. A small particle of sputum or pus is placed between two clean cover-glasses and thus pressed between the thumb and finger into a thin film. This is readily dried and stained as above, the bacillus of tubercle appearing as a delicately-beaded red rod with a background of blue.
Bacteriological Diagnosis. The following points must be ascertained in order to identify any particular micro-organism:
(1) Its morphology, bacillus, coccus, spirillum, etc.; the presence or absence of involution forms.
(2) Motility by the unstained cover-glass preparation ("hanging drop"); note presence of flagella.
(3) Presence of spores, their appearance and position.
(4) Whether or not the organism stains with Gram's method.
(5) The character of the growth upon various media (gelatine, agar, milk, potato, broth); the presence or absence of liquefaction in the gelatine culture; its power of producing acid, gas, or indol.
(6) Whether it is aërobic or anaërobic.
(7) Its colour in cultivation.
(8) If it is a disease-producing organism under examination, its effect upon the animal tissues and the course of the disease should be observed.
There are other points of importance, but the above are essential to a right conclusion.
Diagnosis in Special Diseases:
(1) Diphtheria. This disease may be bacteriologically diagnosed with a minimum of apparatus and equipment. By means of a swab a rubbing from a suspected throat is readily obtained. This may be examined by the microscope, or sub-cultured on favourable medium. Blood serum is perhaps the best, but, as Hewlett remarks, "If no serum tubes can be had, an egg may be used. It is boiled hard, the shell chipped away from one end with a knife sterilised by heating, and the inoculation made on the exposed white surface; the egg is then placed, inoculated end down, in a wine-glass of such a size that it rests on the rim and does not touch the bottom. A few drops of water may with advantage be put at the bottom of the glass to keep the egg-white moist. The preparation is kept in a warm place for twenty-four to forty-eight hours and then examined." The examination, of course, consists in staining and preparing for the microscope and observing the form, arrangement, and characters of the organism or organisms present. A small piece of the membrane may be detached, washed in water, and stained for the bacilli.
(2) Tubercle (Ziehl-Neelsen's stain, vide supra).
(3) Typhoid (Enteric Fever).
Widal's Reaction. This diagnostic test depends upon the effect which the blood of a person suffering from typhoid fever has upon the Bacillus typhosus. The effect is twofold. In the first341 place, the actively motile B. typhosus becomes immotile; and secondly, there is an agglutination, or grouping together in colonies, of the B. typhosus. Neither of these features occurs if healthy human blood is brought into contact with a culture of the typhoid bacillus. There are various ways in which this "serum diagnosis" can be carried out. The simplest and quickest method is as follows: To ten drops of a twenty-four or forty-eight-hours-old neutral broth culture of the typhoid bacillus one drop of the blood serum to be tested is added. The serum and culture are rapidly mixed in the trough of a hollow ground slide (such as is used for the "hanging drop"), and a single drop is taken, placed upon an ordinary clean slide, and a cover-glass superimposed. The positive reaction of agglutination and immotility, if the blood comes from a case of typhoid fever, will probably appear within fifteen or twenty minutes. The fluid culture of typhoid may be taken from an agar culture as well as from broth. In both cases it may be desirable to filter through ordinary filter paper to remove any normally agglutinated masses of bacilli before commencing the test.
In his first experiments Widal used a test-tube in the following manner: The blood to be tested is diluted by one part of it being added to fifteen parts of broth in a test-tube. The mixture is inoculated with a drop of a typical Bacillus typhosus culture. The tube is then incubated at 37° C. for twenty-four hours, after which it is examined. If the reaction be positive, the broth appears comparatively clear, but at the bottom of the test-tube a more or less abundant sediment will be found. This is due to the clumps of bacilli having fallen owing to gravity. If, on the other hand, the reaction is negative, the broth will appear more or less uniformly turbid.
For the apparatus required to carry out the simpler methods of bacteriological work reference should be made to the standard laboratory text-books, which furnish all necessary details. A good microscope, with a 1/12 oil immersion lens, is, of course, essential. This can now be obtained for about £16 (Beck, Swift, Baker, Watson, etc.), and the other necessary apparatus is readily obtainable of Baird and Tatlock, Hatton Garden, E. C., and other makers.
1 The Contemporary Review, November, 1897, p. 719.
2 Some notable exceptions are found in the work of the Bath and West of England Society, Lord Vernon's model dairy, and the Essex County Council Bacteriological Teaching Laboratory.
3 We propose throughout to use the term bacterium (pl. bacteria) in its generic meaning, unless especially stated to the contrary. It will also be synonymous with the terms microbe, germ, and micro-organism. The term bacillus will, of course, be restricted to a rod-shaped bacterium.
4 Migula has recently (1896) suggested that the Schizomycetes should be subdivided into Coccaceæ, Bacteriaceæ, Spirillaceæ (spirilla, spirochæta), Chlamydobacteriaceæ (Streptothrix, Crenothrix, Cladothrix), and Beggiatoa.
5 A one-twelfth oil immersion lens is requisite for the study of the lower bacteria.
6 A flagellum is a hair-like process arising from the poles or sides of the bacillus. It must not be confused with a filament, which is a thread-like growth of the bacillus itself.
7 A "pure culture" is a growth in an artificial medium outside the body of one species of micro-organism only.
8 Some pathogenic germs (suppuration and typhoid) can withstand freezing for weeks.
9 G. J. Romanes, Darwin and After Darwin, vol. ii., 231.
10 It will be observed that there is a marked difference between the effects of dry heat and moist heat. Moist heat is able to kill organisms much more readily than dry, owing to its penetrating effect on the capsule of the bacillus. Dry heat at 140° C. (284° F.), maintained for three hours, is necessary to kill the resistant spores of Bacillus anthracis and B. subtilis, but moist heat at fifty degrees less will have the same effect. It is from data such as these that in laboratories and in disinfecting apparatus moist heat is invariably preferred to dry heat. For with the latter such high temperatures would be required that they would damage the articles being disinfected. Koch states the following figures for general guidance: Dry heat at a temperature of 120° C. (248° F.) will destroy spores of mould fungi, micrococci, and bacilli in the absence of their spores; for the spores of bacilli 140° C. (284° F.), maintained for three hours, is necessary; moist heat at 100° C. (212° F.) for fifteen minutes will kill bacilli and their spores.
11 Water from a house cistern is rarely a fair sample. It should be taken from the main. If taken from a stream or still water, the collecting bottle should be held about a foot below the surface before the stopper is removed.
12 The cubic centimetre (cc.) is a convenient standard of fluid measurement constantly recurring in bacteriology. It is equal to 16–20 drops, and 28 cc. equal one fluid ounce.
13 The gelatine is reduced to liquid form by heating in a water-bath. Before inserting the suspected water it is essential that the gelatine be under 40° C, or thereabouts, in order not to approach the thermal death-point of any bacteria.
14 Micro-organisms in Water (1894).
15 Report on the Micro-organisms of Sewage, Reports to L. C. C., 1894, No. 216.
16 Harben Lectures, 1896.
17 Report on the Metropolitan Water Supply.
18 The methods adopted for making a quantitative and qualitative examination of sewage are precisely analogous to those used in milk research. Dilution with sterilised water previous to plating out on gelatine in Petri dishes is essential (1 cc. to 10,000 cc. of sterile water, or some equally considerable dilution), otherwise the large numbers of germs would rapidly liquefy and destroy the film. Special methods must be used for the isolation of special organisms; phenol-gelatine, Elsner medium, indol reaction, "shake" cultures, Parietti broth, etc., must often be resorted to for special bacteria. Spores of bacteria may always be numerically estimated by adding the suspected water or sewage to gelatine, and then heating to 80° C. for ten minutes before plating out. This temperature removes the bacilli, but leaves the spores untouched.
19 The bacilli of typhoid can live in crude sewage (Klein), but only for a very short period. When sewage is diluted with large quantities of water the case is very different. Bacillus coli flourishes in sewage.
20 Annual Report of the Medical Officer of the Local Government Board, 1897–98, p. 210.
21 John Tyndall, F.R.S., Floating Matter of the Air.
22 Flügge has lately attempted to demonstrate that an air current having a velocity of four metres per second can remove bacteria from surfaces of liquids by detaching drops of the liquid itself.
23 Hewlett and Thomson graphically demonstrated the bactericidal power of the nasal mucous membrane by noting the early removal of Bacillus prodigiosus, which had been purposely placed on the healthy Schneiderian membrane of the nose.
24 Pathological Society of London, Transactions, 1897.
25 Annali d'Igiene Sperimentale, vol. v. (1895), fasc. 4.
26 Public Health, vol. x., No. 4, p. 130 (1898).
27 Flügge, Grundriss der Hygiene, 1897.
28 Zeitschrift für Hygiene, vols. xxiv.-xxvi.
29 Annales de Micrographie.
30 E. A. Schäfer, F.R.S., Text-book on Physiology, vol. i., p. 312.
31 The unorganised ferments are frequently otherwise classified than as above, not according to the locality, but according to the function. The chief are these:—amylolytic, those which change starch and glycogen (amyloses) into sugars, e. g., ptyalin, diastase, amylopsin; proteolytic, those which change proteids into proteosis and peptones, e. g., trypsin, pepsin; inversive, those which change maltose, sucrose, and lactose into glucose, e. g., invertin; coagulative, those which change soluble proteids into insoluble, e. g., rennet; steatolytic, those which split up fats into fatty acids and glycerine, e. g., steapsin.
32 A chemical change obtained by the action of sulphuric or some other acid, or by the influence of diastase.
33 Bacteriology and Infective Diseases, Appendix.
34 E. C. Hansen, Studies in Fermentation (Copenhagen), p. 98.
35 Proc. Royal Soc. of Edin., xxxvii., pt. iv., p. 759.
36 E. A. Schäfer, Text-book of Physiology, vol. i., p. 25 (W. D. Halliburton).
37 "Denitrifying" means reducing nitrates.
38 R. Warington, M.A., F.R.S., Journ. Roy. Agricultural Soc. Eng., series iii., vol. viii., pt. iv., pp. 577 et seq.
39 The saltpetre beds of Chili and Peru are an excellent example of the industrial application of these facts. Nitrates are there produced from the fæcal evacuations of sea-fowl in such quantities as to form an article of commerce. A like form of utilisation of the action of these bacteria was once practiced on the continent of Europe. Economic application is also seen in the treatment of sewage referred to elsewhere.
40 The course of nitrification may be followed by means of chemical tests. 1. The disappearance of ammonia. 2. The appearance of nitrite. 3. Its disappearance. 4. Appearance of nitrate.
41 Professor Warington, in Report IV. (p. 526) of his admirable series of papers on the subject, draws attention to Müntz's criticism that the nitrifying organisms only oxidise from nitrogenous matter to nitrites, and not from nitrites to nitrates. Müntz held that the conversion of nitrite into nitrate is brought about by the joint action of carbonic acid and oxygen. Professor Warington's experiments, however, clearly illustrate that the production of nitrates from nitrites in an ammoniacal solution can be determined by the character of the bacterial culture with which the solution is seeded, and that in a solution of potassium nitrite conversion into nitrate can be determined by the introduction of the nitric organism. Professor Warington still adheres to the opinion, in favour of which he has produced so much evidence, that the formation of nitrates in the soil is due to the nitric organism which soil always contains.
42 British Association for the Advancement of Science, Bristol, 1898, Presidential Address.
43 British Association for the Advancement of Science, Bristol, 1898, Presidential Address.
44 Sir John Lawes and Sir Henry Gilbert (Times, December 2, 1898), have pointed out that the addition of nitrates only would be of no permanent use to the wheat crop. They rely upon thorough tillage and proper rotation of crops as the means of improving the nitrogen value of the soil.
45 Geddes, Nature, xxv., 1882.
46 Sir Henry Gilbert, F.R.S., The Lawes Agricultural Trust Lectures, 1893, p. 129.
47 Ibid., p. 140.
48 This has been denied recently in the official report by the chemist of the Experimental Farm to the Minister of Agriculture at Ottawa (Report, 1896, p. 200).
49 It has already been pointed out that the nitrifying bacteria, though able to live on organic matter, do not require such either for existence or for the performance of their function.
50 Lehmann and Neumann, p. 305.
51 The conditions requisite for an outbreak of enteric fever were, according to Pettenkofer, (a) a rapid fall (after a rise) in the ground water, (b) pollution of the soil with animal impurities, (c) a certain earth temperature, and lastly (d) a specific micro-organism in the soil. These four conditions have not, particularly in England, always been fulfilled preparatory to an epidemic of typhoid. Yet the observations necessary for these deductions were a definite step in advance of mere dampness of soil.
52 Supplement to the Report of the Medical Officer of the Local Government Board, 1887, p. 7.
53 Report of Medical Officer to Local Government Board, 1895–1896, Appendix.
54 H. L. Russell, Dairy Bacteriology, p. 46.
55 Bureau of Animal Industry Reports, 1895–1896.
56 British Medical Journal, 1895, vol. ii., p. 322.
57 British Medical Journal, 1895, vol. ii., p. 322.
58 Journal of Comparative Pathology, vol. x. (1897), pp. 150–189.
59 E. W. Hope, M.D., D.Sc., Report of the Health of Liverpool during 1897, p. 40.
60 S. Rideal and A. G. R. Foulerton conclude, from a series of experiments, that boric acid (1–2,000) and formaldehyde (1–50,000) are effective preservatives for milk for a period of twenty-four hours, and that these quantities have no appreciable effect upon digestion or the digestibility of foods preserved by them (Public Health, May, 1899, pp. 554–568).
61 Report from Wisconsin Agricultural Experiment Station, 1896.
62 Jenner Institute of Preventive Medicine (First Series Transactions).
63 Centralblatt für Bakteriologie, etc., II. Abteilung.
64 A Manual of Bacteriology, Clinical and Applied, p. 397.
65 Hewlett asserts that butter may contain from two to forty-seven millions of bacteria per gram.
66 Such pure cultures for such purposes are in the United States termed "starters," because they start the process of special ripening. For the sake of convenience the term will be used here.
67 The Essex County Council is one of the few public bodies in England which have undertaken pioneer work in this department of industry. Under the leadership of Mr. David Houston, a course of elementary instruction in dairy bacteriology as applied to modern dairy practice is given in the County Biological Laboratory at Chelmsford.
68 Report of Storr's Agricultural Experiments Station, State of Connecticut, 1895.
69 "Observations on Cheddar Cheese Making," Reports of Bath and West and Southern Counties Society, 1898, pp. 163–171. Mr. Lloyd's Reports to the West of England Society since 1892 contain various points respecting the application of bacteriology to cheese-making.
70 Journal of Bath and West of England Society, 1893, 1895, and 1897.
71 New York Medical Record, 1894.
72 British Medical Journal, 1896, ii., p. 760 et seq.
73 Special Report of the Medical Officer to the Local Government Board on Oyster Culture, etc., 1896.
74 Royal Commission on Tuberculosis, Report, 1895, pt. i., p. 13.
75 Ibid., p. 18.
76 British Medical Journal, 1895, vol. ii., p. 513.
77 It should be distinctly understood that this table is merely schematic and provisional. The details of toxin production and its effect are still open to revision and amendment.
78 Sidney Martin, M.D., F.R.S., F.R.C.P., Croonian Lectures delivered before the Royal College of Physicians, June, 1898.
79 It is impossible here to enter into a detailed consideration of the various views held with regard to the formation of antitoxins. It is needless to remark that the whole matter is one of abstruse technicality and intricacy. These antitoxic bodies gradually increase in the blood and tissues, and their action falls into two groups: (a) antitoxic, which counteract the effects of the poison itself; and (b) antimicrobic, which counteract the effects of the bacillus itself. "In one and the same animal the blood may contain a substance or substances which are both antitoxic and antimicrobic, such, for example, as occurs in the process of the formation of the diphtheria and tetanus antitoxic serums" (Sidney Martin).
80 Types of bodies possessing positive chemiotaxis for bacteria are the salts of potassium, peptone, glycerine.
81 Negative chemiotaxis is illustrated in alcohol, and free acids, and alkalies.
82 The friend of Addison and Pope, who married Mr. Edward Wortley Montagu in 1712, and on his appointment to the ambassadorship of the Porte in 1716 went with him to Constantinople. They remained abroad for two years, during which time Lady Wortley Montagu wrote her well-known Letters to her sister the Countess of Mar, Pope, and others.
83 Crookshank, History and Pathology of Vaccination.
84 An exhaustive account of vaccine may be found in the Milroy lectures delivered in 1898 at the Royal College of Physicians by S. Monckton Copeman, M.D.
85 Crookshank, Bacteriology and Infective Diseases; Virchow, The Huxley Lecture, 1898.
86 To shorten this period Dr. Cartwright Wood has adopted a plan by which time may be saved, and 200 cc. injected say within the first two or three weeks. This is accomplished by using a "serum toxin" (containing albumoses, but not ferments) previously to the broth toxin, an ingenious method which we cannot enter into here.
87 At the conclusion of the operation the cannula is removed from the jugular vein, and the wound is closed by the valvular character of the slit in the skin and vein and the elasticity of the wall of the vein. No stitching or dressing is required. Indeed, it is striking to observe in the horse an entire absence of pain throughout the proceedings.
88 The term unit is used as a standard measurement. This means the amount of antitoxin which will just neutralise ten times the minimum fatal dose of the toxin in a guinea-pig (250 grams toxin to kill on the fourth day). If 1 cc. of the antitoxic serum is required for this, one unit is contained in 1 cc.; if 0.01 cc. is sufficient, then 100 units are contained in the cc. Not less than 1500 units should be administered for a dose, and repeated every twelve hours. In severe cases two or three times this amount may be given.
89 The value of antitoxin treatment in diphtheria is discussed in the Brit. Med. Jour., 1899, pp. 197 and 268, by E. W. Goodall, M.D.
90 A detailed study of tuberculosis from its pathological and bacteriological aspect will be found in La Tuberculose et son Bacille, pt. i., Straus, Professeur à la Faculté de Médecine de Paris.
91 For differences of virulence between these conditions of pulmonary tubercle see Lingard, Local Government Board Report, 1888, p. 462.
92 Centralblatt. f. Bact. und Parasit., vol. vii., p. 9.
93 Animal Tuberculosis, p. 129.
94 See the Harben Lectures, November, 1898, by Sir Richard Thorne Thorne, Medical Officer to the Local Government Board; also the Report of the Royal Commission on Tuberculosis, 1896–98.
95 1. Tuberculosis is a disease mainly affecting the lungs (consumption, decline, phthisis) of young adults and the bowels of infants (tabes mesenterica). It may affect any part of the body, and its manifestations are very various. It also affects animals, particularly cattle, by whom it may be transmitted to man.
2. Its direct cause is a microscopic vegetable cell, known as the Bacillus tuberculosis, discovered by Koch in 1882. This fungus requires to be magnified some hundreds of times before it can even be seen. When it gains entrance to the weakened body it sets up the disease, which is an infectious disease, though different in degree to the infectiousness of, say, measles.
3. Trade influence and occupation, in some cases, undoubtedly predispose the individual to tubercle. Cramped attitudes, exposure to dampness or cold, ill ventilation, and exposure to inhalation of dust of various kinds, all act in this way. In support of the evil effect of each of these four conditions much evidence could he produced.
4. Overcrowding has a definite influence in propagating tubercular diseases. The agricultural counties without big towns, like Worcestershire, Herefordshire, Buckinghamshire, and Rutland, are the counties having the lowest mortality from tuberculosis; whilst the crowded populations in Northumberland, South Wales, Lancashire, London, and the West Riding suffer most. Speaking more particularly, the overcrowded areas of London, such as St. Giles', Strand, Holborn, and Central London generally, show very high tubercular death-rates.
5. Tuberculosis is not increasing. During the last thirty years it has shown, with few exceptions, a steady decline in all parts of England. "Consumption" is most fatal in comparatively young people (fifteen to forty-five years), whilst "tabes" and other forms of tubercle are fatal chiefly to young children. These forms have not declined so much as the lung form. The mortality in consumption of males has since 1866 been in excess of that of females. The age of maximum fatality from consumption is later than in the past, which is probably due to improved hygiene and treatment.
6. This decline has been due, not to any special repressive measures—for few or none have been carried out—but to a general and extensive social improvement in the life of the people, to an increase of knowledge respecting tuberculosis and hygiene, to an enormous advance in sanitation, and to more efficient land drainage.
7. Not all persons are equally liable to consumption, some being much more susceptible than others. We have mentioned the predisposing influence of certain trades. There is also heredity, which acts, as we have said, in transmitting a tubercular tendency, not commonly the actual virus of the disease; there is, thirdly, the debilitating effect of previous illness or chronic alcoholism; there is, fourthly, the habitual breathing of rebreathed air; and, fifthly, there are the conditions of the environment, like dampness and darkness of the dwelling. Such influences as these weaken the resisting power of the tissues, and thus afford a suitable nidus for the bacillus conveyed in milk or by the inspiration of infected dust.
8. Consumption is curable if taken in time. In cases where the lungs are half gone, and consist of large cavities, it is obvious that curability is out of the question. But if the disease can be properly treated in its earliest stages, there is considerable likelihood of recovery.
9. The breath is not dangerous, as far as we know, but there is danger from discharges of any kind from any infected part, whether lungs or bowels; for such discharges, when dry, may readily pollute the air, and either the bacilli or spores be inhaled into the lungs.
10. The chief channels of personal infection or the spread of the disease amongst a community are two: (a) dried tubercular sputum (or other tubercular discharges); (b) infected milk or meat. So long as the former remains wet or moist, infection cannot take place. It is, of course, better to destroy it completely. As for milk and meat, boiling the former and thoroughly cooking the latter will remove all danger.
11. The expectoration is infective. This is one of the commonest modes of infection, and to it is held to be due the large amount of respiratory tuberculosis (consumption, phthisis). The expectoration from the lungs must contain, from the nature of the case, a very large number of bacilli. As a matter of fact, a single consumptive individual can cough up in a day millions of tubercle bacilli. When expectoration becomes dry, the least current of air will disseminate the infective dust, which can by that means be readily reinspired. Expectoration on pavements and floors, as well as on handkerchiefs, may thus become, on drying, a source of great danger to others. The discharges from the bowels of infants suffering from the disease also contain the infective material.
12. Milk, though a much more likely channel for conveyance of tubercle than meat, is only or chiefly virulent when the udder is the seat of tuberculous lesions. The consumption of such milk is only dangerous when it contains a great number of bacilli and is ingested in considerable quantity. Practically the danger from using raw milk exists only for those persons who use it as their sole or principal food, e. g., young children. All danger is avoided by boiling or pasteurising the milk.
At the same time there is an increasing amount of evidence forthcoming at the present time which goes to prove that milk is not infrequently tainted with tubercle (see p. 195). The tuberculin test should be applied to all milch cows, and the infected ones isolated from the herd. Milk supplies should be more strictly inspected even than cowsheds.
13. There are several methods by which meat infection can be prevented. In the first place, herds should be kept healthy, and tubercular animals isolated. Cowsheds and byres should be under sanitary supervision, especially as regards overcrowding, dampness, lack of light, and uncleanliness. Public slaughter-houses under a sanitary authority would undoubtedly be most advantageous. Meat inspection should also be more strictly attended to; efficient cooking, and avoidance of "roll" meat which has not been thoroughly cooked in the middle.
14. Consumptive patients may diminish their disease. Dr. Arthur Ransome95a has laid down five axioms of hygiene for phthisical patients which, if followed, would materially improve the condition of such persons. At Davos, St. Moritz, Nordrach and other places where they have been practised, the beneficial change has been in many cases extraordinary:
(1) Abundance of light, nutritious, easily digested food, which must comprise a large allowance of fat; small meals, but frequent;
(2) An almost entirely open-air life, with as much sunshine as can be obtained;
(3) Suitable clothing, mostly wool;
(4) Cleanliness and bracing cold-water treatment;
(5) Mild but regular exercise.
15. Consumptive patients may also assist in preventing the spread of the disease. In the first place, they should follow the hygienic directions just mentioned, because such conditions fulfilled will materially lessen the contagiousness of such patients; next, the expectoration must never be allowed to get dry. A spitting-cup containing a little disinfectant solution (one teaspoonful of strong carbolic acid to two tablespoonfuls of water) should always be used, or the expectoration received into paper handkerchiefs which can be burnt. Spoons, forks, cups, and all such articles should be thoroughly cleaned before being used by other persons. The patient should not sleep in company with another, but occupy, if possible, a separate bedroom.
Isolation hospitals for consumptives, as for patients suffering from diphtheria, are now being established.
16. House influence has some effect, both directly and indirectly, upon tubercular diseases. Damp soils, darkness, and small cubic space in the dwelling-house exert a very prejudicial effect upon tubercular patients. Sir Richard Thorne[95b] has described the favourable house for such persons as one built upon a soil which is dry naturally or freed by artificial means from the injurious influences of dampness and of the oscillations of the underlying subsoil. The house itself should be so constructed as to be protected against dampness of site, foundations, and walls. Upon at least two opposite sides of the dwelling-house there should be enough open space to secure ample movement of air about it, and free exposure to sunlight. Lastly, it should be possible to have free movement of air by day and night through all habitable rooms of the house. It is clear that many inhabited houses could not stand to these tests; but effort should be made to approach as near to such a standard as possible.
17. Sunlight and fresh air are the greatest enemies to infection.
18. Disinfection is necessary after death from phthisis, and should be as complete as after any other infective disease. Compulsory notification of fatal cases and compulsory disinfection have been officially ordered by the Prussian Government. In this country also absolute disinfection should always be insisted upon after phthisis. Walls, floors, carpets, curtains, etc., should be strictly sterilised. Professor Delépine recommends spraying with 1–100 solution of chloride of lime.
[95a] Arthur Ransome, M.D., F.R.S., Treatment of Phthisis.
[95b] Practitioner, vol. xlvi.
96 Journal of State Medicine, vol. iv. (1896), p. 169.
97 For a fuller statement see Trans. Jenner Institute (First Series), pp. 7–32.
98 See Trans. Jenner Institute (First Series), A. G. R. Foulerton, pp. 40–81.
99 Dated 1890–91. The Commissioners were the late Beaven Rake, M.D., G. A. Buckmaster, M.D., the late Professor Kanthack, of Cambridge, the late Surgeon-Major Arthur Barclay, and Surgeon-Major S. J. Thomson.
100 Bacteriology and Infective Diseases (1896), p. 144. Professor Crookshank's Reports to the Agricultural Department of the Privy Council constitute the most complete account of this disease hitherto published.
101 Zeitschr. f. Hyg. und Inf. Krank., xxv.
102 Journal of State Medicine, December, 1897, p. 561.
103 Bacteriology and Infective Diseases, p. 35.
104 British Medical Journal, 1895 (February), p. 353.
105 British Medical Journal, 1896 (August), p. 439.
106 Journal of State Medicine, 1898 (November), p. 541.
107 The measurement of cubic space is of course made by multiplying together in feet the length, breadth, and height of a room.
108 British Medical Journal, 1898 (April), p. 1013.

The Science Series
Edited by Professor J. McKeen Cattell, Columbia University, with the cooperation of Frank Evers Beddard, F.R.S., in Great Britain.
Each volume of the series will treat some department of science with reference to the most recent advances, and will be contributed by an author of acknowledged authority. Every effort will be made to maintain the standard set by the first volumes, until the series shall represent the more important aspects of contemporary science. The advance of science has been so rapid, and its place in modern life has become so dominant, that it is needful to revise continually the statement of its results, and to put these in a form that is intelligible and attractive. The man of science can himself be a specialist in one department only, yet it is necessary for him to keep abreast of scientific progress in many directions. The results of modern science are of use in nearly every profession and calling, and are an essential part of modern education and culture. A series of scientific books, such as has been planned, should be assured of a wide circulation, and should contribute greatly to the advance and diffusion of scientific knowledge.
The volumes will be in octavo form, and will be fully illustrated in so far as the subject-matter calls for illustrations.
G. P. PUTNAM'S SONS, New York & London
THE SCIENCE SERIES
(Volumes ready, in press, and in preparation.)
The Study of Man. By Professor A. C. Haddon, M.A., D.Sc., Royal College of Science, Dublin. Illustrated. 8o, $2.00
The Groundwork of Science. A Study of Epistemology. By St. George Mivart, F.R.S. 8o, $1.75
Rivers of North America. A Reading Lesson for Students of Geography and Geology. By Israel C. Russell, LL.D., Professor of Geology in the University of Michigan. Illustrated. 8o, $2.00
Earth Sculpture. By Professor James Geikie, F.R.S., University of Edinburgh. Illustrated. 8o, $2.00
Volcanoes. By T. G. Bonney, F.R.S., University College, London. Illustrated. 8o, $2.00
Bacteria. By G. Newman, M.D., F.R.S., Demonstrator of Bacteriology in King's College, London. Illustrated. 8o.
The Stars. By Professor Simon Newcomb, U.S.N., Nautical Almanac Office, and Johns Hopkins University.
Meteors and Comets. By Professor C. A. Young, Princeton University.
The Measurement of the Earth. By Professor T. C. Mendenhall, Worcester Polytechnic Institute, formerly Superintendent of the U. S. Coast and Geodetic Survey.
Earthquakes. By Major C. E. Dutton, U.S.A.
Physiography; The Forms of the Land. By Professor W. M. Davis, Harvard University.
The History of Science. By C. S. Peirce.
General Ethnography. By Professor Daniel G. Brinton, University of Pennsylvania.
Recent Theories of Evolution. By J. Mark Baldwin, Princeton University.
Whales. By F. E. Beddard, F.R.S., Zoölogical Society, London.
The Reproduction of Living Beings. By Professor Marcus Hartog, Queen's College, Cork.
Man and the Higher Apes. By Dr. A. Keith, F.R.C.S.
Heredity. By J. Arthur Thompson, School of Medicine, Edinburgh.
Life Areas of North America: A Study in the Distribution of Animals and Plants. By Dr. C. Hart Merriam, Chief of the Biological Survey, U. S. Department of Agriculture.
Age, Growth, Sex, and Death. By Professor Charles S. Minot, Harvard Medical School.
Bacteria. Dr. J. H. Gladstone.
History of Botany. Professor A. H. Green.
Planetary Motion. G. W. Hill.
Infection and Immunity. Geo. M. Sternberg, Surgeon-General U.S.A.