THE
RAY SOCIETY.
INSTITUTED MDCCCXLIV.

LONDON.
MDCCCLIV.
Title: A Monograph on the Sub-class Cirripedia (Volume 2 of 2)
Author: Charles Darwin
Release date: July 25, 2014 [eBook #46408]
Most recently updated: October 24, 2024
Language: English
Credits: Bryan Ness, Emmanuel Ackerman and the Online Distributed Proofreading Team
Transcriber’s Note
Obvious misprints have been fixed. Archaic and unusual words, spellings and styling have been maintained. Details of the changes are in the Detailed Transcriber’s Notes at the end of the book. Hovering over a Greek word will display its transliteration.
This book was published in two volumes, of which this is the second. The first volume was released as Project Gutenberg ebook #31558, available at http://www.gutenberg.org/ebooks/31558. Links to the first volume have not been created in this volume.
Transcriber Added
| Page | |
|---|---|
| Dedication | v |
| Preface | vii |
| Monograph on the Cirripedia | 1 |
| Introduction | 1 |
| On the Names given to the different parts of Cirripedes | 3 |
| Class Crustacea, Sub-Class Cirripedia | 9 |
| On the Sexual Relation of Cirripedes | 23 |
| Order I.—Thoracica | 30 |
| Family Balanidæ | 33 |
| Table of Contents | 33 |
| Structure of Shell | 34 |
| Structure of the Individual Compartments | 43 |
| Structure of the Radii | 45 |
| Structure of the Alæ | 47 |
| Structure of the Sheath | 48 |
| Structure of the Basis | 49 |
| Structure of the Opercular Valves (Scuta and Terga) | 51 |
| Growth of the Whole Shell, and Its Microscopical Structure | 54 |
| Muscles of Sack | 61 |
| Branchiæ | 63 |
| Thorax and Body | 65 |
| Muscular System | 68 |
| Movements and Muscles of the Cirri | 71 |
| Mouth | 74 |
| Cirri | 81 |
| Caudal Appendages | 85 |
| Alimentary Canal | 85 |
| Circulatory System | 87 |
| Nervous System | 88 |
| Eyes and Vision | 93 |
| Acoustic Organs | 95 |
| Olfactory Sacks | 97 |
| Male Organs of Generation | 97 |
| Female Organs of Generation | 100 |
| Metamorphoses and Homologies | 102 |
| Larva, First Stage | 103 |
| Larva, Second Stage | 109 |
| Larva in the Last or Pupal Stage | 110 |
| Act of Metamorphosis | 126 |
| On the Homologies of the Carapace and Shelly Valves | 131 |
| Cementing Apparatus | 133 |
| Affinities, Classification, Variation | 152 |
| Rate of Growth, Exuviation, Powers of Repairing Injuries | 156 |
| Geographical Range and Habits | 159 |
| Geological History | 172 |
| Sub-Family Balaninæ | 175 |
| 1. Genus Balanus | 177 |
| Sections of the Genus | 193 |
| Section A | 194 |
| 1. Balanus tintinnabulum | 194 |
| Var. communis, vesiculosus, validus, zebra, crispatus, spinosus, coccopoma, concinnus, intermedius, occator, d’Orbignii | |
| Varieties | 201 |
| 2. Balanus tulipiformis | 204 |
| 3. Balanus psittacus | 206 |
| 4. Balanus Capensis | 209 |
| 5. Balanus Nigrescens | 210 |
| 6. Balanus decorus | 212 |
| 7. Balanus vinaceus | 213 |
| 8. Balanus Ajax | 214 |
| Section B | 216 |
| 9. Balanus stultus | 216 |
| 10. Balanus calceolus | 218 |
| 11. Balanus galeatus | 220 |
| 12. Balanus cymbiformis | 221 |
| 13. Balanus navicula | 221 |
| Section C | 223 |
| 14. Balanus trigonus | 223 |
| 15. Balanus spongicola | 225 |
| 16. Balanus lævis | 227 |
| Var. nitidus, Coquimbensis | |
| 17. Balanus perforatus | 231 |
| Var. angustus, Cranchii, fistulosus, mirabilis | |
| 18. Balanus concavus | 235 |
| 19. Balanus amphitrite | 240 |
| Var. communis, venustus, pallidus, niveus, modestus, Stutsburi, obscurus, variegatus, cirratus | |
| Varieties | 245 |
| 20. Balanus pœcilus | 245 |
| 21. Balanus eburneus | 248 |
| 22. Balanus improvisus | 250 |
| Var. assimilis | |
| 23. Balanus nubilus | 253 |
| 24. Balanus corrugatus | 254 |
| Section D | 256 |
| 25. Balanus porcatus | 256 |
| 26. Balanus patellaris | 259 |
| 27. Balanus crenatus | 261 |
| 28. Balanus glandula | 265 |
| Section E | 267 |
| 29. Balanus balanoides | 267 |
| Varieties | 270 |
| 30. Balanus cariosus | 273 |
| 31. Balanus declivis | 275 |
| Section F | 277 |
| 32. Balanus Hameri | 277 |
| 33. Balanus amaryllis | 279 |
| 34. Balanus allium | 281 |
| 35. Balanus cepa | 283 |
| 36. Balanus quadrivittatus | 284 |
| 37. Balanus terebratus | 285 |
| 38. Balanus vestitus | 286 |
| 39. Balanus Imperator | 288 |
| 40. Balanus flosculus | 290 |
| Var. sordidus | |
| 41. Balanus bisulcatus | 293 |
| Var. plicatus | |
| 42. Balanus dolosus | 295 |
| 43. Balanus unguiformis | 296 |
| Var. erisma | |
| 44. Balanus varians | 298 |
| 45. Balanus inclusus | 299 |
| 2. Sub-Genus Acasta | 302 |
| 1. Acasta spongites | 308 |
| 2. Acasta sulcata | 310 |
| 3. Acasta cyathus | 312 |
| 4. Acasta undulata | 313 |
| 5. Acasta glans | 314 |
| 6. Acasta lævigata | 315 |
| 7. Acasta fenestrata | 316 |
| 8. Acasta purpurata | 318 |
| 9. Acasta sporillus | 319 |
| 3. Genus Tetraclita | 321 |
| 1. Tetraclita porosa | 329 |
| Var. communis, nigrescens, viridis, rubescens, elegans, communis (young), patellaris | |
| 2. Tetraclita serrata | 334 |
| 3. Tetraclita rosea | 335 |
| 4. Tetraclita purpurascens | 337 |
| 5. Tetraclita costata | 339 |
| 6. Tetraclita vitiata | 340 |
| 7. Tetraclita cœrulescens | 342 |
| 8. Tetraclita radiata | 343 |
| 4. Genus Elminius | 345 |
| 1. Elminius Kingii | 348 |
| 2. Elminius modestus | 350 |
| 3. Elminius plicatus | 351 |
| 4. Elminius simplex | 353 |
| 5. Genus Pyrgoma | 355 |
| 1. Pyrgoma anglicum | 360 |
| 2. Pyrgoma Stokesii | 361 |
| 3. Pyrgoma cancellatum | 362 |
| 4. Pyrgoma conjugatum | 364 |
| 5. Pyrgoma grande | 365 |
| 6. Pyrgoma milleporæ | 367 |
| 7. Pyrgoma dentatum | 369 |
| 8. Pyrgoma crenatum | 370 |
| 9. Pyrgoma monticulariæ | 372 |
| Species dubiæ | 374 |
| 6. Sub-Genus Creusia | 375 |
| 1. Creusia spinulosa | 376 |
| Varieties with the Scuta and Terga calcified together | 380 |
| 7. Genus Chelonobia | 382 |
| 1. Chelonobia testudinaria | 392 |
| 2. Chelonobia caretta | 394 |
| 3. Chelonobia patula | 396 |
| Second Section of the Sub-Family of Balaninæ | 397 |
| 8. Genus Coronula | 397 |
| 1. Coronula balænaris | 415 |
| 2. Coronula diadema | 417 |
| 3. Coronula reginæ | 419 |
| 4. Coronula barbara | 421 |
| Species Dubiæ | 423 |
| 9. Genus Platylepas | 424 |
| 1. Platylepas bissexlobata | 428 |
| 2. Platylepas decorata | 429 |
| Species Dubiæ | 430 |
| 10. Genus Tubicinella | 430 |
| 1. Tubicinella trachealis | 431 |
| 11. Genus Xenobalanus | 438 |
| 1. Xenobalanus globicipitis | 440 |
| Sub-Family Chthamalinæ | 446 |
| 12. Genus Chthamalus | 447 |
| 1. Chthamalus stellatus | 455 |
| Var. communis, fragilis, depressus | |
| 2. Chthamalus antennatus | 460 |
| 3. Chthamalus cirratus | 461 |
| 4. Chthamalus fissus | 462 |
| 5. Chthamalus dentatus | 463 |
| 6. Chthamalus Hembeli | 465 |
| 7. Chthamalus intertextus | 467 |
| 8. Chthamalus scabrosus | 468 |
| 13. Nov. Genus Chamæsipho | 470 |
| 1. Chamæsipho columna | 470 |
| 2. Chamæsipho scutelliformis | 472 |
| 14. Nov. Genus Pachylasma | 475 |
| 1. Pachylasma giganteum | 477 |
| 2. Pachylasma aurantiacum | 480 |
| 15. Genus Octomeris | 482 |
| 1. Octomeris angulosa | 483 |
| 2. Octomeris brunnea | 484 |
| 16. Genus Catophragmus | 485 |
| 1. Catophragmus polymerus | 487 |
| 2. Catophragmus imbricatus | 490 |
| Remarks on Bronn’s List of Fossil Balaninæ and Chthamalinæ | 492 |
| Family Verrucidæ | 495 |
| Genus Verruca | 496 |
| Powers of Excavation | 512 |
| 1. Verruca Strömia | 518 |
| 2. Verruca lævigata | 520 |
| 3. Verruca Spengleri | 521 |
| 4. Verruca nexa | 522 |
| 5. Verruca prisca | 525 |
| Family Lepadidæ | 526 |
| Genus Alcippe | 529 |
| Alcippe lampas | 530 |
| Female | 530 |
| Male | 555 |
| Order II.—Abdominalia | 563 |
| Cryptophialus minutus | 566 |
| Female | 566 |
| Male | 584 |
| Order III.—Apoda | 587 |
| Proteolepas bivincta | 589 |
| Synopsis et Index Systematicus | 606 |
| Synopsis et Index Systematicus Specierum, et recentium, et fossilum | 611 |
| Description of plates | 641 |
| Plate 1. Balanus tintinnabulum | 641 |
| Plate 2. Genus Balanus | 641 |
| Plate 3. Genus Balanus | 642 |
| Plate 4. Genus Balanus | 642 |
| Plate 5. Genus Balanus | 642 |
| Plate 6. Genus Balanus | 643 |
| Plate 7. Genus Balanus | 643 |
| Plate 8. Genus Balanus | 644 |
| Plate 9. Sub-Genus Acasta | 644 |
| Plate 10. Genus Tetraclita | 645 |
| Plate 11. Genera Tetraclita and Elminius | 645 |
| Plate 12. Genera Elminius and Pyrgoma | 646 |
| Plate 13. Genera Pyrgoma and Creusia | 646 |
| Plate 14. Genera Creusia and Chelonobia | 647 |
| Plate 15. Genera Chelonobia and Coronula | 648 |
| Plate 16. Genus Coronula | 649 |
| Plate 17. Genera Platylepas, Tubicinella, and Xenobalanus | 651 |
| Plate 18. Genus Chthamalus | 652 |
| Plate 19. Genera Chthamalus, Chamæsipho, and Pachylasma | 652 |
| Plate 20. Genera Pachylasma, Octomeris, and Catophragmus | 653 |
| Plate 21. Genus Verruca | 654 |
| Plate 22. Alcippe lampas | 655 |
| Plate 23. Genera Alcippe and Cryptophialus | 658 |
| Plate 24. Genera Cryptophialus and Proteolepas | 660 |
| Plate 25. Genera Proteolepas and Balanus | 662 |
| Plate 26. Structure of the Mouth and Thorax | 664 |
| Plate 27. Nervous System and Senses | 666 |
| Plate 28. Cementing Apparatus | 667 |
| Plate 29. Cirri and Larvæ, first stages | 669 |
| Plate 30. Larvæ of Lepas: second and last stages of development | 671 |
| Errata | 674 |
| Index | 675 |
| Plates | 685 |
List of Illustrations.
| Woodcuts on page 3* | |
| Figure 1. Shell | 3* |
| Figure 2. Compartments | 3* |
| Figure 3. Compartments | 3* |
| Figure 4. Compartments | 3* |
| Figure 5. Scutum (internal view of) | 3* |
| Figure 6. Tergum (external view) | 3* |
| Figure 7. Tergum (internal view) | 3* |
| Woodcuts on other pages | |
| Figure 1. Rostrum with two radii, serving in Chthamalinæ for rostro-lateral compartments | 36 |
| Figure 2. Always serving for lateral and carino-lateral compartments | 36 |
| Figure 3. Carina, serving in the Chthamalinæ, also, as a rostrum | 36 |
| Figure 4. Octomeris | 39 |
| Figure 5. Chthamalus | 39 |
| Figure 6. Chamæsipho | 39 |
| Figure 7. Balanus | 39 |
| Figure 8. Tetraclita | 39 |
| Figure 9. Basal edge of wall of compartment in Balanus tintinnabulum | 43 |
| Figure 10. Edge of the radius of Balanus tintinnabulum | 46 |
| Figure 11. Portion of edge of basis of Balanus tintinnabulum | 50 |
| Plates | |
| Plate 1. Balanus tintinnabulum | 685 |
| Plate 2. Balanus | 686 |
| Plate 3. Balanus | 687 |
| Plate 4. Balanus | 688 |
| Plate 5. Balanus | 689 |
| Plate 6. Balanus | 690 |
| Plate 7. Balanus | 691 |
| Plate 8. Balanus | 692 |
| Plate 9. Acasta | 693 |
| Plate 10. Tetraclita | 694 |
| Plate 11. Tetraclita: Elminius | 695 |
| Plate 12. Elminius: Pyrgoma | 696 |
| Plate 13. Pyrgoma: Creusia | 697 |
| Plate 14. Creusia: Chelonobia | 698 |
| Plate 15. Chelonobia: Coronula | 699 |
| Plate 16. Coronula | 700 |
| Plate 17. Platylepas: Tubicinella: Xenobalanus | 701 |
| Plate 18. Chthamalus | 702 |
| Plate 19. Chthamalus: Chamæsipho: Pachylasma | 703 |
| Plate 20. Pachylasma: Octomeris: Catophragmus | 704 |
| Plate 21. Verruca | 705 |
| Plate 22. Alcippe lampas | 706 |
| Plate 23. Alcippe: Cryptophialus | 707 |
| Plate 24. Cryptophialus: Proteolepas | 708 |
| Plate 25. Proteolepas: Balanus | 709 |
| Plate 26. Mouth: Thorax | 710 |
| Plate 27. Nervous System | 711 |
| Plate 28. Cementing Apparatus | 712 |
| Plate 29. Cirri: Larvæ, first stages | 713 |
| Plate 30. Larvæ, last stages | 714 |
THE
RAY SOCIETY.
INSTITUTED MDCCCXLIV.
LONDON.
MDCCCLIV.
BY
CHARLES DARWIN, F.R.S., F.G.S.
THE BALANIDÆ,
(OR SESSILE CIRRIPEDES);
THE VERRUCIDÆ,
ETC., ETC., ETC.
LONDON:
PRINTED FOR THE RAY SOCIETY.
MDCCCLIV.
Reprinted with the permission of the Ray Society
JOHNSON REPRINT CORPORATION
111 Fifth Avenue, New York, N. Y. 10003
JOHNSON REPRINT COMPANY LTD.
Berkeley Square House, London, W. 1
First reprinting, 1968, Johnson Reprint Corporation
Printed in the United States of America
TO
PROFESSOR H. MILNE EDWARDS,
DEAN OF THE FACULTY OF SCIENCES OF PARIS;
PROFESSOR AT THE MUSEUM OF NATURAL HISTORY;
MEMBER OF THE INSTITUTE OF FRANCE;
FOREIGN MEMBER OF THE ROYAL SOCIETY OF LONDON,
OF THE ACADEMIES OF BERLIN, STOCKHOLM, ST. PETERSBURG, VIENNA,
KONIGSBERG, MOSCOW, BRUSSELS, HAARLEM, BOSTON, PHILADELPHIA, ETC.
THIS WORK IS DEDICATED,
WITH THE MOST SINCERE RESPECT,
AS THE ONLY, THOUGH VERY INADEQUATE ACKNOWLEDGMENT
WHICH THE AUTHOR CAN MAKE OF HIS GREAT AND
CONTINUED OBLIGATIONS TO THE
‘HISTOIRE NATURELLE DES CRUSTACÉS,’
AND TO
THE OTHER MEMOIRS AND WORKS ON NATURAL HISTORY PUBLISHED BY
THIS ILLUSTRIOUS NATURALIST.
Having so lately, in my volumes on the Recent and Fossil Lepadidæ, expressed as strongly as I could, and with the utmost sincerity, the obligations under which I lie to very many naturalists, I will not here repeat my thanks, and will only say that the assistance formerly rendered me from so many quarters has been most kindly continued without intermission. The references under the Habitats, in which I may remark the names of Mr. Cuming and of Mr. Stutchbury, and of the British Museum, so often recur, show my deep obligations to these gentlemen and to Dr. Gray, and indeed to most of the British and several Foreign[1] collectors of recent and fossil shells. At the period when the Introduction to this volume was printed, I stated that I did not know whether the Palæontographical Society would publish the few British fossil Balanidæ; the[Pg viii] Council has now honoured me by determining on this publication, so that these species will hereafter be more fully illustrated than they could be in the present volume. I cannot conclude this short preface, without again tendering my most grateful thanks to the Council of the Ray Society for the publication of my two volumes, and for the very kind manner in which they have acceded to all my requests.
DOWN, KENT; July, 1854.
[1] I feel under special obligation to Mr. Dana for several very interesting communications connected with the present subject, and for information derived from his magnificent work on the Crustacea, collected during the United States Exploring Expedition. Also to M. Bosquet, of Maestricht, for the loan and gift of several interesting fossils, described and illustrated with the utmost fidelity, in his beautiful “Monographie des Crustacés fossiles du terrain Crétacé du D. de Limbourg.”
ON
THE CIRRIPEDIA.
My former volume, published by the Ray Society, treated only of the Lepadidæ, one family of the Cirripedia: I was induced to print it from having the materials ready, though this partial publication has been in some respects inconvenient. The Palæontographical Society has done me the honour to publish, with ample illustrations, the fossil species of this same family of Lepadidæ. This present volume completes my work on the sub-class Cirripedia.[2] I had originally intended to have published a small volume on my anatomical observations; but the full abstract given in my former volume, which will be illustrated to a certain extent in the plates appended to this volume, together with the observations here given under the Balanidæ, appear to me sufficient, and I am unwilling to spend more time on the subject. In the volume on the Lepadidæ, I gave the specific or diagnostic characters in English and Latin: I have here left out the latter, inasmuch as I have appended at the end of this volume a Latin Synopsis of all the[Pg 2] species, recent and fossil, of the whole class. To each species is added a reference to the pages and plates of my three volumes, so that the Synopsis will serve as a systematic index to the three: an alphabetical index to the present volume is also given. In the Lepadidæ, I gave an additional specific character, derived from the softer parts of the animal’s body: in the Balanidæ, these parts are more alike in the different species, and I have found it impossible to give a diagnostic character thus derived. In those cases in which a Family contains but one genus, or a Genus but one species, I have assigned my reasons for the institution of such groups, but have given, as heretofore, only a single description in full: it would have been easy to have separated, by analogy, this description into one for the species, another for the genus or for the family; but as I believe such separation and subordination of the characters would have been largely conjectural, I have thought it best to act as I have done, and give, thus saving useless repetitions, only a single description, and leave it for my successors, when more genera or species are known, to separate, with such certainty as is ever possible, the generic from the specific characters.
[2] The number of the British fossil species of the Balanidæ and Verrucidæ in a recognisable state is so small, that I do not know whether it will be considered worth while to publish in the Palæontographical series more detailed illustrations than are given in this volume.
In nomenclature, I have endeavoured rigorously to follow the rules of the British Association, and have never, at least intentionally, broken through the great law of priority. In accordance with the rules, I have rejected, that is, as compulsory, all names given before the introduction of the binomial system in 1758. I have given much fewer synonyms than is usual in conchological works; for it is impossible to recognise with any approach to certainty, several even of the common European forms, in the short descriptions given by most authors; this holds good in many cases in which figures, in appearance excellent, have been added. I assert this the more confidently, from having had the advantage of having gone over some of the Linnean synonyms with Mr. S. Hanley. I may further venture to express my conviction, that giving references to works, in which there is not any original matter, or in which the plates are not of a high order of excellence, is absolutely injurious to the progress of natural history.
NOMENCLATURE OF THE SHELL OF A SESSILE CIRRIPEDE.
Orifice of shell, surrounded by the sheath. Sheath formed by the alæ (a—a.) and by portions of the upper and inner surfaces of the parietes (p—p.)
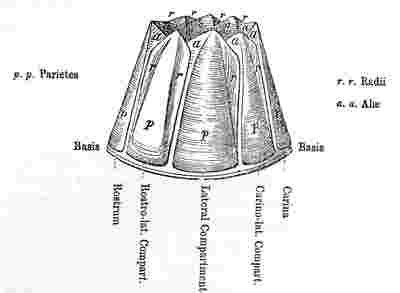
N.B. In Balanus, and many other genera, the Rostrum and Rostro-lateral compartments are confluent, and hence the Rostrum has the structure of Fig. 2.
COMPARTMENTS.
Fig. 2. Compartment with two radii, serving either as a Rostrum or Rostro-lateral compartment.
Fig. 3. serves as a Lateral and Carino-lateral Compartment.
Fig. 4. serves as a Carina or Rostrum.
OPERCULAR VALVES.
Sessile Cirripedes, partly from being attached to surfaces [Pg 3]having very different characters, partly from undergoing a varying amount of disintegration, and partly from unknown innate causes, are extremely variable. Under the head of Variation, in the Family Balanidæ and under the Genus Balanus, I have enlarged on this subject, and have shown that there is scarcely a single external character which is not highly variable in most of the species. As whole groups of specimens often vary in exactly the same manner, it is not easy to exaggerate the difficulty of discriminating species and varieties. It is absolutely necessary in most cases, for mere identification, that the valves of at least one specimen in a group should be disarticulated and well cleaned (for which purpose caustic potash is most useful), so that the internal characters may be examined. Whoever attempts to make out from external characters alone, without disarticulating the valves, the species, (even those inhabiting one very confined region, for instance the shores of Great Britain,) will almost certainly fall into many errors: hence it is, and can thus only be accounted for, that I have not seen one collection of British specimens with all the species, though so few in number, rightly discriminated; and in the large majority of cases, either two or three species, certainly distinct, were confounded together, or two or three varieties, as certainly not distinct, were separated from each other.
In my former volume I have stated that I found it indispensable, in part owing to the extreme confusion of the nomenclature previously used, to attach new names to several of the external parts of Cirripedes. Almost all these names are applicable to the Balanidæ, or sessile Cirripedes, and to the Verrucidæ; but a few additional names are requisite, which, together with the old names, will, I hope, be rendered clear by the accompanying woodcuts. In sessile Cirripedes, the whole of that which is externally visible, may for convenience sake be divided into the[Pg 4] operculum or opercular valves (valvæ operculares), and the shell (testa), though these parts homologically present no real difference. The operculum is seated generally some little way down within the orifice of the shell; but in very young specimens and in Verruca, the operculum is attached to the summit of the shell, and the shell, without the operculum being removed, can hardly be said to have any orifice; though, of course, the opercular valves themselves have an aperture for the protrusion of the cirri.
The shell consists of the basis (called the support by some authors), which is membranous or shelly, and flat or cup-formed, and of the compartments (testæ valvæ), which vary from eight to four in number, and occasionally are all calcified together.
The compartment, at that end of the shell where the cirri are exserted through the aperture or lips of the operculum, is called the carina (fig. 1); the compartment opposite to it is the rostrum,—these two lying at the ends of the longitudinal axis of the shell. Those on the sides are the lateral compartments; that nearest the carina, being the carino-lateral (testæ valva carino-lateralis), that nearest the rostrum, the rostro-lateral, and the middle one, simply the lateral compartment; but these three compartments are rarely present together. The rostro-lateral compartment, which always resembles fig. 2, and may be always known by having radii on both sides, is often absent; and not rarely the lateral and carino-lateral compartments are confounded together, or one is absent; in such cases the compartment that is left is simply called the lateral one. The compartments are separated from each other by sutures, which are often so fine and close as to be distinguished with difficulty. The edge of a compartment, which can only be seen when disarticulated from its neighbour, I have called the sutural edge (acies suturalis).
Each separate compartment consists of a wall (paries), or parietal portion (pp in woodcuts), which always grows downwards, and forms the basal margin; and is furnished on the two sides either with alæ (fig. 4), or with radii (fig. 2), or with an ala on one side and a radius (fig. 3) on the other.
The radius[3] (adopting the name used by Bruguière, Lamarck, and others) differs remarkably in appearance (though not in essence) from the walls or parietal portion, owing to the direction of the lines of growth and the state of its usually depressed surface. In the upper part the radii overlie the alæ of the adjoining compartments: in outline (r, fig. 1, 2, 3), they are wedge-formed, with their points downwards; their summits (and this is often a useful specific character) are either parallel to the basis or as in fig. 1 and 2, oblique. The radii are sometimes not developed.
[3] The radii have been called by Ranzani and De Blainville “areæ depressæ” (the parietal portions of the compartments being the “areæ prominentes”); by Poli, “areæ interjectæ;” by Gray, “sutures;” by Coldstream, “compartments of the second order,” (the parietal portions being those of the first order); by some authors as “intersticia.” I may here add that the scuta are the “ventral valves” of Gray, the “anterior” of Ranzani, and the “inferior opercular” of De Blainville: the terga are the “posterior valves” of Gray and Ranzani, but the “superior opercular” of De Blainville: the rostrum, on the other hand, is the “anterior valve” of Ferussac and the “ventral” of De Blainville; the carina being the “dorsal valve” of the latter author.
The alæ (so called by Dr. Gray) are overlapped by the radii and by part of the walls; they usually extend only about half way down the compartment (a fig. 3, 4, 1); their summits are either parallel to the basis or oblique. The alæ of the several compartments, together with the internal, upper, thickened surfaces of the walls, against a shoulder of which the sutural edges of the alæ abut, have been called (by Dr. Gray) the sheath (vagina). The upper and greater portion of the sheath is marked by transverse lines, caused by the exuviation of the opercular membrane, as that membrane may be called, which unites the operculum all round to the sheath, or upper internal surface of the shell.
The carina has always two alæ, as in fig. 4.
The carino-lateral and lateral compartments have always an ala on one (the rostral) side, and a radius on the other (the carinal) side, as in fig. 3.
The rostro-lateral compartment (when present) has always radii on both sides, as in fig. 2.
The rostrum has normally alæ on both sides, as in fig. 4, but very often from fusion with the rostro-lateral compartments on both sides, it has radii on both sides, as in fig. 2.
The walls of the shell, the basis, and the radii, are in very many cases composed of an outer and inner lamina, united together by longitudinal septa; a set of tubes or pores being thus formed. The points of the longitudinal septa generally project beyond the laminæ, and are denticulated on both sides (see woodcut, further on;) the septa are sometimes branched, several irregular rows of pores between the two laminæ being then formed (see Pl. 7, fig. 3 b, and Pl. 10, fig. 1 g, 1 h).
Operculum, or opercular valves.—These consist of a pair of scuta and a pair of terga. They are joined to the sheath of the shell by the opercular membrane.
Scutum (woodcut 5): this valve is generally sub-triangular, and its three margins are the basal, the tergal, so called from being articulated with the tergum, and the occludent, so called from opening and shutting against the opposed valve. The angles are called from the adjoining margins, as basi-tergal, &c.; the upper angle being the apex. The scutum is ordinarily articulated to the tergum by an articular ridge (crista articularis), running up to the apex of the valve, and by an articular furrow, which latter receives the scutal margin of the tergum. The articular ridge, instead of projecting straight up from the valve, when laid flat on its external surface, often bends over to the tergal side, and is then said to be reflexed. On the internal surface of the valve, there is almost always an adductor pit or cavity (fossa adductoris), for the attachment of the adductor scutorum muscle: this pit is often bounded on its tergal and basal sides, by a ridge, called the adductor ridge (crista adductoris), which, in its upper part, is often confluent with the articular ridge. Beneath the adductor ridge, in the basi-tergal corner of the valve, there is often a lateral-depressor pit (fossa musculi lateralis depressoris), for the attachment of the so-called muscle; and this pit is sometimes furnished with crests.
Tergum, (woodcut 6 and 7):—this valve, also, has three margins, the scutal, basal, and carinal; its upper end, or apex, is sometimes beaked; on the basal margin a spur (calcar) depends; the outer surface of the valve is depressed or longitudinally furrowed (sulcus longitudinalis) in the line[Pg 7] of the spur. The part called the spur is often so broad, that the name becomes not very appropriate. The angles are denominated, from the adjoining margins, as basi-carinal, or basi-scutal angle, &c. On the under side, in the upper part, there is an articular ridge, and on its scutal side, an articular furrow, receiving the articular ridge of the scutum. In the basi-carinal corner of the valve there are often crests for the attachment of the tergal depressor muscle.
Sack, Body, Cirri, Mouth.—A slit-like orifice between the opercular valves leads into the sack, in which the body is lodged. The body consists of the six (perhaps the seven) posterior thoracic segments of the archetype Crustacean; the first of these six segments (or first two, if there be seven segments) is developed on its dorsal aspect into a part, which I have called the prosoma[4] (see fig. 1, c, Pl. 25). There is no abdomen. The thoracic segments support six pairs of cirri. Each cirrus consists of a two-jointed pedicel, carrying two multiarticulated rami. Rarely there are articulated caudal appendages (appendices caudales) on each side of the anus. The prominent mouth consists of a labrum, palpi, mandibles, maxillæ, and outer maxillæ, the latter resembling a lower lip: these organs may be conveniently spoken of, after Milne Edwards, as gnathites. Within the sack, attached to its carino-lateral end, a folded membrane forms the branchiæ. The sheets of ova lying within the sack are called the ovigerous lamellæ.
[4] A discussion on the homologies of the different parts is given under the head of the Metamorphoses of the Balanidæ.
I have often found it convenient to designate the membrane investing the body, lining the sack, &c., by its proper chemical name of chitine, instead of by horny, or other such equivalents; but when covering parts of the shell, for brevity’s sake I have often spoken of it as an epidermis, but I do not believe that such is its nature. When this membrane sends into the body of the animal rigid projections or crests, for the attachment of muscles or any other purpose, I call them, after Audouin, apodemes. For the underlying true skin, I use the term corium.
Relative position of parts.—The centre of the generally flat basis, which is cemented to the supporting surface, is[Pg 8] properly the anterior end, and the tips of the terga, often hidden within the shell, are properly the posterior end of the external covering; but I have found it more convenient to speak of the upper and basal surfaces and aspects, which hardly admit of any mistake. A line drawn from the centre of the basis, along the middle of the rostrum to the tips of the scuta, shows the strictly medio-ventral surface of the shell; and another line drawn from the centre of the basis, along the carina, to the tips of the terga, shows the strictly medio-dorsal line; but from the crooked course of these lines, I have found it far more convenient to speak of the rostral and carinal end or aspect of the different parts of the shell; this is the more necessary with respect to the internal parts of the animal, owing to their remarkable changes of position during the metamorphosis, whence it comes that the dorsal surface of the thorax faces partly dorsally, partly anteriorly or downwards, and partly even ventrally; and the ventral surface of the whole posterior part of the thorax faces upwards or posteriorly; but when we refer these parts to the rostral, carinal, basal, and upper ends of the shell, there can be no mistake. There has moreover been great confusion in these relative terms, as applied by different authors.
When a sessile Cirripede is held in the position in which they have generally been figured, namely with the basis downwards and the scuta towards the beholder, then the right and left sides of the Cirripede correspond with those of the holder.
I have followed the example of Botanists, and added the interjection (!) to synonyms, when I have seen an authentic specimen bearing the name in question.
Every locality, under each species, is given from specimens ticketed in a manner and under circumstances appearing to me worthy of confidence,—the specific determination being in each case made by myself.
Crustacea attached by the anterior end of the head, by cement proceeding from a modified portion of the ovaria; archetype composed of seventeen segments, with the three first of large size, and almost always developed into a carapace, not wholly exuviated, and capable of various movements; antennæ none; eyes rudimentary; mouth prominent, formed by the partial confluence of the labrum, palpi, mandibles, and two pairs of maxillæ; thorax attached to the internal sternal surface of the carapace, generally bearing six pairs of captorial, biramous, multiarticulated limbs; abdomen generally rudimentary; branchiæ, when present, attached to the under sides of the carapace; generally bisexual, when unisexual, males epizoic on the female; penis single, generally probosciformed, seated at the posterior end of the abdomen; oviducts none; metamorphoses complex.
Within the memory of many living naturalists, Cirripedes were universally looked on as belonging to the Molluscous kingdom; nor was this surprising, considering the fixed condition of their shells, and the degree of external resemblance between, on the one hand, Lepas and Teredo, and on the other hand, between Balanus and a Mollusc compounded of a patella and chiton. It is remarkable that this external false appearance overbore, even in the mind of Cuvier, his knowledge of their internal structure, namely, their lateral jaws, articulated appendages, and regular ganglionic nervous system, which now strike us as such conclusive evidence of their position in the great Articulate kingdom. Straus[5] was, I believe, the first who, in 1819, maintained that Cirripedes were most closely allied to Crustacea. But this view was disregarded, until J. Vaughan Thompson’s[6] capital discovery, in 1830, of their metamorphoses,[Pg 10] since which time, Cirripedes have been almost universally admitted amongst the Crustaceans. It is well known, that it is hardly possible to give a definition of this great class, which shall include every member of it; nevertheless, even if the mature Cirripede alone be considered, the following characters, viz. the slight separation of the head and thorax, the latter generally bearing six pairs of appendages, and the being enclosed in a carapace—together with the periodical exuviation of the greater part of the external membranes, would, perhaps, suffice to show that it should be classed amongst Crustacea.
[5] Mémoires du Muséum d’Histoire Nat., tom. v, p. 381.
[6] Zoological Researches and Illustrations.
But it still remains undecided what rank in this class Cirripedes should hold. Before briefly discussing this point, it is indispensable to indicate their essential characters, which I will immediately attempt. For as long as it remained doubtful which was their anterior extremity, which the ventral or dorsal surface; as long as the peduncle was thought by one naturalist to be the legs, by another the abdomen, in a modified condition, it was hopeless to compare Cirripedes with ordinary Crustaceans, and assign to them their due rank.
In the larva in the first stage, an eye and two pairs of antennæ are in process of formation or are developed; here, then, according to the analogy of all Crustaceans, we have evidence of the existence of the first three cephalic segments. The mouth always consists of three pairs of gnathites, and hence again, from analogy, this part may be inferred to be formed of, and supported on, three other segments; making thus far six segments. In two Orders out of the three into which Cirripedes may be divided, namely, in the Abdominalia and Apoda, eight quite distinct segments succeed the mouth; of these the first differs slightly from the seven succeeding segments, and may, I think, be safely considered as forming the seventh (cephalic) segment. The next seven segments resemble each other in all essential respects, and are no doubt the normal, seven thoracic segments. These, in both the above orders, are succeeded by three smaller segments, which differ in structure from the thoracic segments, and must be abdominal. Hence we here have, altogether, seventeen segments. It should,[Pg 11] however, be observed that in the two orders just referred to, each includes only a single species; but I know of no good reason why, on this account, their structure should be valued the less. In the third order, the Thoracica, which includes all common Cirripedes, two segments with their appendages are missing out of the eight that should succeed the mouth; from the open interval in the pupa, between the mouth and first pair of natatory legs, and from some other reasons, I believe that the two missing segments are the seventh and eighth, or last cephalic and first thoracic segments, and that they have coalesced close posteriorly to the mouth.[7] In the order Thoracica, the abdomen is quite rudimentary, though often still bearing caudal appendages; in the pupa, however, of this order, as in the mature animal of the two other orders, it is formed of three segments. Hence I conclude that, notwithstanding the absence of the above two segments with their appendages in the Thoracica, the archetype Cirripede may be safely said to be composed of seventeen segments.
[7] This question and the whole subject of the homologies of the several parts of a Cirripede, will be discussed under the head of the Metamorphoses of the Balanidæ.
In the classification of Crustacea, the relation and number of the segments of the different parts of the body, are viewed both by Prof. Milne Edwards[8] and Mr. Dana,[9] as of the highest importance. I may premise that both these authors divide the Crustacea into Podophthalmia, Edriophthalmia, and Entomostraca; Milne Edwards making a fourth legion, the Branchiopoda, and another division, including Limulus, of equal value to the above four legions altogether; whereas Dana sinks Limulus and the Branchiopoda under his Entomostraca. As far as concerns our present discussion on Cirripedes, the first three divisions, as valued by Dana, will best serve as standards of comparison; but it is not unimportant to our present purpose, as showing how difficult it is to weigh the value of the higher divisions of a Class, to observe the wide difference in opinion of two naturalists, so eminent for their knowledge of the class in question and for their high abilities.
[8] Annales des Sciences Nat., tom. xviii, p. 120, 1852.
In the order Thoracica, including all common Cirripedes, the cephalic and thoracic segments are as much confounded together (but with coalescence and abortion of two middle segments) as in most Podophthalmia; but in the two other orders, the cephalic and thoracic segments are as distinct as in the Edriophthalmia. The number of the segments, however, which strictly appertain to the anterior part of the head and mouth, being only six, is an Entomostracan character; on the other hand, the first pair of cirri in the Thoracica, has some claim from their position, apparent functions, and separation from the succeeding pairs, to be said to belong to the mouth; on which view, the first nine segments would, in function, be cephalic, as in the highest Crustaceans. The fewness of the segments of the abdomen, and their not bearing in two of the orders appendages, is an Entomostracan character.
Cirripedes are permanently attached, even before their final metamorphosis, by a tissue or cement, first debouching through the second pair of antennæ, and, subsequently, in most cases, through special orifices, penetrating the anterior part of the head; this cement proceeds from glands, which certainly are modified portions of the ovarian system. This fact I consider of the highest classificatory importance, for it is absolutely the one single character common to all Cirripedes, besides such as show only that these animals belong to the articulated kingdom, and are Crustaceans. No structure of this kind has hitherto been observed in any other member of the class or kingdom. Even in the Suctorial Entomostracans, which become immoveably attached to the fish on which they prey, the males are free; and the means of attachment, as far as known, are quite different.
Both the first and second pairs of antennæ are absent in the mature animal; for the three terminal segments of the antennæ of the pupa, which may always be found cemented under the centre of the surface of attachment, are functionless, after maturity. The eyes are rudimentary, and are singular from being seated far from the anterior extremity of the head. In their rudimentary state, and in the absence of antennæ, we have characters common with certain Suctorial Entomostracans; and this similarity[Pg 13] apparently arises from the fixed condition of the animals of both groups.
The carapace, which covers the dorsal surface of the larva in the first stage, in the last larval or pupal stage is developed so as to enclose, like a bivalve shell, the whole body. In the mature Cirripede, the whole external covering, whether shell and operculum, or capitulum and peduncle, can be conclusively shown to be the carapace of the pupa, modified. In thus enclosing the mouth and whole body, the modified carapace resembles that of several Entomostracans; but in being apparently formed (as I hope hereafter to show) by the development of the third segment of the head, and in consisting generally of distinct sclerodermic plates, arranged in an imbricated order, there is, I think, a closer resemblance to the same part in some of the Podophthalmia. The carapace, or portions of the carapace, being capable of other movements, besides merely opening and shutting, differs, I believe, from that of all other Crustaceans; as it likewise does[10] in the greater part not being periodically moulted.
[10] The carapace, however, of the Isaura, a Branchiopod, according to M. Joly (‘Annales des Sc. Nat.,’ 2 ser. vol. xvii, p. 293), is not moulted.
Moreover, in all Cirripedes there is another striking peculiarity connected with these parts, namely, the exclusive attachment of the whole thorax or included body to the internal ventral or sternal surface of the carapace and head. In the pupa, the thorax, as in all Crustaceans, opens into, and is continuously united with, the large anterior part of the head; but from the singular fact that the thorax of the young Cirripede is developed not within the thorax, but within the head of the pupa (Pl. 30, fig. 2), with its longitudinal axis placed at right angles to that which it held in the pupal condition (the mouth and the whole exterior being developed conformably with that of the pupa), it comes to pass after the metamorphosis, that the Cirripede is, as it were, internally cut in twain (compare Pl. 25, fig. 1, and Pl. 30, figs. 2 and 3). Thus it is, as will hereafter be more fully explained, that the sack originates, and thus the body becomes attached to the internal ventral surface of the carapace and front of head.
The thorax in two of the Orders bears no appendages, but in all common Cirripedes it is furnished with six pairs of biramous, multiarticulated cirri, which have a peculiar character, different from the limbs of other Crustaceans, not being natatory, ambulatory, or branchial, but “captorial” or fitted for sweeping the water, and thus catching prey.[11] The cirri, at least the anterior pairs, can, besides other movements, lengthen and shorten themselves; and this Milne Edwards[12] states is the case with the Podophthalmia, and is considered by him as an important character. The cirri of the first pair are attached on each side close to the bases of the mandibles, and, as already remarked, have some claim to be considered as maxillipeds or mouth organs. The three or the four posterior pairs of cirri in the Balanidæ, form a series somewhat distinct from the two or three anterior pairs, thus recalling a characteristic feature in the Edriophthalmia.
[11] M. A. Hancock, in ‘Annals and Magazine of Natural History,’ 2d series, 1849, p. 312, speaks of the cirri acting like a prehensile net.
[12] ‘Annales des Sciences Nat.,’ tom. xviii, p. 121, 1852.
The mouth is prominent, and is formed by the partial confluence of the labrum, palpi, and lower segments of the mandibles, and of two pairs of maxillæ; it is capable of movement as a whole; in this respect we are reminded of the Suctorial Entomostracans; but I believe the above type of structure of the mouth is peculiar to Cirripedes.
The alimentary canal is simple, but can be distinctly divided into—(1st) an œsophagus, singular from the bell-shaped expansion of its lower end; (2d) the stomach, which is directed forwards and then doubled back; and (3d) the rectum. There is no distinct liver. The circulation is lacunal. In one family there are well-developed branchiæ, which differ entirely in their homologies and position from these organs in all other Crustaceans. In the nervous system, the sub-œsophageal ganglions vary in concentration from that degree observed in the lower Macroura, to that in the highest Brachyoura; but the supra-œsophageal ganglions are always much less concentrated, and are even embryonic in condition; presenting a difference not observed in other Crustaceans. On the under side of the sub-œsophageal ganglion, two nerves, apparently splanch[Pg 15]nic, arise, and run almost parallel and under the collar surrounding the œsophagus; they are very remarkable from their great size, and from forming a plexus together with a large branch, arising on each side from the collar close behind the supra-œsophageal ganglion,—a structure unlike anything observed in other Crustaceans. The eyes, as already remarked, are rudimentary, and singular from being imbedded at a distance from the anterior end of the animal. In the basal confluent segments of the outer maxillæ there are two orifices, leading into little sacks, which I believe are olfactory organs: again there are two other orifices on each side of the thorax, beneath the first pair of cirri, leading into sacks, with a curious elastic vesicle suspended within them; and these I can hardly doubt are acoustic organs. Of these orifices and organs, there is no trace in the same relative positions in any known Crustacean.
Cirripedes are ordinarily bisexual, in which they differ from all Crustaceans: when the sexes are separate, the males are minute, rudimentary in structure, and permanently epizoic on the females; to these latter facts we have a partial analogy in some of the Suctorial Entomostracans; but a far closer analogy in certain Rotifers, which are considered by many naturalists as Crustaceans; but to the above subject I shall almost immediately have to recur.
The male excretory organ is probosciformed and capable of the most varied movements; it is single and medial; it is seated (in the one instance in which this point can be safely judged of) at the extremity of the abdomen, and therefore near the normal position of the anus; in all these respects there is a very great difference from other Crustaceans, in which the male organs are laterally double, and are not seated at the extremity of the abdomen. In regard to the female organs, the ovarian tubes and cæca inosculate together: there are no oviducts; the ova, connected together by membrane, and so forming the “ovigerous lamellæ,” become exposed by the exuviation of the lining tunic of the carapace or sack, and by the formation of a new tunic on the under side of these lamellæ; a process, I believe, unknown in other Crustaceans.
The metamorphoses are highly complex. The larva in[Pg 16] its first stage bears a very close general resemblance, in having three pairs of natatory appendages, the first being uniramous and the two others biramous, and in having a single eye on its broad anterior front, to the larvæ of most Entomostracans; but I cannot avoid the belief, that this resemblance is only apparent, and not essential; and of false resemblances, how many instances occur in the animal kingdom! In the larva, when first freed from the egg, both pairs of antennæ are in process of formation within envelopes: the mouth is probosciformed and capable of movement, but is destitute of gnathites; it occupies a position between the three pairs of natatory limbs; and these limbs I must believe, for reasons hereafter to be assigned, answer (improbable as I am well aware it must at first appear) to the second, third, and fourth thoracic legs of the archetype Crustacean: the two hinder pairs of limbs apparently soon become captorial, or fitted to secure prey. Now, I cannot find in the published accounts of the larvæ of Entomostracans, any that answer to this description.
The larva in the last stage might be included in the vast class of Entomostracans: the attachment of the eyes to the singular apodemes produced inwards from the basal segment of the great prehensile antennæ, and the development of only the posterior six pairs of thoracic limbs, are its chief peculiarities: but its rudimentary mouth, owing to its transitional or pupal condition, renders the assignment of its proper rank difficult.
Having now given this short comparative sketch of the structure of a Cirripede, I may venture to express strongly my opinion, that the group is formed on a distinct type; as different from the other three or four main Crustacean groups, namely, the Podophthalmia, Edriophthalmia, Branchiopoda, and Entomostraca, as these differ from each other; the differences, moreover, being of the kind considered by the highest authorities on this subject, as the most important. It should be observed that there is no special blending at either end of the Cirripedial series, towards any one of the other main groups of Crustacea; it is hardly possible to take some one Cirripede, and say[Pg 17] that it leads, more plainly than some other Cirripede, into ordinary Crustaceans. Moreover, a great range of structure, as we shall soon briefly show, is included within the group: I can adduce three or four undoubted Cirripedes, very considerably more different from each other, than any two members within the sub-class Podophthalmia, or within the Edriophthalmia, or the Branchiopoda, and quite as different as within the Entomostraca.
The opinion here expressed, that Cirripedes form a sub-class of equal value with the other main Crustacean groups, I am well pleased to find, accords with Mr. Dana’s[13] view, who remarks that this sub-class “has so many peculiarities of structure, that it should be regarded as a distinct type, rather than a subordinate division of the third (or Entomostracan) type.” M. Milne Edwards,[14] after dividing all Crustacea into two groups, divides one of them into four legions; and of one of these, the Entomostraca, he makes the Cirripedes a sub-group. I feel so entire a deference for any opinion on affinities or classification expressed by Milne Edwards, that I differ from him with the greatest hesitation. He does not give his reasons for assigning so subordinate a rank to Cirripedes, but I imagine it is from the nature of their metamorphoses: but if this be the case, I cannot understand why he should assign to his Branchiopods a rank equal to his Entomostracans. Moreover, I must repeat, that I do not believe that the larvæ do resemble the larvæ of Entomostracans and Branchiopods nearly so closely as at first appears to be the case. I may add, that Burmeister[15] has assigned to the Cirripedes a place amongst the Crustacea, almost equally subordinate to that given to them by Milne Edwards.
[13] ‘Crustacea: United States Exploring Expedition,’ p. 1407, 1852.
[14] ‘Annales des Sciences Nat.,’ tom. xviii, p. 120, 1852.
[15] ‘Beiträge zur Naturgesehichte der Rankenfüsser,’ 1834.
That Cirripedes have some special affinity to the Entomostraca, may be inferred from the fewness of the cephalic appendages, the biramous legs, the state of the abdomen, and the form of the carapace. Perhaps in the peculiar state of confluence of the lower segments of the gnathites,[Pg 18] in the aborted antennæ, the rudimentary eyes, and in the minute parasitic males (when such exist), there is a more direct relation to the Suctorial division of the Entomostraca; but some of these resemblances are probably only analogical, resulting from the fixed condition of both groups. It should not be overlooked, that out of the three orders into which Cirripedes may be divided, in the two latter, the mature animal presents hardly any resemblance to an Entomostracan. From the distinct presence in either pupa or mature animal of the fourteen segments of the cephalo-thorax; from the apparent composition of the carapace, as will be subsequently explained; and from the concentrated condition of the nervous system, one is led to glance at the higher Crustacea; and here we shall find amongst the Podophthalmia, one aberrant group of low organisation, namely, that including Phyllosoma, Amphion, &c., in which more points of resemblance to Cirripedes may be detected, than, as I believe, in any other group whatever; for we here see that remarkable elongation of the head in front of the mouth, so eminently characteristic of Cirripedes; we have a carapace overlapping the thorax, which is sometimes free beneath; we have the abdomen sometimes almost obsolete; we have biramous legs: and especially we have the posterior cephalic and the first thoracic appendages more or less rudimentary and obsolete; and this, I infer from Mr. Dana, is a very rare phenomenon, though characteristic of all ordinary Cirripedes, in which the seventh and eighth segments with their appendages have disappeared. In the order including Phyllosoma, &c., namely, in the Macroura, the ganglions which give nerves to the five posterior thoracic limbs, are distinct from the great sub-œsophageal ganglion which supplies the several anterior appendages; this is the case with those Cirripedes in which all the infra-œsophageal ganglions are not concentrated into one. In the Macroura and Brachyoura, the first pair of legs almost always differs in structure from the others, so does the homologous or second cirrus in Cirripedes differ from the four succeeding pairs; in some few Macroura, the second leg is antenniformed, so in some few cases is the homologous (or third) cirrus; J. Vaughan Thompson was even struck by the[Pg 19] resemblance in the curious, doubly pectinated spines on the anterior limbs of Mysis (allied to Phyllosoma[16]), and on those of many Cirripedes: these several latter resemblances may be small, but certainly I do not believe that they are accidental. Now the little group of Crustaceans, which includes Phyllosoma, &c., has lately been placed, by Milne Edwards, as a satellite amongst the Macrourous Podophthalmia; it leads into the Stomopoda, and likewise, as has been noticed by many authors, into the sub-class Branchiopoda, which latter sub-class is considered by Mr. Dana as only a part of the Entomostraca; this group, therefore, exhibits affinities radiating in several directions, and amongst these lines of relationship, one more must, I believe, be added, plainly directed towards the Cirripedia.
[16] M. Martin St. Ange (‘Mémoire sur l’Organ. des Cirripèdes,’ 1835, extrait des ‘Savans Etrangers,’ tom. vi) has compared the mouth of Lepas with that of Phyllosoma, and has given comparative figures; but the resemblance is founded, I believe, on quite false homologies.
One naturally wishes to ascertain how far Cirripedia are highly or lowly organised and developed; but in all cases this, as it seems to me, is a very obscure enquiry. Mr. Dana considers that, in Crustacea, the greater or less centralisation of all the appendages round the mouth is the main sign of high development; on this view, the anterior part of a Cirripede, from being so much elongated, must be considered as very low in the scale; the whole posterior part of the body, on the other hand, is, in ordinary Cirripedes, brought close to the mouth; but this is effected by the abortion of the seventh and eighth segments of the cephalo-thorax and of the whole abdomen, and so, I presume, would not, in Mr. Dana’s estimation, raise the class much in the scale. Von Baer[17] considers that the perfection of the type of any animal is in relation to the amount of “morphological differentiation” which it has undergone; on this view, Cirripedes ought to stand high in the scale, for they differ much morphologically from the type of the class to which they belong; as indeed is shown by the long time that elapsed before their true position, namely amongst the Crustacea, was even suspected; but something more must, I think, be added[Pg 20] to Von Baer’s definition; for, to take as an example the eyes of a Cirripede,—as seen in the first larval stage, there is only one eye, and that most simple; in the pupa there are two, both compound, and furnished with complicated muscles; lastly, in the mature animal there are still two, but of very minute size, often almost confluent, and of the simplest structure; hence, then, there has been much morphological differentiation, but it is almost a contradiction in terms to speak, in relation to such a case, of perfection of type; and what has happened to one organ, might happen to other organs, and so to the whole animal. Lastly, under a physiological point of view, and taking the Balanidæ as the most perfect type of the class, the sub-œsophageal portion of the nervous system is highly concentrated; the organs of sense, excepting the eyes, seem more largely developed than in ordinary Crustaceans; the circulating system is of the simplest kind, being only lacunal; special Branchiæ, however, are developed by the metamorphosis of, as I believe, a special organ, occurring only in the Lepadidæ; the digestive organs are very simple, from not having any distinct liver; the generative system is very low, for both sexes are generally united in the same individual; and the testes and ovaria closely resemble each other. On the other hand, the thoracic limbs are, to a considerable extent, specialised in their structure and functions; only the three posterior pairs strictly resembling each other. Lastly, the dermal and muscular systems are complicated, and not, to use Professor Owen’s term, by mere vegetative repetition, as will be obvious to any one who will study the beautifully constructed and modified carapace—that is the operculum, shell and basis—of a Balanus. On the whole, I look at a Cirripede as a being of a low type, which has undergone much morphological differentiation, and which has, in some few lines of structure, arrived at considerable perfection,—meaning, by the terms perfection and lowness, some vague resemblance to animals universally considered of a higher rank.
[17] English Translation, in ‘Scientific Memoirs,’ 1853, vol. i, p. 228.
It has been seen that I divide the Cirripedia into three orders,—the Thoracica, Abdominalia, and Apoda; between[Pg 21] which the fundamental difference consists in the limbs or cirri being thoracic in the first, abdominal in the second; and entirely absent in the third. For the sake of showing the range of character in Cirripedes, to which allusion has been made, I will briefly indicate the leading differences in each order. In the Thoracica, three families are included,—the Balanidæ, or sessile Cirripedes, the Verrucidæ, remarkable from their quite asymmetrical shell, and the Lepadidæ, or pedunculated Cirripedes. The great difference in external appearance between these three families is known to all naturalists. Even within the one family of Lepadidæ there are great differences in external appearance, as will be admitted on comparison of Lepas, Pollicipes, Conchoderma, &c.; but we have also important internal differences, as in the case of Anelasma, in which the cirri are barely articulated, and are not capable of seizing prey, whilst the mouth is almost probosciformed, with the outer maxillæ and palpi rudimentary: still more important are the differences in Alcippe, in which the cirri of the first pair act as brushes; the second, third, and fourth pairs being quite aborted; and the fifth and sixth pairs consist only of four segments, with one of the two normal rami converted into a crenated, button-like projection, for the sake apparently of triturating food; Alcippe, also, is very remarkable in being destitute of a rectum and anus. In this same genus Alcippe, in Ibla and Scalpellum, there are either separate males or Complemental males, some of which are so utterly abnormal in their characters, that by no definition which I could frame, could they be included even in their proper Order, much less in their proper Family.
In the second order of Abdominalia (Pl. 23 and 24) the seventh or last cephalic segment is quite distinct, and bears rudimentary organs, answering to the first pair of maxillipeds of ordinary Crustaceans, of which organs, and of the segment supporting them, there is no trace in the Thoracica: the seven succeeding thoracic segments are destitute of any appendages; but the three segments of the abdomen bear three pairs of cirri. The mouth is peculiar in the labrum being developed into very large, moveable, lancet-formed organ; and the lower end of the œsophagus is armed with[Pg 22] beautiful discs of teeth, and brushes of hairs,—a structure confined to this order. The male resembles the male of Alcippe; and the latter genus seems to be the connecting link between the Thoracica and Abdominalia. But the most important character of this latter order, in which it differs from Alcippe, and all other known Cirripedes, is in its metamorphoses; all the first changes are merely indicated by changes in form in an egg-like larva, without the development of distinct organs; and the last, or pupal condition, which is attained within the sack of the parent, is very peculiar, by the entire absence of natatory limbs.
The third order of Apoda is the most peculiar of all; it contains, like the last, only one known species: the most acute naturalist, I am convinced, if he had not made the class his special study, would never even have suspected that this animal was a Cirripede. We see much magnified in Pl. 25, fig. 7 a naked, plainly-articulated animal, resembling the larva or maggot of a fly, attached by two threads; and these threads, on analysis, can be clearly shown to be the last rudiment of the carapace, specially modified. The last cephalic, the seven thoracic, and the three abdominal segments, are all equally destitute of appendages. The mouth is suctorial, and constructed on a plan unlike, I believe, anything known in the articulate kingdom; for the mandibles and maxillæ have rotated on their axes, and stand back to back; they can act only by tearing open a slit, and this action is performed in a hood, formed by the confluence of the broad palpi and labrum. Although the œsophagus is distinct, there is no stomach or anus. Lastly, owing to there being no carapace, the ova are developed, differently from in all other Cirripedes, within the thorax.
I will close this preliminary discussion on the confines and type of the sub-class, by recalling attention, now that a sketch has been given of the three Orders, to the remark before made, that a wide range of structure is included within it, and by reurging that the Cirripedia should be ranked, not as one of the subordinate groups, but as one of the main divisions of the Crustacea.
Cirripedes are commonly bisexual or hermaphrodite, but in Ibla, Scalpellum, and Alcippe, members of the Lepadidæ in the order Thoracica, and in Cryptophialus in the order Abdominalia, the sexes are separate. As two of these genera were described in my former volume, and two others (Alcippe and Cryptophialus) are described in this volume, I may as well here give a brief summary of the facts as yet known on this very curious subject. The Males, in the above four genera, present a wonderful range of structure; they are attached in the usual way by cement proceeding from the not-moulted antennæ of the pupa, to different parts, in the different species, of the female. These males are minute, often exceedingly minute, and consequently generally more than one is attached to a single female; and I have seen as many as fourteen adhering on one female! In several species the males are short-lived, for they cannot feed, being destitute of a mouth and stomach. As the females are longer lived, successive crops of males, at each period of propagation, become attached to her. It is the females in the above genera which retain the characters of the genus, family, and order to which they belong; the males often departing widely from the normal type. Some of the males are rudimentary to a degree, which I believe can hardly be equalled in the whole animal kingdom; they may, in fact, be said to exist as mere bags of spermatozoa. So widely do some of them depart in every character from their class, that twice it has happened to me to examine specimens with a little care, and not even to suspect, until a long period afterwards, that these males were Cirripedes.[18]
[18] In my volume on the Lepadidæ (p. 200) in searching for analogies for the permanently epizoic and rudimentary condition of the male Cirripedes, I quoted two cases, which I believe are now known not to be analogous; namely, the Syngamus trachealis of Von Siebold, and the worm-like Hectocotyle, which latter was quite lately supposed to be a male Cephalopod, but has now been ascertained to be only one of the arms of the male wonderfully adapted and organised as a sperm-receptacle. The Asplanchna, the mouthless male of a Rotifer, (p. 292) alone remains for me.
In Scalpellum Peronii, and villosum, the males are but little abnormal, for if classified independently of their sexual relations, they would be considered as immature specimens of a new genus, standing next to Scalpellum; in Scalpellum rostratum, the male would form another and rather more distinct genus. The males, in the latter, are attached to the other sex, between the basal edge of the labrum and the adductor scutorum muscle; but in S. Peronii and villosum they are attached lower down, in the furrow between the two scuta, and are thus protected: in these three species, the internal parts of the male present nothing particular. In Ibla, the males are attached low down within the sack of the female; they may be said to consist of a mouth surmounted on a long peduncle, for there is no capitulum or general covering, and the whole thorax is in a rudimentary condition, the cirri being reduced to two distorted pairs. As these males certainly moult several times and grow a little, they must feed; and as they have no cirri fit for action, they must seize their food by the contortions of their peduncle, which we know homologically consists of the three first segments of the head. The movements of the peduncle must, also, supply those of the probosciformed penis, almost invariably present with other Cirripedes, but here absent. If compelled to class these males without regard to the female, great difficulty would be experienced; we could hardly place in the family of the Lepadidæ, a Cirripede without a capitulum, and without cirri, those very organs which give their name to the class, and with a thorax reduced to the dimensions of a lower lip; yet, if the presence of a peduncle did determine the classifier to place these males amongst the Lepadidæ, then undoubtedly the character of the mouth, &c. would fix their position next to Ibla.
The males of Scalpellum vulgare, ornatum, and rutilum, resemble each other in all essential points, and differ wonderfully in appearance and structure from all ordinary Cirripedes. They consist of a minute flattened bag with a small orifice at the summit, and at the lower end attached by the cemented pupal antennæ. On each side of the orifice, there is a pair of calcareous beads, representing the two[Pg 25] scuta and two terga of ordinary Cirripedes; and between the scuta a minute black eye is generally conspicuous. In S. ornatum the beads, I may remark, on the two sides are not equal; those either on the right or on the left side, being larger than those on the opposite side, so that the animal externally is asymmetrical. Inside, within a tubular sack, the thorax is lodged, supporting four (instead of six) pairs of limbs; and these, instead of forming biramous, multiarticulated, captorial cirri, are reduced to almost a rudiment, supporting a few long sharp spines, which apparently act only as defensive organs. At the end of the thorax there is seated a large abdominal lobe, which does not occur in the other sex. Hence the thorax, though rudimental, has been specially modified. Of the mouth and stomach there is not a vestige. Constructed as these males are, assuredly they have no claim to be ranked amongst the Lepadidæ or pedunculated Cirripedes; nor is it possible to class them in any group whatever of ordinary Cirripedes. In S. vulgare the males are attached, often several together, to the extreme edges of the two scuta, and therefore immediately over the orifice leading into the sack; in S. rutilum and ornatum, they are attached in concavities on the under side of both scuta, just above the depression for the adductor scutorum muscle. In the former of these species, the pit for the reception of the male is formed by shelly matter not having been deposited over a certain space on the under side of the valve; and the pit is converted by a covering of membrane into a pouch. As there are two scuta so there are two pouches, in each of which a male is lodged; hence, according to the Linnean nomenclature, Scalpellum ornatum may be said to belong to Diandria monogynia. As these males, from being mouthless, soon die, they are succeeded by successive pairs; the pupa being led by a wonderful instinct to crawl into the pouch, and there undergo its metamorphosis.
Lastly, the males of Alcippe and Cryptophialus (Pl. 23, fig. 19, and Pl. 24, fig. 19) are remarkable for their similarity to each other, considering the essential dissimilarity of the two females. The females live in cavities which they excavate in the shells of Molluscs, and within which they are attached by a horny disc; this disc is the only part[Pg 26] of the outer integument which is not frequently moulted, and, apparently in consequence, the males are attached to its edges. It results from this position, that the males are protected by being enclosed within the cavity excavated by the female; and it further results, that the males are attached at a considerable distance from the orifice of the sack of the female, into which the spermatozoa have to be conveyed; and to effect this, the probosciformed penis is wonderfully developed, so that in Cryptophialus, when fully extended, it must equal between eight and nine times the entire length of the animal! These males, like those last mentioned of Scalpellum, consist of a mere bag, lined by a few muscles, enclosing an eye, and attached at the lower end by the pupal antennæ; it has an orifice at its upper end, and within it there lies coiled up, like a great worm, the probosciformed penis, and beneath it a single testis, with a single vesicula seminalis. These organs complete the whole organisation of the male; for there is no mouth, no stomach, no thorax, no abdomen, and no appendages or limbs of any kind. Yet all these parts are present in the female. I know of no other instance in the animal kingdom of such an amount of abortion. The whole exterior of these males evidently is composed, as in all ordinary Cirripedes, of the three first cephalic segments; of the fourteen succeeding segments of the archetype Cirripede we have not a vestige, excepting the probosciformed penis, which, from analogy, should arise from the ventral apex of the seventeenth segment, the first three segments of the head being counted in the seventeen. Here, then, fourteen out of seventeen segments have aborted, the tip of the seventeenth having coalesced with the third cephalic segment! I am tempted just to notice the case of Proteolepas, in the order Apoda, as showing, within the limits of the same sub-class, a wonderful amount and diversity in abortion; for in Proteolepas, the three anterior cephalic segments are reduced to the merest rudiment, encasing the cement-ducts, the fourteen succeeding segments being unusually well developed; whereas in the above described males, we have just seen the three anterior segments fully developed, whilst the fourteen succeeding segments[Pg 27] are lost or have coalesced with the others; so that within the same sub-class all seventeen segments of the archetype have almost disappeared.
It may be asked how I know that the several above described rudimentary epizoons are really the males of the Cirripedes to which they are attached. Even if the whole course of the metamorphoses had not been known in three of the cases, the mere fact of these epizoons being cemented by the three terminal segments of their peculiar, pupal antennæ, would have been sufficient to have shown that they belonged to the class of Cirripedes. In nearly every case, I was able to demonstrate, and not in one or two but in many specimens, that these epizoons were males; and as in several instances the spermatozoa were developed, and as, notwithstanding, in no instance was there a vestige of ova or ovaria, it may safely be concluded that they were not hermaphrodites, and therefore required females of some kind. If these epizoic Cirripedes had been independent animals, as they all belong to the same sub-class, and all have such peculiar habits, it might have been expected that they would have shown some special affinity towards each other; but this is not the case; the epizoon of Ibla is more nearly related to Ibla, and the epizoon of Scalpellum more nearly related to Scalpellum, than are these epizoons to each other. If the several epizoons were classed by themselves, they would be grouped in divisions, corresponding with those of the Cirripedes on which they are attached, which is just what might have been expected if these latter were their females. There are, also, many special relations between the male epizoons and the Cirripedes to which they are attached; thus, the mouth of the epizoon of Ibla, is so like the mouth of Ibla, which is peculiar in several respects, that I should easily have recognised it as belonging to a member of that genus. Scalpellum villosum is remarkable as one out of only two or three members of the whole Family, which is destitute of caudal appendages, so is its male epizoon; again, S. villosum is unusually spinose, so is its male epizoon; on the other hand, Scalpellum ornatum is remarkably smooth, so is its male epizoon; and I could give other similar instances. Will it be believed that these[Pg 28] coincidences are accidental, and that the epizoons have no special or sexual relation to the Cirripedes to which they are attached?
One other instance of coincident structure is so important, that it must, even in this sketch, be noticed; the prehensile antennæ of the pupa are most important and complicated organs, and differ in the different genera of the same family; they are preserved in a functionless condition throughout life, and in two instances I was able accurately to compare these organs in the epizoon and in the Cirripede to which it was attached, and they were identical in every particular. The full force of the excessive improbability of this resemblance, and of the above coincidences in structure, on the supposition of the epizoon and its support not being sexually related, will hardly be perceived without referring to the facts given in detail in my former volume.
Lastly, in the case of Cryptophialus (and indirectly in that of Alcippe) the nature of the male epizoon is, I think, actually demonstrated; for I traced both it and the female or ordinary form of Cryptophialus, through the same several larval stages, from the egg, enclosed within the sack of the female, to the pupa and mature animal. Moreover, if the male nature and sexual relation to the supporting Cirripede, be admitted in any one of these epizoons, then so close is the agreement in habits, and to a certain extent in structure, in all the foregoing epizoons, that probably no one admitting one instance would dispute the others, and further evidence would even be superfluous. Indeed, had it not been for the following facts, I should not have brought forward, either here in abstract, or in other places in detail, so many arguments and so much evidence.
I have as yet not entered in detail on the sex of the supporting Cirripede: in Cryptophialus, Alcippe, and in one species of Ibla, I was able to demonstrate in many specimens, that all the male organs, internal and external, were entirely absent; and consequently that these Cirripedes were exclusively female. In Scalpellum ornatum, also, there is no trace of external male organs (the state of the four dried specimens[Pg 29] not allowing the internal organs to be examined), and there cannot be any reasonable doubt that this species likewise is exclusively female. It should be borne in mind that the male organs, external and internal, are most easily discovered, and that in the above cases I had an abundant supply of excellent specimens. On the other hand, in Ibla Cumingii, and in four species of Scalpellum, I was able to demonstrate in the supporting Cirripede the presence of all the male organs, as well as of the female; and in the vesiculæ seminales of several specimens, both in the Ibla and in Scalpellum vulgare, spermatozoa were contained; the male organs, however, not being very amply developed. These species, consequently, are not exclusively female, but are hermaphrodite, though having male epizoons attached to them. This statement, I am well aware, is enough, at first, to cast a doubt on all that I have said; but let any one reflect on the evidence, of which I have here given a summary, and which has been elsewhere given in full, and I think he must admit that at least those epizoons which are exclusively male, and which are attached to Cirripedes exclusively female, are sexually related and form one species; but if he admit this, he cannot possibly escape from the conclusion that some of the other epizoons, for instance that of Ibla quadrivalvis, are the males of the hermaphrodites to which they are attached,—these epizoons not exclusively impregnating the ova of a female, but aiding the self-impregnation of an hermaphrodite. Hence I have called these males Complemental Males, to show that they do not pair with a female, but with a bisexual individual. Nothing strictly analogous is known in the animal kingdom, but amongst plants, in the Linnean class, Polygamia, closely similar instances abound.
In the series of facts now given, we have one curious illustration more to the many already known, how gradually nature changes from one condition to the other,—in this case from bisexuality to unisexuality. Finally, in the four genera so often named, we meet the following several cases, some of them even the most diverse, occurring in closely allied species. (1st), a female, with a single male (rarely with two) permanently attached to her, protected[Pg 30] by her, and capable of seizing, by the movements of its pedunculated body, any minute animals or substances found within her sack; (2d), a female with successive pairs of short-lived, mouthless males, inhabiting pouches on each side under her scutal valves; (3d), a female with many, in one instance fourteen, short-lived males, destitute of mouth, thorax, and appendages, but furnished with a stupendously long male organ, attached to a thickened portion of her outer integuments, but lying within the cavity which she has excavated; (4th), an hermaphrodite with a male attached within the sack, capable of feeding itself, as in the first case; (5th), an hermaphrodite with from one to three males, organised like ordinary Cirripedes, and apparently capable of seizing prey in the common way; and attached between the scuta, and thus protected; (6th and lastly), an hermaphrodite with from one or two up to five or six, short-lived, mouthless males, like those in the second case, attached in one particular spot, on each side of the orifice leading into the sack.
Cirripedia having a carapace, consisting either of a capitulum on a peduncle, or of an operculated shell with a basis. Body formed of six thoracic segments, generally furnished with six pairs of cirri; abdomen rudimentary, but often bearing caudal appendages; mouth with the labrum not capable of independent movements; larva firstly uniocular, with three pairs of legs, lastly, binocular, with six pairs of thoracic legs.
In the sketch of the three Orders given in the Introduction, it will have been seen that the differences in their structure are so great, that it would have been hardly possible to have given a single blended account of the whole Class. But as all common Cirripedes are included in the present Order, here would have been the natural place for a full description of their external and internal structure. Having, however, been necessarily, yet perhaps unfortunately,[Pg 31] led to give, in my former volume, a description of this kind of the Lepadidæ; and as it is necessary to give a similar account of the other great family of the Order, namely, the Balanidæ, I have found it more convenient to make this latter account comparative and supplemental to the former one on the Lepadidæ, and so serve for the Order, rather than attempt to give a separate description in full of it. For this latter plan would have involved much useless repetition, as, on account of the many exceptions and limitations necessary to almost every statement, there is little choice between a description of great length and a mere diagnostic character of the Order, such as I have given above.
The Thoracica may be divided into three very natural Families, of nearly equal value; firstly, the Balanidæ, or sessile Cirripedes, which may be subdivided into two sub-families, also very natural, the Balaninæ and Chthamalinæ; secondly, the Verrucidæ, containing only one genus; and thirdly, the Lepadidæ, or pedunculated Cirripedes. These three families differ from each other, besides in mere external appearance, almost exclusively in the relation of the different portions of their external covering or carapace, and of the muscles moving such portions. In the Balanidæ, the four opercular valves surrounding the orifice leading into the sack, are capable of other movements, besides being opened and shut; whereas all the other valves are immoveably united together. In the Lepadidæ, the valves answering to the opercular valves, are furnished with a muscle only for shutting them; whereas the peduncle answering to the basis in the Balanidæ is capable of various movements. In the Verrucidæ the shell is singularly asymmetrical; only half of the operculum (either the right or the left side, this varying even in the same species) being moveable; the other half being immoveably united to the remaining valves; and the whole shell has only one muscle serving to shut the moveable half of the operculum. All the internal parts and organs are very similar in the above three Families. If, however, the internal structure of one of the two sub-families, into which the Balanidæ may be divided, namely, of the Balaninæ, be compared with that of the Lepadidæ,[Pg 32] several important differences may be detected;—on the one hand, in the Balaninæ, the presence of branchiæ, the extremely complicated cementing apparatus, the difference in structure between the third and succeeding pairs of cirri, the large palpi, the notched labrum, and the laterally double teeth of the mandibles;—and on the other hand, in the Lepadidæ, the presence of ovigerous fræna, caudal appendages, bullate labrum, and often prominent olfactory orifices. But if the Lepadidæ be compared in these several respects with the other sub-family, or Chthamalinæ, which cannot possibly be removed out of the family of Balanidæ, many of these differences break down and disappear, in some or all of the species.
The Lepadidæ include, as has previously been noticed, a much greater range of difference than the Balanidæ; and this is what might have been expected, for it is the most ancient family, and extinction has done its work, separating genera, which, in accordance to analogy, we may suppose were once more nearly connected by intermediate forms. The Lepadidæ, in one sense, may be taken as the type of their order; for they have undergone less “morphological differentiation;” that is, they differ the least from the last larval stage, and seem to give the most general idea of a Thoracic Cirripede. On the other hand, if we mean, as some authors do, by the word type, that form which, in the group in question, has been most modified, and illustrates every peculiarity of its class in the strongest manner, then we must look to the Balaninæ, and to its typical genus, Balanus, for the most Cirripedial form. In this genus the different portions of the carapace differ most, and subserve to a certain extent different ends, and in minute structure are most complicated; here the cementing apparatus, which offers the main characteristic of the whole sub-class, is most complex; here the several pairs of cirri differ most from each other in structure and action; here the peculiar branchiæ (organs apparently derived from the modification of another organ, itself confined to Cirripedes, viz., the ovigerous fræna) are best developed; here the nervous system is most highly concentrated; and, lastly, here we meet with the largest and most massive species of the whole group.
Cirripedia without a peduncle; scuta and terga furnished with depressor muscles; other valves united immoveably together.
| Page | |
| Structure of shell | 34 |
| ”of the individual compartments | 43 |
| ”of the radii | 45 |
| ”of the alæ | 47 |
| ”of the sheath | 48 |
| ”of the basis | 49 |
| ”of the opercular valves (scuta and terga) | 51 |
| Growth of whole shell and microscopical structure | 54 |
| Muscles of sack | 61 |
| Branchiæ | 63 |
| Thorax and body | 65 |
| Muscular system | 68 |
| Movements and muscles of the cirri | 71 |
| Mouth | 74 |
| Cirri | 81 |
| Caudal appendages | 85 |
| Alimentary canal | 85 |
| Circulatory system | 87 |
| Nervous system | 88 |
| Eyes and vision | 93 |
| Acoustic organs | 95 |
| Olfactory sacks | 97 |
| Male organs of generation | 97 |
| Female organs of generation | 100 |
| Metamorphoses and homologies | 102 |
| Larva, first stage | 103 |
| Larva, second stage | 109 |
| Larva, last or pupal stage | 110 |
| Act of metamorphosis | 126 |
| On the homologies of the carapace | 131 |
| Cementing apparatus | 133 |
| Affinities, classification, variation | 152 |
| Rate of growth, exuviation, &c. | 156 |
| Geographical range and habitats | 159 |
| Geological history | 172 |
Almost every one who has walked over a rocky shore knows that a barnacle or acorn-shell is an irregular cone, formed generally of six compartments, with an orifice at the top, closed by a neatly-fitted, moveable lid, or operculum.[19] Within this shell the animal’s body is lodged; and through a slit in the lid, it has the power of protruding six pairs of articulated cirri or legs, and of securing by their means any prey brought by the waters within their reach. The basis is firmly cemented to the surface of attachment. The whole shell, basis, and operculum consists, as we have already seen, of the first three segments of the head, modified into a singularly constructed carapace, which encloses[Pg 34] the mouth and rest of the body. The anterior extremity of the shell is situated in the centre of the basis, where indeed, by due care, the antennæ of the pupa may be always detected; and the posterior extremity is directed vertically upwards.
[19] The best published description of the structure of the shell of a sessile Cirripede, is given by Dr. Coldstream, in the ‘Encyclopædia of Anatomy and Physiology,’ article ‘Cirrhopoda.’
When the shell of a sessile Cirripede or barnacle, for instance, of a Balanus, is first examined, the structure appears extremely complicated; but this can hardly be considered as really the case. The structure will, I think, be best understood by recalling to mind that of Pollicipes,—the oldest known genus, from which, in one sense, all ordinary Cirripedes, both sessile and pedunculated, seem to radiate. I must premise, and the fact in itself deserves early notice, that the homologies of the several parts in the pedunculated and sessile Cirripedes admits of no doubt,—that is, if amongst the pedunculated, the genus Pollicipes, or certain species of Scalpellum, be taken as a standard of comparison.[20] The peduncle corresponds with the basis, as may be clearly seen, if a Pollicipes with a short peduncle, and a Balanus, with a deep cup-formed or cylindrical basis be compared, for the contained parts are similar, and both grow at their upper edges upwards and outwards. Secondly, the valves round the lower part of the capitulum of a Pollicipes, though generally much more numerous, and forming more than one whorl or circle, and not so closely packed together, answer to the compartments forming the shell of a sessile Cirripede; this is shown by their lateral and downward growth, by their upper ends generally projecting freely above the cavity in which the animal’s body is lodged; and in the case Pollicipes mitella, by an actual resemblance in outline, some being triangular, some broad at the upper end, and some sub-rhomboidal, and, lastly, in the manner in which they slightly overlap and indent each other: moreover further resemblances in the relative position and even in the size of the several valves, will hereafter be pointed out between certain sessile genera amongst the Chthamalinæ[Pg 35] and certain genera of the Lepadidæ. Thirdly, the scuta and terga in Pollicipes, so strikingly resemble in manner of growth in position relatively to the animal’s body—in shape—and even in being articulated together, the valves which form the operculum or lid of sessile Cirripedes, that their identity is at once obvious.
[20] Dr. J. E. Gray long ago observed these homologies. If Lepas be taken, the comparison is not quite so simple, owing to the growth of all the valves in that genus being upwards; but in several species of Scalpellum we may see the intermediate steps between the normal downward growth of the valves in Pollicipes, and the abnormal upward growth in Lepas.
It may be well here further to premise, that apparently none of the sutures in the shells of Cirripedes correspond with the articulations between the three archetype cephalic segments, of which the whole shell is formed; or with the eight elemental pieces, of which each separate segment in the archetype crustacean is known to consist. But, as I believe, the several valves in the shell of a Cirripede are homologous, or at least analogous, with the sclerodermic plates,[21] of which the carapace of the Podophthalmia is formed; with this difference, that in the latter they become, after their first formation, united together into a single piece, and are thus moulted as a whole; whereas in Cirripedes, the valves or sclerodermic plates are not moulted, but continue to be added to throughout life.
[21] Milne Edwards, ‘Annales des Sciences Naturelles,’ tom. xviii, (1852), p. 236.
In Pollicipes, there is no difficulty in understanding the growth of the lower valves of the capitulum, especially if a species be taken in which these valves stand a little way apart: at each period of growth, they are added to at their basal edges and a little way up both sides; at the same time, a new membrane connecting them together is formed, the old membrane disintegrating, or being left hanging in tatters to the last zone of growth. Now if we look at the shell of a sessile Cirripede, there is no essential difference in the growth of the compartments or valves; all grow downwards and laterally; but they overlap each other much more laterally than in Pollicipes, and the connecting membrane is in most parts reduced to a mere film jammed in between the valves; but, in the case of the opercular membrane, it still remains wide, and is periodically moulted.
In the annexed woodcut (fig. 1), of the rostrum of Balanus Hameri, the downward growth and the lateral[Pg 36] growth on both sides is plain. The modified sides (rr) for convenience sake, have been called the radii; they invariably overlap the adjoining compartments. The middle part (p), has been called the wall, or parietal portion: in the specimen figured, the walls and radii are distinctly separated, but in some cases, especially amongst the Chthamalinæ, the lines of growth are absolutely continuous from one to the other. In fig. 2 of a Lateral compartment of the same Balanus, we have the same essential structure; but the left side (a) is more protuberant, and is hollowed out in its lower half; it is, also, more distinctly separated from the parietal portion: this side has for convenience been called the ala; it is invariably overlapped by the adjoining compartment: in some few cases, as in Pachylasma, the ala is not hollowed out in its lower part, and from being added to in a straight line along its whole edge, with the lines of growth continuous with those on the wall, it differs hardly at all in appearance from a radius. Lastly, in fig. 3 of the carina, or compartment facing the rostrum, we have alæ (aa) on both sides; these being, as in all cases, overlapped by the adjoining compartments.
p, p, Parietes; r, r, Radii; a, a, Alæ.
Fig. 1, Rostrum with two radii, serving in the Chthamalinæ for rostro-lateral compartments.
Fig. 2, always serving for lateral and carino-lateral compartments.
Fig. 3, Carina, serving in the Chthamalinæ, also, as a rostrum.
Now, the compartments in the shell of every sessile Cirripede, are without exception constructed on the above three simple patterns. In number, they are 8, 6 or 4, or all confluent together.
Considering this simplicity in growth and form of the separated compartments, it seems at first surprising that[Pg 37] the construction and enlargement of the whole shell in Balanus, should long have been viewed as a difficulty. But the radii, from growing against rectangular indentations, or rather furrows, in the opposed compartments, come to be set a little inwards; and their external surfaces assume a very different appearance from the wall-portions of the compartments, which grow against the surface of attachment. In different species, the summits of the radii (and of the alæ) grow either very much more obliquely than in the species figured, or more squarely—that is, they extend from tip to tip of the adjoining compartments, parallel to the basis. In this latter case, and when the surfaces of the radii differ considerably in appearance from the walls, as in Balanus tintinnabulum (Plate 1), I am not at all surprised that the radii should have been described as separate elements, and called “areæ interjectæ,” or “compartments of the second order:” for the shell of this Balanus seems to be composed of six wedges with their points upwards, namely, the parietal portions of the compartments, and of six other narrower wedges, the radii, with their points downwards; and the fact that these latter wedges consist simply of the sides of the parietal portions, modified by growing against the adjoining compartments, is completely masked. I should add, that sometimes the radii are not developed, which simply means that the overlapping lateral edges of the compartments have not been added to during growth.
The alæ are originally developed at the period of the metamorphosis, as slight lateral protuberances in the upper part of the compartments; from being overlapped, and therefore not exposed to external influences, and from growing (as in the case of the radii) against rectangular indentations or furrows in the adjoining compartments, they generally assume an extremely different appearance from the parietes, and might naturally be thought to have a very different nature. But the alæ in all cases (as is obvious in Pachylasma) are nothing but the protuberant lateral edges of the compartments, rendered thin and modified during growth. In order that the margins of the alæ should be received in an indentation, the upper internal surfaces of the walls of the[Pg 38] recipient compartments are thickened all round, excepting where they receive the alæ. This thickened, upper, internal portion of the walls or shell, together with the alæ themselves, form the part called the sheath. The sheath sometimes blends insensibly into the lower parts of the compartments, and then perhaps it would not be thought to be a distinct element; but often it is abruptly separated by an overhanging edge (see Pl. 9, fig. 5 b, 9 b; Pl. 20, fig. 1 b; Pl. 25, fig. 1, K′) from the lower part, and then the sheath greatly complicates the internal appearance, but not the essential structure of the shell. The sheath acts beautifully, like an internal hoop, in strengthening the shell round the orifice, where it is naturally weaker than at the lower or basal end, where it adheres to the surface of attachment: in the upper part of the shell, moreover, the sutures between the compartments do not go straight through, but owing to the alæ projecting and being overlapped, are extremely oblique; or the joints, in the language of carpenters, may be said to be broken.
There is one other point of structure in the shells of the Balanidæ, more especially of species like Balanus tintinnabulum, which adds to their apparent complexity, namely, that the rim or orifice of the shell formed by the upper ends of the compartments, projects considerably above the opercular valves. In a young Balanus, immediately after the metamorphosis, the operculum is attached by the opercular membrane all round to the summits of the compartments, and there cannot be said to be any orifice to the shell itself, but only an orifice or slit between the opercular valves; but during growth, as the compartments are added to at their basal edges, their upper ends are deserted, and cease to enclose the sack, within which lies the animal’s body. Hence the upper ends come to project freely, either quite separately as in some species of Pollicipes, where they cannot be said to form an orifice; or more or less united into a ring so as to form an orifice, as in the different species of Balanidæ. It follows, that to understand the real shape of a Balanus, or rather of the cavity enclosing the animal’s body, all that part of the shell which projects above the opercular membrane, may, in imagination, be removed as[Pg 39] something extraneous, like so many spines; not that I mean to say that these points of shell are dead; on the contrary, they are often porose and penetrated by numerous threads of corium. This upper part of the shell, thus produced so as to form an orifice, no doubt serves to protect the less strong and moveable operculum.
a, Rostrum; b, Rostro-lateral, c, Lateral, d, Carino-lateral compartment; e, Carina.
Horizontal sections through the Shells of the principal genera of Balanidæ, showing the arrangement of the Compartments. Genera 4, 5, and 6 belong to the Chthamalinæ; 7 and 8 to the Balaninæ.
Number and Arrangement of the Compartments.—I have already stated that the shell, in every one of the Balanidæ, consists of eight, six, or four compartments, or of all fused together into a single piece; and that the compartments themselves are all constructed on the three simple patterns of which woodcuts (figs. 1, 2, 3) have been given. They are arranged in a certain definite order. The type arrangement is found amongst the Chthamalinæ, as might have been expected, inasmuch as this sub-family is so closely related to the ancient genera Pollicipes and Scalpellum, whence all the Thoracic Cirripedia may be said to radiate. In Octomeris (fig. 4) the type-arrangement of the compartments, eight in number, is well shown; the rostrum and carina resemble each other, and have alæ on both sides, and therefore are overlapped on both sides: the[Pg 40] rostro-lateral compartments have radii on both sides, and therefore overlap the adjoining compartments on both sides; the lateral and carino-lateral compartments have radii on their carinal, and alæ on their rostral sides; and therefore overlap on one side, and are overlapped on the other side. Now the shell of every other sessile Cirripede differs, I believe, from that of Octomeris, only in the fusion together or abortion of some of the eight normal compartments: in one genus, however, Catophragmus, outer whorls of small compartments, arranged like the lower valves in the capitulum of Pollicipes, are superadded. The genus Chthamalus (fig. 5) differs from Octomeris only in the carino-lateral compartments being aborted, (as will presently be discussed), and hence has six compartments. Chamæsipho (fig. 6) differs from Chthamalus only in the rostro-lateral and lateral compartments being fused together; and hence has only four compartments. In Balanus (fig. 7) and the whole sub-family of the Balaninæ, the rostrum is compounded of the true rostrum, as seen in the type Octomeris, and of the two rostro-lateral compartments; hence this compounded rostrum has radii instead of alæ on both sides, and there are only six compartments. Tetraclita (fig. 8) and Elminius differ from Balanus only in having the carino-lateral compartments absent, and probably aborted; hence there are only four compartments. Lastly, in Pyrgoma, all the compartments are blended together into a single piece.
In Pollicipes, the old type-form of the whole order, and in Scalpellum, we have four valves, (answering to the operculum), surrounding the aperture leading into the sack, and the valves below are arranged in successive whorls, with a strong tendency to alternation. For, the rostrum alternates with, that is faces the interval between, the two scuta; the carina alternates with the two terga; and the upper lateral valves alternate with the scutum and tergum on each side. These four valves, namely, the carina and rostrum, which resemble each other in structure, and the pair of upper latera, which are larger than the other lateral valves, together form the uppermost whorl, or that beneath the scuta and terga. In the next whorl we have the[Pg 41] rostro- and carino-lateral valves, alternating with those above them; and beneath them there are generally other valves, which decrease in size and display the same tendency to alternation. The valves here just specified, namely, the rostrum, carina, and three pairs of lateral valves, in the Lepadidæ, are so much larger, and are so much more commonly present, than the other valves of the capitulum, that to them alone I affixed special names. Now if amongst sessile Cirripedes we look to that genus, viz., Catophragmus, which comes in its whole structure the nearest to Pollicipes, one of the Lepadidæ, we find (as in fig. 4), firstly, a rostrum and carina resembling each other, and a pair of lateral compartments, larger than the other lateral pairs; these four valves alternating with the opercular valves: and, secondly, we find, but forming part of the same whorl, a pair of rostro-lateral and a pair of carino-lateral compartments, which, just as in Pollicipes, are larger than the exterior and lower valves. These lower little valves, I may remark, decrease in size in the successive whorls, and tend to alternate in position, just as in Pollicipes. Observing these several striking points of correspondence in the valves, (and indeed in the whole structure), of Catophragmus and Pollicipes, one is strongly inclined to suspect that in Catophragmus, and therefore in Octomeris and other sessile Cirripedes, although the rostro- and carino-lateral compartments appear to lie in the same whorl with the rostrum, carina, and large lateral compartments, yet that they really belong, as in Pollicipes and Scalpellum, to a lower whorl. Now if a very young shell of Balanus, immediately after the metamorphosis, be examined, the carino-lateral compartments will be found not to have been developed; they first appear after two or three zones of growth have been added to the other compartments; bearing in mind that in Pollicipes and in Catophragmus the lower whorls are added successively during growth, we find in this fact strong confirmation of the view that the carino-lateral compartments normally belong to a whorl beneath that including the rostrum, carina, and lateral compartments. Whether the rostro-lateral, like the carino-lateral compartments, are developed subsequently to the others, I have had no opportunity of ascertaining, and[Pg 42] therefore cannot confirm the above analogical conclusion, namely, that they, also, belong to a lower whorl.
In the sub-family Balaninæ, which includes Balanus (woodcut 7), and Tetraclita (woodcut 8), the shell is characterised by not having rostro-lateral compartments, and by the rostrum having radii: now in Pachylasma giganteum, which undoubtedly belongs to the sub-family Chthamalinæ, at a very early age the rostro-lateral compartments become blended with the true rostrum, making a compound rostrum, exactly like the rostrum in the Balaninæ; distinct evidence of a similar fusion is retained throughout life (Pl. 15, fig. 1) in all three species of Chelonobia, which is undoubtedly a member of the Balaninæ. Hence, I think, we may conclude that in all the genera of the Balaninæ the rostro-lateral compartments are probably not aborted, but are blended with a normal rostrum (resembling that in woodcuts 4, 5, 6), making together a compound rostrum furnished with radii: it must, however, be observed that I could not detect any actual evidence of this fusion in Balanus, even immediately after the metamorphosis. In Chamæsipho (woodcut 6), either the rostro-lateral compartments attain a most unusual breadth, or, as is more probable, they have become confluent with the lateral compartments, which in the Lepadidæ seem to be the most persistent of all the lateral valves. In such genera as Tetraclita and Chthamalus, in which the carino-lateral compartments are absent, they may be fused with the lateral compartments or with the carina; but seeing that they are normally developed later than the other valves, it appears to be the simplest theory to assume, until the contrary be proved, that they are aborted. Finally, the somewhat unexpected conclusion that the shell (not including the operculum) of sessile Cirripedes normally consists of eight valves, four belonging to an upper whorl, and four to a lower whorl, all forced into a single ring, and often more or less fused together, though not strictly proved, is rendered highly probable. I will only further add, that the Basis perhaps represents several whorls of the small valves or scales on the peduncle of Pollicipes, fused together; the comparison of the basis with the calcareous cup, forming the lowest portion of the peduncle in Lithotrya,[Pg 43] which has been made by some authors, I do not think is very accurate, as the cup in Lithotrya seems to have a special relation to the boring habits of that genus.
Fig. 9.
Basal edge of wall of compartment in Balanus tintinnabulum; a, a, outer lamina; b, b, inner lamina; c, c, longitudinal septa uniting the inner and outer laminæ with their ends denticulated.
If the basal margin of a compartment, for instance, of Balanus tintinnabulum, be examined, it appears sufficiently complicated, being composed of an outer and inner lamina, separated by longitudinal septa, which are denticulated at their bases; and the tubes formed by these longitudinal septa are crossed by transverse septa. On the other hand, in some cases, as in the genera Chthamalus and Elminius, each compartment consists of a simple shelly layer. These two extreme states graduate into each other: we have, firstly, on the internal surface, quite irregular points and ridges; these become regular, causing the internal surface to be longitudinally ribbed; then these ribs themselves become finely furrowed on their sides and at their lower ends, producing sharp, minute ridges, the ends of which I have called the denticuli; and, lastly, some of the denticuli on the adjoining longitudinal septa become united into a solid layer, forming the internal lamina of the wall. These denticuli do not generally cover the whole surface of the longitudinal ribs, but leave a portion near the outer lamina of the compartment smooth. The denticulated ends of the longitudinal septa project beyond the basal edge of both the outer and inner laminæ, and enter the mouths of the tubes (where such occur) in the basis, and thus strengthen the shell. The whole of the internal lamina generally is more or less striated longitudinally, thus displaying its origin from the union of the inner edges of the longitudinal septa. I need only further remark that on the internal surface of the outer lamina, between the main longitudinal septa, there are generally (as in the woodcut) smaller longitudinal ridges, which do not reach the inner lamina, and on this account alone are not called septa.
Tubes are formed by the longitudinal septa, between the outer and inner laminæ. These tubes are almost square, and are occupied by threads of corium, which enter at pores left open between the edge of the compartment and that of the basis on which it rests. The tubes extend high up the compartments; but in the uppermost part they are generally cut off by thin, transverse, calcareous septa, deposited by the ends of the threads of corium; a cancellated structure being thus produced. Or the uppermost part of each tube becomes filled up solidly with compact shelly layers, which are always first thrown down on the side of the tube facing the outside, and thus greatly strengthen the shell: in several instances, as in Balanus perforatus and Tetraclita porosa, in which the disintegration of the upper part of the shell is a necessary element in its growth for the enlargement of the orifice, these filled up tubes become exposed. In Coronula and Tubicinella, the tubes in their upper parts are, I believe, crossed only by transverse membranous septa.
Anomalies and exceptions.—In Tetraclita (Pl. 10, fig. 1 g, 1 h) from the branching of the longitudinal septa, several irregular rows of tubes are formed. In certain varieties of Balanus balanoides (Pl. 7, fig. 2 b), and in B. cariosus (Pl. 7, fig. 3 b), slight branching ridges on the internal surface of the walls, seem to answer to the longitudinal septa, and produce, during the downward growth of the shell, extremely irregular cells, and short tubes. In Balanus vinaceus (Pl. 2, fig. 7 d), the internal lamina, instead of being solid, as in every other species, is left cancellated, and thus betrays, much more plainly than usual, its origin in the united denticuli of the adjoining longitudinal septa. In Balanus porcatus, between the main longitudinal septa, there are (Pl. 6, fig. 4 e) what may be called rudimentary and disconnected longitudinal septa. In Coronula and its allies (Pl. 16, fig. 6, and Pl. 17, fig. 4 c) it is the outer lamina of the compartment which is anomalous; for in the two or three lower zones of growth, it forms only a ledge on each side of the longitudinal septa; which ledges, higher up, become confluent, and so form an ordinary outer lamina. In Coronula, also, the wall of each compartment[Pg 45] (see transverse section, Pl. 16, figs. 5, 7) is very remarkable from being deeply folded, the folds being on their internal faces firmly calcified together, and on their external faces closely pressed together (often with a neatly serrated suture), so that the whole nature of the shell might be, as has happened, easily quite misunderstood; and the walls be considered as very thick, instead of being, as is really the case, very thin. In Chelonobia (Pl. 15, fig. 1), however, the walls are truly of such great thickness, that the nature of the relative parts might likewise be misunderstood; in this genus the ovarian tubes enter the walls, extending up between the longitudinal septa, or, as they may here be more naturally called, the radiating septa. I will specify a few more peculiarities worthy of remark:—in some species of the sub-genus Acasta, clefts are left, covered only by membrane, on the lines of suture (Pl. 9, figs. 7 a, 8 a), between the compartments, just above the basis; and in other species the basis is perforated by numerous membrane-covered, minute orifices. In Platylepas, each compartment has one deep inward fold (Pl. 17, fig. 1), somewhat analogous to the three folds in Coronula; this fold is produced into an internal midrib, supporting and rendering convex the membranous basis; in this genus, also, the rostrum, owing to its midrib, is generally thrust a little on one side, and the shell thus rendered asymmetrical. In Chamæsipho scutelliformis the shell is symmetrically perforated (Pl. 19, fig. 4 a) by four apertures. Lastly, in Chthamalus Hembeli and intertextus, after a certain age, the basal edges of the walls become inflected, and continue to grow inwards till they entirely take the place of the true membranous basis.
Radius.—This term, as we have seen, is applied to that side of the compartment, the growth of which is modified, by abutting against and overlapping the adjoining compartment. Hence the structure of the radius is essentially the same with that of the parietal portion of the compartment. When best developed, as in Balanus tintinnabulum, the radius consists of an outer and inner lamina, separated by denticulated septa, extending in horizontal lines parallel to the basis, and is consequently perforated by minute tubes or[Pg 46] pores. The tubes become filled up solidly much more commonly than do the parietal tubes; and the inner lamina, in such cases, is hardly distinct from the outer lamina. The denticuli often fail, or are present only on the lower sides of the septa; and very frequently the edge of the radius can only be said to be crenated. Notwithstanding these frequent anomalies, if a series of species and genera be taken, it is certain that there is, as might have been expected, a close relationship in internal structure, between the radii and the parietes. The edge of the radius is received in a slight furrow (generally marked like a seal, with the impression of the denticulated septa) in the opposed compartment: sometimes the outer edge or lid of the recipient furrow, is so broad as to give the false appearance of a radius having been developed, at least in the lower part of the shell, on both sides of the suture. A crest of corium runs into each suture between the edge of the radius and the furrow in the opposed compartment; and when the radius is permeated by pores (as in woodcut 10), threads of corium branch off this crest, and enter the pores. In the lower part of the shell, these crests of corium project from the corium forming and surrounding the sack; but in the upper part of the shell, above the opercular membrane, and therefore above the sack, the corium is produced up each line of suture as a separate ribbon. In proportion as these ribbons extend more or less near to the summit of the shell, so do the edges of the radii continue to be added to, to a greater or less height from the basis; and consequently their summits become less or more oblique.
Fig. 10.
Edge of the radius of Balanus tintinnabulum. a, outer lamina; b, inner lamina; c, denticulated septa, uniting the two laminæ.
Peculiarities in the Structure of the Radii.—In some of the species of Tetraclita, in which genus the walls consist of several rows of tubes, the radii are likewise perforated by several rows; and in some of the other species (Pl. 10, fig. 1 h), the edge, or disarticulated surface of the radius, is marked by irregularly branching ridges; and these evidently correspond with the branching septa or ridges of[Pg 47] the wall. In Chelonobia, the outer lamina of the radius, as well as of its recipient furrow, is of extraordinary thickness; and this lamina, in C. testudinaria (Pl. 14, fig. 1 a, 5, b, and Pl. 15, fig. 1, f), is modelled into sharp transverse ridges and valleys. In the Chthamalinæ, the radii, like the parietes, are simply solid; and apparently in consequence, for the sake of strengthening the sutures, the edges of the radii, and of the recipient furrow in Octomeris (Pl. 20, fig. 3 a) and in Chthamalus dentatus and Hembeli (Pl. 18, fig. 3 b, 5 a), are neatly dentated. In some other species of Chthamalus (Pl. 19, fig. 1 a), the radii present a slight modification of this structure, the sutures being formed by oblique interfolding laminæ. In the radii of Coronula and Tubicinella, there is a peculiarity, in apparent connection with the fact, that in these genera the parietal tubes are not crossed by transverse calcareous septa, namely, that the pores by which the radii are permeated keep unclosed throughout their length, and open into a special longitudinal tube (Pl. 16, fig. 7, d′), which runs along that margin of the wall, whence the radius arises. In Coronula the wall is of extreme thinness, and in conformity so is the true radius, but that the shell might not thus be rendered very weak, complementary or pseudo-radii are developed on their inner sides (Pl. 16, fig. 7, adjoining the true radii A d, C d, and shaded by distant convex lines). Even in the allied genus Xenobalanus, in which the whole shell tends to become rudimentary, traces of these pseudo-radii (Pl. 17, fig. 4 b, d) can be detected. In Coronula, though the radii (Pl. 16, fig. 7, A d, C d) are, by the above special means, rendered thick, and though the alæ also are thick (C a′, D a′), yet together they do not equal in thickness the folded walls; and consequently, there is left between the radii and alæ square chambers (v), occupied by the branching ovarian tubes.
These project, generally abruptly, from the sides of the upper part of the compartments; they appear from the first growth of the shell; they are overlapped by the radius and by part of the wall of the adjoining compartment; they are thinner, and have, owing apparently to being overlapped, a very different aspect from the parietal portion; but they do not differ from it in essential nature. [Pg 48] They are solid, that is, they are never permeated by pores; but their edges are generally crenated, and there is, in some cases, as in Chelonobia, sufficient evidence that these crenations answer to the horizontal septa on the edges of the radii (also often reduced to mere crenations), and consequently, likewise, to the longitudinal septa of the parietes. In Coronula the edge of each ala consists of a medial ridge, sending off denticulated septa on both sides, and is therefore anomalous as compared with the alæ in other genera, but corresponds in structure with the similarly anomalous radius of Coronula. In order to allow of the growth of the edge of the ala, a fine thread of corium runs up the narrow furrow in which the edge is lodged, proceeding from the corium of the sack. In proportion as this thread runs up higher or lower, so are the summits of the alæ rendered, during growth, less or more oblique.
As the compartments overlap each other, the edges of the alæ would have projected, and the inner surface of the orifice of the shell would not have been smooth and rounded, had not that part of each wall, which does not overlie an ala, been thickened so as to allow of the formation of a shoulder or indentation, against which the edge of the ala fits and abuts. The thickened portions, and the alæ themselves, together form the sheath, of which the use seems to be to strengthen, like a broad internal hoop, the upper part of the shell round the orifice, where naturally it is weak. The sheath is composed of successive, fine, shelly layers, which extend, as the shell is added to at the basal margin, lower and lower down on the inner surface of the walls. The lower edge of the sheath either simply projects a little inwards, or more commonly is formed into a sharp depending ridge, as represented in fig. 1, K′, Pl. 25. In some species of Pyrgoma (Pl. 13, fig. 2 b), the sheath reaches nearly to the bases of the compartments; and in Chelonobia (Pl. 14, fig. 4 e c e), the inner layer of shell surrounding the sack, which seems to correspond more nearly to the sheath than to the inner lamina of the walls, actually rests on the basal membrane. The opercular membrane is generally, but not invariably, attached only a little way above the lower edge of the sheath: at each exuviation, a new opercular[Pg 49] membrane is formed, and is attached to the next lower zone of the sheath; the old membrane being cast off, but a circular slip of it is left, investing the last zone. Hence the whole upper part of the sheath above the opercular membrane, comes to be thus invested; and is marked by circular lines, one above the other, caused by the successive exuviations. This investing membrane often supports rows of minute bristles, directed upwards. Generally, a film of shell is deposited, at the period of the formation of each new opercular membrane, on that part of the sheath which lies immediately beneath. This innermost film or thin layer of shell, on the lines of suture between the compartments, breaks joint, at least in some cases, with the underlying shelly layers,—that is, the suture in this last-formed film does not lie exactly over the suture in the subjacent layers of the sheath. In Tubicinella, the sheath extends down close to the basis; and what is unique in this one genus, the opercular membrane, gradually thinning out downwards, closely adheres to the whole inner surface of the shell. In Tubicinella and in Xenobalanus (Pl. 17, fig. 4 b), the sheath separates easily into separate successive rims of shell; and this structure evidently is for the sake of facilitating the breakage of the upper end of the shell, which, as we shall presently see, is necessary to allow of the increase in size of its orifice.
This, in several genera and species, is composed of simple membrane, and consists of successive, concentric, circular slips, added round the outside, at each period of growth. In some species of Tetraclita and Balanus the basis is calcareous, but diaphanous, very thin, smooth, or somewhat granulated. In other cases it consists of a single calcareous lamina, either smooth, or with ridges radiating from its centre; it is formed of two laminæ, (as is most usual in Balanus,) separated by radiating septa. These septa, as well as the radiating ridges in the case of the single lamina, are homologous with the longitudinal septa of the parietes. The denticulated ends of the latter enter the mouths of the tubes formed by the radiating septa of the basis: threads of corium pass between the denticuli of the parietal septa, and thus enter the basal tubes. The ends of these threads of[Pg 50] corium generally deposit transverse calcareous septa, exactly as within the parietal tubes. When the basis is thick the septa themselves (ccc) between the proper basal tubes, become porose, (or rather cancellated,) and they sometimes expand into a very thick, cancellated layer, separating the outer lamina (a) of the basis from the proper basal tubes, which always lie close under the inner lamina (b). This structure differs only slightly from that seen in the parietes of Tetraclita, in which the branching of the longitudinal parietal septa, produces thick walls, formed of several rows of tubes or pores. With respect to peculiarities in structure of the basis, Balanus lævis offers the most remarkable case; for here, in specimens which have grown crowded together, the whole interior appears sometimes to have become too much elongated or too deep for the animal’s body, and consequently the lower part of the deeply-concave basis has been filled up (Pl. 4, fig. 2 a) by thin, irregular, calcareous diaphragms. In elongated specimens, also, of Balanus balanoides, the shell sometimes appears to have grown too long for the animal’s body; but in this case the membranous basis becomes extremely convex inwards; it still reaches the basal edges of the parietes all round, but in the middle it is raised high above the surface of attachment; yet sometimes threads of the cementing tissue depend from the middle part to the surface of attachment. In Balanus terebratus (Pl. 8, fig. 2 a, 2 b), and in some species of Acasta, the basis is riddled, as previously stated, by numerous, minute, membrane-covered orifices. In B. declivis the membranous basis is always extremely oblique, owing to the rostral end of the shell being twice as high as the carinal and opposite end.
Fig. 11.
Portion of edge of basis of Balanus tintinnabulum, a, a, outer lamina; b, b, inner or upper lamina; c, c, c, porose or cancellated radiating septa.
Regarding the very remarkable means by which the basis of sessile Cirripedes is cemented to the surface of attachment, it will be convenient to defer for a little the description, on account of its necessary length.
These are situated on each side of the slit or orifice leading into the sack; from their shape, their powers of movement, their separation by flexible membrane from the shell, to which they serve as a lid, they appear at first as if they constituted an element very distinct from the shell itself, but this is not the case. They are, together with the opercular membrane, as essentially as the whole of what is externally visible, a part of the modified carapace, of which they occupy the upper or posterior extremity: from tracing the metamorphoses, or even by comparison of a Balanus with Pollicipes, there can be no doubt of the truth of this conclusion. The opercular valves are four in number,—a pair of scuta and a pair of terga; but the latter in Coronula diadema and reginæ, are either aborted or represented by a mere rudiment; and in Xenobalanus both scuta and terga are quite absent. In several cases, more especially in the genus Pyrgoma (Pl. 13, fig. 1 b), the scutum and tergum on each side are calcified together, so that sometimes not even a trace of the line of junction can be discovered. In most cases the scutum is firmly united, being articulated in a manner presently to be described, to the tergum; but in Coronula, Tubicinella (Pl. 17, fig. 3 c), and Platylepas, the ends of these valves are simply approximated.
Scuta.—These valves are important, inasmuch as the animal’s body is attached to them; in Pl. 25, fig. 1, the broken line, surrounding a, b, shows where the body has been cut, in removing the scutum on the near side, the other scutum, S, being left articulated to the tergum, T. In shape the scuta are generally sub-triangular; but in some species of Pyrgoma and in Chelonobia, &c. they are much elongated. The lines of growth are usually prominent; and along the occludent margins the alternate, or sometimes every third or fourth line, is developed into a knob, which produces a serrated edge, serving to lock the two opposed valves together; there is, however, no trace of this structure in Coronula and Tubicinella. In some species of Pyrgoma, a ledge of considerable breadth (Pl. 13, fig. 3 e, &c.) is developed along the occludent margins of the two scuta, as well as of the two terga, giving them an anomalous struc[Pg 52]ture. The Terga differ considerably in outline in the different genera and species: their shape approaches more nearly to a triangle than to any other regular form; but there is generally a projection or spur on the basal margin, on the side towards the scutum. In some species of Pyrgoma, the tergum is of so irregular a shape as to defy description. In most cases, a longitudinal depression or furrow runs down the valve, from the apex to the extremity of the spur; and it not rarely happens that the sides of this furrow become folded inwards and almost closed. The spur probably answers to the basal point of the usually sub-rhomboidal tergum in Pollicipes and Scalpellum.[22] The tips of the terga in some species of Balanus, &c., are specially modified into sharp points or beaks (Pl. 2, fig. 3 b, 3 d), bowed a little inwards, and projecting considerably above the tips of the scuta; this is effected by the medial, uppermost part of the valve being internally thickened and hardened, and then, by the disintegration of the two margins and the external surface, the internal modified portion becomes exposed. The whole valve, also, at least in such cases as in Balanus psittacus, appears to be forced slowly upwards in the articular furrow of the scutum. I am assured, by a competent observer, that the beaks of the terga in B. porcatus can give an object placed within the orifice of the shell a sharp tap.
[22] In comparing the Tergum of one of the Balanidæ with that of a typical member of the Lepadidæ, for instance, that of Balanus with that of Pollicipes, apex corresponds with apex: the extremity of the spur in Balanus corresponds with the basal point of the whole valve in Pollicipes: the scutal margin, (which in Balanus homologically extends down to the extremity of the spur), corresponds with the scutal margin of Pollicipes: the carinal margin in Balanus corresponds with the upper carinal margin in Pollicipes: the basal margin of Balanus on the carinal side of the spur, corresponds with the lower carinal margin in Pollicipes: lastly, (and this is the chief difference), in Balanus there is no appreciable occludent margin, the apex of the valve being brought close to the upper angle of the scutal margin; in Chthamalus, however, there is yet left some remnant of an occludent margin,—which margin in Pollicipes is conspicuous.
The scutum and tergum, with the few exceptions above stated, are articulated together at a large or open angle. The articulation (see Pl. 11, fig. 5 b, c, d, and fig. 6 b, c) is effected by the margin of the tergum being a little inflected, and lodged in a furrow in the margin of the scutum. This[Pg 53] furrow in the scutum has its further border generally prominent and often reflexed or curved over; I have called it the articular ridge; it, also, is lodged in a furrow in the upper part of the tergum, which again is bordered by a ridge, viz., the articular tergal ridge. So that in both scutum and tergum there is an articular furrow, bordered in each case, on one side by the margin of the valve, and on the other side by the so-called articular ridge. In Chelonobia (Pl. 14, fig. 1 b) the articular ridge of the scutum is horny. When, as often happens, the scuta and terga have been much worn, the manner of their articulation (Pl. 18, fig. 1 a) is pretty well shown even from the outside; in this case their external appearance is very different from what it is in those individuals (fig. 1 c) of the same species, which have not suffered disintegration. This articulation of the scuta and terga is prefigured amongst the Lepadidæ, in Pollicipes mitella, and in Lithotrya.
The scuta are brought together by a short, strong, straight, adductor muscle (Pl. 25, fig. 1, a); its attachment leaves (with very few exceptions, as in Tubicinella) a rounded impression, or even pit, on the under side of the valve in its upper part. This pit is frequently bounded, on its lower side, by a sharp ridge, which, though not in actual connexion with the adductor muscle, I have, for convenience sake, called the adductor ridge; it serves apparently to give support to the animal’s body; in some few cases (as in B. psittacus, Pl. 2, fig. 3 c) it is confluent at its upper end with the articular ridge, and converts the whole basi-tergal corner of the valve into a deep cavity. In some of the species of Pyrgoma (Pl. 12, fig. 5 c, 7 b), and in some varieties of Creusia, this adductor ridge is enormously developed, so as to depend far beneath the true basal margin, or that to which the opercular membrane is attached. At the basi-tergal corner of the valve, there is generally a small pit or impression, and sometimes distinct crests, for the attachment of the lateral depressor muscle. At the rostral end there is, also, a small cavity formed by the overfolding of the occludent margin (rarely furnished with crests) for the attachment of the rostral depressor muscle. In the Terga, at the basi-carinal corner, there are usually crests, though[Pg 54] sometimes feebly developed, for the attachment of the tergal depressor muscle. But in Chelonobia, Coronula, Tubicinella, Platylepas, and in some other cases, there are no crests. The crests, when well developed, are furnished with rectangular sub-crests or denticuli on both sides; in fact they resemble, and are probably homologous with, the denticulated ribs or septa in the parietes, radii, and basis. Altogether the scuta and terga are attached, as far as muscles are concerned, to the shell and sack, by three longitudinal pairs.
The opercular valves are added to along their basal margins alone;[23] the animal’s body, together with the several muscles, becoming attached at each period of growth lower and lower down to the valves; this no doubt is effected by the absorption of the upper surfaces of the muscles, and the formation of new fasciæ on their lower surfaces. The opercular membrane, which, though thin and flexible, forms part of the general outer surface of the animal as much as does any portion of the rigid shell, with which indeed it is strictly homologous, is periodically moulted, together with the integuments of the whole included animal. The new opercular membrane is of course each time formed a little larger than the old one. In Coronula and Tubicinella, however, several successive opercular membranes are preserved one over the other, and the outside membrane gradually disintegrates; in these cases the undermost opercular membrane is formed wrinkled and considerably too large, so as to allow of being stretched, before it is finally cast off. In Tubicinella, the opercular membrane runs down, adhering to the inner surface of the shell, to nearly the basis, and hence during the diametric growth of the shell, it is longitudinally split, and is repaired by slips of new membrane, which resemble the radii in form and in direction of the lines of growth.
[23] In some species of Pyrgoma, the ledge (limbus occludens) which is added along the occludent margin of both scuta and terga, and in some species of Balanus a narrow rim, or slight protuberance which is added along the carinal margin of the terga, offer unimportant exceptions to the rule, that the opercular valves grow only at their basal margins.
The basis is added to only round the circumference, and[Pg 55] hence increases in diameter, and, when concave, in depth. The compartments grow at their basal margins, where they are in contact with the basis; hence the shell is added to in height, and, owing to the outward inclination of the compartments, also, in basal diameter; but the compartments likewise, in most cases, grow along both lateral margins, that is, on the edges of the radii and alæ; and hence the upper part of the shell, also, increases in diameter. The orifice of the shell, moreover, thus becomes enlarged. In some cases the shell is destitute of radii, only sutures being present, that is, the compartments do not grow laterally; and sometimes, as in the whole genus Pyrgoma, there are not even sutures, the compartments having been fused together: in both these cases, the shell can increase in diameter only at the base; and the orifice, it might have been thought, would necessarily have remained, to the destruction of the animal, of the same minute size, as when first formed after the metamorphosis: this certainly would have been the case had not the upper ends of the compartments, surrounding and forming the orifice, been nicely adapted always to yield, in a certain limited degree, to the disintegrating influences to which every shell is exposed, but which most Cirripedes can resist; and the disintegration of the narrow end of a conical tube, of course increases the diameter of its orifice. In Tubicinella, in which the shell is furnished with narrow radii, and does increase in diameter from top to bottom, the increase is not sufficient in proportion to the continued elongation of the shell; to compensate for this, the orifice is enlarged at short intervals by the breakage of the upper end of the shell, for which purpose (as explained under the genus) it is evidently constructed. Hence we see that, in certain Cirripedes, decay or disintegration, and breakage, are necessary elements in their growth! It is a remarkable fact, which I cannot explain, that in some species in which the orifice of the shell is usually increased by disintegration, if individuals are so situated that they are not exposed to sufficiently energetic disintegrating influences, as may be inferred from the well-preserved condition of the whole surface of the shell, then the radii become developed, and the orifice is increased in size by the diametric growth of[Pg 56] the upper part of the shell: I have seen instances of this in Tetraclita porosa, and purpurascens, and in Balanus perforatus: it appeared, but of course erroneously, as if the lateral growth of the compartments had been subjected to the will of the animal.
Considering the strength of the shell of sessile Cirripedes, the separation of their compartments one from another and from the basis, during growth, has justly been thought a surprising circumstance. In most Chthamalinæ and in some species of Balanus, however, if the shell be boiled in caustic potash, the compartments fall apart with a touch; this shows that their union is due to animal and probably to organised matter, and the growth of such matter between the opposed edges of the compartments, and their consequent gradual separation, offers no particular difficulty. But in many Balani, boiling in potash for hours does not seem even to weaken, in the least degree, the sutures, which are wonderfully strong—the shell often breaking rather than yield on these lines; if, however, the shell be dissolved in acid, the animalised tissue which is left easily separates on the lines of suture, and if this tissue be boiled in potash, the remnants of the compartments fall quite separate. These facts seem to me to show, that the compartments in such cases are joined along the lines of suture by tissue, which must be in a calcified state, but which, nevertheless, continues to grow by intersusception; in other words, I believe that the tips of the complicated ridges and points interlocking on the lines of suture, are not separated from each other by films of corium or simple animal matter, but are actually united by corium in a calcified, yet still growing condition.
In ordinary Crustaceans, the growth is periodical and sudden; a new and larger carapace, for instance, is formed under the old one, and after the exuviation of the latter, the new one soon hardens, and does not subsequently increase in size; so it is in the case of Cirripedes, with the membranes of the body, and even with certain parts, as the opercular membrane, of the external covering. But a Cirripede cannot, like a crab, crawl into some crevice and remain protected till its shell becomes hardened; hence, probably, it is that[Pg 57] the shell is never[24] wholly moulted. Even if the margins of the opposed compartments and of the basis were to grow rapidly, the shell would necessarily be much weakened on the lines of suture, and unable to withstand the heavy breakers, to which so many species of sessile Cirripedes are exposed. On the other hand, although the margins are thus compelled to grow slowly, they do not grow continuously, as may be seen in the zones of increment on all the valves, corresponding, I believe, with the periods of exuviation of the membranes of the body. A layer of shell, often very thin, seems to be generally deposited over the whole internal surface of the several valves, at the same time that the marginal zones are added; so that the only essential difference in the growth of the external covering, in Cirripedes as compared with ordinary Crustaceans, is that the old shell is not cast off, but adheres to the outside of the new shell, and that the margins are added to (in certain definite directions) slowly yet not continuously, instead of the whole being formed at a single period.
[24] In the genus Alcippe, and in Cryptophialus, the whole of the external membranes are moulted, excepting the surface of attachment; but then these Cirripedes live in cavities which they form for themselves, and are thus protected. In Lithotrya the membrane of the peduncle, with its little valves or scales, is moulted, but here, again, this very part is protected by the tubular cavity, which the animal forms and inhabits. Neither of these three genera belong to the Balanidæ, or sessile Cirripedes, which we are now more especially describing.
If, now, a section of one of the shelly zones of growth be carefully examined, it can in some cases be distinctly seen to be formed of successive, excessively fine laminæ; but the animalised tissue (which differs much in amount in different Cirripedes) left after the shell has been dissolved in acid, exhibits, in most cases, neither laminæ nor any other structure whatever. The shell seems to be the actual pulpy corium, or true skin, in a calcified condition, but generally with its cellular structure modified and much reduced: I have taken a bit of recently-formed shell of Tetraclita and of Coronula, with the corium still adherent to its under surface, and after dissolution in acid, I could not distinguish the part, which had just before existed as shell, from the corium itself. In the case of Coronula, immediately[Pg 58] prior to the period of moulting and growth, I found the unaltered corium so charged, as to effervesce, with carbonate of lime, either in a state of dissolution, or in granules too minute to be visible under the highest powers.
The sutures between the several compartments and the basis are covered by thin membrane, which is continually splitting during the growth of the opposed edges of the underlying shell; but previously to each splitting, a new slip of membrane is, I believe, already formed under the old one; so that the corium is not even momentarily exposed. Owing to this manner of growth, the slips of membrane consist of successive rims united together; in most cases, these soon become abraded from the older parts of the shell, but are sometimes preserved. The last-formed slip of membrane over a suture is homologous with the opercular membrane; and both are strictly analogous with the ring of flexible membrane, forming the joint of the leg of a crab. In the latter case, the flexible membrane and hardened crust are both moulted together: in the opercular membrane, there is a double line of splitting, one close round the opercular valves, and the other at the basal edge of the sheath, and the intermediate portion is moulted, but with a zone of membrane left adherent to the non-moulted valves and sheath: lastly, in the slips of membrane covering the sutures, there is only a single line of splitting, and no portion, I believe, is moulted; the rims of membrane on each side remaining adherent on the compartments and basis, until worn away.
The opercular membrane, when closely examined, exhibits no structure, except that it can sometimes be plainly seen to be composed of successive, numerous, excessively thin laminæ. Occasionally, however, it presents the false appearance of being permeated by parallel and anastomosing vessels: this appearance is clue to one or more of the component laminæ having been wrinkled before a succeeding lamina was thrown down and attached to its under side. If a small piece of an opercular valve of Tubicinella, with the opercular membrane adhering to it, and with the corium under both, be dissolved in acid, it may be clearly made out that the corium under the valve has gone on[Pg 59] being converted into shell, whereas under the opercular membrane it has been converted and condensed into fine constituent laminæ of chitine. Inasmuch as the successive layers of shell, during each period of growth, go on encroaching on those of the membrane, the line of junction between the shell and chitine becomes oblique or bevelled. The membrane on this bevelled line of junction assumes a slightly different aspect to what it has elsewhere; it becomes yellowish or brown, thicker and very much tougher. In many genera it is also furnished with a row of small bristles. At the period of exuviation the opercular membrane separates just outside this modified portion, leaving the latter adherent, as a rim or slip, on the valves. If, however, the opercular membrane be rudely torn off before its proper period of exuviation, it carries with it the as yet continuous, but already modified, slip. A slightly indented line may sometimes be traced before the period of exuviation, showing where the separation will take place: what produces this line I know not. The coloured, thickened, and modified slips of opercular membrane, which are thus retained adhering to the valves, and which together form an investing membrane, have been considered by most authors as the epidermis; but they have no more right to be thus called than has any other part of the opercular membrane. Exactly similar slips of membrane are left investing the sheath. So, again, the membrane which, when well preserved, invests the walls of the shell, is made up, as already stated, of successively adherent slips, which originally covered the lines of suture.[25]
[25] In the case of Coronula there is a peculiarity, described in the last section of this Introduction, (under the head of Cementing Apparatus), namely, that the two or three last-formed, exterior zones of the Basal membrane continue for a period to increase in width; being, as I believe, dragged one from over the other, with fresh laminæ of membrane continually thrown down. In this same genus, and in Tubicinella, the walls of the shell are invested by membrane, which is doubled inwards under their basal edges; and as the latter grow, the investing parietal membrane splits and separates from the basal membrane, and is pulled outwards and downwards. This inflected, often broad border of membrane, seems to me more strictly comparable with the opercular membrane, than with those narrow, thickened rims of yellowish membrane which in other Cirripedes cover the suture between the basal edges of the walls and the basis.
The little bristles above alluded to, which arise from the[Pg 60] slips of membrane left adherent on the opercular valves, sheath, and walls, stand in rows; a row corresponding to each period of exuviation of the opercular membrane. The bristles are generally largest on the opercular valves and sheath; in Balanus tintinnabulum, they are from 1 to 2/1000ths of an inch in length, but they are longer in some other species. I may here mention, as showing the connexion of these bristles with the opercular membrane, that similar bristles occur in B. perforatus, scattered over the surface of that membrane, and are necessarily moulted with it. In the imbedded genera Coronula and Tubicinella, none of these bristles exist. When a portion of valve or shell, furnished with bristles, is dissolved in acid, tough, sinuous, and apparently hollow, threads are seen to run from their bulb-like bases, into and up the corresponding layer, which, before dissolution, existed as shell; and they terminate internally in very fine points, which I believe are united to the underlying corium. These threads, or tubuli,[26] as I have called them in my volume on the Lepadidæ, are, in Tetraclita porosa, about 1/5000ths of an inch in diameter, but only half that size in B. tintinnabulum. On parts of the shell where there are no bristles, similar tubuli penetrate the shelly layers, and come to the surface. The tubuli running to the lowest and last-formed row of bristles, just after a period of exuviation, are so delicate as hardly, or not at all, to be distinguished; in the row above, they are plain and longer, and for the next two or three upper rows they are, in some cases, as in Tetraclita porosa, longer and longer, having been added to during each successive thickening of the valve. These tubuli consist of chitine, and no doubt first existed as threads of corium; they are so tough that they must serve to strengthen the successive layers of shell, but I imagine their chief function is to keep up the vitality of the newly-formed layers of shell. May we not, also, venture to suppose that by their means, some degree of sensibility is given to the bristles?[Pg 61] I need only further remark, that in some species of Balanus and of Chthamalus, the under side of the shell is penetrated by irregular pores, large enough to be visible to the naked eye, into which threads of corium penetrate; but these can hardly be said to appertain to the microscopical structure; and are more nearly related to those pores and furrows, formed by the greater or less development of the longitudinal septa, and in which the threads of corium deposit, or rather become changed into, transverse septa, or solid shelly matter, as previously described.
[26] I regret that I have used this term “tubuli”; for the threads thus designated, I believe, are not the same with the tubuli of Dr. Carpenter, which are not left after dissolution in acid. I have seen tubuli, as called by me, in the shell from the leg of a crab, after having been placed in acid.
In the pupa, the thorax, as we shall hereafter more fully see, is continuous with, and opens into the large anterior end or front part of the head; but during the metamorphosis (Pl. 30, fig. 2), the thorax of the Cirripede becomes, owing to the almost transverse position occupied by the young animal within the pupa, to a great extent internally separated from the anterior end,—which anterior end forms, as we know, either the peduncle or the basis. Hence it comes to pass that the body or Thorax (Pl. 25, fig. 1) is lodged within a sack (f) within the shell. The chitine membrane lining this sack is excessively thin and transparent, but less so in Xenobalanus and Tubicinella; it is obviously continuous with that investing the body of the animal; it is also essentially continuous with the opercular valves and membrane, and consequently with the whole shell. It is periodically moulted. It is lined by corium, as is likewise the surrounding shell; hence the corium is double round the sack, as indeed might have been expected from the shell and opercular valves (at least their upper parts) being formed by the prolongation, as is obvious in the pupa, of the posterior edges of the carapace. Between the two folds of corium, which are united together by transverse ligamentous fibres, branching out at both extremities, like the roots and branches of a tree, we have the longitudinal muscles, which go to the opercular valves; and likewise a layer-like mass of branching ovarian tubes (Pl. 25, fig. 1, g): the ovarian tubes, however, are often confined to the base[Pg 62] of the sack. In Xenobalanus, the two folds of corium are united by longitudinal membranous septa, making a series of quite peculiar, square tubes.
The above-mentioned muscles are attached at their upper ends to the opercular valves, and at their lower ends to the basis. There are, in fact, three pairs, but the pair attached to the basi-carinal angles of the two terga (Pl. 25, fig. 1, i), are almost invariably confluent, forming one great bundle; the second pair is attached to the lateral or basi-tergal corners of the two scuta, and are hidden in the figure; the third pair (h) is attached also to the scuta, to their rostral angles. These muscles can only act as depressores; they are often extremely powerful; they belong to the voluntary class, for they are transversely striped. By their action, the opercular valves are capable of varied slight movements, within the limit allowed by the width of the flexible opercular membrane. By the action of the lateral scutal depressores, the orifice leading into the sack is opened, the movement being generally aided by the protrusion of the cirri. By the sudden contraction of the rostral scutal depressores, the blows which are sometimes given by the beaked terga at the opposite end of the operculum, are probably effected. By the contraction of all three pairs of muscles, the opercular valves are held down with quite surprising force. The valves can be raised only by the action of the animal’s body against the basis.
In Coronula these muscles are more spread out, and do not extend down to the basis; their lower portions, as is likewise the case in Tubicinella, do not exhibit transverse striæ, and hence tend to pass either into the involuntary class, or into ligament. This condition of the muscles, in the above two genera, accords with the little-developed state of their opercular valves. In Xenobalanus, there is no longer any evidence of the muscles being collected into five or six bundles, for they are thinly and almost uniformly spread out, and show in no part transverse striæ. I may add that in much elongated specimens of Balanus balanoides, these muscles become in their lower part ligamentous, and destitute of striæ.
In the Balaninæ, a pair of Branchiæ is always present: they lie on each side, in a somewhat curved position, in the angle between the sides of the shell and the basis. In Pl. 25, fig. 1, they are exactly covered, on the further side, by the body of the animal. They are attached near each other at the carinal end of the sack in a vertical line, and likewise on each side in a transverse line, extending from close beneath the spur of the tergum towards the point of attachment of the body to the scutum. In Balanus, as in the figure (Pl. 25, fig. 3) of B. tintinnabulum, each branchia consists of a medial fold of skin, a little curved conformably with the sack, and slightly tapering towards its rostral and free extremity; but this fold is almost hidden by the vertical sub-folds or membranous ridges, themselves plicated and sub-plicated, which project on both sides: these vertical folds are free at their tips: at their lower attached ends, they are thickest. On the side nearest the wall of the shell, the whole branchia has a bilobed appearance, owing to a very deep indentation caused by the projection of the scutal lateral depressor muscle; the sub-folds on this side are also more plicated. The branchia essentially is an inward plicated fold of the membranes of the sack; for its outer, very thin tunic is continuous with and moulted with that lining the sack; and within it we have two layers of delicate, pulpy, transparent corium, united together (as is best seen in Coronula) by ligamentous fibres, branched at their two ends, all exactly as in the corium surrounding the sack. There are here no distinct vessels, any more than in other parts of the body, but a fluid could easily circulate in the interspaces of the corium. From the large size of this organ, and its simplicity of internal structure, being adapted exclusively to expose a great surface of skin to the water, I do not doubt that it has been correctly considered as a respiratory organ. By the voluntary movements of the opercular valves (i. e. part of the carapace) the water is constantly being pumped in and out of the sack; the movement, indeed, may be[Pg 64] almost compared to the heaving of a man’s chest. Moreover, the branchiæ on each side are attached so closely to the spur of the tergum, that each time the latter is moved, the whole branchia must, I think, be agitated, and the folds opened, as by the action of a lever.
In our two commonest, tidal, sessile Cirripedes, viz. Balanus balanoides and Chthamalus stellatus, I have observed that, when left uncovered by water, they kept the orifice of their operculums a little open, with a bubble of air within their sacks, so that the orifice was in fact closed by a thin septum of water, with air beneath; when disturbed, they closed their operculums with force, and expelled the bubble of air with a clicking noise, which has been noticed by Dr. Coldstream,[27] and has been thought to be made by the movement of the operculum itself. Bal. crenatus, a deep-water species, when out of water, keeps its operculum closed.
[27] ‘Encyclopædia of Anatomy and Physiology;’ article Cirrhopoda.
In Coronula, Platylepas, Tubicinella, and Xenobalanus, each branchia[28] consists of two unequal folds, both plicated on both sides: in the two latter genera, they extend far down the deep and elongated sacks, and hence the area of surface altogether gained is extremely great. In most of the species of Chthamalus, the branchia consist of a small fillet barely plicated: in the allied Chamæsipho columna, they are rudimentary, forming a smooth little pouch only 1/100th of an inch in length: in Chthamalus scabrosus they are quite aborted, being perhaps represented by a slight hairy ridge; but in Chthamalus dentatus, and therefore within the limits of the same genus, the branchiæ (and this seems to me a singular fact) are large, each being composed of two plicated folds, as in Coronula. Tapering filaments situated near the bases of the cirri, such as those occurring in several species of the Lepadidæ, are not found in any sessile Cirripede; but I have observed nearly similar filaments, projecting upwards and inwards at the base of the sack, in several species of Balanus and in Coronula; those which I[Pg 65] examined were simply occupied by delicate corium, and no doubt must aid in exposing a greater surface of corium to the circumambient water.
[28] Burmeister has given a good figure (Tab. 2, fig. 10) of the branchiæ of Coronula, (but the two folds are shaded too unequally), in his ‘Beiträge zur Naturgeschichte der Rankenfüsser.’
In my former volume on the Lepadidæ, I have described the ovigerous fræna occurring on the two sides of the sack, to which the ovigerous lamellæ are attached by a peculiar glandular secretion: in the Balanidæ there are no ovigerous fræna, but the branchiæ just described are identical with the fræna in essential structure and in position; differing only in being placed a little nearer to the carinal end of the sack, and in being generally (but not always) larger and more plicated: seeing this, and that in Alcippe lampas, and in some species of Pollicipes,—the genus which comes nearest to the Balanidæ,—the ovigerous fræna are large and are destitute of glands, and have therefore lost their normal function of supporting the ovigerous lamellæ, I can hardly doubt that the branchiæ in the Balanidæ are the ovigerous fræna of the Lepadidæ in a modified condition; a transformation of function not greater than that of the swimming bladder of a fish into the lungs of the higher Vertebrata.[29]
Parts of the body included within the shell or carapace.—These parts (Pl. 25, fig. 1) consist of the prominent mouth, and of the thorax (c′), with its largely developed portion, called the prosoma (c), and with its appendages. The abdomen is quite rudimentary, being represented merely by a small portion of membrane surrounding the anus, and sometimes inserted like a wedge between the inwardly inflected posterior thoracic segments; in only two genera (Catophragmus and Pachylasma), its nature is rendered somewhat plainer by supporting caudal appendages. The probosciformed penis lies folded under the thorax; and I believe (from what is seen in the anomalous genus Proteolepas), that it normally arises from the ventral[Pg 66] surface of the terminal point of the rudimentary abdomen.[30] The thorax is laterally compressed, the ventral surface being very narrow, with the bases of the cirri placed closely together. It consists, in appearance, of two very different portions; one a soft, more or less rounded bag, which I have called the prosoma; and the other, which supports the five posterior pairs of cirri, is narrower, invested with stiffer membrane, and is more or less distinctly composed of five segments. These segments (Pl. 26, fig. 8) on their dorsal and dorso-lateral surfaces, are generally driven like wedges one into the other, with their points directed anteriorly: on the ventral surface the articulations are transverse. The prolongation (e) of the thin membrane (a) surrounding the anus (b), that is, the rudiment of the abdomen, which sometimes carries caudal appendages, almost divides (in appearance, whether really I know not) the hindermost thoracic segment along the medio-dorsal line, into two parts. I have given the above drawing of these segments, but with the dorsal surface much flattened, in Coronula diadema; in most species of Balanus, however, the wedges formed by one segment being driven into another, are much sharper; on the other hand, in Xenobalanus they are nearly straight and transverse. The three posterior segments are always the most distinct; the two next segments are also distinct laterally, but along the dorsal surface they become, excepting in Xenobalanus and some few other cases, completely confluent. The greater distinctness of the posterior segments is conformable to what takes place in the higher Crustacea. The articulations between the segments are folded inwards, and are formed of thin membrane, which in some cases, as in Coronula diadema, forms a marked contrast with the much thicker, stiffer, and yellowish membrane of the segments themselves; in Balanus tintinnabulum, how[Pg 67]ever, the whole membrane of the five thoracic segments is very thin, excepting small wedge-shaped portions along the medio-dorsal line. The infolded articulations between the segments supporting the three anterior pairs of cirri (at least in the Balaninæ), are much wider than those between the three posterior segments; the former segments, with their cirri, being consequently capable of being moved further apart from each other. Could there have been any doubt as to the distinctness and reality of the five thoracic segments, it would have been set aside by the arrangement of the muscles attached to them, as will presently be described. I need only add, that in many genera there are shield-like swellings at the exterior bases of the pedicels of the posterior cirri, which I for some time thought were the epimeral elements of the thoracic segments; but I now believe them to be parts of the pedicels of the cirri. The basi-exterior margin, moreover, of the pedicel of the third pair of cirri, in many species of the Balaninæ (Pl. 25, fig. 1), is produced as a plate, thickly fringed with fine hairs, half across the dorsal surface of the thorax; serving, apparently, as a brush to clean the sack, or to prevent the ingress of any intruding substance.
[30] Von Siebold and Stannius, in their ‘Anatomie Comparée,’ tom. i, p. 473, and p. 440, (foot-note), consider the articulated probosciformed penis as an elongated abdomen; a view which, at the commencement of my examination, I was tempted to admit; but the position of the caudal appendages on the dorsal basis of the penis, suffice, I think, to show that this view is not correct; for these caudal appendages evidently correspond with those borne on the very extremity of the abdomen in the pupa. Nor, indeed, does the position of the anus accord well with such a view.
The soft, rounded, bag-like portion of the body, which I have called the prosoma, is usually separated by a notch from the five posterior thoracic segments; at its upper end it may be said to carry the mouth and first pair of cirri. The prosoma includes the main part of the stomach and the broad ends of the vesiculæ seminales. It is always clothed by very thin membrane, which in Chthamalus dentatus, is hairy. In Tubicinella and Xenobalanus, the prosoma is much elongated, being produced far down the deep sack. That the prosoma is mainly formed by a great development of that segment (homologically the second thoracic segment) which carries the first pair of cirri, is certain, and I should not have hesitated to have said that it was exclusively so formed, had not the first thoracic segment in the anomalous genus Cryptophialus been developed as a distinct and free segment, not attached to the carapace; showing that possibly in other Cirripedes, the dorsal half of this first thoracic segment may be concerned in the formation of the free prosoma.
Attachment of the Body to the Shell.—The prosoma which carries the posterior thoracic segments, and in appearance the mouth, is the only part of the body which is attached to the general covering (Pl. 25, fig. 1), namely, to the opercular valves. Except through the continuity of the lining membranes of the sack, the body lies free within the walls of the shell. The area of attachment (shown by a sinuous broken line round a and b) extends from about the middle of the two scuta down to their basal margins. As these valves lie obliquely across the orifice of the shell, the animal’s body comes to be suspended almost in the middle of the sack. The two scuta, as we have seen, have the power of opening and shutting a little; and are brought together by the adductor scutorum muscle (a), which is generally very powerful. The body is attached to these valves, round and beneath the adductor, so as to hide it until one of the valves be removed. The attachment is chiefly effected by three pairs of widely expanded, superficial muscles, two pairs of which are spread over the flanks of the prosoma, and the third pair over its rounded (properly dorsal) surface, which lies close to the rostral compartment (A, fig. 1) of the shell. I should have stated, that my chief examination of the attachment of the body to the scutal valves, has been made on Coronula balænaris, and less closely on Balanus tintinnabulum. Within these three pairs of superficial muscles, there are (besides the adductor) no less than five other pairs; of these one long pair is attached at one end to the basal margin of the labrum (e), and at the other end, to the under side, near to the basal margin of the scuta: two other, shorter, parallel pairs of muscles are attached at one end to the interspace of membrane between the basal edge of the labrum and the adductor scutorum muscle, and at the other end, to the under side of the scuta, above the attachment of the first pair: the fourth and shortest pair curls close under the adductor, and is there attached at both ends beneath it. The action of these four pairs of muscles must be to draw back, from the orifice of the shell, the mouth, and that inter[Pg 69]space of body between the basal margin of the labrum and the adductor muscle. This movement I saw in living specimens. The last and fifth pair of muscles is small, but of considerable length; it is a diverging pair, attached at the converging end, above and exteriorly to the adductor muscle; and at the diverging end, low down on the under side of the scuta; I am very doubtful regarding the function of this pair. Altogether we have seen that round and within the fleshy pedicel, by which the body is attached to the scuta, there are no less than eight pairs of muscles. The central space between these muscles is hollow, and here many lacunal channels seem to converge. These muscles receive nerves from the supra-œsophageal ganglions. The interspace above alluded to, between the basal edge of the labrum and the adductor scutorum muscle, occupies a very different position according as the animal’s body is protruded as far as it can be, or is retracted. It is homologically part of the third cephalic segment; and consequently the mouth ought to have stood posteriorly (i. e. above, in the position figured in Pl. 25, fig. 1) to this interspace; yet, in fact, when the animal is retracted within its shell, the mouth usually lies almost directly beneath this interspace and the adductor scutorum muscle.
Besides these muscles of attachment, the prosoma is furnished with several other muscles. There are superficial muscles running up towards the basal margin of the sides of the mouth; and other deeper muscles, to which, I presume, the movements of the mouth, as a whole, are due. The muscles moving the gnathites do not, as far as I could make out, extend beneath the basal edge of the mouth. There are, also, powerful muscles giving movement to the basal segments of the pedicel of the first pair of cirri. Again, there are superficial muscles running to the next succeeding thoracic segment; the anterior ends of which are separated by a clear interspace from the lower ends of the above-described superficial muscles, by which the prosoma is attached to the scuta. On each flank, moreover, but more deeply imbedded, are the long flexor and extensor muscles, presently to be described, running to the five posterior thoracic segments. The last muscle which I need here[Pg 70] mention, is a deep-seated diverging pair, attached near the upper end of the stomach, on its ventral surface, and diverging from this point to the sides of the prosoma high up beneath the mouth. The probable action of this pair, as well as of the three superficial pairs of muscles by which the body is attached to the scuta, is to draw up the whole prosoma towards or from the orifice; and likewise to contract it firmly, so as to serve as a fulcrum for the movements of the five posterior thoracic segments, together with the cirri, which they carry.
The muscles of these five thoracic segments are numerous and powerful; they are also complicated, chiefly owing to the segments on their dorsal and dorso-lateral surfaces being driven, like wedges, one into the other. As far as I could make out, there are on each side three, superficial, dorso-lateral and lateral muscles (generally, if not always, destitute of striæ), which do not cross the articulations, but extend merely from articulation to articulation; and of which the function can be only to contract each separate segment, and consequently to open out the intermediate infolded articulations; the effect of this would be to separate slightly the cirri from each other,—more especially those borne on the two or three anterior segments, between which the infolded articulations are deeper or broader. There are other more deeply imbedded, powerful, long, dorso-lateral extensor, and ventri-lateral flexor muscles, attached at one end within the flanks of the prosoma, and at the other end to the successive segments of the thorax. The action of the former is to straighten and stretch out the thorax; of the latter, or ventri-lateral muscles, to retract it. In tracing these muscles, a fascia could be seen to become attached to a segment, and then this same fascia would run on to the next succeeding segment: the effect of this must be, that each segment can be retracted and protracted either from the prosoma as a fulcrum, or from the antecedent segment as a fulcrum: we have, also, seen that each segment can, by the agency of the superficial, non-striated muscles, contract itself. Hence these thoracic segments are capable of diverse movements, as was very evident when the shell of a living specimen was opened. By one movement in[Pg 71] common, the whole five posterior segments could be drawn back, so as to become even partly imbedded in the prosoma: lateral, twisting or wriggling movements were also quite distinct: the three posterior segments seemed to be capable of less independent movements than the anterior segments; and I observed that the more powerful flexor and extensor muscles did not run into these three posterior segments. The cirri, of course, partake of the movements of the thorax; and in watching, in an uninjured specimen, the alternate, protruding, gracefully sweeping and retracting movements of the posterior pairs of cirri, it was evident that the thorax was the chief agent in their movement. Besides the muscles now mentioned, there are some immediately to be noticed, which extend from within the thoracic segments to within the pedicels of the cirri.
Although the cirri have not been described, it will be most convenient here to treat shortly of their muscles. Each cirrus consists of a pedicel, having a long basal and a short upper segment, supporting two multiarticulate rami. The lower segment of the pedicel can be drawn forward by an adductor muscle, attached low down within this segment, and crossing at right angles (at least in the case of the anterior cirri) the corresponding muscle of the opposed cirrus, on the central, ventral surface of the thorax. This segment can also be drawn back by a muscle springing from the dorso-lateral surface of the thorax, and running only a little way within the segment: I am far from sure that the lower segment does not possess other muscles. The short upper segment of the pedicel can be moved backwards and forwards, as I saw in living specimens, independently of the lower segment; this movement being best seen in the anterior cirri, which are much more often moved independently of each other than are the posterior cirri. The rami are capable, I believe, of being moved backwards and forwards as a whole, by the movement of the few lower segments, which are generally more or less confluent. They can, also, be curled up and uncurled by the combined movement of each separate segment. The uncurling seems to separate the two rami a little laterally. Each ramus, at[Pg 72] least in the two or three anterior pairs, can be moved to a certain extent, independently of the other ramus of the same cirrus; and the few terminal segments, either of both rami or of one ramus, are often a little moved and curled (and this is especially the case with the long anterior ramus of the first pair), without the lower segments or the pedicel being moved.
The flexor and extensor muscles, which, as I believe, move the upper segment of the pedicel (a and b, Pl. 29, fig. 1), are attached at their upper ends to its basal margin, and are thus enabled to draw it a little way down within the lower segment, and so move it. The short flexor muscle (c), which is attached at its lower end within the upper segment of the pedicel, and the longer extensor (d), also, attached within this same lower segment, serve, I believe, to move the lower, partially confluent segments of each ramus as a whole. In the case of these muscles, and of those last mentioned, I am surprised that the extensors (b) and (d) are not attached nearer to the exterior and dorsal surface. Other muscles (e, f) attached at their lower ends within the upper segment of the pedicel, run up each of the two rami to their tips, with some of the fasciæ terminating within each segment: of these muscles, the outer one (f, f) appears to be the extensor, and the inner one (e, e) the flexor. But besides these, there are other short flexor muscles (g, g) which run on the anterior face,[31] from segment to segment, serving to pull the front edge of one segment within the edge of the next lower segment. These muscles differ much in plainness in the several genera: they are very distinct in Coronula. In some specimens of this genus, a few of the articulations between the basal segments of the rami having been obliterated, the short muscles (g, g) running from articulation to articulation were absent, and their presence and nature in the upper segments thus rendered the plainer. The muscular system in the several pairs of cirri seems to be the same, with the exception of[Pg 73] the first pair, in which the muscle answering, as I suppose, to (a), namely, the flexor of the upper segment of the pedicel, is much spread out at its lower end, and is there attached to the exterior surface of the lower segment.
[31] For a considerable time I thought that there were muscles going to the spines, especially to those which arise from the upper dorsal edge of each segment; but I have since ascertained that these are the cases within which new spines, with their lower ends doubled like the fingers of a glove hastily pulled off, are in process of formation.
The backward and forward movements of the segments, both in the rami and in the pedicels of the cirri, are apparently effected, as already noticed, by the outer or inner (as the movement may be) basal edge of one segment being drawn a little way down within the next succeeding lower segment. If, at the same time, both the inner and outer margins of all the segments were drawn one within the other, the whole limb would necessarily be shortened; and I distinctly saw a shortening action, with very slight movement in any other direction, in the first and second pairs of cirri; and I think it almost certain that this movement might be performed by the other cirri. If I correctly understand a statement of Milne Edwards,[32] this is an important fact, as he asserts that only the higher Crustaceans possess the power of shortening their limbs.
[32] ‘Annales des Sciences Naturelles,’ tom. xviii, 1852, p. 121.
When a Cirripede is alive, the action of the cirri is really beautiful: from the position of the thoracic segments, the posterior cirri (three pairs in the Balaninæ and four pairs in the Chthamalinæ) form a sort of semicircle facing the mouth: the anterior cirri stand further apart, and are opposed in pairs to each other, with the first pair pointing beyond the mouth. Together the cirri form a hollow cone, not circular but elongated, with the mouth situated at the lower anterior end. The posterior cirri are protruded, by the movement of the whole thorax, curled up, close along the carinal end of the orifice; as they are protruded, they diverge, both by the movement of their pedicels, and, as I believe, by the separation of the thoracic segments. As the two rami of each separate cirrus are uncurled, they also diverge a little; as do the double rows of spines on the segments in each ramus, by their elasticity. By the movement of the thorax, the cirri are then swept towards the rostrum; and, lastly, they are brought perpendicularly down towards the mouth with a rapid movement, which would be beautifully adapted to catch any object floating or swimming in the water; hence[Pg 74] I have called the action of the cirri, captorial. When the shell of a Balanus is broken open, the second and third pairs of cirri are repeatedly clasped over the mouth with a convulsive movement, in a manner indicating, I think, that their chief function is to seize and carry to the mouth any object entangled by the sweeping movement of the three posterior pairs. The first pair is also well adapted to aid in this seizing action; but I suspect that the long anterior ramus likewise acts as an organ of touch, warning the animal of danger. The mouth being itself moveable as a whole,—the outer maxillæ being capable of a backward and forward sweeping action, and being furnished with orifices apparently olfactory,—the inner maxillæ having more diversified movements,—the toothed mandibles overhanging the œsophagus,—and the œsophagus itself possessing a powerful swallowing movement, are all admirably adapted to secure any prey, when once entangled by the cirri.
The mouth, in the sub-family Chthamalinæ, cannot be distinguished from that of the Lepadidæ, which has been pretty fully described in my former volume. In the Balaninæ, however, the labrum differs considerably in not being swollen; that is, in its outer and inner fold of membrane being close together, and in having a central notch: the palpi are also larger, and the lower teeth on the mandibles, are laterally (Pl. 26, fig. 5) double, as will be more particularly stated under these two sub-families. I have given a drawing (Pl. 26, fig. 1) of the mouth, seen from above, of Balanus perforatus, with the right-hand palpus (d′) and outer maxilla (a′) cut off, in order that the labrum (e), mandibles (c), and inner maxilla (b) might be better shown; the cut-off bases (x, x) of the first cirrus on each side are also shown. In fig. 2 we have the deep supra-œsophageal cavity in Bal. improvisus torn open and laid flat, with the inner surfaces of the labrum (c) and outer maxillæ (a) exhibited, the palpi, mandibles, and inner maxillæ having been removed. Figs. 3 and 4 will presently be referred to; they are parts of the mouth, with the muscles,[Pg 75] &c. removed, of Coronula. The mouth differs extremely little in the different genera and species of the Balanidæ, much less than amongst the Lepadidæ. In the Balaninæ, the crest of the labrum is sometimes hairy, instead of having, as is usual, from two to six teeth on each side of the central notch: in Balanus improvisus (Pl. 26, fig. 2) and eburneus, and in Chelonobia, the crest on each side of the central notch (e′) is furnished with a row of finely graduated teeth. A sub-triangular portion of the inner fold of membrane of the labrum, which overhangs the œsophagus, is always thickened and yellowish; it is also often punctured in patterns (Pl. 26, fig. 2, f), which, I believe, give attachment to little muscles that serve to open the upper end of the œsophagus. Opposite to this thickened, sub-triangular portion of membrane, the thin membrane forming the supra-œsophageal cavity (or the cavity surrounded by the gnathites) is strengthened by a pair of curved ribs (h, fig. 2) of thickened yellowish membrane, running down from the inner bases (a′′) of the bilobed outer maxillæ to the opening of the œsophagus (g): a broad branch from each of these ribs supports the sides of the orifice of the œsophagus; and this branch almost joins on to a slightly thickened rim or bar (f′), which branches off from the upper part of the sub-triangular (f) inner fold of the labrum. This structure, in Bal. improvisus, is represented in Pl. 26, fig. 2, as well as it could be, considering that the deep supra-œsophageal cavity has to be torn open; and then laid flat.
The Palpi (Pl. 26, fig. 6) differ little, except in size, in the different genera, being squarish, more or less elongated, or even approaching to club-shaped: in most of the Balaninæ they are larger even than the mandibles, of which they normally form a part. Their upper margins, especially towards their free extremities, are always thickly clothed with spines; and there is generally a single row, either short (r) or long, of spines of greater length, which arise from a little above, and stand almost in a parallel line to, the basal margin. On the internal surface, there is sometimes a row (t) of very short little spines, which overhang the crest of the labrum. The Mandibles (Pl. 26, fig. 5) have from[Pg 76] three to five teeth; the lower point or angle is generally pectinated. In Coronula and its close allies, there are some small teeth intermediate between the four or five main teeth; and in these genera, though members of the sub-family Balaninæ, the lower teeth exhibit only rudiments of being laterally double.[33] The Maxillæ sometimes have a notch under the upper large pair of spines, and in Octomeris brunnea there is a double notch: in many species of Balanus, the inferior corner stands up like a step (Pl. 26, fig. 7, a): in many other genera and species, the whole edge is straight. In all, or almost all cases, the row of spines on the middle portion is double. The Outer Maxillæ are always bilobed on their inner faces, and are clothed with bristles. On all the gnathites, the bristles are often doubly serrated.
[33] M. Martin St. Ange describes, in his ‘Mémoire sur l’Organisation des Cirripèdes,’ pp. 15 and 32, “une petite langue” in the mouth of Lepas; but I may venture to assert that such does not exist; it is merely the point of union between the outer maxillæ. M. St. Ange, in his comparison of the mouth of Lepas with that of Phyllosoma, compares the mandible of the latter with the palpus of Lepas; the first maxilla of Phyllosoma with the mandible of Lepas; and so on with the other gnathites.
Muscles and functions of the Gnathites, and their confluence.—The outer maxillæ appear at first like a deeply-lobed lower lip, for they reach over almost to the labrum (Pl. 26, fig. 1), and thus partially cover the other organs; they are separately capable of a strong and rapid, to and fro movement, by which no doubt they sweep any prey, entangled by the cirri, towards the other gnathites. Each outer maxilla is furnished with a pair of muscles, apparently a flexor and extensor; there is also a little muscle between the two maxillæ, I presume for the purpose of bringing them together. The outer and inner maxillæ generally stand close together, and in several genera a little way apart, from the mandibles; but there is no trace of any labrum or true lower lip, bounding the mandibles and orifice of the œsophagus. The outer and inner maxillæ and mandibles are not opposed in pairs to each other, but against the thickened inner fold of the labrum; almost in the same manner as the posterior pairs of cirri are not opposed one to the other, but to the mouth.
I have described pretty accurately the muscles of the[Pg 77] mandibles in my former volume, and there given a drawing (Pl. 10, fig. 1) of them. There are four muscles: first, the depressor muscle, which is the largest, and is attached, at its upper end, to ligamentous apodemes under the free toothed portion of the jaw; and at its lower end, spreading considerably out, is attached to a concavity close above the basal margin of the labrum; to understand the action of these muscles, it should be borne in mind that the mandible almost faces the labrum. In some genera, as in Coronula,[34] the swelling near the basal margin of the labrum (Pl. 26, fig. 3, k), caused by the internal concavity for the above muscle, is conspicuous. The depressor muscle is opposed by a small elevator, attached to the mandible close by the depressor; thence it runs upwards, and is united at its upper end to the base of the palpus, at the point where the latter adheres to the labrum: I have ventured to call this muscle the elevator, from being apparently so well fitted for this purpose; but I feel some little doubt, from having observed an apparent slight movement in the palpi of living Balani; and this is the only muscle entering those organs. The free part of the mandible is articulated on a square, thickened piece of membrane, forming part of the side of the mouth (Pl. 26, figs. 3, 4, c1; and Pl. 10, fig. 1, a, b, in my volume on the Lepadidæ); to this square piece of membrane, two short muscles are attached, one above the other, and which ought, in the Plate in my former volume, to have been represented crossing the depressor muscle at nearly right angles; at their further ends they are attached to about the middle of the labrum, where, at least in Coronula (Pl. 26, fig. 3, i), a slight concavity can be detected. The action of these two muscles must be to draw the whole mandible against the labrum; and the depressor muscle might, at the same time, draw the toothed edge downwards, and thus force any prey into the œsophagus.
[34] This is figured by Burmeister in his ‘Beiträge zur Naturgeschichte der Rankenfüsser,’ Tab. 2, fig. 6.
The inner maxillæ are likewise furnished with four muscles, very nearly as figured in my former volume (Pl. 10, fig. 10); namely, two muscles, one inside and the other outside the curious apodeme, which in the Balanidæ (Pl. 26, fig. 7, b′) is[Pg 78] as invariably present as in the Lepadidæ: these two muscles are attached at their lower ends to the outer membrane of the mouth, close to its basal articulation: the outer one of these two muscles would, I presume, act as an elevator, and the inner one as a depressor; the free part of the organ working on the top of the apodeme, like an axe, on a hinge, on the top of a pole. But there is also a larger depressor muscle, in an analogous position with that (i. e. the first-mentioned muscle) of the mandibles; and a fourth muscle, crossing the latter depressor at nearly right angles, and attached (as far as I could make out) on the side of the orifice of the œsophagus, close under the mandibles: the action of this latter muscle would be to draw the whole organ towards the labrum.
I must not conclude my description of the mouth, without drawing particular attention to its peculiar compounded nature. It is prominent, and is capable, as a whole, of movement; it is separated from the body by a fold or articulation, which can be traced all round. It is, as we have seen, composed of a broad labrum and three pairs of gnathites, which latter have only their terminal segments free; and these surround a conical hollow, at the bottom of which lies the opening of the œsophagus. The prominence of the whole mouth appears to result from the lateral fusion of the two basal segments of the three pairs of gnathites. I have examined the mouth of ordinary Crustaceans, and can see no trace of a structure like this. That there has been some union of the parts is indisputable; for the palpi, which in ordinary Crustaceans are quite free, are here firmly united to the upper and outer corners of the labrum; and indeed, at first appear to be more intimately connected with the labrum than with the mandibles. The palpus on its upper and exterior surface, is in direct continuity with the square thickened piece of membrane, on which the mandible is articulated, and likewise with that side of the upper or free portion of the mandible which faces the labrum. This face of the mandible, beneath the toothed edge, is hollowed out or arched (Pl. 26, fig. 5, p), owing to the above-mentioned continuity of its membrane with that of the palpus. On the lower surface, the palpus is[Pg 79] firmly united to the lateral corners of the labrum; or indeed the corners of the labrum may be almost said to be formed by the soft, swollen bases of the palpi: the point of union, when viewed from the outside, is seen to form a knob on the shoulder of the labrum, beneath the level of its crest, and at this knob (Pl. 26, fig. 3, close to d′) several thickened bands in the surrounding membrane unite. The free portion of the palpus stands out transversely behind (i. e. anteriorly to, in a homological sense) the labrum. I suspect that the palpus possibly may consist of two segments, of which the terminal one is free, and the lower one confluent with the labrum.
Before proceeding any further, I should observe that figs. 3 and 4, in Pl. 26, represent the membranes of the mouth of Coronula diadema, perfectly cleaned. In fig. 3, all the front part of the mouth has been removed, the mandible on one side, the labrum with the two palpi, and the œsophagus being alone left, and these are viewed from the inner side; the front part, however, of the supra-œsophageal cavity has been cut away. In fig. 4, the labrum, with the œsophagus, has been removed, whilst the two outer maxillæ, the right-hand inner maxilla and mandible (with the exterior and basal portions, d, d′′, of one palpus) are seen from the outside; but in order that these parts should all be shown, the whole of the right-hand side of the mouth has been spread out, for the teeth of the mandible should have stood in a vertical line between the two outer maxillæ. In the mandibles, the free upper part is separated, by a distinct articulation, from the square piece of thickened membrane (fig. 3, c1) on which it is supported; and this latter is separated by a second articulation from a portion of thickened membrane (c2), the basal edge of which forms the third and lowest articulation, separating the mouth from the body. This basal, thickened portion of membrane curls round and inwards, towards the outer maxillæ or front of the mouth, and its terminal points sometimes even penetrate a little way within the muscles, like apodemes: it is not distinctly separated by any line or suture from the membrane, which forms the whole broad labrum; so that I at first concluded that the labrum dipped under the mandibles, and thus afforded a support on which they were articulated; but this appears so opposed to all[Pg 80] analogy, that it is more probable that the above basal thickened portion of membrane is truly the basal segment of the mandibles, completely confluent with the labrum; and it is, I think, not very improbable that even a large portion of what in appearance belongs to the labrum, namely, those concavities to which the muscles of the mandibles are attached, may, also, be part of the basal segment of the mandibles. Whether or no there really are two segments beneath the upper free portion of the mandibles, which have become laterally confluent with other parts, I must think that the square thickened piece of membrane (c1) represents at least one segment. I may here observe, that Prof. Milne Edwards seems to consider the mandible of the higher Crustaceans as answering homologically to the haunch of the leg; but, according to M. Brullé,[35] there ought to be two basal segments (sous-maxillaire and maxillaire) bearing the proper mandible, and giving rise, on the outer side, to the palpus,—a structure which perfectly corresponds with my view of the mandible and palpus in Cirripedes.
[35] ‘Annales des Scienc. Nat.,’ 3d series, Zoolog., tom. ii, p. 271.
Maxillæ: the point whence the long apodeme (b′, Pl. 26, fig. 4 and fig. 7) arises, according to Audouin’s views, must mark an articulation, and this would separate the upper free segment from the lower segments, which I believe to be laterally confluent with the organs on each side. The thickened membrane, of which the upper free part is formed, extends a little distance beyond the insertion of the apodeme; and this small portion beneath the point of insertion may possibly answer to the square, thickened piece of membrane, or second segment, supporting the mandibles. Beneath it, a rather wide expanse of thin, flexible membrane reaches down to the basal fold surrounding the mouth, and may thus form the third segment.
Outer Maxillæ: the upper free segment has a spinose lobe (a′′, Pl. 26, figs. 2 and 4), on its inner face, which may indicate a lower and second, almost free segment. Passing over this, we have, on the outside of the mouth, beneath the free, upper segments, an expanse of membrane, which, on the side, close to the inner maxillæ, is perforated (Pl. 26, fig. 4, n) by orifices which I believe are olfactory. In some species, as in Bal. eburneus and improvisus, there[Pg 81] is a longitudinal medial suture in this expanse of membrane, which I suppose indicates the lateral confluence of the middle segments of the two outer maxillæ. A short, transverse articulation or fold separates this middle segment (fig. 4, a1) of each maxilla from the third or basal segment; and this latter (a2) is separated from the body by a very distinct fold, which (at least amongst the Lepadidæ) sends inwards a short, medial, tongue-formed apodeme. Here, then, we apparently have, as in the mandibles, two segments under the upper free segment of each outer maxilla, laterally confluent with the adjoining organs. But I must state that, in old specimens, and only in old specimens of Coronula diadema, I have found under the outer maxilla an additional transverse ridge and fold, which plainly shows how easily a mere thickening of the membrane might be mistaken for an articulation. I can, however, hardly persuade myself that the articulated membrane, under the free part of the mandibles, which has now been figured and described, has no homological signification; and the fusion of the palpus and labrum seems too plain to be mistaken. Hence I must conclude that the mouth, in the Cirripedia, does truly exhibit a compounded structure of a very peculiar nature.
There are always six pairs; each biramous and multiarticulated, supported on a pedicel formed of two segments. A shield-like swelling at the exterior bases of these pedicels often appears like another segment; but such, I believe, is not its nature. The five posterior pairs answer to the five pairs of ambulatory legs in the higher Crustaceans; and as in the case of the latter, the three, or the four hindermost pairs almost invariably resemble each other. The first pair, which is homologous with the outer maxillipeds of ordinary Crustaceans, is separated by an interval from the second pair;—though this is not the case with the legs of the pupa, from which the cirri are metamorphosed. These anterior cirri are attached to the lateral edges of the mouth, namely to the thickened rim of membrane, forming the supposed basal[Pg 82] segment of the mandibles. They are capable of more diversified movements than the other cirri: the anterior ramus is always elongated, with the terminal segments more or less tapering, and is directed beyond (or anteriorly to) the mouth: the shorter ramus closely resembles in structure the rami of the second pair. In the Chthamalinæ the second pair, and in the Balaninæ the second and third pairs (as will be more particularly described under these sub-families) differ in structure from the posterior pairs, from which they are separated by a slight interval. The number of segments on the posterior cirri is often great, amounting in Chelonobia even to fifty. Each segment normally is furnished on its inner face, which is usually somewhat protuberant, with from two to rarely eight or ten pairs of long spines or bristles, placed in a double row; the two spines in the lower pairs stand nearer to each other, and are shorter than the spines in the upper pairs. Between each pair of spines there is either a single, very thin bristle, or often a tuft of such. The pairs are directed somewhat upwards, and they diverge when the cirri are uncurled; their function is obviously to entangle the prey. On the dorsal or exterior surface of each segment, close to its upper margin, there is a tuft of spines, often composed of thicker and thinner spines; these, I believe, serve to prevent any creature intruding within the sack. On both sides of the upper margin of each segment, there is generally a row of short, blunt, excessively minute spines, which only deserve notice, inasmuch as it is by their increase in number and size, and by the spreading out of the dorsal tufts, and, lastly, by the increase of the little tuft intermediate between the pairs of spines situated in front, that the segments on the two or three anterior pairs of cirri become covered, like brushes, with bristles. The bristles or spines on the second and third cirri are often, especially in Tetraclita, doubly and coarsely pectinated. The bristles on the pedicels follow the same arrangement as on the rami; namely, being in regular pairs on the posterior cirri, and crowded thickly, like a brush, on the anterior cirri. The segments in the shorter ramus of the first cirrus, and in both rami of the second, and often of the third cirrus, are broader than[Pg 83] the segments of the posterior cirri; they are, also, especially in the genus Balanus, frequently produced in their upper, ante-lateral corners, into remarkable prolongations (see Pl. 29, fig. 4, of the third cirrus of Bal. perforatus), clothed on their inner surfaces, and at their extremities, by numerous bristles. The number of the segments in each cirrus is in some degree variable, and increases with age; this is likewise the case, to a certain extent, with the number of the spines borne on each segment.
As compared with ordinary Crustaceans, I presume the two rami answer to the “tige” and “palpe” of Milne Edwards; and the pedicel (as I have called it) to the two basal segments of the leg.[36] The “fouet” or flabellum does not appear to be developed in any Cirripede; for though the filamentary appendages in certain genera of Lepadidæ, might at first be thought to be of this nature, yet their usual position beneath the basal articulation of the first pair of cirri, and the occasional presence of more than one, proves, I think, that such is not the case.
[36] According to this author’s new nomenclature, the pedicel would consist of the coxopodite and basipodite; the tige would be the ischiopodite and following segments; and the palpe would be the exopodite; the epipodite or flabellum being absent. (‘Annales des Sciences Naturelles,’ tom. xviii, 1852.)
Though the structure of the cirri is very uniform, yet we meet with some peculiarities. In Chelonobia, the segments of the posterior cirri bear only two pairs of main spines; whereas in some varieties of Balanus balanoides, they carry as many as ten pairs in a longitudinal row; but in this latter species, the number of these spines varies, in a singular manner, from six to ten pairs. In Tubicinella, the pairs of spines on the segments of the posterior cirri are arranged so closely one under the other, that they appear almost like a single transverse row. Considering the whole family, the third pair of cirri differs most in structure in the different genera. Thus, in Chthamalus antennatus, the anterior (or outer) ramus (Pl. 29, fig. 3) is thicker and much longer than the posterior (or inner) ramus; the number of the segments in one instance being, in the two rami, 53 and 18; in the longer ramus, the spines are arranged abnormally, tending to form a little circle round each segment; and[Pg 84] the whole ramus may be said to be antenniformed, and I believe acts as an organ of touch: the relative number of the segments, I may add, in the two rami and the arrangement of their spines varies greatly in this species. In two other species of the same genus Chthamalus, we have occasionally the anterior ramus in some degree antenniformed, so that this whole structure is variable. In the allied Chamæsipho columna, it is the posterior or inner ramus which is antenniformed, but this peculiar development is more plainly marked in the case of the second pair of cirri than in that of the third pair. In Tetraclita porosa it is, also, the posterior ramus of the third pair which is antenniformed; in this third pair, and indeed in the other cirri, the relative numbers of the segments vary extremely. A similar structure in the third pair, but in a lesser and variable degree, may be observed in some of the other species of Tetraclita. In Balanus vestitus, also, we have, in the third pair, an analogous structure. It is scarcely possible to believe that the circumstance of the second pair of legs, which answer to the third pair of cirri, being antenniformed in certain decapod Crustaceans, is an accidental coincidence; it must be owing to some special affinity in the two groups.
In Chelonobia, the third pair of cirri is of unusual length compared with the second pair, but does not otherwise differ from the type of its sub-family: in Coronula and its allies, on the other hand, the third pair is very short and broad, as may be seen (Pl. 29, fig. 5) in Xenobalanus: in this latter genus, the front surfaces of the segments of the pedicels (fig. 6) of the posterior cirri, are extremely protuberant, almost as in Scalpellum vulgare.
The last peculiarity in the cirri at all worth mentioning, is in the sub-genus Acasta, in which, differently from in all other known Cirripedes, the anterior ramus of the fourth pair does not absolutely resemble the rami of the fifth and sixth pairs; in most of the species, the spines on this anterior ramus are more crowded together, are larger, and are mingled with some short thick points; and the spines in the dorsal tufts are also longer than in the two posterior pairs of cirri; but in A. sulcata (Pl. 29, fig. 2), and in a lesser degree in A. cyathus and A. purpurata, the front margins of[Pg 85] the lower segments of this anterior ramus, and of the upper segment of the pedicel, are developed into strong, downwardly curved teeth: it is very remarkable that so beautiful a structure should be extremely variable, as it certainly is in Acasta sulcata.
With extremely few exceptions, these are present in all the Lepadidæ and Verrucidæ; whereas amongst the Balanidæ they occur only in the two species of Pachylasma, and in one species of Catophragmus; these being the genera most closely allied to the Lepadidæ, and where, consequently, their presence might have been anticipated. These appendages are seated close together over the anus; they are multiarticulate, each segment being sub-cylindrical, with a few small bristles round its upper edge.
I have not much on this head to add to what I have said under the Lepadidæ. As in that family, the strong internal membrane of the œsophagus terminates in a remarkable, bell-shaped expansion (Pl. 26, fig. 3, g′), which, as observed by M. St. Ange, serves to keep the upper broad end of the stomach expanded. The œsophagus is well furnished with constrictor and radiating muscles for closing and opening it; and it is thus capable of a strong swallowing movement. The stomach runs down to the lower end of the prosoma, and then doubling back on itself extends to the anus. As the prosoma is much elongated in Tubicinella and Xenobalanus, so is the stomach of unusual length in these genera. In several species of Balanus, the upper edge of the stomach is surrounded by from six to eight cæca; these cæca I ascertained, in Balanus perforatus, are branched, and penetrate a considerable way into the body; and some of them at least expand a little at their extremities. Each cæcum, from the manner in which it retained fluid, must, I think, be furnished, at the point where it enters the stomach, with a sphincter muscle. In Tetraclita, Chthamalus, Tubicinella, Coronula, and Xenobalanus, there are no cæca; but in Xenobalanus and Coronula balænaris, there are longitudinal, approximate folds in the upper, broad end[Pg 86] of the stomach, which would serve to expose the food to a greater extent of digesting surface.[37]
[37] The presence and absence of these cæca in genera so closely allied as Balanus and Tetraclita, shows, I think, that these cavities are not of high importance; and I must doubt whether Von Siebold’s view (‘Anatomie Comparée,’ tom. i, p. 445), that these cæca form a passage to a true or isolated liver, such as exists in the higher Crustacea, can be admitted. Cæca are said by Von Siebold to occur in some of the Entomostraca, as Daphnia, Argulus, &c.
As in the case of the Lepadidæ, a transparent, structureless, epithelial tube, composed of chitine, containing more or less digested food, is found, in specimens preserved in spirits, occupying the whole length of the stomach, and where there are cæca, sending branched prolongations into them. It does not extend into the œsophagus or into the rectum. This epithelial tube or model of the stomach, filled with excrement, is expelled by the rectum, whole, that is in a single piece, as I observed in some living specimens of Balanus balanoides: in some specimens, however, of Chthamalus stellatus, the excrement was ejected, perhaps from the animal being confined, in fragments, and the sack thus became befouled. Beneath the epithelial layer, the stomach is lined by a delicate, pulpy and cellular mucus layer, which easily peels off in flakes: this is surrounded by a muscular layer with the fibres closely approximated and transverse; and this by a layer of stronger, longitudinal muscles, but more distant from each other. Lastly, outside this double muscular layer, there is a rather thick, somewhat laminated, pulpy layer, abounding with cells, often nucleated, and frequently containing much oily matter. This structure agrees closely with Dr. C. H. Jones’s[38] account of the external covering of the stomach in Daphnia, and which he believes to be hepatic: as in Daphnia, there does not seem to be any ducts. I may here observe, that within the upper part of the prosoma, but not immediately connected with the stomach, I have often observed much white pulpy substance, permeated by lacunal passages, and exhibiting no structure except some excessively minute cells.
[38] ‘Philosophical Transactions,’ 1849, p. 116. Karsten (‘Nov. Actorum Acad. Nat. Cur.,’ 1845, tab. xx) has excellently figured the testes, as the hepatic glands; and has indicated the ovaria as salivary glands; it is singular that this anatomist overlooked the ducts which lead from his supposed hepatic glands, into the vesiculæ seminales, within which he observed spermatozoa.
The rectum, lined by membrane continuous with that investing the thorax (and seen through it, in Pl. 26, fig. 8, c), extends inwards to about opposite the bases of the third or fourth pairs of cirri. It is longitudinally plaited; the ends of the folds forming a sort of valve where joined on to the stomach. It is coated by circular, transverse muscular fibres: judging from the movements, the anus itself is surrounded by a strong sphincter muscle. The anus opens on the dorsal surface of the thorax (fig. 8, b); but as in the genera, in which caudal appendages occur, it opens under them, the orifice, I believe, is homologically terminal, and owes its dorsal aspect to the aborted state of the whole abdomen, and to the great development of the probosciformed penis; for the anus may be said to be situated on the dorsal base of this organ.
Altogether we see that the alimentary canal is of a very simple structure. The food, judging from the contents of the stomach, seems generally to be composed of infusoria and minute animals: but in the case of Tetraclita, I have been surprised at the size and number of the included amphipod, isopod, and entomostracan Crustaceans, in one case, together with an annelid. I have, also, sometimes seen some confervoid matter within the stomach.
On this subject I can add nothing, except to express my conviction that there is no heart, or true vessels; the circulation being strictly lacunal. A passage has often been quoted from Poli, in which he states that he saw a pulsating organ, close above the anus; but I have seen this movement, which appeared to me to be a convulsive twitching of the sphincter muscle of this orifice. The largest lacunal channel extends down the middle of the rostral compartment of the shell: and this answers to the rostral channel down the peduncle in the Lepadidæ. Large nerves and the main pair of unbranched ovarian tubes (Pl. 25, fig. 1, leading into g) extend along this channel. At the basis (at least in Coronula) this channel joins on to a large circular one, running all round the sack, and sending off branches into the mass of ovarian tubes and cæca.
It has been shown in my former volume, that in Lepas and in some other genera of the Lepadidæ, there are six main ganglions; one supra-œsophageal, and five infra-œsophageal, or thoracic. In Pollicipes, however, there are only four thoracic ganglions, the last ganglion supplying the three posterior pairs of cirri with nerves, whereas in the other genera, the last ganglion supplies only the fifth and sixth pairs of cirri. In this genus, moreover, the lateral fusion of the ganglions has been so complete, that there is no evidence of their having been formed by the union of two. Amongst sessile cirripedes, we discover evidence of much higher concentration even than in Pollicipes. My chief examination has been confined to Coronula diadema, and to Balanus tintinnabulum: and in these genera we find (and the fact appears to me highly remarkable) as high a degree of concentration in the infra-œsophageal ganglion as in any decapod Crustacean, for instance, as in Maia, judging from the figure given by Milne Edwards; for all the nerves, with the exception of those connected with the supra-œsophageal ganglions, radiate from a single great ganglion.[39] The nervous system is, moreover, otherwise complicated.
[39] It must, however, be observed that, according to Mr. Dana, there is in certain suctorial Entomostracans, as in Caligus, only one infra-œsophageal ganglion. Mr. Dana speaks of this as resulting from reduction. In Cirripedes, from the gradation which may be observed from Lepas through Pollicipes into Balanus, the ganglions are certainly not reduced but concentrated. In Van de Hoeven’s figure of the nervous system in Limulus, there is seen to be no chain of thoracic ganglions; all the nerves rising from the circa-œsophageal collar; but this, on the other hand, seems hardly developed into a ganglion.
To begin with Coronula diadema the great infra-œsophageal ganglion (Pl. 27, fig. 1, A) is seated nearly opposite to the anterior margin of the second pair of cirri, which are homologous with the first pair of legs in the decapod Crustaceans. This ganglion shows no trace of any longitudinal medial suture; its shape is hardly discoverable, for it is formed by the union of eleven principal pairs of nerves, besides[Pg 89] several arising from its under surface; in outline, however, it may be said to be divided into a posterior and anterior half; the latter being somewhat heart-shaped, and the posterior half elongated. The nerves going to the five posterior pairs of cirri rise from the posterior margin of the ganglion, and run for some distance in a sheet, parallel and close together; the pair, however, going to the second pair of cirri soon branches off from the others. Each of these nerves enters at the inner and posterior margin of the cirrus to which it belongs, and, at least in the case of the first pair, divides into two branches as it enters. The nerves (Pl. 27, fig. 1, r5, r6) going to the fifth and sixth pairs of cirri are more closely united together than are the others, and appear, till they branch off, like a single large nerve. That which belongs to the sixth cirrus gives off, opposite to the fifth cirrus, a branch (s) nearly as large as itself, which enters the probosciformed penis. I may remark, that homologically this is the only abdominal nerve in any cirripede of the Order. From the under side of the nerves which run to the five posterior pairs of cirri, small branches are given off, extending dorsally into the thorax.
The anterior end of the great infra-œsophageal ganglion is formed by the union of a set of nerves, extending parallel in a bundle in a directly opposite direction to those running to the five posterior pairs of cirri. These nerves consist of an outer larger pair (r1) entering the first pair of cirri; and within these, and rather dorsally to their roots, we have the circa-œsophageal chord (c, c), or collar nerve; between the roots of the latter, and on the ventral surface (or near side of the figure), there are three closely united, small pairs, running to the gnathites, and, as I believe, to the olfactory sacks. From the under (or dorsal) surface of the anterior end of the ganglion, two nerves, larger even than the circa-œsophageal chord, and which I shall call the splanchnic pair (d, d) arise; and the singular course of these nerves will presently be described; between this great pair, there is a single (b) medial nerve, which runs down and branches into that large diverging muscle, which is attached to the upper ventral surface of the stomach. Posteriorly to these three nerves, we have two pairs of much smaller nerves (not[Pg 90] figured), running dorsally into the body, so that we have seven nerves rising from the under surface of the infra-œsophageal ganglion. I need only further add, that on each side of this ganglion, between the nerves going to the first and second pairs of cirri, there is a moderately sized nerve (k), which appeared to run into the muscles of the thorax: a nerve in a similar position is figured by Milne Edwards in Maia.
The circa-œsophageal chord (c, c) nearly equals in length the whole distance from the centre of the main ganglion to the posterior end of the thorax. This collar bows out on each side, where passing the œsophagus (œ), which is seated at its anterior end. From the collar a branch is given off on each side, which I traced as far as between the mandibles and maxillæ; from analogy with other Crustaceans, it perhaps runs to the mandibles. The collar has not a transverse commissure, such as described by Milne Edwards in the Podophthalmia, and as figured by Van de Hoeven in Limulus.
The supra-œsophageal ganglions (B) present a singular contrast with the infra-œsophageal ganglion in their little development, size, or degree of confluence. They lie directly under the basal edge of the labrum. They are laterally quite distinct, and consist merely of a slight enlargement of the circa-œsophageal chord. From the anterior edge of each ganglion, a broad nerve (f) extends for some distance in a straight line, and, on close examination, can be seen to be formed of two nerves closely united, of which the inner and smaller one, after a space, appears to cross over the larger nerve: both become at this point tortuous, and, giving off branches (m, m), form a plexus. The two nerves (f) then bend inwards, and almost touching each other, run down, together with the two ovarian simple ducts, along the rostral compartment of the shell. No doubt, if the smaller branches from these nerves could be traced, they would be seen to form a network over the whole sack; and would therefore enclose, as in a cage, the rest of the nervous system. These nerves correspond, I believe, to the two pair of antennular nerves of ordinary Crustaceans, and hence I will call them by this name.[Pg 91] Just in front, at the outside corners of the two supra-œsophageal ganglions (B), a branch (e′) arises, which I traced to the ends of the adductor scutorum muscle, and to those several muscles which serve to retract the interspace of membrane between the mouth and the adductor.
The pair of great splanchnic nerves above alluded to, which arise from the anterior and dorsal surface of the infra-œsophageal ganglion, are in Pl. 27, fig. 1, d d, (and in fig. 2), laid flat; but in nature they first bow outwards, and then, penetrating deeper into the body, approach each other, and running nearly parallel, pass round the lower end of the œsophagus: their course consequently is nearly similar to that of the circa-œsophageal chord, with this difference, that the outwardly bowed portion is situated near the infra-œsophageal, instead of near the supra-œsophageal ganglion. The splanchnic nerve, a little beyond the supra-œsophageal ganglion, joins a plexus (d′); and into this plexus another large nerve (e) which I will call the supra-splanchnic nerve, sends branches; this nerve takes an almost semicircular bend over the ovarian glands. The supra-splanchnic nerves (e, e), though appearing to spring from the supra-œsophageal ganglions, do really arise, as may be seen by tracing the constituent fibres, from the circa-œsophageal chord. The plexus (d′) lies close to the coats of the upper end of the stomach: several branches, proceeding from it, run further on, but I was able to trace only a few of them: one went (at least in the case of Balanus perforatus), to the adductor scutorum muscle: another branch spread out on the flanks of the prosoma: I strongly suspect that one branch goes to the acoustic sack: it appeared, also, as if some of the small branches entered the second plexus (m), where the inner antennular nerve and ophthalmic nerve cross over the outer antennular nerve.
I have called the nerves (dd, ee) splanchnic and supra-splanchnic, from their course and apparent function in supplying the viscera. In the descriptions of the nervous system of other Crustaceans I can find nothing analogous to my great splanchnic nerve (dd); the so-called supra-splanchnic nerves (ee), which arise from the circa-œsophageal chord, seem to be the analogues of the ordinary splanchnic[Pg 92] nerves, though these latter are always described as uniting into a single medial branch. The plexus (d′) is the cervical ganglion of M. Martin St. Ange,[40] who has likewise indicated the course of my splanchnic and supra-splanchnic nerves; but the plexus, when viewed as a transparent object, hardly appears to me to be ganglionic in its nature. In my former volume on the Lepadidæ, I quite misunderstood the course of these splanchnic nerves.
[40] ‘Mémoire sur l’Organisation des Cirripèdes,’ p. 19.
From the commissure between the two supra-œsophageal ganglions, a straight chord (Pl. 27, fig. 1, g) arises, which terminates in a small ganglion (C), barely exhibiting traces of being formed of two laterally confluent ganglions. This is the ophthalmic ganglion. The chord connecting it with the two supra-œsophageal ganglions is accompanied by a small nerve (h) which runs on to the muscles round the adductor scutorum muscle; the chord is encased by much fibrous tissue, and its dissection is thereby rendered difficult. From the ophthalmic ganglion, on each side, a nerve (i) goes forth and crosses the antennular nerve; these, if I could have traced them, would have been found to run, as may be safely inferred from what is known in Balanus tintinnabulum, to a pair (D, D) of eyes.
In Balanus tintinnabulum, the structure of the great infra-œsophageal ganglion (Pl. 27, fig. 2, A) is essentially the same as described under Coronula. The great pair of splanchnic nerves springing from its under side, are here actually twice as large as the circa-œsophageal chord. The plexus (d′) formed by the splanchnic nerve (d), on each side, with the supra-splanchnic nerve (e), which arises close posteriorly to the supra-œsophageal ganglion, is here much less complicated, but is perfectly distinct; and there was no appearance of the cervical ganglion of M. Martin St. Ange. The chord (g) running from between the two supra-œsophageal ganglions to the ophthalmic ganglion, is nearly as large as the double antennular nerve (f) on each side of it. The ophthalmic chord (which is accompanied in its whole course by a small branch running to the adductor scutorum muscle) terminates in a small ophthalmic ganglion (C), which seems[Pg 93] to be formed by the almost complete fusion of two ganglions. This ganglion is hardly larger than the chord which it terminates: it appeared to me to give rise to more than one pair of nerves, and a single nerve (to my surprise) joined the branch just mentioned, which goes to the adductor scutorum muscle.
From each supra-œsophageal ganglion, two closely united antennular nerves (f) extend, of which the inner one crosses over the main or exterior nerve, nearly opposite to the ophthalmic ganglion, and here forms (m) a plexus. The structure of this plexus I was not able, any more than in Coronula, to make out thoroughly, but I traced quite distinctly a long nerve (i) running from it into what must be considered as the eye. As in the case of Coronula, I traced a nerve on each side from the ophthalmic ganglion into the plexus, where I lost it; and as here in Balanus, I saw on each side of the ophthalmic ganglion a cut off nerve, of about the size of that which runs from the plexus on each side into the eye, I think we may safely conclude that the latter or optic nerve does really arise from the ganglion here called ophthalmic. I may add that the analogy of the nervous system in the Lepadidæ most strongly confirms the view of this latter being the ophthalmic ganglion.
The optic nerve (i), running from the plexus to the eye, is of considerable size; it runs nearly parallel to the main antennular nerve, diverging from it a little. It retains nearly the same diameter throughout; and gives off only one single, small, inner branch. It can be traced beyond the basal edges of the scuta, to just under the upper edge of the transparent opercular membrane, which unites the scuta to the sheath of the rostrum. The nerve itself, at a little distance from its further end, was, in a full-sized specimen, 5/1000ths of an inch in diameter; widening a little, it expands slightly, and abruptly terminates in a circular disc, about 8/1000ths of an inch in diameter, (see Pl. 27, fig. 5). The nerve just beneath this slight expansion, is coated all round by pellets of dark purple pigment-cells, but not actually united into a continuous layer. These pigment-cells are the more conspicuous from the surrounding parts being colourless. I could not make out distinctly any cornea; and I suppose the external[Pg 94] transparent membrane, to which the above slight circular expansion is attached, acts as such. This description very closely agrees with that given of these organs in Bal. rugosus of Gould, (B. crenatus?) by Dr. Leidy,[41] who first discovered the eye in the adult cirripede, but he did not observe the ophthalmic ganglion. These eyes differ from those in some of the genera of the Lepadidæ, only in the greater length of the optic nerve, and by standing laterally further apart from each other.
[41] ‘Proceedings of the Acad. Nat. Sciences of Philadelphia,’ vol. iv, 1848, p. 1. I may add that I have, also, observed the supra-œsophageal and ophthalmic ganglions in Bal. perforatus.
I may here mention that I tried a few simple experiments on the senses of Balanus balanoides, B. crenatus, and Chthamalus stellatus. I found these three species very sensitive to shadows, that is, to an object like my hand passing even quickly, and at the distance of about a foot, between them and the source of the light.[42] They were indifferent to a gradual change from bright to obscure light; but instantly perceived and drew in their cirri, when my hand was passed between the basin in which they were kept and the window, even when this was tried rather late on a dusky evening; and likewise when my hand was passed between them and a single candle. I took, of course, the precaution of passing my hand in other directions, but this never produced any effect. These species are moderately sensible to any vibration in the vessel in which they were kept, but they were indifferent to noises made in the air, or in the water. I found it impossible to touch, under water, an individual shell ever so lightly with a needle, without all the immediately surrounding individuals, when several adhered together, perceiving it, and retracting their cirri: it made no difference whether the one touched had already withdrawn its cirri and was motionless: from this fact, and from seeing that a similar but slighter effect was produced by touching the rock on which the specimens adhered, I infer that the perception by the others of the one being touched, is communicated by vibration.[Pg 95] When an individual was touched under water, not by a needle, but by a pointed camel-hair brush, it generally withdrew its cirri, but the neighbouring specimens took no notice: when touched by a single hair of the brush, no notice was taken, unless the skin of the orifice leading into the sack was so touched. In these trials, it is of course necessary carefully to avoid intercepting the light. I could not make out that cirripedes perceived odours diffused in the water.
[42] I find that this fact was long ago observed by Von Siebold, ‘Anatomie Comparée,’ tom. i, p. 434.
These are situated in the same position as in the Lepadidæ, namely, in a slight swelling on the sides of the thorax (Pl. 25, fig. 1, d′) just beneath the basal articulation of the first pair of cirri. The orifice in Tubicinella and Xenobalanus is slightly produced, or is tubular; the free part in the former genus projecting 5/100ths of an inch. The structure of all the parts is essentially the same as in the Lepadidæ, but I think all are proportionally larger. The external membrane of the body is turned inwards at the orifice, as a short flattened tube, which widens considerably (being, in a middle-sized specimen of Coronula, 4/100ths of an inch in width) before it abruptly terminates. The meatus, as I have called the sack-like cavity which encloses the true acoustic sack or vesicle, is formed of pulpy membrane, and is apparently continuous with the corium of the whole body, but by dissection it can be separated entire. The acoustic vesicle is of various shapes, as we shall immediately see; but in all essential respects it is identical with the same part in the Lepadidæ; it is formed of the same peculiar, soft, elastic, brownish, transparent tissue, which seems to be composed of fine, transverse pillars, becoming towards the outside fibrous, and at their inner ends appearing when viewed vertically from above, like hyaline points. In Coronula diadema, I observed on the outside of the acoustic vesicle, some excessively minute bristles, only 1/3000ths of an inch in length, seated on little eminences. I examined carefully the contents of the vesicle in this species, in specimens well preserved in spirits, and there was nothing within but a very little, thin, pulpy fluid, and a few yellowish nucleated cells, here and there aggregated into small groups. In Coronula, the flattened acoustic vesicle is[Pg 96] elongated, with a somewhat sinuous, but not very irregular margin (Pl. 27, fig. 4), and is without any ridges on the surface; its neck or orifice projects at right angles to the elongated portion, which stands obliquely to the tubular orifice of the meatus. In a moderately-sized specimen of Coronula diadema, the elongated portion of the acoustic vesicle was, 6/100ths of an inch in length. In Tubicinella, the acoustic vesicle is heart-shaped, with the neck attached to its broader end; and the surface is covered by zig-zag ridges. In Balanus tintinnabulum (fig. 3), the acoustic vesicle is almost square at the lower end, with the neck placed at one of the upper corners; on the external surface, there is an oblique prominent ridge or fold, which sends off downwards another ridge; its length, in a large individual, was 5/100ths of an inch.
In all these cases, the acoustic vesicle is mainly attached by its neck, to the upper end of the sack-like meatus; but there is likewise a layer of soft, pulpy, cellular matter, slightly connecting that side of the vesicle which is opposite to the neck, with the walls of the meatus or outer sack. The mouth or orifice of the vesicle is closed by a delicate lid or diaphragm, which can easily be separated; and this diaphragm is formed by the expansion of a large nerve, which here abruptly terminates. In a very large specimen of Coronula diadema I clearly made out the existence of this nerve, and traced its course for some distance from the point where the summit of the meatus and the neck of the vesicle are joined together; the nerve first runs posteriorly, and then turns inwards and doubles back or anteriorly; and I clearly followed it to the antero-lateral sides of the uppermost end of the stomach, where it seemed to enter a ganglion, so that I unfortunately cut it off, but found only a slight plexus, with the cut off nerve apparently running onwards with nearly the same diameter. The diameter is great, fully equalling, in its widest part, that of the circa-œsophageal chord; but it is very much flattened, and so has not nearly so much bulk as that nerve. Before it reached the stomach, it gave off one branch, which ran towards the mouth. The only nerves which, from their size, could, I think, be continuous with this from the acoustic sack, are[Pg 97] the main branches proceeding from that plexus (d′) formed by the interbranching of the splanchnic and supra-splanchnic nerves.[43]
[43] I have always feared that anatomists would reject my view of these organs being acoustic, owing to the absence of otolithes; but I observe that so high an authority as Von Siebold (‘Anatomie Comparée,’ tom. i, p. 433) does not believe that otolithes occur in the acoustic organs even of the highest Crustacea. He considers an “ampoule volumineuse, a parois mince, remplie d’un liquide transparent,” and a “membrane tympanique,” though having a fissure in the centre, as sufficient. I may here remark, that the nerve proceeding from the acoustic vesicle in Cirripedes, and apparently running to the splanchnic nerve, may easily be placed in connexion with the antennular nerves, by the second plexus (m) in figs. 1 and 2, pl. 27. I should infer from Von Siebold’s remarks on his ampoule volumineuse in the higher Crustacea, that my acoustic vesicle answered to the labyrinth in higher animals.
I can add nothing to the account given of these organs under the Lepadidæ: I saw them in all the genera which I examined for this object. In Coronula diadema the orifices are large; they are seated in the usual position (Pl. 26, fig. 4, n), in the confluent segments, beneath the free part of the outer maxillæ, and somewhat exteriorly, or as near as possible to the inner maxillæ. In no sessile cirripede are the orifices produced or tubular, as is the case with several genera amongst the Lepadidæ. I failed, as heretofore, in tracing with certainty the nerve, which appears to enter the base of the sack, to its ganglion.
All the Cirripedes of the family we are now describing, are bisexual or hermaphrodite; and no instance has been observed of the presence of males or complemental males. I have very little to add to the observations made by M. Martin St. Ange and R. Wagner,[44] and to those given in my former volume. The testes seem always to be confined to within the thorax, including the prosoma. With their ducts, they resemble club-moss or[Pg 98] stag’s horns, with the extremities a little enlarged: a figure[45] of a small portion from Balanus perforatus is given in Pl. 25, fig. 2. It is quite surprising how like in structure and appearance the branching ovarian tubes often are to the testes with their ducts; but the latter are of smaller diameter. Two main ducts generally unite just before entering the broad, often reflexed, end of the vesicula seminalis: in Coronula balænaris, however, I observed four ducts entering this receptacle. The two vesiculæ seminales, lying within the thorax and prosoma, are usually very long and tortuous: they are formed of a thin inner tunic, which is strengthened by thicker reticulated lines, and of an outer layer of transverse fibres, which are either elastic, or probably muscular, as they serve to expel the contents with force when the end is cut off. The inner tunic is prolonged up the probosciformed penis, at the base of which the two vesiculæ unite.[46] The contents of the vesiculæ are commonly pulpy and cellular; and from the cells the spermatozoa are developed; soon after their development, they are, as it appears, expelled.
[44] The ‘Report’ on M. Martin St. Ange’s memoir was laid before the Academy of Sciences, July 14, 1834, so that I suppose it was read previously to this date. R. Wagner’s paper was published in ‘Müller’s Archiv,’ 1834, p. 467. Burmeister’s ‘Beiträge zur Naturgeschichte der Rankenfüsser,’ was published this same year, 1834; so that these three authors published almost contemporaneously.
[45] A far better figure is given by Karsten (‘Nov. Act. Acad. Cæs. Nat. Cur.,’ 1845, Pl. 20, figs. 2, 3, 4), but under the erroneous supposition that these organs were hepatic.
[46] In Conchoderma aurita, the ducts, as shown by Burmeister (‘Beiträge,’ &c. tab. ii, fig. 17), unite half way up the probosciformed penis.
I have seen the spermatozoa in Balanus crenatus, perforatus, and balanoides, and in Chthamalus stellatus. The cells, from which the spermatozoa are developed, and which are often found in vast numbers within the vesiculæ, are on an average about 1/5000th of an inch in diameter. The spermatozoa differ remarkably within the vesicula of the same individual, according to their state of development. I have observed in B. perforatus and in the Chthamalus, that the shortest, and therefore, I presume, the youngest (Pl. 29, fig. 7, a), had a globular head with no projection in front: as they increased in size, this head became less in diameter, and a short tapering filament, (a, b,) like the tail, projected out of it. This anterior filament does not lie in exactly the same line with the posterior filament, which is straight as an arrow. In Bal. crenatus, the anterior filament was 1/2000th of an inch in length, and the posterior filament 4/2000th, giving a[Pg 99] total length of 5/2000th: in the longest and best developed specimens of Chthamalus stellatus, the nodular enlargement was much elongated and spindle-shaped, and not above half the diameter it had in the earliest stage; the posterior filament (measured from the front of the enlargement, this consequently being included) was 5/2000th in length, and the front part only 1/4000th, giving a total length of 11/4000ths of an inch. These observations agree pretty well with Kölliker’s;[47] but this author states, that perfectly developed spermatozoa are absolutely without any nodular enlargement: if this be the case, I have never chanced to see the spermatozoa in their perfect condition. Mr. Bate, also, figures some (Pl. 29, fig. 7, c) in this state, without any enlargement.
[47] ‘Annales des Sciences Naturelles,’ (2d series), tom. xix, p. 348. Kölliker refers to Wagner’s paper on the same subject, in Wiegmann’s ‘Archiv,’ 1835, part ii, pl. iii, fig. 9. He also refers to Von Siebold’s observations. Mr. C. Spence Bate has figured, in the ‘Annals and Magazine of Natural History’ (vol. viii, 2d series, 1851), the spermatozoa of Balanus balanoides, perforatus, and of Verruca (Clitia) Strömia, and of these I have given copies, Pl. 29, fig. 7.
The probosciformed penis lies adpressed on the under side of the thorax, with its apex generally projecting between the first and second pairs of cirri. It presents the same ringed or articulated structure as in the Lepadidæ: it arises from an unarticulated projection or support, which also forms the posterior border to the anus. This support often terminates, as first observed by Poli, in a very sharp point; but this point cannot be of much functional importance, for though present in Balanus balanoides, it is absent in the closely allied B. crenatus; in Tubicinella there is only a rudiment of this point; I have not observed it in any member of the Chthamalinæ. The strong, transverse and longitudinal muscles with which the penis is furnished, are attached to this support. The apex or orifice of the penis is, I believe, invariably surrounded by some bristles. Its length varies much, according to its state of contraction or relaxation; and this again, I believe, is dependent on the condition of the male secreting organs. In a small specimen of Elminius modestus, the penis was actually thrice as long as the whole thorax, including the prosoma: in Pachylasma and in Octomeris angulosa, the penis is very short, being equal only to once and a half the length of the pedicel of the sixth[Pg 100] cirrus: in Octomeris brunnea, the unarticulated support is much elongated, being as long as the pedicel of the sixth cirrus, in which respect this organ resembles that of Ibla quadrivalvis, and of no other Cirripede. From the attachment of the penis at the posterior end and on the under side of the anus—from the position of the caudal appendages (where such occur) over the anus—from the position of these same appendages in the pupa—and lastly, from the position of the papilla-like penis in the abnormal Proteolepas, I infer that, homologically, the penis is situated at the apex of the abdomen, on its ventral surface; and that, consequently, this organ cannot be considered as the abdomen itself in a modified condition.
I have scarcely anything to add to the statements in my former volume. These organs consist of the true ovaria, or glandular bodies seated on each side, not far from the basal edge of the labrum; of the main or unbranched ovarian ducts; and of the (Pl. 25, fig. 1, g) ovarian branching tubes and cæca. I traced distinctly in Balanus, Tetraclita, and Coronula, the two main ovarian ducts, running from within the prosoma to the layer of inosculating, branching, ovarian cæca[48] which overlie the basis. In Coronula diadema one of these main ducts was 1/100th of an inch in diameter. Though I traced these ducts near to the grape-like, glandular masses,[49] which I cannot doubt are the true ovaria, I did not succeed in tracing them into actual connection. As in the Lepadidæ, these ovarian glands lie on the sides, near the basal margin of the labrum, and almost under, but rather to the outside of the antennular nerves. The branching and inosculating ovarian cæca form a layer, which corresponds with the mass filling up the peduncle in the Lepadidæ. In Tetraclita they do not cover the whole basis, but are confined to the circumference; they, however,[Pg 101] likewise extend up between the two layers of corium round the walls of the shell, and chiefly in the interspaces between the depressor muscles of the opercular valves. In Chelonobia, they enter between the radiating septa in the thickness of the walls: in Coronula diadema, they extend from over the basal membrane into the six large square chambers (Pl. 16, fig. 7, v) separating the radii and alæ: in Tubicinella they are confined to the basis: in Xenobalanus, they form a layer over the basis and likewise round the upper part of the peduncle-like body, which answers to the shell of other sessile cirripedes.
[48] These are well described in Lepas, by R. Wagner, in ‘Müller’s Archiv,’ 1834, p. 467. Von Siebold, I observe, refers to Burmeister as the first author who discovered the ovarian cæca within the peduncle; I had thought that M. Martin St. Ange had a prior claim.
[49] These are obscurely figured by Karsten (‘Nov. Act. Acad. Cæs. Nat. Cur.,’ 1845, Pl. 20, fig. 1d) as salivary glands; they were so considered by Cuvier and M. Martin St. Ange: I may observe that salivary glands have not been positively recognised in any Crustacean.
As after the most careful and repeated examinations of various Lepadidæ, I was convinced that there were no oviducts, so I have come to a similar conclusion in regard to the Balanidæ; the ova being brought to the surface, by the formation of a new membrane round the sack underneath them, and by the subsequent exuviation of the old membrane. The ova are united together by a most delicate tunic investing each egg; the ovigerous lamellæ being thus formed, as in the Lepadidæ. In the cases of Chthamalus stellatus, Balanus balanoides, and Platylepas decorata, I saw a pair of very distinct but fragile lamellæ. In Xenobalanus, the two ovigerous lamellæ form two sub-cylindrical packets, pointed at their lower ends and often cohering. There are no ovigerous fræna, for the attachment of the lamellæ; the ova being sufficiently well retained, as it would appear, by the well-closed shell. I have elsewhere stated my full belief that it is the ovigerous fræna which have been metamorphosed into the branchiæ of the Balanidæ. Most sessile cirripedes breed when very young; and I have every reason to believe that they breed several times in the year. The ova are ovate, and vary in length from 14/2000th of an inch in Chthamalus, to 19/2000th in some species of Balanus, in which this greater length was owing to a more elongated shape,—up to 25/2000th in some other species of Balanus. The ova are wonderfully numerous, especially in the genus Coronula.
I may here mention the singular case of some elongated specimens of Balanus balanoides, from Tenby, in South Wales: some of these presented nothing abnormal; but in no less than seven specimens, the two, three, or four poste[Pg 102]rior pairs of cirri, either on one or both sides, were in an almost rudimentary condition, being of small size and having a shrunk and wasted appearance. In six out of these seven specimens, the probosciformed penis was quite short and abruptly truncated, as if from abortion. By cutting off the truncated apex, and cleaning the external tissue, I ascertained that it was imperforate, apparently in all the cases, and I am certain of this fact in several of the cases. In three of the specimens, I examined the vesiculæ seminales; in one, I found some spermatozoa, but cohering together in a peculiar manner; in the second, there were no spermatozoa; and in the third, the vesiculæ were shrunk, empty, and quite rudimentary in size. So that these three individuals certainly could not have impregnated their own eggs; nevertheless, within the shell of these very three, there were perfectly developed larvæ: I am led to conclude from this fact, that adjoining specimens in a perfect condition had, by means of their long probosciformed penis, effected the fecundation of their imperfect neighbours. I need only further add, that some out of the above six specimens, with more or less aborted cirri and imperforate male organs, were infested by a peculiar parasite, allied to Bopyrus,[50] and that these specimens did not contain ova.
[50] I have given a short notice on this parasite, in my former volume on the Lepadidæ, in a foot-note to p. 55.
In my former volume, the metamorphoses were described under three principal stages or heads; but whether these three included all the main changes, I was then hardly able to conjecture. But now I have reason to believe that such is the case, for in the genus Cryptophialus, belonging to the Abdominalia, the whole course of the metamorphosis, from the egg to the pupa, takes place within the sack of the parent; and I found, when having, on the coast of South America, numerous specimens to examine, that the egg-like larvæ (Pl. 24, fig. 15-18) could be naturally grouped into[Pg 103] two main stages, but with many transitional intermediate grades (answering to the successive moults in the first stage of ordinary larvæ), before they passed into the third or pupal stage. And the first two stages in these egg-like larvæ of Cryptophialus, clearly seem to correspond with the first two stages in ordinary larvæ; for in both the chief changes are, the shortening of the terminal projection—the increase in size and approximation on the ventral surface of the anterior horns or cases for the antennæ—and the compression of the whole body. In all members of the Thoracica, the metamorphosis seems to run a remarkably uniform course. The larva in the first stage undergoes several moults and slighter changes—how many is not known—before arriving at its second main stage, which has been observed only in one single instance; and judging from Cryptophialus, this second stage passes abruptly by one moult into the pupal stage; and this, certainly, passes abruptly into the Cirripedial or mature stage.
The larvæ in this stage are known, amongst the Balanidæ, in Balanus, Pyrgoma, Coronula, Platylepas, and Chthamalus; and these genera include all the principal forms. Amongst the Verrucidæ they are known in its one genus, Verruca. Amongst the Lepadidæ, in Scalpellum, Ibla, Alcippe, Lepas, Conchoderma, &c.; and in all these genera the larvæ present no important difference—hardly any difference which could be viewed as generic, were these larvæ independent animals,—as may be inferred, chiefly, from Mr. C. S. Bate’s descriptions.[51] The abstract given in my former volume was not accompanied by any illustrations, and I have consequently here given (Pl. 29, fig. 8), a view of the larva, in the first stage before moulting, of Scalpellum vulgare: the natatory legs are not drawn with accuracy, only the relative position of the several organs having been carefully attended to. I have also had copied from Mr. Bate’s memoir, a figure of the larva (Pl. 29, fig. 9) of Balanus balanoides, in its first stage, before moulting, with its ventral surface exhibited; and another figure (with a few trifling alterations made after examining[Pg 104] specimens most kindly sent me by Mr. Bate) of the larva of Chthamalus stellatus (fig. 10), in its first stage, but after moulting once. It should be observed that Mr. Bate has given a drawing of the larva of this latter cirripede, in the first stage, before moulting; and it does not differ essentially from that just referred to (fig. 9), of B. balanoides, but is rather more fully developed. These drawings suffice to show the character of the larvæ in the first stage, both before and after the first moult, and even after the second moult, throughout the Order of Thoracica. The larvæ sometimes undergo their first moult within the sack of their parent, as I have been informed by Mr. Bate, and as I have observed in Coronula.
[51] ‘Annals and Magazine of Natural History,’ vol. viii (2d series), 1851, Plates 6, 7, and 8.
I will now make a few remarks on these larvæ in the first stage, before and after the first moult, supplemental to those in my former volume. Their shape is oval, and the whole dorsal surface is evidently covered by a carapace. It is remarkable that the body exhibits no distinct articulations; those given by Goodsir[52] being certainly erroneous. Commencing at the anterior extremity, the eye varies considerably in the state of its development; in Platylepas decorata it is nearly circular, and in most of the specimens very distinct; whereas in the allied Coronula balænaris, before the first moult, it is very imperfect, but afterwards square and of considerable size. In Balanus galeatus, in the immature larvæ dissected out of the egg, the cellular matter which was in process of conversion into the eye, formed a transverse band, obscurely divided into two portions, and this seems to indicate that the single eye is in fact formed by the confluence of two eyes. In Scalpellum vulgare, this heart-shaped eye lies between a V-shaped muscle, the nature of which I cannot understand, and which has not been represented in (Pl. 29, fig. 8, a). I need only further add, that in Chthamalus stellatus, after the first moult, the eye exhibits, in specimens sent me by Mr. Bate, some appearance of tending to become double.
[52] ‘Edinburgh New Philosophical Journal,’ July, 1843, Pl. 3, 4.
Arising posteriorly to the eye, we see, in Scalpellum vulgare, a pair of minute curved horns (b′), directed back[Pg 105]wards; and within these horns I distinctly saw an articulated organ. These horns are difficult to be distinguished, and probably could not be made out previously to the first moult, in any larva of less size than that of Scalpellum vulgare. But after the first moult, Mr. Bate has seen, in two species of Balanus, in Verruca and in Chthamalus (fig. 10, b), a pair of articulated organs, in this same position, evidently now forming antennæ, and directed anteriorly, and free from any envelope. It is somewhat important, as we shall presently see, to bear in mind that these antennæ first appear within an envelope or horn; and that I detected that they included an articulated organ, before I had heard of Mr. Bate’s observations. These antennæ, from their small size, from being seated internally with respect to the horns containing the other pair of antennæ, and from the position which the latter assume in the later stages of the larva, I believe to be the first or anterior pair. Their position in appearance posteriorly to the large lateral horns, containing the second pair of antennæ, is probably due to the anterior cephalic segments having been driven inwards, the truncated outline of the front of the head, and likewise, probably, the position of the mouth between the bases of the natatory legs being thus caused.
In this same larva of Scalpellum vulgare, within the great lateral horns just alluded to (fig. 8, c), filiform organs, supporting rows of spines, could be distinguished; and these appeared to me to be antennæ. These horns or cases resemble in structure the smaller pair just described; they arise from the ventral surface, and can hardly, therefore, be considered as prolongations of the carapace. After the first moult (fig. 10, c) they are seen to have increased much in length: in some cases they are of considerable length before the first moult, as in Lepas: in the Balanidæ they seem to be generally shorter than in the Lepadidæ; but in Balanus galeatus I found them one third of the entire length of the animal. Whilst within the egg, these horns are adpressed laterally to the body, and so point posteriorly; afterwards they project rectangularly from the sides, or, as in Scalpellum vulgare, are directed somewhat anteriorly. As in the larvæ of all ordinary Crustaceans, as yet known, the an[Pg 106]tennæ are amongst the earliest developed organs; and as the first pair of natatory legs (Pl. 29, figs. 8-10, e) in these Cirripedial larvæ, might so very naturally be thought to be antennæ (as has been remarked to me by Mr. Dana), both from their structure and from their position a little anteriorly to the mouth, I am well aware that to prove my view correct, namely, that these horns are the second pair of antennæ in process of formation, it is not sufficient merely to have seen organs resembling antennæ within them; nor is it sufficient to advance the strictly analogical fact of the first-mentioned pair of antennæ, which in Scalpellum indisputably appear in their earliest condition within an envelope or horn. Further evidence is required, and this is presented in Cryptophialus, in which the lateral horns of the egg-like larva, in its first stage (Pl. 24, fig. 16), can be actually followed step by step until, in the second stage (fig. 17), just before passing into the pupa, the horns are seen to have become larger and more nearly approximated to each other on the ventral surface; and whilst in this condition, I several times dissected out the prehensile antennæ of the future pupa with every character perfectly recognisable. Hence I cannot doubt that in the larvæ of Cirripedes the law of development is, that in their very earliest condition, the small first pair of antennæ are enclosed in cases; and that the large second pair remains thus enveloped until the pupal stage. This conclusion, we shall immediately see, is in harmony with the late development of the succeeding appendages or organs of the mouth, which certainly do not appear in the first larval stage, and are not known to appear even till after the final metamorphosis.[53]
[53] According to M. Joly, (‘Annales des Sciences Naturelles,’ 2d series, tom. xix, p. 59) in the larva of the macrourous Caridina, the natatory legs appear before the gnathites or parts of the mouth; so that in ordinary Crustaceans there is no invariable order of development from the anterior towards the posterior end of the body, as has sometimes been supposed.
The mouth is more or less probosciformed (Pl. 29, figs. 8-10, d), differing considerably in this respect in different species of the Lepadidæ; and this, probably, is due to the larva being born in a more or less mature condition. Its exact position likewise varies, for it arises either between the first or second pairs of natatory legs. It is known, from[Pg 107] Mr. Bate’s observations, to have the power of movement. It is directed posteriorly, the œsophagus extending anteriorly; both these directions being the same as in the mature cirripede. Certainly during these early stages there are no jaws or gnathites; but the margin, answering to the labrum, is furnished with some short, thick, sharp spines, and with hairs. In Scalpellum vulgare the orifice of the œsophagus seems to lie rather beneath the upper prominent spinose edge, which, as just remarked, probably answers to the labrum; but this is one of the species in which the probosciformed mouth, at least before the first moult, is not much developed.
We come, now, to the three pairs of natatory legs: the first (Pl. 29, figs. 8-10, e) has throughout the order only one ramus, whereas the two succeeding pairs (f, g) are biramous. I must here remark that the straight and strong, and the curved plumose spines, with which these limbs, after the first moult, become furnished, now appear to me as more probably prehensile, rather than masticatory as I imagined in my former volume. That these spines are important organs to the larvæ I do not doubt. With regard to the homologies of these three pairs of limbs, my first impression was that they were the mandibles and the two pairs of maxillæ in their earliest condition; but I consider this view as quite untenable, for several reasons; viz., the wide interval between their bases and the mouth itself,—the somewhat variable position of the mouth with respect to the legs,—and the position which the latter occupy in the second larval stage.[54] A far more tenable view is that these three pairs of legs are the three pairs of maxillipeds, in their earliest condition, in accordance with the view of M. Joly[55] on the nature of the three very similar pairs of natatory legs in the larva of Caridina, a macrourous Crustacean. But, in Cirripedes, the three pairs of natatory[Pg 108] legs, in the larva in the first stage, are apparently the very same as the first three pairs, in the larva in the second stage, and in the pupa. And in the pupa the first three pairs, which certainly correspond with the first three pairs of cirri in the mature animal, seem to me, for reasons presently to be assigned, to be the second, third, and fourth thoracic limbs. Hence I am led to the conclusion that the first pair of legs in the larva in the first stage, are homologically the second thoracic (answering to the third pair of maxillipeds in the higher Crustaceans), and that the two succeeding pairs are the third and fourth thoracic limbs; to be succeeded, in the pupal stage, by the fifth, sixth, and seventh thoracic appendages.
[54] Mr. Dana, moreover, has remarked, (‘Crustacea: United States Exploring Expedition,’ p. 1386), “that he knows of no instance of a mandible becoming so completely a leg, as to lose wholly the mandibular function even of its basal portion.”
[55] ‘Annales des Sciences Naturelles,’ 2d series, tom. xix, 1843, p. 34. M. Joly’s observations were made on the Caridina. I owe to the great kindness of Mr. C. Spence Bate, an examination of some larvæ of the allied genus Hippolyte varians, and I found, on dissection, the view of M. Joly, that the three pairs of natatory legs are the maxillipeds, so far strongly confirmed, that they followed closely, with equal intervals, the mandibles and two pairs of maxillæ. The first pair of natatory legs in Caradina, in its earliest condition within the egg, is uniramous, like the first pair in the larvæ of Cirripedes. There is one fact which seems rather strongly opposed to the view of these three pairs of legs in the larvæ of the macrourous Crustaceans being the maxillipeds, which is that Capt. Du Cane (‘Annals of Nat. Hist.,’ 1838, vol. ii, pl. 6, and 7) observed only three pairs of limbs in process of formation posteriorly to the first three pairs, whereas there should be found, in accordance with M. Joly’s view, five pairs, i. e. all five pairs of ambulatory legs. This one fact countenances the view, which I hold on the nature of the legs in the larvæ of Cirripedes during their early stages, namely, that they are the second, third, and fourth thoracic limbs, to be succeeded by only three additional pairs.
Lastly, behind the natatory legs, on the ventral surface, (Pl. 29, figs. 8, 9, i), the body is much produced, and terminates in a horny fork, which, after the first moult (fig. 10, i), becomes much elongated. Anteriorly to this fork, on the ventral surface, there is another fork (l), and again above this I could distinguish, in Chthamalus stellatus, after the first moult, another fork (m), or at least a pair of short thick spines. From the structure of the forked abdomen in the known larvæ of the Podophthalmia, I presume that this portion of the body is the abdomen of the young Cirripede, but it is not at all plainly articulated. After the first moult, the posterior end of the carapace (h), which is always pointed, becomes much elongated and serrated on both sides;[56] reminding one of the structure of the carapace of the so-called[Pg 109] Zoea, or larva of certain Podophthalmia. Situated under this posterior prolongation of the carapace, there is a swelling (n, with long hairs on both sides), which apparently lies on the dorsal surface of the spinose and forked abdomen; here, when the larva is compressed, the cellular and oily contents of the body burst forth; and I suspect that this swelling is the anus, for it is known from the researches of Rathke,[57] that the anus in the higher Crustaceans opens during the earliest periods dorsally.
[56] I suspect that the account given by Goodsir (‘Edinburgh New Phil. Journal,’ 1848) of the posterior points of the carapace and abdomen in the larva of a Balanus, is not quite accurate.
[57] ‘Annales des Scienc. Nat.,’ tom. xx, p. 451.
I have given, from Burmeister,[58] a lateral view (Pl. 30, fig. 1) of the one single specimen, ever observed of a larva in this stage, belonging, as is supposed, to the genus Lepas. The carapace has now greatly altered its character. The two fleshy projections, as so called by Burmeister, by which the larva adhered to the sea-weed, were supposed by this author to include the great prehensile antennæ of the pupa; from my observations, already alluded to, on the two projections (Pl. 24, fig. 17) in the closely analogous egg-like larva, in the second stage, of Cryptophialus, by which it also adheres, I have not the least doubt that this is the case. The small, internal, and anterior pairs of antennæ, are, as it would appear, now aborted. The eye, according to Burmeister, has commenced becoming double; but the two approximate eyes are not as yet compound. The mouth is probosciformed (m), and does not differ much from its condition in the first stage; no gnathites were observed by Burmeister, and they could not be expected to be present, for they are not found even in the pupa. The mouth, which in the larva in the first stage differs in different genera, in being more or less advanced forward, here stands some way anteriorly to the natatory legs, as in the pupal condition. The first pair of legs is uniramous, and the two other pairs biramous; this fact, together with the number of the legs in this second stage being still three, and their structure being not very different, leaves little doubt on my mind that we here have the same three pairs as during the first stage. The abdomen has become much shortened, but still space is left for the development, in the pupa, of the three pos[Pg 110]terior pairs of legs. I may here remark that in the pupa the anterior natatory legs have become, like the others, biramous; but yet, as it were for the purpose of showing their metamorphosis from the uniramous legs of the earlier stages, they have their bristles arranged rather differently from those on the succeeding five pairs of legs.
[58] ‘Beiträge zur Naturgeschichte der Rankenfüsser,’ tab. 1, figs. 3, 4.
I have given a lateral view of the pupa of Lepas australis (Pl. 30, fig. 2), illustrative of the description in my former volume: the specimen is drawn as if transparent, and it was to a certain extent thus rendered by boiling in caustic potash. A sketch of the position of the young Cirripede within the pupa, was made by the camera. At first the drawing will perhaps hardly be comprehended: the darker shaded portion to the left of the letter (b) shows the extent of the sack, with the included thorax and natatory legs of the pupa: to the right of the same letter, if we do not consider the young included Cirripede, the only organs distinguishable in the mass of cellular and oily matter, are the alimentary canal, the cement-glands (t), i. e. the incipient ovaria, and the cement-ducts (t′) which enter the antennæ. A view is also given (fig. 4) of the ventral surface of the pupa; and a transverse section (fig. 7) of the carapace, taken close to the eye-apodemes. On comparison with the larva in the second stage, the changes in external appearance and structure are not very great; the prehensile antennæ are freed from their cases; the two eyes stand further apart; the three posterior pairs of legs have been developed, and a small abdomen has become distinctly separated from the thorax. Before proceeding to make a few additional remarks and corrections to my former description of the pupa, it will be advisable, on account of the importance of the subject, to discuss the homologies of the limbs.
From the presence of eyes and of two pairs of antennæ in the larva, during its earlier stages, the front of the head consists, in accordance with all analogy, of three segments; the mouth, likewise, from being formed of three gnathites (which can be detected by dissection in the pupal state), consists, also in accordance with all analogy, of three segments, making altogether six segments—on the[Pg 111] nature of which I apprehend no objection will be raised. In two out of the three orders into which Cirripedes may be divided, the mouth is succeeded, in the adult animal, by eleven most distinct segments; of which the first (i. e. the seventh cephalic) differs from the succeeding seven thoracic segments; and these seven again differ from the three abdominal and terminal segments. Hence it must be admitted that, as far as the cephalo-thorax of the archetype Cirripede is concerned, it consists, like that of the archetype Crustacean, of fourteen segments, of which eight succeed the first-named six that form the mouth and front of the head; and that, with the three abdominal segments, there are altogether seventeen segments. In the order Thoracica, however, which includes all common Cirripedes, both in the pupa and in the mature animal, only six thoracic segments with their appendages, succeed the mouth, two having been lost; and the question arises which are these two, whether the seventh and eighth, or the thirteenth and fourteenth (i. e. the two terminal thoracic) segments; for there is no reason to suspect any other segments of having disappeared. In my former volume, I inferred, without sufficiently entering into my reasons, that it was the seventh and eighth, i. e. the last cephalic and first thoracic segments, which had disappeared; but I now find that Mr. Dana[59] believes that, in ordinary Crustaceans, the abortion of the segments with their appendages almost always takes place at the posterior end of the cephalo-thorax. Nevertheless, after due deliberation and fresh examination of the pupa, I must retain my former opinion, that it is the last cephalic and first thoracic segments which have either coalesced with the others, or wholly disappeared. In the pupa, the mouth, although functionless, has its place most plainly marked by being slightly prominent, and by the presence of a sort of labrum and of a shrivelled œsophagus, round which latter the gnathites and the new œsophagus of the future young cirripede are in process of formation. Now between the mouth of the pupa and the first pair of natatory legs, there is a space of membrane, equalling, when stretched out, the three succeeding thoracic segments in length and breadth: this interspace, I conceive,[Pg 112] must have some homological signification; here then we have at least an appearance of the abortion of appendages; whereas, at the posterior end of the cephalo-thorax, no such appearance is presented. Moreover this interspace of membrane is divided nearly in the middle by a most conspicuous fold, which, on the view here adopted, would mark the separation of the seventh (cephalic) from the eighth (thoracic) segment; and the interspace and fold are thus simply explained. Lastly, I have shown, in the Introduction (p. 18), that the first and five succeeding pairs of cirri of the mature Cirripede present certain small, but significant, resemblances in structure and in the origin of their nerves, with the outer pair of maxillipeds and with the five pairs of ambulatory legs in the Podophthalmia; which resemblances are all futile, if the cirri belong to the 7th, 8th, 9th, 10th, 11th, and 12th segments of the cephalo-thorax, or those immediately succeeding the mouth; but are full of meaning, if the six pairs of cirri belong, as I believe, to the 9th, 10th, 11th, 12th, 13th, and 14th segments, or the six posterior segments of the cephalo-thorax.
[59] ‘Crustacea: United States Exploring Expedition,’ p. 22.
Before commencing on details, I may premise that I have examined the pupa of Lepas australis, pectinata, fascicularis, and anatifera, of Conchoderma virgata, partially of Dichelaspis Warwickii, of Ibla quadrivalvis, and of Alcippe lampas; and in the Balanidæ, of Balanus balanoides and Hameri. In the pupæ of all these genera there is a most close general agreement in structure, excepting in minute details: I was surprised to find exactly the same slight differences in the spines on the first pair of natatory legs, as compared with the succeeding pairs, in Balanus Hameri, as in Lepas. The abdomen and caudal appendages of the pupa in the abnormal Alcippe, as we shall presently see, offer the only marked exception to this uniformity of character throughout the Thoracica. The outline of the carapace or shell is usually not so blunt at the anterior end, as in the pupa of Lepas australis (Pl. 30, fig. 2); more commonly the shape is that of the pupa of Alcippe (Pl. 23, fig. 16). In Lepas pectinata the two posterior points of the carapace are produced into two short spines. The surface of the carapace in L. australis is lined, as represented in fig. 4:[Pg 113] the colour of this species when alive was blue:[60] in L. fascicularis the surface is punctured: in L. pectinata it is marked with curious points of various shapes, often star-shaped, in parts reticulated, and confluent along the dorsal margin, and in parts lined: in B. balanoides it is very obscurely punctured, and in B. Hameri the punctures pass into lines. The whole of what is externally visible consists of the carapace, for this is produced not only backwards, so as to enclose the thorax and abdomen with their appendages, but also forwards, so as to overhang the whole front of the animal; and the prehensile antennæ, in Lepas, Ibla, Balanus, and probably in all the genera, can be retracted within its lower edge. The protection afforded by the carapace to the antennæ is aided by two crests (Pl. 30, fig. 7, c) parallel to this lower edge. The whole sternal surface is very narrow (fig. 4), and is likewise protected by the carapace; that is, when the two sides are drawn together by the adductor muscle. The shell, however, when thus drawn together, gapes a little at the two ends, at least in the case of Lepas australis. The adductor muscle, if introduced in fig. 4, would have crossed close anteriorly to the basal margin of the mouth; and in fig. 2, its end on the near side would have been attached under the dark cæca, which enter the upper end of the stomach. The adductor is shaped almost like an hour-glass, and so differs from this muscle in the mature Lepas, in which it is of the same thickness throughout. I may here add that the pupa of Lepas australis could swim very rapidly, and often on one side in a circle; it could walk by the aid of its antennæ, but often fell over; being thus locomotive, and, as we shall immediately see, well provided with senses, it cannot be considered as very lowly organised.
[60] I took this species alive in the Southern Atlantic Ocean; and, mistaking it for an independent Crustacean, was much perplexed where to class it. I had overlooked these specimens when publishing my former volume.
Acoustic Organs.—Commencing at the anterior end, two small elongated orifices, 10/6000th of an inch in diameter, (e, fig. 4, Pl. 30), may be seen; these lead, as described in my former volume, into a sack, with a bag suspended in it, which is provided with a large nerve, and which I believe[Pg 114] to be the acoustic vesicle. These orifices occur in the carapace, either in the same position, or a little more posteriorly, in the pupæ of all Cirripedes. In Balanus balanoides they are minute, being only 2/6000th in diameter, but are surrounded with a border: in Conchoderma virgata they are also surrounded by a border: in Lepas pectinata, the orifices are 3/6000th of an inch in diameter, and are very singular from being seated on rounded prominences, causing the carapace to have two short, blunt horns in front. In Lepas australis, and I believe in the other species, the corium round the acoustic orifices is darkly coloured; and these coloured marks can be distinguished for some little time on the peduncle of the young Cirripede, after the metamorphosis, and after the entire organ, together with the whole pupal carapace and eyes, has been moulted. Knowing the connection in the higher Crustacea, of the acoustic organs and the antennæ, and seeing the very backward position (figs. 2 and 4) of the one great pair of antennæ, I have always imagined that these orifices probably marked the normal position of the anterior pair of antennæ, which, since the earlier larval stages, have disappeared. And I now find[61] that Schödler affirms, that in most, if not in all Daphnidæ, there is a black spot in front of the eye, which is connected with an opening in the basal portion of the anterior antennæ, and he concludes that it is an organ of hearing.
[61] Quoted by Dana, ‘Crustacea of United States Exploring Expedition,’ p. 1264.
Antennæ.—These, from their present position, and from standing, in their earlier stages whilst within their envelopes or horns, exteriorly to the small medial pair (since aborted), I believe to be the second pair; and this is Mr. Dana’s opinion. In my former description of these very singular and important organs (Pl. 30, figs. 4 and 8), I have fallen into some considerable mistakes: the two plates or segments (fig. 4, N), of which the posterior margins are inflected as apodemes (n), carrying the eyes, are certainly, as may be clearly seen in the pupa of Alcippe, Pl. 23, fig. 16, and as affirmed by Burmeister,[62] the basal[Pg 115] segments of the antennæ. The second or main segment (formerly called by me the basal segment) has in some species an upper portion of the membrane of which it is composed, next to the body, excessively thin, and separated from the rest of the membrane composing the segment, by an oblique line (fig. 8, o), which I mistook for its articulation with the body.[63] We then come to the disc or third segment; and lastly to the fourth, or ultimate segment. This ultimate segment, generally, has its external corner projecting up, as a step; and this sometimes, as in Dichelaspis Warwickii, gives the appearance of its consisting of two segments; but a careful examination of this part in Ibla, in which the step-like structure is carried to an extreme, makes me believe that there is only one segment.[64] The prehensile antennæ, therefore, like the natatory legs, are formed of four consecutive segments, of which the basal segments give rise to the singular apodemes, presently to be noticed (fig. 7), that carry the great compound eyes. This basal segment, in all Cirripedes, is moulted with the eyes, the three other segments invariably remaining cemented to the surface of attachment.
[62] ‘Beiträge zur Naturgeschichte der Rankenfüsser,’ p. 19. In tab. 1 of this work there are good drawings of the general structure of the pupa of a species of Lepas, probably L. australis. I believe this author was the first who made out the structure of the abdomen of the pupa.
[63] In the table of measurements of the antennæ of the several genera and species of the Lepadidæ (p. 286) of my former volume, the articulation, called by me basal, I now know to be really the articulation between the basal and second segment. In the fourth column, headed “Length from end of the disc to the inner margin of the basal articulation,” the term inner margin really applies to the oblique curved line separating the thin and scarcely visible membrane from the thicker membrane of that segment. These corrections do not in the least affect the object for which the table was given.
[64] In a sketch, sent me by Mr. Dana, of this organ in the pupa of a Lepas from the Antarctic Ocean, I observe that he divides my ultimate segment into two segments.
In the Southern Atlantic I took some specimens of the pupa of Lepas australis, not yet attached, and therefore with the muscles of the antennæ, not having suffered any of that absorption, which they undergo, as soon as the pupa is permanently cemented to some floating object. In my former volume I noticed a pair of strong muscles, attached to the tips of the middle forks (Pl. 30, fig. 7) of the apodemes, and I now find two pairs attached to the bases of the two outer forks, and directed dorso-anteriorly; and two other[Pg 116] pairs, also attached to their bases, but directed dorso-posteriorly, so that altogether there are five pairs of muscles attached to the apodemes; their chief function, I should think, was to draw the antennæ posteriorly and upwards within the carapace; but as the apodemes cannot be moved without the great compound eyes being likewise moved, the muscles probably serve a double purpose. When the pupæ were alive, I noticed that their eyes were constantly kept in a state of vibratory movement. Flexor and extensor muscles are attached at one end to the posterior margin of the basal segment, and at the other end to the second or main segment; other powerful muscles attached to this latter segment, are prolonged by ligaments into the disc. In Cryptophialus I observed that the disc-segment had a movement almost like that of the wrist. Whether any muscles enter the small terminal segment, I know not.
The drawing in Pl. 30, fig. 8, of part of the second segment, of the third or disc segment, and of the fourth or ultimate segment, in Lepas australis, is, I think, very accurate. The second segment articulates on the upper or dorsal surface of the disc, and has the articulation on one side constricted and formed of thin and flexible membrane; the little terminal segment, which is turned outwards at right angles, also, articulates on the disc. That the disc forms a true segment is shown clearly in Cryptophialus (Pl. 24, fig. 18), where the articulation with the second segment is not in such close contact. The disc is either circular, as in Lepas, or hoof-shaped, as in Ibla: in B. balanoides the disc is rather hollowed out on the inner side. It has the power of adhering even to so smooth a substance as glass, placed vertically. It is surrounded by a rim of transparent membrane. On the hinder margin some spines arise from the central and more opaque part: in Lepas australis, there are no less than seven of these spines (fig. 8): in Conchoderma virgata there are only four, in Scalpellum and Ibla only one. When the disc is placed on the surface of attachment, these spines lie parallel to it. The middle part of the disc is, almost always, nearly opaque; and in tracing the cement-ducts from within the body of the pupa, or of the young Cirripede, I in many cases traced them as far as this[Pg 117] point, but here lost them. From this same obscure central part of the disc, in most, if not in all species of the Lepadidæ, spokes radiate, which sometimes are branched, and are not regular, not always even resembling each other on the opposite sides of the same individual. Round the proper membranous border of the disc, a second one may be observed (fig. 8, p), which differs in shape and extent in different specimens: under favorable circumstances, and very high powers, it may be seen to have a reticulated structure, and to be of a very pale brown colour; towards the exterior margin, the reticulations become finer, and are blended together and lost; on the inner margin, the substance forming this membrane may be seen to come out of the spokes. This substance is the cement, which has the power of adhering to whatever substance it grows against; and thus the disc of each antenna becomes cemented down, and soon both the antennæ are surrounded by a common border of cement, which gradually increases, after the metamorphosis, in extent. Occasionally the cement forms little projections, like short spines, on the edges opposite to the orifices of the spokes.
The small terminal segment usually bears on its truncated extremity six spines, some of which are occasionally hooked; in Scalpellum, two spines, rather longer than the others, are borne on a step some way down on the inner side of this segment; but in Lepas, two spines (fig. 8), very much longer than the others, arise from the outer corner of the extremity. These two are very different from the other four borne by this segment, or indeed any other spines on the body; for they are quite flexible, and are furnished with a double row of very long, straight, excessively fine hairs, which seem to be articulated on them—the whole presenting a very beautiful appearance. These spines are of considerable length, and in Conchoderma virgata they even equal in length the whole antenna. I can hardly doubt that these beautiful, plumose, flexible spines, into the thick bases of which the coloured corium could sometimes be seen to enter, serve as feelers. Owing to the facts immediately to be mentioned, I erroneously stated, in my former volume, that there were three long spines.
In three species of Lepas, in Dichelaspis Warwickii, and in Scalpellum Peronii, after having torn the lately-cemented antennæ from the surface of attachment, I observed proceeding from the end of the terminal segment, from between the above two groups of spines, what appeared to be a long narrow ribbon with its end torn off; and which, in the case of Lepas australis, I fancied was one of the plumose, long, flexible spines ripped open. But now that I have examined some of the pupæ of this species before their attachment, I find (as represented in Pl. 30, fig. 8, v′) a flattened tube, ending in a blunt point, and having a peculiar ringed structure. I have noticed similar appendages to the antennæ of specimens just attached of Lepas anatifera. In the Dichelaspis and the Scalpellum, the tube was very long, and seemed, from its torn appearance, to have been firmly attached to the supporting surface. In both these cases, the tube came out from within another slightly larger tube, which had been broken off close to the extremity of the terminal segment of the antenna. In the case of the Lepas anatifera, the tube expanded a little after leaving the antenna. In the Dichelaspis, it had exactly the same diameter as the cement-duct, which could be clearly distinguished within the two lower segments. From these several facts, and from the peculiar appearance of the tube itself, I believe it to be a tube of cement-tissue which thus, sometimes even before the pupa is attached, independently grows outwards. That the cement-tissue can grow outwards and assume definite forms, we know from the singular case of Lepas fascicularis, in which the cement proceeding from several apertures, forms a vesicular float round the peduncle of not only a single individual, but often of a group of specimens: we shall presently find a somewhat analogous fact in the case of Coronula. It is possible that this tube, proceeding from the extremity of the antenna, may be the channel through which cement continues to be poured forth during the continued growth of the above Cirripedes; and the manner in which this is effected, considering how firmly the end of the peduncle is cemented down, has always appeared to me a difficulty. In those pupæ of Lepas australis, which I caught swimming about unattached, it is[Pg 119] surprising that the disc should have been edged with cement, and that a tube, similarly formed, should have grown out of the ultimate segment: it shows, I presume, that the cement-tissue will grow out, whether or no the pupa has succeeded in finding a proper object for attachment. Lastly, I have felt some surprise, in two or three instances in observing some dark purple pigment-cells, like those in the corium, within the terminal tube of cement; and likewise within the spokes of cement in the disc: this is the only fact which causes me the least doubt, whether I have rightly determined the nature of the terminal tube, as wholly formed of cement tissue; or whether it may not be covered by an outer integument, itself lined by true corium, coloured purple.
Finally, I may add, that, excepting in small details, the prehensile antennæ present no difference throughout the Order: I have minutely examined them in several genera of the Lepadidæ; and in the Balanidæ, I have seen them in Coronula, and in several species of Balanus. In B. balanoides I have examined them carefully; they are smaller and thicker than in Lepas, with the second or main segment bowed outwards, carrying its usual single spine; with the disc excised on its inner margin and apparently without the spoke-like vessels for the cement; and with the ultimate segment proportionably longer, and carrying, I believe, six spines, of which two appeared to be longer and more flexible than the other four shorter and somewhat hooked spines. In Coronula balænaris, also, the terminal segment is, proportionably to the others, of large size. Not only throughout the order, but throughout the whole Class, the antennæ are singularly uniform in structure, as will be seen, when the last two orders are described.
Eyes.—These present no difference, except in size, throughout the class; and have been sufficiently described in my former volume. The true basal segments of the antennæ (incorrectly designated formerly as sternal plates or segments) are separated from each other by a deep fold; and are separated from the edges of the carapace on each side by a crest and slight fold (Pl. 30, fig. 7, c; and 4); these folds and crests die out posteriorly, and disappear. The hinder, rounded margins of the basal segments are[Pg 120] inflected inwards, and their corners are produced far up into the body, thus forming the curious UU-like apodemes. These apodemes exist throughout the whole class; and the outer arms always carry the great compound eyes. I noticed, in Lepas pectinata, that the two middle arms are proportionably longer than in L. australis. Owing to the presence of these apodemes, and to certain coloured marks on the adjoining corium, the eyes, though enclosed fairly within the carapace, yet deceptively appear pedunculated, so that even J. Vaughan Thompson was thus deceived. I have already described the several muscles attached to these apodemes, and the constant vibratory movement of the eyes. Whilst the pupa remains a freely swimming animal, the eyes are included, not only within the shell or carapace, but (as would naturally happen) within the corium or true skin lining the carapace; but after the pupa has become attached, preparatory to its final metamorphosis (in the state represented at Pl. 30, fig. 2.), not only are the muscles, as before remarked, which are attached to the apodemes, absorbed, but so is the corium investing the apodemes and the immediately adjoining parts of the carapace. Hence it comes that the new corium of the young Cirripede within, is formed in a deep transverse fold across the whole lower half of the animal, and the apodemes with the eyes are thus, as it were, rejected from within the corium, though still remaining within the carapace. Consequently in this final stage, the eyes and apodemes are very conspicuous from the outside, being seen only through the transparent carapace. I presume that the eyes at this period have become functionless, with the optic nerve divided and absorbed. The eyes, apodemes, and carapace soon afterwards are all moulted together.
The eyes of Cirripedes certainly undergo a remarkable series of changes: in the larvæ in the first stage, there is a single eye, perhaps formed by the confluence of two eyes, occupying the normal position in the front of the head: in the second stage, according to Burmeister, the eye has become double, but the two are as yet simple; they are now situated posteriorly to the second pair of antennæ: in the third or pupal stage, they remain in the same situation, but have[Pg 121] become compound, of great size, and are attached to the apodemes of the antennæ: in the mature and fourth stage, they have moved someway posteriorly, and again have become simple, of minute size, and are either confluent, as in the Lepadidæ, or tolerably far apart, as in the Balanidæ. It must not be supposed that the eye of the mature Cirripede is metamorphosed from the eye of the pupa, for such is not the case; the new eyes and old eyes being formed someway apart, and frequently both can be seen within the pupa (as in Alcippe, Pl. 23, fig. 16) at the same time. It is scarcely possible that the eye of the larva in the first stage, can be changed into the double eyes of the second stage; though these latter may possibly be multiplied into the eyes of the pupa, as both continue to occupy nearly the same position.[65]
[65] Zenker, in his ‘Physiological Remarks on the Daphnidæ,’ (‘Journal of the Microscopical Society,’ 1853, p. 274), speaks of a “tripartite azygous eye” as common amongst Crustacea, and as occurring “in conjunction with the aggregated eyes in Artemia, Argulus, &c.; but as appearing regularly in all the Branchiopoda and Siphonostomata as the earliest visual organ.” Hence I conclude that this azygous eye is the homologue of that single eye which appears in the earliest larval stage of Cirripedes; and that the compound eyes of the cirripedial pupa, answer to the aggregated eyes of Artemia and Argulus, &c., with the difference, that in these latter genera the single eye is retained. See, also, Von Siebold, ‘Anatomie Comparée,’ tom. i, p. 435.
Mouth, thorax, limbs, abdomen.—I have nothing to add regarding the mouth, except to confirm my former account; viz., that it is functionless, consisting merely of crests, which project inwardly between the gnathites of the young Cirripede, and of a shrivelled closed tube representing the œsophagus. In fact the mouth is a model of the outside of the mouth of the young Cirripede. I may remark that some little way beneath the membrane answering to the labrum, a pair of ligamentous apodemes, the use of which I do not know, slightly penetrate the body. The degree of prominence of the mouth varies, but it is far less than in the mature animal. On the limbs I have nothing particular to add: the drawing of the first pair of legs (Pl. 30, fig. 5) is, I think, very accurate: I observed all the spines here figured, on the corresponding leg of the pupa of Balanus Hameri. The five posterior pairs of legs differ only in the outer ramus having five plumose spines, instead of four, and one short simple spine at the exterior angle, making six alto[Pg 122]gether. The legs, in their natural position (fig. 2), have only the terminal segments of their two rami directed posteriorly; and as a consequence the spine (close to i in fig. 5), borne on the penultimate segment of the outer ramus, is directed in the same line with that segment and with the pedicel, namely, anteriorly, and at right angles to the natatory plumose spines. This short spine acts, I imagine, as a defensive weapon; it has been omitted in fig. 2. Of the thorax I need not give, from my notes, any more details. The abdomen (fig. 6) is similarly constructed, as far as I have seen, throughout the order, with the exception of Alcippe (Pl. 23, fig. 17), in which it is composed of only one segment instead of three. In this genus the caudal appendages likewise consist of only one segment, with very short spines. In the pupa of Balanus balanoides, the three spines borne on each caudal appendage are very much more unequal in size than in the pupa of Lepas australis, although in the latter (fig. 6) the inner spine is considerably thicker than the two outer. Whether the three segments of which the abdomen is composed, are the three anterior or three posterior, of the normal seven segments, I know not: on the view that they are the three posterior segments, I presume, according to analogy, that the caudal appendages are borne on the penultimate segment, and that the ultimate segment is here quite aborted.
On the internal viscera I have nothing to add. The cement-duct is represented in Pl. 30, fig. 2, t′, on the near side, running into the antennæ; and I repeatedly traced it, for the duct is very strong, as far as the disc segment; at the other end it joins the cement-gland (t) on the same side of the body. This cement-gland is proved, by the clearest series of facts, to be converted into the incipient ovaria and ovarian cæca. The cement-glands in the older pupæ could be traced as far as the cæca of the stomach, exactly where the ovaria lie in the mature animal; but in some young pupæ, they extended further posteriorly, past the mouth, between the outer and inner membranes of the overlapping carapace. I have faintly shown the course of the stomach, with its two cæca at the upper end; the anus lies between the caudal appendages, at the extremity (above b′) of the abdomen. At[Pg 123] this age there is no trace of the vesiculæ seminales, so conspicuous in the mature Cirripede.
Young Cirripede, whilst within the pupa.—I repeatedly succeeded in dissecting the young Lepas australis out of the pupa; and by the previous action of boiling potash, and by a strong light, I was enabled to make a camera sketch (Pl. 30, fig. 2) of the relative positions of their several parts. The young Cirripede is drawn very faintly, and is best seen by holding the plate in the same position with the mature animal, of which a section is given in my volume on the Lepadidæ, Pl. 9, fig. 4. I may just notice how complicated are the membranes in a longitudinal section taken at this period: we have, 1st, beginning at the back, the horny tissue of the carapace or bivalve shell of the pupa; 2d, the primordial valve (z, in fig. 3) of the young Cirripede; 3d and 4th, two folds of corium; 5th, the membrane of the sack of the Cirripede; 6th, the membrane of the sack of the pupa; 7th, the outer tunic of the thorax of the pupa; 8th, the outer tunic of the thorax of the young Cirripede; 9th, the corium lining the latter membrane; and these nine membranes would be repeated on the opposite side of the section, if it were taken through either side of the shell or carapace, bordering the orifice.
After the exuviation of the outer membranes of the pupa, certain pre-existing coloured marks in the corium, such as those round the eyes and round the acoustic orifices, along the ridge of the back and on the borders of the orifice, &c., are still retained by the young Cirripede, either temporarily or permanently; so that the correspondence of part with part of the external surface admits of no doubt. Moreover, the three terminal segments of the antennæ are invariably retained by the young Cirripede, though in a functionless condition, and into them the outer membrane of the body, and an important organ, viz., the cement-ducts are still prolonged; hence these prolongations must be considered as aborted antennæ. Again, we have seen that the mouth of the young Cirripede is formed under the rudimentary mouth of the pupa, with the new œsophagus, round the old œsophagus, leading into the same alimentary canal. The twenty-four extreme tips, likewise, of the six pairs of biramous[Pg 124] cirri of the Cirripede are formed within the twenty-four extremities of the six pairs of biramous, natatory legs of the pupa. Consequently, in the Cirripede and pupa, thus far, part corresponds with part, notwithstanding that new eyes are formed posteriorly to the old eyes, and new acoustic organs in a quite different position from the old ones; but now we come to a most important diversity in the metamorphosis, or rather to follow Professor Owen,[66] in the metagenesis, of the young Cirripede. Although, as just stated, the extremities of the cirri are formed within the legs of the pupa, yet, from the great length of the cirri, they occupy more than the whole of the thorax of the pupa; so that the thorax of the young Cirripede is not formed within the pre-existing thorax of the pupa, but within that part of the pupa, (homologically a portion of the first three cephalic segments), which lies anteriorly to the thorax. As a consequence of this, the pedicels and lower portions of the cirri, the segments of the thorax and its dorsal surface, all come to occupy a position at nearly right angles to that of the corresponding parts in the pupa: this is shown in Pl. 30, fig. 2. And as a further consequence, (and this is the more important point), the sack, which both in the young Cirripede and pupa is formed by the overhanging and produced portion of the carapace, and which is internally lined by a reduplication of the membrane of the thorax, is necessarily, owing to the changed position of the thorax, altered in extent and carried much further; namely, from extending merely parallel to the longitudinal axis of the pupa (from b to b′), it is now in the young Cirripede, in addition, carried (to s′) almost quite across the inside of the animal. Hence it comes that the young Cirripede is, as I have said in my former volume, internally almost intersected; and its body remains attached only by a small space, (see the broken line, round a and b in Pl. 25, fig. 1, of a Balanus with the shell, &c., removed from one side), to the sternal or ventral, inner surface of the carapace,—the carapace being modified either into the capitulum and peduncle, or into the shell with its operculum and basis. As a still further consequence of this change of[Pg 125] position of the body of the young Cirripede within the body of the pupa, the alimentary canal becomes shortened to fully half its former length. At the same time, the interspace between the mouth and first pair of legs of the pupa, (consisting of the seventh and eighth segments of the archetype), is quite lost in the Cirripede by coalescence. The final cause of the thorax of the young Cirripede not being developed within the thorax of the pupa, probably is, that the cirri may be formed of considerable length, so as to be immediately enabled to seize prey; and that the thorax, which supports the cirri (and this probably is even more important) should be as free as possible within the sack, so as to aid the captorial action of the cirri.
[66] ‘Parthenogenesis,’ pp. 13 and 26.
After these remarks, more especially with regard to the formation of the sack, if any one will look at the sectional drawing of a pedunculated Cirripede in my former volume, or of a sessile Cirripede (Pl. 25, fig. 1) in this present volume, in which latter the shell adds to the complexity, he will perceive the cause of the extreme difficulty in understanding the relative position of the parts throughout the whole class. Even after I had discovered that the prehensile antennæ of the pupa might always be found in the centre of the basis or surface of attachment, and which fact, it might have been thought, should have convinced me that this was the anterior end of the whole animal, yet still I fancied that the prominent mouth represented the entire head, and that the shell was something quite distinct. It is clear that others have been equally perplexed; for that which is the anterior end in the eyes of one naturalist, is the posterior end in the eyes of another; so with the dorsal and ventral surfaces: one naturalist considers the peduncle of the Lepas as the abdomen; another considers it as a pair of metamorphosed, thoracic limbs, &c.! The probable position of the segments of the body of a mature Cirripede, in relation to the three anterior cephalic segments, or carapace, is shown in the diagram (Pl. 25, fig. 6) of the supposed position of the mature Proteolepas within its pupal envelopes. Here, in the diagram, the two segments immediately succeeding the mouth (c), which are the seventh and eighth of the archetype, (for the mouth consists of three segments, and all in front[Pg 126] of the mouth of three other segments), have come to adhere by their dorsal surfaces to the internal surface of the carapace,—that is, of the first three segments, which ought of course to have stood quite in advance of these two segments, and these two segments again ought to have stood in advance of the mouth. The mouth is directed posteriorly, instead of from the body; and the three segments of which it is formed (closed at their anterior end by the labrum), and are very small compared to the relatively monstrously great, three anterior cephalic segments, composing the carapace. To place the segments of the body of Proteolepas in proper sequence, in respect to those of the carapace, and in accordance with the sequence of the archetype Crustacean, it would be necessary, by seizing the extremity of the abdomen (a), to tear the two segments succeeding the mouth from their dorsal attachment, as far back as the basal margin of the labrum; and then pull them till they stood posteriorly to (or in the diagram, above) the mouth; which latter part would, by the same movement, be made to project out at right angles to the ventral surface, and would then be preceded only by the first three, great, confluent segments of the head, which being produced backwards, form the carapace. All that has just been said on the position, in Proteolepas, of the segments of the body in relation to those forming the carapace, I believe to be applicable to all ordinary Cirripedes, with this difference, that in the latter, after the metamorphosis, the two segments succeeding the mouth quite disappear on the ventral surface, and dorsally are either aborted or have coalesced with the adjoining segments.
When the due time for the act of metamorphosis has arrived, the pupal carapace splits along the dorsal ridge, and is cast off, together with the acoustic sacks, the basal segments of the two antennæ, and the great, black, compound eyes, hanging to the UU-like apodemes. The three terminal segments of the antennæ invariably remain cemented to the surface of attachment. The exuviæ usually continue for a time united to the cemented antennæ, but are finally washed away. Besides the split along the dorsal ridge, the[Pg 127] carapace separates, all round the orifice, from the delicate tunic lining the sack and investing the thorax and natatory legs of the pupa; for these membranes are not moulted for some considerable time afterwards. Hence all these inner parts retain for a period the appearance and structure of the natatory pupa, whilst the exterior resembles, in every respect, a fixed and perfect Cirripede.
In my former volume, I have insisted on the important and curious results which ensue from the eye-apodemes penetrating so deeply into the body (see Pl. 30, fig. 7, in which the proportions are more correct than in fig. 2), with the eyes attached exteriorly to their outer arms; for as these apodemes have to be ejected, the external membrane of the young Cirripede (Pl. 30, fig. 2) has to be formed in a deep fold or arch over them, and consequently the membrane on the sternal surface is formed considerably longer than on the dorsal surface. From this it follows, when all the membranes are free and are stretched fully out after the moult, that the whole animal, posteriorly to the cemented-down surface, turns vertically up, and assumes its normal position at right angles to the surface of attachment, and to that which it held in its pupal condition; for the pupa always adheres with its sternal surface parallel to the surface of attachment. A young Lepas, which has just moulted its pupal carapace, and has assumed its proper vertical position, with the cemented antennæ and the surface of attachment remaining as before, is shown at fig. 3, but is drawn on a smaller scale than the pupa fig. 2, out of which it may be supposed to have been excluded. In this fig. 3, it may be observed that the natatory legs and caudal appendages of the pupa have not as yet been moulted. The fact of the stretching out, in the young Cirripede, of the fold of membrane, which in pupa, just before the metamorphosis passes over the apodemes and eyes, is well shown by three darkly-coloured bands in the corium, which in the pupa are curled, but after the moult, are stretched straight out on the peduncle of the young Lepas.
The pupa, and consequently the young Cirripede, from being attached at first by the antennæ, does not adhere by the actual anterior extremity, but by the sternal surface near it;[Pg 128] the anterior extremity, however, soon becomes cemented down, and afterwards, in ordinary cases, ceases to grow. In Cryptophialus, however, and in certain genera of the Lepadidæ, as Alcippe, Lithotrya, and Anelasma, the anterior or basal extremity does continue to grow, and is not cemented down, and therefore comes to be prolonged beyond the original point of attachment; in order to allow of this, the surface to which the Cirripede is attached has to yield, apparently simply to the pressure exerted in the case of Anelasma, but in the three other genera, to the rasping action of the roughened surface of the extremity of the peduncle.
When after a period the pupal membranes of the sack, thorax, and natatory legs are moulted, the cirri of the young Cirripede are curled up, and its thorax is raised towards the orifice, and we have the animal in its ordinary position, and perfect with the exception of a few parts to be further developed or modified. For, instead of calcareous valves, we have at this period only the so-called primordial valves, composed of chitine; and in the case of Lepas australis, some minute spines and some coloured marks on the peduncle, which soon disappear. The muscles, which enter the three terminal segments of the antennæ in the pupa, have to be absorbed and converted into ligamentous threads. In Lepas, the labrum has to become bullate; and the cæca have to increase in number round the upper end of the stomach, and their dark colour and that of the whole alimentary canal has to disappear or be much weakened. The filamentary appendages at the bases of the cirri, which generally contain some of the testes, have to be developed. The probosciformed penis, which at first equals only the pedicels of the posterior cirri in length, and is apparently imperforate, has to increase greatly in length. The testes and vesiculæ seminales have to be formed. And lastly, and this is a more important point, the two gut-formed cement-glands (or incipient ovaria, t, fig. 2, Pl. 30) which, at the period of the moulting of the carapace and eye-apodemes, and when the whole animal was upturned, came to occupy, together with the cement-ducts (t′), their normal position, i. e. nearly parallel to the sternal surface, now undergo further changes. Their upper and posterior ends lying near[Pg 129] the cæca of the stomach, increase in size, but retain nearly the same character, and thus form the two true ovaria; their middle parts become emptied of their cellular contents, and are converted into the two simple ovarian tubes; and their lower ends branch out, inosculate, and form the inextricable mass of ovarian tubes and cæca. The points of junction on each side between the two cement-ducts and the newly branched ovarian tubes, become now developed into the two cement-glands. The cement-ducts, which continue throughout life growing, either still enter the old antennæ and there pour out the cement-tissue, or they pour it out through special orifices formed for this purpose in the lower part of the peduncle. The changes, supervening during the metamorphosis, in the ovaria and in the cementing apparatus, here described, I have no doubt are general throughout the Order.
I have above alluded to the primordial valves; these are beautiful objects when viewed under a high power: they are composed of chitine without a trace of calcareous matter, but prefigure in shape, size, and direction of growth, the shelly valves soon to be formed under and round them. They are composed of an outer membrane, with its margins separated by yellow thickened rims from the membrane uniting the several primordial valves together; and this outer membrane is underlaid by a single layer of generally hexagonal, thickish cells (Pl. 30, fig. 3 a), varying from 1 to 2/6000th of an inch in diameter. These cells seem to contain a nucleus; and they are at first separated from each other by clear interspaces. If a specimen be taken, only a little before the formation of the calcareous valves, one or more layers of membrane, marked by an hexagonal reticulation, can be separated from the lower surface of the main hexagonal network. It is a singular fact, that in those genera in which there are several valves, the primordial valves occur only on five, namely, on the two scuta, two terga, and the carina; and these are the most persistent valves in the several genera. The other valves are prefigured only by brownish membrane, without the hexagonal tissue. In the mature Lepas, the membrane connecting the several shelly valves is not moulted, but dis[Pg 130]integrates; in the primordial valves, however, which stand far separate from each other, this membrane is moulted; and immediately after the first moult, the first layer of shell appears under and a little way beyond each primordial valve; shelly matter likewise appears, at least in some cases, between the cells of the hexagonal tissue. The young shelly valves are connected together, at each successive moult, by narrower strips of membrane, till, in the case of Lepas, the valves when mature come to touch each other (Lepadidæ, Pl. 1, fig. 5). The primordial valves are often preserved for a long time on the umbones, or centres of growth of the five valves, on which they occur, in the same manner as the larva-shell is sometimes preserved on the apex of certain spiral molluscs. Had not Cirripedes gone through so many and such complicated metamorphoses, this last state, when furnished only with primordial valves and with several internal organs only partially or not at all developed, would have deserved to have ranked as a special stage, and not as merely subordinate to the last or pupal condition.
In the Balanidæ, or sessile Cirripedes, the young animal, immediately after the metamorphosis, or still better if dissected out of the pupal carapace, as I succeeded in doing with Balanus balanoides, may be said to be pedunculated; for it is attached by a little disc of cement closely surrounding the antennæ, the rest of the membranous basis forming an almost semi-globular, flexible peduncle. The valves, at this the earliest period, are all membranous, and do not overlap each other. In the Balaninæ they do not present the peculiar structure of the primordial valves of the Lepadidæ; but in the Chthamalinæ, in Chthamalus, I saw traces of this structure. Calcareous valves are soon formed under the membranous valves. The opercular valves, at this early period, are much larger than the valves or compartments of the shell, which are only four in number, for the carino-lateral compartments are not yet formed. The compartments from the first are surprisingly strong, and have their alæ already formed and overlapped by the adjoining compartments; but of the radii there is as yet no trace. The four compartments form a narrow but nearly circular hoop, which, from its relatively large diameter,[Pg 131] tends to draw down the upper or posterior end of the animal, now forming the opercular valves; and as the basis soon becomes throughout cemented to the surface of attachment, the young Cirripede is much depressed. Soon the opercular valves are drawn a little way down within the shell, becoming attached to the sheath, instead of, as at first, to the very summits of the compartments. In regard to the changes which take place in the shell, in the number of the segments in the cirri, and in the number of spines borne on these segments, &c., during the continued growth of the animal, as they are chiefly important for the identification of the species, I will here refer to a discussion on this subject under the head of the Genus Balanus.
In the pupa, the carapace is produced, not only posteriorly, but anteriorly, so as to cover the entire animal, with the exception of a narrow sternal surface (Pl. 30, fig. 4): in front it is notched, where the sternal surface terminates, and from this notch a faint line runs along the dorsal surface, separating its tergal elements. In the young Cirripede, after the metamorphosis, there is no trace of this medial dorsal suture, or of the wider sternal surface. Looking at the several genera of the Lepadidæ, the external covering of the whole peduncle and capitulum is so continuous and of so uniform a nature, that I think we must consider the whole as a carapace, of which the sternal borders have become completely confluent; formerly I was inclined to look at the capitulum alone as formed by the carapace, and at the peduncle as being composed of the two or three anterior cephalic segments, cased only by their own integuments. As far as can be discerned, the carapace in the pupa, and consequently in the Cirripede, consists only of the tergal elements of the segments; and this seems likewise to be the case with the carapace of the Podophthalmia. Until lately,[67] Prof. Milne Edwards doubted whether the carapace in the higher Crustaceans (to which I believe the carapace of Cirripedes must be compared) was formed by the backward production of the third segment,[Pg 132] which bears the second pair of antennæ, or of the fourth, i. e. the mandibular segment; but from the distribution of the nerves, he now argues that it must mainly belong to the third segment. In Cirripedes, the course of the nerves leads to the same conclusion; for the whole shell, sack, and peduncle are supplied with nerves proceeding from the compounded ganglion, which belongs to the second and third cephalic segments.[68]
[67] Compare ‘Histoire Naturelle des Crustacés,’ tom. i, p. 27, with ‘Annales des Sciences Nat.,’ 3d series, tom. xvi, 1851, p. 233.
[68] This conclusion is supported by the structure of Proteolepas: in this Cirripede there is not a vestige of a carapace, and as the whole of the animal in front of the mouth is almost utterly aborted, being reduced to a mere covering to the two cement-ducts, and as, on the other hand, the mouth with the mandibles, though peculiarly modified, is not at all aborted, there is a strong probability, that the abortion of the carapace is connected with the aborted state of the three anterior cephalic segments; and that the carapace in its origin is not any way related to the fourth or mandibular segment.
The whole of the head in front of the mouth, together with the carapace, is, as we know, formed of three segments; and each of these segments, homologically, ought to consist of eight elements; I recall to mind these facts, inasmuch as the transverse separation between the peduncle and capitulum in the Lepadidæ, and between the basis, the shell, and the opercular valves in the Balanidæ, might be thought to be connected with the separation of the three cephalic segments. So again, as in the Balanidæ the shell normally consists of eight compartments, these might be thought to be related to the eight elements of one or other of the three segments. But I see no reason for admitting this view; and in the case of the carina and rostrum, two of the most persistent and important of the compartments, they exactly cover the sutures which ought to separate the two tergal and two sternal elements of the segment. The valves, moreover, often form many more whorls than three, or the number of the true cephalic segments in front of the mouth; and in each whorl the valves tend to stand in tile-like or alternate order, with respect to those in the whorls both above and below, which would not be the case, if they were the eight elements of the segments.
For the true homologies of the sclerodermic plates, or Shelly valves, with which the external covering of Cirripedes is so generally strengthened, we must, I believe, look to the carapace of the Podophthalmia. In these latter, we[Pg 133] find the carapace composed of sclerodermic plates, which, though closely joined and only occasionally separated by sutures, yet in their origin are distinct;[69] they tend, also, to be arranged in alternate or tile-like order. As the animal grows, the old sclerodermic plates, all joined together, are moulted, and new ones, also all joined together, of a larger size, are formed beneath. Now let us imagine the growth to be more gradual but yet periodical, and the new and larger sclerodermic plates, when formed under the old ones, to adhere firmly to them; the older plates would thus be prevented from becoming confluent, and instead of being all moulted together, as is now the case, they would be almost continually separated from each other, owing to the almost continuous increase in size of the new underlying plates. Consequently lines of splitting would run between the several plates, however numerous they might be, instead of there being, as now, a single line of splitting extending down the back. In fact, we should have the identical manner of growth of the shell or carapace, which occurs in Cirripedes. It is on this ground, and from the several points of homological resemblance incidentally mentioned in the last few paragraphs, that, in the early part of this Introduction (p. 13), when discussing the whole class, I stated that I believed that the carapace of Cirripedes presented more real resemblance with the carapaces of the Podophthalmia, or higher Crustacea, than with those of the lower Crustacea, though in mere shape they more nearly resembled the latter.
[69] ‘Annales des Sciences Naturelles,’ 3d series, tom. xvi, pp. 233, 236, 237.
I have already (p. 128) given an account of the manner in which, in the pupa of Lepas, the cement-tissue escapes from the prehensile antennæ, and of the structure of the cement-ducts, and of the cement-glands or incipient ovaria; and likewise of the changes by which these organs assume their ultimate form in the mature Cirripede. In my former volume, on the Lepadidæ, I described the cement-glands and the cement-tissue in several genera,[Pg 134] and I have there shown (singular as the fact is) that the two cement-glands, with their contents, actually consist of ovarian tubes with their contents (for there seemed to be a relation in the state of fulness in both) in a modified condition. In the Balanidæ, I am not able, from the difficulty of the dissection, to confirm these conclusions, excepting in so far that the tubes on which the cement-glands are formed, run into the mass of ovarian cæca; but, I may add, that in the abnormal Proteolepas, belonging to another Order (see the section, Pl. 24, fig. 1), nothing could be plainer than that the membrane of the ovarian sack (b) formed the cement-ducts, and that their cellular contents, which within the sack (a) were in process of conversion into ova, within the ducts were converted into the cement-tissue. This cement, by some unknown power, travels down the ducts, and debouches at the antennæ.
In the Lepadidæ, there are only two cement-glands, which are situated high up in the midst of the ovarian cæca, with one cement-duct proceeding from each: both the glands and ducts increase in size with the age of the animal:[70] the cement issues either permanently from the prehensile antenna, or, after a short period, through apertures in the peduncle, arranged irregularly or in straight lines,—the last formed apertures being furthest from the central and basal point of the peduncle. In the Balaninæ, on the other hand, at each period of growth, a pair of new cement-glands is developed, larger than those last formed, and making, with the older glands, a chain, connected together by what I have called the cement-trunk. The cement-trunk consists of a tube, which generally becomes enlarged just before entering each gland. The glands, the cement-trunk, and cement-ducts, all adhere to the basal membrane or basal shelly plate. Each gland gives rise to two cement-ducts, these often bifurcate, and sometimes repeatedly bifurcate and inosculate before pouring out their contents round the circumference of the basis; and sometimes they all first enter a circumferential cement-duct. The probable cause of the[Pg 135] greater complexity of the cementing apparatus and of the greater number of the excretory orifices in the Balanidæ, compared with the Lepadidæ, is that the entire surface of the broad basis, which answers to the whole peduncle in the Lepadidæ, is firmly cemented down to the supporting object, instead of merely the basal end of the peduncle. The cement issues either in a cellular condition, or more commonly as a fine network, which, at a short distance from the orifices (Pl. 28, fig. 4 a, z), becomes so fine as to form a sheet or layer: I may here recall the fact, that in the cement proceeding from the disc of the antennæ, in some species of Lepas, a similar structure was observed. The cement itself presents the same transparent, brown, laminated, structureless appearance, and the same chemical reaction, as described in my former volume. The cement has the capacity of occupying and filling up all inequalities in the supporting surface; I have seen it, when spread over an encrusting Flustra, present an exact model of every cell; in the case of Coronula, it seems, as we shall immediately see, to have the power of penetrating into, and even almost blending with the epidermis of the supporting Cetacean. The last-formed cement-glands and cement-ducts present a delicate and transparent appearance, and contain cellular matter; whereas the old cement-glands, and sometimes the old cement-ducts, are filled with brownish cement, not acted on by boiling potash. The foregoing remarks are confined to the sub-family Balaninæ, for I have not been able to examine thoroughly the Chthamalinæ, and can only affirm, that in Chthamalus and Pachylasma the cement-ducts repeatedly bifurcate and inosculate, in the same manner as in the Balaninæ. I will now proceed to describe, in some detail, the cementing apparatus in the several following genera.
[70] I had some slight reason to suspect in Pollicipes that new cement-glands were successively formed: this is more probable in this genus than in the others, inasmuch as it is the most nearly related to the Balanidæ.
Coronula.—The cementing apparatus is here more simple than in any other genus of the Balaninæ, and I have studied it more carefully. The basal membrane of Coronula balænaris is figured in Pl. 28, fig. 1 a, and must first be described; its relation to the shell will hardly be understood without looking at the outline of the folded walls of this species, in Pl. 16, fig. 5. The basal membrane closes the central circular hollow, and is continuous with rays (not[Pg 136] represented in Pl. 28) extending under the doubled walls and terminal transverse loops. It has eighteen concave sides, corresponding with the inner ends of the folded walls, for each of the six compartments is trebly folded. The membrane consists of successive, conformable slips (c′, c′), bordered exteriorly by thickened yellowish rims, and internally overlapping (when viewed from the inner side) the few last-formed slips, and then thinning out. The membrane forming each slip is itself laminated. The middle portion, about 1/50th of an inch in diameter, is rather opaque, owing to the slips being so close together. Beyond this central part, the slips suddenly increase in size, but yet have a different shape from the 18-sided outline, which they ultimately assume: this difference is owing to the great changes in shape, as explained under the genus Coronula, which the shell undergoes, when the walls at first assume their folded structure. The walls are invested by longitudinally striated membrane (p, p, p, fig. 1 a), which turns in under their basal edges; and this membrane is united with the basal membrane, by what I shall call the circumferential slip (b), and which is shaded in fig. 1 a, simply for the sake of catching the eye. It is the circumferential slip of membrane which sends rays under the spoke-like folded walls: thin as it is, this slip is yet laminated, but is not bordered by thickened edges. The membrane investing the walls is, like the basal membrane, formed of successive slips with thickened edges, which overlapping (viewed from the inside) the last-formed slips, project beyond them, and so face the edges of the slips in the basal membrane; they are only obscurely indicated in fig. 1 a. The circumferential slip (b) lies over (as viewed from within) both the basal and wall membrane. This whole structure will, perhaps, be best understood by the sectional diagram (fig. 1 b), in which the letters (c′, c′) show the slips of basal membrane; (p) the parietal membrane, coating the outside surface of the walls of the shell, not here represented; (b) the circumferential slip overlying both; and (z, z) the layers of cement, which may for the present be disregarded. In order to allow, of the growth of the shell, the circumferential slip (b) periodically splits in the middle, all round,[Pg 137] in a line exactly conformable to the edge of the last-formed slip of basal membrane; and likewise in straight, medial lines under the spoke-like (cut off in fig. 1 a) doubled walls. I have seen, under a high power, the line of splitting, very shortly after its formation, with the two edges ragged and near together, with an extremely narrow, new circumferential slip just formed, between and over the edges of the previously formed slip. What causes the circumferential slip to split so symmetrically, I cannot say: the opposed edges, after a time, become thickened, apparently from adhering to the underlying layer of cement, as will presently be described. The circumferential slip continues increasing in breadth till the period of its splitting arrives, by which time it has become much broader than the last-formed slip of basal membrane; and after the splitting takes place, the interior half towards the basal membrane, forms a new basal slip all round the basis, and the exterior half adds a new slip to the membrane investing the walls. This latter membrane being inflected under the basal edges of the walls, is, during the growth of their edges, drawn straight down, the newly-formed portion taking the inflected position.
In the sectional diagram, (1 b) the circumferential slip is not yet broad enough to split; when it has become so, it will split under the letter (b). The slips of basal membrane are, as may be seen in fig. 1 a, narrower towards the circumference; but the two or three last-formed slips, are out of proportion narrower than the others; and it is certain, from the comparison of the basal membranes of specimens of different ages, that these will afterwards increase in width.[71] I have seen no other instance, in Cirripedes, of growth in membranes, except at their extreme margins: I suspect that these last-formed slips are pulled, during the downward and outward growth of the shell, a little from over the last-[Pg 138]formed slips, new and larger laminæ being all the time thrown down, so as to prevent any fissure being formed. I also suspect that the gradual increase in width of the circumferential slip itself, is due to the opposed edges of the underlying and last-formed circumferential slip being dragged further apart from each other, new and wider laminæ of membrane being continually thrown down; the new circumferential slip being thus, also, all the time thickened, as well as rendered broader.
[71] In the case of one young shell, I found that the previously-formed circumferential slip must have split, long before it had assumed its proper and ordinary width; for the last-formed slip of basal membrane was of extreme narrowness, and would have to be considerably added to in width, whilst the new and narrow circumferential slip was likewise being added to in width. This slip of basal membrane, though so extremely narrow, had its own cement-ducts and glands.
The central slip or rather disc of membrane, is 3/400ths of an inch in diameter; and this shows the basal diameter of the shell immediately after the metamorphosis. In the middle of this little disc I saw, in several specimens, the prehensile, pupal antennæ; I made out distinctly the ultimate segment with its bristles, and, as I believe, the disc-segment, which was 7/2000ths of an inch in diameter; but this portion was much obscured by the quantity of cement. When the corium is removed from the inner side of the basal membrane, the two chains of glands, extending from exactly over the antennæ in the centre about half way towards the circumference, are conspicuous. The cement-trunk, connecting the glands, is thin, and at the further end is always broken off, by the removal of the corium and overlying layer of ovarian cæca, into which the two cement-trunks enter; and without which removal, nothing could be seen. The two chains of glands form a very large angle, open towards the rostral end of the shell, as represented at fig. 1 c, much enlarged; by a mistake in fig. 1 a, the two have been drawn in a straight line. The cement-trunk increases in diameter in proceeding from the centre to the circumference, and the glands likewise increase in size, at the same time altering somewhat in shape. From near the lower side (the basal membrane being viewed from within) of each gland, two cement-ducts proceed, which pour out their contents beneath the basal membrane. The orifices of the ducts always exactly face the middle folds of the two lateral, and two carino-lateral compartments. In a full-sized specimen, there are from thirty-five to forty cement-glands on each side, always corresponding exactly with the number of slips of basal membrane, including the[Pg 139] circumferential slip, to which the last-formed pair of glands and cement-ducts belong. In correspondence with the great number and narrowness of the central slips of membrane, so are the cement-glands towards the centre numerous and very small. All the glands, in the more central parts, consist of a mere transverse enlargement of the cement-trunk; but the exterior and larger glands, which are more closely packed together, are more globular or pear-shaped; and the two ducts (of which the one on the rostral side is considerably enlarged at its base) do not come out of the gland exactly at the same level. The trunk, connecting the glands, runs straight from one to the other. The ducts proceeding from the outer and older glands, on the carinal side, are much curved (Pl. 28, fig. 1 c). To give an idea of the dimensions of the several parts, I may state that the largest ducts were 3/1000ths of an inch in diameter, and the glands belonging to them nearly thrice as much, measured in the direction of the cement-trunk; on the other hand, some of the ducts from the small central glands had a diameter eighteen times less than that of the largest ducts. Towards the circumference, the ducts that proceed from the older and larger glands are piled one exactly over the other—the last formed being the topmost, and all are imbedded in the corium which overlies the basal membrane: this position of the ducts, one over the other (which could not be well represented in figs. 1 a and 1 c), is owing to their all debouching at the same exact point. But the ducts form the smaller and younger glands, when the shell had a different shape, are spread out, and are all attached to the basal membrane. Altogether, the basal membrane of Coronula, when well cleaned, and examined under a moderately high power, presents the most singular and elegant appearance.
We now come to the cement-tissue: this lies on the under or outer side of the basal membrane; it is not represented in figs. 1 a or 1 c, but only in the sectional diagram, 1 b, by the letters z, z: it presents its usual character and appearance, like solid glue or brown gum, but is obliquely laminated and sub-laminated: it forms a layer, much thicker than the basal membrane itself, being as[Pg 140] much as .004 or .005 of an inch in thickness. It is, however, difficult to ascertain its thickness, from the singular manner in which it penetrates into and almost blends with the epidermis of the whale’s skin; so much so, that for a considerable time I thought (not then knowing anything about the cement of Cirripedes) that this transparent horny substance probably answered to a corn on the human foot produced by pressure. But I soon observed that this horny substance certainly extended into and up the cement-ducts; and this fact first led me to the examination of the whole subject in the several genera of Lepadidæ and Balanidæ. It was not difficult to remove the cement-ducts, leaving small portions of the contained cement projecting freely up as points from the general surface of cement. The cement adheres slightly to the whole basal membrane, but chiefly to the yellowish rims or edges of the successive slips; and it is indeed this adhesion which, I believe, produces the rims; for the circumferential slip, when first split, had very thin ragged edges. The cement also extends under the spoke-like prolongations of the circumferential slip, and likewise some way up the sides of the walls.
The cement-glands, the trunk, and the ducts, except the two, three, or even four last-formed ones, are all filled with an apparently solid thread of transparent, brownish cement, differing in no respect from the cement under the central parts of the basal membrane. In one instance, in which the last-formed pair of glands and ducts had apparently been only just developed, they were so perfectly transparent that I could distinguish them only under certain lights, and I could not perceive any contents. The last-formed glands and ducts always appear very delicate, and include a tube of very delicate tissue, containing more or less of granular matter. The next succeeding pair of glands and ducts are always more opaque, and contain much more granular matter; which, in the next, or next but one, may be seen passing into the state of pale brown, transparent chitine. I have seen some most distinct instances, in which, in the same duct, the part towards the centre of the basis was filled with homogeneous cement, and the part towards the circumference with still plainly granular matter. In the[Pg 141] successive circular slips of cement-tissue, lying attached under the circumferential slip and under the two or three succeeding slips of basal membrane, an exactly analogous series of changes is presented; as indeed might have been expected, as the slips of cement are absolutely continuous with the contents of the ducts. Moreover, if a vertical section be made across one of the last-formed slips of cement, it may sometimes be seen to be apparently in the act of separating into layers, with the lower layers more pulpy, elastic, and white than the upper layers, which are less coherent, and show as yet even still less the character of cement. The cement under the circumferential and adjoining slips, often presents a peculiar wrinkled appearance, in lines conformable with the outline of the basal membrane; but I do not believe that these are real wrinkles, though so like them; they seem to consist of sinuous threads, longer or shorter, sometimes slightly branched, crossing and inter-joined, and composed apparently of faintly coloured cement. I suspect that these threads are formed by the union and subsequent drawing out of aggregations of that matter, which within the ducts is first granular, and then changes into cement; for at the orifices of the ducts these wrinkled threads sweep outwards in curved lines on both sides. The cement in these early stages adheres, with very little force, to the basal membrane; and with only a little more force to the underlying layers of cement; in fact, till it assumes the brown translucent appearance, like solid glue, it hardly seems to act as cement.
How the cement reaches the skin of the whale, will be best understood by referring to the sectional diagram (Pl. 28, fig. 1 b). When the circumferential slip of membrane (b) splits, a new circumferential slip will be formed over it, together with new cement-glands and ducts, and cement (z, z) will issue from four new orifices, and will extend on both sides of these orifices, till the ends meet and become united, thus forming a narrow, 18-sided, continuous, new slip of cement, with 18 spokes proceeding from it. I have not noticed lines of union in the cement of any one slip; but the matter forming each slip, certainly has proceeded from four distinct orifices. Seeing how per[Pg 142]fectly successive layers of cement often become blended together, lines of union or junction, could hardly be expected to be preserved in the same individual layer. When the circumferential slip of basal membrane splits, the underlying slip of cement, which we will call A, does not split, but becomes stretched, so that the newly formed slip of cement, which we will call B, does not reach the skin of the whale. As the new circumferential slip of basal membranes goes on increasing in width, A continues to be stretched, but does not split, till at least another circumferential slip of basal membrane has been formed and has been split, and till B has been also stretched. By this time, the cement-tissue A has assumed its normal structure, and has the power of adhering to the whale’s skin, which it has now reached, owing to the splitting of the under and older slips of cement. At the next period of growth, A itself will split, and B will touch the whale’s skin and adhere to it; and this, also, will ultimately split. It results from this action, that the cement has a stretched, and sometimes even a fibrous appearance, with the lower edges of the layers, of which each slip of cement is formed, thinning out. I have before stated, that the two or three last-formed slips of basal membrane are formed at first too narrow, and apparently have to be dragged outwards, over each other; and it is perhaps owing to this circumstance, and to globules of cement having first adhered to the under surface of the slips of basal membrane, that these slips are studded beneath with parallel little vermiform bodies, sometimes of considerable length, and furnished with heads, all directed outwards. These tapering, vermiform bodies have a considerable resemblance to the threads before mentioned, which give the wrinkled, concentric appearance to the newly-formed layers of cement, and have probably a closely analogous origin: in one case, indeed, it appeared as if some of these concentric threads were in process of being drawn out at right angles to their original course. Lastly, it should be observed, that as the exterior half of the membrane of the circumferential slip, after each splitting, is dragged down, and thus comes to invest the outer surface of the wall of the shell (the wall not being represented[Pg 143] in the diagram, but standing where the letter (p) stands), so it must be with the cement, which thus likewise comes, in an unusual manner, to invest the outer surface of the folded walls of the shell, and attaches them to the skin of the whale,—which latter is always growing upwards, and tending to bury the shell.
Platylepas decorata.—This genus is closely allied to Coronula, and the cementing apparatus is essentially similar. In one specimen, I counted no less than forty-nine slips of basal membrane, each of which, of course, had its pair of cement-glands, and each of the latter its two ducts. The glands consists of a transverse enlargement of the trunk, as in the early stages of Coronula. Neither the glands nor the duct, when old, become filled up with cement, but only the main-trunk. The ducts are very delicate and thin; the larger ones being only 3/10,000 of an inch in diameter. The glands stand some way apart on the two cement trunks; and the latter, instead of being straight as in Coronula, are deeply serpentine; the glands are formed on each bend, so that, though all on one side are connected on the same trunk, they form a double row on each side of the basal membrane. The basal membrane (in the centre of which I distinctly saw the antennæ of the pupa) has six deep bays or excisions, corresponding with the midribs (see Pl. 17, 1 a, 1 d) of the six compartments; and the two ducts from each gland, on the right and left sides, debouch at the heads of the four lateral excisions, exactly opposite the midribs of the lateral and carino-lateral compartments. The later-formed glands, owing to all of them being situated some way apart from each other on the two cement-trunks, lie further from the centre of the basis than do the orifices of their ducts; hence the later-formed ducts are directed a little backwards, or from near the circumference towards the heads of the deep excisions.
Tubicinella.—The cementing apparatus is here less symmetrical; but this, I believe, is chiefly owing to the basal membrane being formed of successively larger discs (not slips) of membrane, thrown down not quite concentrically one over the other; each new disc of membrane seems to cover the last-formed cement-glands and ducts; and there are[Pg 144] as many ducts and glands as there are discs of membrane, all adhering together. In one specimen, it appeared that normally there were four sets of cement-ducts, as in the allied genera of Coronula and Platylepas; but in other specimens, the ducts were distributed very irregularly. In one case the two cement-trunks extended parallel and close together, one of them terminating long before the other. I have given a figure (Pl. 28, fig. 3) of three of the cement-glands, removed from the basal membrane, together with their ducts. The cement-trunk (f f) seemed to be a little enlarged, and to be crossed by septa, as it entered the glands (h), but I could not make out this structure clearly enough to be represented. Whilst young, the cement-glands stand some little way apart from each other; and in the figure given of some of the last-formed glands, they are hardly separate enough. Each gland gives out obliquely, on one side, a cement-duct (c) which I traced in several cases to the margin of one of the discs of basal membrane, where cement issued from it; and on the opposite side, a tapering spur (b), varying in length, which may be called, and I believe really is, a rudimentary duct. Of these spurs we shall meet many instances in other genera. The duct (c) and the spur (b), close to where they entered the gland, in some specimens gave off, at about right angles, short blunt points, or rudimentary branches. This duct and spur correspond, I believe, with the two ducts in Coronula; but besides these, a duct (a) is given off from one end of the gland, from the surface opposite to that at which the cement-trunk enters. This duct (a) is very singular, from always forming a loop (a′), with two spurs projecting from it: these two spurs occasionally spring from a common point: I have seen nothing like this structure in any other Cirripede. This duct (a) runs, like the duct (c), to the margin of its own disc of basal membrane, where it debouches. Besides these ducts, in the best specimen which I examined, there were two other sets of ducts, which were slightly zig-zag, and at each angular bend, a mere knob or point, or at most a short branch, was given off; but this branch seemed never to run to the margin of the basal membrane or to give out cement; whereas the main branch did give out cement. I was[Pg 145] not able to trace these ducts to their glands. In these zig-zag ducts, and in the rudimentary points sometimes observed at the base of the duct (c), and likewise at the base of the spur (b), we see the first indication of that tendency to bifurcation, so strongly characteristic of the cement-ducts in all the genera, excepting those already described, which are allied to Coronula.
Chelonobia patula.—The cementing apparatus is here chiefly remarkable for the thinness and straightness of the main trunk, (f f, Pl. 28, fig. 2), and from the great distance at which the glands stand apart; had another gland been drawn, it would, on the scale here used, have stood exactly under the two upper, (c′ c′) in fig. 1 c. We here see that the trunk (f), before entering the gland (h), has an enlarged portion (g); this, I suspect, is a very general structure. Each gland gives out, on opposite sides, two ducts (a a, b b), larger even than the main trunk; and these ducts bifurcate repeatedly, and inosculate. By this inosculation it is not improbable that all four ducts, proceeding from the two glands of the same age, may be connected together; certainly the bifurcating branches from the same duct thus become repeatedly connected. For the first two or three bifurcations the ducts decrease very little or not at all in diameter; but nearer the circumference they become smaller. The ducts, also, proceeding from the younger and smaller glands, are, of course, proportionably smaller. In one case I was able to count four bifurcations in the duct between the gland and the edge of the basal membrane. It follows from this structure, that the basal membrane, at each period of growth, is cemented down by cement issuing from several orifices; but we shall presently find that in other genera the cement proceeds from many more orifices. In fig. 2 there is represented, by the aid of the camera, a small portion (from the outer (a) to the outer (b) being 12/100ths of an inch in length) of the basal membrane, with all the several cement-ducts adhering to it, which I could distinguish, and drawn of their proper relative sizes; this figure also shows some of the bifurcations, but no inosculation happened to be included in the space here given; the basal membrane itself has not been represented. In taking[Pg 146] a view of a considerable portion of the basal membrane, especially towards the circumference, some parallelism in the branches could be perceived; one set of branches tending to run in the direction of the ray of the circle, and the other set in the line of the circumference.
Elminius Kingii.—The cement-glands here resemble those of Chelonobia, but the trunk does not seem to be enlarged before entering the gland. The glands are situated rather far apart; and the chief peculiarity is, that the trunk connecting the glands is as tortuous as the track of a worm. Each gland gives out two ducts, which bifurcate repeatedly, and often inosculate, making, in parts, an hexagonal mesh-work: some of the branches do not debouch on the basal membrane, but terminate in blunt points. So numerous are the ducts, that the basal membrane may be compared to pieces of paper with the fine fibrous branching roots of some plant dried and heaped on it. Some of these ducts are very regularly jointed, and resemble a conferva,—an appearance which I believe is owing to divisions in the contained cement; other ducts are partially marked by little wrinkles, as presently to be described under Balanus. The cement, instead of, as heretofore, invariably forming a slip round and beneath the circumference of the basal membrane, here often forms little, independent, circular, and irregularly-shaped discs, each of which has issued from a single orifice. I may here add that in two species of Tetraclita I saw the cement-ducts repeatedly bifurcating, with some of the branches inosculating, as in Elminius and Chelonobia.
In Balanus balanoides, which, like all the Cirripedes hitherto mentioned, has a membranous basis, I could only make out an amazing number of bifurcating and inosculating cement-ducts, of very various diameters. The cement-tissue, on the under side of the basal membrane, generally consisted of little circular discs, on an average from 2 to 5/1000ths of an inch in diameter; but there were also globules and short tortuous threads of cement. In some very much elongated and crowded specimens—in which, during the downward growth of the walls, the basal membrane had ceased to reach the surface of attachment, and being thus[Pg 147] suspended had become, as viewed from the outside, deeply concave—the cement had apparently continued to try to reach the rock, and now hung down in the form of two thickish roots, some tenths of an inch in length. These roots were round, and tapered either to a fine or blunt point; one was doubled on itself, and so had become united; in the other, I could perceive five layers or sheaths of the cement-tissue, one within the other; the innermost of these layers, which once, no doubt, formed the outside surface of the root, was only a quarter of its present length.
In Balanus tintinnabulum, the basis is calcareous: when its upper surface is cleaned, dried, and examined under a good light, the numerous larger cement-ducts can be seen, even by the naked eye, or under a weak lens, and present an elegant appearance. These larger ducts run in parallel lines from the two chains of glands towards the circumference. They are all encrusted with calcareous matter, and in the more central parts are hidden under it; at each period of growth, when the basis is added to round the circumference, it would appear that a layer of excessive tenuity of shell is thrown down over the whole surface, just in the same way as in Tubicinella, at each period, a new and larger disc of membrane was thrown down over the pre-existing membranes with their cementing apparatus. The cement-glands, in the middle of the basal plate, seem often to give rise to small abnormal depositions of calcareous matter. When the basis (it is best to take a young specimen) is slowly dissolved in acid, all the cementing apparatus is left uninjured, adhering to the delicate tissue which before existed in a calcified condition. Near the middle I saw the two antennæ of the pupa; and from them the two cement-trunks extended about half-way towards the circumference. These two chains of glands are often placed very irregularly, but they tend to form, as in Coronula, a large angle, open towards the rostral end of the shell. The glands, close to the old antennæ, commence abruptly, of rather large size: the later-formed glands, with their ducts, are in regular order larger than the younger ones, and stand much closer together. After immersion in acid all the glands and ducts appeared empty, instead of the older[Pg 148] ones being, as in Coronula, filled with cement. In one case I counted on each trunk twenty-five glands, besides some smaller obscure ones close to the centre.
In Pl. 28, fig. 4 b, I have given a drawing of two of the cement glands: the cement-trunk (f f) is smooth and apparently cylindrical: it becomes enlarged (at g) before entering the gland: it seems even to be prolonged across the gland under the form of a narrow bar (not represented), which apparently serves to keep the two ends of the trunk, on the two sides of the gland, in their proper relative places and distances. The gland itself is an elongated bag (h), which properly lies exactly over the enlarged portion (g) of the trunk, but in the drawing has been purposely displaced: it gives rise, in the later-formed glands, to a sort of neck (see the upper gland), which is either so long as to deserve rather to be called a duct and which soon bifurcates, or is quite short (see the lower gland) and gives rise to two separate ducts. On the opposite side of these glands, there is a spur (m), of greater or shorter length, which is evidently a rudimentary duct, for in the younger glands it existed as a perfect duct. Moreover, the first-mentioned duct often gives off branches (t′), having an exactly similar appearance with the spur (m). The membrane of which the cement-trunk (f), with the enlargements (g), is composed, is smooth, but that of the glands and of all the ducts, presents a very peculiar appearance, which at first would be called scaled, but more properly perhaps notched,—each notch being apparently formed by a line of thickened membrane, extending obliquely round only a short portion of the tube, and indenting it. The ducts, which I measured, were between 1/3000th and 4/3000ths of an inch in diameter.
In fig. 4 a, I have given a drawing of the two chains of glands, but with only those ducts figured which proceeded from the last-formed pair of glands. The specimen here drawn was old; and it is rare to find the structure of the ducts so simple. From both glands[72] a neck or thick duct arises, which soon bifurcates; one branch runs direct into[Pg 149] the circumferential duct, and the other (t) bifurcates again; of the latter, one branch unites with its fellow from the opposite gland, and then forming a single duct (t′) enters, as do the two other branches, the circumferential duct. Thus, into the latter, five main ducts enter: the position of their points of entrance, with respect to the shell, varies considerably; but I think the five points tend to face the middle of the rostrum, and middle of the two lateral compartments on each side. In some other specimens, in which the ducts were nearly as simple, I observed that the neck or main duct at once divided into three branches, instead of into two, with one soon bifurcating; and on one side a rudimentary branch or spur was given off (above t), indicating a tendency to an additional bifurcation. In the later-formed glands, the ducts proceed only from the outer sides and form the ends of the glands furthest from the centre; but in the earlier-formed and smaller glands of the same individual, other ducts proceed from the inner sides, where in the older glands the spurs (m) are situated: moreover, in the younger glands, all the ducts bifurcate much oftener (how often I was not able to ascertain), before entering the circumferential duct; many of the branches, however, terminating in spur-like points. Now if we imagine twenty or thirty repetitions of the ducts given in fig. 4 a, (independently of the greater complication of the ducts of the younger glands), each a very little smaller than the other, and placed, with the main branches parallel, one over and within the other, we shall gain some insight into the wonderfully complicated structure of the cementing apparatus in this and many other species of Balanus.
[72] It should be observed that fig. 4 b ought to have been drawn with its present upper end downwards, to make it correspond in position with fig. 4 a.
I have as yet only alluded to the circumferential duct (i, i, Pl. 28, fig. 4 a): we have not hitherto met with this duct, but I suspect that the branches which in Chelonobia inosculate, and which seem to run nearly parallel to the circumference of the basal membrane, answer the same purpose of connecting the ducts together, and are, perhaps, strictly homologous. In this, and some other species of Balanus, the last-formed circumferential duct runs round the margin of the upper lamina of the basal plate, close to the basal edges of the walls; and as these latter have[Pg 150] projecting longitudinal ribs, the duct curves a little round each rib; so that the whole duct is formed by as many short inwardly curved portions as the walls have ribs, or longitudinal septa. Between the basal extremities of these parietal, longitudinal septa, the extremities of the radiating septa of the basis project and enter; and along the crests of the latter, little branch-ducts (i′), proceeding from the circumferential duct, extend. In the basis, beneath the tubes formed by the just-mentioned radiating septa, there is a cancellated shelly mass (which, in fig. 4 a, was of unusual thickness), and along the crests of the branching ridges forming this cancellated mass, the sub-branches of the above branch-ducts (i′) run; these soon become so minute as not to be distinguished by the highest powers, and thus form a sheet of cement, which attaches the last-formed zone of the shelly basis to the supporting surface. At what point the membrane forming any one duct ceases, the cement-tissue being alone left, I was not able to ascertain; but the lower parts of the reticulated slip (z, z, fig. 4 a) closely resembled the cement-tissue which surrounds the disc-segment of the pupal antennæ in Lepas australis. The circumferential duct, here and there, forms little loops, as may be seen in fig. 4 a: and often two branches, running along the crests of two adjoining basal septa, proceed from a common point of the circumferential duct. The cement itself, under different parts of the basis, appears as little separate discs, as threads, globules, and as a fine network, but most commonly as simple layers. As each thick zone of shelly matter is added round the basis, the exterior branches of the ducts, between the circumferential duct and the new layer of cement beneath, are fairly imbedded in shell, and are for ever hidden, without, indeed, acid be used for the dissolution of the calcareous matter: so, also, the pre-existing ducts and glands, and the main trunk, would all have been hidden, if the layer of calcareous matter, which, I believe, is thrown down at each period of growth over the entire surface, had not been of excessive tenuity.
I cursorily examined the cementing apparatus in Balanus galeatus, improvisus and crenatus, which have all calcareous bases, and belong to different sections of the genus; and[Pg 151] the structure seemed to be essentially the same. In Bal. galeatus, I found the cement-ducts varying in diameter from 1/4000th to 1/10,000th of an inch in diameter. In B. improvisus, the cement-glands do not differ much from those of B. tintinnabulum; but the cement-ducts bifurcate often before entering the circumferential duct; and the little branches, which proceed from the latter, are very short, and almost immediately, owing to the thinness of the basis, blend into a slip of cement.
I hope to be excused for describing at such length, the apparatus by which sessile cirripedes are permanently attached to a supporting surface; for this is the great leading character of the sub-class, not hitherto observed in any other Crustacean.[73] It is not easy to overstate the singularity and complexity of the appearance of the basal membrane of a Balanus or Coronula: and when we consider the homological nature of the apparatus, the subject becomes still more curious: I feel an entire conviction, from what I have repeatedly seen in several genera of the Lepadidæ, both in their mature and pupal condition, and from what I have seen in Proteolepas, that the cement-glands and ducts are continuous with and actually a part of an ovarian tube, in a modified condition; and that the cellular matter which, in one part, goes to the formation of ova or new beings, in the other and modified part, goes to the formation of the cementing tissue. To conclude with an hypothesis,—those naturalists who believe that all gaps in the chain of nature would be filled up, if the structure of every extinct and existing creature were known, will readily admit, that Cirripedes were once separated by scarcely sensible intervals from some other, now unknown, Crusta[Pg 152]ceans. Should these intervening forms ever be discovered, I imagine they would prove to be Crustaceans, of not very low rank, with their oviducts opening at or near their second pair of antennæ, and that their ova escaped, at a period of exuviation, invested with an adhesive substance or tissue, which served to cement them, together, probably, with the exuviæ of the parent, to a supporting surface. In Cirripedes, we may suppose the cementing apparatus to have been retained; the parent herself, instead of the exuviæ, being cemented down, whereas the ova have come to escape by a new and anomalous course.
[73] Rathke has described (‘Acta Nova,’ 1839, p. 147), in some siphonostomatous crustaceans, a pair of curious organs, which serve to secrete a substance that holds the eggs attached together in a mass to the parent’s body: these organs Rathke has designated by a similar name to that which I have used, namely, the cementing organs or receptacles; they are distinct from the oviducts, but enter them near their external orifices. As in Cirripedes, the cement-glands and ducts are certainly continuous with an ovarian tube; and as they occupy a quite different position in the animal’s body, these organs of Rathke, though in some degree analogous in function, must be homologically distinct.
Under the Order it has been stated that the Balanidæ, are, on the cirripedial type, the highest in the class; that is, they are the most complicated, but not (to use Professor Owen’s term) by mere vegetative repetition. Amongst the Balanidæ, the first section of the genus Balanus may be taken as typical; here we have the structure of the shell extremely complicated, yet beautifully adapted for strength, and for the protection of the included body. The cementing apparatus is here, also, most complicated. I have divided the Balanidæ into two natural sub-families, the Balaninæ and Chthamalinæ, in accordance with certain differences in the structure of the shell and of the animal’s body: that this division is natural, might almost be inferred from the one fact, that all the characters by which the Chthamalinæ differ from the Balaninæ, are those by which the former approaches the family of the Lepadidæ; moreover, certain anomalous characters in the Chthamalinæ, as the supplemental whorls of compartments in Catophragmus, and the presence of caudal appendages in this same genus and in Pachylasma, reveal this same affinity. The only objection which I can see to the separation of the sixteen genera into the above two sub-families, may be drawn from the degree to which they blend together; thus, as far as the shell is concerned, Chelonobia, in one important internal point of structure, tends to assume the character of[Pg 153] the Chthamalinæ; and on the other hand, Pachylasma, a member of the Chthamalinæ, has a shell, which if not examined during its earliest growth, would be placed without doubt amongst the Balaninæ. But it fortunately so happens, that in no one character of the body does Pachylasma approach the Balaninæ more nearly, than do the other members of its sub-family; or Chelonobia approach, in the same respects, the Chthamalinæ. It is only in Chthamalus, of which the shell clearly places it in the sub-family bearing its name, that in some of the species, the less bullate labrum,—the larger palpi,—the lower teeth of the mandibles being laterally double,—and the lower segments of the third pair of cirri being thickly clothed, like the lower segments of the second pair, with bristles—all show that these species make an approach in structure to the Balaninæ.
It will be seen that I have divided the Balaninæ into two little groups, according as whether the branchiæ consist of one or of two plicated folds of membrane, and as whether or not the scutum and tergum are articulated together. I have been greatly tempted to follow Drs. Leach and Gray, who have separated the second of these groups, containing the genera Coronula, Tubicinella, Xenobalanus, and Platylepas, into the sub-family of the Coronulinæ. Certainly these genera have a peculiar aspect in common, and agree in being parasitic and imbedded in the skin of Cetaceans, as is the case with the first three genera, or in that of turtles, manatee, and sea-snakes, as in Platylepas. Though these genera possess several peculiar characters, yet I can find none common to all four, excepting their imbedment in the skin of Vertebrata, their double branchiæ, and their non-articulated opercular valves; and these I do not think of sufficient importance to serve for the separation of a sub-family; for in Chthamalus, one species has double branchiæ, one species has no branchiæ at all, and the other species have single branchiæ; so again in Chelonobia, the scutum having only a horny articular ridge, makes an approach to Coronula and its allies. I may further specify that the folded walls, a singular character common to Coronula, Platylepas and Xenobalanus, fails in Tubicinella; the open tubes and the[Pg 154] imperfect outer lamina of the parietes towards their bases, are characters which fail in one species of Platylepas; the muscles running to the opercular valves being thinly spread out, and partially without transverse striæ, is also a character which fails in Platylepas; the simplicity of the cement-ducts partially fails in Tubicinella; and lastly, the existence of small intermediate teeth on the mandibles, fails in Xenobalanus: hence, I repeat, I have not thought it prudent to admit the sub-family of the Coronulinæ though in many respects a very natural group.
The genera in the Balaninæ and Chthamalinæ are founded chiefly on the number of the compartments (the number being apparently due, as previously explained, to the fusion or abortion of certain of the eight typical compartments); and secondarily, on the nature and even form of the basis, and on the porosity of the walls. In Coronula and its allies, the non-articulated opercular valves and deeply folded walls come into play. As a justification for using these characters in distinguishing the genera, and even to a certain extent in separating the two sub-families, I must call to mind that the shell, with the basis, is not merely a dermal envelope, as amongst Molluscs, but actually consists of the first three segments of the head. The parts of the mouth and the cirri are of very little service in distinguishing the genera,—a singular fact, considering that most of the genera amongst the Lepadidæ could be distinguished by these organs,—though trifling details in their structure sometimes come in useful as specific characters. Balanus, with the sub-genus Acasta; Pyrgoma, with the sub-genus Creusia; Tetraclita, and Elminius, are genera of about equal value; though perhaps the two latter are rather more nearly related together than to the others. Chelonobia is more distinct; it shows some little affinity to the Chthamalinæ, and likewise to the four following genera. Coronula, Platylepas, Tubicinella, and Xenobalanus, are genera quite distinct from the foregoing, and from each other; yet, as we have just seen, palpably allied together. Amongst the Chthamalinæ, Pachylasma, Octomeris, and Catophragmus, are more closely related to each other than to the other two genera of the sub-family; yet Pachylasma,[Pg 155] as far as the shell is concerned, leads into the Balaninæ, and Catophragmus into the Lepadidæ; Octomeris leads towards Chthamalus, and Chthamalus towards Chamæsipho.
Variation.—The discrimination of the species in most of the genera, offers very great difficulties. I cannot too strongly impress on any one intending to study this class, not to trust to external characters: he must separate and clean, and carefully examine the internal structure and form of the compartments, and more especially of the opercular valves. After considerable experience, when numerous varieties of a species have been carefully examined, the eye acquires a sort of instinctive knowledge, by which it can recognise the species, though the character cannot be defined by language; but I have found that no amount of experience with some of the commonest species, will save frequent and grave errors, as long as external characters alone are trusted to. Not only does every external character vary greatly in most of the species, but the internal parts very often vary to a surprising degree; and to add to the difficulty, groups of specimens not rarely vary in the same manner. After having given up several years to the study of this class, I must express my deliberate conviction that it is hopeless to find in any species, which has a wide range, and of which numerous specimens from different districts are presented for examination, any one part or organ,—which from differing in the different species is fitted for offering specific characters,—absolutely invariable in form or structure. I may in one respect even go further, and affirm, that, if in a species, any part or organ differs remarkably from the same part in its congeners, then if many specimens are examined, especially when collected from different districts, such part or organ will be found eminently variable. I may instance the antenniformed third pair of cirri in Chthamalus antennatus, the teeth on the posterior cirri in Acasta sulcata, the terga in Pyrgoma dentatum, the adductor ridge of the scuta in Pyrgoma cancellatum and in Creusia, and other such cases: hence it will not do to found a species on a slight, or sometimes even on a considerable difference, in any single point or organ. On the other hand, I am far from asserting, that if only half-a-dozen specimens of some[Pg 156] rare species of Cirripede be brought from some one quarter of the world, characters beautifully defined may not be readily discovered. In determining what forms to call varieties, I have followed one common rule; namely, the discovery of such closely-allied, intermediate forms, that the application of a specific name to any one step in the series, was obviously impossible; or, when such intermediate forms have not actually been found, the knowledge that the differences of structure in question were such as, in several allied forms, certainly arose from variation,—for instance, in the case of two shells otherwise identical, one being longitudinally ribbed and the other smooth, a character which we know to vary,—but I have always used this argument from analogy with great caution. Finally, as in the large genus Balanus, there is an especial amount of variation, I have there entered in detail on this subject; and I hope that those interested in it, will refer to that discussion, which is almost verbally applicable to some other genera of the family, as Tetraclita and Chthamalus.
In my former volume I have shown that the pedunculated cirripedes grow rapidly; this is likewise the case with the Balanidæ. Mr. Stutchbury informs me that he has seen numerous specimens of Balanus tintinnabulum from 2 to 3 inches in height and from 5 to 6 inches in circumference (and this is nearly the full size which the species attains), on a vessel which had been to sea only during one year. At Coquimbo, in Chile, I have seen a specimen of B. psittacus 1.3 of an inch in basal diameter, and .8 in height, adhering to a chain that had been only six months under water. Poli, also, gives the case of some Balani (probably B. perforatus), which, in about four months, had attained a basal diameter of 1 inch, and a height of 1-1/6th of an inch. Balanus balanoides is a smaller species, and of slower growth; for the late Mr. W. Thompson, of Belfast, found that in three months from July 3d, certain marked specimens had increased from 2-1/2-3 lines to 4-1/2 lines, which is the usual maximum size attained in that locality. From other observations, Mr. Thompson believes that the extreme duration of life of this[Pg 157] species is about two years: whether the other and apparently quicker-growing species, are shorter-lived, I have no means of judging.
In accordance with this rapid growth is the frequency of the periods of exuviation. Mr. Thompson kept twenty specimens of B. balanoides alive, and on the twelfth day he found the twenty-first cast-off integument, showing that all had moulted once, and one individual twice, within this period.[74] This frequency of exuviation, together with the durability of the cast-off integuments, explains the astonishing masses of exuviæ, which Mr. Peach assures me he annually has observed off the coast of Cornwall; they are most abundant in April and May, but he has seen quantities also in September; he could easily, as he tells me, have filled several quart-measures with them. The specimens sent to me consisted chiefly of Balanus balanoides, perforatus, and Chthamalus stellatus. The opercular membrane, with a narrow strip from between the two scuta, and another narrow strip from between the two terga, are moulted together, in connection with the more delicate membrane lining the sack, and investing the plicated branchiæ. This membrane, likewise, is continuously connected with that covering the whole body and its appendages. As I have stated under the Lepadidæ, the inner tunic of the œsophagus, of the rectum, of the olfactory pouches, and that which enters a little way into the acoustic meatus, and the apodemes of the maxillæ, are all moulted. On the cirri and jaws, new spines are formed with their upper ends enclosed within the old spines, but with their lower ends projecting inwards, beyond the bases of the old spines, and inverted like the fingers of a glove hastily pulled off. The membranes of the body, in the act of exuviation, split, I believe, only over the prosoma. How the neat separation of the opercular membrane, from all round the sheath and opercular valves, is effected, I do not fully understand; but it is, probably, analogous to the splitting of the thick carapace of the common crab. I suspect in Coronula, in which genus and its allies the opercular[Pg 158] membrane is not periodically moulted, that the membrane lining the sack is not always thrown off at the same exact time with that of the body. In Chthamalus stellatus, in the act of moulting, the opercular membrane is the last part that separates from the new underlying membranes: I find that this species can moult when kept in a damp box out of water. The new membranes of the body, immediately after the exuviation, are not lax in any extreme degree. The exuviæ of the genus Chthamalus, and of some other genera amongst the Chthamalinæ, can at once be recognised by the divergence of the posterior four pairs of cirri: in the case of Chthamalus stellatus I have also noticed that the animal generally dies with these cirri in the same divergent position. Finally, I cannot doubt[75] that the Triton described by Linnæus was only the exuviæ of some Balanus (probably B. porcatus); Linnæus mistaking the probosciformed penis for the mouth of his imagined distinct animal.
[74] In Daphnia, Straus (‘Mém. du Museum,’ tom. vi, p. 151) found that the individuals moulted every five or six days.
[75] Linnæus speaks of the included body (inhabitant as he calls it) of other Cirripedes, as a Triton; and this, I think, shows that Lesson’s conjecture that the Triton was an Alepas cannot be correct; for Linnæus could hardly have supposed that a pedunculated cirripede inhabited another pedunculated or sessile cirripede.
I have seen a few specimens showing that when the shell has been broken it can be repaired; and this I believe is effected by the growth of a crest of corium between the broken edges, and the subsequent calcification of this crest. Mr. Stutchbury possesses a monstrous specimen of Chelonobia testudinaria, in which one of the lateral compartments on one side has not been developed. The cirri not rarely get cut off, but are, as it appears, soon repaired. I have observed a singular number of examples of the act of reparation in a group of the Australian Balanus vestitus. The manner in which the cirri are repaired seems to me curious: the cut-off end is closed by a rounded scale of yellowish chitine, and then the corium, in the four or five subjacent segments, separates from the external articulated membrane, which now serves only as a case or capsule. The tube of corium thus enclosed, with its contained muscles, shrinks a little, and soon can be perceived to be in process of[Pg 159] dividing into new and smaller segments, in one instance ten in number,—which at the next exuviation would, no doubt, be invested with an external membrane, and be freely exposed. In another instance, the pedicel of a posterior cirrus had been cut off and subsequently closed; in this instance, a whole, immature, miniature cirrus, with the two rami, each having fifteen minute segments, was thus enclosed in what had been the single lower segment of the pedicel. I have seen several specimens of Balanus balanoides, as described under that species, with several of the cirri and the penis truncated; but I believe this was owing to monstrosity, which seemed particularly to affect the male organs of generation; for no reparation seemed to be in progress. In a specimen of Coronula, however, the penis appeared to have been really cut off by accident; it had been closed, by a scab, with concentric lines, like the articular rings on the penis itself; and within the case thus formed, the corium had healed, and had become pointed, but inverted; I presume that the point would, after another exuviation, have been everted, and its length thus increased.
With respect to range, the results arrived at have no particular interest, for the species are not sufficiently numerous; and, what is still more adverse, the genera, with unimportant exceptions, range over the world; so that there is no scale of differences, and it cannot be said that these two regions differ in their genera, and these two only in their species. In all the following remarks, I have trusted exclusively to my own specific identification; and have rejected all assigned localities which appeared from any cause to be doubtful. Sessile cirripedes are found in every sea, from lat. 74° 18′ north to Cape Horn. The area included between the north point of the Philippine Archipelago and the south point of Australia, extending on the right hand to New Zealand, and on the left to Sumatra,—an area, which, though including two distinct Cirripedial regions, is small compared with the surface of the globe,[Pg 160] yet includes a greater number of species, especially of peculiar species, than the whole rest of the known world. This is, probably, in chief part due to the broken nature of the land, affording diversified habitats, and to much of the coast being rocky. Cirripedes, from requiring to be attached, cannot live where the shores and adjoining bottom are sandy or muddy or formed of moving shingle; hence, no doubt, it arises, that there is such a remarkable contrast in the great number of the species inhabiting the bold rocky western coast of South America, and the few species living on the sloping, and often sandy or muddy or shingly, eastern shores of this continent. Hence, also, I believe, it is that not many species have been brought from India. Coral-reefs are not favorable to Cirripedes, consequently but few are known to inhabit the islands of the Pacific Ocean. Where Cirripedes can live, though the species in no district are numerous, the individuals abound in infinite numbers: I have walked over the coast-rocks of the Falkland and Chonos Islands, of Chile and Van Diemen’s Land, fairly encrusted over wide spaces with a continuous layer of Cirripedes, consisting of only two or three species; in the same manner as may be observed on many parts of the shores of Great Britain, and, I believe, of North America.
With respect to the effects of temperature on the range of Cirripedes, no genus (having more than one species) is confined to the torrid zones. Pyrgoma, from being always attached to corals, is, of course, ordinarily found in the hotter seas; but one species ranges from the Cape de Verde Islands in the torrid zone to the southern shores of England and Ireland. Tetraclita is not found in the colder seas, but is numerous in species and in individuals, on the southern shores of Australia and at the Cape of Good Hope. I may here add, that the two genera with the most confined ranges, are Chamæsipho and Elminius; the former has only two species, one inhabiting Australia, and the other the East Indian Archipelago; Elminius has four species, confined to the southern hemisphere, and inhabiting Australia, New Zealand, and South America. To return to the effects of temperature; in Mr. Dana’s great work on Crustacea, an excellent chart is given, in which the isocrymal lines, or[Pg 161] those exhibiting the mean temperature of the waters along their course, for the coldest thirty consecutive days in any season of the year, are given; and which lines Mr. Dana has shown are the most influential in governing the distribution of marine animals. At the isocryme of 68°, Mr. Dana divides the torrid and sub-torrid zones from the several temperate zones; and at 44°, the temperate from the sub-frigid and frigid zones; but as no Cirripedes are exclusively confined to these frigid zones, we may here disregard them. From Mr. Dana’s[76] table of the areas of the torrid and temperate ocean-zones, on both sides of the equator, it seems that they are nearly as 337 to 278, in relative area; and consequently, he remarks, that the marine species in any class, if distributed equally over the two, would be one fifth more numerous in the torrid than in the temperate zones. Now of Cirripedes, taking all the orders, there are at present known 147 species; of these, 7 have doubtful habitats, leaving 140 for comparison. Of these 140, nearly one quarter, or 37, inhabit both the torrid and temperate zones, as above defined; 46 are found exclusively in the torrid, and 57 exclusively in the temperate zones; so that the temperate zones, though less in area, and having, proportionally, even a considerably lesser length of coastline, nevertheless have a preponderance in the number of species. But it should be borne in mind, that there are two great temperate districts, separated from each other by one great torrid district; and, inasmuch as the number of species in any region seems to depend in some degree on the isolation of the sub-regions, we might have expected (the other conditions now being, and the past conditions having been alike), that the two great temperate areas would have contained more species, perhaps doubly more, than the single great torrid area.
[76] ‘Crustacea: United States Exploring Expedition,’ p. 1476 (corrected).
The proportional numbers, above given, are not very widely different, whether we take separately the Balanidæ, the Lepadidæ, or all together. Mr. Dana has shown[77] at length, that generally amongst the Crustacea, the species which he considers of highest rank, belong to the extra-[Pg 162]torrid zones: there seems to me in all such cases to be some degree of vagueness in the attempt to determine which are highest or lowest, but I have already elsewhere stated that Balanus is, probably, the most eminently Cirripedial form, and exhibits in the strongest manner all the characters by which Cirripedes differ from other Crustacea; as this genus is the largest, containing 36 species, of which the habitats are known, I may state that of these, exactly one third, or 12, inhabit both zones; 9 exclusively inhabiting the torrid, and 15 exclusively the temperate zones. According to the proportions of the whole class, the numbers should have been 9 torrid, to only 11.11 temperate; so that evidently the genus Balanus (in one sense typical) inclines towards the temperate regions more strongly than does either the family or the sub-class to which it belongs.
[77] Ibid., p. 1528.
With respect to the relation between the size acquired by the different species of sessile cirripedes, and the temperature of the localities inhabited by them, the genera Chthamalus, Tetraclita, and Balanus, alone can serve for comparison: in Chthamalus much the largest species is found in the temperate zone: on the other hand, the two largest species of Tetraclita are from the torrid zone, though one of them also sometimes ranges into the temperate seas: in Balanus, the largest species, and six other species having a basal diameter sometimes over two inches, inhabit the temperate regions; and two out of these seven species, flourish even in the Arctic seas; whereas, within the torrid zone, there are only three species with a diameter sometimes exceeding two inches, but two of these frequently become very large and massive; so that Balanus, judging from the size of the species, as well as from their range, does not require for its highest development the temperature of the torrid zones.
The greater number of the species of the Balanidæ have wide ranges, as might be inferred from the fact of between one third and one fourth of the total number inhabiting both the torrid and temperate zones; but it should not be overlooked, that those species, as Balanus tintinnabulum, amphitrite, improvisus, and, in a lesser degree, B. trigonus[Pg 163] and Tetraclita radiata, which seem to range over nearly the whole world (excepting the colder seas), are species which are habitually attached to ships, and which could hardly fail to be widely transported. Indeed, it appears to me surprising, that such species as Balanus psittacus and eburneus, which often become attached to vessels, should still be confined, the one to Southern, and the other to Northern America. But some other Cirripedes, which I have never seen attached to vessels, have likewise immense ranges: thus Tetraclita porosa is found in every tropical and warmer sea, and Chthamalus stellatus ranges round the world in the northern hemisphere, and, along the eastern side of America, far south of the equator: Balanus spongicola, and Acasta spongites, extend from the shores of Britain to the Cape of Good Hope: Balanus lævis ranges from Tierra del Fuego to California. I may further remark, that the only two other species of Balanus, and the one Chthamalus, inhabiting Tierra del Fuego, are, also, found on the shores of Peru. But to show the powers of endurance in some species, I may specify the case of Balanus improvisus, which flourishes on the coast of Nova Scotia, amongst the West Indian Islands, in Southern Patagonia, and near Guayaquil. Even more striking is the case of B. crenatus, of which I have seen specimens from latitude 74° 48′ north, from the West Indies, and the Cape of Good Hope! In these two latter localities, however, it seems to be rare, and may have been first transported to them from the shores of Europe, on the bottoms of vessels, to which it sometimes adheres.
The several species of Balanidæ live attached either to coast-rocks, or to objects at various depths, down to, as in the case of Balanus crenatus, 50 fathoms. Balanus balanoides sometimes adheres to rocks or wood so high up, that it is not covered by water during the neap tides. Mr. Thompson has informed me, that he once accidentally kept some specimens of this species out of water for seven days in a warm room, and that they were then quite lively. This species, is very easily killed by brackish water, as are some other species, whilst B. improvisus and eburneus can flourish in it; and at the Falkland Islands, I saw Elminius Kingii attached to rocks at the mouth of a fresh-water[Pg 164] brook, so as to be covered by pure water during the ebb of each tide. Sessile cirripedes adhere to all sorts of objects, floating and fixed, animal and vegetable, living and dead, organic and inorganic. Chthamalus is, perhaps, more commonly attached to rocks than are the other genera. Living Mollusca are, I think, the most frequent objects of attachment: Mr. Cuming has remarked to me, that shells covered by an epidermis, as Patella, Haliotis, and Mytilus, are the greatest favorites. Acasta is always imbedded in sponges, or in the sponge-like bark of Isis; Pyrgoma and Creusia in corals; Chelonobia is attached to turtles, and one species to crabs or very smooth shells; Coronula, Tubicinella, and Xenobalanus, are imbedded in the skin of Cetaceans; and Platylepas in that of manatee, turtles, or sea-snakes.
If we attempt, with our present not very imperfect materials, to divide the globe into provinces, according to the amount of difference in their Cirripedial inhabitants, including all orders and families, and disregarding entirely, as I think we ought, all probabilities or conclusions deduced from the distribution of other tribes of animals, we find that the globe may be divided into the four following great provinces and one sub-province. I should premise, that in the following remarks and tables,[78] the species of Lepas, Conchoderma, Chelonobia, Coronula, Platylepas, and Tubicinella, are excluded, owing to their being attached to floating or swimming objects, and being consequently widely and irregularly distributed.
[78] As the number of Cirripedes in the whole class is not very great, I have given lists of the species in the four main provinces and in the one sub-province.
The first, or North Atlantic province, is that of Europe and the eastern shores of North America, from the arctic regions to lat. 30°: the island of Madeira, part of the north-west coast of Africa, and the whole Mediterranean being included. In this province (the above-named genera being excluded) we have 31 species, of which 22 are not found in any other distant quarter of the globe. As some few of these species range into the West Indies, I have not, on this account, excluded them from the 22 peculiar forms.[Pg 165] Had I included the West Indies[79] in my first province, the total number of species would have been 42, of which 28 would have been peculiar. The coast of Brazil, even as far south as the Rio Plata, might, also, have been included, for I have not seen from it a single species not included in the above 42 West Indian species. So also, by adding a single species, might the west coast of equatorial Africa. The two coasts of South America and Africa, which face each other within the torrid zone, seem to be remarkably barren in Cirripedes. Europe has several more species than the United States, which is inhabited by only ten species, including even the probably imported Balanus tintinnabulum and amphitrite. Of these ten United States species only two are not found in Europe; and both these two range into the West Indies, and as far as the northern shores of South America, and therefore cannot be considered as peculiar to the United States.
[79] The total number of species which I have seen from the West Indies, is 19 or 20; of these, only 6 are peculiar to it, or 8, if the United States be likewise included, the other 12 or 14 species being found in other quarters of the world. Six peculiar species out of 19 or 20, has not appeared to me sufficient to institute even a sub-province.
I have formed my single sub-province for the southern extremity of Africa; for although I know of only 11 species from this comparatively short and uniform line of coast, yet I was not able to group these eleven in any of the main provinces: 5 of the species are peculiar, 1 Australian, 3 European and West Indian, and 2 almost universally distributed.
The second province includes the west coast of North and South America, from Tierra del Fuego to Behring’s Straits: on this enormously long line of coast, only 22 species are known to exist, but of these no less than 15 are peculiar. Of these 15, four are not found south of the torrid equatorial region, and eight are not known to occur north of this same region; so that this long line of coast might have been divided into two sub-provinces, of which the southern would have been the most peculiar; but as eight species are found both north and south of the equatorial region, I have not made this sub-division. Two of the species occurring on the western coast of North America, seem to[Pg 166] represent species found on its eastern coast, and in Europe; thus, Balanus glandula takes the places of B. crenatus, and B. cariosus that of B. balanoides. Not a single species, excepting a few which are also widely distributed over other parts of the world, is known to be common to the east and west coasts of the two Americas.
The third province is that of the East Indian Archipelago, and includes the Philippines, Borneo, New Guinea, Sumatra, Java, Malacca, and the eastern coast of India. Here we find 37 species, of which 24 are peculiar. I may remark, that I have received no species from Madagascar or the eastern coast of Africa; few from India, or from the coast of China; and I suspect, that on most of these coasts, only few exist. Probably our third province will hereafter be found to include the whole Indian Ocean.
The fourth province is that of Australia, including New Zealand: it has 30 species, of which 21 are peculiar. Had the temperate Australian coasts (i. e., those south of the isocryme of 68°) been alone considered, the number of the species would have been probably 25, of which 20 would have been peculiar,—that is, if we admit within the 20, several species which range from the temperate into the torrid zone, but do not extend beyond the Australian shores. Owing to the widely-extended ranges of most Cirripedes, no Arctic or Antarctic provinces can be said to exist.
To recapitulate the above results, bearing in mind that, although the total number of known existing Cirripedes is 147, yet that the habitats of seven are unknown, and that eighteen are excluded owing to their being attached to floating or swimming objects, so that there are only 122 species referred to in the following table:
| Total number of species. | Species confined to the province. | |
|---|---|---|
| (1.) First, or North Atlantic Province, to lat. 30° N. (If the West Indies had been included the numbers would have been 42 and 28) | 31 | 22 |
| (2.) Sub-province of South Africa | 11 | 5 |
| (3.) Second province, or West Coast of North and South America | 22 | 15 |
| (4.) Third province, or East Indian Archipelago | 37 | 24 |
| (5.) Fourth, or Australian province | 30 | 21 |
The least prolific of these provinces contains 22 species, or between 1/5th and 1/6th of the total number of species, and the most prolific between 1/3rd and 1/4th of this same number. In each of these provinces, it is remarkable that the peculiar species are very nearly two thirds of the whole of its inhabitants. These facts, I think, show that the above provinces are natural divisions of the world, as far as their Cirripedial inhabitants are concerned.
As Cirripedes belong to the great class of Crustacea, and as the distribution of the latter has lately been fully discussed by Mr. Dana, it may be worth while briefly to compare my results with his; more especially as they are so very different. I should premise, as perhaps accounting to a certain extent for this difference, that, owing to the wide range of many species, and the almost universal extension of the same genera, my provinces are founded merely on a certain proportion of the species, namely, two thirds, being peculiar or confined to a region of considerable dimensions: whereas, in the case of ordinary Crustaceans, the greater number of the species are distinct even in the sub-provinces, and the provinces are founded mainly on generic differences. Mr. Dana divides the surface of the globe into three great sections, or provinces, the Africo-Europæan, the extent of which is shown by its title; the Occidental, which includes both the east and west coast of both Americas; and the Oriental, including the Indian and Pacific Oceans, with the East Indian Archipelago, and Australia. Thus Mr. Dana entirely separates the Eastern shores of North America from Europe; whereas, as far as their Cirripedial inhabitants are concerned, they are most intimately allied, and form my first or North Atlantic province; and to this, as I have shown, even the West Indies, the coast of Brazil, and equatorial West Africa might have been added. It follows, from this similarity in the Cirripedes on the two sides of the Atlantic, and from their dissimilarity with those on the shores of the Pacific, that the east and west coasts of the two Americas form two quite distinct Cirripedial provinces; though, in the northern half, some connection is shown by a few representative species: on the other hand, Mr. Dana unites both sides of the whole American[Pg 168] continent, into his single Occidental province. The South-African province is not brought out by Mr. Dana so prominently, as I have found necessary. Mr. Dana joins the East Indian Archipelago and Australia into his single Oriental province, and makes New Zealand, as a sub-province, apparently as distinct from Australia, as Australia is from the East Indian Archipelago: whereas I find that the Cirripedes of New Zealand clearly belong to Australia; and that the Australian Cirripedes, especially if the temperate shores be alone considered, are as distinct from those of the East Indian Archipelago, as from those of any other quarter of the whole world. I believe that the provinces deduced from the distribution of Cirripedes, accord better with the Molluscan provinces, than with those given by Mr. Dana for the rest of the great class of Crustaceans.
In the following tables, an asterisk means that the species is not found in any other distinct region of the globe. When found in one of the five provinces, a corresponding number, within brackets, is appended, to show in which province or sub-province it has been found.
(1.) FIRST OR NORTH ATLANTIC PROVINCE: Europe and the Eastern United States, from the Arctic Regions to 30° north latitude.
| Balanus tintinnabulum | (1 to 5). |
| tulipiformis* | confined to Europe. |
| calceolus | Europe and India. |
| galeatus* | confined to North America and West Indies. |
| spongicola | (2) and West Indies (?). |
| perforatus* | confined to Europe, but possibly in the West Indies. |
| amphitrite | (1 to 5). |
| eburneus* | confined to North America and West Indies. |
| improvisus | Europe and North America and (3). |
| porcatus* | Europe and North America. |
| crenatus | ””W. Indies and (2). |
| balanoides* | ”” |
| Hameri* | ”” |
| Acasta spongites | Europe and (2). |
| cyathus* | Madeira and West Indies. |
| Pyrgoma anglicum* | confined to Europe, but ranges as far at least as the Cape de Verde Islands. |
| [Pg 169]Xenobalanus globicipitis* | confined to Europe. |
| Chthamalus stellatus | Europe and North America and (3 and 4). |
| Pachylasma giganteum* | confined to Europe. |
| Verruca Strömia | Europe and Red Sea. |
| Spengleri* | Madeira. |
| Pœcilasma aurantia* | ” |
| crassa* | ” |
| Dichelaspis Lowei* | ” |
| Oxynaspis celata* | ” |
| Alepas minuta* | Europe. |
| parasita* | Europe and Atlantic Ocean. |
| Anelasma squalicola* | Europe. |
| Alcippe lampas* | ” |
| Scalpellum vulgare* | ” |
| Pollicipes cornucopia* | ” |
Here we have 31 species, of which 22 are not found in any other great region of the world.
(2.) SUB-PROVINCE: Africa, South of lat. 30°.
| Balanus tintinnabulum | (1 to 5). |
| Capensis* | |
| spongicola | (1) and West Indies (?). |
| amphitrite | (1 to 5). |
| crenatus | (1) and West Indies. |
| Acasta spongites | (1). |
| Tetraclita serrata* | |
| rosea | (4). |
| Chthamalus dentatus* | also on West Coast of Africa. |
| Octomeris angulosa* | |
| Scalpellum ornatum* |
In this small region we have only 11 species, of which five are peculiar: Balanus Capensis and Tetraclita serrata, seem to be representatives of B. psittacus of S. America and of T. porosa of that and several other regions.
(3.) SECOND PROVINCE: West Coast of South and North America, from Tierra del Fuego to Behring’s Straits.
| Balanus tintinnabulum | north and south of the equator (1 to 5). |
| psittacus* | south. |
| vinaceus* | ” |
| trigonus | north and south (4 and 5). |
| lævis* | ” |
| concavus | ”(4 and 5). |
| pœcilus* | south. |
| improvisus | south (and north?) (1). |
| [Pg 170]nubilus* | north. |
| glandula | north, and Southern Pacific Ocean. |
| cariosus* | north. |
| Tetraclita porosa | north and south (4 and 5 and W. Indies). |
| Elminius Kingii* | south. |
| Chthamalus stellatus | north. |
| cirratus* | south. |
| scabrosus* | ” |
| fissus* | north and south. |
| Hembeli* | north. |
| Verruca lævigata* | south. |
| Pollicipes elegans* | south and north. |
| polymerus* | ” |
| Cryptophialus minutus* | south. |
Here we have on this long line of coast, 22 species, of which 15 are peculiar.
(4.) THIRD PROVINCE: Indian Archipelago (including the Philippines, Malacca, Borneo, Sumatra, Java, and New Guinea, and eastern coast of India).
| Balanus tintinnabulum | (1 to 5). |
| Ajax* | |
| navicula* | |
| stultus | and West Indies. |
| trigonus | (3 and 5). |
| concavus | (3 and 5). |
| amphitrite | (1 to 5). |
| patellaris* | |
| amaryllis | (5). |
| quadrivittatus* | |
| Acasta lævigata | and Red Sea. |
| fenestrata* | |
| purpurata* | |
| sporillus* | |
| Tetraclita porosa | (3 and 5) and West Indies. |
| costata* | |
| vitiata | (5). |
| cœrulescens | Pacific Ocean. |
| radiata | (5) and West Indies. |
| Pyrgoma cancellatum* | |
| grande* | |
| milleporæ* | |
| crenatum* | |
| monticulariæ* | |
| Creusia spinulosa | and West Indies. |
| Chthamalus stellatus | (1 and 3). |
| intertextus* | |
| Chamæsipho scutelliformis* | |
| Octomeris brunnea | |
| [Pg 171]Pœcilasma fissa* | |
| eburnea* | |
| Dichelaspis Warwickii* | |
| Ibla Cumingii* | |
| Scalpellum rostratum* | |
| Pollicipes mitella* | |
| Lithotrya Nicobarica* | |
| truncata | and Pacific Ocean. |
Here we have 37 species, of which 24 are peculiar to this province.
(5.) FOURTH PROVINCE: Australia (including New Zealand).
| Balanus tintinnabulum | (1 to 5). |
| nigrescens* | |
| decorus* | |
| trigonus | (3 and 4). |
| concavus | (3 and 4). |
| amphitrite | (1 to 5). |
| amaryllis | (4). |
| allium* | |
| vestitus* | |
| imperator* | |
| Acasta sulcata* | |
| glans* | |
| Tetraclita porosa | (3 and 4). |
| rosea | (2). |
| purpurascens* | |
| vitiata | (4). |
| radiata | (4) and West Indies. |
| Elminius plicatus* | |
| simplex* | |
| modestus* | |
| Chthamalus antennatus* | |
| Chamæsipho columna* | and Pacific Ocean (?). |
| Pachylasma aurantiacum* | |
| Catophragmus polymerus* | |
| Alepas tubulosa* | |
| Ibla quadrivalvis* | |
| Scalpellum Peronii* | |
| Pollicipes spinosus* | |
| sertus* | |
| Lithotrya cauta* |
Here we have 30 species, of which 21 are peculiar.
The ancient history of the Balanidæ is a brief one. No secondary species has hitherto been discovered; in my monograph on the fossil Lepadidæ[80] I have shown that the negative evidence in this case is of considerable value, and consequently that there is much reason to doubt whether any member of the family did exist before the Eocene period. The existence of a Cretaceous Verruca is an apparent exception to the rule, as this genus has hitherto always been ranked amongst sessile cirripedes; but Verruca, as we now know, must be placed in a family by itself, quite distinct from the Balanidæ. Balanus is the oldest genus as yet known; it first appeared in Europe and North America, during the deposition of the eocene beds; and was at that time, as far as our information at present serves, represented by very few species. In South America, one species of Balanus abounds in individuals in the ancient Patagonian tertiary formation. I have seen, in the British Museum, specimens, said to have come from the eocene nummulitic beds near the mouth of the Indus, belonging to the second section of the genus. Generally, the extinct forms belong to the last section of this genus, which has the parietes not permeated by pores. During the miocene and pliocene ages, sessile cirripedes abounded. No extinct genus in this family has hitherto been discovered. It is singular, that though the Chthamalinæ approach much more closely than do the Balaninæ to the ancient Lepadidæ, of which so many species have been found fossil even in the older Secondary formations, yet that only one species of one genus of this sub-family has been hitherto found in any deposit; and that species is the still existing Pachylasma giganteum, in the modern beds of Sicily. During the epoch of the Glacial deposits in Scandinavia, Scotland, and Canada, the still existing species seem to have abounded; and they attained larger average dimen[Pg 173]sions than the same species now do on the shores of Great Britain, or even on the shores of the northern United States, where the average size seems larger than on this side of the Atlantic.
[80] Since the note to page 5 of that work was written, I have been informed that the so-called cretaceous Tubicinella maxima is not a Cirripede.
Under the genus Balanus, I have given my reasons for never naming species in this large and difficult genus, without examining the opercular valves: it has been owing to this, as it appears to me, proper want of caution, that there are so many nominal species. Thus it is made to appear in catalogues, that the tertiary seas abounded with species of Balanus to an extent now quite unparalleled in any quarter of the world. Bronn,[81] for instance, in his invaluable ‘Index Palæontologicus,’ gives the names of 35 Balani, found fossil in Europe, and I have not counted those found only in alluvial deposits, as they would certainly be the same as the still living species. Now I know only 11 recent Balani on the shores of all Europe, from the North Pole to lat. 30°; and of these I doubt whether B. balanoides and improvisus have been found fossil. In the Red Crag there is one extinct Balanus: in the Coralline Crag, which seems to have been very favorable to the existence of Cirripedes, there are six species of Balani, of which two are absolutely extinct, and one does not occur in any neighbouring sea: in the Eocene formations the species seem to have been rare, and I have seen only one, and that is an extinct form. Taking these several facts into consideration, and bearing in mind that Cirripedes usually range widely, I do not believe, if all the specimens of Balani hitherto found in the several tertiary formations, from the eocene to the glacial deposits, throughout Europe, were collected together, they would amount to 20 species. I have myself seen, in a recognisable state, only 12 fossil species, of which five are extinct, or not found in any neighbouring sea: I think it probable that three other recent species, viz. B. tulipiformis, perforatus, and amphitrite, may occur in the Mediterranean formations; and this would make 15 species. Therefore in the above estimate of 20 species, five are allowed for[Pg 174] species existing in European collections, but not hitherto seen by me; and this, I believe, is a very full allowance. Consequently, even on the supposition that the five species just admitted as possibly existing in cabinets, and that the other five extinct species, which I have seen and examined, have all been previously named by other authors, a supposition excessively improbable, even then there would be 15 superfluous names in Bronn.
[81] To save any other person, interested in fossil Cirripedia, going through the several works quoted by Bronn, I have given some remarks on his list of species, in an appendix at the end of the Balanidæ.
The following short table shows how Cirripedes, including all three Families, were represented in Great Britain, throughout the several TERTIARY STAGES.
| Name. | Living species but found fossil in some tertiary deposit. | Mammilliferous crag, and glacial deposits. | Red crag. | Coralline crag. | Eocene. |
|---|---|---|---|---|---|
| Balanus tintinnabulum | * | * | |||
| calceolus | * | * | |||
| spongicola | * | * | |||
| concavus | * | * | * | ||
| porcatus | * | * | * | ||
| crenatus | * | * | * | * | |
| Hameri | * | * | * | ||
| bisulcatus | * | * | |||
| dolosus | * | ||||
| inclusus | * | ||||
| unguiformis | * | ||||
| Acasta undulata | * | ||||
| Pyrgoma anglicum | * | * | |||
| Coronula barbara | * | ||||
| Verruca Strömia | * | * | * | * | |
| Scalpellum magnum | * | ||||
| quadratum | * | ||||
| Pollicipes reflexus | * | ||||
| Total, 18, recent and extinct, found fossil in Great Britain, in some tertiary deposit | 9 | 5 | 8 | 10 | 3 |
As affording some standard of comparison by which to compare the number of fossil species, at any period, in relation to the number of species probably existing in the neighbouring seas during the same epoch, I may state that there are now living and propagating on the shores of Great Britain, 18 species belonging to the three Families[Pg 175] included in the above table. I have not counted three species, in the genera Alcippe and Conchoderma, which, from the minuteness of their valves, it is hardly possible would be found fossil. On the other hand, I have included in the 18, five species of Lepas, which from floating and being oceanic, are more likely to be cast up on beaches, than to be imbedded in sedimentary deposits; so that 13 would, perhaps, be a safer number, as a standard of comparison. Now in the coralline crag, which seems to have been eminently favorable for the existence and subsequent preservation of Cirripedes, and which has been so well worked, only nine fossil species, as may be seen in the table, have been as yet discovered.
Shell with the rostrum having radii, but without alæ; lateral compartments all having alæ on one side and radii on the other side; parietes generally either porose, or longitudinally ribbed on their inner surfaces.
Mouth with the labrum notched in the middle, not swollen; palpi large, almost touching each other; mandibles generally with the lower teeth laterally double; third pair of cirri with their segments resembling those of the second pair.
Scutum and tergum articulated together, or overlapping each other; each branchia composed of a single plicated fold.
Genera—Balanus; Acasta; Elminius; Tetraclita; Pyrgoma; Creusia; Chelonobia.
Scutum and tergum (when both are present) not overlapping each other; basis membranous; parietes often deeply folded, with the outer lamina, towards the basis, generally[Pg 176] imperfect; each branchia composed of two plicated folds; shell attached to living vertebrata.
Genera—Coronula; Platylepas; Tubicinella; Xenobalanus.[82]
[82] At the end of the volume a Synopsis is given, which will serve as a systematic index for the discovery of generic and specific names.
The Balanidæ may be divided into two sub-families; namely, the Balaninæ and Chthamalinæ; and, in the former, the genera, as we see, may be very naturally grouped into two sections. The Balaninæ differ from the Chthamalinæ, as far as the shell is concerned, in the rostrum having radii but no alæ, all the lateral compartments having both radii and alæ; on the other hand, in the Chthamalinæ, the rostrum has alæ, and the rostro-lateral compartments radii on both sides, and therefore no alæ. These differences probably arise, as already explained, from the perfect confluence, in the Balaninæ, of the true rostrum with the rostro-lateral compartments. In Chelonobia, belonging to the Balaninæ, we see an intermediate state, with the fusion not quite effected: on the other hand, in one genus amongst the Chthamalinæ, namely, Pachylasma, we must look to the shell at a very early age, to find the rostrum with its alæ, distinct from the rostro-lateral compartments. In Tetraclita, Elminius, and Creusia, the carino-lateral compartments are aborted, or possibly confluent with the lateral compartments, making altogether only four: in Pyrgoma all the compartments are fused together and form a solid ring. The sub-genus Acasta is, in one sense, very natural, as it includes species most closely allied: in another sense it is far from natural, as some of the species can hardly be distinguished from those species of Balanus, which live attached to Gorgoniæ: I almost regret I did not merge the species of Acasta into Balanus. In the Balaninæ generally the parietes are either porose, or are furnished on their internal surfaces with regular ribs, representing the longitudinal parietal septa, which in other species form the tubes or pores; there are, however, many exceptions to this rule in several species of Balanus, in Acasta and Elminius,[Pg 177] all of which have the parietes of their shells internally quite smooth, or only irregularly roughened with points.
Looking to the animal’s body, in the Balaninæ, the labrum is always notched in the middle, and is never swollen or bullate, for the outer and inner folds of membrane of which it is composed lie close together. The palpi are large, so that their tips almost touch each other. The mandibles, generally, have their lower main teeth laterally double. Of the cirri, the third pair invariably much more closely resembles, in its whole structure, and in its action, the second than the fourth pairs; and it is also generally separated by a small interval from the fourth pair.
I have already under the Family sufficiently entered on the relations of the Balaninæ to the Chthamalinæ, and of the genera, one to the other, so that I need not here add anything.
I can point out no difference in habits or geographical distribution between the Balaninæ and Chthamalinæ.
[83] The name Balanus was used, almost as at present, by Lister and Hill, before the introduction of the binomial system. Since that period the first two authors, as far as I know, who used this name, were Da Costa, in his ‘Hist. Nat. Test. Brit.,’ in 1778; and Bock, in the ‘Naturforscher,’ for the same year; Bock, however, applied it to a Chelonobia.
CONOPEA (pars generis). Say. Journal Nat. Sc. Philadelphia, vol. ii, part ii, 1822.
MESSULA (do.). Leach. Zoological Journal, vol. ii, 1825.
CHIRONA (do.). J. E. Gray. Philosoph. Transacts., 1835, p. 37.
Compartments six; basis calcareous or membranous; opercular valves sub-triangular.
Distribution.—Mundane: in the warmer seas.
General appearance.—The shape of the shell in the different species varies from depressed conical to cylindrical;[Pg 178] the latter form being generally assumed when specimens are crowded together; but some species, as B. balanoides, crenatus, and lævis, seem more subject than others to be thus affected. The colour is either white, generally tinted by the yellowish or brownish epidermis, or any colour intermediate between bright pink and rich blue, purple being the prevailing tint. The persistence of the so-called epidermis is very different in different species, being even sometimes highly variable in the same species. The surface is either smooth or more commonly folded longitudinally, or sharply ribbed. The orifice differs in form from diamond-shape to trigonal; the carinal end, owing to the shape of the carina, being always sharper or narrower than the rostral end. The size of the orifice, in proportion to the shell, varies accordingly as the latter is more or less conical or cylindrical. The orifice is either entire or more or less deeply toothed, in proportion to the degree of obliquity of the summits of the radii and alæ. The radii almost always have smoother surfaces than the parietes. In some few species the radii are not developed, the sutures being marked only by fissure-like lines; in others they are very narrow, and in this case their upper margins are generally rounded and smooth, instead of being straight and jagged. The carino-lateral compartments are usually much narrower than the lateral compartments, occasionally in an extreme degree, as in B. allium. The shell is generally strong, sometimes to a wonderful degree; but the strength and thickness vary in the individuals of some of the species. By the action of hot caustic potash, the compartments in several species, such as B. Hameri and crenatus, separate on a touch; in others, they adhere so strongly as to prove that the sutures must be calcified together. In this genus we have the largest known sessile cirripede, viz., the B. psittacus, and on the other hand many small species; but it is very difficult, except in well-known species, to ascertain the average or even the maximum dimensions.
Scutum.—This valve is almost triangular, with the basi-tergal corner more or less rounded off. The prominent lines of growth are sometimes crossed by longitudinal striæ. Internally, the articular ridge projects to a very different[Pg 179] degree in the different species; its lower end is sometimes (as in B. lævis, Pl. 4, fig. 2 c) produced downwards as a small, sharp, free style; there is always an articular furrow receiving the inflected margin of the tergum. There is always an impression left by the attachment of the adductor scutorum muscle; and often its lower side is bounded more or less closely by a sharp adductor ridge, running some way down the valve; this ridge is occasionally almost confluent, in its upper part, with the articular ridge, and in this case sometimes it forms, together with the inflected tergal margin, a large tubular cavity, running up, as in B. psittacus (Pl. 2, fig. 3 c), almost to the apex of the valve. Almost invariably there is a slight pit or depression for the lateral depressor muscle; sometimes within the depression there is a little ridge, as in B. perforatus and nubilus (Pl. 4, fig. 3 a, and Pl. 6, fig. 2 a); and in the case of B. vestitus, flosculus, and imperator (Pl. 8, figs. 3 a, 4 a), there are regular crests for this same purpose. The rostral depressor muscle is usually attached in a small pit formed by the folding over of the lower part of the occludent margin: in B. imperator (Pl. 8, fig. 4 a) there are regular crests for its attachment, and traces of them may be discovered in B. vestitus.
Tergum.—This valve is more nearly triangular than any other shape, with the spur more or less prominent. The apex generally projects a little above the level of the scutum; in some species it consists of a triangular and solid, in others (Pl. 2, fig. 3 b) of an almost cylindrical, extremely sharp, inwardly curved, and very prominent beak. This beak is generally purple; it is sometimes hollow, and occupied by a thread of corium. Its formation, and the apparent sliding up of the whole tergum, so as to project above the scutum, has been described under the family. From an account given to me by a person who kept B. porcatus alive, the beaks appear to be used, when the operculum is touched, as an organ of defence,—the animal striking with them. The tergal margin is more or less inflected; and the carinal margin is convex in different degrees, and, in some species, is added to by upturned zones of growth. The basal margin either forms a nearly straight line on opposite sides of the spur, or more commonly slopes towards it in various[Pg 180] manners. The spur, or basal projection, is rarely placed in the middle of the basal margin, generally near, sometimes close to the basi-scutal angle; it varies much in length and breadth, and is sometimes even half the width of the valve. The surface of the valve is almost always more or less depressed, sometimes so much as to form a deepish furrow, the “longitudinal furrow,” which extends from the apex to the extremity of the spur. When the furrow is deep, its sides, as the specimen grows old, almost always become folded inwards, so as to touch, and then the furrow becomes converted into a closed fissure: in this latter case the folded sides generally form a central crest on the spur. Internally, in the middle of the upper part of the valve, the articular ridge is more or less prominent, forming the carinal margin of the articular furrow, in which the articular ridge of the scutum is lodged; occasionally, however, this articular ridge can hardly be said to exist. In most species the tergal depressor muscle is attached to sharp crests on the basi-carinal corner of the valve, but these are almost obliterated in other species.
Compartments.—The external appearance of the shell has already been described. In the most typical species, the parietes consist of an outer and inner lamina, separated by strong longitudinal septa; these septa are denticulated on both sides at their bases, but only close to the inner lamina; in fact the inner lamina is apparently formed by the union, thickening, and production, of some of the denticuli. As it is not the innermost of the denticuli on the basal edges of the longitudinal septa, which thus become united into a solid layer, the longitudinal septa form slightly projecting, longitudinal ribs on the inner lamina. These internal ribs are longitudinally striated; in old specimens they often become obliterated, especially in the upper part of the shell. The parietal tubes or pores (occupied by threads of corium) are generally square and large; but in B. Ajax they are very small, and in B. glandula often extremely minute. In the upper part of the shell, and sometimes low down, they are generally crossed by thin, transverse, calcareous septa: in some species, as in B. perforatus, and in some varieties of B. amphitrite, the upper ends of the[Pg 181] tubes are filled up solidly with shell. In some varieties of B. crenatus and of amphitrite, the longitudinal septa, near the outer lamina, divide, thus giving rise to a very imperfect row of outer short tubes. In B. vinaceus (Pl. 2, fig. 7 d) the inner lamina is cancellated instead of being solid, which is caused by the basal denticuli of the longitudinal septa being simply united together by their ends and crossed by transverse septa, instead of being consolidated into a mass. In several species, as in B. Hameri, the walls consist only of the outer lamina with longitudinal ribs, no inner lamina having been formed; the ribs here evidently answer to the longitudinal septa in the foregoing species. In B. flosculus and imperator the walls are solid, their basal margins being formed of irregular, elongated points, and little ridges (Pl. 8, fig. 4 c), which apparently prefigure the more regular longitudinal ribs or septa. In B. balanoides the walls are generally either nearly smooth and solid, or irregularly cancellated; in B. cariosus (Pl. 7, fig. 3 b) two or three rows of short irregular tubes are formed by unequally branching septa, almost as in the genus Tetraclita.
The Radii, in all the species, are constructed essentially on the same plan as the parietes; thus, in the typical forms, there is an outer and inner lamina, with septa, which, near the inner lamina, are furnished with denticuli on both sides; hence the radii are permeated by pores or tubes, like the parietes; but this holds good only in the first section of the genus, for, in the other species, the tubes are filled up quite solidly. The denticuli on the septa often occur only on one side, or disappear altogether; and, lastly, the septa themselves often appear merely like little teeth, or disappear altogether as in B. Hameri, or occur only near the bases of the radii, as in B. amaryllis. A slight furrow in the compartment, against which each radius abuts, is generally marked by the septa and their denticuli. In regard to the alæ, their lateral or sutural edges are either thin and smooth, or, more commonly, finely crenated or ribbed. The little transverse crenations are homologous with the septa in the radii and parietes. The edges of the alæ are usually received in a furrow. The diametric growth of the shell is[Pg 182] effected by the growth of the radii and alæ, and chiefly by that of the former. The sutural and lateral edges of both radii and alæ are added to, either quite up to their summits, or only low down, and during the continued growth of the shell, lower and lower down; in accordance with this difference in growth, the summits of the radii and alæ become either very oblique, or they extend parallel to the basis, that is, from tip to tip of the adjoining compartments. When the radii and alæ are added to, as is most usual, above the level of the opercular membrane, and therefore above the sack, ribbons of corium run up the sutures from the sack, higher or lower, according to the height to which, in the different species, the edges of the radii and alæ continue to be added to. The obliquity of the summits of the radii and alæ varies, in some cases, in the same species. It often happens that when the summits of the radii are very oblique, the summits of the alæ are but little so; and the converse; both, however, are often either equally oblique, or both have square summits. The sheath extends either one third or more than half down the shell; its basal margin often (Pl. 25, fig. 1, K′) freely depends or overhangs the inner lamina of the walls.
Basis.—In typical species the basis is calcareous, and consists of an upper and lower lamina, separated by radiating septa, forming pores. In the same manner as the septa of the parietes sometimes, though rarely, become irregularly divided near the outer lamina, forming outer pores, so it is, but in a much more marked degree, with the basis. The basis in such cases becomes extremely thick, and consists of an upper, thin lamina, with the regular radiating septa and pores, and of an underlying, thick, cancellated mass, which seems wholly to result from the dividing and sub-dividing of the septa. The basal radiating pores, like the parietal pores, are closed at intervals by calcareous transverse septa. The basal points of the parietal septa enter the orifices of the basal pores, and the threads of corium pass into the latter, between the denticuli of the parietal septa. In some species, as in B. crenatus and Hameri, the basis is perfectly solid, the upper lamina being absent, just as in some species, the internal lamina of the[Pg 183] parietes is absent. In B. flosculus the basis is calcareous, but consists of so excessively thin a film as hardly to be distinguished: it presents, moreover, as also is the case with B. imperator, a beaded structure. Again, in some few species, as in B. balanoides, the basis is simply membranous. When the basis is thin, it is always flat, and is closely moulded to the irregular surface of attachment; and in this case, when specimens are crowded together, their elongation is effected exclusively by the growth of the walls; but, when the basis is thick, it sometimes becomes, in crowded groups, deeply, but irregularly, cup-formed, or cylindrical, as in B. psittacus and perforatus. In B. allium, however, which inhabits massive corals, the basis is as regularly concave or cup-formed as in the genus Pyrgoma: in B. calceolus and its allies, and in some varieties of the fossil B. inclusus, the basis is boat-formed, with its lower surface deeply grooved longitudinally from clasping the stem of the Gorgonia or other zoophyte, to which it was attached. In certain varieties of B. lævis it is very remarkable that the deeply cup-formed basis becomes, owing apparently to the whole shell having grown too deep for the animal, half-filled up with irregular, calcareous, transverse plates (Pl. 4, fig. 2 a), resting one upon the other by irregular points or pillars. The cementing apparatus has been sufficiently described under the Family.
Mouth.—The labrum is always notched; sometimes it has no teeth, but generally there are three on each side; in B. balanoides there are five or six on each side; and in B. improvisus and eburneus there is a whole row of teeth (Pl. 26, fig. 2, e′), graduated in size, on each side of the notch. The palpi are large, with their apices nearly touching, and furnished with long spines. The mandibles have, as it appears, normally, five teeth, but the two lower teeth are always small and often rudimentary, and almost confluent with the inferior, sometimes spinose angle. The maxillæ have either a simple edge, or a notch under the pair of large upper spines, or the lower part forms (Pl. 26, fig. 7) a step-formed projection: there are generally two lower spines, placed singly or not in pairs, larger than the[Pg 184] others, with the exception of the uppermost pair. The outer maxillæ are, on their inner faces, obscurely divided into two lobes.
Cirri.—The rami of the first pair are unequal, the shorter one sometimes not being more than half the length of the other ramus: the segments of the shorter ramus are broad, and are, together with the lower segments of the longer ramus, thickly clothed with spines; in some species, as in B. perforatus, the anterior surfaces of the segments, more especially of the shorter ramus, and of both rami of the second pair are produced (Pl. 29, fig. 4), so as sometimes to form very remarkable projections. The segments of the second and third pairs are always thickly clothed with spines, as also are their pedicels. The third pair is rather longer than the second; but in B. vestitus and imperator it is much longer, and is otherwise somewhat different. The dorsal and basal margin of the pedicel of the third pair, in some of the species, as in B. tintinnabulum, is produced backwards on the thorax, and forms a membranous plate fringed with fine spines. The three posterior and longer pairs of cirri have from three to rarely eight or ten pairs of long spines on each segment, with generally one or two minute spines in the middle between each pair: their pedicels have a regular double row of spines.
The penis is long and hairy: in most of the species there is, at its dorsal basis, a small, sharp, flattened, imperforate projection; first observed by Poli: but this is sometimes absent, as in B. crenatus, though present in the closely allied B. balanoides; and its presence is variable in B. tintinnabulum. All the species have large plicated branchiæ. The base of the sack in several species is furnished with inwardly projecting filamentary appendages. In B. perforatus, crenatus, and improvisus, and I believe in other species, the upper part of the stomach is furnished with a circle of branching cæca.
On the variation of the species; their arrangement and affinities; value of the characters used; changes during growth.—Owing to the great variation in external characters, to which almost all the species are subject, and likewise to[Pg 185] the genus being a very natural one, that is, to the species following each other in close and natural order, it is not easy to exaggerate the difficulty of identifying the species, except by a deliberate examination of the internal and external structure of each individual specimen. Every one who has collected sessile cirripedes must have perceived to what an extent their shape depends on their position and grouping. The surface of attachment has a great effect on that of the shell; for as the walls are added to at their bases, every portion has at one time been in close contact with the supporting surface; thus I have seen a strongly-ribbed species (B. porcatus) and a nearly smooth species (B. crenatus) closely resembling each other, and both having a peculiar appearance, owing to their having been attached to a pecten. Dr. Gray has pointed out to me specimens of B. patellaris, curiously pitted like the wood to which they had adhered; and numberless other instances might be added. Quite independently of the effect produced by the surface of attachment, the degree to which the longitudinal folds and ribs are developed on the parietes, is variable in most of the species, as in B. tintinnabulum, vestitus, and even in B. porcatus; the presence or entire absence of these ribs often surprisingly alters the whole aspect of the shell. The persistence of the so-called epidermis is in some degree variable; and in B. lævis we have groups of specimens absolutely naked, and others uniformly clothed with a brown membrane. Again, some species in certain localities are all subject to the disintegration of the entire outer lamina of the walls; and in such cases (as with B. perforatus) there is not the smallest resemblance between the corroded and perfect specimens. The size of the orifice, and consequently of the operculum, compared with the shell itself, varies accordingly as the shell is more or less conical or cylindrical; in the latter case, the summits of the radii are generally more oblique and the aperture consequently more deeply toothed than in the more conical varieties. Size is a serviceable character in some cases, but very many specimens are required to ascertain the average or maximum size of each species, for there is no method of distinguishing a half-grown from a[Pg 186] full-grown specimen; and I believe, as long as the individual lives, so long does it go on moulting and growing. Colour is of very considerable service; though the precise tint varies greatly in almost every species; and what is a far more serious evil, the majority of the species have their white or nearly white varieties, the latter sometimes as numerous as the coloured ones: in B. perforatus, lævis, flosculus, amphitrite, and in several other species, the common white varieties are eminently deceptive.
Besides the slight variation in the obliquity of the summits of the radii and alæ, dependent on the more or less cylindrical form of the shell, in some species, as in B. tintinnabulum, amphitrite, improvisus, trigonus, and porcatus, their obliquity also varies occasionally from unknown causes, and thus greatly affects the general appearance of the shell. In some few species, as in B. perforatus, the radii are often either not at all developed, or are of very variable width; in others, when the shell has become cylindrical, or when very old, the radii cease to grow, and from the disintegration of the whole upper part of the shell, with the continued growth of the lower part, the radii at last come to exist as mere fissures: I have seen instances of this in B. psittacus, nigrescens, and porcatus. Nevertheless, the obliquity of the upper margin, and the breadth of the radii are useful characters; and still more useful is the fact whether the upper margins are smooth and arched, or straight and jagged. The fact of the terga being more or less beaked is useful: as is, likewise, the presence of striæ, or furrows, or rows of pits, radiating from the apices of the scuta; but to ascertain the presence of these marks, it is almost invariably necessary to take out the scuta, clean, and examine them with a lens; these ridges and furrows, moreover, in some species, as is strikingly the case with B. tintinnabulum, and in less degree with B. trigonus, lævis, and concavus, appear and disappear, and vary without any apparent cause.
Now if we reflect that form, size, state and nature of the surface, presence of epidermis, relative size of the orifice, presence of longitudinal ribs, tint, and often the existence of any colour, are all highly variable in most of the species; and that the obliquity of the summits of the[Pg 187] radii, and the presence of longitudinal striæ on the scuta, are variable in several species; we shall perceive how difficult it must ever be to distinguish the species from external characters. As some evidence of this, I may mention that, after having described nearly 40 species, and when my eye was naturally able to appreciate small differences, I began carefully to examine varieties of B. tintinnabulum, amphitrite, improvisus, porcatus, vestitus, &c., without even a suspicion that they belonged to these species, at that time thoroughly well known to me; yet in the cases here referred to, there could be no doubt, when a perfect series was examined, that the specimens were only varieties. From this cause the labour of naming a collection is great. Let no one attempt to identify the species of this genus, without being prepared to disarticulate, clean, and carefully examine with a microscope the basis and parietes, and both the under and upper surfaces of the opercular valves; for I feel convinced, that he will otherwise throw away much labour. Moreover, in many cases, it is almost necessary, on account of the variability of the characters, to possess several specimens. From these facts, I have not hesitated to form my sections on characters which require close examination, though I would gladly have seized on external characters, could I have found such even moderately constant.
The least varying, and therefore most important characters, must be taken from the internal structure of the parietes, radii, and basis: not that these characters are absolutely invariable; thus the porosity of the parietes is slightly variable in B. glandula, and highly variable in the fossil B. unguiformis; it is also highly variable in B. balanoides, but under a systematic point of view this is unimportant, as the section including this latter species is well defined by the membranous basis. The porosity of the basis is in some degree variable in B. nubilus, improvisus, and patellaris; and in B. flosculus we see signs of a passage from a calcareous to a membranous basis. Characters derived from the general shape, and from the ridges and depressions on the under side of the scuta and terga, especially of the scuta, are highly serviceable. The terga, indeed, in many species, as in B. amphitrite, vary con[Pg 188]siderably, and are affected by the general shape of the shell. Unfortunately the differences are not very great between the scuta of the different species. The cause of the opercular valves offering more useful characters, as far as outline is concerned, than do the walls of the shell, is no doubt due to their being almost independent of any influence from the nature of the surface of attachment. Even the ridges and depressions on the under side of the scuta, which are in direct connection with the muscles and soft parts of the animal, vary to a certain extent: thus the length and prominence of the adductor ridge is decidedly variable in B. concavus and tintinnabulum, and in a less degree in B. lævis; the size and form of the little cavity for the lateral depressor muscle varies in many species; so does the exact shape and degree of prominence of the articular ridge. There is one character in the terga, which at first would be thought very useful, namely, whether an open longitudinal furrow, or a closed fissure runs down the valve from the apex to the spur; but it is found that the furrow almost always gradually closes up during growth; and as a consequence of this, the width of the spur compared to that of the whole valve, as well as its distance from the basi-scutal angle, and the form of its basal extremity, all vary in some degree. The length of the spur sometimes varies considerably, as in B. concavus and amphitrite. The parts of the mouth are only occasionally serviceable; for the teeth on the labrum, and the state of the lower teeth on the mandibles, and the presence of a step-formed projection at the lower angle of the maxillæ, are all often variable. The relative lengths of the two rami of the first pair of cirri, the degree of protuberance of the segments, and the number of pairs of spines on the segments of the posterior pairs of cirri, are sometimes useful; but the relative lengths of the cirri, and more especially the numbers of pairs of spines on the posterior cirri, are apt to vary. Finally, I must express my deliberate opinion, that every part and organ, internal and external, in Cirripedes, is liable to some amount of variation in some of the species.
I must now point out the principal changes which super[Pg 189]vene during growth, and which cannot properly be called variations. In the first place, I think, it is scarcely possible to recognise a species when under the 1/10th of an inch in diameter. In some cases, as in var. d’Orbignii of B. tintinnabulum, the shell is invariably coloured when old, but quite white when very young. Generally the tints become very much darker with age. Some species, which usually or invariably have, when mature, longitudinally folded walls, as B. flosculus and balanoides, are perfectly smooth in youth. The walls in B. eburneus, when young, have white, hyaline, longitudinal lines, and are naked; whereas, with advancing age, these lines disappear, and the subsequently formed shell becomes covered with membrane. The summits of the radii are apt to be oblique in the young of B. Capensis, psittacus, and tintinnabulum, whereas they are generally quite square in old specimens. In B. eburneus, cariosus, and in a lesser degree in B. psittacus, the scuta become longitudinally striated only with age. On the other hand, in very young specimens of B. tintinnabulum, the scuta sometimes are deeply impressed by little pits placed in rows. I have already alluded to the longitudinal furrow on the tergum so entirely changing its character, owing to the edges becoming, during growth, folded inwards; this likewise causes a decrease in the proportional breadth of the spur. In old specimens of B. flosculus, var. sordidus, the whole tergum is much more elongated than in young specimens. The basal margin of the sheath is hollow beneath in the young of B. cariosus and of some other species, but in the old it is continuous with the inner surface of the walls. The inner lamina of the parietes generally loses, to a certain extent, its longitudinally ribbed character in old age. The basis is solid, instead of being porose, in very young specimens of B. improvisus. In all the species, the carino-lateral compartments, in early age, are very narrow in proportion to the width of the lateral compartments; and in all, at this early period, the operculum is large in proportion to the whole shell. The number of spines on the edge of the maxillæ, the number of segments in all the cirri, and the number of spines on each segment, are few in early youth, and go on increasing with[Pg 190] each successive exuviation: the pedicels of the cirri are long in proportion to the rami at this same early age.[84]
[84] In some specimens of Balanus perforatus I made the following enumeration of the number of segments in the cirri:—
| Basal diameter of shell 1/20th of an inch. | Basal diameter of shell 1/5th of an inch. | Medium sized specimen about 3/4ths of an inch in basal diameter. | |
|---|---|---|---|
| First cirrus shorter ramus | ? | 11 | 17 |
| Second cirrus | 4 or 5 | 9 | 13 |
| Sixth cirrus | 9 or 10 | 19 | 31, in another 36 |
In the specimen 1/5th of an inch in basal diameter, each segment of the posterior cirri carried five pairs of spines; whereas, in full-grown specimens, there are six or seven pairs. In the 1/20th of an inch specimen, on the inner maxillæ, there were no spines between the upper large and the lower large pair of spines; whereas, in the 1/5th of an inch specimen, there were five intermediate spines, and in larger specimens nine or ten spines.
Notwithstanding the difficulties now enumerated, I hope that, owing to having examined a vast number of specimens of the most varying species, I have not fallen into very many errors. I have endeavoured to err on the side of making too few instead of too many species. In those cases, however, in which I have seen only a few specimens, I have been sometimes compelled to decide without sufficient evidence.
I would gladly have divided this genus, already including 45 species, into smaller genera; but so far from being enabled to do so, I have been compelled to form my Sections (immediately to be given) on characters not absolutely invariable, and far from obvious. I was particularly anxious to separate the elongated species with a boat-formed basis, which are attached to Gorgoniæ, and which form the genus Conopea of Say, but I was unable to effect their separation even as a sub-genus; for B. navicula and cymbiformis graduate in the most insensible manner through B. galeatus (the type of Say’s genus) and B. calceolus into B. stultus, and this into B. Ajax; yet this latter species has even been described as a mere variety of the typical B. tintinnabulum! Indeed, so insensible is this graduation, that the first and second sections of the genus are hardly distinct. I fully[Pg 191] admit, that if B. stultus and Ajax had never existed, B. calceolus and its three allies might have formed as natural a little group, though difficult to be characterised, as does the sub-genus Acasta; or perhaps this group and Acasta might have been combined together. These same species, viz., B. calceolus and its allies, are intimately allied to B. terebratus and inclusus, which are contained in the last section (F) of the genus; and this shows that Dr. Gray’s proposed genus Chirona, including the species with non-porose parietes in sections (E) and (F), could hardly have been instituted, even if the porosity of the parietes had not been variable in B. unguiformis, balanoides, and glandula. My fourth section (D), founded on the basis not being porose, is perhaps the weakest of the divisions, as may be seen in the list of exceptions appended to the sectional headings.
The arrangement of the species is, I think, as natural as a linear one could be made: every one knows how irregularly and in how many directions the lines of affinity in every natural genus branch out. Some few species stand rather isolated, as B. declivis; and B. allium, cepa, and quadrivittatus in a little group by themselves. I have already shown how the species in the first and second sections (A and B) blend into each other; and that the blending species are likewise allied to some in the last section (F); furthermore, I shall have occasion to show that these same species can hardly be separated naturally from the sub-genus Acasta. The first section, moreover, passes into the third (C) by B. tulipiformis; and the third into the fourth (D) by B. improvisus, nubilus, corrugatus, and patellaris: the fifth and sixth (E and F) sections are closely connected together by B. cariosus and flosculus; and these two sections blend into the fourth (D) through B. unguiformis and balanoides, and lastly, into the third (C) section by B. dolosus and improvisus.
The genus, as we have just seen, is hardly separated from the sub-genus Acasta; by B. allium it tends to pass through the sub-genus Creusia into Pyrgoma; and by B. imperator and flosculus it graduates into Elminius and Tetraclita.
Geographical Distribution.—This, which is much the largest genus of sessile cirripedes, has its species scattered over the whole world, from the arctic regions, in lat. 74° 48′, where we have B. crenatus and porcatus, throughout the tropical seas, to Cape Horn, where B. flosculus adheres to the coast-rocks. Many of the species have individually very wide ranges; thus B. tintinnabulum and amphitrite are found throughout the warmer seas; but the wide distribution of these species may be partly due to their frequent adhesion to ships’ bottoms: B. crenatus ranges from the frozen seas, in lat. 74° 48′ north, to the West Indies and Cape of Good Hope—a wonderful endurance of the most opposite climates. Balanus improvisus, again, extends from Europe to Nova Scotia, thence southward to Patagonia, and up the western coast of S. America, someway north of the Equator. Most of the species have considerable ranges; thus of the six species found on the eastern shores of northern America, five of them occur in Great Britain. Of the thirty-six species of which the habitats are known, exactly one third, or twelve, inhabit both the torrid and temperate zones, these being divided by the isocryme of 68°; nine are found exclusively in the torrid, and fifteen exclusively in the temperate zones. Within the warmer latitudes, and especially in the southern hemisphere, Tetraclita and Elminius to a certain extent supplant Balanus. In depth, the species range from the upper limits of the tidal zone to even fifty fathoms. Balanus improvisus and eburneus are able to survive in brackish water. The different species are attached to various surfaces—rocks, shells, timber, floating objects, sea-weed, lamelliform corals, Milleporæ, Gorgoniæ, and even to sponges. Mr. G. B. Sowerby has remarked[85] that in the species from the southern hemisphere it is the basis, and in the species from the northern hemisphere it is the parietes, which are elongated, when the individuals, from being crowded together, become cylindrical; but this is erroneous; B. perforatus, in the northern hemisphere, sometimes has an elongated basis; but no doubt the basis of our commonest species, as B. balanoides, crenatus, and porcatus,[Pg 193] from being either membranous or thin, does not become cup-shaped; whereas this structure is conspicuous in B. psittacus and lævis, the two commonest species in southern South America.
[85] Darwin’s ‘Geology of S. America,’ p, 264.
Fossil Species.—Having already given, under the Family, some account of the geological history of sessile cirripedes, short as it is, I here only allude to the subject in order to state my conviction that species cannot be satisfactorily distinguished in a fossil state, and rarely in a recent state, without an examination of the opercular valves. Nothing, indeed, could have been easier than to have affixed names to many groups of specimens, having different aspects, but to feel sure that these were really distinct species requires better evidence than can be afforded by the shell, without the operculum. No doubt, in some of the smaller sections of the genus—for instance, in that characterised by a membranous basis—it would have been possible to have distinguished some or several fossil species; but such have not as yet been found. When the specimens are much fossilised, it is, indeed, difficult to make out the primary points of structure—namely, whether the parietes, radii, and basis are porose: to do this it is sometimes necessary to rub down, polish, and carefully examine, a transverse section of a piece of the shell.
Parietes, and basis, and radii permeated by pores.
Parietes and basis sometimes permeated by pores, sometimes not; radii not permeated by pores; shell elongated in its rostro-carinal axis; basis boat-shaped, attached to Gorgoniæ and Milleporæ.
Parietes and basis permeated by pores; radii not permeated by pores.
Parietes permeated by pores. Basis and radii not permeated by pores.
Basis membranous.
Parietes and radii not permeated by pores. Basis sometimes permeated by pores, sometimes not, sometimes excessively thin and hardly distinguishable.[86]
[86] The following species might, owing to variation, or to the obscurity of their structure, without care, be classed in the wrong sections:—in A, Balanus Ajax, from living attached to Milleporæ, is sometimes elongated in its rostro-carinal axis, and from having its radii only finely porose, it might be classed in B: on the other hand, Bal. stultus is sometimes but little elongated, and the basis hardly boat-formed, and hence might be classed in A. The distinction between all the species in (B) and the sub-genus Acasta is artificial.
In sections C and D, I have seen one specimen of B. spongicola with a solid basis, and very young specimens of B. improvisus are thus characterised, and therefore these species are liable to be placed in the wrong section, D: Bal. nubilus, also, has part of its base non-porose, and therefore likewise might be placed in D: on the other hand, the circumference of the basis in B. patellaris is often porose, and hence this species, which belongs to D, might be placed in C.
In Bal. glandula, in D, the parietes of the compartments, without several were examined, might be thought to be solid, and therefore this species might be wrongly placed in F; on the other hand, the fossil B. unguiformis, in F, often has porose parietes, and such specimens would naturally be placed in D. Lastly, without care, B. flosculus might be thought to have a membranous basis, and so get placed in E.
The genus Pachylasma, without an examination of the animal’s body, might easily get misplaced in the wrong genus, amongst the species in the last section (F) of Balanus, yet there can be no doubt that Pachylasma belongs to the Chthamalinæ.
LEPAS TINTINNABULUM. Linn. Syst. Naturæ, 1767.
---- ------------ Ellis. Phil. Transact., vol. 50, 1758, Tab. 34, figs. 8 and 9.
---- ------------ Chemnitz. Neues. Syst. Conch., 8 B. (1785), Tab. 97, figs. 828-831.
BALANUS TULIPA. Bruguière. Encyclop. Meth., 1789; sed non B. tulipa alba, in Chemnitz; neenon B. tulipa, O. F. Müller, Zoolog. Dan.; neenon B. tulipa, Poli, Test. ut. Siciliæ.
------ ---- G. B. Sowerby. Genera of Recent and Fossil Shells, Tab. Genus Balanus.
LEPAS CRISPATA (var.) Schröter. Einleitung Conch. vol. iii, Tab. 9, fig. 21.
---- SPINOSA (var.) Gmelin. Linn. Syst. Nat.
---- TINTINNABULUM, SPINOSA, CRISPATA ET PORCATA. W. Wood. General Conchology, 1815, Pl. 6, figs. 1, 2. Pl. 7, figs. 4, 5. Pl. 8, figs. 1-5.
BALANUS TINTINNABULUM. Chenu. Illust. Conch.[87]
------ D’ORBIGNII (var.) Chenu. Illust. Conch., Tab. 6, fig. 10, sed non Tab. 4, fig. 13.
------ CRASSUS (Foss.) Sowerby (!). Min. Conch., 1818. Tab. 84.
[87] Chenu gives several admirable figures of this species; but he confounds some forms certainly distinct under this name, for instance the B. tulipiformis of this work.
Shell varying from pink to blackish purple, often striped and ribbed longitudinally: orifice generally entire, sometimes toothed. Scutum with the articular ridge broad and reflexed. Tergum with the basal margin generally forming a straight line on opposite sides of the spur.
Var. (1) communis (Pl. 1, figs. a, b, f supra; Pl. 2, figs. 1 a, 1 c, 1 d, 1 e, 1 i, 1 k): conical or tubulo-conical; smooth or moderately ribbed longitudinally; colours varying from purplish-pink to blackish-purple; often in obscure longitudinal stripes; orifice of shell rounded-trigonal.
Var. (2) vesiculosus (young) (Pl. 2, fig. 1 h): exterior surface of the scuta impressed with small square holes, arranged in two or more rows, radiating from the apex of the valve.
Var. (3) validus (Pl. 1, figs. c, f infra): globulo-convex; coarsely ribbed, ribs flexuous; either smooth or rugged; pale chocolate purple or pink; shell extremely strong; orifice almost circular.
Var. (4) zebra (Pl. 1, fig. g): conical; rich chocolate purple with broad snow-white ribs; sheath bright chesnut colour; summits of alæ oblique; orifice almost circular.
Var. (5) crispatus (Schröter) (Pl. 1, fig. h): pale blueish or pinkish-purple, with irregular rough projections, or with short, sharp, needle-[Pg 196]like points; scuta with their exterior surface either plain, or with radiating lines formed of hood-like projecting points.
Var. (6) spinosus (Gmelin) (Pl. 1, fig. i): globulo-conical or cylindrical; shell rather thin, with long, upcurved, nearly cylindrical, very sharp points; colours very pale; attached to other specimens, and to Lepas anatifera.
Var. (7) coccopoma (Pl. 1, fig. d; Pl. 2, fig. 1 f, 1 l, 1 o): globulo-conical; orifice small, rounded; walls generally smooth, thick; intense rose-colour, sometimes most faintly striped longitudinally with varying shades of pink; radii tinged with purple; scutum sometimes as in var. communis, sometimes with its basi-tergal corner much cut off, with the adductor ridge prominent, the pit for the depressor muscle deep, and the articular ridge broad and hooked; tergum sometimes as in var. communis, sometimes with a broader spur, placed nearer to the basi-scutal corner of the valve.
Var. (8) concinnus (Pl. 1, fig. e; Pl. 2, fig. 1 g): globulo-conical; walls finely ribbed; dull purple, tinged and freckled with white; scutum, with a broad, hooked, articular ridge, with an extremely sharp plate-like adductor ridge, and with a cavity, bordered by a plate, for the rostral depressor muscle; tergum as in var. 1.
Var. (9) intermedius: radii with their summits slightly oblique; parietes pale blueish purple, with narrow dark purplish-blue longitudinal lines; sheath with the internal surface of the rostrum and lateral compartments much darker colored than the internal surface of the carina and carino-lateral compartments; scuta and terga as in var. communis.
Var. (10) occator (Pl. 1, fig. k; Pl. 2, 1 b): radii with their summits slightly oblique; parietes smooth, or ribbed, or spinose; very pale blueish-purple, with narrow darker longitudinal lines; sheath with the internal surface of the rostrum and lateral compartments dull blue, whilst the corresponding parts of the carina and carino-lateral compartments are white; scuta with small, sharp, hood-formed points, arranged in straight radiating lines; terga with the spur placed at either its own width, or less than its own width, from the basi-scutal angle.
Var. (11) d’Orbignii (Chenu) (Pl. 1, fig. l; Pl. 2, 1 m, 1 n): radii with their summits oblique, and the orifice of the shell rather deeply toothed; shell conical or tubulo-conical, smooth, or rugged; colour dull purplish-lilac, with the tips of the parietes and a band along one side of the radii quite white; sheath rather darker at the rostral than at the carinal end; scuta as in var. 1; terga as in var. occator.
Habitat.—West coast of Africa, on mytili; Madeira, on rocks; West Indies; Cape of Good Hope, on a patella and on kelp; mouth of the Indus; East Indian Archipelago; Sydney, Australia, attached to Lepas anserifera, adhering to a floating cane; Peru; Galapagos Islands; West Mexico; California. Extremely common on ships’ bottoms arriving from West Africa, India, and China, often associated with B. amphitrite.
Fossil Red Crag, England; Mus. S. Wood and J. de C. Sowerby.—Touraine (?); Mus. Lyell.
General Remarks.—This, the first species of Balanus, is, perhaps with the exception of B. amphitrite, the most difficult and variable in[Pg 197] the genus. There are some other species which vary quite as much in external appearance; for instance, B. perforatus; but B. tintinnabulum also varies in far more important points, as in the proportions and structure of the opercular valves. The difficulty in determining whether or not the differences are specific, is wonderfully increased by whole groups of individuals varying in exactly the same manner. I have seen three most distinct varieties taken from the bottom of the same vessel, so that I did not at first entertain the least doubt that they were three distinct species. I may mention, as showing the vacillations which I have experienced on this subject, that beginning with the impression, that the above three varieties were really distinct species, after going over the several immense collections of specimens placed at my disposal, I came to the conclusion that the above three, and several other forms presently to be described, were only varieties; yet after an interval of some months, having to look at some of these specimens again, I could not but think that I had come to a false conclusion, and so went into all the details again, and satisfied myself that I had followed a right course; after another interval, I had to repeat the same process, and even now I can never look at a group of the beautifully coloured shells with their small rounded orifices of var. coccopoma attached to the Avicula margaritifera, or again at var. d’Orbignii, with its toothed orifice and white tips to the compartments, without thinking that they must be specifically distinct from the dull-coloured specimens with large entire orifices so common on ships’ bottoms; yet I can produce a full series of intermediate forms, and I can further show, in each variety, that the several points of difference by which each is characterised, are variable. I may be permitted to add, in order to show that it has not been from indolence that I have combined so many forms, that I had named and already written out full descriptions of most of the varieties, before determining to sacrifice them.
Seeing that B. tintinnabulum and amphitrite are the two most variable species in the genus, more especially in the important characters derived from the opercular valves, and knowing that these species are attached so very frequently to ships’ bottoms, one is led to suspect that their extreme tendency to vary may be due to their being exposed to varying and peculiar conditions, whilst transported to new and distant localities. It is even just possible, as may be inferred from the facts given in the Introduction (p. 102) in regard to certain monstrous specimens of Bal. balanoides having been apparently impregnated by adjoining individuals, that the varieties may interbreed, and so produce numerous intermediate forms. Whether or not this could take place, I am inclined to look at these two species, as in an almost analogous condition with our domestic animals, which give rise to such infinitely numerous varieties. It appears to me probable, that several of the varieties keep true to their peculiarities, as long as they continue to breed in the same locality; but that when their larvæ become attached to ships’ bottoms, and are thus transported and exposed to new conditions, they give rise to new and ever-varying varieties. I will first give a full description of the more common forms of B. tintinnabulum, which undoubtedly belong to the same species,[Pg 198] only alluding to the less frequent points of difference, and then separately describe the more marked varieties.
General Appearance.—Shape of shell generally tubulo-conical, or conical, or globulo-conical, rarely depressed. Orifice either large and rounded-trigonal, or small and oval, either entire or less frequently toothed. Surface quite smooth, or longitudinally ribbed; ribs of variable strength, not unfrequently flexuous or branching, sometimes roughened with blunt or sharp projecting, irregular points, or more rarely with almost cylindrical, upturned, long spines; the simple longitudinal ribs are generally most strongly marked in young specimens. Colour, generally varying from pink, to pink tinged with purple, to dark, inky purple, more or less striped, longitudinally, with white or pale tints; rarely the shell is of the brightest rose-colour, either uniform or longitudinally striped; sometimes it is pale purplish, or dark blue; and sometimes dark chocolate-purple: the ribs, when present, are generally more or less white, sometimes snow-white. That there is much variation in colour, and in the prominence of the longitudinal ribs, is quite certain, as the two sides of the same individual sometimes differ greatly in these respects. The radii are generally rather darker coloured than the parietes, but sometimes they are lighter, even in the darkest tinged specimens. The surfaces of the radii are occasionally finely plaited in lines parallel to the basis. In some infrequent varieties the radii have oblique summits, making the orifice of the shell to be toothed. The sheath is generally feebly coloured, but sometimes bright chesnut-brown, and sometimes blueish. The strength of the shell varies considerably; some of the globulo-conical varieties are extremely massive. Size; basal diameter of largest specimen very nearly three inches; height of the highest specimen three inches.
Young specimens are apt to have a peculiar aspect; for their shell is often strongly ribbed longitudinally, and the summits of their radii are sometimes oblique. Their scuta are sometimes deeply pitted in radiating lines. Their colours are generally pale. I have seen specimens attached to kelp from the Cape of Good Hope, with their parietes white and ribbed, and their radii mottled with pinkish-purple; I have seen other young specimens from the Galapagos Archipelago, of a uniform grayish-blue.
The Scuta generally have their lines of growth moderately prominent; occasionally they are longitudinally striated, with the lines of growth flexuous and upturned at intervals into small, sharp, hood-liked projections, which are symmetrically arranged in straight lines radiating from the apex of the valve; I have seen this structure in some specimens of var. crispatus and in var. occator (Pl. 2, fig. 1 b); and I have noticed an intermediate state in var. communis. The degree to which the basi-tergal corner of the valve is rounded off varies much even in var. communis (Pl. 2, fig. 1 a, 1 d, 1 e). The articular ridge (1 c, 1 e) is broad and much reflexed; and often, but not always, distinctly hooked (1 f). The adductor ridge is confluent with the articular ridge, and runs straight down the valve, bounding the cavity for the depressor muscle; generally the adductor ridge is blunt, and so little prominent as barely to deserve notice; but I have seen it sharp and prominent in[Pg 199] one specimen of var. communis, and it is generally prominent in var. coccopoma (1 f), and most remarkably so in var. concinnus (1 g). The cavity for the lateral depressor muscle is generally very slight; but in the two vars. just mentioned, and sometimes in var. communis, it is deep. In var. concinnus (1 g) there is a remarkable plate developed for the attachment of the rostral depressor muscle. The scuta are coloured either dull-purple or reddish, or striped longitudinally white and blue. The surface is sometimes externally depressed in the line of the adductor ridge; and in young specimens there is sometimes, along this line, a chain of pits (1 h), as in full-grown specimens of B. trigonus and lævis.
The Tergum (Pl. 2, figs. 1 i to 1 o) is broad, with a generally closed longitudinal furrow; this furrow is open in young specimens, and it is often, but not always, open in rather large specimens of var. occator; it is always open in var. d’Orbignii (Pl. 2, fig. 1 m), and sometimes in var. concinnus. Apex barely beaked, except in var. spinosus, in which it is sensibly produced. Spur placed either very nearly in the middle of the basal margin, or when least medial, it stands at above its own width from the basi-scutal angle; yet in some specimens of var. occator the spur is less than its own width from this angle. The basal margin (1 i), on opposite sides of the spur, either forms a nearly straight line, or the scutal portion descends lower than the carinal portion, and curves very regularly towards the spur; this is the case in var. d’Orbignii (1 n), and in some specimens of var. occator. The carinal half of the basal margin generally forms an angle with the spur of only a little above a rectangle. The spur varies a little in length and breadth, but never exceeds one fourth of the greatest breadth of the valve. The scutal margin is broadly inflected, the inflected portion forming either a right angle, or somewhat less than a right angle, with the exterior surface of the valve. Internally the articular ridge is prominent, and is either considerably or slightly curved; it extends down either about half, or three fourths, of the length of the valve. The spur is produced for a considerable distance up the internal surface of the valve as a prominence. The crests for the tergal depressor muscle are very feeble. In one specimen, in which both the shell and operculum had undergone much disintegration, the scuta and terga were calcified together.
Compartments: their exterior appearance has been already described. The parietal tubes are not large; they are generally crossed by transverse septa in their uppermost part; but they are sometimes almost solidly filled up by dark shelly layers. The internal surface of the parietes is more or less plainly ribbed; in old specimens, however, it generally becomes smooth. The radii have their septa denticulated on both sides; and they are porose, that is, the interspaces between the septa are not filled up solidly. The radii generally extend from tip to tip of the adjoining compartments, that is, their summits are parallel to the basis; but in three not common varieties, viz., vars. intermedius, occator, and d’Orbignii (the former of which at least must, without the smallest doubt, be ranked as a mere variety), their summits are oblique. I have occasionally met with specimens of var. communis with oblique[Pg 200] radii; and this is not very infrequent in young shells. Exteriorly the radii are generally smooth, but sometimes finely ribbed horizontally, owing to the projection of the septa. The alæ have their sutural edges smooth; their summits are usually parallel to the basis, but they are often much broken; in var. zebra, however, in every specimen which I examined, the summits were oblique. The sheath varies much in colour: in var. occator, and in a less degree in var. intermedius and var. d’Orbignii, the portion lining the rostrum and lateral compartments is much darker than the other parts of the sheath. The Basis generally has a thick, underlying, cancellated layer. Sometimes the basis (Pl. 1, fig. b) is irregularly cup-formed.
Mouth: labrum with four or six minute teeth: mandibles with five graduated teeth; inferior point more or less spinose. Maxillæ, either with or without a small notch, beneath the upper pair of spines; in the lower part there are two spines longer than those immediately above them. Cirri, the first pair has the rami unequal, in the proportion of about 19 segments in the longer ramus, to 16 in the shorter. The segments in the latter have their anterior surfaces very protuberant. The second pair is short, with the anterior surfaces of the segments protuberant. On the thorax (Pl. 25, fig. 1), on each side, at the bases of third pair of cirri, there is a projecting membranous plate fringed with fine bristles. The three posterior pairs have their segments shield-shaped in front, generally bearing four pairs of spines, of which the lower pair is minute; between these pairs there are some minute spines. In some young specimens from the Cape of Good Hope, and in var. concinnus, I found six pairs of spines on the segments of the posterior cirri.
Geographical Distribution.—This species is extremely common over the whole of the warmer seas. It ranges from the Island of Madeira to the Cape of Good Hope, and on the west coast of America, from Monterey, in lat. 37° N., in California, to Peru. It is attached to rocks and sub-littoral shells, to floating timber, to kelp, and to Lepas anatifera. It is attached in wonderful numbers to ships’ bottoms arriving at our ports, from West Africa, the West Indies, the East Indian Archipelago, and China. It is generally associated with B. amphitrite and amaryllis. I have already stated that, on the bottoms of vessels, the different varieties are generally grouped together; and this makes me believe that they are local. In Mr. Stutchbury’s collection there are numerous specimens taken from a ship which first went to the west coast of Africa for guano, and then to Patagonia for the same object, and it was interesting to see the manner in which numbers of B. psittacus, a Patagonian species, had become attached on the African B. tintinnabulum. The varieties from the west coast of America seem eminently peculiar; we there find var. coccopoma and concinnus; and a blueish, rugged variety, with peculiar opercular valves.
Geological History.—I have seen specimens in Mr. S. Wood’s collection from the Red Crag of England, which, though not accompanied by opercular valves, I cannot doubt belong to this species. The specimens named by Mr. Sowerby, in the ‘Mineral Conchology,’ as B. crassus, and which I have seen through the kindness of Mr. J. de C.[Pg 201] Sowerby, also belong to this species. I further believe that a specimen in Sir C. Lyell’s collection, given to him by M. Dujardin under the name of B. fasciatus from Touraine, is likewise B. tintinnabulum.
With respect to var. communis, I have nothing to remark. The second, var. vesiculosus (Pl. 2, fig. 1 h), is confined to young specimens, and may, perhaps, be due to a want of calcareous matter. With respect to var. validus (Pl. 1, figs. 1 c, 1 f infra), I may observe that some of the coarsest and strongest specimens which I have seen were said to have been attached to a surface of iron. I have seen two large lots of var. zebra (1 g), taken by Mr. Stutchbury, from the bottoms of ships, arriving from Bengal and China, and in both cases associated with var. communis, and in one case with var. coccopoma. I at first thought that this variety, zebra, was specifically distinct, but now I feel no doubt, that it is a mere variety; its body was in every respect identical with that of var. communis.
Nor have I any doubt that var. crispatus, of Schröter (Pl. 1, fig. h), is only a variety, although the scuta in some specimens have a peculiar appearance, externally like these valves in var. occator (Pl. 2, fig.1 b): the scutum is here broader and flatter than in var. communis, and the adductor ridge is very feebly developed, but we shall see how variable this ridge is in all the varieties: externally, the sharp, hood-like points formed by the upturned lines of growth, have a very remarkable appearance, from being arranged in quite straight radiating lines. This structure is evidently caused by the same tendency which produces on the walls the sharp, upturned, irregular points; but it is singular that the scuta are smooth in some specimens with very rough parietes; and, on the other hand, bristling with the symmetrically arranged, hood-like projections, in other specimens on which I could with difficulty detect only a few exceedingly minute points on the walls. In var. communis I have seen a few specimens with a slight tendency in the scuta to become striated longitudinally. The tergum in var. crispatus presents no difference from that in var. communis. Some of the roughest and best characterised specimens of var. crispatus appear to have come off copper-sheathed vessels.
I believe var. spinosus of Gmelin (Pl. 1, fig. i), has been correctly considered by me as a variety, but I have unfortunately seen only one set of specimens with their opercular valves preserved: these were attached to Lepas anatifera. The colour of the shell varies from reddish-purple to nearly white; the radii are sometimes quite white; the walls are slightly ribbed. The scuta in the above specimens, externally were smooth; the adductor ridge was rather more distinct from the articular ridge than in any other variety; and the terga rather more plainly beaked. The tubular, up-curved, calcareous spines sometimes occur only on one side of the shell, and often only in the lower part. These spines are often coloured brighter than the walls. Their presence cannot be accounted for (any more than the state of the scuta in the[Pg 202] foregoing var. crispatus) by the nature of the surface to which they are attached; for I have seen one set attached to a large rugged specimen of B. tintinnabulum, and another to the very smooth valves of Lepas anatifera. I believe that this form is almost always associated with var. communis, which is an argument that it is only a variety.
To var. coccopoma (Pl. 1, fig. d) I alluded in my introductory remarks as having so strongly the aspect of a distinct species. I possess a beautiful group, with a globulo-conical, smooth shell, of the finest rose colour, with a rather small, rounded orifice. These specimens were attached (mingled with B. trigonus) to Avicula margaritifera, from, as Mr. Cuming believes, Panama. I can never look at this set of specimens without doubting the correctness of the determination at which I have arrived. In the British Museum there are two sets of specimens taken off a vessel, on the west coast of South America, almost identical in external appearance with those in my possession, but rather more rugged. Mr. Stutchbury has sent me some specimens from a ship, direct from China, which are rather paler pink, and more striped, and come near to some ordinary varieties of B. tintinnabulum. The scuta (Pl. 2, fig. 1 f) in the above three sets of specimens agree in having the adductor ridge more developed, and the pit for the lateral depressor muscle deeper than is usual. The tergum (1 l, 1 o) in most, but not in all these specimens, has a rather broader spur; and some of the specimens have the carinal portion of the basal margin considerably hollowed out; the spur, also, is placed nearer the basi-scutal angle than in ordinary cases. On the other hand, in Mr. Cuming’s collection, there are two specimens taken off a vessel, identical in external appearance with the foregoing, but which have scuta and terga in every character exactly as in var. communis; hence I am compelled to consider all these specimens as mere varieties.
Var. concinnus (Pl. 1, fig. e) is, perhaps, the most remarkable of all the varieties; I have seen three sets of specimens from the west coast of South America,—all identical in appearance, having longitudinally-ribbed walls, either rosy or of a dull purple, striped and freckled in a peculiar manner with white. I have, however, seen an approach to this colouring in some few specimens of var. communis; and the shell itself offers no other peculiarities. The scutum (Pl. 2, fig. 1 g) resembles, in general shape, that of var. coccopoma; but the adductor ridge is here much sharper and more prominent; and the rostral depressor muscle, instead of being lodged in a little cavity formed by the folding over the occludent margin, has, in addition, a small plate on the under side, which tends to convert the pit into a tube. The tergum exactly resembles that of var. communis. The segments in the sixth cirrus bear six, instead of four, pairs of spines,—a circumstance which I have noticed only in some young specimens of var. communis, from the Cape of Good Hope. From these several peculiarities, until quite lately, I resolved to keep this form specifically distinct; but I have finally concluded that they are not sufficient. For firstly, I have seen a scutum in var. communis (Pl. 2, fig. 1 d), with the adductor ridge nearly as sharp; and this ridge is always strongly pronounced in var. coccopoma; secondly, with respect to the plate for the rostral depressor muscle,[Pg 203] although I have not seen this in any other variety, yet in Bal. concavus a closely analogous plate, situated in the lateral depressor cavity, is highly variable, and I am not willing to found a new species on one minute point of structure,—a structure which is variable in another species of the same genus.
I have seen some cylindrical and conical specimens of B. tintinnabulum, from the coast of Mexico and California, only noticeable, as far as the shell was concerned, from being rugged, and of a dull blueish-purple; but which had opercular valves exactly like those of var. coccopoma, and therefore, as far as the scutum is concerned, approaching closely in structure to var. concinnus,—all three from the west coast of America. Hence I was at one time led to believe that there existed a species on this line of coast, which represented B. tintinnabulum, and which varied in external shape and colour in an analogous manner to that species. But as the opercular valves in var. coccopoma are sometimes identical with those of var. communis, and as this is always the case with the tergum of var. concinnus, and as the shell itself presents no differences, it is scarcely possible to admit the existence on the west coast of America of this supposed representative of B. tintinnabulum.
With respect to var. intermedius I have little to say in addition to the character given above: I have seen only two groups of specimens in Mr. Cuming’s collection: the chief interest in this variety is that it shows that the next form must be ranked as a variety, and not as a distinct species.
Of var. occator (Pl. 1, fig. k) I have seen several specimens, mostly taken off the bottoms of vessels, and one specimen, marked in Mr. Cuming’s collection “South Seas.” After having carefully examined these specimens, I came to the conclusion, that the slightly oblique radii—the general colouring, and more especially that of the sheath—the scuta (Pl. 2, fig. 1 b), with their sharp hood-like points, in radiating lines—and the terga, with the spur so near to the basi-scutal angle, were amply sufficient to distinguish it as a good species. Subsequently, however, I found that the scuta in var. crispatus presented, both externally and internally, exactly the same peculiar appearance. In var. intermedius, I found the summits of the radii equally oblique, and the general colouring nearly the same, and more especially a close approach to the singular circumstance of the sheath differing in colour towards the opposite ends of the shell. So that the position of the spur of the tergum was the chief remaining character; and this evidently varied considerably in the four or five specimens examined by me, being either its own width, or much less than its own width, from the basi-scutal angle: the outline, also, of the small portion of basal margin, between the spur and the basi-scutal angle, likewise varied much, being either angularly indented, or gradually curved down towards the spur: so also the tip of the spur varied in shape. The longitudinal furrow is unusually apt, in this variety, to remain open. We know that the position of the spur varies considerably in var. communis. Hence, although the spur, on an average, lies closer to the basi-scutal angle in this than in any other variety, even than in[Pg 204] var. d’Orbignii, it would, I conceive, be preposterous to found a species on this one character. In the animal’s body, every part agrees perfectly with that of var. communis.
Lastly, we come to var. d’Orbignii (Pl. 1, fig. l): until quite recently I did not even suspect that this form was only a variety of B. tintinnabulum: I have examined a great number of specimens in Mr. Stutchbury’s collection, which had come attached on a vessel from Java, and likewise a few other specimens in other collections. They all closely resemble each other in shape, and even in size, and differ only in tint of colour, and in the surface being either very smooth, or longitudinally ribbed, sometimes with rugged, sharp points. From this circumstance—from the peculiarity of the tint, with the tips of the parietes and one side of the radii perfectly white—and from the obliquity of the summits of the radii, I was led to think this form specifically distinct. But the colour does not differ from that of some other varieties of B. tintinnabulum; the circumstance of the colour being uniform or not striped, is common to the sub-varieties of several varieties, and the white tips to the parietes, and the white borders to the radii, result simply from the shell, whilst young, having been wholly white, and this is not rarely the case with var. communis. Dismissing, therefore, colour, it will be found that hardly any other characters remain by which this form can be separated from var. occator; in both the summits of the radii are oblique, in both the sheath is coloured in nearly the same manner, in both the opercular valves, especially the terga (Pl. 2, figs. 1 m, 1 n), resemble each other; the scuta, however, are smooth in var. d’Orbignii and rough in var. occator. This latter form, certainly, cannot be specifically separated from var. intermedius, and this assuredly is only a variety of B. tintinnabulum. Hence I am led to conclude that Balanus d’Orbignii of Chenu, peculiar as its whole aspect is, must be ranked only as a variety of B. tintinnabulum; its oblique radii resulting from the same cause, whatever that may be, which has given this structure to var. intermedius and occator; and its peculiar colouring to having been exposed (owing probably to having been transported on vessels) to different conditions, whilst young and old.
BALANUS TULIPIFORMIS EX CORALLIO RUBRO. Ellis.[88] Philosoph. Transactions, vol. 50 (1758), tab. 30, fig. 10.
LEPAS TULIPA. Poli. Test. utriusque Siciliæ, tab. 5, fig. 1. et 6 (1791).
BALANUS TINTINNABULUM (var.) Chenu. Illust. Conch., tab. 3, f. 5.
[88] According to the letter of the Rules of the British Association, Ellis’s name ought to be retained, as it was published in 1758, the same year during which the 10th edition of the ‘Systema Naturæ’ appeared, in which edition the binomial method was first used. But as Ellis himself did not then know of, or follow this method, it might be disputed whether, according to the spirit of the law, his name ought to stand. The only other name given to this species is that of tulipa, affixed by Poli in 1791, but this name had been previously used by Müller in 1776, and by Chemnitz in 1785, for another species, the B. Hameri of this work; and likewise, also previously to Poli, by Bruguière in 1789, for still another species, viz., B. tintinnabulum of this work: under these complicated causes of confusion, I think it is highly advisable to adopt Ellis’s name. I may add that the B. tulipa of Mr. G. B. Sowerby is the B. tintinnabulum of this work. It is possible that the B. conoides of Brown, ‘Illustrations Conch.’ (1st edit. pl. 6, fig. 7), may be our present species; but without details of structure it is hardly possible to identify, in many cases, the species of Balanus.
Shell dark rose-coloured, sometimes tinged with purple; orifice toothed. Scutum externally very smooth, covered by membrane. Tergum with distinct crests for the depressor muscles.
Hab.—Sicily, Malta, Malaga, (associated with B. perforatus), Madeira. Often growing in clusters and associated with Pachylasma giganteum. Attached to Millepora aspera, oysters, and other shells. According to Poli, an inhabitant of deep water; yet in mus. Cuming there are two fine specimens attached to the always floating Lepas anatifera. Mus. Lowe, Macandrew, Stutchbury.
General Appearance.—Shell tubulo-conical or conical: orifice large, toothed, approaching to pentagonal. Surface moderately smooth, naked. Colour rosy, or tile-red, with a slight tinge of purple; or beautiful rich purple. Radii nearly as dark as, or darker than, the parietes. The portion of the alæ seen externally is generally white. Internally the whole shell is nearly white. Generally the tints outside vary in transverse fasciæ; sometimes there are very fine, dark, longitudinal lines. Largest specimen (from Malta), 1.4 of an inch in basal diameter; usually full-sized specimens are about three quarters of an inch in basal diameter.
Scuta (Pl. 2, fig. 2 a, 2 c) very smooth, with the growth-ridges very little prominent, sometimes there are obscure traces of longitudinal striæ; surface covered by an unusually thick and persistent yellow membrane: valve narrow, with the upper part commonly reflexed: the basal margin forms, with the occludent margin, a smaller angle than is usual: the tergal margin of the valve is rectangularly inflected, instead of being, as is usual, merely bowed inwards. Internally, the articular ridge is rather prominent. The depth of the slight pit for the lateral depressor muscle is variable; it sometimes includes a minute, central, longitudinal ridge.
Terga (2 b, 2 d): the longitudinal furrow is deep, with the sides folded in; the spur is placed at about its own width from the basi-scutal angle; it is moderately long, with its lower end obliquely rounded off; but the length, breadth, and precise outline of the lower end varies a little. The basal margin on the opposite sides of the[Pg 206] spur, forms a nearly straight line, but with the portion on the carinal side very slightly hollowed out. Crests for the depressor muscle are well developed.
Compartments.—The radii and alæ always have their summits oblique: the sutural edges of the radii are deeply penetrated by pores between the strongly denticulated septa: the sutural edges of the alæ are quite smooth: the tubes in the parietes are crossed in the upper part of the shell by septa. Basis tubular, with an underlying cancellated mass.
Mouth.—Labrum with the teeth either absent or very small: mandibles with the fourth and fifth teeth rudimentary: maxillæ with a small notch under the two upper spines; near the lower angle, two spines, one beneath the other, are larger even than the upper pair; beneath the lower pair, there is a tuft of fine spines. Cirri, segments protuberant in one ramus of the first cirrus and in both rami of the second cirrus; posterior cirri with the segments short and broad, each bearing three pairs of spines, with a small intermediate tuft.
Affinities.—This species in all essential respects comes very near to the three last varieties of B. tintinnabulum, which have the orifices of their shells toothed. The smoothness of the scutum, with its persistent epidermis,—its peculiar shape,—its small and not reflexed articular ridge,—together with the crests on the tergum for the depressor muscles, are sufficient diagnostic characters. Even in general habit and tint of colour, this species has a different aspect from B. tintinnabulum. In some respects B. tulipiformis leads into the species included in the third section of the genus.
LEPAS PSITTACUS. Molina. Hist. Nat. Chile (1788), vol. i, p. 223.
BALANUS PICOS. Lesson. Zoolog. Voyage de la Coquille (1829).
------ TINTINNABULUM (var. c). Ranzani. Mem. di Storia Nat. tab. 3, fig. 1-3 (1820).
------ CYLINDRACEUS. Lamarck, in Chenu. Illust. Conch. Tab. 4, fig. 17, Tab. 5, fig. 7, sed non var. (c.) in Lamarck, Animaux sans Vert., (1818).
------ PSITTACUS. King and Broderip. Zoolog. Journal, vol. v (1832-1834), p. 332.
Shell, pale dirty pink; orifice hexagonal. Scutum with the articular ridge very small, confluent with the very prominent adductor ridge, forming a tubular cavity, which extends up to the apex of the valve. Tergum with the apex produced, needle-like, purple: spur placed at less than its own width from the basi-scutal angle.
Hab.—Peru, Chile, Chiloe, Patagonia. Fossil in an ancient tertiary deposit, Coquimbo; and in a superficial, recent bed at S. Josef, in Patagonia.
General appearance.—Shell either almost cylindrical or steeply conical, generally flesh-coloured, sometimes pale pink; surface either smooth (when not disintegrated) or sometimes with the parietes distinctly and rather strongly ribbed, with the ribs distant from each other: I have seen six or seven ribs on the rostrum alone. The orifice in the most perfect specimens is nearly equilateral and hexagonal. The radii generally are very broad, but occasionally quite narrow, and even linear. The basis is generally deeply and irregularly cup-formed.
Size.—This is the largest species in the family: I have seen a specimen six inches in length and three and a half in diameter; and another specimen no less than nine in length, though only two and a half inches in diameter.
Scuta.—In full-sized specimens the surface is finely striated longitudinally, caused by the lines of growth being minutely sinuous; but in young specimens, until they attain a basal diameter of above half an inch, the surface is smooth. The valve is transversely arched, a line of flexure running from the apex to the basal margin, at about one third of the width of the valve from the tergal margin. The basal margin is curved nearly continuously, and extends nearly half-way up the valve; hence the basi-tergal corner is largely rounded off. The articular ridge is but little prominent, and is not reflexed: the articular furrow is very narrow. The adductor ridge consists of a sharp, much-projecting plate, running down close to the basal margin, and is confluent with the lower part of the articular ridge. This plate and the inflected tergal margin of the valve, together form a large and deep cavity, which extends up almost to the apex of the valve. The depressor muscle is attached in the middle, at the lower, open end of this cavity.
Terga.—These are strongly beaked, the beak being from one third to one fourth of the total length of the valve, including the spur: the beak is very sharp, somewhat flattened, and bowed; when young, and when well preserved, it is coloured purple: it is penetrated by a fine tubular cavity, occupied by a thread of corium, which extends about half-way up it. The whole valve is narrow, being about thrice as long as wide. The spur is also long and narrow; it is seated at less than its own width from the basi-scutal angle. The scutal margin is not much inflected. The longitudinal furrow has its sides, in full-grown specimens, closely folded together. The basal margin slopes down on both sides to the spur. There are no crests, or only traces of them, for the attachment of the depressor muscle. Internally, the spur is prolonged, as a prominent ridge, upwards to the beak, and serves as an articular ridge. In the middle, in the upper part (Pl. 2, fig. 3 d), between this articular ridge and the carinal margin, there is a second narrow ridge, which extends from the lower part of the beak half-way down the valve, and then dies out. The space between these two ridges, and the ridges themselves, are coloured purple, and consist of[Pg 208] harder shell than the rest of the valve; hence, when the outer surface and the adjoining scutal and carinal margins disintegrate, this part remains, and so forms the beaked, purple apex.
Compartments.—The parietal tubes are unusually large in proportion to the size of the shell, and run up to the summit without any transverse septa: the longitudinal septa are strongly denticulated. The radii are penetrated by large tubes; their septa are very strongly denticulated, and the denticuli themselves often subdivide and branch out at their extremities. The sutural edges of the alæ are smooth, or with a high power can just be seen to be crenated. The radii are generally very highly developed, so that their summits are even wider than the bases of the parietes; but, on the other hand, in some few large specimens, the radii are either very narrow or absolutely linear. In these latter cases, the diametric growth has nearly or altogether ceased, whilst the walls of the shell have continued to be added to at their bases, their summits at the same time suffering disintegration; and thus the orifice has increased in size.
Basis generally, and occasionally very deeply, cup-formed. An unusually thick cancellated layer in most cases forms the under side of the basis.
Mouth.—Labrum apparently without teeth, or with very minute ones: mandibles with three teeth, of which the third is thicker than the first or upper one: the fourth and fifth teeth are confluent with the inferior angle. The maxillæ have a small notch under the upper pair of spines; inferior part projecting and supporting two spines, placed one below the other, and equalling in size the upper pair. Cirri: the rami of the first cirrus are unequal by four or five segments; shorter ramus and both rami of the second cirrus with the segments extremely protuberant: posterior cirri not much elongated, with the segments rather broad, supporting six pairs of spines.
General Remarks.—This, which is much the largest known species of the genus, ranges from Peru (Arica being the most northern spot, whence I have seen specimens), along the coast of Chile, where it is very abundant at a few fathoms’ depth, at least as far south as Southern Chiloe; it is said by Captain King to attain the largest size at Conception. On the coast of Eastern Patagonia, I dredged up this species from nineteen fathoms, in lat. 49°. In lat. 42° (S. Josef), on the same eastern coast, I found fossil specimens in beds of sand upraised between eighty and one hundred feet. In the tertiary formation at Coquimbo, in Chile, it occurs in the middle bed, associated with the recent B. lævis, and with various mollusca, all of which are apparently extinct, indicating that the formation is of considerable antiquity. In the living state, on the coast of Chile, it is often associated with B. lævis. As it frequently adheres to large specimens of the Concholepas, it must sometimes be an inhabitant of shallow water. I have seen one specimen attached to Mytilus Magellanicus. Mr. Cuming believes that about six fathoms is the usual depth at which it lives. Numerous specimens are often congregated together into great masses. Mr. Stutchbury has some interesting specimens which he procured from a ship that had first sailed to Ichaboe, on[Pg 209] the coast of Africa, and afterwards to Patagonia; consequently numerous specimens of B. psittacus had become attached on B. tintinnabulum, and subsequently during the voyage home, some few of the latter again had adhered on B. psittacus: the contrast in the paler colour and hexagonal orifice of this species, with the darker tints and more trigonal orifice of B. tintinnabulum was striking. At Coquimbo, in Chile, I procured a specimen of B. psittacus, attached to a chain cable which had been in the water only six months; this specimen measured 1.3 of an inch in basal diameter, and .8 in height: this shows a rapid rate of growth. Lastly, I may mention that it is asserted by Molina, and I am assured by Mr. Cuming that the statement is perfectly correct, that this Balanus, when cooked, is universally esteemed as a delicious article of food.
BALANUS CAPENSIS ORE OBLIQUO. Ellis. Phil. Transact., vol. 50 (1758), Tab. 34, fig. 14.
Shell shaded, and often longitudinally striped with bright pink. Scutum as in B. psittacus. Tergum with the apex produced and needle-like, white: spur placed at its own width from the basi-scutal angle.
Hab.—Cape of Good Hope. Attached to stems of Fuci, Algoa Bay. Mus. Brit. and Bowerbank. Attached to a Patella, Mus. Darwin, Mus. Cuming, and Stutchbury. Attached to floating kelp, Lagulhas Bank, Mus. James Ross, associated with B. tintinnabulum and spongicola.
This species comes extremely close to the South American B. psittacus, and I should hardly have attached a specific name to it, had I not examined many specimens, young and old, of the true B. psittacus, from Peru, Chile, and Eastern Patagonia, and found them all identical in the few, apparently trifling points, in which that species differs from B. capensis. The animal’s body and the shell agree in every respect, excepting that the shell is decidedly pinker, being often most distinctly and prettily striped longitudinally with pale and bright pink. In some of the specimens the basis is cup-formed: in some, the broad radii are pale pink, in others they are quite white, and in this latter case a singular aspect is given to the pinkish varieties. In very large specimens (and I have seen one fully two inches in basal diameter) the pink colour is extremely feeble, and the whole shell has a very rugged, disintegrated, coarse, and sometimes dirty appearance: in most of these large specimens the walls are more massive than in B. psittacus, and the orifice of the shell rather smaller; in some, however, the walls certainly are of unusual thinness.
The Scuta differ from those of B. psittacus only in the basi-tergal[Pg 210] corner not being so much rounded off, and consequently in the articular ridge, which is rather more reflexed, descending in proportion lower down the valve: the cavity at the basi-tergal corner is in proportion broader. The valves in the two species differ, also, but only in young specimens, in the occludent half being tinted, both externally and internally, purple, whereas in B. psittacus the whole valve, at all ages, is white. In the terga the spur is removed fully its own width from the basi-scutal angle, whereas it is not half this distance in B. psittacus. The scutal margin is here much more inflected. In B. psittacus there is a remarkable patch of purple on the inside of the valve, between the articular ridge and a second special ridge; of this purple patch there is here no trace, consequently the beak or apex is white. The beak, also, is less prominent. The special ridge, just alluded to, here runs much nearer to the articular ridge, and is less prominent: indeed, in old specimens, it is often almost obliterated. Finally, the whole valve, in proportion to the Scutum, is rather broader.
I have seen a young specimen, about a quarter of an inch in basal diameter, with the orifice of the shell toothed owing to the obliquity of the summits of the radii; and this gave the shell a very peculiar aspect. The largest well-coloured specimen which I have seen is 1.2 of an inch in basal diameter; but in Mr. Cuming’s collection there are two rugged, disintegrated specimens, two inches in basal diameter, and two and a half in height. Some specimens, 1.3 in basal diameter, in Mr. Stutchbury’s collection, are remarkable from the radii having been obliterated—the shell being merely divided by six sutures, as we have seen is likewise sometimes the case with large specimens of B. psittacus.
This species is evidently a South African representative of the South American B. psittacus.
BALANUS NIGRESCENS. Lamarck, (1818) in Chenu. Illust. Conch., Tab. 4, fig. 16.
------ GIGAS. Ranzani. Memoire di Storia Nat., 1820, Tab. 3, fig. 5, 6, 7.
------ ---- De Blainville. Dict. des Sc. Nat., Tab. 116, fig. 2, 2 a.
Shell cinereous, tinted with pale or blackish blue, or wholly white. Scutum with the articular ridge terminating downwards in a small, sharp, free point: adductor ridge prominent. Tergum with the apex produced and needle-like.
Hab.—Swan River, West Australia, Mus. Brit., attached to sandstone. Attached to sandstone and to each other in a group, Mus. Cuming. Twofold Bay, S. E. Australia, attached to tidal rocks and Patellæ, Mus. Darwin.
There can be no doubt that this species is the B. nigrescens in Chenu, who had access to Lamarck’s original specimens; and there can be equally little doubt that it is the B. gigas of Ranzani, collected, during Baudin’s expedition, at King George’s Sound: it is essentially allied to B. psittacus, but in external appearance strikingly resembles some of the varieties of B. tintinnabulum.
General Appearance.—Shape tubulo-conical: walls smooth, sometimes longitudinally ribbed: colour ashy-gray tinged with blue, but many specimens are dark purplish-blue, owing to the disintegration of the outer lamina, and consequent exposure of the almost solidly filled up, dark blueish parietal tubes; on the other hand, some specimens are quite white. Ranzani describes the colour as earthy-violet, which is very characteristic of some of the specimens. The orifice is apt to be rather small, compared to the size of the specimens, and tends to be hexagonal. The radii are often rather narrow. The opercular valves are tinted pale blue. The basal diameter of the largest specimen is two inches, and its height two and a quarter.
The Scuta have their basi-tergal corner much rounded off, as in B. psittacus, so that the tergal margin does not extend more than half down the valve. The surface is somewhat prominent, along a line running from the apex to the point of chief curvature in the basal margin. The surface is not striated. Internally, the articular ridge is little prominent, and not reflexed; the lower end depends as a free, sharp style or point. The adductor ridge is moderately sharp, and stands some little way distant from the articular ridge: it is produced downwards, and forms a moderately deep and large cavity for the depressor muscle; but this cavity is not closed, and does not extend up, as in the two last species, to the apex of the valve.
Terga, narrow, with a sharp, prominent, needle-like beak. Spur, long, narrow, placed at less than its own width from the basi-scutal angle: the basal margin on both sides slopes down to the spur: the scutal margin is not inflected. Internally, the articular ridge is very feebly developed, but extends down close to the basi-scutal angle. On the under surface in the upper part of the valve, there is a short, very slight ridge, extending on the carinal side, near and parallel to the articular ridge; this slight ridge plays an important part, as in the two foregoing species, in the formation of the beak or apex. Crests for the depressor muscle are hardly distinguishable.
The Walls appear to vary in some degree in strength and thickness; as is likewise the case with the opercular valves. In some of the thinner specimens, the parietal tubes are large, and the longitudinal septa are furnished with small, sharp denticula. The tubes are often thickly lined or almost filled up solidly with blue shell; they are not crossed by transverse septa.
The Radii vary in width; externally they are often finely ribbed transversely, at other times they are smooth; their septa are fine and[Pg 212] thin, with their delicate denticuli not extending to the outer lamina: they are very porose. The alæ have their summits parallel to the basis; their sutural edges are most finely crenated. The sheath is blueish, excepting the wedge-like portions of the alæ which have been added during the diametric growth, and these are white.
Mouth: labrum without teeth: mandibles with five sharp teeth: maxillæ with the edge straight. Cirri, first pair with the rami very slightly unequal; segments of the shorter ramus and of both rami of the second pair protuberant: posterior cirri with the segments shield-shaped in front, bearing four pairs of spines, of which the upper pair is much longer than the lower pairs; each pair has a small intermediate tuft of minute spines.
Parietes pale pink; radii rather darker. Scutum with a small articular ridge. Tergum with the longitudinal furrow very shallow and open; basal margin on both sides sloping towards the spur.
Hab.—New Zealand. Mus. Brit., and Flower: attached to shells.
General Appearance.—Shell conical or tubular, with a large rhomboidal orifice; very pale pink, but tinted yellowish from the persistent epidermis, and sometimes faintly striped longitudinally; radii and sheath of rather a darker pink; scuta in themselves white, though lined by purple corium; the carinal half of the tergum pink. Walls extremely smooth. Largest specimen above one inch in basal diameter.
Scuta, with the finest striæ radiating from the apex; growth-ridges moderately prominent; articular ridge small; there is a very slight and blunt adductor ridge: the hollow for the lateral depressor muscle is rather narrow and deep.
Terga, with the apex slightly prominent or beaked; the longitudinal furrow is of very little depth; on its scutal margin there is a narrow, rounded, slightly prominent ridge, which, however, appears more like a furrow than a ridge. Spur moderately long and blunt; placed at half its own width from the basi-scutal angle; the basal margin on both sides of the spur, slopes gently towards it. Internally, the articular ridge is pretty well developed; the scutal margin is not much inflected; the carinal portion of the under surface of the valve is rough; the crests for the carinal depressor muscle are entirely absent.
Compartments.—Walls moderately strong; parietal tubes small, with transverse septa in their upper ends; inner surface of the walls much less strongly ribbed than is usual. Radii broad, with their summits parallel to the basis; their septa are strongly denticulated. Alæ[Pg 213] with their summits oblique; their sutural edges are barely crenated. Basis, thin, flat, or cup-formed. Body unknown.
Affinities.—In general appearance this species comes near to B. psittacus; but in all essential characters it comes much closer to the following species, from which, however, it can easily be distinguished by colour, and by the inner lamina of the parietes not being cancellated.
Shell purplish dark brown: inner lamina of the parietes cancellated. Scutum finely striated longitudinally. Tergum with the longitudinal furrow shallow and open; basal margin on both sides sloping towards the spur.
Hab.—West Coast of South America. Mus. Cuming.
General Appearance.—Shell conical, with a large, rhomboidal orifice; walls rather thin, coloured, together with the radii and operculum, dark purplish-brown; sheath nearly colourless. Walls smooth, slightly irregular, very finely striated longitudinally. Basal diameter of largest specimen .8 of an inch.
Opercular Valves, unusually smooth, that is without prominent growth-ridges. Scuta, finely striated longitudinally, with the sharp striæ closely approximate. The teeth on the occludent margin are sharp, and stand some way apart from each other. Internally, the whole surface is remarkably flat and smooth: the articular ridge is of moderate breadth, and slightly reflexed: there is no adductor ridge, and the oval depression for the lateral depressor muscle is extremely slight. Terga, with the longitudinal furrow very slight; the bottom of this furrow is feebly striated longitudinally, and there is a trace of a fine, rounded ridge on the scutal margin, as in B. decorus. The basal margin slopes on both sides towards the spur, which is of moderate length and breadth, with its lower end truncated and parallel to the carino-basal margin; the spur stands at about once and a half its own width from the basi-scutal angle. Internally, the valve is lined by very dark, purplish-brown corium; the articular ridge is prominent; in the upper part of the valve, parallel to the articular ridge, there are two or three feeble ridges; there are no crests for the tergal depressores.
The Parietes, though moderately thick, yet are light and fragile; the denticuli at the bases of the longitudinal septa are prominent, and those on the adjoining septa are united together, making a network (Pl. 2, fig. 7 d), but the interspaces between them are not filled up by solid calcareous matter (as is the case with every other species of the genus), but are only crossed at successive levels by fine transverse calcareous septa; the internal lamina thus becoming cancellated, and, though thick, fragile. Hence, in a transverse section of the parietes, the ordinary parietal tubes or pores are seen to be lined on their inner sides[Pg 214] by five or six rows of very minute pores. I have not seen any other instance of this structure. The internal lamina is ribbed, as usual, on its inner surface, by the projection of the longitudinal septa. The ordinary parietal tubes are open, to nearly the summit of the shell. The radii are rather thin, and unusually fragile; their summits are parallel to the basis: their septa, as seen on the sutural edges, are extremely thin and denticulated on both their upper and lower surfaces, on the side towards the internal lamina: towards the external lamina, the septa are simple, and the small square pores thus formed, are open or not filled up. The alæ have their summits extremely oblique, being added to very little during the diametric growth of the shell; the narrow margin, however, which is thus added, is coloured red, the rest of the sheath being nearly colourless: the sutural edges of the alæ are smooth. The basis has a thick, underlying, finely cancellated layer of shell.
Animal’s body unknown.
A young specimen, .2 of an inch in basal diameter, differed from the above in being of a much paler purplish-brown. This species is distinct from all its congeners, in its peculiar colour, and likewise in the structure of the inner lamina of the parietes. As already stated, it comes nearer to B. decorus than to any other species.
BALANUS TINTINNABULUM (var.) Chenu. Illust. Conch., Tab. 2, fig. 8.
Shell globulo-conical, often elongated in the rostro-carinal axis, pale pink, smooth, extremely massive: parietal pores, close to the basal margin, circular and very small. Scutum with the articular ridge broad and reflexed.
Hab.—Philippine Archipelago, attached to Millepora complanata, Mus. Cuming. Mus. Brit. and Stutchbury.
General Appearance.—Shell globulo-convex, sometimes much elongated in its rostro-carinal axis; smooth; walls excessively strong, massive, and heavy. Orifice oval, rather small in proportion to the size of shell, this being chiefly due to the infolding of the upper part of the rostral compartment. Parietes pale pink, feebly tinted with purple: radii either paler, or tinted of a bright chesnut-brown: sheath rich purplish chesnut-brown. Basal diameter of the largest specimen nearly 3-1/2 of an inch; height 2-3/4: another specimen had a basal longitudinal diameter of 2.9 of an inch, and a transverse diameter of only 1.6; this great difference in the two diameters being caused by the prolongation of the basal portion of the rostrum in the line of the branch of the Millepora, to which the shell had adhered; the height of this same specimen was[Pg 215] 1.5; and the diameter of the orifice, both transversely and longitudinally, .75 of an inch.
Scuta, broad, feebly tinted with pink; exterior surface rough, with sharp hood-formed projections, arranged in straight lines radiating from the apex; an inflected portion of the valve along the tergal margin is not roughened. Internally (Pl. 3, fig. 1 d), the articular ridge is broad and reflexed. An adductor ridge can hardly be said to exist, but a slight prominence borders the gentle hollow in which the lateral depressor muscle is attached. The basal margin, on its inner face, is slightly toothed. Tergum white, with the narrow part of the valve, on the scutal side of the spur, rough with the little projecting hoods, like those on the scutum; the other and larger half is smooth: spur rather long, narrow, placed at twice its own width from the basi-scutal angle; on the carinal side, about half of the basal margin slopes down towards the spur. The longitudinal furrow is either quite or nearly closed. Internally, the spur is produced upwards on the valve, as a prominence: the articular ridge is not very prominent. There are no crests for the tergal depressor muscle.
Altogether the opercular valves strikingly resemble those of B. tintinnabulum, but all the characters above mentioned have not been observed in any one variety of this species; perhaps var. coccopoma comes nearest, both in the external appearance of the shell and in the structure of the opercular valves, to B. Ajax.
The Compartments are remarkably compact and solid; the parietal tubes are cylindrical and quite minute even close to the basis; they extend, however, nearly up to the top of the shell; the parietal septa at the basis are thick, and with blunt denticuli; the thickness of the walls in the upper part of the shell is excessive; in the lower part, it is also unusually great, owing to the thickness of the inner lamina, and hence the ribs, generally formed by the projection of the longitudinal septa on the inner lamina, are here visible only close to the basis. The radii are rather wide; their summits are parallel to the basis; the septa on their sutural edges are thin, straight, and closely approximate, and most symmetrically furnished with little denticuli of equal sizes on both sides: the interspaces are nearly filled up solidly, but with some pores still left open. In the upper part of the shell, the radii, like the walls, are of extraordinary thickness: the septa are transverse and horizontal, as seen externally by slight variations in the colour of the radii; internally, as seen in a vertical section of the shell, the septa dip inwards at an angle of above 45°. The alæ are thin, and have their summits oblique: their sutural edges are smooth. The pores in the basis are crossed by numerous transverse septa, and there is an underlying cancellated layer: the internal surface is very smooth.
Animal’s body unknown.
The strength of this Balanus is truly remarkable; and when, by repeated blows, a specimen which I was examining at last yielded, the radii broke sooner than separate at their sutures. In most of its characters, this species approaches B. tintinnabulum, and I believe has been included by Chenu as one of its varieties; but it comes almost equally near to B. stultus, to which it is much more closely allied in[Pg 216] its habit of being attached to Milleporæ. By a close and unbroken chain of affinities, B. Ajax, through B. stultus, is connected with B. calceolus and its allies in section (B), which live attached to Gorgoniæ. Some of the specimens of B. Ajax, are almost as much elongated in their rostro-carinal axis, as are the species in section (B); and there is an affinity in the same direction in the smallness of the pores in the radii of B. Ajax; indeed, had the basis in this species been generally more boat-or cup-formed, I should have placed it as the first species in section (B), instead of, as at present, the last species in section (A). The intermediateness of the characters of B. Ajax has been one chief cause why I have rejected the genus Conopea, which was instituted by Say for the species living attached to Gorgoniæ.
Parietes and basis sometimes permeated by pores, sometimes not: radii not permeated by pores: shell elongated in its rostro-carinal axis: basis boat-shaped: attached to Gorgoniæ and Milleporæ.
Parietes and base porose: shell white, or faintly tinged with purple. Scutum with the basal margin protuberant in the middle. Tergum with the longitudinal furrow closed in the upper part: spur not closely adjoining the basi-scutal angle.
Hab.—Attached to Milleporæ, Singapore, Mus. Cuming. West Indies,[89] Mus. Brit.—Mus. Stutchbury.
[89] This specimen in the British Museum was purchased at the sale of the Rev. L. Guilding’s collection, and therefore it is not certain that this habitat is correct; but as it was sold in the same lot with a Cirripede certainly West Indian, and as the main collection was made in the West Indies, this habitat may, I think, be trusted.
I have considerable doubts whether it would not have been more correct to have placed this species in the last section, instead of where it now stands; it certainly is more[Pg 217] closely allied to B. Ajax, especially in its operculum, than to the following species; yet the fact of the radii not being permeated by pores does not permit of its admission into the last section; and both in habits and structure it undoubtedly comes very near to the following species. Those varieties which are not much elongated, and which have the basis nearly flat, would certainly, if considered by themselves alone, not have gained admission into our present section.
General Appearance.—Shell conical, somewhat globular, more or less elongated in the rostro-carinal axis, owing to the basal production of the rostrum. Orifice, rather small, entire, oval, pointed at the carinal end. Radii moderately broad, with their summits parallel to the basis. Colour dirty white, often faintly tinged with purple; sheath, pale purplish-blue. Surface extremely smooth; the parietes are generally covered (as viewed through a lens) by a very thin, yellowish epidermis, giving to the whole a glistening, granular aspect: the radii are generally destitute of this epidermis, and are therefore of a dead white. The basis is concave, and sometimes deeply cup-formed; it is, however, not symmetrical; sometimes it is flat. Basal diameter of largest specimen, including the basis itself, 1.5 of an inch in the longitudinal axis; transverse diameter, 1 inch; the inequality in the length of the two diameters is rarely so great as in this unusually large specimen.
Scuta, externally very convex, with the growth-ridges extremely prominent; basal margin sinuous, the middle portion being prominent; this is best seen in young specimens (Pl. 3, fig. 2 d). Internally, the articular ridge is broad and reflexed. The adductor ridge in the upper part is almost confluent with the articular ridge; it runs down to the most prominent point of the basal margin; in young specimens it is sharp and prominent; in old specimens it is very blunt and little prominent. There is a rather deep hollow for the lateral depressor muscle. In young specimens there is a small, depending, blunt tooth at the basi-tergal angle, which helps to make the basal margin more deeply sinuous.
Terga, with the longitudinal furrow closed, except on the spur itself, where it is open. The spur is moderately long and broad, but varies in breadth; it is placed at rather less than its own width from the basi-scutal angle; its lower end is obliquely rounded; the basal margin on the opposite sides of the spur, together form a nearly straight line. The whole valve is rather broad. The crests for the tergal depressores are barely developed.
The Compartments have rather large parietal tubes; the septa are coarsely denticulated at their bases; the internal lamina is smooth, except close to the basis. The radii have their summits parallel to the basis; their sutural edges are formed of rather thick septa, which stand at an unusual distance apart from each other, and have perfectly symmetrical, minute denticuli on each side. The interspaces between the septa are filled up solidly to within a short distance of the surface; but yet not so[Pg 218] completely as in the following species, and as in those in the succeeding sections of the genus; this is what might have been expected from the close affinity of B. stultus to B. Ajax, in which latter the radii are still permeated by pores, though smaller than is general in the species of our first section (A). The alæ have their summits extremely oblique, and their sutural edges, I believe, smooth. Basis porose, with an underlying, finely-cancellated layer.
Mouth: labrum with six small teeth; mandibles with the 3d tooth blunt; the 4th minute, and the 5th almost confluent with the inferior angle. Maxillæ with the edge straight and simple. Cirri partly destroyed; on each segment of the sixth pair there were five pairs of spines.
BALANUS CALCEOLUS KERATOPHYTO INVOLUTUS (?) Ellis. Phil. Trans., vol. 50 (1758), Tab. 34, fig. 19.
LEPAS CALCEOLUS (?) Pallas. Elench. Zooph., p. 198, (sine descript.) (1766).
CONOPEA OVATA (?) J. E. Gray. Annals of Philosophy, vol. x, 1825.
Parietes and basis porose. Scutum with the pit for the lateral depressor muscle small and deep.
Hab.—Attached to Gorgoniæ, West Coast of Africa. Tubicoreen, near Madras, (Dr. Johnston), associated with B. navicula. Mediterranean (?). Mus. Brit., Cuming, Stutchbury.
Fossil. Coralline Crag; Mus. S. Wood.
I must premise, with respect to the nomenclature of this and the three following quite distinct species, that in the published descriptions no allusion is made to any one of the characters by which alone they can be distinguished: hence I have been guided by geographical probabilities in assigning the specific name of calceolus to the present species, as Ellis’s specimens came from the Mediterranean; and that of galeatus to the North American and West Indian specimens, as Linnæus’ original specimens (according to a statement by Spengler) came from the West Indies. I have assigned new names to the two remaining East Indian species. I may here add that Spengler (‘Skrifter af Naturhist.’ 1 B, tab. 6, fig. 3, 1790) has described, under the name of B. cassis, an allied form attached to the Gorgonia placomus from the seas of[Pg 219] Norway; but I do not believe that it is the same with our present species.
General Appearance.—The degree of elongation of the shell in its rostro-carinal axis varies considerably (3 a, 3 b): the elongation is due to the production of the rostrum and of the corresponding end of the basal cup. These two portions of the shell always form together an angle, and sometimes an acute angle, whereas in all the many specimens which I have seen, the carina and the carinal end (or heel) of the basis together form a straight line; yet I should not be surprised if this end of the shell was sometimes produced. The surface of the shell is smooth, or sometimes marked with very minute projecting points: it is almost always covered by the horny bark of the Gorgonia. The colour is either dull purplish-red or dull purple, with obscure longitudinal stripes, and often more or less transversely banded with white. The rostrum is either white or very feebly tinted, being always paler than the rest of the shell: the radii are usually paler than the parietes, and are sometimes white: the carinal end of the basal cup is tinted of the same colour with, but rather paler than, the compartments. The orifice is rather small compared to the shell, and nearly heart-shaped. The carino-lateral compartments are about one-third of the width of the lateral compartments. The shell is very strong, and the sutures resist the action of boiling caustic potash. The largest specimen which I have seen was .7 of an inch in extreme length, and under .25 in extreme breadth.
Structure of the shell and basis.—The parietes are permeated by quite distinct pores,—a character sufficient by itself to separate this from the following species; the longitudinal septa forming the tubes are slightly denticulated at their bases. The radii have their summits quite square, extending from apex to apex of the adjoining compartments. The alæ have oblique summits. The sutural edges of the radii have approximate septa, which are obscurely denticulated: the interspaces are filled up solidly, so that the radii are not porose. The basis is distinctly porose, by which this species can be distinguished from B. navicula and cymbiformis. The basis has a deep furrow on the under side, from clasping the thin horny axis of the Gorgonia: the basal point of the rostrum is also notched from the same cause, and, as a consequence, its upper surface becomes slightly furrowed along its whole length.
The Scuta have an articular ridge but moderately prominent, and only slightly reflexed; the basi-tergal corner is rounded off; there is no adductor ridge; there is a small, rather deep, distinct pit for the lateral depressor muscle. Terga; externally the surface is considerably depressed in the line of the spur. The spur is between half and one-third of the width of the valve: its lower end is square and truncated, or in some degree rounded; it is sometimes (3 e) dentated with a few, minute, sharp teeth. The articular ridge is but slightly developed; the crests for the depressor muscle are very feeble.
Animal’s body unknown.
LEPAS GALEATA (?) Linnæus. Mantissa altera Holmiæ, 1771.
CONOPEA ELONGATA. Say.[90] Journal of Acad. Nat. Sci. Philadelphia, vol. ii, part 2, p. 323, 1822.
[90] If I have assigned the specific title of galeatus to the wrong species, yet Say’s name of elongatus ought not strictly to be admitted; as the Lepas elongata of Gmelin is a Balanus,—probably a variety of Balanus crenatus. I may add, that as the Lepas galeata of Schröter (‘Einleitung in die Conch.’ &c.), was attached to a Gorgonia from the East Indies, it cannot be our present species, but probably is one of the three other allied species, which all occur in India.
Parietes not porose; basis porose. Tergum, with the apex square, caused by the great development of the articular ridge.
Hab.—Charlestown, South Carolina; Florida; West Indies; Central America; attached to Gorgoniæ; Mus. Brit., Agassiz, Cuming, Stutchbury.
General Appearance.—This and the two following species come so close in general appearance to the last, that it will be quite superfluous to do more than describe the few points of difference. The shell and basis are generally quite as much elongated as in the last species, and sometimes much more so, owing to the carinal end (fig. 4 a), with the corresponding portion of the basal cup, being produced like the rostral end, into a flattened, sharp point: I have seen a specimen in this state .9 of an inch in length, and only .25 in breadth in the broadest part. In many specimens, however, the shape is exactly as in B. calceolus; but the rostrum seems less usually furrowed from clasping the stem of the Gorgonia. The colour is paler, pinker, and more distinctly striped longitudinally than in B. calceolus; I have, however, seen some not-striped, purple specimens (and one transversely freckled with white) from the West Indies. The parietes are strongly-ribbed internally, and are not permeated by pores. The radii have their sutural edges crenated. The basal cup is permeated by pores.
The Scutum differs from that in the last species, only in the pit for the lateral depressor muscle, being much shallower, and less defined, and in the apex being truncated. The Tergum is remarkable from its broad, square, truncated summit, which underlies the whole broad apex of the scutum: the square summit of the tergum is formed by a great and peculiar development of the uppermost part of the articular ridge. The spur is a little narrower than in B. calceolus.
Mouth: on the crest of the labrum there are two teeth on each side of the central notch. The mandibles have five teeth, of which the two lower are very small. The maxillæ show a trace of a notch under the upper large pair of spines; near the inferior angle there are two long spines. Cirri: in the first pair, one ramus is nearly twice as long as the other: the segments are not very protuberant. There is a sharp point at the dorsal basis of the penis. The branchiæ are of moderate size, and plicated on one side.
Parietes and basis not porose. Scutum and Tergum with very small articular ridges. Tergum broad, almost equilateral.
Hab.—Attached to a Gorgonia, Tubicoreen, near Madras, (Dr. Johnston). Hab. unknown, Mus. Cuming.
General Appearance.—I have seen only two specimens, kindly sent me by Dr. Johnston, and a single specimen in Mr. Cuming’s collection. In most points this species agrees with the two last species. The shell (excepting the rostrum), and even the opercular valves in Mr. Cuming’s specimen were of a very fine purplish-red; in the other specimens they were feebly tinted purple. The parietes are strongly ribbed internally, and are not permeated by pores. The basal cup is not porose, but its inner surface is ribbed in lines radiating from the centre, and in both these respects this species differs from the two foregoing. The Radii are rather narrow; they are paler coloured than the parietes; they have their sutural edges plainly crenated. The alæ have extremely oblique summits; the narrow rim added during the diametric growth of the shell is white, the rest of the sheath being, in Mr. Cuming’s specimen, finely coloured like the parietes. Basal diameter of the longer axis of the largest specimen, .4 of an inch.
Scutum, rather narrow, with the basi-tergal corner much rounded off; externally the lines of growth are little prominent. Internally, the articular ridge is extremely little developed, and not at all reflexed; there is no adductor ridge; there is a minute pit for the lateral depressor muscle, placed almost on the edge of the valve. The Tergum is broad, forming (the spur being excepted) an almost equilateral triangle. The articular ridge is remarkably little prominent, and placed close to the scutal margin. The spur is nearly half as broad as the valve, with its extremity or basal margin in one case obliquely truncated, and in another case nearly square.
Animal’s body unknown.
Parietes and basis not porose: carino-lateral compartments very narrow, and of nearly the same width from top to bottom: radii with their sutural edges smooth. Scutum externally striated longitudinally.
Hab.—Attached to Gorgoniæ, Tubicoreen, Madras (associated with B. calceolus), Dr. Johnston. Hab. unknown, Mus. Brit. and Darwin.
This is a very distinct form, though nearer to the foregoing than to the other species. Its separation from the sub-genus Acasta is quite artificial; its affinity to this sub-genus is shown by its weaker shell, non-porose parietes and basis; by the radii having their sutural edges smooth, and their summits not quite square; by the carino-lateral compartments being very narrow; by the less elongated basis, not furrowed, from not clasping the branches of the Gorgonia; and by the longitudinally striated scuta; nevertheless, from the similar habits, and from the graduated structure in the five foregoing species, it cannot be removed out of the genus Balanus. I have seen three sets of specimens of this species.
General Appearance.—Shell, sometimes with the rostrum, and sometimes with the carina, and corresponding portions of the basal cup, elongated; but not, apparently, to so great a degree as in the foregoing species; basis not furrowed, from not clasping the branches of the Gorgonia. Colour pale blueish-purple, with the radii whiter. The surface is studded with small calcareous points. The carino-lateral compartments are very narrow, not more than one tenth of the width of the lateral compartments; they are, moreover, scarcely wider at the base than at the summit. The summits of the radii are, apparently, a little oblique, or at least not so square as in the foregoing species. The shell is not nearly so strong as in the last three species; and the compartments separate by gentle force, and from the action of caustic potash. The largest specimen was .4 of an inch in basal diameter.
Internally, the parietes are not very strongly ribbed, or they are almost smooth, and there are no pores. The basis is concave and smooth within, and is not porose. The sutural edges of the radii are quite smooth, or sometimes they exhibit, in the lower part, mere traces of septa,—a character by itself sufficient to separate this from the foregoing species. The alæ have oblique summits, and the rather narrow portion added during the diametric growth of the shell, is white.
The Scutum, externally (6 d), has raised striæ, radiating from the apex; valve rather thick; internally, the articular ridge is but slightly prominent, and its lower end is rounded off: the depression for the lateral depressor muscle is slight; between this depression and that for the adductor muscle, the surface of the valve is prominent. Tergum, somewhat beaked; externally, the surface is depressed in the line of the spur: the carino-basal margin slopes towards the spur.
Animal’s body unknown.
Parietes and basis permeated by pores. Radii not permeated by pores.
Parietes ribbed, mottled purplish-red; orifice broad, trigonal, hardly toothed. Scutum thick, with from one to six longitudinal rows of little pits. Tergum without a longitudinal furrow; spur truncated, fully one third of width of valve.
Hab.—Java; East-Indian Archipelago; Peru; West Columbia; California; Sydney; New Zealand. Mus. Brit., Cuming, Stutchbury, Dunker, &c.
General Appearance.—Shell conical, generally depressed; orifice broad, triangular, almost equilateral; walls coloured or only mottled with purplish-pink, having either irregularly branching, or regular, longitudinal ribs, which are generally white. The radii are pale pink, or nearly white: the opercular valves have either their upper parts, or nearly their whole surface, clouded with pinkish-purple: the epidermis is not persistent: the walls are moderately strong: the largest specimen was one inch, but generally full-grown specimens are about half an inch in basal diameter.
The Scuta have the lines of growth highly prominent. From one to five or six rows (7 b, 7 c) of nearly circular, or transversely oblong, deep pits, extend down the middle of the valve; rarely there is not even one row; in this latter case, the valve is not striated longitudinally. These little pits are caused by one or more deep longitudinal furrows, crossed by the lines, or rather ridges, of growth. In the same group of specimens, I have seen individuals with three, five, and six rows; and even a few specimens with only one row, or none at all. The outline of the valve is elongated, with the apex slightly reflexed: the inner surface is protuberant, sometimes to a remarkable, but variable degree. The articular ridge is not very prominent, but it extends fully half-way down the valve, and generally ends in a small free point. There is a short adductor ridge, and a deep narrow pit or cleft for the lateral-depressor muscle. Terga, externally smooth, flat, with scarcely a trace of a longitudinal furrow; spur broad (7 e, 7 f), varying from half to one third of the width of[Pg 224] the valve, with the end truncated, situated either near or quite close to the basi-scutal angle. The crests for the depressor muscles are moderately well developed.
Compartments.—The parietal tubes are, in their upper parts, filled up solidly, without transverse septa. The radii generally have their summits slightly oblique, and this is almost always the case with the radii of the rostrum; the other radii sometimes extend from tip to tip of the parietes, and are parallel to the basis; rarely the radii are considerably oblique. The septa of the radii are very obscurely denticulated, and the interspaces between them are filled up solidly. The alæ have their sutural edges thin and smooth.
Mouth.—Labrum with three teeth close together on each side of the central notch: mandibles with four teeth, the fourth being small, the fifth either absent or scarcely distinguishable from the inferior angle: maxillæ without any notch, with the two lower spines rather longer than the others. Cirri: In the first pair, one ramus is only half the length of the other; in the second pair, both rami are short and about equal in length; in the posterior pairs, the segments, which are not protuberant, bear four pairs of spines, of which the three lower pairs are short.
This species is widely-distributed, and where found seems to be common. It is generally attached to shells of mollusca, but I have seen it also attached to wood. I have found it associated with B. tintinnabulum, var. concinnus, and coccopoma, with B. psittacus, improvisus and amphitrite, and with Elminius modestus.
Young specimens bear a considerable resemblance to certain young varieties of B. tintinnabulum, and can indeed be distinguished from them only by a careful examination of the opercular valves; for it should be borne in mind, that in certain cases the scuta in B. tintinnabulum are pitted with little cavities. This species in some respects is, I think, allied to B. porcatus, but it is far more closely related to B. spongicola, and can be discriminated with difficulty from certain varieties of this latter species. In Mr. Cuming’s collection, there is a group of small specimens, crowded between some older specimens, which are remarkable from the shell being oval in a transverse section,—from the smoothness of the walls,—and from the absence of pits on the scuta; yet there could be no doubt that these specimens belonged to our present species.
BALANUS SPONGICOLA. Brown’s Illustrations of the Conchology of Great Britain (1827), pl. 7, fig. 6: 2d edit. (1844), pl. 53, figs. 14-16.
Parietes generally smooth, sometimes longitudinally folded; coloured pink: orifice toothed. Scutum longitudinally striated. Tergum, with the apex produced, without a longitudinal furrow; spur truncated, about one third of width of valve.
Var. with the walls slightly folded longitudinally.
Hab.—South coast of England, and Tenby in South Wales, often imbedded in sponges; attached also to shells and rocks in deep water; Mus. Brit., Jeffreys. Algiers, on Mytili and Serpulæ, with B. perforatus, Mus. Mac Andrew. Madeira, with B. tulipiformis, Mus. Lowe. Lagulhas Bank, Cape of Good Hope, on detached kelp, with B. Capensis, Mus. Sir J. Ross. Imbedded in sponge with Acasta spongites, Mus. Bowerbank. Var. West Indies.
Fossil in Coralline Crag, Mus. S. Wood.
General Description.—Shell tubulo-conical; orifice of moderate size, rather deeply toothed; colour dull pink, or purplish, or dark flesh-colour; sometimes the radii are paler, sometimes of the same colour with the parietes. Surface smooth when well preserved, having transverse rows of minute spines. In the West Indian variety the walls are slightly or much folded, but I will describe this form separately. Size of largest specimen (Mus. Jeffreys), .6 of an inch in basal diameter.
Scutum, with fine ridges radiating from the apex, and with the lines of growth, crenated: internally, the articular ridge is small, adductor ridge short and barely distinct: there is a rather deep and narrow pit for the lateral depressor muscle. The whole valve is much thinner than in B. trigonus, which in most respects it closely resembles. Tergum, with the apex pinkish purple, produced or beaked, but the beak is not needle-like, as in B. psittacus and its allies, for the carinal margin is perfectly preserved up to the tip. Externally the valve is nearly flat, for the longitudinal furrow is very shallow. The spur is about one third of the width of the valve; its lower end is abruptly truncated: in European specimens (1 b) the whole basal margin, on the carinal side, slopes down to the spur in a straight line, which, together with the sharpness and production of the basi-scutal angle of the spur itself, gives to the whole valve a peculiar appearance: in the specimen (1 c) from the Lagulhas Bank, the basal margin on the carinal side is a little more hollowed out, but it is quite impossible to doubt about the specific identity of these specimens: in the West Indian variety (1 d) the basal[Pg 226] margin on the carinal side forms a distinct but obtuse angle with the spur. In all cases the crests for the depressor muscles are very feebly developed.
The Compartments have their radii developed to a rather varying degree, with their summits oblique; hence the orifice is toothed: the sutural edges of the radii have their septa barely denticulated; the sutural edges of the alæ are smooth. The basis, as with the other species of this section, is permeated by pores; yet I found one specimen, from the Cape of Good Hope, with the basis apparently solid, thus offering a very singular anomaly. In the specimen imbedded in sponge, the basis, as viewed externally, is concave; whereas in Acasta, which always inhabits sponges, the basis is highly convex or hemispherical.
The Mouth and Cirri resemble those of B. trigonus, and I can point out no distinguishing character.
With respect to the variety from the West Indies, I have seen two sets of specimens differing somewhat in external appearance, one set attached to a coral from St. Vincent’s, and another set to an Avicula from an unknown locality; at first I described these specimens, with some hesitation, as a distinct species, and I am very far from sure whether this would not have been the more correct course, although I am unable to point out any sufficient diagnostic characters. This form differs from the ordinary B. spongicola, in the walls being more rugged, stronger, and slightly or deeply folded longitudinally; in this latter case the shell in external aspect differs much from ordinary specimens of B. spongicola; but this is a variation so common that I dare not place any reliance on it. The colour is more purple; the summits of the radii perhaps rather less oblique. In the scuta the only difference is that the articular ridge seems rather longer, and the adductor ridge perhaps more prominent: in the terga, as already remarked, the basal margin on the carinal side does not slope so straight into the spur. These differences I consider all too slight to be of specific value. The difficulty in determining the nature of this variety is added to by its approach to B. trigonus in all those points in which it departs from the ordinary B. spongicola, so that for a short time I was even tempted to consider both these species as varieties of one form. But until B. trigonus is found with its scutum longitudinally striated, and with its tergum beaked, it can hardly be confounded with B. spongicola; for it should be observed that when in B. trigonus the rows of little pits disappear from the scuta, as sometimes happens, though rarely, yet these valves do not become longitudinally striated.
Balanus spongicola occurs, mingled with B. tulipiformis, in the Mediterranean, and by the external characters of the shell alone cannot be distinguished from that species; but the striated scuta and beaked terga suffice to separate them. Again, this species, at the Cape of Good Hope, occurs mingled with B. Capensis, and from the non-striped young varieties of that species, it can, externally, be distinguished only by the beak of the tergum not being sharp like a needle. I have seen a single, perfectly characterised specimen, with its opercular valves preserved, found by Mr. S. Wood in the Coralline Crag at Sutton, mingled with B. inclusus.
BALANUS LÆVIS. Bruguière. Encyclop. Meth. (1789), Pl. 164, fig. 1.[91]
------ DISCORS. Ranzani. Mem. di Storia Nat., 1820, Tab. 3, figs. 9 to 13.
------ COQUIMBENSIS. G. B. Sowerby, in Darwin’s Geology of South America (1846), Tab. 11, fig. 7.
[91] M. Deshayes, in his descriptions of the plates, considers this figure, I have no doubt erroneously, as that of B. perforatus, of Bruguière. The B. Coquimbensis of Sowerby, is a different species from the B. Coquimbensis, of Chenu, ‘Illust. Conch.,’ tab. 6, which latter is unknown to me.
Shell covered by brown membrane, or naked and white or pale purple; orifice small; radii very narrow. Scutum with one or two deep longitudinal furrows.
Var. nitidus (fig. 2): shell not covered by membrane, white or pale purple: orifice but slightly toothed: scutum generally with two furrows. Hab.—Chile, as far south as Concepcion; Peru; California.
Var. Coquimbensis (fig. 2 a): with the basal cup partly filled up with thin, irregular, calcareous layers, making a cancellated mass. Fossil, and recent.
Hab.—Strait of Magellan, ten to twenty fathoms, attached to shells; often entirely surrounding pebbles, forming globular masses; associated with Verruca lævigata. Chile and Peru, (generally var. nitidus), often attached to Balanus psittacus. California. Very common.
Fossil in an ancient tertiary formation (middle bed) at Coquimbo, Chile. In a recent deposit (var. nitidus) at the height of 1000 feet at Valparaiso; with Human remains at San Lorenzo, Callao, Peru.
I may premise that, having myself collected this species from the same locality, the Strait of Magellan, where no allied species occurs, attached to the same Mytilus and associated with the same Verruca, I feel confident that it is the B. lævis described by Bruguière; and there can hardly be any doubt that it is the B. discors of Ranzani. With respect to the old tertiary specimens from Coquimbo, named B. Coquimbensis by Sowerby, they differ from the recent in no respect, except in being considerably larger; and therefore I cannot consider them specifically distinct. At first I was unwilling to believe that the specimens with[Pg 228] a single very broad longitudinal furrow, and those with two rather broad, or with one narrow furrow, on their scuta, could belong to the same species; but I soon found that all these varieties occurred mingled together, and that they differed in no other respect whatever. Generally, however, all the individuals in the same cluster had the same variety of scutum,—thus adding one more to the many instances amongst cirripedes of variations common to whole groups of specimens. Still more unwilling was I to believe that var. nitidus and the common variety could belong to the same species. Their general aspect is totally unlike: var. nitidus has a smooth, clean, naked shell, either white or pale purple, somewhat globulo-conical, often with a nearly entire orifice; whereas the other common variety generally has a more steeply conical shell, with a toothed orifice, and is covered by a dirty brownish membrane. Moreover, though I have seen hundreds of specimens from Tierra del Fuego, I have not seen one specimen of var. nitidus, or even of an approach to it in appearance; and, on the other hand, var. nitidus is the common form in Chile and Peru; though I have seen one or two specimens of the membrane-covered variety from Valparaiso. Such facts strongly induced me to believe that these forms were specifically distinct; but upon careful examination I could find no other or more important differences than those just specified. Some specimens from northern Chile are in an intermediate condition; and from Concepcion, in the south of Chile, where the climate approaches in character to that of the more southern parts of the Continent, there are many specimens, in so intermediate a condition that I know not whether or no to rank them under var. nitidus. Thus I became convinced that these forms are only varieties. At Concepcion, some few specimens are pale purple, and yet are wholly invested by thick brown membrane, thus uniting the two extreme varieties. From California I have seen both varieties, but I do not know which is most common there. With respect to the great difference in aspect between the specimens from northern Chile and Tierra del Fuego, we shall hereafter see a strictly analogous case in Balanus flosculus.[Pg 229] Finally, I may add that B. lævis seems to represent in the southern hemisphere and on the west coast of North America, the B. perforatus of Europe and Western Africa.
General Appearance.—Shell conical, sometimes slightly globular; surface smooth (that is, not folded), either naked, and in that case white or pale purple, or covered by dirty yellowish-brown membrane. Orifice small, more or less toothed, rarely exceeding one third of the basal diameter. Radii very narrow, often not developed, the six sutures forming in all cases deep and narrow clefts. The largest recent, but much depressed, specimen which I have seen (from the Strait of Magellan) was three fourths of an inch in basal diameter; specimens growing congregated are often much elongated. I have seen one with the basal cup between two and three times as deep as the height of the compartments. Of the ancient tertiary specimens, the largest had a diameter of three fourths of an inch, and a total length of actually two inches (fig. 2 a); another of these fossils had a basal cup in depth equalling four fifths of the entire length of the shell and basis.
The scutum has either one very broad and deep longitudinal furrow (2 b), or two moderately broad and deep (2 e), or two narrow and deep, or less frequently one narrow and inconspicuous longitudinal furrow (2 f); rarely there is not one furrow; sometimes there are none towards the apex, whilst furrows have been formed in the lower part of the valve. In young specimens the furrows extend down to the actual basal margin, but in old specimens they often fall short of this, and, as a consequence, the furrows become crossed by one, two, or three calcareous ridges, which ridges at successive periods formed the basal margin of the valve. The external surface is covered by yellow membrane; and fragments of several successive opercular membranes are often attached to the zones of growth.
Internally the articular ridge is not very prominent, but is remarkable (2 c, 2 d) from its lower point being produced into a long, sharp, sub-cylindrical, free style (like the hinge of a common gate), which is generally broken off in disarticulating the valve from the tergum. The adductor ridge is either sharp and prominent or blunt: it extends up the middle of the valve nearly to the apex, and downwards it trends a little towards the occludent margin. The pit for the lateral depressor muscle is minute but deep: the basal margin is sometimes hollowed out under this pit. Sometimes there is a distinct, but blunt ridge, caused by one of the furrows outside, parallel to the adductor ridge, and placed between it and the little pit for the lateral depressor; in this case, the basal margin, as viewed internally, is rendered sinuous (2 d), as is best exhibited in the great fossil specimens from Coquimbo.
Tergum (2 g).—Spur of moderate length and breadth, with its lower end obliquely truncated and rounded. The longitudinal furrow has its edges somewhat folded in. The basal margin on the carinal side of the spur is sometimes a little hollowed out. The crests for the depressor muscles are well developed; but the corner of the valve supporting them is extremely thin, and is often imperfectly calcified.
Compartments.—The parietal tubes are not crossed by transverse[Pg 230] septa, but in their upper parts are filled up solidly. The radii are always very narrow, with their summits oblique, though to a variable degree: their sutural edges have fine and closely approximate septa, with minute denticuli: the sutural edges are received in a furrow, on the opposed compartment, of unusual depth; hence the lines of suture run, in the lower part of the shell, almost exactly in the middle between each two compartments. The alæ are added to above the level of the opercular membrane.
The Basis is often thick, with an underlying layer, largely cancellated or honeycombed. When many specimens grow crowded together, the basis is generally deeply cup-formed, or even sub-cylindrical; and equals as much as four fifths of the length of the entire shell. In such cases, in some few recent specimens, and in all the large or even quarter-grown old tertiary specimens, but not in the quite young fossil specimens, a structure is presented, which I have not seen in any other Cirripede, namely, the basis (Pl. 4, fig. 2 a) is filled up for one third, or even for more than half its depth, by successive, separate, calcareous, transverse layers or septa. It would appear as if the basal cup had grown too large for the animal’s body, and so required filling up. The layers are thin and fragile; a single layer never stretches across the whole shell; each is irregularly mammillated or blistered, with the convex surfaces generally directed upwards; the layers are furnished on their under sides with little pillars and short ridges, resting on the layers beneath; it rarely happens that the supports of one layer lie directly over those of another, though this is sometimes the case. In a vertical section, the mass formed by these irregular layers has a coarsely cancellated structure. This structure, although confined to this one Cirripede, is not so anomalous as might at first be thought, for in most species of the genus, each time that the circumference of the basis is added to, an excessively thin calcified film is thrown down over its whole inner surface; and in any of these species, if the films had been formed thicker and had rested only on certain points, instead of over the whole underlying layer, the cancellated structure above described would have been produced.
Mouth: the labrum is either destitute of teeth, or has two or three very minute teeth. The palpi have a tuft of very long spines at their ends. The third tooth of the mandibles is thicker and larger than the two upper ones. The maxillæ have either a nearly straight edge, or the inferior corner is obliquely truncated, and projects much beyond the rest of the edge. In the Cirri, none of the segments are very protuberant: in the first pair, one ramus is nearly twice as long as the other: in the posterior pairs, the segments are not much elongated, but each supports seven pairs of spines.
Var. nitidus: with respect to this variety I have little to add to my preliminary remarks on its peculiar appearance, owing to its smooth, naked condition, and pure white or pale purple colour. This colour, when examined through a lens, is seen to consist of very fine longitudinal stripes; and is produced by the calcareous matter within the longitudinal parietal pores being thus coloured. Generally the scuta have two longitudinal furrows; but I have seen a scutum of one perfectly characterised specimen with only a single broad furrow, like that[Pg 231] which frequently occurs in the membrane-covered variety. Var. Coquimbensis, as before stated, differs only in its greater size: the scutum, in the one specimen examined, had two broad longitudinal furrows; neither it, nor the tergum differed from certain varieties now found on the coast of Chile.
BALANUS PERFORATUS. Bruguière. Encyclop. Meth., 1789, Tab. 164, fig. 12 infra.
LEPAS ANGUSTA. Gmelin. Syst. Naturæ, 1789.
---- ORE ANGUSTIORE. Chemnitz. Vol. viii, Tab. 98, fig. 835.
BALANUS CORNUBIENSIS CONICO ORE MINORE. Ellis. Phil. Trans. vol. 50, 1758, Tab. 34, fig. 16.
LEPAS BALANUS ET FISTULOSUS. Poli. Test. Siciliæ (1795), Tab. 4, fig. 5, Tab. 6, fig. 1.
BALANUS COMMUNIS. Pulteney. Dorset Catalogue, 1799.
------ ---- Montagu. Test. Brit., 1803.
LEPAS ANGUSTATA. Wood. General Conchology, 1815, Pl. 6, fig. 5.
BALANUS CRANCHII. Leach (!). (B. Blainvillii in Tab.) Encyclop. Brit. Suppl., vol. iii, 1824.
------ ---- Brown. Illust. Conch., 1827, Pl. 7, fig. 9, 10, and 2d Edit., Pl. 53, fig. 9-12.
------ PERFORATUS. Chenu. Illust. Conch., Tab. 3, fig. 9, Tab. 6, fig. 15.[92]
[92] I have very little doubt regarding any of these references: I have no means of ascertaining the priority, within the same year, of Gmelin and Bruguière, but have given it to the latter, as perforatus is much the best known specific name. English conchologists seem generally to suppose that the B. communis of Pulteney and Montagu is the B. porcatus of this work; but I have not the smallest doubt that I have given it rightly as a synonym of the present species; the indistinctness of the compartments, the multitude of fine ridges, the smallness of the orifice, the longitudinal furrow on the terga, the colour, size, and habitat, all given by Pulteney or Montagu, will agree with no other British species. The Lepas balanus of Poli, which is certainly a synonym of our present species, has been erroneously considered by several authors to be the same with the L. balanoides of Poli, which latter undoubtedly is the B. amphitrite of this work.
Shell pale purple, or white, or dirty ash-colour; smooth, or, from being corroded, finely ribbed longitudinally; sheath purple; orifice generally small; radii generally narrow or absent. Scutum, internally, with a short minute ridge, parallel and close under the prominent adductor ridge. Tergum with the apex somewhat produced.
Var. angustus (Gmelin) Pl. 5, fig. 1 a: pale dull purple or white; orifice small or of moderate size; radii very narrow or moderately wide, white or pale purple, with oblique summits.
Var. Cranchii (Leach) Pl. 5, fig. 1 b: corroded, covered with fine longitudinal ridges owing to the exposed, filled-up, parietal tubes; dark dirty ash-colour, with a tinge of purple: radii not developed, or very narrow with oblique summits; orifice small.
Var. fistulosus (Poli) Pl. 5, fig. 1 d: shell cylindrical, white or dull purple; orifice of moderate size or small; basis deeply cup-formed.
Var. mirabilis, Pl. 5, fig. 1 c: bright purple; radii white, very broad, with their summits parallel to the basis; orifice entire, large.
Hab.—Southern shores of England; South Wales; Mediterranean; Western Africa, southward to Loanda, in 9° S.; West Indies (?). Generally adhering to rocks at a low tidal level; in one case attached to the floating Lepas Hillii, Mus. Jeffreys.
This is a well-marked species, and in its essential characters does not vary much; but owing to the shell being almost as often white as purple,—to its being remarkably subject to disintegration,—to its often becoming cylindrical,—to the radii being either not at all, or slightly, or moderately, or largely developed, and consequently to the orifice of the shell varying in size, the general external appearance of the different varieties is singularly diversified; but when a series of specimens is examined, it is easy to see how one form passes into another.
General Appearance.—Shell conical, with the orifice oval, unusually small, being generally only from one third to half of the basal diameter; sometimes moderately large; in one single instance as wide as the basis. Radii, often represented by mere lineal fissures, or they are narrow, or sometimes moderately wide. Colour pale, dull purple, sometimes lilac, often passing into a dead pure white: the same individual will occasionally have one part of its shell white, and another purple: the purple tint almost invariably is nearly uniform, or not in stripes. The radii are generally white, when the whole shell is purple, but sometimes they are pale purple: the sheath is apparently always coloured of a fine claret-purple, with the triangular portion of the alæ, added during diametric growth, generally white, but sometimes purple. The surface is quite smooth, but very often, especially on the shores of England, whole groups of specimens (excepting the very young ones,) have had the outer lamina of the parietes entirely corroded and removed; in this case the shell assumes a dirty, more or less dark, ash-colour, feebly tinted with purple, and the whole surface, owing to the exposure of the solidly filled-up parietal tubes, becomes finely striated, or covered with very narrow, longitudinal ridges. When specimens are crowded together they often become cylindrical, and[Pg 233] much elongated, owing to the basis becoming deeply cup-formed: I have seen specimens, half an inch in diameter in the widest part, one inch and a half in height, the walls forming only a third of this. The largest specimen which I have seen (from the southern shores of England) had a basal diameter of 1.2 of an inch; some very steeply conical specimens were .9 of an inch in height, and .8 in basal diameter.
Scuta, externally, slightly convex; growth-ridges approximate, moderately prominent. Internally (Pl. 4, fig. 3 a) the articular ridge is moderately developed, with the lower end produced downwards into a freely depending, flattened style, somewhat variable in size, but not so long as in B. lævis, and easily broken in disarticulating the valves. The adductor ridge is very prominent, running from almost the apex of the valve, close to the articular ridge, to near the basal margin. The basi-tergal portion of the valve is converted by the adductor ridge into a rather deep cavity, within which there is a short, sharp, and minute ridge, close and parallel to the adductor ridge, and bounding the impression left by the lateral depressor muscle: this insignificant ridge was present in every specimen; it occurs only in very few other species, as in B. nubilus and cariosus. The thickness of the valve sometimes varies a little, and when thick the adductor ridge does not appear quite so prominent. Tergum, with the apex moderately beaked and produced; beak triangular in section, coloured dark purple, as is the upper internal surface of the valve; the longitudinal furrow is deep, and has its edges folded in, and even quite closed. The spur is moderately long and narrow; but its width varies a little (Pl. 4, fig. 3 b, 3 c), and consequently it stands at either rather above or at twice its own breadth from the basi-scutal angle: its lower end is either bluntly pointed or square, and generally is feebly toothed on the under-side. The basal margin of the valve generally slopes a little, on both sides, towards the spur. Internally, the scutal margin is but slightly inflected: the articular ridge is but slightly prominent, and but little curved; in the upper part of the valve there are generally several very minute ridges, parallel to the articular ridge, on the side towards the scutum. The internal surface of the spur itself is sometimes concave. The crests for the carinal depressor muscle are barely developed. It may here be mentioned that on the opercular membrane many long spines stand rudely arranged in rows.
Parietes: the parietal tubes have not transverse septa; but are solidly filled up in their upper parts by dark-purple layers of shell. The radii, as already stated, are either not at all developed, or are extremely or only moderately narrow, with their summits more or less oblique: in Mr. Cuming’s collection, however, there is an unique specimen, var. mirabilis (Pl. 5, fig. 1 c) with the aperture of the shell as wide as the basis, with bright purple parietes, and white, very broad radii, having their summits parallel to the basis. The septa of the radii are finely denticulated, and the interspaces are filled up solidly. The alæ have very oblique summits, and their edges are finely crenated. Basis, flat, or deeply cup-shaped; there is often an underlying, coarsely-cancellated layer.
Mouth: labrum finely hairy, but without any teeth; mandibles, with the 4th tooth small; the 5th confluent, with the sometimes smooth, sometimes pectinated inferior angle. Maxillæ, rather broad, with a slight notch under the upper pair of spines. Cirri, first pair, with one ramus, having 29 segments, and above one third longer than the shorter ramus, having 17 segments; these latter segments are remarkable by the extent to which their upper front surfaces are laterally produced into projections, twice as long as the breadth of that portion of the segment which is articulated to the adjoining segment. These projections have a double row of serrated spines on their upper edge, and a beautiful radiating bundle at the end; the projections decrease in length, both in the upper and lower segments. The second cirrus (Pl. 29, fig. 4) has the segments (13 in number, in the same individual with the segments above enumerated) of both rami produced in the same singular manner as in the first pair. The third pair have only inverted conical segments, coloured darker purple than the other cirri. The sixth pair had in the same individual 31 or 32 segments, and therefore one or two more than in the longer ramus of the first pair.[93] The segments in the posterior cirri have their anterior faces shield-shaped, and bear 6 or 7 pairs of spines, with some minute intermediate spines. There is the usual point at the dorsal basis of the penis.
[93] Under the Genus (p. 190) I have given the numbers of the segments in the cirri of this species at successive ages.
Range.—This species is common on the southern shores of England and in the Channel Islands: the largest specimens which I have seen came from these quarters. The most northern point whence I have seen specimens, is Tenby, in South Wales. This species is common throughout the Mediterranean; I have seen specimens from Malaga, Sicily, Algiers, and Smyrna; thence it ranges down the western coast of Africa, as far south as the Gambia and Loanda, in 9° south latitude. I believe British specimens are more often corroded than those from further south. Amongst some old, ill-kept specimens in a box in the British Museum, marked “Kingston, Jamaica,” there were some of this species: also I received some specimens, marked “S. America,” from Mr. G. B. Sowerby: again, Ellis, in Phil. Trans., vol. 50, part 11, gives a figure (Tab. 34, fig. 15) of some specimens from the West Indies, which I believe to be B. perforatus: hence, it is in some degree probable that this species, like B. tintinnabulum, and amphitrite, and improvisus, may be found on both sides of the equatorial Atlantic. Balanus perforatus is attached, together with B. tulipiformis, trigonus, amphitrite, Chthamalus stellatus, and Pollicipes cornucopia, usually to rocks, near the lower limit of the tidal level; but I believe, from specimens kindly sent me by Mr. Mac Andrew, that it is frequently obtained by dredging; one specimen was even marked 30 fathoms. According to Poli, it is sometimes attached to the bottoms of vessels; and I have seen a specimen adhering to the floating Lepas Hillii.
Affinities.—This is a distinct species, closely allied to no other species, but comes nearest to B. lævis, which is its representative in[Pg 235] Southern America, and on the whole west coast of that continent. It is allied to that species, and differs from most other species, in the general form of the shell, its small orifice, narrow radii, and often deeply cup-formed basis. It agrees to a certain extent in the colouring, though the purple here is much more prevalent, and is not confined to the shelly matter filling up the parietal tubes. It agrees with that species in the general structure of the scutum; but the two or three deep, longitudinal furrows are here absent; and the minute ridge, parallel to and almost under the adductor ridge, is a peculiarity confined to this and very few species in the genus. The terga differ from those of B. lævis, chiefly in the spur being narrower, and in the apex being beaked. Lastly, the highly protuberant segments of the one ramus in the first cirrus, and of both rami in the second pair, are here remarkable. With regard to the varieties, I have nothing to add to their short diagnostic characters above given.
BALANUS CONCAVUS. Bronn. Italiens Tertiär-Gebilde (1831) et Lethæa Geognostica, b. ii, s. 1155 (1838), Tab. 36, fig. 12.[94]
------ CYLINDRACEUS, var. c. Lamarck. Animaux sans Vertèbres (1818).
LEPAS TINTINNABULUM. Brocchi. Conchologia Sub-Appen., t. ii, p. 597 (1814).
[94] I suspect that B. pustularis, miser, and zonarius, all figured by Münster, in his ‘Beiträge,’ b. iii, Tab. 6, may be this species.
Shell longitudinally striped with white and pink; or dull purple; sometimes wholly white. Scutum finely striated longitudinally; internally, adductor ridge very or moderately prominent.
Hab.—Panama; Peru; S. Pedro in California; Philippine Archipelago; Australia. Mus. Brit., Cuming, Stutchbury, Aug. Gould.
Fossil in Coralline Crag, England; Mus. Brit., S. Wood, Bowerbank, Lyell, J. de C. Sowerby, Tennant. Sub-Appennine formations, near Turin, Asti, Colle in Tuscany, Mus. Greenough, &c. Tertiary beds, near Lisbon, Mus. D. Sharpe and Smith. Bordeaux (?) Mus. Lyell. Tertiary beds, Williamsburg; and Evergreen, Virginia, Mus. Lyell. Maryland, Mus. Krantz. Recent formations[95] near Callao, Peru, Mus. Darwin. Red Crag (Sutton) Mus. S. Wood.
[95] I procured this specimen from the Island of S. Lorenzo, off Callao; it was imbedded, together with seventeen species of recent shells and with human remains, at the height of eighty-five feet.
This species has caused me much trouble. Looking first to the recent specimens, I examined several from Panama[Pg 236] and California, which, though differing greatly in colour, resembled each other in their scuta having the adductor ridge extremely prominent, and in having (Pl. 4, fig. 4 a), an almost tubular cavity for the attachment of the lateral depressor muscle,—characters which at first appeared of high specific value; but I soon found other specimens from Panama in which these peculiarities were barely developed. I then examined a single specimen from the Philippine Archipelago, resembling in external appearance one of the Panama varieties, but differing in the scuta being externally strongly denticulated in lines instead of being merely striated,—in the adductor ridge being far less prominent,—and in the spur of the tergum being broader and more truncated; I therefore considered this as a distinct species. I then examined a single white rugged specimen from the coast of Peru, which differed from the Philippine specimen in the shape of the well-defined denticulations on the scuta, and in some other trifling respects, and in the segments of the posterior cirri bearing a greater number of spines; with considerable doubt, I also named this as distinct. But when I came to examine a large series of fossil specimens from the Coralline Crag of England, from northern Italy, from Portugal, and from the southern United States, I at once discovered that the form of the denticuli on the scuta was a quite worthless character,—that in young specimens the scuta were only striated,—that the prominence of the adductor scutorum ridge and the depth of the cavity for the lateral depressor muscle varied much (as in the case of the recent specimens), owing apparently to the varying thickness of the valve,—that in the terga the spur varied considerably in length and breadth, the latter character being in part determined by the varying extent to which the edges of the longitudinal furrow are folded in,—and lastly, that in young specimens the basal end of the spur is much more abruptly truncated than in the old. Hence I have been compelled to throw all these forms, originally considered by me as specifically distinct, into one species. I must repeat that this considerable variation in the prominence of the adductor ridge, and in the depth of the pit for the lateral depressor muscle—the pit in some cases becoming even tubular—is a very unusual circumstance.
With respect to the fossil specimens[96] from the above-stated several distant localities, I consider them as certainly belonging to one species, though varying considerably in several points of structure. When compared with the recent specimens, they differ from them in often attaining a considerably larger size; in the parietes being often, but not always, longitudinally ribbed; and in the radii often having more oblique summits. On the other hand, considering the many points of identity between the fossil and the recent specimen, I have concluded, without much doubt, that they ought all to be classed together. I may remark that, in the Coralline Crag specimens, the spur of the tergum (Pl. 4, fig. 4 d), is unusually long and narrow; it is broader and shorter in the Italian specimens (4 e), and variable in this respect, in the United States specimens; the scuta of the Lisbon specimens are remarkable for the greater prominence of the adductor ridge, and for the depth of the lateral depressor cavity. Some of the specimens from all the several localities are identical with the recent ones from the coast of Peru. The walls of the shell in the Coralline Crag specimens, are generally ribbed longitudinally. I have entered into the above particulars, on account of, in the first place, its offering an excellent example how hopeless it is in most cases to make out the species of this difficult genus without a large series of specimens; secondly, as showing how the characters alter with age; and thirdly, as a good instance of the amount of variation which seems especially to occur in most of the species which have very extensive ranges.
[96] These will be fully illustrated in the monograph on the Fossil Balanidæ, to be published by the Palæontographical Society.
Some of the pink-striped Panama varieties, though having a somewhat different aspect, can be distinguished from certain varieties of B. amphitrite only by their scuta being longitudinally striated,—a character in this species variable in degree, and in most cases of very little value. Some of the other recent varieties are sufficiently distinct from B. amphitrite; and the great fossil Coralline Crag specimens, which stand at the opposite end of the series of varieties, with their ribbed walls, very oblique radii, and coarsely striated scuta, are extremely unlike B. amphitrite. With respect to the no[Pg 238]menclature of the present species, I have little doubt that I have properly identified the Italian fossil specimens with B. concavus of Bronn, who has given a very good figure of this species in his ‘Lethæa Geognostica;’ it must, however, be confessed that the longitudinal striæ on the scuta are not there represented. Considering the large size and frequency of this species in Europe and in the United States, it has probably received several other names, besides the two incorrect synonyms, quoted at the head of this description. I should add that the true B. cylindraceus (not var. C) of Lamarck, according to the plate given by Chenu in his ‘Illust. Conch.,’ is the B. psittacus of South America. I have seen in collections specimens of B. concavus labelled as B. tulipa of Poli (B. tulipiformis of this work),—a very natural mistake, without the opercular valves be carefully examined.
General Appearance.—Shell conical, often steeply conical; orifice rather small, with the radii narrow, and generally in the fossil specimens very oblique; surface generally smooth, sometimes rugged, and in the coralline crag specimens generally ribbed longitudinally, the ribs being narrow. Colour various, either dull reddish-purple with narrow nearly white, or wider dark longitudinal bands; or, again, pale rosy-pink with broad white bands; or lastly, wholly white. The radii are either darker or paler than the parietes. The opercular valves are either dark purple or nearly white. Pale pink and white stripes are visible on some of the Italian and Portuguese tertiary specimens; and in most of the fossils the sheath is tinged dull red.
Dimensions.—The largest actually recent specimen which I have seen, from the Philippine Archipelago, had a basal diameter of 1.2 of an inch; the Peruvian pleistocene specimen is 1.7 in diameter; specimens from the crag and from the Italian deposits, however, sometimes slightly exceed two inches in basal diameter, and three in height.
Scuta: these in young and moderately-sized specimens are striated, sometimes very faintly, but generally plainly, causing the lines of growth to be beaded; but often, in large and half-grown specimens, the lines of growth are extremely prominent, and being intersected by the radiating striæ, are converted into little teeth. As the striæ often run in pairs, the little teeth frequently stand in pairs, or broader teeth have a little notch on their summits, bearing a minute tuft of spines. In very old and large specimens, the prominent lines of growth are generally simply intersected by deep and narrow radiating striæ. In one case, a single zone of growth in one valve was quite smooth, whilst the zones above and below were denticulated. The valve varies in thickness, which I think influences the prominence of the lines of growth and the depth of the striæ. These striæ often affect the internal surface of the basal margin, making it bluntly toothed. The articular ridge is rather[Pg 239] small, and moderately reflexed: the adductor ridge (as already stated,) varies remarkably; in most of the Panama specimens, it is extremely prominent, and extends down to near the basal margin; in other specimens it is but slightly prominent, especially in some of the fossil specimens from Virginia. The cavity for the lateral depressor, also, varies greatly; it is often bounded on the side towards the occludent margin by a very slight straight ridge, which occasionally folds a little over, making almost a tube; this, at first, I thought an excellent specific character, but far from this being the case, the cavity often becomes wide, quite open, and shallow.
Terga, very slightly beaked; the surface towards the carinal end of the valve, in some of the fossil specimens, is very slightly striated longitudinally. There is either a slight depression, or more commonly a deep longitudinal furrow, with the edges folded in and touching each other, extending down the valve to the spur, and causing the latter to vary in width relatively to its length. When the furrow is closed in, the spur is about one fourth of the entire width of the valve, and has its lower end obliquely rounded, and stands at about its own width from the basi-scutal angle: when there is only a slight depression and no furrow (as is always the case with young specimens), the spur is broader, equalling one third of the width of the valve, with its lower end almost truncated, and standing at about half its own width from the basi-scutal angle. But the absolute length of the spur, also, varies considerably; it is often very long, compared to the whole valve. The basal margin on the carinal side is sometimes slightly hollowed out; when the furrow is closed, this latter side slopes towards the spur. Internally, the articular ridge and crests for the tergal depressor muscle are moderately prominent.
Parietes, the longitudinal septa sometimes stand near each other, making the parietal pores small. The radii have oblique summits, but to a variable degree; their septa are unusually fine, and are denticulated on their lower sides; the interspaces are filled up solidly. The alæ have their summits very oblique, with their sutural edges nearly or quite smooth. In most of the fossil specimens, and slightly in some of the recent specimens, the surface of the sheath presents an unusual character, in a narrow, longitudinal, slightly raised border, running along the sutures, on the carinal side of each compartment.
Basis thin, porose; sometimes with an underlaying cancellated layer.
Mouth: labrum with six teeth: mandibles with the fourth and fifth teeth small, either sharp, or blunt: maxillæ with a straight edge, or with the inferior part slightly prominent. Cirri with the rami of the first pair unequal by four or five segments: the segments in the shorter ramus are extremely protuberant. The segments in the second cirrus only moderately protuberant: but all the specimens were in bad condition, and it appeared as if, in the Panama specimens, the segments of the second cirrus were more protuberant than in the Philippine Island specimens. In the posterior cirri there are from three to five pairs of spines on each segment: even amongst the Panama specimens some had three and some four pairs, and a white Panama specimen had five pairs of spines.
All the recent specimens which I have seen, were, with one exception, attached to various shells and crabs, and to each other. The Peruvian specimen was associated with B. flosculus. The tertiary specimens are often congregated together into great masses. Including the recent and fossil specimens, this species encircles the globe. During the miocene period it seems to have been the commonest existing sessile cirripede; now, it does not appear to be common, excepting, perhaps, at Panama: Mr. Cuming procured only one specimen from the Philippine archipelago.
LEPAS RADIATA. Wood’s General Conchology (1815), Pl. 7, fig. 7.
---- MINOR? Wood’s General Conchology (1815), Pl. 7, fig. 6.
---- BALANOIDES. Poli. Testacea utriusque Siciliæ (1795), Tab. 5.
BALANUS BALANOIDES. Risso. Hist. Nat. de l’Europe Merid., tom. iv, 1826.
Shell longitudinally striped with purple or pink; sometimes with the stripes confluent; sometimes wholly white. Scutum internally with a prominent broad adductor ridge.
Var. (1) communis: (2 e, 2 h, 2 l,) nearly white, with pale or dark violet-coloured longitudinal stripes: epidermis rarely persistent: shell either thin or thick: radii white or freckled with reddish mahogany colour, with their summits either oblique, sometimes in a high degree, or nearly parallel to the basis: basal point of spur of the tergum either square or bluntly pointed. Hab. Mediterranean, W. Indies, S. Africa, Philippine Archipelago, New South Wales.
Var. (2) venustus: (2 a,) white or pale pink, with narrow bright pink, or broad pinkish-purple stripes; orifice either much dentated or nearly entire. Tergum with the carinal half of the basal margin sometimes much hollowed out. Hab. W. and S. Africa, Ceylon.
Var. (3) pallidus: (2 c, 2 k,) white, with or without a yellowish persistent epidermis; sometimes with the edges of the compartments tinted purple: radii moderately oblique: tergum generally narrow, with the spur sharp, and the basal margin on its carinal side much hollowed out. Hab. W. Africa, Madagascar, Red Sea.
Var. (4) niveus: (2 f,) white, with longitudinal hyaline lines; epidermis not persistent. Hab. W. Indies, Florida, S. Africa, &c.
Var. (5) modestus: upper part of shell white, lower part uniform blueish-gray, opercular valves as in Var. (1). Hab. unknown.
Var. (6) Stutsburi: (2 d, 2 i, 2 m, 2 n, 2 o,) white, with or without pinkish-purple stripes, which are often confluent, rendering the lower part of[Pg 241] the shell of a uniform purplish tint; epidermis persistent: radii very narrow: tergum narrow, spur sharp, varying in form and in exact position; carinal margin sometimes highly protuberant; basal margin on the carinal side of the spur generally, but not invariably, much hollowed out. Hab. West Africa.
Var. (7) obscurus: (Pl. 5, fig. 2 g,) with narrow, approximate, obscure and often almost confluent, slaty, or pale purplish-brown, or dark slate-coloured stripes. Hab. West Indies, Australia, and unknown.
Var. (8) variegatus: with narrow, approximate, dusky, claret-coloured stripes, transversely freckled with white; shell conical; walls very thin: scutum with the adductor ridge small. Hab. New Zealand.
Var. (9) (an. spec.?) cirratus: (fig. 2 b,) shell very pale purplish-brown, with faint, more or less plain longitudinal stripes, transversely freckled with white; walls thin: scuta with the lines of growth beaded: basis, in specimens growing in groups, irregularly cup formed: maxillæ with the inferior corner extremely prominent. Hab. Mouth of Indus, Australia, Philippine Archipelago.
Hab.—Warmer temperate and tropical seas; extremely common; Mediterranean, Smyrna, Sicily, Coast of Portugal; West Coast of Africa, River Gambia, West Indies, Demerara, Natal, Madagascar, Red Sea, Mouth of the Indus, Ceylon, Philippine Archipelago, East Indian Archipelago, Pacific Ocean, east coast of Australia, New Zealand; extremely common on ships’ bottoms; often attached to floating timber, canes, &c.; often associated with B. tintinnabulum; attached to pebbles and various shells.
With respect to the nomenclature of this extremely common species, which is widely distributed in all the warmer seas (excepting, as far as I have seen, on the west coast of America), there is some difficulty. I have no doubt that it is the Lepas radiata of Wood (1815), but Bruguière, in 1789, gave this same name to a Balanus which he had not seen, but which is figured in Chemnitz, Tab. 59, fig. 842. I should have thought that this also had been the present species, but Spengler, in describing (Skrifter af Naturhist. Selskabet i, B. 1790) this individual specimen, which he calls L. purpurea, states that it is 13 lines in basal diameter; now this is a size which is never acquired by B. amphitrite; and the description, habits, and size, would apply equally well to the species which I have called B. amaryllis; but when no notice is taken of such points of importance, as whether the walls are permeated by pores, whether the radii are smooth-edged, whether the scuta are striated, it is impossible to identify with any approach to certainty sessile Cirripedes; and the[Pg 242] names given ought, in my opinion, to carry little weight with them. With respect to Lamarck’s Balanus radiatus (1818), the synonyms quoted exhibit some great and inextricable confusion. The B. radiatus, again, of Risso, is a fossil and apparently distinct species. There can be no doubt that the present species is the Lepas balanoides of Poli, (and of several authors who have followed him), and equally little doubt that the present species is not the true L. balanoides of Linnæus, which has a membranous basis, and which I have not seen from the Mediterranean. Under these circumstances I have concluded that less confusion would be caused by giving a new name to this species than by taking that of Wood, which ought not to have been used by him, considering Bruguière’s previous adoption of it.
Under the head of B. tintinnabulum I have alluded to the great variation of B. amphitrite, which consists not only in a vast diversity in the colouring and in the general aspect, but likewise in the degree of obliquity of the summits of the radii, in the form of the terga, and slightly in that of the scuta. In order to show that it has not been from indolence that I have put so many forms together, I may state that I had already named and fully described in detail eight of the following forms as species, when I became finally convinced that they were only varieties: it would require at least thirty figures, which I have not the power to give, fully to illustrate the transitional forms. As with B. tintinnabulum, the deception is wonderfully enhanced by whole groups of specimens from the same locality exactly resembling each other, and sometimes differing from other groups attached to the very same object. If a person were to get together only some fifty or sixty specimens from only half a dozen different localities, he would almost certainly come to the same conclusion, as I at first did, that several of the varieties are true species; but when he gets several hundred specimens from all quarters of the globe, he will find, to his trouble and vexation, that character after character fails and blends away by insensible degrees, and he will be led, as the more prudent course, to include, as I have done, and I hope rightly, all under one specific name. I have experienced[Pg 243] more doubt regarding the last variety, cirratus, than on any other, on account of its peculiar colouring, and from the basis being often irregularly cup-formed. Under B. concavus I have remarked how closely some of its varieties approach to B. amphitrite, and it is to this last variety that they approach; almost the only difference being that the scuta in B. concavus are longitudinally striated. Yet some of the varieties of the two species are so distinct that it would be puerile to class them together. I will only add, that after studying such varying forms as B. tintinnabulum and amphitrite it is difficult to avoid, in utter despair, doubting whether there be such a thing as a distinct species, or at least more than half a dozen distinct species, in the whole genus Balanus.
As with B. tintinnabulum, I will first give a full description of the more common forms, alluding only to each less frequent variation, and then separately describe briefly the more marked varieties.
General Appearance.—Shape conical, either steep or considerably depressed; sometimes tubular; orifice either nearly entire or deeply toothed, not large, varying from rhomboidal to rounded-trigonal. Surface of shell smooth, never ribbed, generally naked, but occasionally the yellowish epidermis is persistent; in the same individual, I have seen all the lower part of the shell thus covered and the upper part naked, the line of separation being defined. The colour varies much, even sometimes considerably on the same individual; generally white or pale gray, with dull violet-coloured, longitudinal, moderately broad stripes; these stripes are sometimes equidistant, but more usually they are arranged so as to leave broad white spaces; the stripes fade away by endless variations, the edges of the compartments and the carinal end of the shell longest retaining any colour, until we have a uniformly white shell, generally covered with a yellowish epidermis; or the white is longitudinally marked with hyaline lines; this latter variety has a very peculiar aspect, and I did not doubt it was specifically distinct, until, in a number of specimens on a ship from the West Indies, I got the most perfect series, and another scarcely less perfect series from the Mediterranean, graduating into common coloured varieties. Rarely the dull violet or purple stripes become approximate and dark, so that the whole shell is tinted of a brownish slate-colour, occasionally freckled with white. Again, we have another set of very pretty varieties, with a white or very pale pink ground, with either narrow bright pink or broad pinkish-purple stripes. Again, I have seen numerous specimens of a variety, var. Stutsburi, from the west coast of Africa, in which the upper part of the shell is white, and the lower part shaded with[Pg 244] pinkish or dark purple approximate stripes, which often become confluent; in one group, the whole shell being thus uniformly coloured, without any vestige of stripes. I have seen another group from an unknown locality, in which the lower part of the shell was uniformly blueish-gray. A variety from Australia has narrow approximate dark claret-coloured stripes, transversely freckled with white. Lastly, in the variety cirratus, the whole shell is very pale purplish-brown, with indistinct longitudinal brownish stripes, transversely freckled with white lines. I considered this as a distinct species, until quite lately finding forms which I could not possibly determine whether to class as B. cirratus or amphitrite.
The radii are generally snow-white, or freckled with a bright mahogany tint, or rarely clouded with purple, or in the pink varieties with pink. The scuta are dull purple or pink, generally with a white band along their tergal margin; often, however, they are white, with merely one or two purple fasciæ. The thickness or strength of the shells varies much; some specimens attached to a floating cane, from Natal and the Philippine Archipelago, were extremely strong; others, from the Mediterranean and Australia, and some tubular varieties from the West Indies, were very thin, translucent, and fragile. Size: large specimens generally attain a diameter of from half to three quarters of an inch in basal diameter; and I have seen one or two specimens an inch in diameter.
Scutum; sometimes the surface is very smooth, but generally the growth-ridges are moderately prominent; the latter are occasionally very finely beaded, and this seems always the case with var. cirratus. Internally, the articular ridge is prominent and reflexed: the adductor ridge is sharp, very prominent, and straight; it runs parallel to the occludent margin; close to its lower side there is often a depression (Pl. 5, fig. 2 i), sometimes bounded by a slight ridge, as if for the attachment of a muscle, but there certainly is no muscle here: rarely the adductor ridge is only slightly prominent: there is a small and shallow little pit of variable depth for the lateral depressor muscle.
Tergum (2 k-2 o); this valve is here far more variable than in any other species: in the commonest purple-striped forms (2 l), the valve is rather broad, the basal margin lies in nearly a straight line on the opposite sides of the spur, which is placed at rather less than its own width from the basi-scutal angle; the spur is rather short, and in width about one fourth of the entire valve; its lower end is either bluntly pointed or more commonly nearly square (2 k) and parallel to the basal margin: in young specimens it is generally sharper than in older ones. Externally, in the line of the spur, there is either a slight longitudinal depression, or more rarely a deep furrow. The carinal margin is more or less convex, and is formed by upturned lines of growth: the scutal margin is broadly inflected. Internally, the articular ridge in the upper part is very prominent: the crests for the tergal depressor muscle are moderately prominent, but very variable. Sometimes the carinal portion of the basal margin is slightly hollowed out. In var. Stutsburi (2 m, 2 n, 2 o), and in some white varieties, which differ most in the shape of the tergum from the commoner varieties, the whole[Pg 245] valve is narrower, the spur is much sharper and narrower, the carinal half of the basal margin is much hollowed out and slopes down towards the spur, with the crests for the depressor muscles depending beneath the basal margin, and with the carinal margin sometimes extremely convex or protuberant. But the shape and position of the spur, and the outline of the carinal half of the basal margin vary much in nearly all the varieties.
Compartments.—The upper parts of the parietal pores are either filled up solidly with, generally coloured, shell, or they are crossed by thin transverse calcerous septa: the longitudinal parietal septa occasionally bifurcate at their bases close to the outer lamina, making an irregular outer row of minute pores. The Radii have their septa rather fine, and finely denticulated on both sides, but sometimes only on the lower side; the thickness of the septa varies a little; the interspaces are filled up solidly; the summits of the radii are jagged and oblique, and usually form an angle of about 45° with the basis, not being added to above the level of the opercular membrane; but not rarely they reach up much higher, and are very nearly parallel to the basis, extending from tip to tip of the compartments. Again, in some ordinary varieties, and always in var. Stutsburi, the summits of the radii are extremely oblique, the radii themselves forming a mere border to the compartments to which they belong. In no other species have I seen so great an amount of variation in the form of the summits of the radii. The alæ, in like manner, have their summits either very oblique, not being added to above the opercular membrane, or they are only slightly oblique; it often happens that in those specimens in which the summits of the radii are nearly parallel to the basis, the alæ are very oblique, and the converse: in other individuals, both radii and alæ have equally oblique summits. The sutural edges of the alæ vary in thickness, being either very thin and obscurely crenated, or moderately thick and ribbed. The basis is porose; but I have never seen an underlying cancellated layer of shell, as is so common in several species.
Mouth: labrum, with from four to eight, generally with six, little teeth: mandibles with three teeth, and two minute lower teeth, or mere knobs: maxillæ with the edge straight, or with the inferior part forming a slightly step-formed projection. Cirri: the rami of the first pair are unequal by three or four segments, but in some specimens by five or six segments, with the front surfaces of the segments in the shorter ramus extremely protuberant. The second pair of cirri are short, with the front surfaces of the segments moderately protuberant: the third pair have a tuft of bristles at their bases on the thorax. The segments in the sixth pair have from four to six pairs of spines on the segments; equal-sized specimens seem to vary in this latter respect. There is a small sharp projection on the dorsal base of the penis.
With respect to var. 1, communis, I have nothing further to remark, except that I have seen specimens identically similar from the Mediterranean,[Pg 246] Natal, the Philippine Archipelago, and Sydney; at the latter place it is said to be rare, but in most places it is the commonest variety, and is often attached to ships’ bottoms. Of var. 2, venustus, I have seen specimens from the west coast of Africa, Natal, and Ceylon, in groups by themselves, and associated with var. communis; it is much less common than var. 1. The third variety, pallidus, is not uncommon; I have seen many specimens from the bottoms of ships, from the West Indies, and the west coast of Africa. Of the var. 4, niveus, I have seen the most perfect graduated series passing into var. 1, both from the West Indies, Florida, and the Mediterranean: I have seen other specimens from the Red Sea and Madagascar. Of the var. 5, modestus, I have seen only one group from an unknown locality; it is only remarkable from its uniform colouring. The var. 6, Stutsburi, is more remarkable than the foregoing; until quite lately I did not doubt that it was specifically distinct; but as I have seen every character graduate into other varieties, I am now convinced that it is not a true species: all the specimens which I have seen have come on shells, or on ships’ bottoms, from West Africa. Of var. 7, obscurus, I have seen three or four groups of specimens from unknown quarters, both on pebbles, shells, and on cork (probably from the Atlantic ocean); and likewise some specimens taken from the bottom of Her Majesty’s ship “Fly,” on the east coast of Australia; these latter are intermediate in character with the next var. variegatus; from the Australian seas, which I at first ranked as an undoubted species, but I have subsequently failed in discovering any sufficient diagnostic character. Lastly, of var. cirratus, I have seen several groups of specimens from India and the Philippine Archipelago, and a group intermediate in character between this and the first and third varieties, from Australia; I retained this variety owing to its peculiar freckled, pale brown colouring and beaded scuta (of which, however, I have seen decided traces in the common variety), as a distinct species, after I had given up all the foregoing forms. I entertain some doubts whether I have now acted right; but when I found some specimens which, I found it impossible to decide, whether to rank as amphitrite or cirratus, I determined to take the more prudent course, and sink the latter as a species. This variety, also, seems to connect B. amphitrite and concavus very closely.
Shell dull red, freckled with white. Scutum internally without an adductor ridge; tergum with the spur, sharply truncated, almost one third of width of valve.
Hab.—West coast of South America, Mus. Cuming; attached to an Avicula.
The appearance of the fragile shell, in the one group of[Pg 247] specimens which I have seen, leads me to suspect that they may have grown under unfavorable circumstances. This species differs considerably in general aspect, but not much in essential characters, from B. amphitrite; the absence, however, of an adductor ridge to the scutum, and the sharply truncated spur of the tergum, are sufficient to distinguish them. In the opercular valves this species comes near to B. vinaceus, also from the west coast of South America; but the striated scuta of that species, the cancellated inner lamina of the parietes, the general colouring, and square porose radii, are amply diagnostic characters.
General Appearance.—Shell fragile, tubulo-conical, orifice large, passing from diamond-shaped into oval. Colour fine dark rose, freckled with transverse, sharply pointed, fine zig-zag white lines: the pink is also so arranged as to obscurely give to the walls a longitudinally striped appearance: radii generally rather whiter than the walls, and similarly freckled: terga similarly freckled: scuta dull red, with a white band along the scutal margin. Basal diameter of largest specimen half an inch.
Scutum, externally smooth: internally, articular ridge moderately developed, slightly reflexed: there is no adductor ridge: there is a distinct pit for the lateral depressor muscle. Tergum, with the scutal margin unusually prominent, toothed: longitudinal furrow shallow, the edges apparently having no tendency to fold in: spur short, barely one third of width of valve, with the lower end sharply truncated, parallel to the basal margin: articular ridge and crests for the depressores moderately prominent.
Compartments.—Walls very fragile, with the outer lamina not thicker than the inner lamina. Radii fragile, broad, with their summits moderately oblique; their sutural edges have the septa plainly denticulated on both sides, with the interspaces filled up solidly nearly to the tips of the septa. Alæ, with their summits more oblique than those of the radii; their sutural edges smooth. Basis with an underlying cancellated layer. Mouth: labrum with three unusually large teeth on each side of the notch: mandibles with the fourth tooth tolerably well developed, the fifth being confluent with the inferior angle. Maxillæ simple. Cirri,—first pair with one ramus longer by about four segments than the other ramus, which has considerably protuberant segments: second pair with segments only moderately protuberant: sixth pair with segments much elongated, but bearing only four pairs of spines.
BALANUS EBURNEUS. Aug. Gould (!). Report on the Invertebrata of Massachussetts, 1841, fig. 6.
Shell yellowish white. Scutum striated longitudinally: tergum with the spur truncated, the basi-carinal margin generally much hollowed out, and the carinal margin protuberant in the upper part.
Hab.—United States, from about lat. 42° to Charlestown; West Indies; Honduras; Venezuela; attached to shells and floating wood. Attached to ships’ bottoms from Trinidad and Jamaica, associated with B. tintinnabulum, amphitrite, and improvisus. Brackish water, Salem, Massachussetts, according to Mr. Stimpson. Mus. Aug. Gould, Agassiz, Stutchbury, Cuming, W. Dunker, &c.; very common.
General Appearance.—Shell conical, or almost tubular; white, with the surface very smooth, covered by thin yellowish epidermis, but with the radii naked. Orifice large, passing from rhomboidal into pentagonal, moderately toothed. Average full size, about one inch in basal diameter; I have seen a specimen 1.3 in basal diameter, and the same in height.
Scutum, plainly striated longitudinally: the teeth on the occludent margin small. Internally, the upper surface is roughened: the articular ridge is prominent, and either slightly or not at all reflexed: the pit for the adductor muscle is distinct; the adductor ridge is prominent in a variable degree, and is almost confluent with the articular ridge. In one specimen from Beverly Bay, U. S., the scuta were extraordinarily disintegrated, and I could perceive no trace of the external radiating striæ. Tergum, with the basal margin on the carinal side of the spur sometimes deeply (Pl. 5, fig. 4 b), and sometimes only slightly (fig. 4 d), and rarely hardly at all, hollowed out: when much hollowed out the valve may almost be said to be two-pronged, with the carinal prong narrower than the spur. There is no distinct longitudinal furrow, but the whole scutal margin projects above the general surface of the valve. In the carinal margin, in the upper part, there is a remarkable convexity or protuberance in the same plane with the valve, from which it is separated by a very slight and narrow ridge. The spur is about one fourth of the width of the valve, with its lower end abruptly truncated. Internally, the upper surface is much roughened with finely crenated ridges: the distinct crests for the depressores cover the whole of the so-called carinal prong.
Compartments; the radii and alæ have their summits oblique, sometimes a little rounded, but not smooth. The septa on the sutural edges of the radii are remarkably fine, and closely approximate; the[Pg 249] denticuli are excessively minute. The sutural edges of the alæ are most delicately crenated; the alæ are largely added to during the diametric growth of the shell, and above the level of the opercular membrane. The parietal pores are square and rather large: they are crossed by transverse septa almost close down to the basis: the longitudinal septa have tolerably large denticuli at their bases. The pores in the basis are crossed by numerous transverse septa. When specimens grow in a group, the basis is sometimes irregularly cup-formed.
Mouth: labrum serrated with small teeth, decreasing in size downwards, on each side of the central notch. Mandibles with the third tooth rather thick and blunt, and with the fourth and fifth knob-like. Maxillæ, with the inferior part projecting much beyond the rest of the edge, and bearing two long single spines: between these two spines and the large upper pair, there are, in a full-sized specimen, about seven pairs of moderately long spines, feathered on their sides. Outer maxillæ thickly clothed with very fine spines, and remarkably prominent.
Cirri: first cirrus, with one ramus having twenty-six segments, and longer by ten segments than the shorter ramus, which has sixteen segments: the shorter ramus, and both ramii of the second pair, have their segments remarkably protuberant in front; the protuberance, in the upper segments, equalling in length the supporting part of each segment: rami of the second cirrus unequal in length by five segments. Third cirrus with the segments only slightly protuberant; rami considerably longer than those of the second cirrus: at the dorsal base of the pedicel of this third cirrus there is no tuft of fine hairs, as is common in many other species. Sixth pair, with the upper segments elongated, bearing from six to seven pairs of spines; dorsal spines short, thin, and few.
Affinities: in external appearance of the shell, this species can hardly be distinguished from some of the white varieties of B. amphitrite; and there is a considerable resemblance, in some of the varieties, in the opercular valves; but the longitudinally striated scuta of B. eburneus suffice to distinguish these certainly very distinct species. Equally, or even more like externally, is this species to the B. Hameri, so that I have received from an eminent naturalist in the United States both species mingled in the same lot, all bearing the same name of B. eburneus; but when the internal structure of the shell is examined, the species are at once seen to be far removed from each other. Still more close is the affinity of this species to B. improvisus, both in internal and external characters: it agrees with this species in the singular habit of being able to live in brackish water: these two species are the only ones which have the labrum serrated with teeth, graduated in size, on each side of the central notch. In the case of young specimens of the var. assimilis of B. improvisus, an inhabitant of the same seas with B. eburneus, the diagnosis is most difficult without long practice; for in the young of eburneus, the compartments are only partially covered by yellow epidermis, and have a striped appearance, the radii are sometimes very oblique, the scuta externally have[Pg 250] not acquired their longitudinal striæ, and internally the adductor ridge lies not so close to the articular ridge as it does subsequently; hence I for some time mistook the var. assimilis of B. improvisus for the young of B. eburneus. But I found in the latter, that the rami of the first pair of cirri, are always, even in the earliest youth, more unequal in length, and that each segment of the posterior cirri bears a greater number of pairs of spines, there being, even in very minute specimens, seven pairs. Moreover, after having examined scores of specimens, I found I could almost always distinguish the two species by the smoothness and curvature of the summits of the radii of B. improvisus; I entertain no doubt whatever about the distinctness of the two species; indeed, when both are mature, besides the greater size, striated scuta, &c. of B. eburneus, their general aspect is very different.
Shell white: radii narrow, with their upper margins smooth, slightly arched, very oblique. Tergum with a longitudinal furrow; spur with the end rounded.
Var. assimilis, with longitudinal white hyaline lines.
Hab.—England, Scotland, Belgium (?), Nova Scotia, United States, West Indies, Rio Plata, Southern Patagonia, Guayaquil, West Colombia; attached to wood, shells, rocks, ships’ bottoms, from low tidal level to twenty fathoms depth.
General Appearance.—Shell conical, with a rather large diamond-shaped orifice, moderately or but little toothed; very smooth; walls never folded longitudinally; white, with an extremely thin pale-yellow persistent epidermis. The radii are very narrow, with their summits very oblique, rounded, and smooth; the epidermis is generally more persistent on the radii than on other parts, and this is exactly the reverse of what is common with B. eburneus. The specimens from nearly fresh-water in the R. Plata (hereafter to be mentioned), are brownish, and have undergone a remarkable degree of corrosion, the outer lamina of the walls having been entirely removed to near the base; hence the external aspect of these specimens is wholly different from ordinary individuals. The var. assimilis has also a very different appearance, owing to the dead white of the walls being relieved by narrow approximate longitudinal hyaline lines, corresponding with and caused by the longitudinal parietal septa being externally visible through the outer lamina of the parietes; the epidermis on the radii is also of a rather brighter yellow. The largest specimens which I have seen are those from the Plata, and those attached to a ship from the West Indies, and they had a basal diameter of .6 of an inch: from .4 to .5 of an inch is the more usual full average size.
Scuta, with the lines of growth but little prominent: articular ridge[Pg 251] prominent, but little reflexed: adductor ridge straight and very prominent, varying a little in its distance from the articular ridge; there is scarcely any depression for the lateral depressor muscle; the upper internal surface of the valve is roughened with ridges. Terga, with a moderately deep longitudinal furrow; spur short, rather narrow, with the end rounded, placed at less than its own width from the basi-scutal angle; in the Rio Plata specimens the spur is close to this angle: the basal margin is generally straight on opposite sides of the spur, but sometimes on the carinal side it is a little hollowed out. The lines of growth are upturned along the carinal margin, which consequently is a little protuberant, but to a varying degree. The crests for the depressores are extremely distinct and prominent. In the varieties having the basi-carinal margin hollowed out, and the carinal margin protuberant, there is a marked resemblance to the peculiar tergum of B. eburneus.
Walls: the parietal pores are tolerably large, and are crossed by numerous transverse septa: the longitudinal septa are very finely denticulated at their bases, but occasionally almost smooth. The radii are, as stated, extremely narrow, and very remarkable from their smooth rounded edges; their septa are barely denticulated. The alæ are remarkably protuberant; they have their summits much less oblique than those of the radii, and sometimes they are almost parallel to the basis: their sutural edges are coarsely crenated. Basis, flat, thin, permeated by pores, but the pores do not generally run to the very centre; they are, as usual, crossed by transverse septa.
Mouth: the labrum is the most remarkable part; on each side of the central notch there are generally two teeth; and on the two sides of the notch itself nine or eleven smaller teeth, decreasing regularly in size downwards till they become so minute as to be hardly visible even under the compound microscope; thus, in the two specimens closely examined, there were altogether twenty-two and twenty-six teeth on the labrum. Mandibles with the two inferior teeth reduced to mere knobs: maxillæ with the lower part of the edge bearing two large spines, and generally, but not always, forming a step-formed projection. Cirri: the ramii of the first pair are but slightly unequal; in one specimen examined there were fifteen segments in one ramus and twelve in the other: segments very protuberant in front. Second cirrus with the segments only slightly protuberant; segments thirteen. Third cirrus longer than the second pair, with the rami rather unequal in length: there is a tuft of long spines on the basal segment of the pedicel of this cirrus. Fourth cirrus twenty-two segments. Sixth cirrus, in the same individual, thirty-four segments: on each of these segments there are five or six pairs of spines. I may specify that the longer ramus of the first cirrus of a large Rio Plata specimen had twenty-four segments.
Varieties, affinities.—When I first met with the var. assimilis, misguided by its general aspect, I did not doubt that it was specifically distinct; I was strengthened in this view by the absolute identity of several hundred specimens attached to two vessels from Jamaica and Trinidad, in the West Indies, with one specimen from Charlestown, in[Pg 252] the United States, sent me by Prof. Agassiz, and with several in three lots from the western tropical shores of South America: yet on close examination I can point out no one distinguishing character, either in the shell or animal’s body, excepting the longitudinal hyaline lines on parietes, due to the septa being externally visible. The presence of similar lines is variable in white vars. of B. amaryllis and amphitrite, and they are seen in very young specimens of B. eburneus: hence it is impossible to consider so trifling a character as specific; moreover, lately I have seen a British specimen with hyaline lines, and some few other specimens in an intermediate condition. Under the head of B. eburneus, I have stated that although that species and B. improvisus, which in the West Indies are associated together, are most readily discriminated when old, yet when young, they so closely resemble each other that the eye requires much practice to separate them. On account of this species and B. crenatus being sometimes associated together on the shores of England, I have pointed out under B. crenatus, the relative diagnostic characters of the two. The chief affinity of B. improvisus is certainly towards B. eburneus; but in the narrow, oblique, rounded, and smooth-edged radii, there is a relationship shown to the species in the last section of the genus, such as B. amaryllis, and more especially to the fossil B. dolosus: so close is the resemblance in the external appearance of the shell, and in the structure of the opercular valves, to the latter species, that I for some time did not discover their distinctness. Balanus improvisus has hitherto been overlooked by naturalists, and has probably been confounded with B. crenatus or balanoides.
Range and habits.—This species, as far as my experience goes, is commoner on the shores of Kent than on other parts of England: the first specimens which I met with, I owed to the kindness of Mr. Metcalf, they were attached to wooden stakes from Herne Bay, together with a single specimen of B. crenatus: I have seen other specimens from near Woolwich, from the Kentish oyster-beds, from Sandwich, and from Ramsgate. The only other British specimens which I have seen are from the River Itchen, in Hampshire, and from Loch Shieldaig, in Ross-shire (Mus. Jeffreys), from a depth of twenty fathoms. This species is often attached to wood. At Ramsgate, the specimens were attached to a small coasting vessel, and they must have been immersed five or six feet; they were associated with B. crenatus, and with a few of B. balanoides. In the Brit. Mus. there are specimens collected by Mr. Redman, from Nova Scotia, in North America. When her Majesty’s ship Beagle was beached at Santa Cruz, in Southern Patagonia, numerous specimens were found adhering to her copper bottom, some so small as to show that the species breeds in those latitudes. Near Monte Video, in the estuary of La Plata, I found many large, but much corroded specimens, adhering to some rocks in a small running stream of perfectly fresh water. The rise of the tide is here small, but at high water the specimens apparently were for a short time covered by the waters of the estuary, here itself only brackish, and occasionally almost fresh. I took home some specimens, and placing them in perfectly fresh water they continued for many hours expanding and retracting their cirri with perfect regularity and vigour. Here then we[Pg 253] have a Balanus capable of living in fresh water, and likewise in the saltest seas: even brackish water is a deadly poison to several, probably to most, species of the genus; but this, as we have seen, is not the case with the allied B. eburneus. The water, I may add, at Woolwich, on the Thames, whence I have received B. improvisus, must at times be very brackish. I have already incidentally mentioned that the var. assimilis was attached in great numbers, associated with B. eburneus, tintinnabulum, and amphitrite, on vessels from the West Indies: one specimen sent me by Prof. Agassiz, from Charlestown, was attached to a specimen of B. eburneus; and, lastly, I have seen three sets of the same identical variety attached to shells from Guayaquil (in Mus. Brit. and Cuming), and from West Colombia. Here, then, we have the same species with an enormous range, from Nova Scotia and Great Britain to South Patagonia; and, which is the case with scarcely a single mollusc, it lives both on the eastern and western tropical shores of the South American continent.
Shell white, rugged: basis in parts imperfectly porose. Scutum with the articular ridge minute; adductor ridge prominent, forming a deep pit for the lateral depressor muscle: tergum with an internal patch of purple; apex produced, purple.
Hab.—California, Mus. Brit. and Aug. Gould; associated with B. glandula, and attached to wood.
I have seen two specimens of this species, brought by Lady K. Douglas from California; and two from Monterey, sent me by Dr. Aug. Gould. This is a very distinct species, coming nearer to B. porcatus than to any other species: it is also allied to B. cariosus. In the basis being in parts solid or not permeated by pores, it has claims to be placed in the next section, in which I at one time included it.
General Appearance.—Shell conical, rugged, sometimes furnished with sharp longitudinal ribs; dirty white. Orifice not large, oval, toothed. Radii rather narrow, with their summits oblique, much jagged. Basal diameter of largest specimen 2.1; height only 1.3 of an inch.
Scuta, broad, with the lines of growth prominent; internally, articular ridge very little prominent, sometimes hardly developed, but thick, ending downwards in a small free point. Adductor ridge[Pg 254] prominent, blunt, produced straight downwards, making a deep longitudinal cavity for the lateral depressor muscle; in some specimens this cavity is almost arched over, so as to tend to be tubular, with a short ridge in the middle (Pl. 6, fig. 2 a): in other specimens there is no trace of this tubular structure. Terga, with the apex beaked, beak triangular, dull purple; the longitudinal furrow is so shallow as hardly to exist. The basal margin slopes down on both sides, with a nearly equable curvature towards the spur; hence the spur is broad in its upper part, and narrow at its obliquely truncated lower end. Internally, there is an elongated dark purple patch: the shallow articular furrow is of quite remarkable breadth; the articular ridge is medial, and the inflected scutal margin is not wide. The internal surface of the spur is formed into a ridge, which runs a little way up the valve, and is sometimes partially separated from the spur itself (fig. 2 c), making the basal extremity toothed or double. The crests for the depressores are pretty well developed.
Walls, moderately strong: inner lamina slightly ribbed: the denticuli on the bases of the parietal longitudinal septa are sharp: I could not see any transverse septa in the parietal tubes. The radii are rather narrow; their summits are remarkably jagged and very oblique; the septa are plainly denticulated on both sides, but chiefly on the lower side; each septum itself, towards the inner lamina of the radius, branches and divides: the interspaces are filled up nearly solidly. The alæ have apparently their summits less oblique than those of the radii: their sutural edges are finely crenated. The lower edge of the sheath is hollow underneath. The basis is flat; it is rather thin, and imperfectly porose; in parts it is not at all porose, in others it is traversed only by very minute pores: there is nevertheless, in some parts, even where the upper layer is not porose, an underlying, cancellated layer.
Animal’s body unknown.
Shell white, longitudinally folded; radii narrow. Scutum internally without an adductor ridge.
Fossil, Sub-Appennine formations; Colle in Tuscany; Mus. Greenough.
I have seen only two specimens of this species attached to rock, collected by Mr. Greenough, at Colle, and kindly given by him to me. The species comes near to the living B. crenatus, also found fossil in deposits of this same age; it differs, however, distinctly from that species, in having its basis permeated by pores, and, in a less degree, in the sutural edges of the radii being more plainly crenated: the[Pg 255] opercular valves of the two species closely resemble each other. This may be the B. stellaris of Bronn, but it is futile attempting to identify the species of this genus merely by external characters, even when aided, as in this case, by an excellent drawing of the shell.
General Appearance.—Shell conical, with broad rounded longitudinal folds; orifice of moderate size, oval; radii narrow, with their upper margins oblique; but the summits of both specimens had been much broken. Colour, as it appears, originally white. Basal diameter of largest specimen 3/4 of an inch.
Scuta, with the upper portion much reflexed; the articular ridge is very prominent, and the articular furrow of great width; when the valve is viewed from the outside the articular ridge is very conspicuous: there is no adductor ridge. Terga, with the longitudinal furrow very slight; the spur is from one third to one fourth of the width of the valve, and its basal end is blunt and almost truncated; it stands about half its own width from the basi-scutal angle. Internally, the articular ridge is very prominent, and the articular furrow narrow and deep, extending down the valve in the line of the spur.
Parietes: the parietal tubes are remarkably large, and I think this can hardly be an individual peculiarity: the tubes are crossed by many transverse septa, close down to the basis. The radii are narrow, and have jagged, oblique summits: their sutural edges have very distinct septa, barely denticulated, with the interspaces filled up solidly. The alæ have oblique summits; I was unable to make out the structure of their sutural edges. The Basis is very distinctly permeated by pores, which are crossed by transverse septa.
The shell and opercular valves of B. corrugatus so closely resemble the same parts in B. crenatus, that I should not be much surprised at seeing the two species graduating into each other, if a larger series of specimens, from beds intermediate in age between the Sub-Appennine formations and the present time, were obtained. If indeed the basis of B. crenatus were permeated by pores, the two species could hardly be discriminated.
Parietes permeated by pores. Basis and Radii not permeated by pores.
BALANUS PORCATUS. Emanuel da Costa. Hist. Nat. Test. Brit., p. 249 (1778).
LEPAS BALANUS. Linn. Syst. Naturæ (1767).
---- ------ Born. Testacea Mus. Cæs. Desc., Tab. 1, fig. 4, (1780).
---- ------ Chemnitz. Syst. Conch., 8 Band., Tab. 97, fig. 820, (1785).
BALANUS ARCTICA PATELLIFORMIS. Ellis. Philosoph. Transact., vol. 50, Tab. 34, fig. 18 (1758).
------ SULCATUS. Bruguière. Encyclop. Method., Tab. 164, fig. 1 (1789).
LEPAS COSTATA and BALANUS. Donovan. British Shells, 1802-1804, Tab. 30, fig. 1, 2.
LEPAS SCOTICA. W. Wood. General Conchology, Pl. 6, fig. 3, sed non Lepas balanus, Pl. 7, fig. 3, (1815).
BALANUS ANGULOSUS. Lamarck (1818), in Chenu, Illust. Conch., Tab. 11, fig. 11.
------ TESSELATUS. Sowerby (!). Mineral Conchology, Tab. 84 (1818).
------ SCOTICUS. Brown. Illust. Conch. Great Britain, Pl. 7, fig. 2, sed non, Pl. 6, fig. 9 et 10 (1827); 2d edit., Pl. 53, fig. 1-3, 22, 23 et Pl. 54, fig. 1-3.
------ GENICULATUS. Conrad. Journal Acad. Philadelphia, vol. 6, part 2, p. 265 (1830), Tab. 11, fig. 16.
------ ---------- Aug. Gould (!). Report on the Invertebrata of Massachussetts, fig. 9 (1841).
Shell white, generally sharply ribbed longitudinally: radii with their summits almost parallel to the basis. Scutum longitudinally striated: tergum with the apex produced and purple.
Var. (a): Walls without longitudinal ribs. Mus. Brit., Cuming, Stutchbury, Jeffreys.
Hab.—South shores of England, Ireland, Scotland, Shetland Islands, Iceland, Davis’s Straits, 66° 30′ N.; Lancaster Sound, 74° 48′ N. (Mr. Sutherland). Maine and Massachussetts, United States. China (?). In deep water, common on shells, crustacea, and rocks, sometimes imbedded in sponges.
Fossil in the glacial deposits of Scotland, Uddevalla, and Canada; in the mammaliferous and Red Crag of England; Mus. Lyell, Sowerby, S. Wood, &c.
General Appearance.—Shell conical, somewhat convex; white, sometimes tinted yellowish, from the thin investing membrane; the produced tips of the terga are purple: the parietes of each compartment have from two to four strong, prominent, sharp, straight longitudinal ribs; these are sometimes irregular, and rarely, as will presently be described, they are absent. The radii are smooth and of considerable breadth; their summits are nearly parallel to the basis or only slightly oblique: hence the orifice is entire; it is rather small and ovate, being broad at the rostral end, and very sharp and narrow at the carinal end.
Dimensions.—The largest specimens which I have seen from Great Britain or Ireland, have been 1.3 of an inch in basal diameter: in Mr. Cuming’s collection, however, there was one much depressed specimen from the Shetland Islands, 2.1 in basal diameter: a regularly conical specimen from the coast of Massachussetts attained a nearly equal diameter; out of the glacial deposits in the Isle of Bute, Scotland, several specimens had this same diameter, namely, two inches, and were even more steeply conical, being 1.85 in height; some glacial specimens from Uddevalla and Canada, in Sir C. Lyell’s collection, were 1.7 in basal diameter. Hence, it appears, as we shall presently see is likewise the case with B. crenatus and Hameri, that northern specimens, and those from the United States and from the glacial deposits, often exceed in dimensions those from Great Britain or Ireland.
Scutum: the lines or ridges of growth are broad and prominent; they are divided into square beads by fine striæ, radiating from the apex: and hence the valve is longitudinally striated. Internally, the articular ridge is extremely little prominent; the adductor ridge, or what must be called such, runs straight down under the articular ridge, making a deep longitudinal pit for the lateral depressor muscle. Tergum: the apex is a little produced, and coloured purple, as well as the upper internal surface of the valve; there is no longitudinal furrow, only a very slight depression: the spur is placed close to the basi-scutal angle; it is rather long, and measured across the upper part, is half as wide as the valve: its lower end is truncated and rounded; the basal margin slopes towards it. Internally, a very small portion of the scutal margin is inflected: the articular furrow is shallow and broad: the crests for the depressores are feeble. In young specimens the spur is bluntly pointed.
The Parietes (4 e) have large square parietal tubes: in the upper part these are filled up solidly without transverse septa: the longitudinal septa are finely denticulated at their bases, and the denticuli extend[Pg 258] unusually close to the outer lamina. In very young specimens the inner lamina of the parietes is ribbed, in lines corresponding with the longitudinal septa, as is the case with most species of the genus; but in medium and large-sized specimens, there are between the ribs, thus produced, from one to four smaller ribs, which do not correspond with any longitudinal septa; they are finely denticulated at their bases, and may be considered as the representatives of longitudinal septa which have not been developed and reached the outer lamina. I have seen no other instance of this structure, namely, the presence of a greater number of ribs, on the inner lamina of the walls, than there are longitudinal septa. The radii have their summits generally parallel to the surface of attachment, as is usual in the first section of the genus, but sometimes they are slightly oblique: the septa sometimes rudely branch a little, but they exhibit scarcely a trace of denticuli: the interspaces are filled up quite solidly. The alæ have their summits very oblique; their sutural edges are finely crenated.
Basis, rather thin, translucent, not permeated by pores; obscurely furrowed in lines radiating from the centre: the circumference is marked in a peculiar manner by the longitudinal septa, and by the tips of those intermediate, denticulated ribs, which occur on the inner lamina of the parietes.
Mouth: labrum with six teeth: mandibles with the fourth and fifth teeth small and rudimentary: maxillæ, with a small notch under the upper pair of spines; in the lower part there is a single large spine. Cirri, dark brownish purple, making a singular contrast with the white operculum and shell; first pair, with one ramus, having twenty-six segments, and about twice as long as the shorter ramus, having twelve or thirteen segments, with their front surfaces protuberant. In the second pair the segments are but little protuberant: third pair about one third longer than the second pair: sixth pair, elongated, having in the same individual forty-six segments; these segments have shield-shaped fronts, bearing five pairs of spines, with some minute intermediate bristles. There is the usual point at the dorsal base of the penis.
Range: Geological History.—This species is common on the shores of Scotland and Ireland; the most southern point of Europe whence I have happened to see a specimen is Tenby, in South Wales: but I have no doubt it is found further south; and Mr. Jeffreys, who knows this species well, has found it common on the extreme southern shores of England. In the United States, it is found on the shores of Maine and Massachussetts: northward, I have seen specimens from Iceland, from Davis’s Straits, and from Lancaster Sound, in lat. 74° 48′ north; these latter I owe to Sir J. Richardson. It is an inhabitant of deep water; in Mr. Thompson’s collection there are several specimens from the Bay of Belfast, marked twenty-five fathoms, and one group said to have come from “about fifty fathoms, on the coast of Antrim:” one specimen from Cape St. Anne, Massachussetts, is marked as having come from only five fathoms. This species is commonly associated, on both sides of the Atlantic, with B. crenatus, and sometimes with B. Hameri and Verruca Strömia: mollusca, such as pectens, modioli, and oysters, offer the most usual surfaces of attachment: I have, however, seen many[Pg 259] specimens on crustaceans, on rocks, and even on the roots of the larger sea-weeds. This species is very common in the glacial deposits of Uddevalla, of Skien in Norway, and of Canada, and is associated with the same species as in the living state: I have seen, also, specimens from the same formation in the Island of Bute, Scotland. I have seen numerous specimens from the mammaliferous crag, and a few from the Red Crag of England. I owe to the kindness of Mr. J. de C. Sowerby an inspection of the original specimens of the B. tesselatus of the Mineral Conchology, which is certainly the present species.
Affinities.—This species is very distinct from every other; it comes nearest, as shown in all the characters derived from its opercular valves, to B. nubilus, and in this latter species we have seen the basis plainly tending to lose its pores and thus become solid. B. porcatus is perhaps allied in some degree to B. trigonus, and slightly to B. crenatus. The rather broad radii, with their summits hardly oblique, give this species a very different aspect from those species of the genus amongst which it must be placed.
Varieties.—A conical specimen, sent to me from the coast of Massachussetts, is remarkable from the radii not having been at all developed, being represented by mere fissures. I have seen a few specimens of var. (a), (one collected by Sir E. Parry in the arctic seas) which had a remarkably different aspect from the common forms, but which, after a careful examination of the opercular valves and of the animal’s body, I feel convinced are not specifically distinct: they are characterised by the walls being smooth and absolutely destitute of the external longitudinal ribs; by the shell being more cylindrical, with broader radii, and with the orifice larger and more rhomboidal; the walls and radii are much thinner, and the internal lamina is less plainly ribbed: the beak of the tergum is not purple. As most of these specimens had grown in a group crowded together, the difference of shape, and perhaps the thinness of the walls, is thus explained. In a specimen from Davis’s Straits, in Mr. A. Hancock’s collection, most of the above characters are in an intermediate condition; there are only a few external longitudinal ribs on the parietes; and the terga have not purple apices. In Mr. Cuming’s collection there are some fine, brilliantly white specimens (without opercula) from the coast of China; these have thin walls and radii, and the walls are not longitudinally ribbed, but they are not smooth: the orifice is not large, nor the shape of the whole shell cylindrical. It is just possible that these latter specimens may be a distinct and representative species, but I do not think so.
LEPAS PATELLARIS, (Gmelin). Spengler. Schriften der Berlin. Gesellschaft, &c. b. i (1780), Tab. 5; Chemnitz, Neues Syst. Couch., Tab. 98, fig. 839.
Shell depressed; brown, generally with obscure longitudinal[Pg 260] violet stripes: radii (in full-grown specimens) with their summits rounded and surfaces finely ribbed parallel to the basis: basis sometimes permeated by imperfect pores. Scutum internally with an adductor ridge.
Hab.—Bengal, on wood, Mus. Brit.; on a shell, Mus. Stutchbury; Philippine Archipelago (young specimen), Mus. Cuming. According to Spengler, on the Coromandel and Malabar coasts.
General Appearance.—Shell depressed, sometimes much depressed: orifice elongated, rhomboidal, but little toothed; surface smooth, but in old specimens sometimes with the walls slightly folded longitudinally. The radii are rather narrow, with their summits oblique; in old specimens their summits are rounded, and their whole surface finely ribbed parallel to the basis. Colour, in old specimens dirty brown, tinged with violet, sometimes in longitudinal bands, and with whiter irregular marks in the upper parts owing to disintegration: in young specimens the walls are regularly banded longitudinally, with violet-brown and dirty white; the radii being generally of a paler dirty red or violet. Basal diameter of largest specimen .9 of an inch.
Scuta, externally rather smooth; internally, articular ridge prominent, reflexed, with the lower edge hollowed out so as to be slightly hook-formed: adductor ridge small; there is a slight pit for the lateral depressor. Tergum, with the spur bluntly pointed, placed at about its own width from the basi-scutal angle; there is no longitudinal furrow, only a slight depression; carinal margin arched and protuberant: internally, articular ridge extremely prominent, running down in the direction of the middle of the spur: crests for the tergal depressores well developed.
Parietes, with the pores rather large; the internal lamina is very strongly ribbed, the ribs being but slightly denticulated at their bases: the parietal pores do not appear to be crossed by transverse septa: sheath closely attached to the walls. The radii have jagged oblique summits forming an angle of about 45° with the horizon; in old specimens they become more oblique and narrow: and are then very remarkable from their summits being arched and rounded, with a crenated edge, and with their whole surface transversely ribbed in horizontal lines; this is likewise the case with the recipient furrow in the opposed compartments: in young specimens the radii are externally quite smooth: the septa on the sutural edges are bluntly denticulated; the interspaces being filled up solidly. The alæ have their summits oblique, but much less oblique than the summits of the radii; their sutural edges are very finely crenated.
Basis thin, either quite solid, that is, not permeated by pores, but only furrowed in lines radiating from the centre, or permeated by pores towards the circumference, the pores being of very small diameter;—so that we here have an important character variable within the limits of the same species. Base flat, and this holds good, as remarked by Spengler, even when the specimens are attached to cylindrical pieces of wood.
Animal’s body unknown.
Affinities.—In the basis being sometimes permeated towards the circumference by pores, and by the colouring (the other species in this and the next section being dirty white), B. patellaris has almost as strong a claim to be ranked in the last as in the present section: in the rounded summits of the radii, and in the state of the basis, it, perhaps, shows more affinity to B. improvisus than to any other species; it is, however, almost equally allied to B. glandula.
B. CRENATUS. Bruguière. Encyclop. Method. (des Vers) 1789.
LEPAS FOLIACEA, var. a. Spengler. Skrifter af Naturhist. Selskabet, b. i, 1790.
---- BOREALIS. Donovan. British Shells, Pl. 160 (1802-1804).
B. RUGOSUS. Pulteney (?) Catalogue of Shells of Dorsetshire, 1799.
— ------ Montagu (?) Test. Brit. 1803.
— ------ Gould (!). Report on Invertebrata of Massachussetts (1841), fig. 10.
B. GLACIALIS (?) J. E. Gray. Suppl. Parry’s Voyage, 1819.
B. ELONGATUS (!), CLAVATUS (!), Auctorum variorum.
Shell white: radii with their oblique summits rough and straight. Scutum without an adductor ridge: tergum with the spur rounded.
Hab.—Great Britain, Scandinavia, Arctic Regions as far as Lancaster Sound, in 74° 48′ N. (Mr Sutherland); Behring’s Straits (Captain Kellett); United States; Mediterranean; West Indies, (Mus. Brit.); Cape of Good Hope, (Mus. Krauss). Generally attached to shells and crustacea in deep water; sometimes to ships’ bottoms. Very common.
Fossil in glacial deposits of Scandinavia and Canada, Mus. Lyell; in the mammaliferous, and Red, and Coralline Crags, Mus. S. Wood, J. de C. Sowerby, Bowerbank; Miocene formation, Germany, Mus. Krantz.
I find, in most collections, this species confounded with B. balanoides; I have even seen the two species, placed by Leach, on the same tablet in the British Museum: B. balanoides is, moreover, generally confounded with Chthamalus stellatus; nor has any one hitherto separated the present species from B. improvisus. On the other hand, trifling varieties, both of B. balanoides and B. crenatus, have commonly been considered as specifically distinct. From these[Pg 262] facts it will be seen in what confusion our commonest British species of Balanus have been left. After due deliberation, I have little doubt that this is the B. crenatus of Bruguière, and probably the B. rugosus of Montagu, but this latter author omits all reference to the really important diagnostic characters between this species and B. balanoides. The B. crenatus is certainly the B. rugosus of Dr. Aug. Gould. In various collections, I find specimens of B. crenatus, when coming from the arctic regions, called B. glacialis, arcticus, and borealis; though I have not met with an authentic specimen of the B. glacialis of Gray (‘Supp. Parry’s Voyage,’ 1819, p. ccxlvi), I have little doubt that it would prove to be the present species.
General Appearance.—White, usually of a dirty tint, from the yellowish or brownish persistent epidermis: conical, generally (fig. 6 a) with the parietes rugged and irregularly folded longitudinally; but sometimes much depressed and extremely smooth (6 b); often cylindrical and very rugged; occasionally club-shaped (6 c), the upper part being much wider than the lower: specimens in this latter condition sometimes have extremely narrow parietes, like mere ribs, and wide radii. The orifice in the cylindrical varieties is often most deeply toothed. The radii are generally narrow, and have jagged oblique summits; but not infrequently they are so narrow as to form mere linear borders to the compartments. The orifice is rhomboidal, passing into oval, either very deeply or very slightly toothed.
Dimensions.—The largest British specimen which I have seen was only .55 of an inch in basal diameter: specimens from Greenland and the northern United States frequently attain a diameter of three-quarters of an inch, and I have seen one single somewhat distorted specimen actually 1.6 of an inch in basal diameter. The specimens from the glacial deposits of Uddevalla and Canada appear, on an average, to attain as large or larger dimensions than those from the United States: on the other hand, the specimens from the mammaliferous and Red Crag are smaller, the largest being only .35 in basal diameter. When individuals have grown crowded together, their length is often twice, and even occasionally thrice, as great as their greatest diameter; thus I have seen a Greenland specimen 1.6 of an inch in length, and only .75 in diameter. In the British Museum there are some arctic specimens, one and a half inch in length, only half an inch in diameter at the summit (fig. 6 c), thence tapering downwards to a blunt point.
Scuta; the lines of growth are but little prominent: the surface is generally covered by disintegrating membrane. The upper ends are usually a little reflexed, so that the tips project freely as small flattened points. Internally, the articular ridge is highly prominent and somewhat reflexed: there is no adductor ridge, but a very distinct impression for the adductor muscle: the depression for the lateral depressor[Pg 263] muscle is small, but variable. The terga are rather small: the spur is short, and placed at rather less than its own width from the basi-scutal angle; the basal margin slopes a little towards the spur, of which the lower end is rounded or bluntly pointed in a variable degree. There is no longitudinal furrow, hardly even a depression. Internally, the articular ridge is very prominent in the upper part; the crests for the tergal depressores are well developed, but variable.
Compartments.—The internal carinal margin of each compartment, from the sheath to the basis, generally, but not invariably, projects a little inwards beyond the general internal surface of the shell, in a manner not common with the other species of the genus: the basal edge of this projecting margin rests on the calcareous basis, and is crenated like the basal edges of the longitudinal parietal septa. The whole internal surface of the shell is ribbed, but the ribs are not very prominent. The parietal tubes are large, and are crossed in the upper part, and often low down, by transverse thin septa: the longitudinal parietal septa are only slightly denticulated at their bases; occasionally they divide at the basis close to the outer lamina of the parietes, making some short outer subordinate pores. In the circular furrow beneath the lower edge of the sheath, there are sometimes little ridges, dividing it into small cells: sometimes, however, this furrow is filled up by irregular knobs of calcareous matter. The radii are always rather narrow, and often they form mere linear ribbons of nearly uniform width along the edges of the compartments. Their summits or edges are always more or less irregular and jagged: they form an angle with the horizon of generally above 40°. Their septa are fine, and barely or not at all denticulated. The alæ have oblique summits: their sutural edges are rather thick and distinctly crenated. Basis flat, calcareous, very thin, with the surface slightly marked by radiating furrows, which furrows answer to the radiating pores that occur in the bases of most species. In a club-shaped arctic specimen, one inch and a half in length, the summit being half an inch and the base only one fifth of an inch in diameter, the basis was still calcareous, thick, and not permeated by pores.
Mouth: labrum with six teeth: mandibles with the fourth tooth minute or rudimentary, and the fifth generally confluent with the inferior angle. Maxillæ with generally, but not invariably, a small notch under the upper pair of great spines. Cirri, first pair with the rami very unequal in length, one ramus being nearly twice the length of the other; in a large specimen having a cylindrical shell the proportional numbers of the segments in the two rami of the first cirrus were ten to twenty-three; in a small conical specimen the numbers were only eight to thirteen. The second cirrus has only two or three more segments than the shorter ramus of the first pair: the third cirrus has one or two more segments than the second; but it is nevertheless decidedly longer than the second. On the dorsal surfaces of both segments of the pedicel of the third cirrus, there is a tuft of fine spines. The segments of these three pairs of cirri are not much protuberant in front. The segments of the posterior cirri have, each, four, or five, or six pairs of spines. Penis, with a straight, sharp, short point on the dorsal basis.
Range, habits, &c.—I have received specimens from all parts of the coast of Great Britain and Ireland, generally attached to crustacea and mollusca, and never hitherto from rocks uncovered by the tide. This species is also attached to floating timber, sticks, fuci, and occasionally to pebbles at the bottom of the sea. Mr. Thompson has sent me specimens from twenty-five fathoms depth in Belfast Bay: others on a Pinna from about fifty fathoms on the coast of Antrim; others from between three and six fathoms attached to Laminaria digitata: there is a specimen in Mr. Jeffreys’ collection marked forty-five fathoms. It is often associated, both on the coasts of America and Britain, with B. porcatus, and though these species are so distinct, yet when both have their surfaces similarly affected by being attached, as is often the case, to large Pectens, it is not at first easy, by external characters, to distinguish them, except by close inspection of the terga, which in B. porcatus are beaked and purple. The B. crenatus is sometimes associated in deep water with B. Hameri. At Ramsgate, in Kent, I saw a rudder of a ship, in which the two or three upper feet were thickly coated with B. balanoides, and the two or three lower feet with B. crenatus and improvisus mingled, together with a few of B. balanoides: occasionally vessels are thickly encrusted with this species, but I have never seen an instance of its concurrence with B. tintinnabulum and amphitrite—the commonest species on ships coming from the south. I have seen specimens from Greenland, Baffin’s Bay, the coast of Labrador, and other specimens marked simply, “Arctic regions,” and, again, others from the shores of Maine and Massachussetts. The arctic specimens, and those from the northern United States, are larger than the British. I have seen one single minute specimen on a crab, marked as having come from the Mediterranean. In the British Museum, amongst some specimens of B. eburneus, ticketed as having been sent from Jamaica, there was a small group of specimens, differing in no one essential respect from the common varieties of B. crenatus: at first I concluded that this was an erroneous habitat, and that the specimens had really come from the United States, where B. eburneus, is found as well as in the West Indies: for it appeared to me exceedingly improbable that an animal which can exist in lat. 75° N. should inhabit the hot shores of Jamaica: but subsequently I have received a specimen from Prof. Krauss, collected by himself in Algoa Bay, which is perfectly characterised, and even has the little cells in the furrow under the sheath: so that I am compelled to admit this enormous range and capability of resisting the most extreme climates. That this species should live in the tropical seas is the more surprising, as the large size of the specimens in the northern seas and in the glacial deposits, might fairly have been supposed to have indicated special adaptation for a cold climate. The great geographical range of this species accords with its range in time from the present day to the Coralline Crag period.
The specimens from the glacial deposits which I have examined, chiefly in Sir C. Lyell’s collection, are very fine and large; they are often associated, like the now living individuals, with B. porcatus and Hameri: they come from the well-known formation of Uddevalla and[Pg 265] from Canada. There are well-characterised specimens in the mammaliferous Crag, at Bramerton and near Norwich, in Sir C. Lyell’s collection, and from Sutton and other places in the Red Crag of the eastern shores of England: these specimens are decidedly not only smaller than the glacial, but than the recent English specimens; for the largest Crag specimens which I have seen had a basal diameter of only .35 of an inch. The specimens which I have seen from the Coralline Crag, and some others sent me by Krantz from the miocene formation of Flonheim bei Abzei, in Germany, had not their opercular valves, yet I cannot doubt, considering how few species there are in the present section of the genus, that I have rightly identified them.
Diagnosis.—Under the head of B. balanoides I shall make a few remarks on the diagnosis between that and the present species; as B. improvisus is found on the British shores, sometimes mingled with B. crenatus, I may observe that, externally, the only difference consists in the edges of the radii in B. improvisus being much smoother and rounded, and in the whole shell being less rugged. Internally, in B. improvisus the porose basis, the presence of an adductor ridge on the under side of the scutum, the graduated teeth on each side of the central notch in the labrum, and the little inequality in length of the rami of the first pair of cirri, are clearly and amply diagnostic.
Shell white; parietes with the internal lamina generally strongly ribbed longitudinally, with the pores imperfect and small, sometimes in part absent; radii narrow, with their summits rounded. Scutum with an adductor ridge; tergum with the spur truncated and rounded.
Habitat.—California, Mus. Cuming, Aug. Gould; attached to shells and wood, together with B. nubilus. Southern Pacific ocean, attached to Pollicipes polymerus; Mus. Brit.
General Appearance.—Shell steeply conical, or cylindrical and elongated; dirty white; walls rugged, longitudinally folded; radii narrow, with their summits very oblique and rounded; orifice toothed. Basal diameter of largest specimen half an inch.
Scutum, resembling externally that of B. crenatus; rather broad, surface smooth; articular ridge very prominent, and articular furrow very wide; hence, when the summits of the opercular valves are worn down, the two scuta together form a square projection indenting the two terga, as in B. balanoides. Internally, there is a small adductor ridge, on the lower side of which there is a pit, as if for a muscle. The depression for the lateral depressor muscle is small, but variable. Tergum without any longitudinal furrow, and hardly a depression: spur[Pg 266] broad, with its lower end truncated and rounded; internally, articular ridge very prominent; crests for the depressores well developed.
Compartments:—The internal surface of the parietes is smooth in the upper part beneath the sheath, but generally very strongly ribbed in the lower part, the ribs being plainly denticulated at their bases; in other specimens, the ribs are very small, and even in parts quite obsolete. The parietal pores are short and imperfect, sometimes reduced to an extremely minute size, to be detected only when the walls are broken across near the basal edge, and most carefully examined; occasionally not even a trace of a pore exists. Hence in this respect, this species offers a singular case of variation. The radii are narrow, and of nearly the same width from top to bottom; their very oblique summits, when well preserved, are smooth and rounded; their sutural edges are ribbed or crenated with extremely fine, smooth septa; the recipient furrow is plainly marked by these septa. The sutural edges of the alæ are crenated; their summits are less oblique than those of the radii.
Basis, thin, finely furrowed in lines radiating from the centre; margin sometimes deeply sinuous.
Mouth: labrum with the central notch rather widely open, with four teeth on each side of it: palpi with very short spines along their inner margins: mandibles with the fourth and fifth teeth forming mere knobs: maxillæ small, with a mere trace of a notch under the two great upper spines. Cirri; first pair with the rami unequal by three or four segments, the longer ramus being only one quarter of its own length longer than the other ramus. Second pair short, with the segments (and those of the shorter ramus of first pair) somewhat protuberant. Third pair with the rami one third longer than those of the second pair. Sixth pair with the upper segments elongated, and bearing six or seven pairs of spines.
Affinities.—This species in general appearance closely approaches B. crenatus and balanoides, and it is related to them in many essential parts, such as in the opercular valves. It agrees with B. balanoides, and differs from B. crenatus, in the smallness and imperfection of the parietal pores, and in the radii having rounded summits; it agrees with B. crenatus in the structure of its basis, and in the prominent longitudinal ribs on the internal surface of the parietes, and differs from that species in the spur of the tergum being squarer, and in the scutum having an adductor ridge.
Range.—From the appearance of the Californian specimens, I suspect that they had adhered to tidal shells and to wood. The specimens in the British Museum, adhering to Pollicipes polymerus, consist of two lots, one of unknown origin, and the other certainly brought from the southern half of the Pacific Ocean by Sir James Ross: it deserves notice, that the Pollicipes polymerus, the supporting object, ranges from California to the southern Pacific Ocean.
Basis membranous.
LEPAS BALANOIDES. Linn. Fauna Succica, 1746, et Syst. Naturæ, 1767.
---- -------- O. Fabricius. Fauna Groen., p. 424, 1780.
---- -------- ET CLIVATUS. Montagu (!). Test. Brit., 1803.
BALANUS VULGARIS (?) Da Costa. Hist. Nat. Testacea, Pl. 17, fig. 7, 1778.
------ OVULARIS ET ELONGATUS. Aug. Gould (!). Report, Invertebrata of Massachussetts, figs. 7 and 8, (1841).
------ PUNCTATUS, CYLINDRICUS, ELONGATUS, FISTULOSUS CLAVATUS. Auctorum variorum. Sed non B. punctatus, Bruguière, Encyclop. Method., et non B. punctatus, Montagu, Test. Brit.
Parietes either solid, or cancellated, or rarely formed by a single row of pores. Tergum, with the spur bluntly or sharply pointed.
Var. (a) with the parietes permeated by tubes; spur of tergum sharply pointed; segments in the posterior pairs of cirri, bearing from eight to ten pairs of spines.
Habitat.—Great Britain, France, Norway, Shetland Islands; Greenland, according to O. Fabricius; North America, in lat. 66° 34′ N.; Labrador; Nova Scotia; Massachussetts, Delaware. Extremely common, attached to rocks, shells, and wood, within the tidal limits.
I have no doubt that the present species is the Lepas balanoides of Linnæus; though O. Fabricius is the only author who gives, in his “Fauna Groenlandica,” a sufficient description for the species to be recognised with certainty. I believe this also is the B. balanoides of Bruguière, though he is in error, as far as my experience goes, in stating that[Pg 268] the basis is ever calcareous. I have little doubt, also, that this is the B. vulgaris of Da Costa. The B. balanoides, in its corroded and therefore punctured state, is certainly the B. punctatus of most British collections; but I do not believe it is the B. punctatus of Montagu, which I have scarcely any doubt is the Chthamalus stellatus, so often found in the southern shores of England, and even in some of the best arranged collections, mingled with our present species.
General Appearance.—The shell, in middle-sized and old specimens, is almost invariably folded longitudinally and irregularly; it is either dirty white or very often pale brown, and punctured from the outer lamina having been corroded, to which action it is extremely subject. In very young specimens, the surface is usually quite white and smooth. The shell is sometimes much depressed; generally conical, but when crowded together, cylindrical or club-shaped, one specimen being even more than five-and-a-half times as long as wide. In Mr. Jeffreys’ collection there is a specimen 2.5 of an inch long, .45 in diameter at the summit, only .2 in the middle, and rather more than .2 near the base. Another specimen was 1.8 in length, its greatest diameter being .35 of an inch at the summit. On the other hand, I have seen a very depressed variety, with deeply folded walls, in Mr. Thompson’s collection from near Dublin, which was no less than four times as wide as high; so that the difference in proportion of height and greatest width, in the two extreme specimens, was nearly as 10 to 1. Occasionally, from some unknown cause, isolated specimens become cylindrical. The orifice of the shell, in the much elongated specimens, is generally deeply toothed. The radii are always narrow, sometimes extremely narrow, and have their summits smooth and rounded.
English specimens do not usually attain half an inch in basal diameter; I have, however, seen one from near Yarmouth .9 of an inch in diameter. Specimens from Massachussetts seem rather larger than the average size of British specimens, many being .6 of an inch, and one specimen a whole inch in basal diameter.
The opercular valves so closely resemble those of B. crenatus, that the description is necessarily comparative; in some cases they could hardly be discriminated; generally, owing to the disintegration to which this species is subject, the tips of the scuta are worn off, and hence the articular ridges together form (Pl. 7, fig. 2 a) a square projection, indenting the two terga; but I have examined young specimens and others when not disintegrated, in which the opercular valves, viewed externally, presented no difference whatever from those of B. crenatus. The scuta, however, are, I think, generally rather thicker, with the growth-ridges more prominent, and with the tips certainly less reflexed than is usual with B. crenatus. Internally, the articular ridge is rather less prominent: there is no distinct adductor ridge. The terga are often rather narrower in proportion, and this especially holds good[Pg 269] in the elongated varieties; in these latter, there is occasionally a moderately deep longitudinal furrow: the spur is often exactly the same shape as in B. crenatus, but it is apt to be rather longer (Pl. 7, fig. 2 c) and more pointed: in var. (a) it is pointed (fig. 2 d) in a very remarkable manner. Internally, the articular ridge is decidedly more prominent than in B. crenatus; the crests for the tergal depressor muscles are either well developed or almost absent. From this description it will be seen, how singularly the opercular valves of the common varieties of these two species resemble each other. I may mention that in some of the much elongated specimens, the muscles going to the opercular valves partially lose their transverse striæ, and become ligamentous.
The Parietes are either quite solid, or more commonly are permeated by minute pores, or by small irregular square tubes (Pl. 7, fig. 2 b), which only run up each successive zone of growth, for very short distances, giving to the shell a cancellated structure, which from corrosion is often externally visible. In the rather rare variety (a) the parietes are permeated by regular tubes, extending up to the apices of the compartments, but crossed by transverse septa. The longitudinal septa, when such can be said to occur, in no case are denticulated at their bases. The internal surface of the parietes is either quite smooth or is traversed (Pl. 7, fig. 2 b) by very slight anastomosing ridges, but never, even in var. (a), by regular longitudinal ribs, as in most other species. The carinal margin of each compartment, on the inside, projects, as in B. crenatus, inwards, beyond the general surface of the shell, and running down, rests on the basal membrane. The lower edge of the sheath is rarely hollow beneath. The walls are lined by purplish, or pale brown, or sometimes by almost black corium; numerous tubuli penetrate the under sides of the walls and opercular valves; and it is the intersection of these tubuli that gives the punctured appearance to the often corroded surface of the shell. The radii are narrow, generally very narrow; they have their upper and outer margins, as seen externally, very oblique, rounded and (when well preserved) smooth; their sutural edges are either quite smooth, or sometimes just perceptibly pitted, like the basal margin of the walls, or occasionally furnished with globular or arborescent little ridges. The alæ are also very oblique, but to a variable degree, sometimes only slightly oblique: their sutural edges are either smooth or obscurely crenated. Basis, membranous; in some much elongated specimens, during continued growth, the basal edges of the compartments approach each other so closely as almost to touch, so that the whole shell becomes pointed at the bottom; but on careful inspection I have never failed to find, even in the most pointed specimens, a minute basal membrane; in other much elongated varieties, in which the shell has apparently become too large for the animal’s body, the basal membrane, instead of being flat, becomes drawn up deeply inwards, so as to touch the surface of attachment only close round the basal edges of the shell.
Mouth: labrum with the teeth on each side of the central notch unusually variable in number; I have seen specimens with only two on each side, with four on each side, with five on one side and four on[Pg 270] the other, with five on one side and none on the other, and with six on both sides; hence the total number ranges from four to twelve. Mandibles, with the fourth and fifth teeth small, or quite rudimentary. Maxillæ, with scarcely even a trace of a notch under the upper pair of spines. Cirri; first pair, with one ramus one third or one fourth longer than the other; in one specimen the number of segments were nine and sixteen in the two rami: second and third cirri short, very nearly equal in length, having in the just-mentioned specimen respectively ten and eleven segments; the sixth cirrus in this same specimen had twenty-five segments, each segment being about as long as broad, and supporting six pairs of spines. In the singular variety (a) the posterior cirri are more elongated, and each segment supports seven or eight, and in one case even ten pairs of spines! the third pair is also in this variety proportionally rather longer. At the base of the third pair there is a tuft of fine spines. The penis has not, as in B. crenatus, a point at its dorsal basis. The branchiæ are very little plicated.
Of the varieties having much elongated, club-shaped, hour-glass shaped, and depressed shells, there is no necessity to say anything in particular. With respect to the remarkable variety (a), I at first named and described it as a distinct species: I have received two lots, both from North America, one being sent me by Professor Agassiz from Cape Cod. These agreed in having the parietes permeated by regular tubes; in having the spur of the tergum most sharply pointed; in the third pair of cirri being proportionally longer compared with the second pair; in the sixth pair having more numerous segments, namely, three times as many as in the third pair; in the segments of the posterior cirri being more elongated, and especially in the number of pairs of spines on each segment—amounting in one case even to ten, a number unparalleled in other cirripedes. It may naturally be asked why I have not retained so well marked a form as a distinct species? In the first place, I found the most remarkable character in var. (a), namely, the number of pairs of spines on the posterior cirri variable, there being in one lot seven or eight pairs, and in the other lot nine or ten pairs on each segment. Secondly, all the characters by which this variety differs from the common B. balanoides, are those which are variable in the latter; this is especially the case with the structure of the parietes, and in a lesser degree with the spur of the tergum. Thirdly, I found a specimen in Mr. Cuming’s collection, from Sweden (so that this var. (a) is not confined to North America), in which the cirri quite resembled those of the American specimens, but the spur of the tergum was in an intermediate condition as compared to that of ordinary varieties; and the parietal tubes were of unequal sizes, and scarcely more regular than sometimes in the true B. balanoides. And lastly, I have seen specimens from Ayrshire, with the parietes permeated by regular tubes, but with the tergum in an intermediate condition, and with the segments of the posterior cirri not more numerous or more elongated than in B. balanoides, supporting only six or seven pairs of spines, that is only one more than is common with B. balanoides; so that it was impossible to decide whether to rank the Ayrshire[Pg 271] specimen under var. (a) or under the common form, so that I was compelled to give up var. (a) as a species.
Monstrous individuals, with the male organs aborted: Parasite.—Amongst some specimens, chiefly elongated ones, sent to me from Tenby, in South Wales, I found no less than seven individuals with some of the posterior cirri distorted, unequal on the opposite sides, and in an almost rudimentary condition, and in each case with the penis truncated, without any muscle entering the stump, which was absolutely imperforate: the vesiculæ seminales were much shrunk; in one case without any zoosperms; in another case with headless zoosperms cohering in an unusual manner; hence it is certain that these individuals were functionally only female, and could not impregnate their own ova; yet in two instances the ova had been impregnated, no doubt by neighbouring perfect individuals, for they contained well-developed larvæ. Several of these monstrous individuals were infested by one, two, or three curious crustaceans, which have been described by Mr. Goodsir,[97] as the male of the Balanus; but these supposed males are females, and were distended with ova containing almost mature larvæ; I believe that they are the females of the unnamed genus, belonging to the family of Ioniens, described by Mr. Goodsir, which live parasitic within the sack (as I likewise found) of the same individual Balani.
[97] ‘Edinburgh New Philosophical Journal,’ July, 1843.
Diagnosis.—I have seen several specimens of this species and of B. crenatus, absolutely undistinguishable in external appearance. I may specify one of B. balanoides, imbedded in an alcyonidium, and one of B. crenatus, imbedded in a sponge, and therefore neither at all abraded. Generally, the tips of the scuta in B. crenatus are a little reflexed, whereas in B. balanoides, when the shell has been at all disintegrated, the tips form a square projection locked into the terga. Bal. crenatus never assumes the punctured appearance so common in B. balanoides. Very young specimens of the latter can be distinguished by their dead white colour and smoothness. The edges of the radii are almost always smoother than in B. crenatus, and they are never so wide as is sometimes the case with B. crenatus. When a specimen is disarticulated, our present species can at once be distinguished from B. crenatus (and from B. improvisus), by its membranous basis, and by the solid or cancellated walls, which are rarely permeated by regular tubes or pores; and the walls when porose are not internally ribbed. I have already pointed out the few very trifling points, in which the opercula of the two species differ. The mouth and cirri offer likewise very few differences: in B. balanoides there are often more teeth on the labrum than in B. crenatus; the rami of the first cirri are perhaps here rather less unequal; the second and third pairs of cirri are certainly in most cases more equal in length; and lastly, the segments of the sixth cirri, even in the common varieties, bear, in equal-sized specimens, more pairs of spines than in B. crenatus. We shall see that in habits, with regard to depth, the two species differ, B. balanoides inhabiting much shallower water than B. crenatus.
Range, Habitats, &c.—This species is extraordinarily abundant within[Pg 272] the tidal limits round the shores of Great Britain, and apparently of the northern United States. Besides numerous specimens sent to me from very many English localities, the late Mr. W. Thompson, of Belfast, kindly placed in my hands his very large collection; from these materials it appears that B. balanoides is the only tidal species in the northern parts of our island; but in the south and south-west, it is associated with the Chthamalus stellatus and Balanus perforatus. I doubt whether this species ever lives below the lowest tides; the case of a few specimens being mingled with B. improvisus and crenatus, (mentioned under the latter species,) at the bottom of a rudder of a small vessel, about six feet deep, is hardly an exception, for the water would there be troubled and aërated almost as in a breaker; and on this very rudder the upper two or three feet were coated exclusively by the B. balanoides. This species lives on rocks at both the uppermost and lowest limit of the tides; I am informed by Mr. Thompson, that he has seen specimens attached to a spot not covered by water during neap-tides. As a proof of its tenacity of life, Mr. Thompson informs me that he accidentally kept some specimens in a box, in a warm sitting-room, and found them alive seven days afterwards. This same most accurate observer finds, however, that B. balanoides is very susceptible to brackish water; he says, “that having kept some specimens alive for a week in excellent health, the water being changed once in thirty-six hours, they were one day killed instantly by some water, though brought from the same part of the estuary as usual, having been rendered brackish by much rain having lately fallen.” I may recall the fact, that B. improvisus lives daily for hours in absolutely fresh running water.
The B. balanoides lives attached, often continuously coating many square feet of the surface, to rocks, pebbles, wooden-piers, littoral shells and ulvæ. The most northern point whence I have received specimens, is in lat. 66° 34′ in North America, collected by Mr. Sutherland; and the most southern point is Delaware Bay, in the United States, in lat. 39°; I do not believe that this species extends into the Mediterranean, for Ranzani (Mem. di Storia Nat.), who particularly attended to the nature of the basis, was not acquainted with any Balanus having a membranous basis; and Poli (Test. Ut. Siciliæ,) describes only two species thus characterised, and these are manifestly Chthamali.
With respect to the rate of growth of this species, I am indebted to Mr. W. Thompson for the following note:—
“Sept. 29, 1848.—I examined a great number of Balani, in reference to the growth made by them during the present season, and found it to average three lines in diameter, and at most four lines. I saw a few minute specimens, only one line in diameter, showing that the species continued to breed until lately: these latter were probably not more than four weeks old. The young of the present year are plainly distinguished from the older ones, by their pure white colour and fresh appearance. Judging from the size of this year’s specimens, and of[Pg 273] the older ones on the same stones, I am of opinion that the term of life of the species is two years. Of the older shells, which I examined and found living in the spring, nine tenths are now dead, the walls only remaining, the opercular valves having been washed away.”
Mr. Thompson goes on to say, that the individuals which had, on July 3, a basal diameter of from two and a half to three lines, had attained, by the 30th of September, a diameter of four and a half lines, this being here the maximum size of the species.
LEPAS CARIOSA. Pallas. Nova Acta Acad. Scient. Petrop. tom. ii (1788), p. 240, Tab. 6, fig. 24.
Parietes thick, formed by several rows of unequal-sized pores. Tergum narrow, with the apex beaked, and spur sharply pointed.
Hab.—Columbia River, west coast of North America, Mus. Brit. and Cuming; Behring Straits (Capt. Kellett); the Kurile Islands, according to Pallas. Attached to shells, and to each other in groups.
General Appearance.—Shell steeply conical, with a rather small oval orifice; or cylindrical, with a large rhomboidal and little toothed orifice. Colour dirty white. Surface either simply rugged, or more commonly covered by numerous, narrow, extremely prominent, longitudinal plaits; from the manner in which these overlap each other, the shell almost appears as if thatched with straw. The upper corroded part of the shell usually exhibits a cancellated and finely punctured surface. The radii are generally very narrow, forming towards the base of the shell a mere narrow ribbon to each compartment, and often hardly distinguishable; but in one specimen they were of considerable width: in the former case, the alæ are often widely exposed. The largest specimen which I have seen was 1.5 of an inch in basal diameter, but Pallas gives 2.2 as the measurement of a specimen from the Kurile Islands.
The opercular valves are united to each other and to the shell by unusually strong membrane; and the upper parts of both valves, in all the specimens seen by me, have been much disintegrated. The Scutum, in old specimens, is faintly striated longitudinally, but in some there is hardly a trace of this: the occludent margin is furnished with a few large knobs, not corresponding with every alternate line of growth (as is usual with other species), but with every fourth or fifth line. Internally, the articular ridge is moderately prominent (in young specimens more prominent) and reflexed. The adductor ridge is sharp and[Pg 274] prominent; in the upper part it is confluent with the articular ridge, but in young specimens can be seen to be distinct; in the lower part it borders a large deep cavity for the lateral depressor muscle, in the middle of which there is a very slight longitudinal ridge; this cavity sometimes is almost closed or arched over in its upper part. In one specimen, the basal margin of the scutum was deeply hollowed out in the middle. The Tergum is remarkably narrow, with its apex produced into a triangular beak, hollow within, and sometimes faintly tinged purple. A deep, closed, longitudinal furrow runs down the valve. The spur is long, remarkably narrow, and pointed. Internally, the spur is produced up the valve as a ridge: the inflected scutal margin, and the prominent articular ridge, are both nearly straight, and parallel to the spur. The crests for the depressores are sharp and very prominent.
The Parietes are very thick and strong: unlike every other species of the genus, they consist of several very irregular rows, of unequally sized, round or angular tubes (3 b). These tubes or pores are generally short, and are at frequent intervals crossed by transverse septa; they often rather deserve to be called cells than tubes. New tubes are formed along the inner as well as along the outer lamina. They are lined by dusky purple corium. The internal surface of the parietes is smooth in the upper part, and in the lower, it is reticulated by slight, irregularly branching ridges. The carinal internal margin of each compartment projects a little, as in the case of B. crenatus and balanoides. The lower edge of the sheath is either hollow beneath, or is united to the walls. The radii in one specimen were broad, with slightly oblique, jagged summits; generally they are extremely narrow, forming more ribbons along the lower edges of the compartments, barely extending up as high as the sheath. They can sometimes hardly, or not at all, be seen, until the shell is disarticulated: in rather young specimens the sutural edge is sometimes quite smooth; in old specimens the lower part of the edge has coarse arborescent septa, with the interspaces filled up solidly, whilst the upper part is smooth. The alæ are conspicuous from the outside, owing to the little development of the radii; but owing to the diametric growth not having been great, the part added during such growth is narrow; the summits of the alæ are only slightly oblique: the sutural edge is coarsely crenated, with the teeth denticulated or slightly arborescent.
Basis membranous.
Mouth: labrum with only four very minute teeth: mandibles with four teeth; the third tooth broader than the first; the fourth small. Maxillæ, with the two upper spines placed on a slight prominence, beneath which there is a small notch. Cirri of a very dark colour (much injured): the segments of the first, second, and third pairs very broad and short, protuberant in front, and most thickly clothed with spines; the third pair is very little longer than the second pair: the sixth pair (in a large specimen above one inch in basal diameter) had the segments broader than long, each furnished with seven pairs of spines.
Affinities.—This species, though very distinct, evidently comes near to B. balanoides, especially to var. (a). By merely doubling or[Pg 275] trebling the irregular rows of short tubes in the walls of B. balanoides, with their reticulated inner lamina and longitudinally folded outer lamina, we should have the structure exhibited in B. cariosus. We have seen, also, that in var. (a) of B. balanoides, the spur of the tergum is remarkably sharp, as in B. cariosus. This species, also, in a very marked manner approaches in many characters, especially in the opercular valves, in the cirri, and to a certain extent in the shell and basis, to B. flosculus, and even in external appearance to var. sordidus of the latter—an inhabitant of the opposite extremity of the continent, namely, of Tierra del Fuego. Again, the tergum to a certain extent, and the scutum in a singular manner, resemble these valves in B. nubilus, showing an unequivocal affinity to that species. With respect to the most remarkable character of the species, namely, the several irregular rows of tubes or pores in the walls, it deserves notice that in B. crenatus, which is certainly closely allied to B. balanoides, the longitudinal septa sometimes divide near the outer lamina, thus giving rise to a few additional tubes. Of the above several species, to which our present species is allied, B. flosculus stands in the next section, and B. nubilus and crenatus in the last: hence we see that B. cariosus has singularly divergent affinities. The peculiar structure of the parietes, together with the general appearance of the shell, made me at the first moment suppose I was examining a Tetraclita (or Conia of Leach); hence, also, it has arisen, that Lepas cariosa of Pallas has often been quite erroneously given as a synonym of Tetraclita porosa.
Parietes solid; rostrum nearly twice as long as the carina or carino-lateral compartments, hence the basis is oblique. Tergum with the spur truncated, half as wide as the valve.
Hab.—West Indies; Mus. Brit.—Jamaica, imbedded in a sponge; Mus. Cuming.
This is a remarkable species; when first seeing it imbedded in numbers in a sponge, I did not in the least doubt but that it was an Acasta: on examination, however, it is found to have a membranous basis, and therefore cannot by the definition enter into that sub-genus, to which, however, it is very closely allied. It differs from other sessile cirripedes very remarkably in the rostrum being nearly twice as long as the carinal compartments, so that the basis is always very oblique, or placed almost on one side; in this elongation of the rostrum, although in a[Pg 276] different direction, we are reminded of B. calceolus and its allies; and these latter we know can hardly be separated from certain species of Acasta. Hence the position of our present species in this section, is not natural; but I am unable to place it elsewhere, without breaking down every definition: it should stand somewhat isolated, on one side of a line of affinity connecting Balanus calceolus and navicula with Acasta purpurata.
General Appearance.—The shell is thin, fragile, smooth, and white, but covered to a considerable extent by a brown membrane, which on the sheath and opercular valve is of a bright tint, and clothed with bristles. Viewed laterally, the rostrum is seen to be considerably bowed, and from its being nearly twice as long as the other compartments, with its lower end bluntly pointed, the basal margin of the whole shell is rendered very oblique, forming a slightly concave line. The lateral compartments are rather longer, and about one third broader than the carino-lateral compartments. The rostrum, from terminating downwards in a blunt point, instead of being square or truncated, as in all other Cirripedes, and from the upper end being, as is usual, pointed, has, when disarticulated from the other compartments, the shape of a boat. The parietes are not at all porose: their internal surface sometimes shows traces of longitudinal ribs, but sometimes there are none. The radii are narrow, with their summits very oblique, and their sutural edges smooth. The sutural edges of the alæ are likewise smooth. The largest of Mr. Cuming’s specimens was .2 of an inch in diameter; but a disarticulated specimen in the British Museum must have been larger, having a rostrum .3 in length. The Basis is membranous.
The Scuta are rather convex; they have their lines of growth approximate, most finely crenated, so as to be very feebly striated longitudinally. Internally, the articular ridge is pretty well developed, its lower edge being very oblique; there is barely an adductor ridge: the pit for the lateral depressor muscle is deep. The spur of the Tergum is placed close to the basi-scutal angle of the valve; it is about half as wide as the valve, with the lower end truncated: sometimes it may be rather said to be bluntly pointed, owing to its carinal side sloping up to the basal margin. The articular ridge is pretty well developed. The crests for the depressor muscles are barely discernible.
Animal’s body unknown.
Parietes and radii not permeated by pores; basis sometimes permeated by pores, sometimes not permeated, sometimes excessively thin and hardly distinguishable.
LEPAS HAMERI. Ascanius. Icones rerum naturalium, Tab. 10, 1767.
---- TULIPA. O. F. Müller. Prodromus. Zoolog. Dan. 1776; sed non L. tulipa, in Poli, Test. ut. Siciliæ; neenon B. tulipa, in Bruguière, Encyclop. Method; neenon B. tulipa, in Sowerby, Genera of Shells.
---- TULIPA ALBA. Chemnitz. Syst. Conch. Tab. 93, fig. 832.
---- FOLIACEA. Spengler. Skrivter of Naturhist. Selskabet, 1 B. 1790.
BALANUS CANDIDUS. (Tab. emendata) Brown. Conch. Great Britain (1827), Tab. 6, figs. 9 and 10, et 2d edit. Tab. 54, figs. 9-12.
------ TULIPA. Lyell.[98] In Phil. Transact., 1835, p. 37, Tab. 2, figs. 34-39.
[98] Sir C. Lyell remarks that this is apparently the B. Uddevallensis (Linn.) of Swedish lists of fossils. Prof. E. Forbes has shown (Mem. Geolog. Survey of England, vol. 1, p. 364) how this name arose, from a short description, prior to the introduction of the binomial system, “Lepas quæ Balanus Uddevallensis,” given by Linnæus in his Wast-Gotha Resa, in 1747.
For the reference to Ascanius’ work, which is on the binomial system, and subsequent to the 10th edit. of Linnæus, in 1758, I am greatly indebted to Mr. Sylvanus Hanley. Had it not been for this gentleman, I should have used Müller’s name of B. tulipa as the first name.
Shell white: radii with their oblique summits smooth and arched; sutural edges smooth: basis solid. Scutum feebly striated longitudinally: tergum with the spur narrow.
Hab.—Coast of Yorkshire; Scotland; Galway, Ireland; Isle of Man, and Anglesey, twelve fathoms. Generally in deep water; not very common. George’s Bank, Massachussetts, United States; Mus. Aug. Gould. Iceland, Finmark, and the Faroe Island, according to Spengler. Attached to crustacea,[Pg 278] mollusca, stems of fuci, and stones; often associated with B. porcatus and crenatus.
Fossil.—In glacial deposits at Uddevalla in Sweden, and Beaufort in Canada; Mus. Lyell. Banks of the Dwina, Russia; Mus. Murchison. Greenland, “in blue clay,” according to Spengler. Red Crag (Sutton) Mus. S. Wood.
General appearance.—Shell tubulo-conical, very smooth, white, generally more or less covered by yellow thin membrane: orifice large, sub-triangular: radii moderately broad, with their more or less oblique summits slightly rounded and smooth; from this circumstance the shell has been justly compared to the half-opened flower of a white tulip. Specimens often exceed an inch in basal diameter; I have seen one from Scarborough two inches in diameter and one and three quarters in height: another specimen was 1.6 in diameter and 3 in height. The specimens in the glacial deposits seem even to have acquired larger dimensions, one from Uddevalla being nearly four inches in height.
Scuta, elongated, flat, feebly striated longitudinally: internally, articular ridge short, moderately prominent: adductor ridge, confluent in the upper part with the articular ridge, running straight down and forming a rather large cavity for the lateral depressor. Terga feebly striated longitudinally, with a longitudinal furrow, having the sides, in old specimens, partly closed in: the basal margin slopes much towards the spur, which is rather long and narrow, with its end rounded: it is placed at about its own width from the basi-scutal angle. Internally, articular furrow narrow; crests for the depressores moderately prominent, but in a variable degree.
Compartments: these are unusually thin, and separate easily: the parietes are finely ribbed longitudinally on their insides; the bases of these ribs being just perceptibly denticulated. Radii, with their summits oblique (usually at about an angle of 45°), slightly arched and quite smooth: the smoothness is produced by the edge being a little inflected: sutural edge quite smooth, without even a trace of septa or denticuli. Alæ oblique, generally rather less oblique than the summits of the radii: sutural edges smooth, with an excessively fine linear furrow running along the edge, a little towards the inner side, and filled with a yellow ligamentous substance: a furrow of this kind I have seen in no other species.
Basis, solid, not permeated by pores; either smooth, or slightly furrowed in lines radiating from the centre.
Mouth: labrum with scarcely perceptible minute bead-like teeth thinly scattered along the edge. Palpi and outer maxillæ rather sparingly clothed with hairs. Mandibles with teeth rather sharp; the fourth and fifth teeth small, but well developed; inferior angle pointed with fine spines. Maxillæ with a deep notch under the two upper great spines. Cirri, the first pair is short, with rami of nearly equal length: the segments are not protuberant in front either in the first or second pairs. In the posterior cirri, the segments bear four pairs of spines, with a tuft of rather long intermediate spines: in young specimens there are only three pairs: the spines in the dorsal tufts are short and thin.
When the shell is disarticulated, this species cannot be confounded with any other; but judging by external characters alone, it may[Pg 279] sometimes be very easily confounded with B. eburneus, and I have received the two species under this one name from Massachussetts: generally B. Hameri may be distinguished from B. eburneus by the smoothness of the summits of its radii, and by the so-called epidermis being of a darker yellow.
With respect to the fossil specimens from the glacial deposits, I have little to add; I have seen one from Uddevalla, as already remarked, four inches in height, and a lateral compartment broader by one fourth than the same compartment in any recent specimen. As Sir C. Lyell remarks (Phil. Transact.), the compartments are always found separated, which is accounted for by their weak union in a recent state. This species, when fossil, is usually associated with its deep-water congeners B. porcatus and crenatus, as at the present day.
I must here mention that I have examined a considerable number of separated compartments, without opercular valves, brought from Barbados, in the West Indies, showing the existence there of a closely allied or possibly identical species. The only difference which I can point out in these compartments is, that the parietes are rather thicker, and the radii rather narrower, with more oblique summits: some of the compartments are two inches in length. It seems very improbable that the true B. Hameri should extend to the West Indies, but after what has been seen in the case of B. crenatus, this is possible.
Shell striped or clouded with pinkish-purple, or quite white; radii narrow, with their oblique summits smooth or arched: basis porose. Scutum plainly striated longitudinally: tergum with the spur narrow.
Var. (a):[99] bright rosy pink, not distinctly banded longitudinally. Hab. North-east coast of Australia.
Var. (b): snow white, glossy; orifice deeply toothed.
[99] This variety perhaps is the B. roseus of Lamarck, as figured in Chenu, ‘Illust. Conch.’ Tab. 2, fig. 9; but as Lamarck does not even notice such conspicuous external characters as the longitudinal striæ on the scuta, and the smooth rounded edges of the radii, it is impossible to identify his species.
Hab.—Mouth of the Indus; East Indian Archipelago; Philippine Archipelago; Moreton Bay, and the north-east coast of Australia. Attached frequently on ships’ bottoms, associated with B. tintinnabulum and amphitrite. Sometimes attached to Gorgoniæ with B. calceolus.
General Appearance.—Shell steeply conical, with the orifice sub-rhomboidal, moderately large, very slightly, or deeply notched: surface very smooth: white, longitudinally banded with pinkish or leaden purple, with sometimes a purplish, sometimes a yellowish tint, the[Pg 280] latter owing to the persistent epidermis; the bands are pale, and often fade away in the lower, and sometimes in other parts of the shell; the epidermis is generally more persistent on the narrow rounded radii than on the parietes, and hence the radii are generally yellowish. The opercular valves are pale dull purple: the sheath is darker purple, with the exception of the portions of the alæ added during the diametric growth, which are of a dead white, and are externally conspicuous. The scuta are striated longitudinally. I may remark, that, excepting the narrowness of the radii, with their quite smooth, rounded and very oblique summits, some specimens are hardly distinguishable, in external aspect, from varieties of B. amphitrite. If the specimens from the north-east coast of Australia, of which I have seen many (but unfortunately only one small one had its opercular valves), form, as I fully believe, merely a variety; it is characterised by its nearly uniform beautiful rosy pink, without any distinct longitudinal bands: of these specimens I have seen one two inches in basal diameter, and three in height: of ordinary duller-coloured striped specimens, the largest was 1.7 in basal diameter. Of the perfectly white var. (b), I have seen several specimens, the largest being .6 of an inch in diameter: these have a somewhat peculiar aspect, but I have met with only one specimen with opercular valves, and that was extremely young: I at first considered this form as specifically distinct; but I can point out, after careful examination of the whole shell, operculum, and internal animal of the young specimen, no sufficient diagnostic characters.
Scutum, plainly striated longitudinally, with the striæ dividing the prominent lines of growth into squarish beads: internally, the upper part of the valve is roughened: the articular ridge is short, remarkably little prominent, and not reflexed; the adductor ridge is blunt and little prominent; sometimes it is almost confluent with the articular ridge: there is a deep but variable depression for the lateral depressor muscle; and in young specimens of var. (a) it was almost absent. Tergum: the surface exhibits traces of longitudinal striæ: there is a deep longitudinal furrow, with the sides folded in and quite closed in full grown specimens: the scutal margin is considerably curved towards the scutum. The spur is long and narrow, with the end bluntly pointed, placed at rather above its own width from the basi-scutal angle; the basal margin slopes but little towards the spur: the crests for the depressores are feebly developed.
Parietes: their internal surfaces are strongly ribbed longitudinally, with the basal ends of the ribs coarsely denticulated, and with the denticuli extending close to the outer lamina. The radii are generally narrow, but their width varies; their summits are very oblique, smooth, rounded, and inflected, with the lines of growth, in the uppermost part, curving inwards; their sutural edges, in the upper inflected portion, are quite smooth, without septa; in the lower and larger portion, the edge is crenated with excessively fine teeth or septa, not denticulated: the radii, like the parietes, have no inner lamina: the recipient grooves in the opposed compartments are smooth, and are in the lower part of the shell of unusual depth. The alæ, differently from the radii, generally have their summits very slightly[Pg 281] oblique, but sometimes they are highly oblique: their sutural edges are most finely crenated. The basis is generally flat, sometimes cup-formed; it is permeated by pores, crossed by transverse septa; and sometimes there is an underlying cancellated layer.
Mouth: labrum with either six very small teeth, or with none. Mandibles (Pl. 26, fig. 5), with the third tooth a little thicker than the first; fourth and fifth teeth small, but quite distinct. Maxillæ (Pl. 26, fig. 7), with the inferior part forming a square step-formed projection, bearing, one behind the other, two spines as long as the upper pair; in a young specimen of var. (a) this step-formed projection was absent.
Cirri: first pair with the rami unequal by about four segments: the shorter ramus has the segments very protuberant in front, thickly clothed with strongly serrated spines; the second cirrus has segments moderately produced; the third has them produced only in a slight degree. The pedicels of the second and third cirri have dorsal tufts of spines, but not a hairy plate prolonged over the thorax. The posterior cirri have segments broader than long, bearing only two pairs of nearly equally long spines; and between each pair there is a small intermediate tuft. The penis has the usual basi-dorsal point.
B. amaryllis is a distinct and well-defined species, more nearly related to B. Hameri than to any other form.
Shell faintly tinged with purple: radii broad, with their summits not oblique: basis not porose. Scutum with the lines of growth crenated: tergum with the spur extremely short, truncated, broad as half the valve.
Hab.—Raine’s Islet, Barrier Reef, Australia, Mus. Stutchbury. Hab. unknown, attached to and coated by Porites. Mus. Brit.
General Appearance.—Shell conical, smooth, but with the lower part sometimes narrowly ribbed in lines corresponding with the internal longitudinal ribs; tinted pale peach-blossom purple, owing to the sheath being finely so coloured; or wholly white. Radii broad, white, square on the summit, hence orifice entire, ovate passing into rhomboidal. The parietal portion of the carino-lateral compartments extremely narrow, about one eighth of the width of the parietes of the lateral compartments. Basis concave, partially imbedded in the coral. Largest specimen .35 of an inch in diameter.
There are some specimens in Mr. Cuming’s collection which appear to belong to this species, and are certainly very closely allied to it, but not having the opercula, cannot be identified positively; the shell is[Pg 282] flatter, with the walls strongly ribbed up to the orifice, which is more rhomboidal: the basis is much more cup-formed and more deeply imbedded in the coral; but these differences by themselves are by no means sufficiently diagnostic.
Scutum: the lines of growth are crenated, causing the surface to be very obscurely striated longitudinally: the articular ridge is very prominent, as can be best seen from the outside, and runs down the whole length of the tergal margin with a very regular curve, and hence differs from the articular ridge in the foregoing species. The adductor ridge is either absent, or very indistinct, and parallel to the articular ridge: there is a deep little pit for the lateral depressor muscle. Tergum (7 d), with the apex somewhat produced or beaked, and tinged purple: external surface almost flat, without any longitudinal furrow: scutal margin curved. Spur very short, placed quite close to the basi-scutal angle of the valve; broad as half the valve; lower end square. Internally, the articular ridge is prominent only in the uppermost part of the valve: crests for the depressores very feeble.
Parietes: their internal surface is very strongly ribbed longitudinally, the ribs being coarsely denticulated at their bases, and finely fluted along their sides. The sheath is transversely ribbed, and clothed with an epidermis furnished with transverse rows of fine hairs. The radii are of a dead white, whereas the parietes are translucent; the summits are parallel to the basis; they are broad; the radii of the carino-lateral compartments appear extraordinarily broad, owing to the narrowness of the parietal portion: the sutural edges are furnished with coarse septa, which are sinuous, irregular, and obtusely denticulated; the interspaces are filled up solidly. The alæ are thin, with their sutural edges almost smooth, and their summits oblique: in some specimens, during the diametric growth, a mere, almost thread-like ribbon is added to their sutural edges. Basis slightly cup or saucer-shaped; moderately thick, permeated by fine pores, and generally ribbed in lines radiating from the centre. The walls and basis adhere together very firmly.
Mouth: labrum with six teeth: mandibles with five teeth; the three upper teeth being sharp, narrow, and unusually prominent; the two lower teeth minute and sharp; maxillæ without a notch. Cirri much injured: first pair with one ramus apparently one third longer than the other: segments not very protuberant: the posterior cirri have elongated segments with five pairs of spines.
Affinities.—This species is very distinct from all the foregoing: in the carino-lateral compartments being so narrow, and tending, as we may suppose, to become aborted; in the form and structure of the whole shell, and in its habits, this species shows an affinity and passage to the coral-inhabiting sub-genus Creusia, which has only four compartments. There is also a close affinity to the sub-genus Acasta. This species is so closely allied to the following, that I at one time felt some doubts whether they ought to have been specifically separated; it is also probably closely allied to B. terebratus, but the materials hardly suffice for judgment: it is also related, though less obviously, to B. vestitus.
Shell dirty reddish-purple, steeply conical: radii narrow: basis obscurely porose. Scutum with the lines of growth crenated: tergum with the spur truncated, broad as half the valve, and depending beneath the basi-scutal angle as much as half its own breadth.
Hab.—Japan, attached to an Isis, Mus. Cuming. Attached to an oyster, Mus. Stutchbury.
As already stated, this species comes in all essential respects very near to the last, though differing much in appearance; I have seen two sets of specimens, and two sets of B. allium, and there was no variability or passage in the points in which they differed; hence I must consider them as specifically distinct.
Shell, steeply conical, strongly but bluntly ribbed longitudinally; coloured either all over dull reddish purple, or with the upper part only pinkish purple: in one set of specimens, the yellow epidermis was partially persistent. Radii narrow. Orifice small, ovate. The wall of the carino-lateral compartment is very narrow. The internal surface of the parietes is ribbed, but finely, and only in the lower part. The septa, on the sutural edges of the radii, are finer than in B. allium. Basis flat, obscurely permeated by pores. The largest specimen is .25 of an inch in basal diameter.
Scuta: these are longitudinally and finely striated; the basi-tergal corner is more rounded off than in B. allium, and the articular ridge is not nearly so prominent: internally, the adductor ridge is rather more prominent. The Tergum is rather broader: its apex is produced into a minute sharp point: the scutal margin is straight; the spur is broader, and measured from the basi-scutal angle of the valve, considerably longer; namely, as long as half the width of the basal margin of the spur, whereas in B. allium it is only about a quarter as long as the basal margin of the spur: the lower edge of the spur is not here so directly transverse to the longitudinal axis of the valve as in B. allium: the external surface is not so flat as in that species, and a depression runs down to the basi-scutal angle of the spur.
Considering the difference in the shape and appearance of the shell, with its narrow radii and small orifice; considering the less strongly ribbed internal lamina of the parietes, the finer septa on the sutural edges of the radii, the slight[Pg 284] difference in outline in the scuta and terga, more especially the greater length of the spur, I conceive I am right in ranking this form as a distinct species, though assuredly it is very closely allied to B. allium, and even still closer to the following B. quadrivittatus.
Shell steeply conical, having four longitudinal gray bands placed crosswise: radii with their summits oblique: basis thin, solid. Scutum, with the lines of growth smooth; no distinct pit for the lateral depressor muscle: tergum as in B. cepa.
Hab.—East Indian Archipelago, attached to lamelliferous corals, and associated with Pyrgoma grande and Creusia spinulosa, Mus. Brit. and Stutchbury and Darwin. Philippine Archipelago, attached to a Tetraclita, Mus. Cuming.
I have seen four sets of specimens of this species, taken in four different places, one set containing above twenty individuals, and all resembled each other exactly: nevertheless, this species comes so close to B. cepa, that I am somewhat doubtful about its specific distinctness.
General Appearance.—Shell smooth, or slightly folded, steeply conical; white, with four longitudinal bands of pale brownish-gray colour, namely, on the rostrum, the carina, and the two lateral compartments: the carino-lateral compartments are very narrow and almost white: the four brownish-gray bands are darkest in the upper part of the shell, though always rather faint, and die out towards the base: they can sometimes be seen to be formed of several narrow longitudinal stripes; the tint shows a trace of containing purple. The orifice of the shell is small, rhomboidal, and not quite entire, owing to the obliquity of the summits of the moderately broad radii. In structure, the shell, radii, and alæ resemble those in the last species. The basis, however, does not appear to be permeated by pores. Basal diameter of largest specimen .25 of an inch.
The Scuta most closely resemble those of B. cepa, but the lines of growth are not crenated, and internally there is only a very minute pit for the lateral depressor muscle, placed almost on the edge itself of the valve. The Tergum hardly differs at all from that of B. cepa, but is perhaps of rather greater breadth.
The Mouth does not differ from that in the last two species. In the Cirri, the three posterior pairs have elongated segments, bearing only[Pg 285] three pairs of spines, of which the lowest pair is minute: in B. allium, and I believe in B. cepa, there are five pairs of spines on each segment.
This species differs from the last only in the peculiar colouring, smoother walls, more oblique radii, solid basis, and more especially in the scuta having the lines of growth not crenated, and internally, in the pit for the lateral depressor muscle being so very minute and placed on the basi-tergal edge of the valve. The posterior cirri, also, I believe, differ in the number of the spines which the segments support; nevertheless, I cannot feel confident about the specific distinctness of B. quadrivittatus.
Shell white, strongly ribbed longitudinally, with the basal margin produced into long points: basis concave, not permeated by pores, but strongly ribbed externally in radiating lines; the interspaces between the ribs being riddled by minute rounded apertures, often placed in double rows.
Hab.—Unknown, Brit. Mus., attached to a lamelliferous coral.
I have in this instance broken through my rule of not describing a Cirripede without examining the opercular valves; but the species here named is so peculiar, that it would have been a fault to have passed it over. There is but a single specimen in the British Museum, without, as just stated, the operculum, and of course without the animal’s body.
Shell, white, depressed, conical, somewhat elongated in its rostro-carinal axis; orifice rather small, pentagonal, toothed, elongated. Parietes rather thin, with extremely prominent longitudinal ribs, produced at the basal edge into long spikes: the internal surface is also ribbed, but less strongly than the outside. Radii rather narrow, with oblique, not smooth summits; sutural edges very finely and obscurely crenated. Alæ with their summits extremely oblique. Lower edge of sheath closely attached to the walls. The carino-lateral compartments are rather narrow.
Basis, slightly concave or saucer-shaped; the circumference is pro[Pg 286]duced into long spikes, corresponding with those on the basal margin of the parietes: these projections equal half the semi-diameter of the shell. The internal surface of the basis has slightly prominent, rounded ridges; and the external surface has extraordinarily prominent, sharp ridges, radiating from the centre; the edges of the external ridges are irregular, notched, and knobbed. I have seen in no other species external ridges on the basis or surface of attachment; and what is more remarkable, the interspaces between the ridges are penetrated by small rounded apertures, of irregular shape and unequal sizes; and these are generally arranged in an irregular double row, and externally are closed by the membrane, which clothes the basis. In the sub-genus Acasta, the basal cup is sometimes penetrated by similar holes, but these seem never to extend over the whole basis, and are very variable; nevertheless, in some specimens of Acasta spongites from the Cape of Good Hope, portions of the basis closely resembled, except in the absence of the radiating ridges, the structure here described, but the holes were not arranged in any definite order. The internal surface of the parietes in Acasta sporillus presents a somewhat analogous appearance, but the pits do not penetrate through the walls. This species, I have no doubt, is closely allied to the sub-genus Acasta, and to Balanus navicula with its allies, and, but much less closely, to B. allium with its allies. Indeed, had B. terebratus inhabited a sponge, I should have been compelled to have ranked it in the sub-genus Acasta.
Shell pinkish-purple or white, clothed by an orange-coloured membrane; radii represented by mere fissures; basis solid. Scutum, with a sharp, curved adductor ridge; with crests for the lateral depressor muscle: tergum, with the spur short, truncated, one third of width of valve.
Hab.—New Zealand, New South Wales, Mus. Brit. and Stutchbury; attached to shells.
General Appearance.—Shape conical, often steeply conical; orifice small; radii not developed, represented by mere fissures. The walls are smooth, or slightly, or strongly ribbed longitudinally. The shell itself is of a fine peach-blossom pink, or nearly white, but it is generally covered by a thick yellow or brownish-orange epidermis. Opercular valves pinkish, but similarly covered. Basal diameter of largest specimen .7 of an inch.
Scuta, with the lines of growth closely approximate; surface somewhat convex. Internally, the articular ridge is very little prominent, but runs far down the tergal margin; in some specimens, however, it is shorter and more prominent. The adductor ridge is strongly prominent, is curved towards the rostral angle, and runs down nearly to[Pg 287] the basal margin. The rostral depressor muscle is lodged in a small cavity, formed, as usual, by the overlapping of the occludent margin; within this cavity there are either tolerably distinct little crests, or merely traces of them, for the attachment of the muscle. The lateral depressor muscle is attached to several quite distinct crests, seated in a concavity beneath the adductor ridge. Tergum rather narrow, with the apex produced or beaked; the beak is purplish and flat. There is a slight rounded longitudinal furrow, or depression. The spur is fully one third of the width of the valve: it is short, with the end truncated, and placed close to the basi-scutal angle; the basal margin on the carinal side slopes gently towards the spur. Internally, the scutal margin is scarcely at all inflected, and the articular ridge is very little prominent: the crests for the tergal depressores are pretty well developed.
From the points here enumerated, it is clear that the opercular valves are articulated together much less strongly than is usual with most species, excepting B. allium and its allies. It is remarkable that in this species the terga are united to the sheath, not, as is usual, by a single opercular membrane, but by five or six, one above the other, the upper membranes not having been exuviated as each new lower one was formed. The minute spines on the membrane lining the sheath are rather larger and more numerous than is usual, and to the base of each spine a tubulus of unusual diameter runs, imbedded in the shell.
The Walls, internally, have unusually numerous, narrow, approximate, strongly prominent, longitudinal ribs, denticulated at their bases, and inserted into the furrows on the borders of the basis: in old specimens these internal ribs die out in the upper part of the walls. The Radii are not developed in any of the many specimens which I have seen, and the edges of the compartments on both sides of each suture are equally marked by slight, irregular ridges or knobs, answering to the septa and their recipient furrows, in the species with ordinarily developed radii. There is very little diametric growth, the orifice being gradually enlarged by the disintegration of the upper ends of the walls; the alæ, however, in some specimens, do grow a little along their lateral or sutural edges, so that some little diametric growth must be effected. The summits of the alæ are very oblique; their sutural edges are plainly crenated. The sheath descends about half way down the walls. The Basis is flat, not permeated by pores, but deeply furrowed in lines radiating from the centre.
Mouth: labrum sometimes with no teeth, sometimes with four minute teeth; mandibles with four teeth, of which the third is blunt and rather large; the fourth is a mere knob. Maxillæ; there is, as usual, an upper pair of large spines (beneath which there is sometimes a small notch), but all the lower spines, instead of standing as usual in pairs, form a single row. Cirri; first pair with the rami remarkably unequal in length, one ramus having twenty-two segments, and being more than twice the length of the other, having only nine segments: these segments, and likewise those of the second and third pairs, have an inverted conical shape; and they are all less thickly clothed with spines than is usual. The second pair is short, about as long as the[Pg 288] shorter ramus of the first pair, and has ten or twelve segments. The third pair is above twice as long as the second pair, and contains twenty-four segments: this very unusual length is owing to the presence of numerous thin tapering upper segments, unlike those generally found in Balanus, in the third pair of cirri, and apparently serving as feelers. These upper tapering segments are of an inverted conical shape, and support on their upper margins two very small tufts of spines, one behind and one in front: on the segments lower down these tufts increase in size, and the spines are more spread out, so that in the basal segments, the tufts in front form on the upper margin two or three crowded rows of bristles. The three posterior pairs of cirri have elongated segments, which bear on their upper half three pairs of spines; of these the lowest pair is minute, and the middle pair is only one third of the length of the upper pair. The sixth cirrus, in the same individual as before, contained twenty-seven segments in each ramus, that is only three more than in the third cirrus! I must observe, that the cirri in all the specimens were irregular, often distorted and monstrous; and therefore, probably, there is considerable variation in the proportional numbers of the segments in the cirri.
At the base of the penis there is a minute, knife-edged, triangular projection. The branchiæ are rather narrow, pointed, and not very large.
Affinities.—This is a very distinct species, as shown by the peculiarities in the cirri, by the absence of radii to the shell, and by the presence of crests for the attachment of the lateral scutal depressores. With the exception of this latter character, the opercular valves clearly show, that B. vestitus is allied to B. allium, cepa, and quadrivittatus. In some respects this species manifests an affinity to B. imperator, which latter has its third pair of cirri nearly similar to those of B. vestitus.
Shell internally imperial purple; parietes thick, with their internal basal edges rough with irregular points and ridges; radii narrow; basis very thin, solid. Scutum, with crests for the rostral and lateral depressor muscles; tergum, with the end of spur rounded.
Hab.—New South Wales, Sydney, Port Stephens, Moreton Bay; attached to sandstone-rocks and shells, at low-water line; Mus. Brit., College of Surgeons, Cuming, Stutchbury.
Shell conical, very thick and very strong; longitudinally sulcated more or less strongly; whole thickness of shell beautifully coloured rich violet, or more strictly “imperial purple;” externally the surface, from disintegration, is generally whitish; internally the colour is best developed: the narrow radii and the thin basis are white. The largest[Pg 289] specimen which I have seen was one and three-quarters of an inch in basal diameter, the walls close to the basis being, in this instance, actually .3 of an inch in thickness.
Operculum thick and strong, covered by yellowish-brown epidermis; internally, the shelly substance is either all of the richest purple or yellowish-white, tinged, especially in the upper part, with purple. Scuta, with the apex beaked and somewhat reflexed; articular ridge very thick, little prominent; articular furrow very narrow, the impression made by the adductor muscle is seated very high up the valve; there is hardly an adductor ridge, but the surface of the valve is angularly prominent in a curved line, running from the articular ridge to near the rostral angle of the valve. At the rostral angle, the occludent margin is not folded inwards, as is generally the case, but the surface is flat, and is marked by four or five crests for the attachment of the rostral depressor muscle. There are other crests for the lateral depressor muscle. Tergum, with the apex somewhat produced and beaked, but blunt; longitudinal furrow shallow; spur of moderate breadth, with its lower end rounded; the basal margin on the carinal side of the spur slopes towards it. Internally, articular ridge moderately prominent. Crests for the tergal depressor well developed.
Parietes, solid, thick, with the basal internal edge (4 c) formed of short ridges, or flattened and irregular points, which in very old specimens are but little prominent; inner surface, finely, closely, and irregularly ribbed longitudinally, but in some specimens nearly smooth. The radii are nearly white; they are narrow, sometimes hardly at all developed, and have their summits very oblique and jagged; exteriorly, they are sulcated in a transverse direction, and sometimes form oblique steps, from having been formed layer over layer: their sutural edges are formed of irregularly branching crests or septa. The alæ have their summits very oblique; their sutural edges are thick and crenated: the part added during diametric growth on the inner surface is smooth, and has a different appearance from the transversely ribbed portions of the sheath, of which the alæ form a portion. The lower edge of the sheath is hollow beneath. The carino-lateral compartments are very narrow.
Basis calcareous, thin, white, sometimes opalescent, apparently formed by an aggregation of very minute calcareous beads, with no trace of furrows radiating from the centre.
Mouth: labrum hairy, with apparently some very minute teeth; mandibles, with the fourth and fifth teeth small and rudimentary; maxillæ rather broad, with a narrow and rather deep notch under the two great upper spines: outer maxillæ with the lower lobe very large.
Cirri: first pair, with the rami unequal by several segments: second pair, with the rami unequal in length by about six segments: third pair elongated, with the segments very numerous, almost equalling those in the sixth cirrus; upper segments of both rami much elongated, each with only a circle of spines; segments in the above first three pairs of cirri only slightly protuberant. Posterior cirri elongated, with the upper segments bearing three pairs, and the lower segments four pairs of main spines, between which there is a small intermediate tuft.
Affinities.—This noble Balanus, in all the characters derived from its opercular valves, and from its cirri, is closely allied to the last species: in the structure, however, of the shell and of the basis, it comes closer to the following, B. flosculus. The crests on the under side of the scutum, for the lateral depressores, are confined to these three species; and the crests for the rostral depressores are confined to B. imperator and vestitus, but they are generally rudimentary in the latter. The internal basal structure of the parietes is singularly like that of Chelonobia caretta, though there is no other special affinity to that genus. In the nature of basis; in the structure, to a certain limited extent, of the walls of the shell; in the narrowness of the carino-lateral compartments; in the elongation of the third pair of cirri; in the crests for the rostral and lateral scutal depressores, B. imperator comes nearer to the genus Tetraclita than does any other species of Balanus.
Shell purple or dirty white, with the internal basal edges of the parietes rough with irregular points and ridges; radii narrow or absent; basis excessively thin, in appearance absent. Scutum with crests for the lateral depressor muscle; tergum very narrow, with the spur pointed.
Var. sordidus. (Pl. 8, fig. 5 b) shell globulo-conical, dirty white, with numerous sharp, narrow, longitudinal folds or ridges.
Hab.—Peru and Chile; generally attached to the Concholepas Peruviana, or to Balanus psittacus, and associated with Chthamalus scabrosus. Var. sordidus, inhabits Tierra del Fuego, attached to littoral shells, wood, and rock, associated with Ch. scabrosus.
General Appearance.—Shell either extremely much depressed and irregular, or globulo-conical, or more rarely cylindrical and elongated. Walls, either with a few rather broad, smooth, irregular, longitudinal folds, or, in var. sordidus, with numerous sharper and more prominent longitudinal ridges; basal margin very sinuous. Colour, either fine rich peach-blossom purple, or so pale as to be almost white; or in var. sordidus dirty white, generally stained greenish from confervoid matter. Orifice small, oval, entire. Radii very narrow, white, or not at all developed, and with even the sutures not distinguishable. The purple coloured varieties, with the narrow white radii, the small oval orifice, and folded walls, have a very pretty appearance, which is far from the case with var. sordidus. The largest specimens attained a basal diameter of .6 of an inch, but this is an unusual size: I have seen a cylindrical specimen of var. sordidus one inch in length.
The opercular valves are united to the sheath by unusually strong membrane: internally, their upper parts are stained purple. Scuta, these vary considerably in breadth, some being even broader than in[Pg 291] Pl. 8, fig. 5 c, and others as much elongated as in fig. 5 d: these latter come from an elongated cylindrical shell. The valve externally is unusually convex: the apex, also, projects freely to an unusual degree. Internally, the articular ridge is moderately prominent: the adductor ridge is prominent and much curved: in the upper part it either lies close to, or at some little distance from the articular ridge. The lateral depressores are attached to several little crests, occupying a cavity, (often bordered above by a little ridge) close beneath the adductor ridge. Tergum, extraordinarily narrow and elongated; beak triangular, purple: a longitudinal furrow runs down the valve: spur narrow, long, bluntly pointed, lying close to the basi-scutal angle of the valve: the scutal margin is nearly straight and parallel to the spur. Internally, the articular ridge is prominent: the crests for the depressores are moderately well developed: the upper part of the valve is marked by a purple patch, bounded on one side by the articular ridge, and on the other side by a very slight special ridge. There is some variability in the narrowness of the whole valve, and in the sharpness of the spur.
Parietes.—The under surface, in the more depressed varieties, is roughened in a remarkable manner nearly or quite up to the sheath, with very irregular, projecting, and branching ridges, and sometimes with depending points. These ridges and points are granulated on their under surfaces. The roughened surface in the more conical varieties is confined to the basal inner margin. This structure is nearly the same as that in B. imperator, represented (Pl. 8, fig. 4 c), but the little ridges are here more apt to be concentric, instead of radiating. The lateral edges of the compartments on the inside, especially the carinal edges, project inwards beyond the inner surface of the shell. The sheath is but little hollow on its under side. The diametric growth of the shell seems to be quite capricious; in the same group, some individuals thus increasing, and others not at all. When the radii are developed they are narrow and white, with their upper margins nearly if not quite parallel to the basis: their sutural edges are formed by large, irregular, branching teeth or septa. The alæ, also, have their sutural edges coarsely crenated; and when the shell increases by diametric growth, they are added to above the opercular membrane, so as to be nearly square at top.
Basis.—When a shell is removed from the surface of attachment, and inspected even under a lens, there appears to be no basis whatever, either adhering to the shell, or to the supporting surface: but when a more careful examination is made, with a higher power, an excessively thin, translucent, calcareous, irregular layer, or rather film, can almost always be discovered. This would be more easily distinguished if the specimens had adhered to rock instead of to the rugged shells of molluscs. At one time I thought the basis was partially membranous, for I have certainly scraped off small fragments of membrane from the supporting surface; but these, when examined under the compound microscope, seemed always to consist of a thin sheet of the yellow cementing tissue; and in some instances, a brittle film of shell, representing the true basis of the cirripede, still adhered to the upper surface of the membranous bits of cement. Nevertheless, so imperfect is the[Pg 292] calcareous basis, that I should not be surprised if portions of a true membranous basis did really in some cases exist.
Mouth: labrum with the notch wide, generally with a few little teeth; mandibles with three teeth, and some inferior knobs; maxillæ notched. Cirri, first pair with one ramus shorter by three segments than the other ramus. Second and third pairs short, of nearly equal length, thickly clothed with spines; segments very little protuberant. Posterior cirri, having elongated segments, supporting seven pairs of spines.
Var. sordidus.—This form is very common on the tidal shores of the Strait of Magellan, and of the southernmost parts of Tierra del Fuego, near Cape Horn: it lives attached to rocks, mytili, and logs of wood, and is associated with Chthamalus scabrosus. It almost certainly is the most antarctic form of the genus Balanus. If I were guided by external appearance alone, I should certainly separate this form specifically from B. flosculus, but, as will be seen in the following description, the differences consist only in var. sordidus being much duller and rather differently coloured, in the longitudinal folds being sharper and more prominent, and in the whole shell being rather more globular, and on an average rather larger; but in the true B. flosculus there is considerable variation in all these respects, as there likewise is in var. sordidus; thus some of the cylindrical varieties of the latter have less prominent ridges than even B. flosculus. In general appearance I have seen some nearly, but not exactly, intermediate forms; therefore, I do not feel positive that these forms may not be specifically distinct, but have failed, after careful examination, to find any sufficient diagnostic characters. Moreover, in the case of Balanus lævis, I was led to believe that there is an equal and somewhat analogous amount of variation in the specimens inhabiting Tierra del Fuego and northern Chile; and in this case I was enabled to show the existence of strictly intermediate forms in the intermediate districts.
The shell in var. sordidus is generally globulo-conical, dirty white, frequently with a green tinge, from the growth of confervoid matter. Orifice small. The exterior surface is covered with numerous prominent, narrow, sharp ribs or folds, the basal margin being serrated with projecting points where the folds terminate. When the radii are not developed, the sutures are very often obscure. Internally, the shell is faintly tinted of a port-wine purple. In all points of structure this form is identical with the true B. flosculus. In some few specimens the whole exterior surface was disintegrated and smooth; and this is generally the case with the upper parts of the shell. Some other specimens, which had grown crowded together on wood, had become cylindrical, and consequently the orifice was as large in diameter as the shell, namely, half an inch: in some of these cylindrical varieties the sheath was entirely soldered to the walls. The largest specimens which I have seen were .6 of an inch in diameter; and above one inch in height.
Affinities.—This species, in its opercular valves, even in such trifling characters as the strength of the opercular membrane, and in its cirri, approaches closely to B. cariosus. We even see on the under side of[Pg 293] the scutum, in that species, a single little ridge, foreshadowing, as it were, the crests for the lateral scutal depressores, so remarkable in our present species. In the structure of the shell and of the basis, B. flosculus is much more closely related to the last species, or B. imperator. If it had been possible to have arranged the species in a single line, B. flosculus ought undoubtedly to have been placed between B. cariosus and imperator.
BALANUS SULCATINUS (?) Nyst, apud D’Omalius (sine descript. aut tabulâ), Geologie de Belgique, 1853.[100]
[100] I am indebted to M. Bosquet for a specimen, bearing this name and reference, found in the ‘Systeme Bolderien’ of Dumont, (miocene according to Sir C. Lyell) at Bolderberg. The specimen consists of a rostrum, with a portion of the base attached; and as these parts are in some degree characteristic, I fully believe this specimen to be the B. bisulcatus. I hope hereafter to give in the Palæontographical Series fuller illustrations of this and the following fossil species.
Radii with their upper margins oblique and smooth; sutural edges smooth: basis permeated by large pores. Scutum narrow, with from two to four longitudinal furrows: tergum with the spur very short, broad as half the valve.
Var. plicatus, with the walls deeply folded; radii narrow, with their upper margins very oblique.
Fossil in Coralline Crag, Ramsholt, Gedgrave, Sutton; Mus. S. Wood, Bowerbank, J. de C. Sowerby. Rauville, dans le Cotentin, Mus. G. B. Sowerby. Var. plicatus, Coralline Crag, Sutton, Mus. S. Wood. Bolderberg, near Hasselt, Belgium, Mus. Bosquet.
General Appearance.—Shell conical or tubulo-conical, often rather globose; walls frequently thin, either very smooth, or deeply plicated longitudinally: occasionally the same specimen is smooth in the upper part, and strongly plicated in the lower. The Radii in the larger specimens are wide, and with their upper margins only slightly oblique; in the smaller, they are narrower and much more oblique, but in each case their upper margins are smooth and slightly bowed. Colour apparently originally nearly white, but with the alæ generally, in the smaller specimens, clouded with a dark tint: the radii are usually striped feebly in longitudinal lines. Basal diameter of largest specimen .8 of an inch; but this seems to have been an unusual size.
Scuta: narrow, with the basal margin forming an unusually small angle with the occludent margin; surface slightly convex, with lines[Pg 294] of growth approximate, moderately prominent; on the tergal half of the valve, two distinct rather broad furrows, with sometimes a third, and even a fourth, nearer to the occludent margin (Pl. 8, fig. 6 a), extend from the apex down the valve, and give it a very peculiar appearance: the furrows near the tergal margin are the deepest. Internally, the upper part of the valve is roughened with small points: the articular furrow is unusually wide: the articular ridge is very prominent and but little reflexed, with the lower end almost abruptly cut off: the adductor ridge is prominent, but short: there are small deepish pits for the rostral and lateral depressores.
Terga (6 b), broad, flat, with a slight narrow prominent rim along the scutal margin, which margin is slightly bowed. The basal margin on the carinal side of the spur slopes so gradually towards the spur, that the latter is barely distinct, and is very short, not depending nearly half its own width beneath the basi-scutal angle: it is broad, namely, measured across the upper part, as broad as half the valve; its basal end is obliquely rounded off on the carinal side; it is placed close to the basi-scutal angle. The carinal margin of the valve is just perceptibly bowed, and is formed by rectangularly upturned lines of growth. Internally, the upper part of the valve is rough; the articular ridge is prominent; the crests for the tergal depressor muscles are moderately well-developed.
Parietes, not porose; internally, the ribs are smooth, with their basal edges very finely or barely denticulated. The radii (as already stated) are of variable breadth; they have their upper margins either very slightly or highly oblique, but always smooth and rounded: their sutural edges are quite smooth, or sometimes, with a strong lens, traces of transverse striæ, representing septa, can just be discovered. The alæ have their upper margins very oblique; their sutural edges are, in the large specimens, quite smooth; in the younger ones, plainly crenated; the recipient furrow being clearly marked with these teeth. Basis plainly porose.
Varieties.—It is certain that there are longitudinally plicated specimens of this species, and that the obliquity of the upper margins of the radii also varies a little: nevertheless some of the deeply plicated specimens undoubtedly have a very different aspect from the ordinary varieties, and do really differ in the sutural edges of the alæ being crenated, and in the greater narrowness and obliquity of the radii; but these points are all commonly variable. I have not seen any large specimens of the variety, plicatus, so as to compare them with the large specimens of the normal form, yet I can hardly entertain any doubt, considering their agreement in so many important points, that I have rightly treated these forms as mere varieties; it is unfortunate that none of the specimens of the var. plicatus seen by me have had opercular valves, as their presence would have removed all shadow of doubt.
Affinities: this is a strongly characterised species, and nearly allied only to the following species, B. dolosus. The furrows on the scuta in some degree resemble those on B. lævis, but there is no alliance with that species. It is certain that amongst recent species, the chief affinity is with B. Hameri and amaryllis.
Radii with their upper margins oblique and smooth; sutural edges smooth: basis permeated by large pores. Tergum with the spur not very short, broad as one third of valve.
Fossil in Red and Mammaliferous Crag, England; Mus. S. Wood, Bowerbank, Lyell, J. de C. Sowerby, Henslow, &c. Mammaliferous Crag, Postwick, near Norwich, Mus. Lyell.
This species so closely resembles B. bisulcatus, both externally and in all the essential characters of the parietes, radii, and basis, that it is quite superfluous to describe over again these parts. The specific characters are derived from the opercular valves, which present well defined distinctions, found by me constant in several specimens of both species. B. dolosus, like B. bisulcatus, has quite smooth and deeply plicated varieties, often adhering to the same univalve. The ribs on the inner surfaces of the parietes are remarkably prominent. I think the upper margins of the radii are in this species rather more oblique than in B. bisulcatus. The sutural edges of the radii are marked by the finest striæ, representing septa. The sutural edges of the alæ are generally distinctly crenated. The basis is often slightly cup-formed, and very plainly porose: its surface is marked by radiating ridges. The orifice of the shell is large, and elongated, especially in young specimens. The basal diameter of the largest specimen is .4 of an inch.
The Scuta have no trace of the two or three longitudinal furrows so conspicuous on these valves in B. bisulcatus, and which, in that species, run down from the apex of the valve; this fact showing that the furrows occur in quite young individuals. The whole valve is not quite so narrow as in B. bisulcatus, but otherwise agrees with it in shape: internally, there is hardly any difference: the articular furrow is not so wide: the articular ridge is very prominent, and abruptly truncated at its lower end: the adductor ridge is also prominent; it here runs a little higher up the valve than in B. bisulcatus. The Tergum differs more in the two species: the spur is not so broad; measured in its upper part, it is only about one third of the entire width of the valve, instead of being[Pg 296] half as wide as the valve: it is considerably longer, depending beneath the basi-scutal angle more than half its own width: the basal margin of the valve on the carinal side, does not slope so gradually into the spur: the occludent and carinal margins are slightly arched, as in B. bisulcatus. Internally, the surface is rough, the articular ridge is prominent, and the crests for the tergal depressores are well developed,—all as in B. bisulcatus. It is remarkable, how generally the opercular valves have been preserved in this species in its fossil condition, as compared with most other species of the genus.
It is not easy to distinguish by external characters the rugged varieties of this species from B. crenatus; indeed, the only difference is that the furrows receiving the edges of the radii, generally, exhibit in B. crenatus a slight impression of the septa, which are entirely absent in B. dolosus. By internal characters, such as the non-porose parietes, and porose basis, our present species widely differs from B. crenatus.
BALANUS UNGUIFORMIS. J. de C. Sowerby (!). Mineral Conchology (sine descriptione) Tab. 648, fig. 1, (Jan. 1846).
------ ERISMA. J. de C. Sowerby (!). Ib., fig. 2.
------ PERPLEXUS. Nyst, apud D’Omalius (Sine descript. vel Tab.), Geologie de la Belgique, 1853.[101]
[101] I am much indebted to M. Bosquet for specimens bearing this title, from Klein Spauwen, which certainly appear to me, as far as can be judged by the separated compartments, without the opercular valves, to belong to our present species.
Parietes thin, sometimes permeated by pores; radii with their upper margins oblique; sutural edges very finely crenated: basis solid. Tergum with the spur narrow, bluntly pointed.
Var. erisma, with the walls longitudinally folded or ribbed.
Fossil in the Eocene formation, Isle of Wight, Colwell Bay; Hordwell; Barton, (Chama Bed); Headon; Bembridge. Bergh, near Klein Spauwen, Belgium (?). Attached to various shells and wood. Mus. J. de C. Sowerby, E. Forbes, F. Edwards, Charlsworth, T. Wright, Bowerbank, Tennant, Bosquet.
This species, the most ancient one as yet well known in the genus, presents to the systematist a most unfortunate peculiarity, in the parietes being almost as often as not permeated by small pores: I have seen no other instance, except to a limited degree in B. glandula, of this character being variable, and hence it must be still considered of high classificatory value, in so varying genus as Balanus. Owing to this varying condition of the parietes, together with the basis being quite solid, our present species has as good a claim to be ranked in the last as in the present section; indeed, I think it has more affinity to B. crenatus and glandula in the last section, than to any other recent forms: I have placed it in its present place, owing to its intimate affinity to B. varians, in which the parietes seem always to be solid; and partly, I believe, because all the first specimens examined by me exhibited no traces of parietal pores. Owing to the kindness of Mr. F. Edwards, I have seen the original specimens, excellently figured by Mr. J. de C. Sowerby in the ‘Mineral Conchology:’ I can perceive no difference between B. unguiformis and erisma, excepting that the walls in the latter are longitudinally folded,—a character we know to be variable in so many species. In both varieties, the parietes are sometimes porose and sometimes solid. The smaller specimens, however, figured in the ‘Mineral Conchology’ to the right hand, may possibly be a distinct species, as I infer from the narrowness of their radii.
General Appearance.—Shell, tubulo-conical, sometimes considerably elongated and sub-cylindrical: surface either very smooth, or slightly folded, or deeply folded so as to be strongly ribbed longitudinally: orifice rather large, rhomboidal, narrow at the carinal end, toothed, but not deeply: walls rather thin and fragile: radii of moderate width, with their summits oblique, not quite smooth. Basal diameter of largest specimen, about three quarters of an inch.
Scuta, with the external surface smooth: there is a trace of a furrow running down the valve from the apex, near to the occludent margin, and this is only worth mentioning from the analogous furrows in B. bisulcatus. Internally, the upper surface of the valve is roughened: the articular ridge is very prominent, and slightly reflexed: there is no distinct adductor ridge; there is a slight but variable depression for the lateral depressor. Tergum, with the longitudinal furrow shallow; spur moderately long, about one fourth or one fifth of the width of the[Pg 298] valve; placed at about its own width from the basi-scutal angle; basal end bluntly pointed; the basal margin on the opposite sides of the spur forms a nearly straight line; the carinal margin has an extremely narrow border formed by upturned lines of growth. Internally, the surface is roughened with little points: the articular ridge is prominent: the crests for the tergal depressores moderately prominent.
Parietes: the longitudinal ribs on the internal surface are either feebly, or, in the lower part, strongly developed; their basal ends are only just perceptibly denticulated. As already stated, in about half the specimens, there were no traces of parietal pores; in the other half there were either distinct or obscure pores; the pores are circular, generally of unequal sizes, and never large; in the same individual they would sometimes be almost wholly absent in some of the compartments, and quite plain in the other compartments. The Radii are either moderately wide or rather narrow, and have their upper margins very oblique, and not distinctly arched, and not quite smooth: their sutural edges are very finely crenated, the teeth or septa not being denticulated. The upper margins of the alæ are rather less oblique than those of the radii: their sutural edges are barely crenated. The basis is thin, and without any trace of pores; the upper surface is sometimes furrowed in radiating lines.
B. VARIANS. G. B. Sowerby, in Darwin’s Geolog. Observ. on South America, (Sept. 1846), Tab. 2, fig. 4, 5, 6.
Parietes moderately thick: radii with their upper margins very oblique; sutural edges almost smooth, or finely crenated: basis finely porose. Tergum with the spur small, narrow, bluntly pointed.
Hab.—Port St. Julians, Patagonia; ancient Tertiary formation. Eastern plain of Tierra del Fuego (?).
This species comes so close to B. unguiformis, that I have some doubt whether they ought to be specifically separated: the whole shell is stronger, and the basis can be seen to be porose when a polished section is made: the spur of the tergum is smaller, more pointed and more medial, but these latter differences may be due to mere variation. Should B. varians and unguiformis prove to be the same species, the latter name has the priority.
General Appearance.—Shell moderately strong and thick; shape conical or tubular, or even inverted conical; orifice moderately[Pg 299] toothed, large, sub-trigonal; walls either smooth or longitudinally folded; the elongated specimens are most apt to be smooth. The Radii are narrow and oblique. Diameter of largest specimen above three-quarters of an inch.
Scuta, with the lines of growth moderately prominent; the internal surface of the valve has been ill preserved; but a very prominent, hardly reflexed, articular ridge, can be distinguished, as well as the absence of an adductor ridge. Terga, with no distinct longitudinal furrow running down the valve: spur short, bluntly pointed, narrow, about one fifth or one sixth of width of valve; placed at above its own with from the basi-scutal angle; the basal margin, on each side close to the spur, curves towards it. Internally, all that can be distinguished, is that the articular ridge is prominent.
Parietes; their inner surfaces appear to have been nearly smooth; the absence of parietal pores could be made out only by polishing a transverse section. The Radii are narrow, and have their upper margins very oblique and rather smooth: in the elongated varieties the sutural edge appears to be almost absolutely smooth; in the conical specimens it is slightly crenated, the septa being apparently not denticulated. In living species we have a similar variation in the state of the sutural edges of the radii, in B. balanoides and crenatus the edges being much smoother in much elongated specimens than in other varieties. The alæ have their upper margins less oblique than those of the radii, with their sutural edges barely crenated. The basis is either flat, or, in the elongated specimens, deeply cup-formed; in section it can be seen to be finely and irregularly porose.
Shell reddish-brown: radii broad, with their upper margins not oblique, or only moderately oblique; sutural edges with plainly denticulated septa: basis porose. Scutum without an adductor ridge; tergum with the spur rather narrow.
Var. (a) (Pl. 8, fig. 10 b, 10 c), with the shell elongated in its rostro-carinal axis; basis narrow, clasping the stem of a zoophyte; lateral compartments much broader than the almost linear rostrum, carina, and carino-lateral compartments.
Var. (b), with rough longitudinally folded walls, and with the summits of the radii forming an angle of about 45° with the basis.
Fossil in Coralline Crag, Sutton and Gedgrave; attached to foliaceous Bryozoa; Mus. S. Wood, Bowerbank. Var. a, Coralline Crag, Sutton, attached to cylindrical branches of corals; Mus. S. Wood. Var. b, attached to shells, Osnabruck, Hanover, Mus. Lyell; Bünde, Westphalia, Mus. Krantz.
My materials consist of a beautiful series of specimens in Messrs. Wood and Bowerbank’s collections; but unfortunately only a single young specimen had its opercular valves preserved. Not one specimen of the very curious variety (a) had opercular valves, yet I cannot feel any doubt about its being only a variety, caused by its attachment to a thin cylindrical branch of a coral, instead of to a foliaceous Bryozoon; it will, however, be convenient to give a separate description of this very remarkable form. With respect to var. (b), both sets of specimens came to me with the name B. stellaris, of Bronn; but as Bronn distinctly states, that in his species the parietes are porose, and as such is not here the case, this cannot possibly be that species: these specimens did not possess their opercular valves, and therefore cannot be identified with certainty.
General Appearance.—Shell conical, with the orifice rather large, and rhomboidal. The surface is very smooth, except in var. (b.) from the Continent, in which it is rugged and longitudinally folded. The colour is ochreous-brown (chiefly no doubt derived from the imbedding substance) tinged with red. The radii often have a much darker and more distinct red tint; they are sometimes longitudinally striped with dirty white. The radii are broad, with their summits straight, and very slightly oblique; in var. b, however, they slope at an angle of about 45°. Basal diameter of largest specimens .6 of an inch; but this is an unusual size.
Scuta (from a young individual), with the growth ridges little prominent. Internally the articular ridge is moderately prominent, with its lower end very obliquely rounded off; there is no adductor ridge; there is a minute pit for the lateral depressor muscle. Terga, with a slight longitudinal depression extending down to the spur: spur short, with its lower end almost square or truncated, about one fourth of width of valve, and placed at about half its own width from the basi-scutal angle. Internally, the articular ridge is prominent; the crests for the tergal depressores are feebly developed.
Parietes, moderately thick and generally strongly ribbed internally, without parietal pores. Radii, wide, with their upper margins straight, not smooth or rounded, and very slightly (or, in var. b moderately) oblique; their sutural edges have well-developed septa, which are denticulated: the interspaces between the septa are filled up solidly. The alæ have their upper margins oblique: they are only slightly, and sometimes not at all, added to above the level of the opercular membrane: their sutural edges are smooth. The basis is thin, but plainly porose.
Var. (a).—With respect to this remarkable variety, any one would at first think it specifically distinct. The shell is much compressed, or elon[Pg 301]gated in the rostro-carinal axis, sometimes to a great degree; I have seen a specimen .25 of an inch in this axis and only .1 in its broadest part; but this is a very unusual degree of elongation. The most remarkable character is the extraordinary narrowness of the carina, the carino-lateral compartments, and of the rostrum, compared with the great breadth, especially along the basal margin (Pl. 8, fig. 10 b, 10 c), of the lateral compartments. The radii are of unusual breadth. The tips of the rostrum and of the lateral compartments are a little arched in, tending to make the shell somewhat globular. The true basis is extremely narrow (fig. 10 c): it is deeply grooved, from clasping the thin, cylindrical stem of the coral to which it had adhered; and I have seen specimens in which the opposite edges of the groove had met, a tube having been thus actually formed. From the grooved basis, and from the elongation of the shell in the rostro-carinal axis, this species presents so close a general resemblance to Balanus calceolus, and its allies, that I have seen it in a collection arranged on the same tablet with a fossil specimen of B. calceolus. Notwithstanding the above several strongly-marked characters, by which this variety differs from the ordinary form, there is a resemblance in colour and aspect, which though difficult to be described, made me from the first suspect that the two were specifically identical. In no point of real structure is there any difference, excepting that, perhaps, the pores in the basis are here rather smaller; but this might arise from the little development of the peculiar basis. Having come to this conclusion, I was interested by finding a specimen in Mr. Wood’s collection, which had originally fixed itself (judging from the form of the basis) on a cylindrical stem, but which had subsequently grown on to an adjoining flat surface; consequently, one side of the shell presented all the peculiar characters of the present variety, whereas the other side, at the rostral end, was undistinguishable from the ordinary form. The unequal development of the rostrum on the two sides was very striking, and clearly showed how great an effect could be produced by the nature of the surface of attachment.
This singular variety cannot be considered accidental, in the sense in which this term may be applied to some varieties: the larva evidently fixes itself intentionally, in a certain definite position, on the branch of the coral (when a branch is chosen), exactly as in the case of Balanus calceolus, or Scalpellum vulgare. But when other species of Balani occasionally fix themselves on branched corals, their position seems to be accidental and unsymmetrical; thus among the symmetrically elongated specimens of the present species, I found one specimen of Balanus bisulcatus, which had evidently been attached in an almost transverse position to a branch, and had thus become much distorted; so, again, I have seen specimens of the recent B. amaryllis attached irregularly to a Gorgonia, in the midst of the symmetrically elongated shells of Balanus navicula, an ally of B. calceolus.
This variety does not seem to attain so large a size as the ordinary form.
Affinities.—This species is allied to the two last described fossils, namely, B. varians and unguiformis, but is perhaps more nearly related[Pg 302] to the recent B. allium, an inhabitant of the Barrier Reef of Australia. The longitudinally folded variety (b) can hardly be distinguished by external aspect, or even by the opercular valves, from B. crenatus; but when the shell is disarticulated, the porose walls and non-porose basis of B. crenatus, allow of no mistake in the diagnosis of the two species.
ACASTA. Leach. Journal de Physique, tom. lxxxv, 1817.
Compartments six; parietes and basis non-porose: basis calcareous, cup-formed, not elongated, attached to sponges, or rarely to the bark of Isis.
Distribution mundane; imbedded in sponges and the sponge-like bark of Isis.
This sub-genus, in one sense, is a very natural one, inasmuch as all the species are closely allied in essential structure, in general appearance, and in habit. On the other hand, in the structure of the shell, in all the characters derived from the opercular valves and animal’s body, Acasta cannot properly be distinguished generically from some species of Balanus; thus B. navicula and cymbiformis agree in the parietes and basis not being porose and in all other essential respects, differing only in the shell being more elongated in the rostro-carinal axis and in being attached to Gorgoniæ instead of to sponges; yet we shall see that Acasta purpurata lives imbedded in the bark of Isis, so that even the habit of being imbedded in sponges fails. Balanus terebratus would have been ranked as an Acasta had it inhabited sponges. On the other hand, some species of Balanus inhabit sponges, as is often the case with B. spongicola, and always with B. declivis: but both these species are distinguished easily from Acasta, the former by its porose walls and basis, and the latter by its membranous basis; it may, however, be reasonably doubted whether such differences ought to be considered as even sub-generic. The most important character of Acasta probably consists in the anterior ramus of the fourth pair of cirri, differing slightly[Pg 303] in the arrangement of its spines, and in some other points, from the rami of the two posterior pairs of cirri,—a character not as yet observed in any other cirripede. Had not the genus Acasta been already founded and extensively admitted, certainly I should not have formed it; but considering the close similarity in habits, aspect, and structure, of the nine species of Acasta, and considering the already large size of the genus Balanus, I hope I may stand excused for admitting Acasta as a sub-genus.
General Appearance.—The shape varies from nearly globular to that of a somewhat flattened acorn, the orifice being often a little contracted from the inward curvature of the tips of the parietes. In A. spongites, however, the orifice is generally widely open; and, on the other hand, in A. sporillus, the orifice is reduced to a mere pore. The usual tint is pale reddish, but A. purpurata is purple, and A. sporillus purplish-brown. The surface is either smooth, or is shagreened with minute points, as in A. sporillus, and fenestrata, and in some specimens of A. sulcata; and in all the species, except A. sporillus and fenestrata, many individuals are furnished with elongated, curved, sharp, shelly points, like those in var. spinosus of Balanus tintinnabulum. The summits of the radii, which are generally of moderate breadth, are more or less oblique; their surface is often marked by lines parallel to the basis, instead of by vertical lines corresponding with the lines of growth, as in most species of Balanus. The carino-lateral compartments vary in proportional breadth in the different species: in A. sporillus, they tend to become rudimentary, and in this species (Pl. 9, fig. 9 b) their basal margins, or rather points, do not reach down to the basis. The species are all small, A. glans and undulata are the largest, being sometimes .55 of an inch in basal diameter.
Opercular Valves.—These differ in no generic respect from those of Balanus. The Scuta are striated longitudinally in several of the species: the adductor ridge is barely developed in any, being most prominent in A. cyathus. The articular ridge is prominent in A. fenestrata and purpurata. In the Terga, the spur is either truncated and very broad, or moderately narrow and bluntly pointed: the surface of the valve is often depressed, and in[Pg 304] A. spongites and fenestrata it is furrowed in the line of the spur. The articular ridge and furrow are well developed in A. fenestrata and purpurata. The crests for the tergal depressor muscles are either absent or very feebly developed.
Structure of the Parietes and Radii.—The parietes are not porose; internally, they are either smooth, or slightly, or strongly ribbed in longitudinal lines; the presence of these ribs, which are homologous with the parietal septa in Balanus, is variable even in the same species. In A. sporillus the inner surface is curiously reticulated. The sutural edges of the radii are either smooth, or very slightly crenated by the septa, in lines parallel to the basis. The upper margins of both radii and alæ are always more or less oblique. The radii sometimes do not extend down to the basis; and in this case, as will presently be described, apertures are left in the lower half, between the compartments. In A. glans and lævigata the internal margin of the wall of each compartment, from the sheath to the basis, projects inwards, forming inside the shell as many double ridges (Pl. 9, fig. 5 b), as there are compartments, namely, six: a nearly analogous structure occurs in certain species of Balanus. The basal edge of the sheath, in most of the species (5 b, 9 b), depends freely, and is hollow beneath, but this is always a variable point.
Basis.—The base is either saucer or cup-shaped, but in A. cyathus it is almost flat; it is generally symmetrical and smooth, with the lines of growth closely approximate. In A. fenestrata the basis is commonly as deep, as the shell is high. The edge, in several of the species, is crenated with minute teeth or notches; and these are so large in some specimens of A. sulcata and cyathus, as to make the edge almost pectinated. In A. glans, and in a lesser degree in A. lævigata, there are six knob-like teeth (fig. 5 a), corresponding with the points of junction, between the basal edges of the inwardly prominent margins of the six compartments, and the basis: in those specimens, in which the six teeth are largely developed, six ridges produced by their successive development, extend down towards the centre of the basal cup. When the basal cup is dissolved in acid, there is but a little animalised tissue and an external membrane, formed as usual in slips, and furnished with blunt[Pg 305] little external points (apparently representing spines), each of which has a short tubulus extending to the corium. Although I dissolved the basis of three specimens, I could not distinctly make out any cement; nor did I see any cement-ducts; yet these are readily distinguished, after the dissolution of the basis in acid, in Balanus, Elminius and Tetraclita. There can be no doubt that the young shell must at first be cemented to a fibre of the sponge; but I suspect that the cementing-tissue is not subsequently formed, owing to the support afforded by the growth of the enveloping sponge. As some species of Balanus are habitually or occasionally imbedded in sponges, it is important to observe, that the species of Acasta are not only imbedded, but are attached to the fibres of the sponge: but even this character, as we have already seen, is not sufficient to distinguish the genus Acasta from Balanus, for B. declivis is attached exclusively to sponge.
Perforations in the Shell.—Calcification seems often to fail to a certain extent in this genus: the basal cup in most specimens of A. spongites, and in some of A. glans and other species, is irregularly perforated by numerous minute orifices, closed only by the external membrane, and filled up inside by pulpy corium. In some specimens of A. spongites, from the Cape of Good Hope, parts of the basis were riddled like a sieve. I have seen similar perforations in the parietes of a few specimens of A. glans. In some specimens of A. sulcata, the radii do not extend quite down to the basal edge of the walls (Pl. 9, fig. 2 a), and in consequence a small cleft, closed only by membrane, is left between the compartments, for a little space above the basal cup. In A. fenestrata (fig. 7 a), and in a lesser degree in A. purpurata (8 a), not only do the radii not extend to the basal cup, but the parietes either on one or both sides of each suture are hollowed out, so that six, large or small, elongated, membrane-covered openings are formed, which extend from beneath the sheath down to the basal cup. These openings, which I have not seen in any other genus, will be more fully described under the respective species.
Mouth.—The parts of the mouth are identical in the several species, and present no generic differences from[Pg 306] those in Balanus. The outer maxillæ and palpi appear unusually prominent: the labrum is deeply notched, with no teeth, or very obscure teeth on each side. The mandibles have five teeth, but the fifth is sometimes confluent with the inferior angle. The maxillæ are not notched; and carry one or two spines, near their inferior angle, nearly as large as the upper pair. The outer maxillæ are bilobed.
Cirri: in the first pair, the rami are very unequal in length, the one ramus being from half to one-third of the length of the other. The segments in the second and third pairs, are not so much broader, or so much more crowded with bristles, in comparison with the three posterior pairs, as is the case with most species of Balanus. The three posterior pairs, except in A. purpurata, are much elongated, and the long thin segments bear four, and sometimes only three, pairs of spines, which are generally doubly and finely serrated, or even feathered. The most remarkable fact respecting the cirri, is, that in A. spongites, sulcata, cyathus, and glans, the fourth pair, instead of being identical in structure, as in all other genera, with the fifth and sixth pairs, has, on its anterior ramus, the pairs of spines more crowded together, with the little intermediate spines, and those in the dorsal tufts, a little longer than in the sixth cirrus; and between the pairs of spines, there occur some straight, upwardly pointed, very minute, and very thick spines or teeth. And, what is still more remarkable (as will hereafter be described in detail), in certain specimens, but not in all, of A. sulcata, the front surfaces of the lower segments on the anterior ramus, are developed into thick, small, downwardly curved, hook-like teeth; this likewise is the case with the upper segment of the pedicel (Pl. 29, fig. 2),—a most elegant, mandible-like organ for the prehension of prey being thus formed. The variability of such beautifully contrived teeth is very surprising. Some similar teeth occur on the segments of the anterior ramus of the fourth cirrus, but not on the pedicel, in A. cyathus. A few teeth resembling the above, but thicker, occur on the segments of the anterior ramus of the same cirrus, in A. purpurata.
Branchiæ, &c.—In A. spongites, I found the branchiæ rather small, with transverse plications. The muscles of the[Pg 307] sack, which run to the opercular valves, seemed rather feeble in most of the species. The penis in several species was remarkably long, and in A. spongites I noticed the straight projecting point at its dorsal base, as is common in Balanus.
Affinities, &c.—At the commencement of the description of the genus, I gave my reasons for keeping Acasta distinct as a sub-genus from Balanus. The species are particularly troublesome to identify, not only from the great variability of the most obvious characters, but from the very close general external appearance of most of the species, and the consequent necessity for cleaning and disarticulating at least one specimen in every group. The shape, however, of the shell and basis, and the state of their disarticulated edges, whether smooth, crenated, or toothed, here offer more serviceable, though still very variable, characters for the identification of the species, than is usual with sessile cirripedes; and this, probably, is in part due to the almost free or unattached condition of the whole shell, suspended, as it were, in the midst of sponges, which they inhabit. The opercular valves, on the other hand, are less serviceable than usual.
Range, Habitats, &c.—The species are found all over the world, excepting in the very cold latitudes.[102] Acasta lævigata ranges from the Red Sea to the Philippines; A. spongites from the south of England and Wales to the Cape of Good Hope; and A. cyathus from Madeira to the West Indies; most of the other species seem to have rather confined ranges. The East Indian Archipelago seems to be the metropolis of the genus, for here A. lævigata, fenestrata, purpurata, and sporillus, are all found. Of these four species, A. purpurata lives imbedded, not in sponges, but in the sponge-like bark of an Isis; and I think it probable, that A. sporillus may have somewhat analogous habits. The same species often lives imbedded in different kinds of sponge; thus, I have seen A. lævigata and cyathus in apparently three kinds, and A. spongites in, as I believe, four kinds. The genus existed during the miocene period, in the Coralline Crag, under a form closely allied to A. spongites.
[102] I am greatly indebted to Mr. Bowerbank for his great kindness, in looking over his immense collection of sponges from all parts of the world, and sending to me all the specimens of Acasta he could find imbedded in them.
LEPAS SPONGITES. Poli. Testacea utriusque Siciliæ, 1795, Tab. 6, fig. 3-6.
BALANUS SPONGEOSUS. Montagu. Test. Brit. Suppl., 1808.
------ SPONGITES. De Blainville. Dict. Sc. Nat., Pl. 116, fig. 3.
LEPAS SPONGIOSA. Wood’s General Conchology, p. 47.
ACASTA MONTAGUI. Leach, in Lamarck, Animaux sans Vertèbres, 1818, et in Encyclop. Brit., Supplement, 1824, vol. 3, Pl. 57.[103]
------ ------ J. E. Gray. Annals of Philosophy, New Series, vol. 10, Aug. 1825.
------ SPONGITES. Philippi. Enum. Mollus. Siciliæ, vol. 2, p. 211, 1844.
BALANUS MONTAGUI. Brown’s Illustrations of Conchology (2d edit. 1844), Pl. 53, fig. 24-26.
[103] As the plate to the Supplement to the ‘Encyclop. Brit.’ is marked as engraved in 1817, I presume Dr. Leach gave a proof to Lamarck, thus enabling him to publish this species four years before the Supplement itself appeared.
Carino-lateral parietes about one sixth of width of lateral parietes: inner surface of the parietes generally ribbed feebly: scutum with the articular ridge abruptly cut off at its lower end: tergum with the spur rounded-truncated, about one third of width of valve.
Hab.—South coast of England; South Wales, (Tenby); Portugal; Naples; Sicily; Cape of Good Hope.
This species and the three following, have caused me much doubt and trouble. At first I took the view here adopted, namely, that they were distinct: more careful examination made me run them altogether under one name. Finally, after still further deliberation, and the examination of a few additional specimens, I concluded there was the least chance of error in classing them separately. I may here mention, that in some sponge from the Cape of Good Hope, this species was mingled with Balanus spongicola.
General Appearance.—The shape is usually that of a cup, the orifice being, in most cases, rather large, and deeply notched, owing to the great obliquity of the radii and alæ. The surface is generally smooth, but furnished with sharp calcareous projections. The colour is pinkish, and chiefly in the upper part of the shell; the lower part is often yellowish from the preserved epidermis. The parietes in the carino-lateral compartments are always narrow, being only one sixth or one seventh of the width of the parietes in the lateral compartments. The radii are not very wide, never equalling in width the parietes. The basis is moderately deep, and sometimes very deep, being even occasionally curled like a horn on one side. The specimens from Lisbon and Naples are a little larger than any British specimen which I have seen; the former being .3 of an inch in basal diameter.
Scuta: these are striated longitudinally in close lines, generally plainly, but to a variable degree. The whole valve is flat, thin, and rather elongated, with barely a trace of an adductor ridge: the articular ridge is short and rather prominent: it terminates downwards abruptly, and this does not appear to be the case in the two following species. Terga: these are small compared with the scuta, they are slightly beaked: the spur is truncated but rounded, more especially on the carinal side; it is rather more than one third of the width of the whole valve. The articular ridge and crests for the depressor muscles are feebly developed.
Structure of the Parietes and Radii.—The state of the inner surface of the parietes varies much; generally they are slightly ribbed close to the basis, the ribs sometimes extending up to the sheath; rarely the surface is quite smooth. The edges of the radii are slightly crenated. The upper internal surfaces of the radii, where overlying the alæ, are usually marked by feeble undulating lines, nearly parallel to the basis. The alæ have very oblique summits.
Basis: this is generally of a regular cup-like form, and about two thirds as deep as the shell is high; sometimes it is pointed at the bottom and distorted. The edge is feebly crenated, and rarely quite smooth. It is often penetrated by small rounded irregular holes; and I have seen specimens from the Cape of Good Hope with parts like a sieve.
Cirri: in the first pair, the anterior ramus is nearly thrice as long as the posterior ramus. The second cirrus is short, with one ramus longer by three or four segments than the other ramus; the terminal segments are truncated. The third cirrus is about one third longer than the second cirrus. In the anterior ramus of the fourth pair, the regular pairs of spines are rather crowded together in the upper part of each segment, and the intermediate little spines and dorsal tufts are rather long in comparison with those of the fifth and sixth pairs of cirri: moreover, amongst the regular pairs, a few very minute and thick spines, pointing upwards, could be perceived. So that we here have the very unusual case of the fourth cirrus not exactly resembling the fifth and sixth pairs; and we shall see, in the following species, that this same anterior ramus of the fourth cirrus presents in addition another very extraordinary character. In the sixth cirrus there are on each segment four pairs of spines.
ACASTA SULCATA. Lamarck. Animaux sans Vertèbres, 1818.
------ ---- Deshayes, in Guerin, Magasin de Zoologie, 1831, Tab. 24.
Carino-lateral parietes about one sixth of width of lateral parietes: inner surface of the parietes generally ribbed strongly: basis with the edge strongly crenated: orifice of shell rather small: tergum with the spur generally truncated and nearly half as wide as valve.
Var. (a) (fig. 2 b): with the walls externally ribbed.
Var. (b) (fig. 2 a): with small membrane-covered clefts between the edges of the walls, close above the basal cup.
Hab.—Sydney, Port Fairy, Moreton Bay in lat. 27° S., New South Wales; Southern Australia; and, according to Lamarck, Western Australia, in lat. 25° S. Mus. Brit., Cuming, Bowerbank, &c.
I am almost ashamed to admit this species, so small are its differences compared with A. spongites; yet I think that it probably is a distinct form.
In general appearance and character this species comes very near to A. spongites. As in the latter, the parietes of the carino-lateral compartments are narrow.[104] The orifice seems always to be smaller. Internally, the parietes are generally much more strongly ribbed, and the edges of the basal cup more plainly crenated. The sheath is generally coloured of a brighter pink, sometimes tinged with orange. The average largest specimens (from .3 to .5 of an inch in basal diameter) are a little larger than the largest European specimens: I have seen one specimen from Moreton Bay .4 in basal diameter, and from the basal cup being very deep, actually .75 in height. The scutum has the articular ridge not so prominent and not so abruptly cut off at the lower end, as in A. spongites: on the other hand, the adductor ridge seems rather more prominent; but these differences are trifling. In the tergum the breadth of the spur (fig. 2 c, 2 d) varies in specimens taken out of the same branch of sponge; some can hardly be distinguished from the[Pg 311] same valves in A. spongites, but generally the spur is broader and squarer.
[104] In Mr. Cuming’s collection there is a specimen, from Sydney, which I fully believe to be the present species, but cannot be positively sure, as the opercular valves have been lost, which is very remarkable from the walls of the carino-lateral compartments being reduced to the thickness of a mere thread, not one twentieth of the width of the lateral compartments; we here see the structure of Acasta sporillus prefigured.
This species presents some remarkable varieties: in one specimen, from Sydney, the parietes were externally ribbed longitudinally (fig. 2 b), the ribs being roughened with minute points, giving to the shell an elegant appearance; and this without doubt is the A. sulcata of Lamarck, procured by Peron at Shark’s Bay, lat. 25° S., on the opposite of the Australian continent: some specimens from Port Lincoln, in South Australia, were not ribbed, only smoothly striated in longitudinal lines; although both sets of specimens had almost smooth scuta, and were thus different from common specimens, yet there could be no doubt, from their similarity in all other points, that they did not differ specifically from them, though the latter had their scuta striated, but not their parietes. Hence we see that there is no relation between the striæ on the parietes and on the scuta. The Port Lincoln specimens, and some others, were remarkable from the radii not extending down to the basal cup, a minute cleft, covered only by membrane, being thus left along the sutures, low down between the parietes (fig. 2 a); we shall see this singular structure strongly developed in A. perforata. Owing apparently to these clefts, the edge of the basal cup, exhibited traces of six knob-like teeth, like those characteristic of A. glans.
Cirri.—The cirri resemble those of A. spongites, with the exception that the segments on the posterior pairs bear only three main pairs of spines. With respect to the anterior ramus of the fourth pair of cirri, the following very singular facts were observed:—in a specimen from New South Wales (var. with the tergum having a narrow spur), on two or three of the lower, but not on the lowest, segments, the front margin was produced or developed into two or three minute, thick teeth, slightly curved like hooks downwards: in other specimens from New South Wales (var. with the tergum having a broad spur, and inhabiting the same branch of sponge with the last-mentioned variety), there was no trace of these teeth. But again, in two other specimens with the tergum having a broad spur (collected by different persons, near Sydney), and in another from South Australia, this structure was carried to an extreme, for in these (as represented, Pl. 29, fig. 2) there were beautifully formed teeth on the fourteen lower segments (the twelve upper being without them), and likewise on the upper segment of the pedicel. These teeth are graduated in size on each segment; they are admirably adapted for securing any prey; and, in fact, they convert each segment into a mandible-like organ. On the segments, on which these teeth are well developed, some of the regular pairs of spines are aborted.
Diagnosis.—Finally, this species, if it be, as I believe, distinct, differs from A. spongites only in the internal surface of the parietes being more strongly ribbed and brighter coloured,—in the edge of the basal cup being more plainly crenated,—in the articular ridge of the scutum being of a different shape,—in the spur of the tergum being often broader,—in the segments of the posterior cirri having only three pairs of main spines,—and, lastly, in the occasional presence of the hook-like teeth on the anterior ramus of the fourth cirrus.
Carino-lateral parietes about one fourth of width of lateral parietes: radii wider than the parietes: basis nearly flat, small: tergum with the spur truncated, half as wide as valve.
Hab.—Madeira, Mus. Lowe, and Bowerbank. West Indies, Mus. Stutchbury. Hab. unknown, Mus. Brit.
I feel more confidence in this case, than in that of A. sulcata, that we here have a distinct species, though at one time I treated it only as a marked variety of A. spongites. I rely chiefly on the great proportional width of the radii of all the compartments, and on the width of the carino-lateral compartments, compared with the lateral compartments, and on the general shape of the shell, which differs considerably from that of the two previous forms: in consequence of Acasta being attached to and imbedded in a yielding substance, such as sponge, I believe external form to be of more value as a specific character in this genus, than in most sessile cirripedes. I have examined specimens taken out of the yielding Spongia officinalis and out of an unusually compact sponge, and they resembled each other in every respect.
General Appearance.—Colour pale pink, or that of flesh: basis remarkably flat and rather small, with the walls above bulging out a little. The radii are very wide, being wider than the parietes to which they belong: the orifice is generally rather large. The parietes of the carino-lateral compartments vary from one third to one fourth of the width of the parietes of the lateral compartments. Basal diameter of largest specimen .35 of an inch. Internally, the parietes are generally more strongly ribbed than in A. spongites.
The Opercular Valves are large, owing to the form of the shell. The Scuta present no particular character, and are not distinguishable from those of A. sulcata; but the adductor ridge is perhaps rather more developed. The Terga (Pl. 9, fig. 3 c) are nearly as large as the scuta, and this is an unusual circumstance; the spur is more than half as wide as the valve; it is placed not quite close to the basi-scutal angle; on the carinal side, the basal margin of the valve slopes a little towards the spur. I may mention, that in several specimens from Madeira, the scuta and terga, on one side, had grown to a monstrous thickness.
Cirri: these resemble, in every respect, those of A. spongites, with the remarkable exception that on the anterior ramus of the fourth cirrus, several segments were furnished with the beautiful downward curved, mandible-like teeth, as in A. sulcata; but differently from in that species, there were none on the upper segment of the pedicel. I should have thought this an excellent specific character, had not these teeth been so extremely variable in A. sulcata.
Finally, I think this species is more nearly related to A. sulcata than to A. spongites.
Shell, apparently, as in A. spongites, but larger: scutum marked by longitudinal ridges, often in pairs, with the intermediate furrows rather wide: spur of tergum nearly half as wide as valve.
Fossil in Coralline Crag (Sutton), Mus. S. Wood.
I owe to Mr. Wood the inspection of a fine suite of separate valves. Owing to the shell never having been found entire, its general shape is not known, and, what is of more consequence, the relative proportional width of the parietes of the carino-lateral compartment is unknown. I have (but with doubt) given it a distinct specific name, owing to the peculiar character of the furrows on the scuta, and to the large size of the whole shell. In its other characters it comes nearest to A. spongites, excepting in the spur of the tergum, which resembles that of A. sulcata.
The compartments appear to have been rather smooth externally. The radii are not wide, as in A. cyathus; and the basis is cup-formed. Internally, the parietes are feebly ribbed, as in A. spongites. Judging from the dimensions of the separated valves, this species must have equalled and perhaps exceeded the size of the largest living species, namely, A. glans, from Australia. Hence we may infer, that the basal diameter probably exceeded .55 of an inch: I may add, that the largest European specimens of A. spongites, from Naples and Portugal, are only .3 of an inch in basal diameter.
Scuta.—These seem to resemble the scuta of A. spongites in all respects, except in the longitudinal ridges standing much further apart, and, consequently, in the furrows being much wider: each ridge is generally double. Although there is a good deal of variability in the character of these ridges in A. undulata, and likewise in A. spongites,[Pg 314] I have not seen any form intermediate between them. It must, however, be confessed, that this is an extremely variable character in many sessile cirripedes. In the Terga, the spur is about half the width of the whole valve, and therefore rather wider than in A. spongites.
ACASTA GLANS. Lamarck. Animaux sans Vertèbres, 1818.
Parietes internally quite smooth, with the lateral margins of each compartment inwardly prominent: basis with the edge rarely crenated, but furnished with six inwardly prominent teeth: scutum strongly striated longitudinally.
Var. (a) with the edge of the basal cup finely crenated.
Hab.—New South Wales, Southern Australia; Mus. Brit., Stutchbury, &c.
This fine species seems to be extremely common, imbedded in an open porose sponge on the eastern and southern shores of Australia. It is very distinct from the other species, with the exception of the following A. lævigata, which, with some hesitation, I have allowed to remain specifically separated.
General Appearance.—Excepting in its larger size, this species differs in external appearance but little from A. spongites; its colour is pale dirty reddish. The surface is generally studded with small calcareous points. The parietes of the carino-lateral compartments are about one fourth of the width of the parietes of the lateral compartments, and therefore proportionally of the same width as in A. cyathus. The largest specimen which I have seen, was .55 of an inch in basal diameter.
Scuta.—These are slightly narrower, thicker, and more convex than in A. spongites: they are strongly striated in longitudinal lines. The articular ridge is very feebly developed. Terga: in full-grown specimens, the spur is half the width of the whole valve, and is truncated; its basal edge being parallel to the basal margin of the valve. The articular ridge and crests for the depressor muscles are very feebly developed.
Internal structure of the parietes.—The inner surface of the parietes is quite smooth, without even a trace of ribs or teeth. But the most important character is that the internal lateral margins on both sides of each compartment, from the sheath to the basis, project inwards and form a rim; so that when the shell is viewed from within (Pl. 9,[Pg 315] fig. 5 b, representing the lateral and carino-lateral compartments, and part of the carina), the six sutures are seen to be strengthened by six double columns.
Basis.—This is moderately cup-formed. The edge, in order to meet the basal points of the inwardly projecting lateral margins of the six compartments, has six knob-like teeth. These are placed at unequal distances, for two on each side stand near each other, owing to the narrowness of the carino-lateral compartments. The degree of their development varies extremely; when most developed, as in the specimen figured (Pl. 9, fig. 5 a), each tooth is bifid and a little hollowed out, so as to receive the points of the two inwardly projecting margins which form each suture. Ridges, more or less prominent, running from each of the six marginal teeth, extend towards the centre of the cup. These six teeth cannot be seen from the outside. The edge of the cup is rarely crenated; but I have seen two instances in which this was the case.
Cirri.—In the first pair, the rami are not quite so unequal as in A. spongites; the longer ramus being about twice as long as the shorter. In the third pair, there are some very minute, thick, upwardly-pointing spines, which I did not notice in A. spongites. In the fourth pair, the spines are a little more crowded, with longer dorsal tufts, than in the sixth pair; and they are mingled with some very minute, thick, upwardly pointing spines. In young individuals, there are only three pairs of main spines on the segments of the sixth pair, instead of four pairs.
ACASTA LÆVIGATA. J. E. Gray (!). Annals of Philosophy, (new series), vol. 10, Aug. 1825.
Parietes internally quite smooth, with the lateral margins of each compartment inwardly prominent: basis with the edge strongly crenated, and furnished with six inwardly prominent teeth: scutum feebly striated longitudinally, or smooth.
Var. (a), epidermis coloured dull orange.—Red Sea.
Hab.—Red Sea, Philippine Archipelago; Mus. Brit., Cuming, &c.
This species, of which I have examined many specimens from the above two and other unknown localities, agrees in all essential points of structure with A. glans, and consequently I for some time classed them together; but the characters, though usually of small value, by which this form differs from A. glans being apparently constant, I[Pg 316] have with some doubt allowed it to remain specifically distinct. These characters are, firstly, the much smaller size of the whole shell in A. lævigata; secondly, the edge of its basal cup being always crenated, which seems to be a rare accident in A. glans; thirdly, though of secondary importance, the scutum being here less plainly striated; and, lastly, the spur of the tergum being of less breadth, and of a more rounded outline; on the other hand, it must be confessed, that when small specimens of A. glans are taken, there is hardly any difference in the spurs of the terga.
General Appearance and Structure of Shell.—The surface of the shell is often very smooth, but is sometimes studded with some small sharp calcareous points. The colour is white, or pale reddish-brown; but in the specimens from the Red Sea, the tint is more orange, with the upper part of the shell white. The orifice of the shell is unusually small. The largest specimen which I have seen was only .25 of an inch in basal diameter, and therefore less than half the size of A. glans. The internal surfaces of the parietes are smooth, with the two lateral margins inflected, as in A. glans. The edge of the basal cup has six knob-like teeth, like those in A. glans, but smaller; and, in addition, it is finely crenated.
Scuta: these differ only in being less plainly striated in longitudinal lines; indeed, some specimens show hardly a trace of this structure. Terga; these valves, in some varieties (Pl. 9, fig. 6 b) can hardly be distinguished from those of equal size from young individuals of A. glans; other varieties have the spur (Pl. 9, fig. 6 a) not truncated, but broadly pointed, and therefore of considerably different shape.
Neither in the mouth, nor cirri could I detect any difference with A. glans.
Shell reddish, with six large, membrane-covered apertures between the sutures, above the basal cup: carino-lateral parietes half as broad as lateral parietes; internally, parietes and edge of basis smooth; tergum with the articular ridge short and prominent; spur pointed.
Hab.—Philippine Archipelago, Mus. Cuming.
General Appearance.—Shell rather elongated or tubular; with the upper part reddish, and the surface roughened with very minute points. The basal cup is generally as deep as the shell is high, ending down[Pg 317]wards in a blunt point, often curved to one side. The summits of the radii, as usual, are oblique. The parietes of the carino-lateral compartments are about half as wide as the parietes of the lateral compartments, and are therefore of greater proportional width than in the foregoing or any other species of the genus. The large membrane-covered openings, or, as they may be called, windows, presently to be described, between the lower halves of the compartments, is much the most remarkable character of this species. The largest specimen which I have seen was only .23 in diameter, and .6 of an inch in height, measured from the basal point of the cup to the tips of the compartments.
Scuta.—These barely exhibit a trace of longitudinal striæ. The valve is rather thick and convex. The basi-tergal angle is much rounded off. Internally, the articular ridge is thick and rather prominent. Terga: the valve is furrowed in the line of the spur: the spur is pointed and rather long; it is distinctly separated from the basi-scutal angle of the valve, and the basal margin on the two sides of the spur forms a straight line. The articular ridge is prominent, and short.
Structure of the Parietes, Radii, and Basis.—The parietes are internally quite smooth down to its basis. The edges of the radii are also smooth, as is the edge of the basal cup. The alæ project less than usual. The radii are of moderate breadth, they extend downwards only a little below the sheath, namely, about half way down the shell, where they terminate, as usual, in a point. The increase in width, during growth, of the radii, and their not extending down to the basis, would necessarily cause a gap between the opposed edges of the walls, in the portion beneath the radii; but besides this, the edges of the walls themselves, beneath the radii, and on the opposed side beneath the alæ, are hollowed out, but on the latter side or beneath the alæ sometimes in a lesser degree. The result of this is, that the compartments, in their lower halves, are separated from each other by membrane-covered windows or apertures, arched at their upper ends, and of considerable size, namely, about as wide as the parietes of the carino-lateral compartments. I have only further to remark, that during the downward growth of the parietes, the apertures increase in size, but at the same time become closed up at their upper ends; and the arched layers of shell added at these upper ends, assume a very different aspect from the rest of the parietal surface,—appearing like two wedges, with their points upwards, let in, on one side of the suture, between the ordinary parietal surface and the radius, and, on the other side of the suture, between the ordinary parietal surface and the recipient furrow of the radius.
The animal’s body was in a bad state of preservation; but, as far as I could make out, the cirri resembled those of A. glans.
Affinities.—This species differs from A. glans in not having the internal margins of the compartments projecting inwards. It differs from all the ordinary varieties of A. spongites, in the smoothness of the basal edges of the parietes and of the edge of the cup; in the greater width of the carino-lateral compartment, though this is a variable point in A. spongites; slightly in the shape of the scuta and terga; and, lastly, in the large, membrane-covered openings between the compartments.
Shell dull blueish-purple, with six small, membrane-covered apertures between the sutures, close above the basis: tergum with the articular ridge very short and prominent; spur very broad and rounded.
Hab.—Sumatra; Philippine Archipelago; imbedded in the bark of an Isis; Mus. Cuming, Stutchbury, Brit.
This species is perfectly distinct from the others, as shown by its general appearance, its habits, and the structure of its opercular valves: it is allied to A. fenestrata, in having membrane-covered apertures between the compartments, and in some respects in its opercular valves: it is also allied to A. sulcata and cyathus in the parietes being often internally ribbed, in the basal cup having a crenated edge, and in the anterior ramus of the fourth cirrus being furnished with the minute hook-like spines.
General Appearance.—Sub-globular, slightly compressed, with a rather small orifice; smooth, but sometimes furnished with sharp shelly points; dull purple, more or less dark, with the upper parts of the walls often white. The radii are rather narrow, and generally white, with their summits only slightly oblique, but variable in this latter respect. The parietes of the carino-lateral compartments are narrow, being only one sixth of the width of the parietes of the lateral compartments. In some specimens there are membrane-covered apertures of considerable size, in others mere narrow clefts, between the basal halves of the compartments. The basal cup is moderately deep. The largest specimen was only .16 of an inch in basal diameter.
Scuta, rather broad, externally convex, not longitudinally striated: articular ridge prominent, short, not extending down above one third of the length of the valve. Depression for the adductor muscle deep. On the internal surface, near to the rostral angle, a rather large purple spot of corium adhered to the valve. Terga, broad, externally rather convex: scutal margin protuberant: carinal margin slightly inflected, or furnished internally with a rim: articular ridge prominent, very short, not extending down above one fourth of the valve. Spur very broad, rounded, confluent with the basi-scutal angle of the valve.
Internal Structure of the Parietes, Radii, and Basis.—The parietes, internally, are either quite smooth, or more commonly ribbed, with the[Pg 319] basal edge in consequence crenated; the ribs are either placed at an unusual distance from each other, and consequently are few in number, or are pretty close together. The edge of the basal cup is either quite smooth, or closely crenated, or distantly toothed, in conformity with the state of the internal surface of the parietes. The radii have nearly smooth edges, with their summits more or less oblique. They sometimes extend down only three fourths, or only two thirds, of the length of the shell, and the margins of the parietes under the radii being a little hollowed out, the sutures are converted into clefts or apertures (of course covered by membrane) like, but not so large as, those in A. fenestrata. The margins of the parietes are hollowed out only on the side of the radius, and not on both sides of the sutures, as is most usual in A. fenestrata. In some specimens the radii extended down close to the basal cup, and only very minute clefts were left between the opposed edges of the parietes.
In the animal’s body the only noticeable character was, that on the anterior ramus of the fourth pair of cirri, some of the segments were furnished with very broad and thick, small, downwardly curved, teeth or hooks, like those described in certain varieties of A. sulcata; but they are here stronger and thicker. The segments in the three posterior pairs of cirri are not so much elongated, as in the other species.
Shell purplish-brown, with the parietes internally strongly ribbed and reticulated: carino-lateral compartments extremely narrow, not extending down to the basis.
Hab.—Sooloo Islands, East Indian Archipelago; Mus. Dana.
I am indebted to Mr. Dana, the distinguished naturalist of the United States Antarctic Expedition, for two specimens of this interesting species, which, in the singular reticulated structure of the inner surface of the walls, and in the almost rudimentary condition of the carino-lateral compartments, not extending down to the basal cup, is very distinct from the foregoing species. I have used Mr. Dana’s very appropriate MS. name of sporillus. The specimens were dredged up, lying quite loose and unattached at the bottom of the Sooloo sea; the one which I opened, must have long lain dead; but Mr. Dana assures me that some were living, and he has sent me drawings of parts of the mouth and cirri: I am much surprised at this circumstance;[Pg 320] for analogy would have made me believe that this species must have been imbedded in some sponge-like body, such as the bark of a zoophyte, and that it could not have lived unattached. I may add that a small fragment of a brown leathery substance adhered to the upper end of one of the two specimens, and this seems to indicate attachment.
General Appearance.—Shell shaped like a pointed acorn; slightly flattened; orifice extremely small; surface very finely punctured, covered by a purplish-brown epidermis, with transverse stripes of different shades, and with the apex dark; according to Mr. Dana, when fresh, the colour was purplish-carmine. Radii narrow, white. The carino-lateral compartments are extremely narrow; the wall-portion (fig. 9 b) forming a mere linear rib, terminating downwards in a sharp point, which does not reach the basal cup: hence this compartment evidently tends to become rudimentary. The basal cup is moderately deep and pointed. Basal diameter .16; height, from the bottom of the cup to the top of the shell, .24 of an inch.
Scuta: narrow, with the upper part produced; not striated longitudinally; coloured by a pale purple, longitudinal band. Internally, there is scarcely a trace of an articular ridge, which, in the other species, is always more or less developed. Terga with the spur bluntly pointed; nearly the whole basal margin, on the carinal side, slopes towards the spur.
Internal Structure of the Parietes, Radii, and Basal Cup.—The parietes are strongly ribbed internally; and these ribs are connected by very narrow, less prominent, transverse, slightly branched ridges, giving a reticulated structure to the inner surface. Between several of the main longitudinal ribs, in the lower part of the shell, new ribs may be seen in process of formation, and these tend to convert the reticulated structure into a double row of minute cells. I have not met with an exactly similar structure in any other cirripede; but I have no doubt that the little transverse ridges are homologous with the transverse calcareous septa in the parietal pores of many Balani, in the same manner as the internal longitudinal ribs, in this and other species of Acasta, are homologous with the longitudinal septa forming the above pores. The edge of the basal cup is pectinated with teeth, which lock into the teeth formed by the ends of the internal parietal ribs. The radii are narrow, and have smooth edges. The alæ project beyond the parietes to a remarkably small extent. The sheath is free, or hollow beneath. I have already described the almost rudimentary condition of the carino-lateral compartments; this is best exhibited in an internal view of the two compartments, as given in Pl. 9, fig. 9 b.
Animal’s body unknown to me: from Mr. Dana’s drawing the three posterior pairs of cirri seem to have been much elongated: and the rami of the first pair, as usual, unequal in length.
M. Deshayes has given an indifferent figure and imperfect description of ACASTA TUBULOSA (Guerin, Magasin de[Pg 321] Zoologie, 1831, Tab. 39; and Guerin, Iconographie du Règne Animal, Mollusques, Tab. 38, fig. 4, but here by a misprint called A. spinulosa); it is utterly impossible to recognise the species of this genus from such materials.
TETRACLITA. Schumacher. Essai d’un Nouveau Syst., &c., 1817.
CONIA. Leach. Journal de Physique, tom. 85, 1817.[105]
ASEMUS. Ranzani. Memoire di Storia Naturale, 1820.[106]
POLYTREMA. De Ferussac. Dict. Classique d’Histoire Naturelle, 1822.
LEPAS. Gmelin. Systema Naturæ, 1789.
BALANUS. Bruguière. Encyclop. Method., 1789.
------ Lamarck. Animaux sans Vertèbres, 1818.
[105] From a note by the Editor, it appears that Schumacher’s essay appeared before the number of the Journal containing Leach’s paper, so that Schumacher’s name must be adopted.
[106] I have not seen a complete copy of this work, and give the title from a catalogue; the running heading of the part containing the Cirripedia, is “Opuscoli Scientifici.”
Compartments four, sometimes externally calcified together: parietes permeated by pores, generally forming several rows. Basis flat, irregular, calcareous, or membranous.
Hab.—Throughout the tropical and warmer temperate seas.
General Appearance.—The shell is conical, more or less depressed, and very rarely, even when growing in crowded groups, becomes cylindrical or elongated. The orifice is seldom large, generally diamond-shaped or oval. The colour is either nearly white or purple, occasionally even inky black, or very dark green, and sometimes of a pale pink peach-blossom. The surface is sometimes smooth, but more commonly longitudinally ribbed; the outer lamina of shell is very often wholly corroded away, excepting close to the basis, leaving the solidly upfilled parietal tubes exposed: these give the shell a striated appearance, or they appear like flattened tapering points adpressed to its surface (Pl. 10,[Pg 322] fig. 1 b): Lamarck attempted to express this appearance, by using the specific name of stalactiferus. The colour of the shell depends, to a considerable extent, on the colour of the shelly matter in these exposed parietal tubes. We shall presently see that the corrosion and disintegration, to which some of the species are so liable, plays an important part during their growth. The radii are either well developed, as in most of the species; or they are entirely absent, as in the great majority of specimens of T. porosa and serrata. In many individuals of T. porosa and purpurascens not only are the radii absent, but the four compartments are calcified together without any trace, on the external surface, of the sutures. The largest specimen which I have seen of T. porosa, which is the largest species, was two inches in basal diameter, and nearly one inch and a half in height.
Scuta.—These valves are sub-triangular, and either longitudinally or transversely elongated. Externally, the growth-ridges are moderately prominent, and in T. costata and cœrulescens they are crossed by longitudinal striæ. Along the occludent margin, the inflected extremity of each alternate growth-ridge is generally much thickened,—a set of teeth being thus formed, by which the two valves are locked together. In T. porosa, this character is variable, for sometimes every alternate ridge, and sometimes only two or three ridges, separated from each other by several growth-ridges, are thus developed into teeth. The articular ridge is either moderately prominent, or is extremely prominent, as in T. cœrulescens; but the lower edge in no case depends as a free, hinge-like style, as sometimes in Balanus. The adductor ridge is generally well developed and distinct from the articular ridge: in T. purpurascens it is very blunt: in T. serrata it is united to the articular ridge half way up it, thus forming a deep tubular cavity running up to the apex of the valve: in T. cœrulescens, the adductor ridge is very short, and is united to, or almost continuous with, the lower end of the articular ridge, a small sub-cylindrical tubular cavity being thus formed. Small crests exist for the attachment of the rostral and lateral depressor muscles, in most of the species, excepting T. purpurascens and costata, in which, however, more especially in the former, there are,[Pg 323] instead of crests, minute pits for the attachment. These crests vary much in prominence in the same species.
Terga.—These valves present no essential differences from those of Balanus; they are sometimes beaked, and the beak is hollow and occupied by a thread of corium, as in that genus. The external surface of the valve is often depressed in the line of the spur, but there is never a longitudinal furrow with the edges folded in, as in Balanus. The spur is very short in T. purpurascens. In T. radiata, the articular ridge is remarkably prominent. The crests for the depressor muscles are well developed in all the species. The shape of the terga is variable in nearly all the species, and greatly so in T. porosa.
Compartments.—Owing to there being only four compartments, the lateral pair are large; the size of the carina relatively to the rostrum varies, according as its alæ have been added to during diametric growth. The walls are very thick, and are composed of numerous tubes, in some species as many as fourteen or fifteen rows being exposed on the basal margin (Pl. 10, fig. 1 g). The tubes are generally angular, and slightly elongated in the ray of the circular shell; sometimes they are nearly circular and small. New tubes are formed only at the basal edge of the outer lamina, by the bifurcation of the septa which form the tubes. In very young specimens there is only a single row of tubes; and in T. rosea this holds good throughout life: in this species (fig. 3 d) the tubes, in the single row, are large and quadrangular, and the outer lamina of shell is strengthened by numerous, small, internal, longitudinal plates. I believe the branching septa, which separate and form the parietal tubes, correspond with the longitudinal septa in the more simple walls of Balanus. The tubes become solidly filled up, in their upper parts, with hard, and generally coloured shelly matter. The degree to which they are filled up differs in the different species; the external side of each tube is always first thus coated. The thin outer lamina of shell, in several of the species, commonly disintegrates and disappears; the upfilled parietal tubes being thus exposed. The inner lamina of the walls is generally smooth, but in T. radiata it is longitudinally ribbed, as in most species of Balanus. The sheath is generally dark-[Pg 324]coloured; its lower edge does not project or overhang the inner lamina, as is usual in Balanus, excepting in T. serrata, and in some few varieties of T. porosa.
The Radii, when developed, are either narrow or broad, with their summits either oblique or extending in a straight line from the top of one compartment to that of another. In T. serrata, I have not seen a single specimen with the radii developed; in T. porosa, they are very seldom developed, and then, apparently, only in quite young specimens, in which they are narrow; in T. purpurascens, they seem to be about as often developed as not, and when present they are broad; in T. costata, cœrulescens, and radiata, they are always largely developed. In some specimens of the species, in which the radii are not developed, even the sutures do not reach the external surface; the outer lamina of the parietes being continuous all round, so that the shell seems formed of a single piece. Even in such specimens, the four compartments, viewed from within, can be seen to be distinct; and the sutures can generally be traced across the whole thickness of the parietal tubes; in this latter case, when the sutures are broken open, the radii are seen to be represented (Pl. 10, 1 h) by a few small sinuous ridges. Owing to the disintegration of the upper and outer part of the shell, and the consequent exposure of parts of the sheath and alæ, the radii sometimes appear as if developed, when such is not the case. With respect to the internal structure of the radii, they are formed, in T. purpurascens and costata, of tubes, like those of the parietes, and therefore according to the normal plan; whilst in the other species they are formed by longitudinal sinuous ridges, sending out on each side irregular denticuli; and the interspaces between the ridges are filled up solidly during the growth of the radii, in all the species, except in T. radiata, in which they are left to a considerable extent open. These sinuous ridges, with their denticuli, homologically represent the branching septa which form the parietal tubes. The edges of the alæ are crenated in all the species, except in T. costata.
Diametric growth.—When first examining groups of T. porosa, in none of which the radii had been developed, and in which, consequently, the shell could not have grown in[Pg 325] diameter, but only at its basal margin, I was at first unable to comprehend, how the upper part of the shell and the orifice could have acquired their proper proportional width. The young shell, at its first formation, starts with an orifice so small that a pin could hardly be inserted in it; and this in many individuals is never increased in diameter by the diametric growth of the shell; but in place of this, as the conical shell is added to at its base, the whole upper part disintegrates and wears away, the orifice becoming thus enlarged. We thus see that the corrosion and wearing away of the upper part of the shell is a necessary element in its growth. The development of the radii, which in some of the species, as in T. purpurascens, at first seems to be quite capricious, really depends upon the fact, whether the specimens have been exposed to disintegration; for I have almost always found that when the outer lamina of shell has been well preserved, the radii have been developed, and the orifice has been enlarged by their growth, instead of by the wearing down of the upper part of the conical shell.
Basis.—This consists of a very thin, flat, though irregular, translucent, calcareous plate, which towards the edges is sometimes membranous. In T. purpurascens, the basis is entirely membranous. When a portion of the calcareous base is dissolved in acid, a tissue is left, composed of several laminæ, to which numerous bifurcating cement-ducts are attached: even before dissolution, these delicate bifurcating ducts can just be perceived by the aid of a simple lens.
Mouth.—The several organs present no particular characters. There are generally three teeth on each side of the notch in the labrum. The palpi usually have parallel sides, but are club-shaped in T. purpurascens and costata. The mandibles have generally four teeth, but there are five in T. vitiata, and only three in T. costata. The maxillæ are notched. The outer maxillæ are bilobed in front.
Cirri.—The segments of the three posterior pairs usually support only three pairs of main spines, but there are four pairs in T. vitiata and costata: between each pair, there is either a tuft of fine spines, or a single fine spine. The rami of the first cirrus are unequal in length. In the third cirrus, the posterior ramus is sometimes much elongated, but some[Pg 326]times both rami are short and blunt. Some of the segments in the third cirrus often support very coarsely and doubly pectinated spines. Under the head of T. porosa, it will be seen to what a remarkable degree the relative numbers of the segments in the several cirri vary, even in specimens taken out of the same cluster.
The Branchiæ are well developed (at least in T. porosa and costata), as a large, plicated, tapering fold, with a small second fold on the inner side at the base. In T. porosa the stomach is destitute of cæca. The vesiculæ seminales in this same species are large, with their broad, blunt ends reflexed. The ovarian tubes surround the sack, and cover the basal plate.
Distribution and Habitats.—This genus is confined to the tropics, and to the warmer parts of the temperate seas: in the southern hemisphere, it ranges south, to the Cape of Good Hope and to Van Diemen’s Land: in the northern hemisphere it does not appear to range so far; I do not know of any authentic case of a species having been found in the Mediterranean, or on the shores of the United States, north of the West Indies. Tetraclita porosa is found round the whole world; T. radiata, also, has a very wide range, inhabiting the West Indies, the East Indian Archipelago, and New South Wales. This latter species, as well as T. cœrulescens, is often attached to the bottoms of ships, adhering to Balanus tintinnabulum. The several species live attached to tidal rocks, to littoral shells, or to massive corals. I have met with two instances, in the West Indies and the Philippine Archipelago, of T. porosa adhering to wood. Tetraclita porosa seems to feed chiefly on crustaceans: the number and the size of the amphipods, isopods, and entomostracans, together with an annelid, in the stomachs of some specimens from South America, was quite surprising. As many as five species occur in the same region, in the eastern half of the world; thus on the shores of New South Wales, we have T. porosa, vitiata, radiata, purpurascens, and rosea; in the Philippine Archipelago, we have T. porosa, vitiata, costata, and cœrulescens.
I have not seen any species of this genus fossil.
Variation.—The species vary in shape nearly, but not[Pg 327] quite as much as in Balanus, for we very rarely here see cylindrical varieties; but in the same species, we have extremely depressed, steeply conical, and convex forms; the orifice varies in relative size; the state of the surface, whether ribbed or smooth, whether well preserved or corroded, the upfilled parietal tubes being thus exposed, varies more than in Balanus. The colour also varies; specimens of T. porosa (Pl. 10, fig. 1 a to 1 f) being dark purple, or even inky black, white, pale pinkish-purple, and green: as far as I have seen, the pinkish varieties of T. porosa are confined to the eastern half of the world. Even the ridges and crests on the under side of the scutum vary in some degree; and the tergum in T. porosa varies considerably, and in some of the other species, slightly, in general shape: I believe that the tergum becomes narrow and elongated, when the shell is steeply conical, with the orifice of small diameter. After having spent several days in disarticulating and closely examining the many specimens of T. porosa in my possession, I concluded that its varieties formed at least four species, these being in external appearance extremely distinct; but ultimately the many intermediate forms compelled me to unite all into a single species. Again, I may instance the conical, ribbed variety of T. purpurascens (Pl. 11, fig. 1 b), with the outer lamina of the shell preserved and with the radii widely developed, as having not the smallest resemblance to the other common depressed variety (fig. 1 a), having a granulated surface, produced by the exposed tips of the upfilled parietal tubes, and without a trace of the radii or even of the sutures; I may add that, according to the characters used by some authors, these two varieties would be classed in two separate genera.
To distinguish the species of this genus, the chief reliance must be placed (as in the case of most other sessile cirripedes) on the general outline of the opercular valves, and on the ridges and crests on their under sides: the internal structure, however, of the radii, and in a lesser degree of the parietes, must not be overlooked. I have not found the parts of the mouth, or the differences in the cirri, of much service; and it will be shown under T. porosa, that the[Pg 328] relative numbers of the segments in the several cirri, and even their shape, is extraordinarily variable.
Affinities.—All the species are pretty closely allied, and there is no ground for making any sectional division of the eight species, more especially not on the ground whether or no the radii are developed. Tetraclita purpurascens, taking into account all the characters, including the animal’s body, is perhaps the most distinct species in the genus, but it is closely allied to T. costata. In the well-marked character of the parietes being formed of a single row of large tubes, T. rosea differs from all the other species. The genus is closely allied to Balanus; I can point out no difference in the animal’s body, nor any constant difference in the opercular valves. The ridges and crests on the under sides of the scuta are more prominent in Tetraclita; and all the species, excepting two, have crests (though sometimes very slight) for the attachment of the rostral depressor muscle, and these occur only in two species of Balanus: crests, also, for the attachment of the lateral depressor muscle are common in Tetraclita, but rare in Balanus. The main character, however, of the genus, as compared with Balanus, is derived from the existence of only four compartments, and in a lesser degree from the several rows of parietal pores; but, as just stated, T. rosea has only a single row, and, on the other hand, in Balanus crenatus, there is a slight tendency exhibited, in the dividing of the longitudinal septa near the outer lamina, to form a second row of parietal pores; and in B. cariosus, moreover, we actually have two or three rows of irregular and very short pores. The thin, yet solid calcareous basis which occurs in all the species of Tetraclita, excepting T. purpurascens, resembles the basis in Balanus flosculus and imperator, but I suspect that the structure of the cement-ducts is different in the two genera.
LEPAS POROSA. Gmelin. Syst. Naturæ, 1789.[107]
BALANUS SQUAMOSUS. Bruguière. Encyclop. Method., 1789, Pl. 165, fig. 9, 10.
LEPAS FUNGITES. Spengler. Skrivter af Naturhist. Selskabet, 1 B., 1790.
---- POROSA. Wood’s General Conchology, Pl. 9, fig. 4, 1815.
TETRACLITA SQUAMULOSA. Schumacher. Essai d’un Nouveau Syst., &c., 1817.
BALANUS STALACTIFERUS. Lamarck. Animaux sans Vertèbres, 1818.
------ ------------ Chenu. Illust. Conch., Pl. 4, fig. 6, 7.
ASEMUS POROSUS. Ranzani. Memoire di Storia Naturale, Tab. 3, fig. 32-35.
CONIA POROSA. Sowerby. Genera of Recent and Fossil Shells, Plate, 1823.
---- ---- Leach. (sine descript.) Encyclop. Brit. Supplement, vol. 3, 1824, Tab. 57.
[107] As Gmelin and Bruguière published the same year, I do not know which comes first, but I have adopted the best known name. Most authors give, amongst their references, Chemnitz, vol. 8, Tab. 98, fig. 836, 837, but the accompanying description is more applicable to T. serrata of this work than to T. porosa; without a figure or description, however, of the under side of the scutum it is impossible to decide. Several authors, also, give Lepas cariosa of Pallas, as a synonym; this, however, is the Balanus cariosus of this work.
Radii rarely present, when present narrow; even the sutures often absent: shell steeply conical, with the surface generally corroded, and having a stalactiferous appearance.
Var. (1) communis (Pl. 10, fig. 1 a): outer lamina of shell almost wholly removed; the portion preserved, as well as the exposed parietal tubes, gray, or pale dirty brown, or dull purple.
Var. (2) nigrescens (Pl. 10, fig. 1 c): outer lamina of shell almost wholly removed; the portion preserved, and the exposed parietal tubes, very dark purple or inky black.
Var. (3) viridis: outer lamina of shell almost wholly removed; the portion preserved, and the exposed parietal tubes, green; under surface of opercular valves clouded green.
Var. (4) rubescens (Pl. 10, fig. 1 b): outer lamina of shell almost wholly removed; the portion preserved, and the exposed parietal tubes, pale reddish-purple, or peach-blossom purple; under surfaces of opercular valves often reddish purple; terga often rather narrow, with the spur somewhat pointed, and with the basal margin on the carinal side sloping, or not making an angle with the spur.
Var. (5) elegans (Pl. 10, fig. 1 d): outer lamina preserved, excepting sometimes near the summit of the shell; white, tinged with yellowish brown from the epidermis; surface strongly ribbed longitudinally; orifice rather small; sheath reddish-purple; terga narrow, with the basal margin sloping as in var. rubescens.
Var. (6) communis (young) (Pl. 10, fig. 1 e): radii developed, very narrow; outer lamina of shell preserved, gray or dull purple; surface slightly ribbed longitudinally.
Var. (7) patellaris (Pl. 10, fig. 1 f): radii developed, very narrow, white; outer lamina of shell preserved, generally reddish-purple; steeply conical, with the orifice extremely small; surface smooth, with longitudinal white ribs. Terga very narrow, with the spur sharply pointed, and with the basal margin on the carinal side sloping towards it, or not making an angle with it. Scuta, with the adductor ridge very prominent. Attached to a ship’s bottom.
Hab.—West Indies, Brazil, West Colombia, Panama, Galapagos Archipelago, California, Philippine Archipelago, China, East coast of Australia, Red Sea; generally attached to tidal rocks, sometimes to shells, sometimes to wooden posts. Very common.
General Appearance.—This, the widest-distributed and much the commonest species of the genus, varies greatly in external appearance. The usual shape is steeply conical, but some individuals are much depressed. In the common varieties the outer lamina of shell has been removed even close to the basal edge; the upfilled parietal tubes being thus exposed (fig. 1 b), as flattened adpressed points. These points are largest in large specimens, but they vary somewhat in size in specimens of equal growth. When the outer surface is preserved, it is generally ribbed longitudinally, but is sometimes quite smooth. The most general colour is dirty gray or dark purple; but many specimens are pale pinkish-purple, owing to the exposure of the parietal tubes upfilled with shelly matter of this tint: there are also, as given under the characters of the vars., black, white and green varieties. The sheath is always tinted by the prevailing colour. The radii are rarely developed, but generally the four sutures are distinct; sometimes these are externally quite obliterated, the shell, as seen from the outside, consisting of a single piece like a patella or fissurella. When the radii are developed they are very narrow, with their upper edges oblique: their development seems always coincident with the more or less perfect preservation of the shell, and their function is to enlarge the orifice; the enlargement being usually effected by the disintegration and removal of the whole upper part of the conical shell. The size of the orifice varies considerably; in the seventh variety it was extraordinarily small: in outline it varies from oval to rounded trigonal or rhomboidal; in some specimens, with the radii well developed, it was rounded pentagonal.
Size.—Tetraclita porosa is the largest species of the genus; I have seen specimens attached to a large pebble of granite in the British Museum, which measured two inches in basal diameter, and nearly one inch and a half in height.
Scuta: these are sub-triangular and generally a little elongated, but they vary slightly in relative breadth (fig. 1 i, 1 l), and likewise in the degree to which the basi-tergal angle is rounded off. The under surface is clouded with dull or pinkish-purple, or with green, or is nearly white. The articular ridge is not prominent, and the articular furrow is narrow. The adductor ridge is prominent, and runs upwards for some distance close and parallel to the articular ridge; and sometimes it extends nearly or quite up to the apex of the valve; in one single specimen the adductor ridge had an abraded appearance, and was very little developed. The crests for the rostral and lateral depressores are sharp and distinct. Along the occludent margin, the ends generally of the alternate lines of growth are enlarged into knobs, serving to lock the two valves together; but in many specimens only two or three knobs, at intervals of several lines of growth (fig. 1 b), are developed.
Terga: when the upper end of the valve is not corroded, there is a distinct beak, hollow within for a thread of corium. The scutal margin is not much inflected, and the articular ridge not very prominent. The spur is placed quite close to the basi-scutal angle of the valve, so that there is no basal margin on that side of the valve. The width of the valve and of the spur, and the acumination of the extremity of the latter, varies in a remarkable manner. In the broad and commonest variety (fig. 1 k), the basal margin of the valve form an angle of about 130° with the carinal side of the spur, and the basal end of the spur is broad and truncated. In the less common and narrow variety (1 m), the basal margin in some extreme cases forms very nearly a straight line with the carinal side of the spur; and the spur itself is bluntly pointed: in var. 7 it is sharply pointed.
Structure of the parietes and radii.—In all cases the four sutures are quite distinct, from top to bottom, on the internal lamina of the shell, and generally they run through the whole thickness of the walls, and are visible externally. Often they do not quite reach the outer lamina, and then the shell externally consists of a single piece, like a patella. Sometimes the sutures can be traced running through the parietal tubes only for a short distance from the internal surface; where they cease, the two walls of the suture become fused together. When a perfect suture is split open, the radius is represented (fig. 1 h) by a few narrow, sinuous ridges, sending out on each side little branches or denticuli; these are received into corresponding furrows in the opposed compartment. These ridges run nearly parallel to each other, and somewhat obliquely, from the outer lamina of the shell to the basis. When the radii are developed, their edges are similarly formed, by sinuous denticulated ridges, with the interspaces between them filled up solidly. The alæ are but little prominent.
The mouth does not deviate from the generic type. The cirri are remarkable from the variability in the several pairs of the relative numbers of their segments, as shown in the following table. The segments do not correspond even on opposite sides of the same individual, as may be seen in the two lower lines of the table. I believe that variability to this degree is very uncommon in other cirripedes, though, as stated in the Introduction, the number of the segments always increases with[Pg 332] the growth of the individual. The terminal segments in the longer rami of both the first and third cirrus are antenniformed,—being elongated, and of a different shape, with fewer bristles, compared with the basal segments of the same cirri. It is apparently these terminal segments which are particularly liable to vary in number. In both rami of the third cirrus, some of the segments, from the sixth to the eleventh inclusive, (counting from the bottom), more especially the eighth, ninth, and tenth, carry a few spines coarsely and doubly pectinated; but as some of the adjoining segments carry spines which may be called doubly serrated, it is not easy to draw an exact line of demarcation. Sometimes, though rarely, a few of the nearly terminal segments in the second cirrus are furnished with similar, doubly pectinated spines.
| First cirrus. | Second cirrus. | Third cirrus. | Sixth cirrus. | ||||
|---|---|---|---|---|---|---|---|
| Shorter ramus. | Longer ramus. | Shorter ramus. | Longer ramus. | Shorter ramus. | Longer ramus. | Either ramus. | |
| Specimen from Pernambuco, Brazil (var. 1) | 9 | 19 | 10 | 11 | 15 | 27 | 26 |
| Another specimen from the same cluster (var. 1) | 12 | 23 | 12 | 14 | 14 | 19 | 23 |
| From the Galapagos Archipelago, (var. 1) | 10 | 16 | 10 | 11? | 11 | 16 | 25 |
| From the Philippine Archipelago, (var. 4) | 10 | ? | 10 | 10 | 11 | 19 | 18 |
| From California (var. 5) | 15 | 24 | — | — | — | — | 22 |
| From the bottom of a ship (var. 7), young | — | — | 9 | 9 | 10 | 17 | 14 |
| From South America (var. 1) British Museum | 11 | 18 | 11 | 12 | 13 | 21 | 22 |
| Another specimen from the the last specimen | 16 | 25 | 13 | 18 | 15 | 23 | 27 |
| Same individual, but the cirri from the opposite side of the body | 15 | 28 | 15 | 21 | 16 | 23 | — |
Varieties.—Under the generic description, I have stated that after having spent some time in examining a very large suite of specimens of T. porosa, I concluded that at least four of the varieties were true species. It so happened that all the specimens which I first examined of the var. (4) rubescens, had the narrow sloping terga, and scuta with only two or three great teeth on their occludent margins; but ultimately, in a group thus characterised, I found one or two individuals with terga precisely of the shape of those in var. (1) communis. Again, in a group of dull purple specimens of var. communis, a few had the narrow sloping terga, and scuta with teeth on their occludent margins, intermediate in size and number between the varieties with only one or two[Pg 333] great teeth, and those with every alternate growth-ridge enlarged into a tooth. Hence var. rubescens completely broke down as a species.
With respect to var. (5) elegans (a M.S. specific name of Leach) I inferred at first, from external appearance alone, that it was distinct; the outer lamina of the shell and even the epidermis is preserved; the surface is strongly ribbed, and the whole shell, excepting the sheath, is nearly white; the terga are narrow, with a sloping basal margin as in var. (4) rubescens. Whole groups of specimens are thus characterised. But as var. communis is often white, and as the surface, when the outer lamina is preserved, is generally, as we shall presently see, ribbed longitudinally, the differences in var. elegans are quite unimportant.
The (6th) var. differs from var. communis only in the narrow radii having been developed, and consequently in the orifice being pentagonal, and in the outer, longitudinally ribbed lamina of the shell having been preserved. In the same group of specimens, I have seen every intermediate stage between this and the common form. It must not, however, be supposed that the young of var. communis always pass through these stages, for such is not the case. In the genus Balanus it has been seen, how capricious in some species is the development of the radii.
Of the other varieties, enumerated at the beginning, no further mention is required, excepting with respect to var. (7), the most singular of all. I have seen only three specimens, sent to me by Dr. Aug. Gould, of Boston, United States, and these from the appearance of their bases I have no doubt had been attached to a ship,—the only instance which I have met with, in the present species. The shell is steeply conical, with the orifice so small as to be reduced to a mere pore; the radii are extremely narrow and white; the shell is thin, with the surface smooth, but ribbed longitudinally and regularly; the outer lamina of the shell and the epidermis are perfectly preserved; the upper part of the shell is reddish purple, which dies away towards the base: careful examination of the apex shows that at the first growth the young shell was blueish-green. The terga are narrow, with a sloping basal margin, as in var. rubescens, but with the point of the spur sharper. I have formerly remarked that the shape of the terga seems influenced by the size of the orifice. The lower edge of the sheath depends freely: I have seen no other instance of this latter structure, so common, but so variable, in Balanus, in the present species, except to a partial extent in one distorted specimen, in Mr. Stutchbury’s collection, adhering to Balanus tintinnabulum, and probably taken from a ship’s bottom. I may add that this distorted specimen was remarkable from its radii being wider than in any other instance,—from its smooth uncoloured surface without longitudinal ribs,—and from the perfect preservation of the epidermis over its entire surface. Although Dr. Gould’s specimens, in external aspect, are absolutely and entirely different from the common varieties of T. porosa, there are so many intermediate forms, and the differences are so little important that I feel no hesitation in attributing them to variation, consequent on the individuals having been exposed to unusual conditions, attached to the bottom of a ship.
Shell dark greenish-gray, with narrow, longitudinal, serrated ribs: radii absent: scutum with the adductor and articular ridges forming a cavity, which runs up to the apex of the valve.
Hab.—Cape of Good Hope; Algoa Bay; attached to sandstone and to Patellæ; Mus. Brit., Cuming, and Stutchbury.[108]
[108] I have seen three separate lots of this species all from the Cape of Good Hope; one lot was collected by Dr. Krauss, at Algoa Bay, and I strongly suspect is the species described by him in his ‘Südafrikanischen Mollusken’ as Conia porosa. If the species, figured by Chemnitz, and mentioned in a note (p. 329), under T. porosa, be the present species, the specimens probably did not come from Tranquebar, on which point Chemnitz speaks only from memory. I have seen one specimen ticketed New South Wales, it is possible, considering the case of T. rosea, that this may be correct, but I should like to have further confirmation before giving it as a habitat.
General Appearance.—Colour dark greenish gray; form steeply conical; surface covered, especially in the lower half of the shell, by numerous, narrow, sharp, longitudinal ridges, but slightly prominent, and serrated or transversely divided into small teeth: when the shell has been much disintegrated, the upper part of the surface consists of the exposed, smooth, rather large, upfilled parietal tubes. I have seen no instance of the development of the radii; sometimes even the sutures are with great difficulty distinguishable, though I believe they always reach the outer surface; sometimes the sutures are wide from the disintegration of the edges of the compartments. Orifice rounded or oval.
Scuta.—The scuta and carinal half of the terga are blueish-green. In the scuta neither the articular ridge or furrow are much developed: the adductor ridge is prominent, and is united to the articular ridge, about half way up the latter, thus forming a rather large, triangular cavity, which runs up to the apex of the valve.
The Terga are beaked. The spur, measured across the upper part, is half as wide as the valve; it is bluntly pointed; it is placed quite close to the basi-scutal angle of the valve, so that there is no basal margin on that side; it curves towards the scutum, its extremity extending beyond the basi-scutal angle.
Structure of the Shell and Radii.—The parietal tubes are rather large, especially those adjoining the inner lamina of the walls. The shell is of singularly little specific gravity, which is due to the parietal tubes not being filled up with shelly matter nearly to so great an extent as in the other species; even in the uppermost part the tubes are not solidly filled up, only their external sides are thickly coated with greenish-black shell, which by corrosion becomes grayish. The radii, as stated, are not developed: the shell breaks with singular facility along the sutures, and the radii are then seen to be most feebly represented[Pg 335] by a few very small branching ridges. The alæ have their edges plainly crenated. The sheath is dark green, with the lower edge free.
The Mouth presents no particular characters. With regard to the Cirri, I am doubtful whether any confidence can be placed in the numbers of the segments being constant; but I may state that the second cirrus contained thirteen and sixteen segments in its two rami; the third cirrus only fourteen in both rami; and the sixth cirrus twenty-six segments in both rami. Whereas in every specimen of T. porosa, the longer ramus of the third cirrus contained more segments than either ramus of the second. About half the segments, namely, those in the middle of both rami of the third cirrus, are furnished with coarsely and doubly pectinated spines, like those in T. porosa.
Affinities.—Upon the whole, this species is more nearly allied to T. porosa than to any other. In the cavity formed by the union of the adductor and articular ridges, it is allied to T. cœrulescens. This species differs from all, in its little specific gravity, consequent on the parietal tubes being only slightly filled up, and in the peculiarly serrated, narrow, approximate ridges on the external surface of the walls. The character derived from the adductor ridge, just alluded to, is remarkable. In the shape of the terga, in the absence of radii, and in the structure of the body, this species approaches closely to T. porosa.
CONIA ROSEA.[109] Krauss (!). Die Südafrikanischen Mollusken, Tab. 6, fig. 28, 1848.
BALANUS CUMINGII. Chenu. Illust. Conch., Tab. 4, fig. 5.
[109] I am greatly indebted to Professor Krauss for having sent me, for examination, the unique specimen collected by himself in Algoa Bay. There can be no doubt of the identity of the African and Australian specimens. It is a singular circumstance that the same species should occur in these two distant places, and, as far as at present known, not in the intermediate, more tropical coasts.
Shell dirty white, tinged with pink; parietes formed by a single row of large tubes: radii generally narrow: tergum with the spur rather short and broad.
Hab.—New South Wales, Moreton Bay in lat. 27°, Port Jackson, and Twofold Bay; South Africa, Algoa Bay. Attached, in Australia, to littoral rocks and shells; often associated with T. purpurascens, Chthamalus antennatus, and Catophragmus polymerus; Mus. Brit., Cuming, Krauss, Darwin, Stutchbury.
General Appearance.—Shell steeply conical, often rather convex; dirty or brownish white, feebly tinted with pink; external surface generally much disintegrated, and having in the upper part a pillared appearance, owing to the exposure of the upfilled, large, square, parietal[Pg 336] tubes, and, in the lower part, a striated (and sometimes serrated) appearance, from the exposure of the parallel, approximate plates, with which the outer lamina of the shell is internally strengthened. In only a few young specimens, the whole outer lamina of the shell was well preserved; and in these the surface was very smooth, and even glossy, giving to the specimens a quite different aspect; even in partially corroded specimens, the lower part of the shell sometimes is quite smooth. Generally, the radii are developed; in most specimens they are narrow, but sometimes of moderate width; their summits are oblique, and their edges often notched or toothed. The recipient furrow, in each opposed compartment, is often almost as wide as the radius itself, and is equally notched. In some much corroded specimens there were no radii. Basal diameter of largest specimen, 1.1 of an inch.
Scuta, generally thick, sometimes very thick, with the external surface usually much corroded: articular furrow rather wide; articular ridge not very prominent; adductor ridge prominent. The rostral depressor muscle is attached in a small oblong pit, sometimes including little crests; and the lateral depressor muscle is attached to what may be described either as three or four parallel furrows or crests.
Terga, with the spur placed close to the basi-scutal angle, so that there is no basal margin on that side; spur short, with its lower end truncated and rounded; broad, even exceeding, when measured across the upper part, half the width of the valve. Articular furrow wide. Apex not beaked.
Structure of Walls and Radii.—This species differs from all the others of the genus in having only a single row (fig. 3 d) of parietal tubes; these are large, quadrangular, but elongated in the ray of the circle. They are not filled up, even at the very top of the shell, but they become thickly lined all round with compact shelly matter. When the surface of the shell is disintegrated, these upfilled tubes greatly affect, as already stated, the external appearance. The outer lamina near the basis is internally strengthened by longitudinal, sharp, approximate ridges or plates, which, also, often affect, after corrosion, the external appearance. The radii have their sutural edges formed by a set of narrow, branching ridges or septa; the ends of which, seen externally, often give a notched outline to this edge; the recipient furrows in the opposed compartments are deep, and their edges likewise are often notched: the interspaces between the branching ridges are filled up solidly. The alæ have their edges coarsely crenated. The lower edge of the sheath is not free.
The mouth and cirri present no particular characters: the third cirrus has both its rami elongated, with the terminal segment tapering. In the three posterior pairs of cirri, the tufts of little spines between the main pairs are rather large.
Affinities.—This species has no particular affinity with any other. The circumstance of there being only a single row of parietal tubes is not so important a difference as might at first be thought, inasmuch as in the other species, during their quite early youth, the walls are formed of only a single row of tubes or pores.
LEPAS PURPURASCENS.[110] Wood’s General Conchology, p. 55, Pl. 9, fig. 42, 1815.
BALANUS PLICATUS. Lamarck. Animaux sans Vertèbres, 1818.
------ ------ ET PUNCTURATUS. Chenu. Illust. Conch., Tab. 4, fig. 3 et 12.
CONIA DEPRESSA (!). J. E. Gray. Appendix, Dieffenbach’s Travels in New Zealand, 1843 (sine descript. vel figurâ).
[110] The descriptions given by Wood and Lamarck are fuller and more accurate than is usual in the case of Cirripedes, and I have no doubt regarding these two names. The Conia depressa of Dr. J. E. Gray is, as I know from having seen the original specimens, the young of this same species; the name is unaccompanied by any description or figure.
Shell depressed, pale purple or dirty white, with the surface longitudinally ribbed, or corroded and granulated: radii or even sutures none, or radii well developed and broad, with their summits parallel to the basis: basis membranous: scutum transversely elongated: tergum small, with the spur extremely short and rounded.
Hab.—Sydney, New South Wales; Flinder’s Lagoon, Sir C. Hardy’s Island, Barrier Reef; King George’s Sound, Western Australia; Van Diemen’s Land; New Zealand, adhering to Pollicipes spinosus; Mus. Brit., Cuming, Stutchbury, Darwin, &c. China (?) attached to Pollicipes mitella, Mus. Brit. and Stutchbury. Generally attached to tidal rocks, sometimes to shells. Very common.
General Appearance.—Shell generally much depressed, in a few cases rather steeply conical, in one single instance cylindrical, but not much elongated. Colour, when alive, pale, but fine purple; I presume, judging from some dried specimens, sometimes dirty white. The state of the surface varies remarkably: about half the specimens (fig. 1 a) which I have seen, had the outer lamina of shell quite removed, and the surface granulated, owing to the projecting and exposed tips of the upfilled parietal tubes; the radii are not developed, and often even there is no trace of the four sutures; the rather large orifice is somewhat rounded, and the two scuta, with their surfaces disintegrated, have their middle parts deeply indenting the terga. The shell, in the other and perhaps more common condition (fig. 1 b), has the outer lamina preserved, and is longitudinally ribbed with generally at least five or six ribs on each compartment: the radii are here very wide, and extend from tip to tip of the compartments, so that their summits are parallel to the basis; they are gene[Pg 338]rally covered by a brownish epidermis, thickly clothed with little spines; the orifice is neatly diamond-shaped; the apices of the opercular valves meet at a common point: these specimens are almost always smaller and younger than the granulated specimens. Altogether the specimens in the two opposite states have, in their external appearance, nothing in common, and no one, without careful examination, would ever suspect that they were specifically identical; this, however, was proved by the intermediate forms, and, in one instance, three of the compartments had their surfaces granulated, and were entirely destitute of the radii, whilst the fourth by some chance had been preserved from corrosion, was longitudinally ribbed, and had its epidermis-covered radius fully developed. The difference in the appearance of the opercular valves, in the two states, is simply owing to the degradation of their upper parts in the granulated specimens.
The basal diameter of the largest specimen was one inch, but the height only .35 of an inch.
Scuta, transversely elongated, so that the basal margin is nearly twice as long as the tergal margin: articular ridge very little prominent; articular furrow wide but shallow; adductor ridge very blunt, slightly prominent, sometimes almost absent, almost parallel to the basal margin: there are no distinct crests for the rostral or lateral depressor muscles, but some small irregular pits for the latter. In one young specimen, the lines of growth were crenated, showing a tendency in the valve to become longitudinally striated, as in the allied T. costata. In some young and immature specimens, the basal margin was deeply sinuous.
Terga, small in area, not above half that of the scuta: spur extremely short, broad, placed close to the basi-scutal angle of the valve, so that there is no basal margin on that side of the spur. The lower end and sides of the spur form one uniform curve. Articular ridge barely developed. Crests for the tergal depressores sharp and prominent.
Structure of the Shell and Radii.—The walls are very thick, and the parietal tubes small and numerous; there are sometimes from twelve to fifteen rows of tubes in the thickness of the wall. The tubes in their whole upper part are filled up solidly; and, as we have seen, are often exposed by disintegration. In very young specimens, of that size in which in T. porosa there would be only a single row of parietal tubes, there were here two or three rows. The development of the radii, as we have seen, is very capricious; the sutures even sometimes being lost. The radii, when developed, are broad, square on the summit, and covered by brownish hirsute epidermis: internally they are formed of tubes like those forming the parietes; in this respect differing from all the species except the following, T. costata. The tubes in the radii run obliquely down towards the basis; instead of in a transverse line, directly towards the opposite compartment, as might have been expected from the structure of the radii in Balanus. The alæ have their edges finely crenated. The sheath in all the specimens which I have observed is colourless; its lower edge is not free. The corium entering the parietal tubes, and lining the opercular valves, the mouth, and the anterior cirri, is generally of an extremely dark purple colour.
The Basis is entirely membranous, in which respect this species differs from all the others in the genus.
Mouth: all the trophi are unusually hairy or spinose. The labrum is deeply notched and apparently destitute of teeth on the crest. The palpi are club-shaped or enlarged at their extremities. The mandibles have the fourth tooth rudimentary.
In the Cirri, the second and third pairs are remarkably short and blunt. In one specimen the two rami of the first cirrus had respectively six and sixteen segments; those of the second, six and seven; those of the third, seven and seven; and those of the sixth cirrus twenty rather elongated segments, with a small tuft of spines between each main pair of spines.
Affinities.—This species differs from all, in its membranous basis, and in its transversely elongated scuta. It resembles T. costata, and differs from all the other species, in its radii (when developed) being square on the summit, and in being formed of tubes,—in the smallness and number of the parietal tubes,—in the absence of crests on the under side of the scuta for the rostral and lateral depressor muscles,—in the shortness and rounded form of the spur to the terga,—and, lastly, in the club-shaped palpi and small size or absence of the fourth tooth in the mandibles. T. purpurascens differs from T. costata in those points, namely, in its membranous basis and transversely elongated scuta, in which it differs from all the other species, and, moreover, in its scuta not being plainly striated longitudinally, in having more ribs on the external surface of the parietes of its shell, and in having only three pairs of main spines on the three posterior cirri.
Shell depressed, whitish, generally with ten very prominent longitudinal ribs: radii broad, with their summits parallel to the basis: basis calcareous: scutum externally striated longitudinally: tergum with the spur short and rounded.
Hab.—Philippine Archipelago, Mus. Cuming. Attached to various shells, within the tidal limit.
General Appearance.—Shell whitish, probably tinged, when alive, with reddish-purple; depressed; surface perfectly preserved, smooth, but having longitudinal very prominent ribs, almost invariably ten in number; namely, three on both the rostrum and carina, and two on the two lateral compartments, with ten corresponding projections round the basal margin. Orifice passing from rounded-trigonal to diamond-shaped. The radii are very broad and square at the summit, and extend from tip to tip of the compartments. Basal diameter of largest specimen under half an inch, generally from .3 to .4 of an inch.
Scuta, of the usual sub-triangular shape, and not transversely elongated, as in T. purpurascens. External surface striated longitudinally; in many specimens there is a medial depression, or a row of very small pits, such as occur on the scuta of Balanus trigonus and lævis. The adductor ridge is moderately developed, and runs nearly parallel to the occludent margin; there are no crests for the rostral and lateral depressor muscles.
Terga: these in area equal two thirds of the scuta: the spur is short and rounded, and placed as described under T. purpurascens; but the articular ridge seems to be more prominent than in that species.
Structure of the Shell and Radii.—The parietal tubes are small, and very numerous, as in T. purpurascens. The radii are wide, square on the summits, but not so conspicuously covered by hirsute epidermis as in that species. Internally, the tubes forming the radii are smaller, and run more transversely than in T. purpurascens, that is in the normal course, as in Balanus. The edges of the alæ are nearly or quite smooth. The Basis is as distinctly calcareous, as in the other species of the genus.
Mouth: the trophi are not so hairy as in T. purpurascens; the labrum seems destitute of teeth; the palpi are club-shaped at their ends; the mandibles have only three teeth. The second and third cirri are not so short and blunt relatively to the others as in T. purpurascens. In the posterior cirri, the elongated segments carry four main pairs of spines, between which there is no intermediate tuft of fine spines.
The Affinities of this species have been fully pointed out under the last and closely related species. In external appearance, T. costata can at first hardly be distinguished from those young and pale-coloured varieties of T. purpurascens, which have their external surface not corroded, and their radii well developed.
Shell white, generally tinged in the upper part with pink; surface irregular: parietal tubes very irregular: radii moderately wide, with their summits oblique: alæ with very thick crenated sutural edges: tergum with the spur not joined to the basi-scutal angle; spur with its extremity equably rounded.
Hab.—Philippine Archipelago; Barrier Reef, (Raine’s Islet), Australia; Mus. Cuming and Stutchbury. Attached to massive corals, to coral-rock, to a Tridacna, and to Tetraclita cœrulescens.
General Appearance.—Shell conical, moderately steep: white, generally with a tinge of pinkish-purple in the upper part, owing to the[Pg 341] exposure of the tips of the upfilled parietal tubes. The lower part of the surface is generally well preserved, and is formed by very irregular, branching, longitudinal, slightly prominent, ridges. Radii of moderate width, with their summits oblique. Orifice rather large, rounded trigonal.
Scuta, rather narrow, with the upper part acuminated: the external surface generally much disintegrated, and marked with some irregular blotches of dark red. Articular ridge not prominent; articular furrow rather deep: adductor ridge, distinct from the articular ridge, pretty well developed, as are the crests for the rostral and lateral depressor muscles. In young and well preserved scuta, there is an external, medial, hyaline band, corresponding with the hollow under the adductor ridge, and caused by the thinness of the valve along this line.
Terga: externally, the carinal half is longitudinally and very feebly striated. Internally, the articular furrow is very wide, but shallow, and of unusual length, owing to the preservation of the upper part of the valve; the articular ridge is not prominent. The spur does not actually join, as in all the foregoing species, the basi-scutal angle, but is separated from it by a short piece of basal margin; its two sides are more nearly parallel than is usual, and the end is regularly rounded. It is always rather narrow, though the width varies considerably (fig. 3 d, 3 e). It extends in the same straight line with the middle of the articular furrow. The terga, though not possessing any striking characters, differ considerably in appearance from those of the other species.
Structure of the Shell and Radii.—The parietal tubes are remarkable from their irregular shapes, and unequal sizes (fig. 3 b),—hardly two resembling each other. Sometimes a single elongated tube will reach across the whole thickness of the walls. The septa between the tubes are rather thick and rugged. The tubes are generally darkly coloured from the adhering corium; they are solidly upfilled, but only in the uppermost part, by dark chocolate-red shelly matter. The radii are formed by irregularly branching ridges, with the interspaces filled up solidly. The square edges of the alæ are much thicker than in any other species, and are furnished with transverse ridges, which are sometimes slightly branched. The inner lamina of the walls near the basis, in most of the specimens, is irregularly and longitudinally ribbed, the ribs being longitudinally striated. The sheath and the upper part of the inner lamina of the parietes are clouded with chocolate-red.
The animal’s Mouth and Cirri were ill-preserved; but I was able to make out that the labrum had some strong teeth, and that the mandibles were furnished with five teeth, a greater number than in any other species. The palpi had parallel sides as usual. In the sixth cirrus, the segments had four pairs of main spines, instead of the usual number of three.
Affinities.—This species does not appear to be particularly related to any other one: perhaps it is rather nearer to the two following than to the foregoing species. The irregular parietal tubes, thick-edged alæ, form of terga, five teeth to the mandibles, and four pairs of spines to the segments of the posterior cirri, are its chief characteristics.
LEPAS CŒRULESCENS. Spengler. Skrivter af Selskabet, 1 Bind., 1790.[111]
[111] The longitudinally folded walls, as described by Spengler, the blue colour, the habitat, namely, associated with B. tintinnabulum from the East Indies, and more especially the expression “Valvulæ operculi cardine dentato mobilis,” apparently referring to the highly prominent articular ridge of the scutum, leave little doubt on my mind that I have rightly named the present species.
Shell with the upper part tinged greenish-blue, longitudinally ribbed: radii moderately wide, with their summits oblique: scutum with a small adductor and extremely prominent articular ridge, united together and so forming a small sub-cylindrical cavity: tergum with the spur not joined to the basi-scutal angle.
Hab.—Philippine Archipelago, attached to Balanus tintinnabulum; attached to a ship’s bottom, and to Balanus tintinnabulum, both from the Pacific Ocean; attached to a massive coral, and associated with T. vitiata, and therefore from the tropical eastern seas; Mus. Brit., Cuming, Stutchbury.
General Appearance.—Shell conical, sometimes depressed; surface with rather broad, smooth, longitudinal ridges; whitish, with the upper part greenish-blue, sometimes very feebly tinted with pink; radii white, or mottled with blueish-green, or with pink. When the outer lamina of shell has been corroded, the upfilled parietal tubes, of a dull blueish-gray colour, are exposed. The radii are moderately wide, with their summits very oblique. In basal diameter, one specimen was 1.8, and another 1.5 of an inch.
Scuta, externally furrowed very slightly in a longitudinal direction, causing the lines of growth to become a little sinuous. The valve is strong and thick; and the epidermis, when preserved, is hirsute with spines. The articular ridge is extraordinarily prominent; it projects, as measured from the external surface of the valve, to an amount equalling half the width of the valve in its widest part. The adductor ridge is very short, and is united to the bottom of the articular ridge, thus forming a small, nearly cylindrical tube, which runs up to near the apex of the valve. The inflected occludent margin is broad and coarsely toothed. The crests for both depressor muscles are not very prominent. When the scuta and terga are articulated together, owing to the great projection of the articular ridge of the scutum, its upper part is separated from the tergum (fig. 4 b), by a remarkably wide and deep,[Pg 343] fissure-like hollow. The scutum, in some distorted specimens from a ship’s bottom, was narrow in proportion to its ordinary breadth.
Terga.—These are large. Externally, there is a broad, longitudinal depression, bounded on each side by a ridge. The carinal half is feebly striated longitudinally. A large upper portion (fig. 4 b) projects freely, and stands, when the two valves are articulated together, above the apex of the scutum, but does not form a beak as in T. porosa. Internally, the tergal margin is widely inflected; the articular furrow is very deep, but the articular ridge is not prominent. The spur is not very broad; it is separated from the basi-scutal angle by a small space of basal margin, which forms a straight line with the basal margin on the opposite side of the spur. The end of the spur is truncated, and is parallel to the basal margin.
Structure of the Shell and Radii.—The walls are thick: the parietal tubes are rather large and regular; they become solidly filled up in their upper parts. The sutural edges of the radii are formed by unusually narrow, sinuous ridges, sending off delicate denticuli on each side: the interspaces between these ridges are solidly filled up. The crenated edges of the alæ are rather thick. Neither the mouth nor the cirri offer any noticeable character; but I may observe, that, in the mandibles, the third and fourth teeth are close together; and that, in the three posterior cirri, the tufts of small spines between the main pairs are small.
Affinities.—This species is very distinct from the others, with the exception of the following T. radiata, to which it is in several respects allied. The under side of the scutum, with its great articular ridge, and the cylindrical tube formed by the union of the latter with the short adductor ridge, affords the most noticeable character.
CONIA RADIATA.[112] De Blainville. Dict. Sc. Nat., 1816-1830, Pl. 164, fig. 5, 5 a, (sine descript.)
---- LYONSII. G. B. Sowerby. Genera of Recent and Fossil Shells, Plate, 1823, (sine descript.)
[112] The synonymy of this species is complicated. De Blainville gives no description under the article Conia, published in 1818, or in the vol. published in 1822; but I believe, from the figures of the opercular valves, that I have correctly identified this species with his C. radiata. Mr. Sowerby gives no description of C. Lyonsii, or any figure of the opercular valves, but his drawing of the shell is much better than de Blainville’s, and I believe it is the same species. Whether de Blainville’s or Sowerby’s plate appeared first I cannot ascertain. In the second edition of Lamarck, Conia radiata of Blainville is given as a synonym to Balanus radiatus of that work; but this is quite erroneous. I may add that if de Blainville’s name does not apply to the present species, it must to T. cœrulescens, and as the latter is the older name it will be permanent. In this case, T. radiata might be allowed to stand as my own name, considering that Mr. Sowerby’s figure is imperfect and is not accompanied by any description. At first I thought that the present species might be the Lepas mitra of Spengler, (‘Skrivter af Naturhist. Selskabet,’ 1790, Tab. 6, fig. 5), but the parietes are not described as porose; and the folds on the walls are too broad; on the other hand, his description of the opercular valves makes me think this may be the same species.
Shell white, with numerous approximate longitudinal ribs: radii broad, with their summits slightly oblique, internally porose: tergum with the articular ridge extraordinarily prominent, with the spur not joined to the basi-scutal angle.
Hab.—West Indies, adhering to Balanus eburneus and to Lepas anserifera. New South Wales, adhering to Tetraclita porosa. Attached to Balanus tintinnabulum, on a ship’s bottom from Sumatra; not rarely attached to Balanus tintinnabulum on ships’ bottoms; Mus. Brit., Stutchbury, and Cuming.
General Appearance.—Shell white, rather steeply conical, with numerous, approximate, rather narrow, longitudinal, rounded ribs on the walls: in a specimen half an inch in diameter, there were from eight to twelve ribs on each compartment. The outer lamina of shell seems always well preserved. Orifice rounded, trigonal. Radii white, smooth, broad, with their summits only slightly oblique. I have seen one specimen 1.2 in basal diameter, but quarter of an inch is a common size, and very young specimens are unusually frequent in collections.
Scuta broad, externally not striated longitudinally. The articular ridge is prominent, and the furrow deep, but not in so great a degree as in T. cœrulescens. The adductor ridge is only slightly prominent; it extends upwards only a little way above the lower end of the articular ridge, and does not form with the latter a cavity. There are no crests for the rostral depressor muscle, but there is a little pit, formed by the folding over of the occludent margin.
Terga.—These valves, when articulated with the scuta (fig. 5 b), project above them to an extraordinary degree, and are separated from them by a deep, fissure-like hollow, caused by the remarkable prominence of the articular ridge of the terga. The upper part of the tergum is not beaked, and does not project freely much above the sack. The valve is large; externally there is a rounded longitudinal furrow. The tergal margin is broadly inflected. The articular furrow is deep, and the articular ridge far more prominent than in any other sessile cirripede, for it projects, as measured from the outside surface, more than half the width of the valve; and consequently the valve, when viewed vertically from above, almost appears as if formed by the union of three plates, viz., the articular ridge, and the outside surface on each side of the spur. The spur is of moderate width, with the corners rounded: it is placed near, but not close to the basi-scutal angle, so that there is on this side a small portion of basal margin,[Pg 345] forming nearly a straight line with the margin on the carinal side. In some young specimens, about the tenth of an inch in diameter, from the West Indies and from New South Wales, the spur (the position of which I found varied a little in some other specimens) was placed nearly in the middle of the valve, and very nearly at right angles to the basal margin; it is possible that these may be a distinct species, but without larger specimens to judge from, I think it more probable that this difference in the tergum is due to variation and youth.
Structure of Shell and Radii.—The parietal tubes are commonly elongated in the ray of the circle: the septa are rather thick, and strongly crenated at their basal edges. The inner lamina of the walls is strongly ribbed longitudinally. The broad radii have their sutural edges formed by ridges, with numerous and closely approximate denticuli: the interspaces between the main ridges are not soon filled up, and at the bottom, each interspace usually terminates in a pore or tube; so that the radii are not solid, as in most of the foregoing species, but porose. The alæ have their edges crenated.
Basis, calcareous, of unusual thickness; the inner, or upper surface, is striated from the centre in rays, corresponding with the ribs on the inner lamina of the walls. This striated or furrowed structure in the basis, shows a tendency to its becoming tubular or porose, as may be inferred from analogous cases in Balanus.
Animal’s body unknown.
Affinities.—This species is rather more closely allied to the last than to any other. There is a close analogy in the peculiar manner in which the scuta and terga are articulated together in the two species: in this species it is effected by the great development of the articular ridge of the tergum, and in T. cœrulescens by that of the scutum. The internally striated calcareous basis, and the internal tubular interspaces between the denticulated ridges of the radii, are peculiar characters. The white colour, the narrowly and closely ribbed parietes, and the broad radii, give this species an aspect, by which it can be easily recognised.
ELMINIUS. Leach. Zoological Journal, vol. 2, July, 1825.
Compartments four: parietes not porose. Basis membranous.
Distribution, Southern temperate seas.
General Appearance.—Shell conical, with a strong tendency in most of the species to become cylindrical: orifice[Pg 346] generally large. Walls either thin and smooth, or thick and plicated longitudinally. Colours various, pale purple, greenish, white, and, in E. plicatus, owing to the exposure of an intermediate lamina of shell, bright orange-yellow. Radii, either of considerable width, with their summits oblique and rounded, as in the first two species of the genus, or very narrow, as in the last two species. Elminius plicatus is the largest species, and is sometimes one inch in basal diameter. The outer surface of this latter species is occasionally much corroded.
Scuta: these are of the usual shape; in E. Kingii and modestus there is no adductor ridge and no crests for the depressor muscles; in E. plicatus and simplex, on the other hand, there is a well developed adductor ridge and crests for the lateral depressor muscles; in some individuals, also, of E. plicatus there are small crests for the rostral depressores.
The Terga are remarkable for their variability in all the species; in many specimens of E. Kingii and modestus the basal margin on the carinal side of the spur is deeply hollowed out. The width and acumination of the spur varies in all the species. In E. plicatus and simplex this valve is remarkably like that of Tetraclita porosa. In some specimens of E. Kingii the terga and scuta are firmly calcified together.
Structure of the Parietes and Radii.—As in Tetraclita, the two lateral compartments are necessarily broad. The parietes are never porose, but consist, in appearance, of a single layer of shell. In E. modestus the basal internal edges of the parietes are smooth, but in the other species they are striated longitudinally with short ridges, or sometimes with sub-cylindrical projections. In those specimens of E. plicatus, which have externally suffered much corrosion, the walls have been rendered extremely thick, by the inward production of these ridges or plates; and in this case the ridges are not confined to the basal edges, but extend upwards close to the sheath. The basal surfaces of the walls in these latter specimens resemble those of Chelonobia, but the walls in that genus have an internal lamina, which here is not the case. The radii are wide in E. Kingii, and of moderate width in E. modestus, with their summits oblique and smoothly rounded, and their sutural edges not in the least[Pg 347] crenated. In E. simplex they are extremely narrow, smooth-edged, and rounded: in E. plicatus they are narrow, and in this species alone the sutural edge is sinuous, and sends inwards short ridges or teeth. The alæ, in all the species except this last, are likewise smooth-edged. The lower edge of the sheath depends, more or less freely, in all the species, except in E. Kingii.
Basis, membranous in all four species. In E. modestus, the true basal membrane is extremely thin, and is divided into concentric slips: on its inner surface there are attached numerous cement-ducts, varying from 1/3000 to 1/2000 of an inch in diameter, repeatedly trifurcating, rarely forming hexagonal or quadrangular loops, and with the branches placed approximately parallel to each other. Beneath the true basal membrane there is a complicated layer of cement, in the form of a network, or of separate tubes, or in beads and patches. In E. Kingii, the basal membrane presented a wonderfully complicated appearance, in part due to the cement forming a mass of inosculating fibres; many of these fibres seemed to end in circular discs of cement.
Neither the Mouth or Cirri offer any noticeable generic characters, as distinct from Balanus and several other genera. The Branchiæ, in E. plicatus, are well developed and moderately plicated. In E. modestus they are small, not plicated, but with a rounded sinuous margin: in a specimen having a basal diameter of 25/100, the branchiæ in total length were only 4/100 of an inch. At the bottom of the sack I observed some inwardly pointed, tapering filaments, such as occur in Balanus. In this same species I measured the ova, which were unusually elongated, being 19/2000 in length; I may add, that the probosciformed penis was actually thrice the length of the animal’s body in some small but mature specimens (with ova), having a shell with a basal diameter of 16/100 of an inch.
Distribution and Habitats.—This genus is remarkable, inasmuch as it is not distributed over the whole globe: three of the species occur very commonly on the shores of New South Wales, Van Diemen’s Land, and New Zealand; not extending, as far as I can judge, much north of Sydney: the fourth species is confined to South America, ranging[Pg 348] from the Falkland Islands and Tierra del Fuego, as far north as Chiloe. Elminius, therefore, appears to be strictly a southern genus. Elminius Kingii and modestus represent each other on the American and Australian continents; so I believe E. plicatus, in New Zealand, represents E. simplex in New South Wales and Van Diemen’s Land. The species are all attached to tidal rocks and shells. E. Kingii is sometimes attached to floating wood. At the Falkland Islands, the last-mentioned species adhered to some rocks, in a running brook of fresh water, at most eighteen inches under high-water mark, so that for the greater part of each tide it was exposed to absolutely fresh water. At Sydney I found E. modestus adhering to oysters in a muddy lagoon, almost separated from the sea, and apparently very unfavorable for cirripedes.
Affinities.—This genus can be distinguished from Tetraclita only by the four compartments not being porose, and by the basis being always membranous; whereas, in Tetraclita purpurascens alone it is membranous. Elminius Kingii and modestus, on the one hand, are closely allied together, as are E. simplex and plicatus on the other hand. The last-named species, in the characters of its opercular valves and in its shell, comes nearest to Tetraclita. In T. rosea we have seen that there is only a single row of parietal tubes, and the outer lamina of the shell is strengthened (as, indeed, it is in most of the other species of the genus, and in Balanus) by small longitudinal plates or ridges, which are similar and homologous to those on the internal basal edges of the parietes in three of the species of Elminius; so that the difference in the structure of the parietes, in Tetraclita and Elminius, is small.
ELMINIUS KINGII. J. E. Gray. Zoological Miscellany, p. 13, 1831.
------ LEACHII. King and Broderip. Zoological Journal, vol. 5, 1832-1834, p. 334, and appendix to King and Fitzroy’s Voyages.
------ ------ G. B. Sowerby. Genera of Recent and Fossil Shells, Plate.
Shell smooth, gray or dirty white: radii broad, smooth-edged: scutum without an adductor ridge; tergum with the spur distinct from the basi-scutal angle: scutum and tergum sometimes calcified together.
Hab.—Tierra del Fuego, Falkland Islands, Chiloe. Attached to tidal rocks and sometimes to floating timber; Mus. Brit., Darwin, &c.
General Appearance.—Shell fragile, either steeply conical with a large orifice, or sub-cylindrical; surface smooth, grayish or white, with large portions covered by pale brown epidermis. Radii broad with their summits oblique, smooth, slightly arched, exhibiting a large surface of the alæ. The alæ usually have their summits much less oblique than those of the radii; the portion added during diametric growth is of a dead white colour. The growth ridges on the scuta are very little prominent, and are crossed by a very obscure band of blueish-gray. The largest specimen which I have seen was .8 in basal diameter, and the longest cylindrical variety .55 of an inch in height.
The Scuta are remarkable for not having any adductor ridge or crests for the depressor muscles; the articular ridge is prominent, but it is short, not extending down half the valve. I have mentioned under the genus, that in many specimens at the Falkland Islands the scuta and terga were calcified together.
The Terga are rather small: the basal margin on the carinal side of the spur is always hollowed out, but to a very variable degree, as may be seen in the three figures (6 c-6 e); this margin is generally dentated with one or two little points; and an inner lamina of shell sometimes depends beneath the outer lamina, to which the opercular membrane is attached, as may be seen in the figure (6 d) of the external surface of the valve. The crests for the depressor muscles are well developed. The tergal margin is broadly inflected, and the articular ridge prominent, making the articular furrow deep. The spur is rather narrow, and is either (6 d, 6 e) bluntly or sharply pointed. The basal margin on the scutal side of the spur, is hollowed out, but to a variable depth.
Structure of the Parietes and Radii.—The parietes are thin; at their internal basal edges they are finely striated in longitudinal lines. The radii are solid, with quite smooth edges; they are generally covered by the epidermis. The sutural edges of the alæ are likewise smooth, these are added to largely during the diametric growth; and their summits, as already stated, are much less oblique than the summits of the radii. The internal surface of the shell is smooth, and is tinted pale dull purple. The lower edge of the sheath can hardly be said to be free. The carinal margins of the compartments project a little inwards.
Mouth: the labrum is deeply notched, and supports five little teeth on each side; the palpi are thickly clothed with spines on their inner sides; the mandibles have five or only four teeth: the maxillæ are notched, and the outer maxillæ bilobed.
Cirri: the first pair has one ramus nearly twice as long as the other. The three posterior cirri are elongated, and each segment supports five or six pairs of long spines, with a few minute intermediate bristles.
Shell folded longitudinally, greenish or white: radii of moderate breadth, smooth edged: scutum without an adductor ridge: tergum narrow, with the spur confluent with the basi-scutal angle.
Hab.—New South Wales; Van Diemen’s Land; New Zealand; very commonly attached to littoral shells and rocks; associated with Balanus trigonus and vestitus; Mus. Brit., Cuming, Stutchbury, Darwin.
General Appearance.—Shell conical, generally rather steep, occasionally depressed: walls longitudinally folded, sometimes very deeply, sometimes only to a slight degree: colour dull greenish or white. Radii of moderate width, with their summits very oblique, smooth and slightly arched: alæ much exposed, with their summits less oblique than those of the radii: the portion added to the alæ during the diametric growth differs much in appearance from the other portion. The scuta have the growth ridges but little prominent; they are crossed by a faint longitudinal band of gray. The largest specimen out of the many which I have seen, was under .4 of an inch in basal diameter.
Scuta, destitute of an adductor ridge and of crests for the depressor muscles: the articular ridge is moderate; but the articular furrow is rather wide: the internal occludent margin is much thickened. The Terga are narrow and small; they are somewhat variable in shape, caused by the degree to which the basal margin is hollowed out (fig. 1 c-1 e), and likewise by the extent to which the upper end of the valve has been worn away. No spur is apparent, for it is confluent with the basi-scutal angle of the valve. The articular ridge is very prominent, and runs down to the basi-scutal angle; and as the valve in this part is extremely narrow, with the spur not developed, it here assumes a channelled structure. The basi-carinal corner of the valve is furnished with rather feeble crests for the depressor muscles, and in those varieties in which the basal margin is much hollowed out, this part is remarkably narrow.
Structure of the Parietes and Radii.—The internal basal edges of the parietes and the sutural edges of the radii and alæ, are all smooth. The lower edge of the sheath depends freely. In the green varieties the colour is most distinct on the internal surface of the shell. The four compartments separate very easily when the shell has been ill preserved in spirits, or after a very short immersion in caustic potash.
Mouth, as in E. Kingii, excepting that there are only three teeth on each side of the notch (which is deeper) on the labrum. The[Pg 351] cirri resemble those of E. Kingii; the segments in the sixth pair are equally elongated, and bear five or six pairs of spines.
Affinities.—This species is closely allied to its South American representative E. Kingii; the differences consist in its smaller size, often greenish colour, more folded walls, and narrower radii: the internal basal edges, also, of the parietes are here smooth, instead of being striated, as in E. Kingii. The terga present even more obvious differences, in their narrowness, channelled under surface, and in the absence of the spur, or more properly in its confluence with the basi-scutal angle of the valve.
ELMINIUS PLICATUS. J. E. Gray. Appendix to Dieffenbach’s Travels in New Zealand, p. 269, 1843.
Shell deeply folded longitudinally, corroded, coloured in parts orange: radii very narrow, with their edges sinuous, and slightly dentated: scutum having an adductor ridge.
Hab.—New Zealand; New South Wales(?). Attached to rocks, often coated by Chamæsipho columna; Mus. Brit. and Cuming.
General Appearance.—Shell tubulo-conical, or conical, rarely depressed; strong, rugged, coloured in parts bright orange; deeply plicated longitudinally, but with the upper parts corroded and smooth. Orifice large. The sutures are indistinct and almost obliterated; the radii, when most developed, are narrow. Some specimens have their whole surface deeply corroded; in which case they are finely striated longitudinally, or pitted, and are of a gray or brown colour. The largest specimens are one inch in basal diameter, but one depressed specimen was 1.3 in diameter; another was rather under one inch in diameter, and one inch in height.
Scuta; beginning with the common tubulo-conical and not much corroded specimens, the valve (fig. 2 c) is moderately elongated, but in a rather variable degree. A prominent adductor ridge runs, from a little above a middle point of the basal margin, along the slightly prominent articular ridge: the articular furrow is moderately wide. There are distinct crests for the lateral depressores. In the conical, corroded specimens, the scuta (fig. 2 e) are considerably broader, with the articular ridge much more prominent, and the furrow wider: in one such specimen, there were crests for the rostral depressor muscle.
The Terga, in the commoner variety, resemble those of Tetraclita porosa; the spur adjoins the basi-scutal angle of the valve: the articular ridge is moderately prominent, and the furrow moderately deep. The valve is beaked, with an unusually large internal tube for the thread of corium: the beak, however, is often worn away. In the depressed much corroded specimens, the terga (fig. 2 f), like the scuta,[Pg 352] are broader and shorter than in the commoner variety; and the spur more especially is broader. The scutal margin is much more widely inflected, and the articular ridge much more prominent; consequently the articular furrow is much deeper.
Structure of the parietes and radii.—The orange or yolk-of-egg colour, which is so conspicuous a character in the present species, is due to a layer of shell between the inner and outer lamina, and is exposed only by the corrosion of the latter. Hence the very base of the shell is not of this colour; nor are the uppermost and still more deeply corroded portions, for here the orange-coloured layer has been removed. The sheath is orange-coloured, and the operculum, to a certain extent, is similarly tinted. The epidermis on the parietes, where preserved quite close to the basis, supports remarkably strong spines, about 1/100th of an inch in length. The basal internal edges of the walls are rather coarsely striated with irregular short ridges and sub-cylindrical points; and the walls in most of the specimens are regularly and deeply folded, which, with the little ridges, gives the appearance represented in fig. 2 b, Pl. 12. I have stated, under the Genus, that in the corroded and depressed specimens, the walls are rendered extremely thick by the inward production and upward extension of these same ridges and points; the under surface of the shell acquiring almost the appearance of Chelonobia caretta. The Radii are often not developed, even the sutures being obscure; when most developed, they are narrow, with the outer lamina along the growing edge sinuous, giving to the sutures a crenated appearance. The sinuosities on the growing edge generally send inwards short ridges or septa, like those on the sutural edges of the radii in most Balanidæ, but of which there is no trace in the other species of Elminius. In very minute, colourless specimens, about the 1/20th of an inch in diameter, the radii are quite smooth-edged. The alæ have their edges strongly crenated. The lower edge of the sheath depends freely.
Mouth: the labrum shows some tendency to be bullate; the notch is broad and shallow: the palpi have a thick brush of bristles on their inner sides. The mandibles have four or five teeth. In the maxillæ, the upper spines above the broad notch, are very strong. In the outer maxillæ, the two lobes are widely separated.
Cirri: in the first pair, one ramus is about one fifth longer than the other. In the third pair, the posterior ramus is one fourth longer than the anterior ramus, and its terminal segments are tapering, each having a single circle of bristles: the other segments, and those of the shorter ramus, support many coarsely pectinated spines. In the sixth cirrus, the segments are protuberant in front, and carry four pairs of stout spines, with a tuft of fine bristles between them.
Affinities.—This species differs considerably from the first two of the genus. In several characters it approaches nearer than the other species to Tetraclita, especially to T. porosa;—namely, in the scutum having an adductor ridge and crests for the lateral depressores, in the whole form of the tergum, in the thick walls liable to much corrosion, in the narrow radii, and in their edges, as well as those of the alæ, being crenated; and, lastly, in the character of the cirri, more especially of the third pair, with its coarsely pectinated spines. It also approaches, in all its characters, Balanus imperator and flosculus.
Shell ribbed longitudinally, dirty white; radii extremely narrow, smooth-edged; scutum having an adductor ridge.
Hab.—New South Wales (Sydney and Twofold Bay); Van Diemen’s Land; tidal rocks, often attached to other Cirripedes, and associated with Balanus nigrescens, Tetraclita purpurascens, Catophragmus polymerus; Mus. Brit., Cuming, Stutchbury, and Darwin.
This species, of which I have seen specimens from the above three localities, all exactly agreeing with each other, is perhaps the Australian representative of E. plicatus, which seems to be confined to New Zealand.[113] In all essential points it comes so near that species, that I shall make the greater part of my description comparative.
[113] I am bound to state that I have seen two specimens of E. plicatus marked Sydney, and one marked Moreton Bay, but in both cases the collectors had visited New Zealand, so that a wrong habitat by mistake might easily have got attached to the specimens in question.
General Appearance.—In external appearance there is considerable difference from E. plicatus, for E. simplex is generally of a regular conical shape, of a dirty-white colour, with the surface well preserved, having moderately wide, not very prominent longitudinal ribs. The orifice is rather small and pentagonal. The radii are extremely narrow or linear, with quite smooth edges; the sutures, however, are always very distinct, and in the upper part, the alæ are generally rather widely exposed, as viewed from the outside. The largest specimen which I have seen was .7 of an inch in basal diameter.
The opercular valves are closely similar to those of E. plicatus, but the scutum is generally a little more elongated, and the articular furrow not so deep: in accordance with this last fact, the articular ridge in the tergum is not so prominent as in E. plicatus; but we have seen that these several characters are highly variable in E. plicatus. The slope of the basal margin of the tergum towards the spur varies in the present species, in a strictly analogous manner, as it does in Tetraclita porosa.
Structure of the Parietes and Radii.—The parietes are not so thick as in E. plicatus; internally they are tinted pale purple; when broken transversely, a row of microscopically minute orange-coloured dots can generally be distinguished between the outer and inner laminæ; and these evidently represent the orange-coloured layer in E. plicatus. The sheath also exhibits a faint tinge of orange. The radii are very[Pg 354] narrow, and are quite smooth-edged, differently from in E. plicatus. The edges of the alæ barely exhibit a trace of being crenated.
In the body I could perceive no difference from E. plicatus, excepting that in the third pair of cirri the two rami are like each other, and do not support any coarsely pectinated, only serrated, spines; but after what we have seen on the variability of these very same characters in Tetraclita porosa, I dare not trust to them. The three posterior pairs of cirri, also, seem here to be more elongated in proportion to the others, than in E. plicatus.
Affinities.—It is certain that this species is most closely allied to E. plicatus; but as I have seen many specimens of the latter brought by different persons from New Zealand, and as I have observed in them no approach to the characters of E. simplex, which, in specimens from three localities, also appear to be constant, I have considered the two forms as specifically distinct. The present species differs from E. plicatus, in its white, conical, moderately ribbed, well preserved, smaller shell; and more especially in the orange-coloured intermediate lamina of E. plicatus being here represented only by microscopically minute dots. But the radii being smooth-edged, is the most important differential character, though in E. plicatus, during its earliest growth, whilst still immature and colourless, the radii are likewise smooth-edged.
PYRGOMA. Leach. Journal de Physique, tom. 85, 1817.
BOSCIA. Ferussac. Dict. Classique d’Hist. Naturelle, 1822.
SAVIGNIUM. Leach. Zoological Journal, vol. 2, July, 1825.
MEGATREMA. Ib. Ib.
ADNA. Ib. Ib.
DARACIA. J. E. Gray. Annals of Phil. (new series), August, 1825.
CREUSIA. De Blainville. Dict. Sc. Nat., Pl. 116, 1816-30.
NOBIA. G. B. Sowerby, junr. Conchological Manual,[114] 1839.
[114] The name, Nobia, is given in this work on the authority of Leach, but this must be a mistake, probably caused by some MS. name, (that fertile source of error in nomenclature), having been used.
Shell formed of a single piece; basis cup-formed, or sub-cylindrical, attached to corals.
Distribution, imbedded in corals, chiefly in the tropical seas, round the world.
I feel no hesitation in including the above several genera in one genus. In external appearance the P. monticulariæ (Pl. 13, fig. 5 a), which forms the genus Daracia of Gray, is the most distinct, but it is so intimately allied to the two, indeed to the three, foregoing species, that it cannot be separated from them. Of the first five species, P. grande, conjugatum, and cancellatum, form a graduated series, but with the steps very distinct, the chief difference being in the length of the spur of the tergum; for the fact of the scutum and tergum being calcified together in P. grande and conjugatum, and distinct in P. cancellatum, is certainly unimportant, as may be inferred from what we shall see in comparing together the last four species of the genus, and from what we shall see in Creusia spinulosa. The three above-mentioned graduated species are connected with the last four species of the genus, by several points of resemblance between P. grande and crenatum. The first two species, namely, P. Anglicum and Stokesii, are the most closely related together, and may indeed possibly be identical; these two, in all the characters derived from the opercular valves, resemble Balanus and other ordinary forms, and for this very reason they have some claims to be generically separated from the other species of Pyrgoma; for in these latter, the opercular valves seem to have broken loose from all law, presenting a greater diversity in character than do all the other species of Balaninæ and Chthamalinæ taken together.
General Appearance.—The shell consists of a single piece, generally without any suture, even on the internal surface; and this is the case, at least in P. Anglicum, in extremely young colourless specimens: nevertheless, in some specimens of this very species, and of P. conjugatum, there were traces of two, but only two, sutures on the sheath, one on each side towards its carinal end. The shell is much depressed or actually flat; and I have seen specimens even slightly concave; in P. Anglicum, however, the shell is steeply conical. The outline is generally oval; but in P. monticulariæ it is extremely irregular. The surface is generally furnished with more or less prominent ridges, radiating from the orifice, which is oval and small; sometimes, as in P. monticulariæ, excessively small. The[Pg 356] colour is white, or pinkish-purple. Most of the species are small, but I have seen specimens of P. grande three quarters of an inch in diameter in the longer axis, and, including the deep, almost tubular, basis, more than three inches in length or depth.
Opercular Valves.—In three species, viz., P. conjugatum, (Pl. 12, fig. 7 c), grande, and monticulariæ, the scuta and terga, on each side, are calcified together so perfectly, that there is no trace of a suture or line of junction: in P. milleporæ, these valves are generally slightly calcified together, but with the suture distinct. The Scuta differ so much in shape in the different species, that little can be said of them in common: in P. Anglicum and Stokesii they resemble those of Balanus; but in the other species they are much more elongated than is usual, and this is carried to an extreme in the last four species; this elongation is due to a great increase in breadth, as may be inferred from the position of the apex of the valve, and from the direction of the lines of growth. But the two most remarkable characters are, first,—the extraordinary development of the adductor ridge, so that, in P. conjugatum, cancellatum (Pl. 12, fig. 5 c), grande, and crenatum, it extends considerably beneath the basal margin, being produced, in the first two species, at the rostral angle, into a point; at the tergal end of the valve, the adductor ridge, when thus much developed, blends into the articular ridge. The second very remarkable character is the addition of a special ledge along the occludent margin of the scutum, and along the carinal margin of the tergum, which I will call the occludent ledges (limbus occludens), and which serve to close the orifice leading into the sack. The occludent ledge is small in P. grande, and is clothed with thick yellow spines, giving it a brush-like appearance: in P. crenatum and dentatum it is largely developed, the ledge on the scutum being articulated with that on the tergum, as shown in Pl. 13, fig. 4 a, 4 b,—the ledges being here and elsewhere marked by little bristly points. In P. monticulariæ, however, this ledge arrives at its maximum development (Pl. 13, fig. 5 f), the rest of the valve (the scuta and terga being here, as in P. grande, calcified together) being reduced to a mere basal edge or border. Excepting for the adductor[Pg 357] muscle, the depressions or crests for the other muscles, both on the scuta and terga, are hardly developed.
Terga: these, as in the case of the scuta, differ so much in shape in the several species, that little can be said of them in common. In P. Anglicum and Stokesii, they are of the normal shape; in P. cancellatum (Pl. 12, fig. 5 d) this, to a certain extent, is likewise the case, but the spur is produced to a quite extraordinary length. In P. grande (Pl. 13, fig. 1 b) there is no distinct spur, and the whole valve is square. In P. milleporæ (2 f) there is no spur, and the valve is arched and triangular. In P. crenatum the spur is broad, rounded, and depressed (fig. 4 b), with the carino-basal portion of the valve reduced to a mere border, barely distinguishable from the great occludent ledge. In P. monticulariæ there is no spur, and the whole valve forms a mere border to the occludent ledge; and, lastly, in P. dentatum, the valve is extremely variable in shape (fig. 3 c, 3 d, 3 f), and on its internal surface (fig. 3 g) there is an inwardly projecting, most singular and anomalous, tooth. Hence we see how wonderfully variable the terga are in this genus.
Structure of the Walls.—The shell consisting, as has been stated, of a single piece, is generally thick. From the close alliance between this genus and Creusia, it is probable that the shell, if examined immediately after the metamorphosis, would be found to show traces of being formed of four compartments. The walls are either solid or porose; their basal margin is formed by strong crenated ridges, answering to the longitudinal septa in Balanus; but these in P. monticulariæ are modified into a very irregular surface. The internal surface of the shell is generally smooth, or only slightly ribbed. The sheath has its lower edge free in P. Anglicum and Stokesii, and in a slight degree in P. milleporæ, but in the other species it is closely attached to the walls. In P. monticulariæ the sheath might easily be overlooked. In P. Anglicum, grande, and crenatum, it descends almost to the basal margin of the depressed shell, and as the opercular valves and membrane are attached to the lower edge of the sheath, the animal’s body necessarily comes to be almost exclusively lodged in the cup-formed basis. In P. grande and conjugatum, the lines of growth in the sheath are bent[Pg 358] upwards on each side, at points corresponding with the line of union between the scutum and tergum, in a manner I have not seen in any other cirripede; and this sometimes gives the appearance of two lateral sutures. I may here remark, that the manner of growth in Pyrgoma is almost the converse of that in Balanus, Tetraclita, and other allied genera; for in these latter, the basis increases in diameter, and the shell chiefly in height; whilst in Pyrgoma, the shell, from being so flat, increases almost exclusively in diameter, whereas it is the basal cup which is added to in height or depth.
Basis.—This in all the species is more or less regularly cup-formed or sub-cylindrical. In P. grande it penetrates the coral to a surprising depth; in P. monticulariæ it is irregular in outline, corresponding with the shell. The basis is generally almost wholly imbedded in the coral; but this is not the case with P. Anglicum, in which the basis is generally exserted, as it is in a slight degree in P. grande. The shelly layer forming the basis, in most of the species, is very thin, and is finely plicated owing to its edge folding between the ridges or septa that form the basal edges of the shell; this is very conspicuous in P. cancellatum. The basis is not permeated by pores, except in P. Anglicum. In some sessile cirripedes a cleft, covered only by membrane, may be observed all round between the lower edge of the shell and the basis; a cleft of this nature is rather conspicuous in P. crenatum, so that small portions of the septa on the internal surface of the walls can be seen from the outside. In P. monticulariæ, an analogous structure, developed to an extreme degree, presents a very different and unique appearance; the shell is nearly flat, and the smooth outer lamina does not nearly reach to the circumference, a wide border being thus left exposed, which is roughened (Pl. 13, fig. 5 a, 5 c, 5 d, 5 e) by the exposure of the irregular septa. I have not seen a fresh specimen, but there cannot be a doubt that this border is properly covered by membrane.
Animal’s Body.—From some cause, perhaps from the corals in which the species of Pyrgoma are imbedded, long remaining damp, the internal organs are generally badly preserved. I have received, in spirits of wine, only P. Anglicum, but I have examined dry specimens, in a tolerable[Pg 359] condition, of P. milleporæ and crenatum. As neither the mouth nor cirri, in these three species, offer any noticeable characters, distinct from those in Balanus or Acasta, my ignorance of these organs in the other six species is not important. In the above three species the labrum is deeply notched, with about three teeth on each side of the notch, except in P. milleporæ, in which the number is six. In all, the mandibles have five teeth, the two lower ones being small: the maxillæ are not notched: the outer maxillæ are bilobed. In the Cirri, the rami in the first pair are very unequal in length, the segments being slightly protuberant in the shorter ramus. On the segments of the posterior cirri there are four pairs of spines in P. Anglicum, and three pairs (of which the second and third are short) in P. milleporæ. At the dorsal basis of the penis there is a small straight projecting point.
Affinities.—The species (with the exception of the first two) are much more distinct from each other, and more easily determined than is usual with sessile cirripedes; it is, however, quite useless to attempt naming the species without disarticulating and cleaning the opercular valves. Although these valves differ so greatly in some of the species from those of Balanus and the allied forms, the genus itself, as a whole, does not differ much, except in the shell not being divided into compartments, and in the basis being cup-formed and not generally permeated by pores,—these latter characters being in common with Acasta. With respect to the absence of separate compartments, it should be remembered, that in the same species of Tetraclita we have individuals with the four compartments distinct and furnished with radii, and other individuals without any trace of a suture externally,—the outer lamina of shell (though not the inner) having become completely confluent all round. At the commencement of this description, when giving my reasons for uniting the several proposed genera into one genus, I gave a sketch of the affinities of the species: I have only to add, that the following sub-genus Creusia is closely, perhaps too closely, allied to Pyrgoma.
Geographical and Geological Distribution.—Most of the species are inhabitants of the hot coral-growing zones, in both the eastern and western hemispheres, but more especially,[Pg 360] as it would appear, in the East Indian Archipelago. From the habits of the corals, most of the species must be inhabitants of shallow water. Pyrgoma Anglicum, however, is an inhabitant of deep water on the southern shores of England, whereas at St. Jago, in the Cape de Verde Islands, I myself collected it adhering to a Caryophyllia, within the tidal limits. This same species existed on the shores of England during the Coralline Crag period; and at this epoch it attained a larger size than at present. Two species of the genus, according to Sismonda, are found in the tertiary beds of Piedmont. Mr. Stutchbury, who is so well acquainted with recent cirripedes, informs me that he has for many years examined fossil Secondary corals, in the expectation of finding imbedded, species of this genus or of some allied form, but without success. The same species of Pyrgoma is by no means always confined to the same coral: I have seen P. crenatum on four or five different corals, and P. Anglicum on at least three kinds: on the other hand, I have seen P. milleporæ only on the Millepora complanata (a member, as I believe, of the vegetable kingdom), and P. monticulariæ on a Monticularia from near Singapore.
PYRGOMA ANGLICA. G. B. Sowerby. (sine descript.) Genera of Recent and Fossil Shells, fig. 7, No. 18, Sept. 1823.
MEGATREMA (ADNA) ANGLICA. J. E. Gray. Annals of Philosoph. (new series), vol. x, Aug. 1825.
PYRGOMA SULCATUM. Philippi. Enumeratio Molluscorum Siciliæ, Tab. 12, fig. 24, (1836).
------ ANGLICA. Brown. Illustrations of Conchology, (2d edit., 1844), Tab. 53, fig. 27-29.
Shell steeply conical, purplish red: orifice oval, narrow: basis permeated by pores, generally exserted out of the coral: scutum and tergum sub-triangular.
Hab.—South coast of England and of Ireland, (12 to 45 fathoms, Forbes and MacAndrew); Sicily; Madeira; St. Jago, Cape de Verde Islands; generally[Pg 361] attached to the edge of the cup of a Caryophyllia, in deep water, but at St. Jago within the tidal limits; Mus. Brit., Cuming, Lowe, &c.
Fossil in the Coralline Crag, Ramsholt; Mus. S. Wood.
General Appearance.—Shell steeply conical, slightly compressed, the lower part with rounded, approximate, radiating ribs: colour dull purplish-red: orifice oval, small, and narrow. The basis is not deeply conical, and occasionally is even flat. Generally it stands exserted; but in the Coralline Crag specimens, it is almost wholly imbedded. Externally it is furnished with ribs corresponding with those on the shell. The largest recent specimen which I have seen, from St. Jago, was .22 of an inch in basal diameter; but some few of the British specimens are nearly as large, and one of the fossils from the Coralline Crag a little larger.
The Scuta and Terga are of the ordinary shape of these valves in Balanus and its allies. Scuta triangular, with the basal margin a little curved and protuberant: adductor and articular ridges distinct from each other, moderately prominent: there is a small hollow for the lateral depressor muscle. Terga triangular, with the spur rather narrow, moderately long, placed near, but not confluent with the basi-scutal angle of the valve. The basal margin forms an angle rather above a right angle with the spur. Internally, the articular ridge and crests for the depressor muscles, feebly developed.
Internal Structure of the Shell and Basis.—Internally, the shell is ribbed more or less prominently. The lower edge of the sheath, which is reddish, and extends far down the walls, seems always to project freely. In several specimens there were on each side, at the carinal end of the shell, a trace of a suture, which could be perceived only on the sheath. The basis appears always to be permeated by minute tubes or pores, though these are sometimes rather difficult to be seen.
MEGATREMA STOKESII. J. E. Gray. (sine descript. aut figurâ) Annals of Philosophy, (new series), vol. 10, Aug. 1825.
Shell moderately conical, pale-purplish red; orifice oval: basis not permeated by pores, deeply imbedded in the coral: scutum and tergum sub-triangular.
Hab.—Imbedded in the Mycedia (Agaricia) agaricites; therefore from the West Indies;[115] Brit. Mus. and Stutchbury.
[115] I am greatly indebted to Mr. Dana for having named for me the coral in which this species was imbedded, and informing me that it is a West Indian species.
This species comes so close to the last, that I am not sure that I have acted rightly in retaining it, but I think that it is distinct; and in this case, it is the representative, on the other side of the Atlantic, of P. Anglicum of our own side. It will be sufficient to point out the few points of difference. The shell is much more depressed, with the orifice oval, larger, and not so narrow. It is apparently of a paler red, and the radiating ribs perhaps not so prominent. The basis offers the most important difference, being deeply imbedded in the coral; and there is not the least appearance of the thin shelly layer, of which it is composed, being permeated by pores, as, we have seen, is always the case with P. Anglicum. As in this latter species, the sheath here depends freely. The opercular valves are closely similar; but in the scutum, the adductor ridge occupies a rather more central position; and in the tergum, the basal margin is more inclined towards, or forms a greater angle with, the spur: these differences, by themselves, I consider quite insufficient to characterise a species; but conjoined with the flatter shell, the larger orifice, the more deeply imbedded and non-porose basis, they may, I think, be admitted as specific. In dimensions, this species seems to attain a slightly larger size than P. Anglicum, for several specimens were .22 of an inch in diameter.
PYRGOMA CANCELLATUM. Leach (!). Encyclop. Brit., Supplement, vol. 3, Pl. 57, 1824.
------ LOBATA. J. E. Gray (!). Annals of Philosophy, (new series), vol. 10, 1825.
CREUSIA RAYONNANTE. De Blainville. Dict. Sc. Nat. (sine descript.), Pl. 116, fig. 7, a, b.
Shell with the circumference generally lobed: scutum elongated, with the adductor ridge descending far below the basal margin, and produced at the rostral end into a square point; tergum with the spur four times as long as the upper part of the valve.
Hab.—Imbedded in a Gemmipora, probably from the West Indies;[116] Mus. Brit. and Cuming.
[116] In this case I am again indebted to Mr. Dana for naming the coral; he informs me that the genus is found in the Pacific and East Indies; the specimen sent he believes is the G. cinerascens.
Appearance and Structure of Shell.—Shell nearly flat, sometimes tinted dull dirty purple, with the surface marked by slight, broad, approximate ridges, the ends of which form considerable projections, giving to the shell, when not too much encrusted by coral, a lobed border. In young specimens (.15 of an inch in diameter) some of the points projected half as much as the semi-diameter of the shell, giving it a radiating appearance. The orifice is oval and rather small, but of variable size. The shell is thick, and, near the outer lamina, is penetrated by pores: the internal surface is smooth. The sheath (fig. 5 b) extends down but a short distance from the orifice; it is closely attached to the walls: the lines of growth, at a point on each side, bend a little downwards (instead of upwards, as in the two following species), and hence the lower edge of the sheath irregularly projects downwards on each side. The basal cup, internally, is plicated, the hollows corresponding with the projecting, longitudinal, parietal septa, which form the lobed border of the shell. The largest specimen which I have seen, was rather above .4 of an inch in diameter.
The Scuta and Terga are not calcified together: they are both much elongated. Scutum (fig. 5 c, 5 e).—For the first time in the genus, or indeed in the family, this valve presents a remarkable character in the adductor ridge being immensely developed, so as to project far below the ordinary basal margin. At the rostral end, it at first appears to project even more than it really does, for the toothed occludent margin is in fact a prolongation of the true valve, as distinct from the adductor plate. Excluding this very narrow, prolonged, occludent margin, the adductor plate projects for a length equalling the rest of the valve. Along the tergal margin, the adductor plate is united to the articular ridge; and at the rostral end, it is produced into a square tooth, whence a square-edged ridge extends on the surface of the plate upwards to the ordinary basal margin of the valve. The exact shape of this adductor plate varies a little, as does the degree to which it is closely attached to the ordinary basal edge of the valve. The valve, as distinct from the adductor plate, is narrow, with the basal margin regularly curved. The articular ridge is very prominent.
Terga extremely narrow, linear, consisting in chief part of the spur, which is fully four fifths of the entire length. Externally (fig. 5 d) the valve is furrowed, with the edges more or less folded in along the spur. The upper or ordinary part of the valve is about one third wider than the spur. The basi-carinal angle is sharp, owing to the basal margin being a little hollowed out. A special plate of shell (fig. 5 f), hollow under its basal edge, runs from the carinal margin to the articular ridge, which latter is situated in the middle of the valve, and against[Pg 364] which the articular ridge of the scutum abuts. The spur is central; its end is bluntly pointed. The total length of the tergum rather exceeds that of the scutum, the produced adductor ridge being included in the latter.
Affinities.—Under P. grande I shall make a few remarks, showing that in several characters P. cancellatum and grande are at opposite ends of a short series, with P. conjugatum intermediate between them.
[117] Dr. Gray thinks this is the Pyrgoma stellata, of Chenu, (‘Illust. Conch.’); it may be so; but the figure given of the shell will do equally well or rather better for the Pyrgomum dentatum of this work, and for some varieties of P. crenatum. Without a careful description of the opercular valves, it is really impossible to recognise, with any approach to certainty, the species of this genus.
Shell nearly flat with approximate radiating ridges: scutum and tergum calcified together without any suture: scutum with the adductor ridge descending below the basal margin, and produced at the rostral end into a point: tergum with the spur about as large as the upper part of valve.
Hab.—Red Sea; Brit. Mus. and Cuming.
Appearance and Structure of Shell.—Shell white, or with a tinge of purple; nearly flat, with moderately prominent, narrow, approximate ridges, radiating from the orifice, which is oval, rather narrow, and not very small. In the largest specimens the ridges are less prominent than in the figure given. The walls are thick, and not at all porose: the sheath extends down almost to the base of the shell, and its lower edge is closely attached to the walls: on each side, towards the carina, there is a trace of a suture, and the lines of growth on the sheath are here a little upturned. The basis is deeply imbedded and internally furrowed; the calcareous layer forming it, is thin. The largest specimen was .4 of an inch in the longer diameter.
The Scuta and Terga (fig. 7 b, 7 c), in all the specimens which I have seen, are calcified together, with no trace of a suture. There is, however, a slight furrow, which, I believe, marks the normal line of separation between the two valves; and in the following description this is assumed to be the case. The Scutum has the adductor ridge greatly developed, so as to project below the ordinary basal margin to a distance as great as the height of the valve. At the rostral end, this adductor ridge or plate is produced into a point; and at the tergal end, it is blended with the articular ridge, and united to the inner face of the[Pg 365] tergum. That portion of the scutum which corresponds with the valve in ordinary cases, and alone is externally visible as long as the operculum is united by the opercular membrane to the sheath, is narrow, with the basal margin considerably hollowed out: the occludent edge is formed into thick teeth. The Tergum is elongated, rather exceeding in length the scutum, the latter being measured from the apex to the rostral projection of the adductor plate. The surface of the valve is depressed in the line of the spur, with the basal end of the latter bluntly pointed. A very slight flexure (fig. 7 c) on the basal margin indicates where we may believe the spur to commence, showing that it rather exceeds in length the whole upper part of the valve. The lines of growth obscurely indicate a tendency to the formation of a slight “occludent ledge” along the carinal margin. Traces are just visible of crests for the attachment of the tergal depressor muscles.
NOBIA GRANDIS. G. B. Sowerby, junr. (sine descript.) Conchological Manual, fig. 29, 1839.[118]
CREUSIA GRANDIS. Chenu. Illust. Conch., Tab. 1, fig. 2 a, sed non, fig. 2.
[118] It is quite possible that this may be the Balanus duploconus of Lamarck, but with such a character as the following, who can recognise a species? “B. testæ parte supremâ univalvi, indivisâ, convexâ: inferiore turbinatâ, non clausâ: aperturâ ellipticâ. L’exemplaire est sans opercule et incomplet.”
Shell moderately convex, nearly smooth: scutum and tergum calcified together without any suture: scutum furnished with a small occludent ledge, with the adductor ridge descending below the basal margin: tergum square without a spur.
Hab.—Singapore and East Indian Archipelago; Mus. Brit., Cuming, Stutchbury; imbedded in two kinds of coral.
Appearance and Structure of Shell and Basis.—The shell is conical, though to a variable degree, and sometimes is much depressed. The surface is smooth, with only traces of narrow approximate ridges. The colour is white, often with a tinge of dark purple. The orifice is oval, and moderately large. The shell and a small portion of the basis usually stand exserted above the coral. The walls are of variable thickness; when thick, the pores, by which they are permeated, are but[Pg 366] little apparent; sometimes there is more than a single row of pores. The points of the septa on the basal edge of the shell are small. The internal surface of the shell is smooth. The sheath is closely attached to the walls, and descends nearly to the basis; on each side its lines of growth are slightly upturned. The basis is deeply cup-formed or cylindrical, and in section oval like the shell; it penetrates the coral to a very remarkable depth,—in one instance to three inches. The shelly layer forming it, is thin, finely furrowed, and not permeated by pores. This is the largest species in the genus; one specimen was three quarters of an inch in its longer diameter, and above three inches in length.
The Scuta and Terga are calcified together, without any trace of a suture; the line of junction can be inferred only from the analogy of P. conjugatum, in which species the valves have a more normal character, and are separated by a slight furrow. It may be seen in the figure (Pl. 13, fig. 1 d) of the right and left hand opercular valves, viewed from vertically above in their proper relative positions, how abnormal their appearance is, which is partly caused by the spinose occludent ledges, presently to be described, but chiefly from the carinal margins of the two terga not being straight and parallel, as in all other cirripedes, and therefore not meeting each other, as is usual. In other genera, only the upper part of the carinal margin of the two terga can be opened for the exsertion of the cirri, the lower portion being united by membrane; but here, I have little doubt, from the position of attachment of the adductor muscle (fig. 1 c), so close to and almost on the terga, that the whole length of the carinal margin of the two terga is free or disunited for the protrusion of the cirri. This opening between the two terga evidently cannot be closed, but is probably filled up, and the animal thus protected, by the dorsal surfaces of the curled-up cirri; such, I believe, being likewise the case with some pedunculated cirripedes, as with Conchoderma. The scutum has a large adductor plate, which extends some little way (namely, about one quarter of the height of the valve), below the ordinary basal margin. This latter margin is slightly sinuous, and a little hollowed out towards the tergal corner of the valve. I believe that the ridge, which runs down to the basi-scutal corner of the tergum, though appearing to be part of the scutum, really belongs to the tergum. The adductor scutorum plate is not, as in the last two species, produced into a point at the rostral angle; at the tergal end it blends into the under surface of the tergum. The occludent margin is coarsely toothed. Rather on the under side of this margin and in the upper part, there is a narrow occludent ledge, which extends up beyond the apex of the valve, and thence runs a little way along the carinal margin of the tergum. This ledge is thickly clothed with strong, yellowish-brown spines, and hence appears like a brush. It is remarkable that the cavity for the adductor scutorum muscle is situated almost on the tergum.
The tergum is of large size, and nearly square; it is, in appearance, separated from the scutum by a ridge running up to the apex. The basal margin forms a right angle with the carinal margin, along which latter margin the lines of growth are upturned, and blend into the occludent ledge, which is common to the two valves. There cannot be[Pg 367] said to exist any spur, the whole basal margin being almost straight; nevertheless, on close examination, the ridge which in appearance separate the scutum and tergum, may, I think, be safely considered as one side of the spur (which, it should be remembered, has in all ordinary cases a longitudinal furrow or depression), and the other side of the spur is, apparently, very feebly indicated by a slight flexure in the middle of the basal margin. Hence, if the spur had been developed, it would probably have been half as wide as the valve. There are no crests for the tergal depressor muscles.
Affinities.—The present species, with the last two, form an interesting series. Pyrgoma grande and conjugatum, however, are more closely allied to each other than to P. cancellatum. In the scutum, the whole valve is least elongated, with the adductor plate least developed, in P. grande, and most elongated, with the adductor plate most developed, in P. cancellatum. In the outline of the tergum the range of shape is quite remarkable; in P. conjugatum, which stands between the other two species, the spur is rather long, whereas in P. grande there is no spur at all—a very unusual circumstance—and in P. cancellatum, at the other end of this short series, the spur attains a length wholly unparalleled in any other cirripede.
[119] From external aspect I suspect this species to be the Creusia madreporarum, Leach (?), as given in Chenu, ‘Illust. Conch.,’ Tab. 1, fig. 6. But I feel sure that Leach has nowhere published this name; and it may be observed that Chenu gives it with a mark of doubt. The shell in its imbedded state is only figured; the opercular valves are not given; and no descriptive details are added. Under these circumstances I have not adopted this name; I have, perhaps, been in some degree influenced by the fact that this species, judging from the many specimens examined by me, is never imbedded in madrepores, but exclusively in millepores, so that Chenu or Leach’s specific name of Madreporarum is singularly inappropriate.
Shell with the orifice narrowly ovate: sheath dark purple: Scutum much elongated: tergum triangular, convex, without a spur.
Hab.—Philippine Archipelago (Mindoro Island), Mus. Cuming. Mus. Brit., Stutchbury, &c. Imbedded in Millepora complanata, sometimes associated with Balanus Ajax.
Appearance and Structure of Shell.—Shell oval, flat, coloured pale dull purple, or white, with slight and narrow ridges, radiating, from the orifice, which is not quite central, but is placed rather nearer to the carinal than to the rostral end of the shell. The orifice is small and narrow; the carinal end being rounded, and the rostral end narrower and sharper,—this being the exact reverse of the usual shape of the[Pg 368] orifice in the Balanidæ. The walls are thick, and are formed of large square tubes. The internal surface of the shell is smooth. The sheath (fig. 2 b) is much more elongated than the shell, for at the rostral end it extends to the basal margin, and at the carinal end nearly to it, whereas on the two sides it is some way distant from the base. The orifice is considerably out of the centre of the sheath, being placed nearest to the carinal end. The sheath has its lower edge slightly prominent or free; the lines of growth are neither turned up nor down on the two sides, as in the last three species. When nearly full grown the sheath is coloured dark purple, but when young it is white, hence the upper part is white, surrounded by an oblong purple ring, and this is surrounded by the white shell. The basal cup is deep, and internally nearly smooth. The largest specimens were .3 of an inch in their longer diameter. Great masses of the Millepora are absolutely studded with this Pyrgoma, with usually more specimens on one side of the plate or branch than on the other. They stand in approximately parallel positions, the broad or carinal ends of the orifices pointing upwards.
Scuta.—The scuta and terga are closely united, and are often (perhaps always) in some slight, though variable degree, calcified together; and hence they often break, rather than separate, at the line of articulation. The external fissure or line of junction between them (fig. 2 c) is oblique to the longer axis of the scutum; in the uppermost part of the valve it is sometimes almost obliterated. The two valves together are nearly as long as the sheath, and consequently much longer than the orifice of the shell. The scutum is much elongated, being fully four times as broad as high. The valve narrows towards the rostral end, but in a variable degree: the basal margin is hollowed out a little (but to a variable amount), close to the basi-tergal corner. Along the occludent margin a slip of the valve, widening downwards, is a little bent inwards, and this inflected portion is separated from the rest of the surface by a slightly angular ridge, running from the apex to near the rostral angle. Internally, at the basi-tergal corner, a slight ridge, parallel and close to the basal margin (and which can be seen only when the basal edge of the valve is held upwards), represents the adductor ridge, which we have seen so largely developed in the last three species, and shall again see in P. crenatum. The articular ridge (fig. 2 d) is extremely prominent, consisting of a more or less rectangular shoulder.
Terga: these are rather small compared to the scuta: they are triangular and much arched: there is no trace of a spur. Internally (fig. 2 f), the articular ridge is central: there are some vestiges of crests for the depressor muscle.
Affinities.—This well-marked species, in the tendency of the opercular valves to be soldered together, and in the remarkable absence of a trace of a spur to the tergum, is allied to P. grande, but it is more closely allied to the three following species.
Scutum much elongated, with a tooth-like articular projection: tergum convex, irregularly triangular, sometimes with an imperfect spur, and on the internal surface with an inwardly projecting tooth; scutum and tergum furnished with an occludent ledge.
Var. (1), 3 c, 3 g: tergum, with a sharp internal tooth, projecting rectangularly inwards.
Var. (2), 3 d: tergum, with a broad blunt internal tooth, depending beneath the spur-like portion of the valve.
Var. (3), 3 f: tergum, with the basi-carinal end of the valve truncated, with a small blunt internal tooth projecting rectangularly inwards.
Hab.—Red Sea; Mus. Brit. and Cuming. Also associated with Pyrgoma crenatum, and attached to Meandrina spongiosa.[120]
[120] Mr. Dana informs me that he believes that this coral comes from the West Indies; though the specimens originally described by him had no label. If this be so, both Pyrgoma dentatum and crenatum have very wide ranges.
Appearance and Structure of Shell.—Shell nearly flat, oval, white or pink, with rather distant prominent ridges radiating from the moderately large (for the genus) oval orifice. The ridges are often obscured, and apparently sometimes almost obliterated by the encrusting coral. Shell permeated near the outer lamina by short imperfect pores: internal surface smooth: sheath inconspicuous, descending rather more than half way down the walls; lower edge closely attached to the walls. Basis deep. Diameter of largest specimen .3 of an inch.
Scuta: these are elongated, but to a very variable degree, some specimens being quite three times, and some barely twice as broad as high. I observed this same variability in two sets of specimens, differing, as we shall presently see, in the form of their terga: it depends in part, but not wholly, on the varying width of the occludent ledge, which is sometimes only a fourth, and sometimes half as high as the rest of the valve. This is the first species in the genus in which the occludent ledge—a structure peculiar to the genus—has been amply developed. The basal margin of the valve is slightly sinuous, and a very little hollowed out near the basi-tergal corner; it is also very slightly reflexed, the reflexed portion being separated from the upper part of the valve by a very slight depression or even furrow. I notice this slightly reflexed portion, simply as indicating a well marked feature in the basal margin of the following closely allied species. Internally (3 g), the adductor ridge is thick and slightly prominent, but does not descend[Pg 370] beneath the basal margin: it blends into the articular ridge, which here projects in a remarkable manner and degree (3 b, 3 e, 3 g), like a rounded tooth. This tooth is in part a development of the occludent ledge; it varies much in shape. The line of junction between the scutum and tergum is nearly straight, and nearly at right angles to their longer axes. In some specimens the scuta and terga are partially calcified together.
Terga: in three sets of specimens the terga differed considerably, but as in every other respect there was the closest resemblance, I do not doubt that these are merely varieties. In all three, the valve is rather small, irregularly sub-triangular in shape, and externally somewhat convex; in all three, there is an occludent ledge, of variable width as in the scuta; and in all three, there is an internal tooth-like projection, of variable form, unlike anything I have seen in any other cirripede. In the first variety (Pl. 13, fig. 3 c, 3 g), the basi-carinal corner of the valve is bluntly pointed, and a slight flexure separates this portion of the valve from the other and scutal half, which latter thus exhibits some tendency to be converted into a spur: on the internal surface (3 g) of this spur-like portion of the valve, there is a rather long, sharp tooth, which projects rectangularly inwards; it is flattened in a plane at right angles to the longer axis of the scutum and tergum together: it cannot be seen from the outside. In the second variety, the shape of the valve is not very different (Pl. 13, fig. 3d), excepting that the flexure, separating the basi-carinal corner of the valve from the spur, is deeper; but on the internal face of the spur, the tooth is far broader than in the first variety, and is flattened quite differently, viz., in a plane nearly parallel to the surface of the valve, and instead of projecting rectangularly inwards, it depends beneath the basal edge of the so-called spur, and can be seen from the outside. In the third variety (fig. 3 f), the whole carinal end of the valve is cut off, and there can hardly be said to be any trace of a spur, yet a slight furrow apparently marks the line of separation between the basi-carinal portion of the valve, here become very narrow, and the broad, irregular part, which would have formed the spur had such been developed: on the internal surface of the latter portion of the valve, there is a very small, blunt, slightly flattened tooth, projecting inwards, and more resembling that in the first than that in the second variety.
Affinities: under P. crenatum I will point out the diagnosis and relationship between this and that species.
PYRGOMA CRENATUM. G. B. Sowerby. Genera of Recent and Fossil Shells, (No. 218, Sept. 1823), fig. 1 to 6.
Scutum much elongated, with the adductor ridge descending below the reflexed basal margin: tergum with a broad[Pg 371] depressed spur: scutum and tergum furnished with a wide occludent ledge.
Hab.—Philippine Archipelago; Singapore; Mus. Brit., Cuming, Stutchbury; sometimes associated with Creusia spinulosa.
Appearance and Structure of Shell.—Shell not distinguishable from that of P. dentatum; nearly flat, oval, white, sometimes pale pink, with rather distant prominent ridges radiating from the moderately large oval orifice. Shell solid, or permeated near the outer lamina by short imperfect pores: internal surface smooth. The sheath descends nearly to the base of the walls; it is but little conspicuous, and its lower edge is closely attached to the inner surface of the shell. Basis deep. Diameter of largest specimen under .3 of an inch.
Scuta: in this species the scutum is more abnormal than in any other Cirripede in the whole family: this is owing both to the adductor ridge descending far beneath the basal margin, and to the great development of the occludent ledge; hence the middle and very narrow portion of the valve alone answers to the scutum, as seen in other genera. The whole valve, including the adductor ridge and the occludent ledge, is narrow, being more than twice as broad as high; but the proportional width varies, owing chiefly (as in the last species) to the varying width of the occludent ledge. This ledge commences a little way from the rostral point of the valve, and gradually widening, extends to the apex, where it is either as high or twice as high as the rest of the valve. It is articulated by a convex surface, and by a hollow on its under side with the occludent ledge of tergum. The scutum cannot be said to have any tergal margin; without, indeed, the articular surface of the occludent ledge be thus called. The basal margin is curved, and considerably reflexed, of which peculiarity we have seen a vestige in the last species: the reflexion is not well shown, owing to the foreshortening of the reflexed edge, in fig. 4 a: this reflexed edge not being shown, causes the lines of growth to appear as if they ran more transversely to the longer axis of the valve, than they really do; for they run nearly as in the scutum (fig. 3 e) of P. dentatum. The direction of these lines of growth is of importance, for they show that the elongation of the scuta is due to an inordinate increase in their breadth, as compared to the same valves in ordinary species. The adductor ridge, having a sinuous margin, runs from near the apex to near the rostral angle: it descends below the basal margin about as far as the height of the true valve, excluding the occludent ledge. Of course this adductor ridge or plate lies beneath the membrane connecting the opercular valves with the sheath, and is concealed by it, as long as these valves remain within the shell. The edge of the occludent ledge is straight, but the edge of the middle portion of the valve, that is of the true valve, is much bowed.
Terga (fig. 4 b): these are of so irregular a shape that they can hardly be described; they may, however, be said to consist of two portions joined together, of which the lower portion is a little elongated transversely, of somewhat variable shape, with part of its surface considerably[Pg 372] depressed (compared with the rest of the valve), sometimes being even concave: this concave portion apparently answers to the spur in other cirripedes. At the scutal corner of the valve there is a shoulder (perhaps answering to the inflected scutal margin in an ordinary tergum), which locks into a hollow on the under side of the occludent ledge of the scutum. The internal surface of the so-called spur is rounded and convex. The upper part of the tergum is in main part formed by a great occludent ledge; but this, on its lower side, is bordered by a narrow irregular slip, which, as shown by the lines of growth, represents the whole of the ordinary valve, excepting, of course, the spur already described. The occludent ledges of both valves support some fine spines.
Affinities.—Observing how extraordinarily the terga varied in P. dentatum, and that the shells were identical in that and the present species, it occurred to me at first that they might, perhaps, be both extreme varieties of one form: but in the scutum of P. crenatum, the invariably great development of the adductor plate,—the marked manner in which its basal margin is reflexed,—the absence of a tooth-like articular projection,—and again in the tergum of P. crenatum, the invariably large size of the concave spur, without any internal tooth, altogether convince me that the two species must be considered as distinct. This species is allied to P. grande, in the scutum of that species having an occludent ledge, though small, and a great adductor plate. I have only further to remark, that the figure of the opercular valves, given in Sowerby’s Genera of Recent and Fossil Shells, is so good, that there can not be the least doubt about the present identification.
PYRGOMA (DARACIA) MONTICULARIÆ. J. E. Gray (!). Zoological Miscellany, p. 6, 1831.
Shell of an irregular shape, with a roughened exterior border: orifice minute, circular: scutum and tergum both much elongated, calcified together without any suture, both furnished with a broad occludent ledge.
Hab.—Singapore; Mus. Brit., Cuming, and Stutchbury. Sometimes associated with Creusia spinulosa.
Appearance and Structure of Shell.—Shell dull white, very irregular in outline, sometimes rounded, more often unequally elongated, and frequently star-shaped,—the projections being quite irregular. Whole shell nearly flat, but with the central part saddle-backed, or formed into a more or less prominent ridge, extending in the line of the longitudinal axis of the animal’s body: the circumferential portions[Pg 373] of the shell not unfrequently are a little recurved upwards. Orifice extremely minute, circular. The outer lamina of shell, which is smooth, does not extend to the circumference, and consequently a rather broad, nearly equal border, which is rough, surrounds the whole shell. I have no doubt that, when the shell was alive, this border was covered by a membrane, which, in drying, has curled up and been lost, in the same manner as the strictly analogous but narrow open seam between the basal edges of the shell and the basis in some cirripedes (as in the last two species of Pyrgoma) is protected. The roughened border can sometimes be plainly seen to be formed of normal (Pl. 13, fig. 5 e) longitudinal septa having crenated edges, with shorter septa between the longer ones; but more often the septa are so irregular, and so much branched (5 d), that the whole resembles a mass of moss. Why the outer lamina of the shell in this one species does not nearly reach the circumference of the walls, I cannot conjecture. The extremely irregular, depressed shape of the shell, with the minute circular orifice, and the singular rough circumferential and often slightly reflexed border, together give to this species so peculiar an aspect, that until close examination I did not believe that it was a cirripede. The extreme irregularity of shape depends in great part upon the irregular growth of the Monticularia, in which it is imbedded.
Internally (fig. 5 b) the walls are smooth, but they are perforated by many quite irregular, small orifices, which have the appearance of having been formed by some boring animal, but really serve, as I believe, to admit threads of corium into certain irregular pores which penetrate the shell. The sheath descends but a very short distance from the orifice: it is closely attached to the walls, and might easily be overlooked. The basis is deep, of an irregular outline, like that of the shell, and is formed by a very thin shelly layer. The largest specimen which I have seen, measuring from the extreme projecting points, was .4 of an inch in diameter.
The Scuta and Terga (fig. 5 f) are calcified together without any trace of a suture; together they form a bow with the two ends curled rather abruptly inwards; they are both extremely narrow, but furnished with an occludent ledge, twice or thrice as high as the proper valves themselves. This occludent ledge, which is finely hirsute, begins at about one third of the length of the scutum from the rostral angle, and runs to near the basi-carinal angle of the tergum. The scutum itself is curved, with a slip, along the true occludent margin (best seen at the rostral end), lying in a different plane from the rest of the valve, much in the same way as in the scutum of P. milleporæ. The adductor ridge descends a very little below the basal margin of the valve, and extends for nearly its entire length: this adductor ridge makes the proper valve even narrower than it at first appears. The Tergum is extremely narrow, forming merely a border to the occludent ledge; but it is not short, being about two thirds of the length of the scutum. There is no trace of a spur; indeed, the valve is rather narrower where the spur should have stood, than it is at the basi-carinal end. The scuta and terga are calcified together by their apices.
Affinities.—Although this species, as above stated, differs so remark[Pg 374]ably in external appearance from the other species of the genus, and, indeed, of the whole family, yet the shell in no one essential point of structure materially differs from its congeners; and if we compare the opercular valves with those of the three last species, we shall be struck with their close, yet graduated, affinity: in P. milleporæ the scuta and terga tend to become calcified together, and the rostro-occludent end of the scutum is bent into a different plane from the rest of the valve. In P. dentatum we have an occludent ledge exactly as in the present species; but in that species the adductor plate is less developed than in P. monticulariæ; on the other hand, in P. crenatum, the adductor plate is more developed than in P. monticulariæ. If in P. crenatum we were to remove the spur from the tergum (and it is much less developed in P. dentatum; and in P. milleporæ it is entirely absent) this valve would be almost identical with that of P. monticulariæ. Under these circumstances I consider it impossible to separate the present species as a distinct genus.
The Daracia Linnæi of J. E. Gray (Annals of Philosophy, new series, vol. 10, 1825), was published without description or figure.
The Megatrema semicostata of G. B. Sowerby, junr. (Conch. Manual, fig. 33, 1839), is not described, and is very indifferently figured without the opercular valves, and therefore can never be recognised.
There is an admirable figure of a Pyrgoma, without any specific name, in the great ‘Description d’Egypte,’ but from the want of some details, I cannot positively recognise the species; I am inclined to believe that it is the P. dentatum of this work.
M. Chenu, in his grand Illustrations Conchyliologiques, has given most beautiful figures of several species of Pyrgoma, and of Creusia, but unfortunately, from the opercular valves not having been figured, I find it impossible to recognise them. The new species are Pyrgoma stellata (on which I have appended a note under P. conjugatum), and P. spongiarum and P. corymbosa of Valenciennes.
CREUSIA. Leach. Journal de Physique, tom. 85, July, 1817.
Compartments four, furnished with radii; basis cup-formed: attached to corals.
Distribution, imbedded in corals throughout the tropical seas.
Creusia is closely allied to Pyrgoma; and had not this genus already been adopted by several authors, I should not, I think, myself have formed it; though, no doubt, it harmonises well with some of the other genera of the family, which are perhaps all too intimately related. Creusia differs from Pyrgoma only in the shell being separated by sutures, into four compartments, with well developed radii: in other respects, such as in general habit, in the cup-formed imbedded basis (not permeated by pores), in the opercular valves, in the characters derived from the mouth and cirri, there are no generic differences. This affinity is more particularly evident when Creusia is compared with the first few species of Pyrgoma: indeed, for a short time, I was inclined to consider var. 10 of Creusia as identical with Pyrgoma conjugatum. With respect to the species of Creusia, I confess I have been much perplexed in determining whether there be only one, or half a dozen. The latter conclusion would almost certainly be arrived at if only a few specimens were examined; and it might naturally be thought that some of the species were extremely well marked; the difficulty of drawing any line between varieties and species, begins only when some hundreds of specimens, from various parts of the world, are disarticulated, cleaned, and carefully examined. Creusia, in this respect, offers a striking contrast with Pyrgoma, in which nearly all the species are strongly characterised. The shell differs very little in the several varieties or species of Creusia; and the most marked difference, namely, whether the walls are permeated by irregular pores or not, seems certainly quite[Pg 376] variable. It is in the opercular valves, which in other genera offer by far the most reliable character, that we encounter the chief cause of perplexity; for the characters thus derived, though at first appearing very distinct, blend into each other, and are not accompanied by any well marked differences in the shell. Only a few of my specimens have any habitat; but the geographical range, as far as it does go, throws no light on the question which forms to regard as species and which as varieties. As is generally the case with cirripedes, the variations are local, so that the greater number of specimens imbedded in the same coral resemble each other. Under these circumstances I have thought it best, after repeated examinations of a very large suite of specimens, to describe separately each variety, without attaching any name to it; but I will first make a few general remarks on the structure of the shell. If I do not thus throw much light on the subject, I shall at least not burden it with error. I believe that the species will be definitely made out only by persons resident in the coral-bearing zones. I have given copious illustrations of the opercular valves; for, if my view be correct, this genus offers a curious and striking case of variation; if, on the other hand, I am wrong, the drawings, I hope, will aid others in coming to a more correct conclusion.
CREUSIA SPINULOSA. Leach (!). Encyclop. Brit. Suppl., vol. 3, Pl. 57, 1824.
CREUSIA SPINULEUSE. De Blainville. Dict. Sc. Nat., Pl. 116, fig. 6.
CREUSIA GREGARIA. G. B. Sowerby (!). Genera of Recent and Fossil Shells, No. 18, Sept. 1823.
---- GRANDIS. Chenu. Illust. Conch. Tab. 1, fig. 2, sed non fig. 2 a and b.
Hab.—Philippine Archipelago, China, Singapore, Java, Red Sea, West Indies; imbedded in various corals; Mus. Brit., Cuming, Stutchbury, Dunker, &c.
General Appearance.—The shell is oval, generally flat, sometimes conical, with narrow and approximate ridges radiating from the orifice[Pg 377] (fig. 6 a). The ridges, however, are sometimes distant from each other, and considerably prominent, projecting round the basal border. The orifice is either neatly diamond-shaped or oval. The four compartments are quite distinct; the radii are generally white, of considerable width, and with their summits not oblique. The colour is either white, or pale pinkish-purple; but in var. 11, bright pink. Even in the white specimens, when well preserved, the sheath is generally, but not always, either pale or dark purple. The largest specimen which I have seen, from the West Indies, was above half an inch in diameter; but from .3 to .4 of an inch is the more usual full size. I believe that the size, as well as the great variability of the present species, is partly determined by the rate of growth of the various zoophytes in which the specimens are imbedded, for the shell has to keep on nearly a level with the surface of the coral.
Structure of Shell and Basis.—The walls are internally ribbed; the ribs being usually prominent, sometimes to such a degree as to deserve to be called plates. The outer lamina is of variable thickness, and the prominence of the internal ribs appears in considerable part to depend on the extent to which the outer lamina has been thickened from within. In many specimens, instead of the interspaces between these internal ribs or longitudinal septa being solidly filled up, separate and successive laminæ have been deposited, by which the shell is rendered cancellated or porose; but the pores are very irregular; and sometimes they form two or three irregular rows one behind the other: this structure seems eminently variable. The edges of the radii are formed by crenated, and occasionally branched, septa. That part of the alæ, which is added during the diametric growth of the shell, is very thin. The lower edge of the sheath seems always to be free. The shelly layer, forming the basis, which is deeply cup-formed, is thin, more or less finely furrowed in radiating lines, and not permeated by pores.
The opercular valves will be best described under the following eleven varieties.
Var. (1), Pl. 13, fig. 6 a, 6 b, 6 c, 6 d.—Hab. Java, and probably several other districts.—I will first describe a typical sub-variety, by which I mean a sub-variety not presenting any extreme character. The scutum is of a sub-triangular shape, with the basi-tergal corner much rounded off (6 d), and generally but not always hollowed out in a rather remarkable manner. The adductor ridge is considerably prominent, and extends high up, parallel to the articular ridge, which latter is rounded and prominent, but to a variable degree. Near the rostral angle there is sometimes a small tooth, or only a trace of one, depending beneath the basal margin; this tooth we shall hereafter see much more developed. The Tergum is about two thirds of the width of the scutum. It is often slightly beaked, but this is more conspicuous in some of the following varieties. The spur is about half the width of the valve, and its basal end is truncated, and nearly parallel to the basal margin of the valve, but the truncated form passes insensibly into a rounded outline. The shell in this variety is generally thick and is not permeated by pores; the orifice is diamond-shaped. But in another sub-variety the walls of the shell are always, or nearly always, permeated by pores, and the[Pg 378] tergum is very much narrower, with the spur sharper, so that at first I concluded that these two sub-varieties were specifically distinct: we shall, however, soon see in var. 2 and in var. 4, that no confidence whatever can be placed in the exact breadth of the tergum, or in the porosity of the walls; hence I have been driven to consider the two varieties just mentioned as merely sub-varieties.
Var. (2), fig. 6 e, 6 f, 6 g.—Hab. China; Red Sea.—The shell is almost invariably permeated by pores, sometimes arranged in two or three very irregular rows. In some specimens the scutum exactly resembles that in var. 1, but with the tooth near the rostral angle often rather larger: in other specimens the scutum is much more elongated transversely (fig. 6 e), with the adductor ridge more medial, and the basal margin not at all hollowed out at the basi-tergal corner of the valve. The tergum, here, is the remarkable feature, being sometimes excessively narrow, with a long sharp spur, which often, but not always, terminates in a needle-like point. In other specimens, from the same coral and certainly belonging to this same variety, the valve is not so narrow (6 g), and the spur not so pointed; consequently (as in several analogous cases in other cirripedes) it is impossible to draw any line of distinction between the specimens with the narrow and broad terga.
Var. (3), fig. 6 h [Creusia gregaria of G. B. Sowerby!]—Hab. Unknown.—The scutum presents here exactly the same considerable range of variation as in var. 2. The tergum is broad, as in var. 1, but the spur is rounded, and from not being placed so immediately close to the basi-scutal angle of the valve, gives to it a rather different aspect. The breadth of the spur varies; an extreme variety is given in fig. 6 h.
Var. (4), Pl. 14, fig. 6 i, 6 k, 6 l.—Hab. Philippine Archipelago; West Indies.—The scutum here presents the same sub-varieties as heretofore, excepting that I have not seen any so much elongated transversely. The shell is covered either with slight, closely approximate ribs, as in the foregoing varieties, or with more distant and more prominent ribs. In specimens taken out of the same branch of coral the walls were either porose or solid. Sometimes the sheath is bright purplish-pink. It is the tergum, again, which presents a remarkable range of difference; for the longitudinal depression or furrow which in the former varieties was quite open, here has its edges more or less folded inwards, and is sometimes quite closed. This same variation has been commonly observed in many species of Balanus, in which it appears to be dependent on the age of the individual; but this does not appear to be the case in the present genus. As a consequence of the greater or less folding in of the two sides of the furrow, the spur is rendered more or less narrow and pointed, and thus becomes removed to a greater or less distance from the basi-scutal angle of the valve. Further, as a consequence of this folding in, the internal surface of the valve along the line of the external furrow, is raised into a longitudinal ridge. The length of the spur varies considerably. In some very young individuals, the basal margin descends lower on the scutal than on the carinal side of the spur. In one set of specimens (fig. 6 l), a plate extended from the carinal margin to near the central longitudinal ridge just mentioned: a similar structure was described under Pyrgoma cancellatum.
Var. (5), Pl. 14, fig. 6 m.—Hab. Unknown.—We have seen in vars. 2 and 3 that the scutum varies considerably in shape: here it is unusually narrow, with the adductor ridge almost touching the articular ridge. There is no little tooth near the rostral angle, and the basi-scutal corner is not hollowed out. The tergum also varies; in some individuals it is truncated and like that figured of var. 1, but rather more rounded; in other specimens (from the same branch of coral) the basal margin so blends into the spur that the latter can hardly be discriminated (fig. 6 m); in other respects the outline resembles pretty closely that of one of the sub-vars. (fig. 6 f) of var. 2. The shell is not porose; it is thick, with strong internal ribs, and resembles that of var. 1; but it is of a pale purplish colour.
Var. (6), an. spec.? fig. 6 n-6 q.—Hab. Philippine Archipelago.—This is a very remarkable variety; we have, imbedded in the same coral, and with shells absolutely identical, specimens with the scutum having three distinct but graduated forms. Firstly, a scutum transversely elongated, in all external respects like some of the varieties mentioned under vars. 2 and 3, with no rostral tooth, and not hollowed out at the basi-tergal corner, but with the adductor ridge more prominent. Secondly, a scutum of the same general shape, but with the adductor ridge so much developed (fig. 6 n) as to descend slightly beneath the basal margin, and to be seen when the valve is viewed externally; there is a very slight tooth near the rostral angle (as in some former sub-varieties), and which can be here rather more clearly seen than hitherto, to be formed by the adductor ridge (closely united to the external surface of the valve) extending thus far, and being here produced a little downwards. Thirdly (6 p), we have the adductor ridge immensely developed, descending far below the basal margin of the ordinary valve; and the basal margin at the basi-tergal corner is angularly and deeply hollowed out. The appearance of the valve is widely different from that in the first sub-variety, yet it is impossible to separate the first and second sub-varieties, and it is almost equally certain that the third sub-variety is only an exaggeration of the second. The lower edge of the adductor ridge, in the third sub-variety, varies a little in outline; it is deeply sinuous, and is produced at the rostral angle into a point, of which we have heretofore seen only a feeble representation. It would appear as if the great development of the adductor plate had caused the exterior ordinary surface of the valve to shrink or be less developed. There is a striking resemblance in the structure here described with that in Pyrgoma cancellatum and conjugatum. The terga belonging to the above scuta, also, vary; the spur being sometimes square (6 o), and sometimes bluntly pointed: when the spur runs in the same exact line (6 q) with the scutal margin of the valve, a peculiar aspect is given to it, but this is by no means always the case. Both opercular valves are often partially coloured pinkish-purple. The shell is not porose; it is thin, with remarkably prominent internal plates; it is apparently always of small size, which I attribute to this variety inhabiting a hard thin plate-like coral. The sheath is bright pinkish-purple, of which we have had instances in some of the other varieties; and the shell itself is sometimes pinkish. Taking the scutum of the[Pg 380] first sub-variety, together with the commonest accompanying variety of tergum, I find it quite impossible to assign to it a specific character; if, on the other hand, we consider the scutum of the third sub-variety by itself, nothing can appear more distinct; but I must repeat, there can be hardly a shadow of doubt that the three sub-varieties of scutum here described, graduate into each other, and are specifically identical.
Var. (7), fig. 6 r.—Hab. Probably Philippine Archipelago, associated with Balanus quadrivittatus.—There can be hardly any question of this being specifically identical with the last variety. It inhabits a different coral. All the specimens were of small size. The walls are not so thin, and the internal ribs not so prominent as in var. 6. The sheath is either white or dull purple; I can, in short, point out no difference in the shell from the typical var. 1. The scutum is not so much elongated transversely as in var. 6, and the basi-tergal corner is more cut off,—in which respect it resembles the common varieties. The adductor ridge is largely developed, so as to be just visible when the valve is viewed from the outside, in a degree between the first and second sub-varieties of var. 6: but the most singular character is the larger development of the tooth near the rostral angle, and this was the case in the same degree in all the specimens which I examined. The tergum resembles that rather unusual sub-variety of var. 6 (fig. 6 q), which has the scutal margin and the one side of the spur forming a straight line. It appears to me that it would be absurd to consider these slight differences, in parts unquestionably subject to much variation, as specific, when we are almost forced to admit that the much greater differences in the three sub-varieties of var. 6, are not of specific value.
Var. (8).—Hab. Unknown, Mus. Cuming.—I have seen only a single specimen of this, and refer to it on account of var. 11. The shell is rather steeply conical, with distant and prominent ribs; the radii are narrow; the walls are not permeated by pores; the colour is pale purple. Altogether its external appearance is very different from that of the foregoing varieties; but the scuta are identical with those of var. 1, excepting that the rostral tooth is rather larger, being nearly as large as in the last, var. 7. The tergum precisely resembles that in some specimens of var. 2. Hence this variety differs from the first two varieties only in the shade of colour, the external shape, and the greater prominence of the external radiating ribs of its shell. All these characters are variable in the several foregoing varieties, and they have been found, as yet, insufficient to discriminate species in any genus of sessile Cirripedes.
Var. (9), Pl. 14, fig. 6 s [C. spinulosa, of Leach (!)]—Hab. Unknown.—The shell is undistinguishable by a single character from many specimens of the first, third, and fourth varieties; it is not permeated by pores. The scutum and tergum, with the exception of the one striking difference of their being calcified together without any trace of a suture, are identical with those of var. 3, as may be seen by comparing the figures 6 h and 6 s. Hence to separate this form specifically from var. 3, we should have to rely solely on the calcifi[Pg 381]cation or union of the scuta and terga; but we have seen this is a point which is variable in Elminius Kingii, Pyrgoma milleporæ, and in some species of Balanus. The serial affinities, moreover, in Pyrgoma, clearly show that this is a character of no great importance. I must add that in several specimens of several of the varieties, the scuta and terga were so closely joined, that until careful examination, I was unable to detect the suture separating them; such being the case it must be quite unimportant for any functional purpose, whether or not the valves are calcified together. I feel, consequently, hardly any doubt that I have acted right in treating the present form as a mere variety.
Var. (10) an. spec.? fig. 6 t.—Hab. Unknown.—This variety bears nearly the same relation to var. 6, as the last variety did to var. 3. The shell is rather stronger than in var. 6, with the internal ribs not so prominent; and except in being tinted pale dull purple, it differs in no respect from the shell of var. 1. If we imagine the scutum and tergum in the third sub-variety of var. 6 (6 p, 6 q), in which the adductor ridge descends far beneath the true basal margin of the valve, to be calcified together, without any suture, we shall produce almost the identical valves of the present variety. The scutum, however, here is not quite so much elongated transversely, and the occludent margin is spinose and is furnished with large teeth; these two characters give the valve a somewhat different aspect, and hence I am more doubtful than in the foregoing case, whether this form may not be specifically distinct. I must, however, state that in Tetraclita porosa, I ascertained that the teeth on the occludent margin of the scuta were even more variable than here is supposed to be the case; and as for the shape of the valve we have seen what wonderful variation there is in var. 6. The tergum in this variety is about intermediate between the two common forms, in the sub-varieties of var. 6. As for the calcification of the two valves together, we have seen, under the last variety, how little important a character it is.
Var. (11) fig. 6 U, 6 u [Creusia grandis, of Chenu, Tab. 1, fig. 2, but not fig. 2 a and b].—Hab. Singapore, associated with Pyrgoma monticulariæ.—This variety is very closely related to the last. The shell, however, has a very peculiar aspect, which made me for some time think it must be specifically distinct. It is of a much brighter pink than in any of the foregoing varieties; the surface is marked with very prominent, distant ribs, and the radii are narrow, in which latter points, together with the tint (though here brighter), this variety cannot be distinguished from var. 7. The shell, however, is permeated by several rows of pores, in which respect it resembles the shell in var. 2, and some specimens of vars. 3 and 4. In the opercular valves there is a close general resemblance with those of the last var. 10; the tooth, however, near the rostral angle, is not so prominent; and in the tergum, the spur is more truncated, shorter and broader than in var. 10, and closely resembles that in var. 1 and 2. But I cannot consider any of the points here specified of much weight.
The foregoing descriptions show how singularly the affinities of the several varieties interlock in the most complicated manner. Hereafter[Pg 382] some one may, perhaps, succeed in grouping several of these forms as species; but I am sure he ought not to attempt it without possessing a very large suite of specimens, or without the great advantage of comparing some two or three of the forms, fresh in their native site.
Species dubiæ.—Under Pyrgoma, I have stated that though Chenu, in his ‘Illust. Conch.,’ has given beautiful external figures of the shells, imbedded in the coral, yet from the want of details on the opercular valves and on the structure of the shell, I cannot recognise his species. So it, likewise, is with Creusia. Chenu gives the following new species; C. radiata, multistriata, decorata, and striata. The C. madreporarum, I suspect to be the Pyrgoma milleporæ of this work, as there stated. The C. grandis no doubt is the P. grande of this work, the Nobia grandis of Sowerby. The name Creusia Childreni is given by Dr. Gray, without description or figure, in the ‘Annals of Philosophy,’ vol. 10, new series, 1825.
CHELONOBIA. Leach. Journal de Physique, tom. 85, (1817).
CORONULA. Lamarck. Animaux sans Vertèbres, 1818.
-------- Ranzani. Memoire di Storia Naturale, 1820.
-------- De Blainville. Dict. des Sciences Naturelles.
ASTROLEPAS. J. E. Gray. (Klein) Annals of Philosophy, (new series), vol. 10, (1825).
Compartments extremely thick, six; but one of them, the rostrum, internally is composed of three rudimentary compartments, united together: basis membranous: scuta narrow, united to the terga by a horny articular ridge.
Distribution, throughout the tropical, and warmer temperate seas of the whole world; attached to turtles, crustacea, or smooth gasteropod molluscs.
This is a distinct and well-defined genus. Several authors have confounded it with Coronula, but this has been owing to an entire misapprehension of the structure of the shell in the two genera. In Coronula, the parietes are very thin, and are so deeply folded as to appear like rays or septa connecting the outside of the shell (the ex[Pg 383]panded ends of the folds) and the internal surface of the shell, but the open spaces between the folds are occupied by the epidermis of the Whale, and are external to the cirripede. In Chelonobia, the parietes are remarkably thick; hence the plates or septa, connecting the outer and inner lamina, are of unusual length; and the spaces between them, though of course internal with regard to the cirripede and occupied by the ovaria, have been compared to the spaces external to and between the folded walls of Coronula. There is but little special affinity between these genera; and I regret that they have come to be placed one after the other in this work: but the elongated opercular valves,—the thick and double opercular membrane,—the weak depressor muscles,—and the peculiar manner in which the scutum is articulated by the aid of a horny projection to the tergum, may indicate some real but slight affinity to Coronula; the many points of difference, however, in the structure of the shell and of the opercular valves, and especially in the cementing apparatus of the basal membrane, and in the branchiæ, all prove that the genera are very distinct. The singular structure of the rostrum, which, in fact, consists of three compartments externally blended together, and which three correspond in all essential respects to the rostrum and two rostro-lateral compartments in the Chthamalinæ, offers a very striking point of identity with that sub-family; but neither in the mouth, cirri, or other part, can I detect any other evidence of this relationship. Having so far discussed the affinities of the genus, I may add, that the three species, though decidedly distinct, are closely and nearly equally related to each other.
General Appearance.—The shell is generally depressed, and broadly oval or almost circular; in C. testudinaria and caretta, it has a massive appearance: the surface is generally smooth, or, when disintegrated, finely striated: the colour is white. The six compartments do not differ much in size: the rostrum is rather larger than the carina, and the lateral compartments, than the carino-lateral compartments. It is remarkable that in C. caretta (Pl. 14, fig. 2), even in specimens which have not grown crowded together, the compartments are almost invariably placed rather un[Pg 384]symmetrically, the rostrum and carina not exactly facing each other.[121] The shell, though so thick and massive, yields easily along the lines of suture. The radii are moderately wide, or narrow, or not at all developed, being represented by mere sutures: in this latter case, in C. caretta, the orifice of the shell is enlarged, in the same manner as we have seen in some species of Balanus and Tetraclita, by the gradual wearing away of the upper part of the shell. In C. testudinaria, the radii have a singular notched structure (fig. 1 a), and the whole shell a star-like appearance. The orifice is not filled up by the elongated opercular valves,—a considerable extent of opercular membrane being visible on the two sides. The largest specimen which I have seen, namely, of C. testudinaria, was nearly two and a half inches in its longer diameter.
[121] In Mr. Stutchbury’s collection there is a specimen of C. testudinaria in which there are only five compartments, one of the lateral compartments having been aborted; of this I have seen no other instance in any genus.
Structure of the Parietes.—The parietes are of unequalled thickness, especially in the first two species of the genus. From the outer lamina (see Pl. 14, fig. 4, and the section in Pl. 15, fig. 1), numerous vertical plates extend inwards, alternately to a less or greater distance, some of them reaching to the inner lamina: these answer to the longitudinal parietal septa in other genera, and the elongated cavities between them (which occur in C. testudinaria and patula) answer to the parietal tubes or pores. The radiating plates or septa have their sides finely channelled, and their basal edges generally slightly sinuous and always finely toothed. The interspaces between the plates in the uppermost part of the shell are filled up solidly, and, in C. caretta, even down to near the basis: in this latter species, the plates are irregular and much broken up, so as in parts to consist of little, separate, flattened points. In C. patula, the inner lamina of the parietes (b, in fig. 4. Pl. 14) can be best made out to be distinct from the sheath (e c e in fig. 4, and c′ in the section of C. testudinaria, fig. 1, Pl. 15). The sheath in this genus descends in a very remarkable manner to the basal membrane, and has its basal edge toothed like the basal edges of the radiating septa. The inner lamina[Pg 385] itself does not descend to the basal membrane. In C. testudinaria, the inner lamina is of great thickness; but in the section, (fig. 1) owing to its having been taken high up, the inner lamina, (b), is not distinct from the shelly matter deposited between the septa. In C. caretta, the line of separation between the inner and outer laminæ can in no part be distinguished, owing to the interspaces between the septa having been solidly filled up, close down to the basis.
Sheath.—The layer of shell surrounding the internal cavity (e c e, fig. 4, Pl. 14), and extending down to the basal membrane, I have called the sheath, owing to its being distinctly continuous with the innermost layer in the upper part of the shell, to which the opercular membrane is attached: this can be best seen by examining the alæ in the separated compartments of C. patula. The sheath is not only remarkable from thus descending to the basal membrane, but in C. testudinaria and patula from its lower edge being perforated by arched channels (under c, in fig. 4), allowing thick ribbons of corium to reach the interspaces between the radiating septa. There is one central arch or channel in the middle of the lower part of the sheath of each compartment, and one on each line of suture, the sheath being a little hollowed out on both sides of the sutures. As the rostrum, as far as its internal structure is concerned, consists of three compartments, we have altogether in the shell eight compartments and eight sutures, and consequently altogether sixteen arches through the lower part of the sheath, allowing sixteen thick ribbons of corium to penetrate the parietes, and thus likewise reach the radii. There is, however, sometimes a little variation in the number of these arched channels. The upper part of the sheath is transversely marked by zones of growth, to the lower one of which the opercular membrane is attached. The line of attachment is not low down the sheath.
Radii and Alæ.—The radii, when the compartments are disarticulated, present a remarkable structure, from appearing to consist of a distinct inner and outer portion. The radius normally consists of an inner and outer lamina, united by septa parallel to the basis; but here the inner portion is formed by a central ridge (a a, fig. 5, Pl. 14), send[Pg 386]ing off on both sides little septa, often sub-branched; it is of nearly uniform width; and there is no distinct inner lamina. The outer portion (b), which often equals or exceeds in thickness the inner portion, is, in fact, the normal outer lamina, developed to an unparalleled degree. In most Cirripedes the edge of the radius is received in a slight furrow in the opposed compartment, the lid of which furrow is narrow, and matches the outer lamina of the radius; here the lid of the recipient furrow is very broad, and resembles the outer lamina of the radius in all its characters. In order to allow of growth between the thick opposed edges of the outer lamina of the radius and the lid of the recipient furrow, the two surfaces are finely dentated (look in fig. 5, under the pits, marked b), almost like the crown of a molar tooth; thus allowing films of corium to enter. The structure here described is common, in a greater or less degree, to all three species, but is best seen in C. testudinaria. In this species, moreover, (fig. 5, b), the outer lamina, instead of being smooth and of either equal or gradually increasing thickness from top to bottom, is generally, but not always, (fig. 1 a), deeply pitted or notched in transverse lines, the outer lamina being thus rendered alternately thicker and thinner, and so formed into transverse ridges and valleys. Hence the lines of suture become toothed, the points of the teeth facing each other, and not interlocking. In the transverse section, fig. 1, Pl. 15, of the same species, taken high up across the shell, (f) is the pitted outer lamina, and (e) the inner portion of the radius. Although the radii are thus specially added to in thickness, they are not so thick as the very thick walls, and hence the lines of suture form furrows more or less deep. The alæ are of moderate thickness, and have their sutural edges crenated by fine transverse septa.
Rostrum.—I have already alluded to the peculiar compounded structure of this compartment, unlike anything we have as yet seen.[122] The thin outer lamina is quite continuous, and shows no trace of the triple nature of the compartment; as may be seen by comparing the drawing[Pg 387] (Pl. 14, fig. 1 a) of the shell of C. testudinaria, with the transverse section (Pl. 15, fig. 1) of the same species: in this latter figure, a is the outer lamina, and B A B the three compartments of the rostrum. But when the outer lamina is worn away, as is always the case with the upper part of the walls in C. caretta (Pl. 14, fig. 2), the two fissures separating the three compartments of the compounded rostrum, are plainly exhibited on the outside of the upper part of the shell. On the internal surface, the sutures separating the three compartments are always open, except in the upper part of the sheath, above the attachment of the opercular membrane, where they are partially obliterated by a thin continuous layer of shell. That these three portions of the rostrum are in their essential nature compartments, is well shown in C. patula and testudinaria, by the sheath or inner lamina having loopholes or channels (such as before described) in the middle of each, and on each line of suture. From the number of these channels, seven altogether, (the two between the compound rostrum and lateral compartments being counted,) the sheath of the compound rostrum is reduced to mere flattened pillars between the several channels. By slight violence, the rostrum breaks into the three portions; and the sutures between them are found to be marked on both sides by sinuous, slight, calcareous ridges, those on opposite sides locking into each; these represent the septa on the edges of the radii and alæ and their recipient furrows, in the ordinary compartments. The outline of the middle compartment of the three (best seen in section Pl. 15, fig. 1, A), much resembles that of the carina; in fact, if we suppose the growth of the two alæ of the carina to have been arrested, no essential difference can be pointed out: in this rudimentary compartment, therefore, we have a rostrum, characterised as in the sub-family of the Chthamalinæ. In the two little rostro-lateral compartments, moreover, (B B), a slight swelling on the side opposite to the large existing radius, shows that if the development of these compartments had not been prevented, each, probably, would have had, exactly as in the Chthamalinæ, a radius on both sides. In the introduction to the Balanidæ, I have argued, from the structure here described and from some[Pg 388] other facts, that in the Balaninæ the rostrum is composed of the three anterior compartments, which we see in the Chthamalinæ, indissolubly united together; hence in Chelonobia the middle one of the three partially-blended compartments is properly the rostrum, and those on the sides the rostro-lateral compartments.
[122] My attention was first called to this peculiar structure of the rostrum by Mr. Stutchbury.
Basis.—The basal membrane extends under the thick walls to the outside. I was not able to make out the whole cementing apparatus. The main trunk is remarkable from its small diameter, (f f in fig. 2, Pl. 28), and from the distance at which the cement-glands stand apart. Each gland gives rise to a pair of cement-ducts, which tend to run in parallel groups; these ducts repeatedly bifurcate, occasionally inosculate, and decrease in diameter; they debouch and allow the cement to escape at numerous points, placed at irregular distances, on the edges of each new slip of the basal membrane.
Opercular Valves.—These are elongated; they do not fill the orifice of the shell; they are attached by a strong opercular membrane a little way down the sheath. The opercular membrane is generally double, for the last-formed membrane is not immediately moulted as soon as a new one is formed, as generally happens in Balanus. Externally, the valves are marked by rather rugged, broad zones of growth. The Scuta are elongated in the line of the orifice of the sack; the occludent margin (Pl. 14, fig. 1 b) is much inflected, and generally sinuous; along this inflected portion a distinct square-edged ridge runs, which widens from the apex downwards. The depression for the adductor muscle is very deep: there are no pits or crests for the other muscles. When the scutum is thoroughly cleaned and all the membrane removed by caustic potash, the tergal margin is marked by a slight articular ridge and furrow. This articular ridge is very remarkable from supporting a prominent, flattened crest (fig. 1 b), composed of hard, yellow, horny membrane, which overlaps the inner surface of the tergum, and exactly corresponds, in shape and purpose, with the calcareous articular ridge, when best developed in other genera. Beneath the basal and generally slightly sinuous exterior margin of the valve, that is, the margin to which the opercular mem[Pg 389]brane is attached, a slight ledge depends (fig. 1 d), which narrows off towards the rostral end of the valve. I should have thought that this had been a depending adductor ridge, as in several species of Pyrgoma, had there not been a nearly similar ledge beneath the middle part of the basal margin of the tergum. The Tergum is mitre-formed, with the summit more or less truncated, and with the carinal margin generally more sloping than the scutal margin: near the carinal margin there is a slight furrow (fig. 1 d), sometimes difficult to be distinguished, with the lines of growth curving down to it on each side, and consequently with a very slight, but variable, corresponding projection on the basal margin. This furrow and slight projection, there can be hardly any doubt, represent the spur, though here placed close to the carinal instead of to the scutal margin, as in other genera. The tergum has a small articular ridge, against which the overlapping horny articular ridge of the scutum abuts. There are no crests for the tergal depressor muscles. Altogether the scuta and terga are very peculiar. A portion dissolved in acid exhibits no tubuli. On the opercular membrane there are no hairs. The rostral depressor muscles of the scuta are singularly feeble, each consisting of only one or two, or sometimes three or four, very thin ribbon-like fasciæ; the lateral depressores of the scutum are twice as strong as the rostral depressores; and the tergal depressores a little stronger than the lateral depressores. All these muscles are plainly marked with transverse striæ.
Mouth.—The labrum is not in the least bullate, which character, as well as some others that follow, I specify on account of the apparent affinity of Chelonobia to the Chthamalinæ, as indicated by its rostrum. The crest of the labrum is notched, and on each side of the notch there is a long row of teeth, which, however, are sometimes worn away. The mandibles have five main teeth; all excepting the first being laterally double. The maxillæ are not notched. The outer maxillæ are bilobed on their inner sides.
Cirri.—The four posterior pairs, in proportion to the size of the animal, are much elongated, and are remarkable from the number of their short segments. The rami of the first cirrus are a little unequal in length. The second[Pg 390] cirrus is moderately short, with its segments rather broad and protuberant, and thickly clothed with spines. The third cirrus is of unusual length, being but little shorter than the fourth pair; its segments, however, are broad, and are thickly clothed with spines, as are the two segments of its pedicel: hence there is no real approach to that important character of the Chthamalinæ, namely, the similarity of the third with the three posterior pairs of cirri. The numerous segments of the fourth, fifth, and sixth pairs of cirri each support only two pairs of main spines; between each of these pairs there is a little tuft of fine intermediate spines; the upper of the two tufts on each segment is the longest. In a specimen of C. patula, in which there were fifteen segments in one ramus of the second cirrus, there were fifty segments in either ramus of the sixth cirrus. At the exterior bases of the pedicels of some of the anterior cirri, there are large tufts of finely plumose, delicate hairs.
Branchiæ.—These are of large size: they consist of a single fold, much plicated and sub-plicated.
Ovaria.—The ovarian tubes run into the parietes, and fill up the interspaces between the radiating septa.
Range, &c.—The three species seem to range together, over the tropical and warmer temperate seas of the whole world. C. patula and testudinaria are found in the Mediterranean, and the former at Charleston, in the United States; I have not heard of specimens from any point further north. C. testudinaria and caretta live attached to turtles; whilst C. patula always adheres to crustacea, to large and smooth gasteropod mollusca, and, I believe, sometimes to ships’ bottoms. I have not heard of the discovery of any fossil species.
Attachment.—Chelonobia patula leaves no impression on the crabs and shells to which it is attached. I have seen only a few specimens of C. testudinaria attached, and the carapaces of the turtle were not at all, or scarcely at all, indented by them. The case is very different with C. caretta, in which the shell, even of young specimens, is always, as far as I have seen, imbedded to some depth, and occasionally to a very great depth in the tortoise-shell. From the extreme hardness of the tortoise-shell, when dry, the imbedment[Pg 391] appears more wonderful even than it really is. The younger shells have the appearance of having grown from within the carapace, and then of having burst through it, almost like little volcanos. I have seen only one very young shell (1/10th of an inch in external diameter) attached, and here there was nothing to countenance an idea which at one time occurred to me, namely, that the larva perhaps fixed itself in some little crack or cavity in the carapace, and there underwent its metamorphosis. I believe that the imbedment is effected simply by the sharp, growing, basal edges of the walls of the shell indenting the surface, and finally rupturing the outer laminæ of the tortoise-shell, through that same force by which the tender radicle of a plant penetrates very hard ground. As soon as the surface was once ruptured, the shell of the Chelonobia, growing outwards and downwards, would easily, like a wedge, turn up the laminæ of the tortoise-shell; and their ragged ends would surround the Chelonobia, as is seen actually to be the case. In the genus Coronula and its allies, which are attached to Cetaceans, we shall presently see, that the epidermis immediately under the downward growing shell, and apparently in consequence of the pressure thus exerted, is thinner than in the surrounding parts. In two specimens of Chelonobia caretta, imbedded much more deeply than usual—in one of which half the basal edge of the shell had fairly cut through the carapace, and in the other was on the point of effecting this—the tortoise-shell manifestly thinned out towards the line of yielding; now this, I suppose, must be attributed either to absorption, or to the living tortoise-shell being actually stretched till rendered transparent and ready to burst or until bursted. On the latter view of the tortoise-shell having been stretched, we must further suppose that the pressure has prevented fresh layers of tortoise-shell being deposited under the old and yielding laminæ. In one of the above two specimens, the walls of the Chelonobia were deeply folded, nevertheless the laminæ of the tortoise-shell followed every curvature, showing that, though now so rigid, during the slow imbedment of the Cirripede it must have been sufficiently pliant. A shell attached, as these two specimens were, could never be removed, and, whether[Pg 392] dead or alive, would remain for ever imbedded in the tortoise-shell. Dr. R. Ball, of Dublin, informs me, that he has seen specimens in which the shell of the cirripede not only had penetrated the carapace, but likewise the underlying bone, and had even entered some way into the body of the turtle: it is well known that the tusk of a boar or the horn of a ruminant, when curving in abnormally, will sometimes penetrate deeply into the bones of the face or head; and this, I believe, is effected, not by the fracture of the bone, but by the absorption of the point pressed on: I conceive a similar process must have taken place in the curious specimens examined by Dr. Ball.
LEPAS TESTUDINARIA. Linn. Syst. Naturæ, 1767.
VERRUCA TESTUDINARIA. Ellis. Phil. Transact., vol. 50, 1758, Pl. 34, fig. 12.
BALANUS POLYTHALAMIUS. Bock. Naturforscher, Stuch. 12, 1778, fig. 9.
LEPAS TESTUDINARIA. Poli. Testacea Utriusque Siciliæ (1795) Tab. 5, fig. 9-11.
CORONULA TESTUDINARIA. Ranzani. Memoire di Storia Naturale, Decade 1, (1820).
-------- ---------- De Blainville. Dict. des Sciences Nat., (1824), Tab. 117, fig. 2.
CHELONOBIA SAVIGNII (?) Leach. Encyclop. Brit. Suppl., vol. 3, 1824.
ASTROLEPAS ROTUNDARIUS. J. E. Gray. Annals of Philosoph. (new series), vol. 10, 1825.
Shell conical, depressed, heavy: radii rather narrow, depressed, generally notched on both sides.
Hab.—Mediterranean; west coast of Africa; north-east coast of Australia; Low Archipelago, Pacific Ocean; west coast of Mexico, 23° N. Attached to turtles.
It is impossible to feel sure which of the three species of the genus Linnæus had in view, when describing his[Pg 393] Lepas testudinaria; but as Spengler has well discriminated the following species under the specific name of caretta,[123] and Ranzani the third species under patula, the present name may, without question, be retained for the following species. In several respects this species is intermediate between C. caretta and patula, but it can most conveniently be described first.
[123] I am indebted to Dr. J. E. Gray for having guided me to this identification.
General Appearance.—Shell strong, globulo-conical, depressed; outline broadly oval; surface smooth, generally well preserved, but when disintegrated, upper part finely striated; colour dead white; orifice oval elongated, rather exceeding in length one third of the longer basal diameter of the shell. The radii are rather narrow, and deeply depressed; they have their summits square: their outer lamina, as explained under the genus, on both sides of each suture is in most specimens divided into teeth, the points of which face and touch each other. These teeth or notches give quite a peculiar appearance to the shell, and alone suffice to discriminate this species; they are sometimes blunt and partially obliterated, but it is rare to find a specimen in which some few teeth do not occur in some one of the six radii. I have, however, seen two or three specimens with all six radii perfectly smooth; in one of these the general shape of the shell, without the aid of any internal characters, almost sufficed to show that it belonged to the present species; but in another specimen, which had unusually narrow radii, and the whole surface of which had undergone considerable disintegration, and was consequently striated, could only be distinguished from C. caretta by internal characters. I have seen several specimens having very irregularly shaped compartments, but generally the appearance of the whole shell is highly symmetrical, like a star; and the genus was appropriately named by old Klein, Astrolepas. In some specimens, in Mr. Cuming’s collection, from the Low Archipelago, in the Pacific, taken off the toe-nail of a turtle, the shape was almost cylindrical; the shell almost resembling that of Coronula diadema. The largest specimen which I have seen was 2.3 of an inch in basal diameter, but only .55 in height.
Structure of Shell and Radii.—After the full generic description, the only point to which I need allude is, that the radiating, parietal septa, as well as the descending sheath, are much thicker than in C. patula, and that their basal edges can be plainly seen by the naked eye to be dentated with numerous points. The thickness of these plates and of the sheath varies considerably. In C. caretta, I may add, the plates are broken up into many separate points, and in this species the descending sheath is not generally perforated, excepting at the sutures, by loop-holes for the entrance of ribbons of corium. The[Pg 394] sheath and inner lamina of the parietes in C. testudinaria, taken together, fully equal one third of the entire thickness of the walls. Flattened cavities or tubes run up between the parietal septa for about two thirds of the height of the inside of the shell. With respect to the radii, I have only to add, that the thickness of the inner portion relatively to the generally notched outer lamina, varies considerably.
Opercular Valves (fig. 1 b-1 d).—These closely resemble each other in all three species, and have been almost sufficiently described under the genus. The scutum has its occludent margin always sinuous, but to a variable degree. In one specimen there was a deep little pit at the rostral end for the rostral depressor muscle. The terga vary considerably in shape, according as the basi-carinal corner is more or less truncated. The external furrow, marking the position of the rudimentary spur, varies much in distinctness, chiefly depending upon its nearness to the carinal margin of the valve.
Mouth and Cirri.—The labrum sometimes has its teeth worn away. The second pair of cirri, relatively to the others, is shorter and thicker than in the two following species.
LEPAS CARETTA. Spengler. Skrifter, Naturhist. Selbskabet, Bd. 1, 1790, Tab. 6, fig. 4.
BALANUS CHELYTRYPETES. Hincks (!) sine descript., Annals of Nat. Hist., vol. 5, p. 333.
CORONULA SULCATA. Chenu. Illust. Conch., Tab. 1, fig. 1.
Shell globulo-convex, extremely massive and heavy; upper part worn, sub-striated: radii either not developed or very narrow: parietes without cavities extending up between the interrupted, radiating septa.
Hab.—West coast of Africa; northern Australia; common. Attached to, and generally imbedded in, the carapaces of turtles.
General Appearance.—Shell extremely strong, massive, heavy, broadly oval, globulo-convex, though sometimes depressed; dirty white: surface in the upper part always worn and disintegrated, hence finely striated; in the lower part a little rugged, slightly folded, and occasionally, in deeply imbedded specimens, much folded or ribbed. Orifice oval, not at all angular, rather exceeding in length one third of the basal diameter of the shell. Radii either not at all developed, or very narrow; but even in the former case the six lines of suture are plain; and in the latter case the radii lie at some depth beneath the surface of the shell. It is remarkable that the compartments are hardly ever arranged symmetrically, the rostrum and carina not[Pg 395] facing each other exactly; and this holds good in specimens attached separately, without any apparent cause for this want of symmetry. The largest specimen which I have seen was 2.1 in basal diameter; and this, which was a steeply conical individual, was 1.1 in height; and therefore nearly twice as high as an ordinary large specimen of C. testudinaria.
Structure of Shell and Radii.—The descending sheath and radiating septa are of very variable thickness, and have their basal edges finely dentated. The septa are not continuous, from the circumference to the sheath, in unbroken plates, but are irregularly divided into separate, often short portions, and even occasionally into mere points. The sheath differs from that of the other two species in having loopholes for the entrance of ribbons of corium only on the eight lines of suture, and not (with rare exceptions) in the middle of each compartment: this is evidently due to fewer filaments of corium being here sufficient to supply the less deep interspaces between the radiating septa; for in this species there are no flattened cavities or tubes running far up the shell. The inner lamina of the walls cannot be here distinguished, for a solid, flat, calcareous surface extends from the circumference, between the radiating septa, to the sheath. The sheath, had it not been from the light thrown on this part by the other species, would have certainly been mistaken for the inner lamina of the walls. The absence of the flattened cavities or tubes extending up the parietes, seems to be the least varying character; and serves to distinguish this species from those worn and massive specimens of C. testudinaria, which have narrow and not-notched radii.
In specimens in which the radii are not developed, no vestige of the outer lamina can be detected, the lateral faces of the adjoining compartments being closely pressed together; but in specimens in which the radii have been developed and have grown, the outer lamina of course is present, and is extremely thick, with the growing edge having a branched and mammilated surface, as in C. testudinaria, but with the external surface not pitted or notched as in that species. The inner portion of the radius, whether or not developed, has nearly the same appearance, consisting of rather thick transverse septa, branching from a central ridge, which is sometimes obscure.
The Alæ are remarkably little prominent, as least in those specimens in which the radii are not developed, so that Spengler seems to have thought that the structure of the shell was essentially different from that of Balanus, which certainly is by no means the case. The edges of the alæ are very thick, nearly as thick as the inner portion of the radius.
The Opercular Valves hardly present any essential difference, compared with those of the other species; but the occludent margin of the scutum is apt to be more sinuous, and its rostral end blunter and squarer. The carinal end of the tergum is also squarer than in any common variety of C. testudinaria; the external furrow or spur, near the carinal margin, is very indistinct, and even sometimes is quite absent.
Neither the mouth nor cirri present any deviations from the generic character.
CORONULA PATULA. Ranzani. Memoire di Storia Naturale (1820), Tab. 3, fig. 25-28.
ASTROLEPAS LÆVIS. J. E. Gray (!). Annals of Philosophy (new series), vol. 10, 1825.
VERRUCA CANCRI AMERICANI. Ellis. Phil. Trans., vol. 50, 1758, Pl. 34, fig. 13.
Shell steeply conical, very smooth and light; orifice large, generally exceeding half the basal diameter of the shell: radii broad, smooth, only slightly depressed.
Hab.—Mediterranean; Gambia, West Africa; Charlestown; Jamaica; Honduras; Brazil; Australia. Attached to Crustacea, smooth univalve shells, and apparently to ships’ bottoms.
General Appearance.—Shell white, very smooth, of little specific gravity, steeply conical, but not high; orifice broadly oval, polygonal, very large, namely, generally exceeding half the basal diameter of the shell. The summits of the compartments are usually perfectly preserved, pointed, and often a little recurved. The radii are rather broad, very smooth, with their summits slightly oblique and arched: they are seated only a little below the general level of the parietes. I have seen one specimen rather more than one and a half inch in basal diameter, but this is an unusual size; this species not growing to so large a size as the two foregoing forms.
Structure of Shell and Radii.—The walls are here thinner than in the two foregoing species; and the basal surface of a compartment rarely equals half the basal diameter of the internal cavity of the shell, measured transversely to its longer axis. The radiating septa are also much thinner, generally sinuous, and so finely dentated along their basal edges, that the teeth can be clearly perceived only by the aid of a lens. The interspaces between the septa run up to nearly the summits of the compartments, and hence the lightness of the whole shell. The inner lamina of the parietes is here not so thick (fig. 4), and is more distinct from the descending sheath than in the foregoing species. The sheath is thin, like the radiating septa; the medial loophole in each compartment, for the entrance of a filament of corium, is much wider than in C. testudinaria, for it is generally as wide as the bordering plate on either hand; and in not a few specimens, the medial loophole is so wide as hardly any longer to deserve being so called, for the descending sheath is reduced to mere flattened pillars or legs on the sides of the sutures. Although the parietes are here not nearly so thick as in the two foregoing species, yet as the radii stand but little beneath[Pg 397] the level of the parietes, the outer lamina of the radius has a considerable thickness, and is sometimes separated from the inner portion of the radius by an interval, in the same manner as in C. testudinaria; and the growing edges of the outer lamina exhibit traces of the same mamillated structure as figured in that species.
The Opercular Valves are apt to be rather narrower and more elongated than in the foregoing species, with the occludent margin of the scutum generally, but not always, only slightly sinuous: externally the scutum is sometimes feebly striated longitudinally. In the tergum (Pl. 14, fig. 3 b) the longitudinal furrow is generally plainer, from being more distant from the carinal margin, and the rudimentary spur itself is more prominent.
The mandibles usually have five narrow teeth; but I have seen one specimen having only four teeth, and with the inferior angle truncated. The cirri present no particular character.
[Scutum and tergum (when both are present) not overlapping each other, or articulated together; basis membranous; parietes often deeply folded, with the outer lamina, towards the basis, generally imperfect; each branchia composed of two plicated folds; shell attached to living vertebrata.]
CORONULA. Lamarck. Annales du Museum, tom. 1 (1802).
DIADEMA. Schumacher. Essai d’un Nouveau Syst., &c., 1817.
CETOPIRUS (SED NON CORONULA). Ranzani. Memoire di Storia Naturale (1820).
POLYLEPAS. J. E. Gray, (Klein). Annals of Philosophy, (new series), vol. 10, 1825.
Compartments six, of equal sizes: walls thin, deeply folded, with the folds forming cavities, open only on the under side of the shell: opercular valves much smaller than the orifice of the shell.
Distribution, mundane, imbedded in Cetacea.
This genus and the three following, namely, Platylepas, Tubicinella, and Xenobalanus, have very considerable claims to be separated as a sub-family, as has been proposed by Drs. Leach and Gray. Although in structure and habits they are certainly closely related together; yet only few characters can be predicated of all four in common,—some characters failing in one genus and some in another. All four, however, differ from the foregoing genera in the opercular valves not being articulated together, being simply united by tough horny membrane; but Chelonobia makes some approach in this respect. All four have the branchiæ composed of a double fold; but this can hardly be considered a character of much weight, as may be inferred from the remarks made on this subject at p. 153 of the Introduction, where the differences of these four genera from the other genera of the family are discussed at some length. Altogether I have been led to conclude, though with much hesitation, that these genera had better not be separated as the sub-family of Coronulinæ.
Our present genus, Coronula, is closely related to Platylepas, and likewise to Xenobalanus, though this latter genus is so very different in external aspect that it might easily be mistaken for a pedunculated Cirripede. Lamarck and some other authors have placed the species of Chelonobia under Coronula, but this has arisen, as explained under that genus, from a misapprehension of their structure; the folded very thin walls in Coronula having been compared with the radiating septa of the very thick walls in Chelonobia. I may further add, that Coronula has been divided into two genera by Ranzani, on palpably insufficient grounds.
General Appearance.—The three recent species of this genus have a very handsome and striking appearance. The shell is highly symmetrical, owing to the six compartments being of the same size and having exactly the same outline. The general shape is either depressed or, as expressed by the name, like a crown. The walls are longitudinally and slightly ribbed, owing, as we shall presently see, to their wonderfully folded structure; and the surface is marked by very fine longitudinal striæ, crossed by finely beaded lines[Pg 399] of growth. Hence the walls offer a strong contrast in appearance with the six, smooth radii, of equal breadth. The symmetrically hexagonal, or rounded-hexagonal orifice of the shell is closed by a thick, nearly horizontal membrane, supporting, towards the rostral end, the small opercular valves, with a slit, having protuberant lips, in the middle, for the protrusion of the cirri. The opercular membrane is attached all round, but a short distance beneath the summit of the shell. In regard to size, I have seen a specimen of C. diadema two inches in height and two and a half in diameter.
Structure of the Shell.—The structure at first appears singularly complicated, and quite unlike that of any other Cirripede; but the whole results simply from the folding of the very thin walls, which in all essential respects are constructed like those in Balanus. In a young specimen of C. balænaris, having the orifice of the shell 2/10ths of an inch in diameter, I found the upper part of each compartment only slightly sinuous, not more so than is common in many varieties of Balanus, but more symmetrical, for each compartment had three slight furrows, making, for the whole shell, eighteen furrows. The ensuing changes during the growth of the shell will be best understood by looking at the diagrams a, b, c, in fig. 10, Pl. 15, which are supposed to represent the basal margins of the walls of a single compartment: (a) shows the simply sinuous wall of the young shell. As the shell grows, the furrows rapidly grow deeper and deeper (b), and wider and wider; at the same time, the folds or ridges between the furrows gradually become drawn out at their ends into transverse loops (c), the extremities of which ultimately become closely pressed together—the furrows being thus converted into cavities, extending from the top to the base of the compartments. This structure in the mature shell will be best understood by looking at the transverse or horizontal section (Pl. 16, fig. 7) of the rostral end of the shell of C. diadema, in which species the folding of the walls is simpler than in C. balænaris or C. reginæ: the walls (e) are represented by a double line connected by little cross lines,—(h) being one of the transverse loops at the outer end of one of the folds,[Pg 400] and (f′) one of the cavities between the folded walls, open at the bottom of the shell, and occupied by the epidermis of the Whale. The walls of the compartments, as here represented, (A being the rostrum, C C the lateral compartments, D D the alæ of the carino-lateral compartments,) are separated from each other by the broad radii; but if the section had been taken low down near the basis, the end of the folded wall of one compartment would have been separated from that of the adjoining compartment only by the close suture: this will be understood by a glance at the entire shell, given in fig. 3, Pl. 15. The rostrum of the same species, viewed from the inside, is shown at fig. 1, Pl. 16; here it may be observed, that the basal margin (e e′ e′′) of the folded wall is extremely oblique, the outer portion having extended downwards much more than the inner portion: this obliquity is more clearly shown in the lateral view (fig. 2) of a lateral compartment, for this figure will equally well serve for a lateral view of the rostrum (fig. 1), if the ala (a′) be supposed to be removed. The section of the rostrum in fig. 7, (A), will now be intelligible in relation to the view of the rostrum given in fig. 1, if it be borne in mind that the section has been taken high up, near the letters a, b, c′′ in fig. 1.
As above stated, the folds or ridges in the young shell become more and more transversely drawn out at their ends into the transverse loops, till the latter join and touch each other. In C. balænaris the lines of junction are simple, though very close; in the other species, the ends of the transverse loops, where touching, are finely and elegantly toothed, and thus locked together. These teeth appear single when the shell is viewed either externally or internally, but when the walls along the lines of junction are forced apart, they are seen really to consist of transverse rows of minute teeth. These teeth are less distinct, forming only sinuous ridges in C. barbara (fig. 6, Pl. 15); in all cases the teeth are formed by the modification of the very minute beads, which ornament the lines of growth on the external surface of the shell. In the rostrum of C. diadema, as seen internally (fig. 1, Pl. 16), the serrated and closely-fitting lines of junction (f) between the ends of the[Pg 401] transverse loops of the folded walls are plainly shown. It is the more or less rounded surfaces of the transverse loops which give to the external surface of the parietes its longitudinally ribbed structure: the ribs are plainest in C. diadema, fig. 3, Pl. 15. The shell, in fact, as seen externally, consists of but an extremely small portion of the external surface of the whole length of wall, being exclusively formed of the transversely looped ends of the radiating folds, together with the radii. Owing to the ends of the transverse loops being so closely pressed together, the furrows are practically converted (as already remarked) into cavities, open only on the under side of the shell, and extending from the oblique bases of the compartments up to their apices; and these are invariably filled by the black epidermis of the Whale. Owing to this circumstance, the skin of the Whale has been mistaken by some authors for parts of the Cirripede! In C. diadema, in which the summit of the shell is often a little disintegrated, the whale’s skin is often there exposed, forming three black spots at the top of each compartment. It should always be remembered that these flattened and deep cavities are furrows, which homologically ought to be open in longitudinal lines along the external surface of the shell, from the top to the bottom.
As the shell increases in diameter, each of the original eighteen transverse loops, forming the exterior surface of the shell, increases in breadth; and they would have had to increase extremely, had not some of the transverse loops become, during growth, divided into two or three new transverse loops, in a manner strictly analogous with the first formation of the eighteen folds in the young shell. In Pl. 15, fig. 7, 8, 9, we see how one of the circumferential transverse loops, by the formation of a medial furrow, or rather bay, becomes developed into two transverse loops; and it is rather important to observe that three new loops might equally well have been contemporaneously formed. By the repeated formation of new circumferential loops and the consequent formation of new folds, the wall of the shell, when old, especially in C. balænaris, becomes folded in a wonderfully complicated manner, as may be seen in Pl. 15, fig. 5, which is an exact tracing of the extreme basal edge of the wall of[Pg 402] a shell of C. balænaris; to perceive the full amount of complication, it is advisable to trace the wall of any one of the compartments, from one suture (s) to another. In this figure the sutures are purposely drawn a little open. It may be seen that the new transverse loops, and consequently the new folds of the walls, have been, in this species, mostly formed in symmetrical order, on both sides of the six sutures; this results from the transverse loops on the sutures almost always giving rise contemporaneously to three new transverse loops. In C. diadema the transverse loops on the sutures usually divide into only two new loops, one on the rostral side and one still remaining at the suture; hence the folding of the walls in this species is much less symmetrical. The number, however, of the transverse loops and the exact pattern of the folding is variable in all four species of the genus. I may further add, to show the complication of the folds, that in a shell of C. balænaris, having a basal diameter of two inches, and which had the walls as little folded as ever they are, yet I found, by careful measurement, that the entire basal edge of the wall, if stretched straight, would have extended for a length of fifty-two inches! Therefore, if the wall had not been folded, but had been simply circular, as in ordinary cirripedes, the basal diameter of the specimen would have been between sixteen and seventeen inches!
The central membranous basis is flat, but the bottom of the folded walls of the shell is concave, which is caused by the outer ends of the folded walls having grown at a greater rate than the inner ends. The concavity is deep in C. diadema; in C. balænaris it is much less so, and here the inner hood-like ends of the folded walls are rather abruptly, but in a variable degree, produced downwards, generally even slightly beneath the level of the circumference of the shell; this fact is of interest in relation to the peculiar, depending, spur-like processes in the genus Platylepas. A lateral view of a compartment in both these species, is given in Pl. 16, figs. 2 and 3; and by supposing in each case a compartment to stand opposite, at a distance which may be judged of from fig. 5, the vertical sectional outline of the whole shell will be understood: in fig. 3 of[Pg 403] C. balænaris, however, the inner hood-like ends of the folded walls are not produced so much downwards as is usual.
In the same manner, as the outside of the shell consists of the transversely expanded ends of the folded walls, pressed closely together, so the cavity in which the animal’s body is lodged, is formed by the inner and less closely joined ends of the folds, lined by the thick sheath (a, fig. 1 and 7, Pl. 16), which latter extends down very near to the basal membrane. The cavity for the body, is small compared to the whole shell; in C. diadema it is deeply cup-formed, with a small, flat, membranous bottom or basis; in C. balænaris it is wider and shallower, with a broader bottom, and with the upper edges of the walls more inflected. In both species, the thick membrane connecting the opercular valves to the shell, is attached all round near the summit of the sheath. The uppermost portion of the sheath is not marked by concentric lines, as in most of the Balanidæ, owing to the opercular membrane not being, as we shall presently see, regularly moulted. A portion of a single wall, when closely examined, is found to be formed of an outer and inner lamina, united by longitudinal septa, and is thus permeated by minute, square, longitudinal pores,—exactly as in the normal structure of Balanus. The walls are extremely thin, and are striated longitudinally, owing to the slight projection, on both the inner and outer surfaces, of the longitudinal septa; they are thicker in the part forming the external transverse loops, being here, in C. balænaris, as much as 15/1000ths of an inch in thickness; but when forming one side of the spoke-like folds, the thickness is only 6/1000ths of an inch. The inner lamina is thicker, contrary to what is usual, than the outer lamina; the sharp tips of the longitudinal septa project a little beyond either lamina, giving to the basal edge of the wall a serrated outline. It is singular that the thin outer lamina is first formed as a rim or ledge on each side of the longitudinal septa; these ledges being not closely united for some little space up the wall, as is represented in the enlarged drawing of a bit of the basal wall of C. diadema, Pl. 16, fig. 6. The open clefts thus left are, of course, covered by the so-called epidermis, for[Pg 404] otherwise the included threads of corium would have been exposed. Each fresh period of growth, in the case of C. diadema (fig. 6) and reginæ, and to a certain extent in C. balænaris, is marked by little knobs on the longitudinal, slightly prominent, septa, and this prefigures an analogous strongly marked structure in Tubicinella. A fine thread of corium runs up each pore to the summit of the compartment; for these pores are not, as in Balanus, cut off by transverse calcareous septa, or have their upper ends solidly filled up with shelly matter. As, however, the summit of the shell in Coronula is sometimes disintegrated, the threads of corium within the pores would have been exposed, had not each thread formed for itself, as I suppose, a transverse membranous septum near the summit of the shell; at least this is the case with the larger pores of the radii. The walls, where closely pressed together in the spoke-like folds, are disunited at the extreme base, but above this they are firmly calcified together. A ribbon of corium runs along the basal edge of each spoke, and sends threads of corium up the parietal pores on each side, and its upper edge serves to deposit calcareous matter (homologous with the layers of the sheath) and thus to unite the two walls firmly together. In C. diadema, the walls of the terminal transverse loops are simply calcified together like the spoke-like portions; but in C. balænaris the opposite sides of the loops are united by septa (see the transverse section in Pl. 15, fig. 2 a), making from five to eleven longitudinal tubes within each transverse loop; these tubes being larger than the parietal pores. When a piece of the shell is dissolved in acid, no tubuli can be discovered, which may be accounted for by the thinness of the walls; nor are there any spines on any of the external membranes. The number of the pores, in the parietes of a moderately-sized specimen of C. balænaris, I calculated was at 3400, each occupied by a thread of corium springing from the eighteen branched ribbons, diverging from the corium, surrounding the base of the sack. To this number must be added between 300 and 400 larger threads of corium running up the tubes in the transverse loops; and no less than about 2300 fringes and threads occupying the pores in the six radii: thus we see that the dermal system in Coronula is wonderfully complicated.
Radii.—The radii are very wide in C. diadema. In all the species their summits are square or parallel to the basis. Their internal structure is remarkable: as the walls in this genus are extremely thin, so are the proper radii, for in fact they consist in this and all cases, as we know, of one margin of the wall modified by its lateral growth against the opposed compartment. But as the radii in Coronula are not plicated, like the walls, the shell would have been excessively weak along the six lines of suture, had not the radii been strengthened by numerous sinuous plates, springing from the inner lamina of the proper radius, and running downwards, attached to the folded wall of the compartment to which the radius belongs, and with their free edges pressed against the folded wall of the opposed compartment. These plates give out short transverse denticuli, making altogether a beautiful structure, as is best seen in Pl. 16, fig. 3, but also in fig. 2 and 4, and d in fig. 1. In the section, fig. 7, the proper radius (d) is seen to be continuous with the wall (e), and to be very thin, in fact forming but a small portion of the compound radius: it is formed of an outer and inner approximate lamina, separated by septa, which are nearly horizontal, and which consequently cannot be shown in the transverse section fig. 7. The outer lamina of the radius is imperfect, or does not reach quite to the suture, leaving the septa a little exposed (imperfectly shown in Pl. 16, fig. 3), exactly as is the case with the outer lamina of the parietes, at the basal margin of the shell. In all common Balanidæ, a ribbon of corium runs up each of the six sutures, and sends in fine threads between the septa of the radii, but here a thread of corium runs up a minute, cylindrical pore, situated on the line of junction between the radius and the wall whence it arises; and from this longitudinal thread the finer threads spring which pass between the horizontal septa of the proper radius: this cylindrical pore is rather large in C. balænaris, but excessively small in C. diadema (see a black dot (d′) in section, fig. 7), and is solidly filled up in the upper part of the shell. The plates (fig. 3) which run down from the inner lamina of the proper radius, and form the greater part of its thickness, are occupied by fringes of corium, extending up from a ribbon of corium,[Pg 406] running along each suture, like those which run along the bases of the spoke-like folded walls. This difference in the origin of the ribbons of corium, occupying the interspaces between the plates and the pores in the proper radii, shows the essential difference between the latter and the thick inner portions of the compound radii. In C. balænaris (fig. 3), the compound radius extends from the outside of the shell to the sheath and to near the basal edges of the folded walls: in C. diadema (fig. 2), it does not extend so far inwards and downwards; and in C. reginæ (fig. 4), even still less so. It ensues from this circumstance that when, in these two latter species, a transverse section is made across the middle of the shell, a large chamber (v, fig. 7), occupied by the ovaria, is found on each line of suture (s s), separating the radii (d) and alæ (a).
The Alæ in C. diadema (section, Pl. 16, fig. 7, a′) and C. reginæ, are of remarkable thickness, nearly equal to that of the radii, and in C. balænaris, of considerable thickness; this is evidently to give strength to the shell, which is weakest along the lines of suture, notwithstanding that the radii have been specially thickened. The edge of each ala presents a miniature resemblance of the edge of the radius, namely, a central ridge sending off on both sides sinuous plates, themselves denticulated. In C. balænaris, the ala rests almost entirely on the inner surface of the compound radius; but in the other two species, in which, as already stated, the radii and alæ are separated by chambers, the ala rests on a plate (c′′ in fig. 1, 4, and 7), which extends from the top of the radius to the bottom of the sheath, narrowing downwards (c′′, fig. 1), and is a specially developed portion of the sheath for the radius to rest on. In C. diadema and reginæ, the sides of the folded walls, at the ends of the chambers (v, in section, fig. 7), are strengthened by the deposition of layers of shell in connexion with the sheath. The sheath extends close down to the basal membrane in C. balænaris, and does not project freely: in the other two species it depends freely, but does not run quite so low down. I have only further to remark, that the sutures, though very strongly united, are not calcified together; for they easily separate after the action of caustic potash.
The Basis (Pl. 28, fig. 1 a-1 c) is membranous and is[Pg 407] formed of concentric slips: each slip has eighteen angles, corresponding with the open ends (see Pl. 16, fig. 5) of the eighteen folds of wall. A ray of membrane runs under each of these folds, being prolonged from the basal membrane; but these rays can hardly properly be called parts of the basal membrane, for they split at each period of growth along the middle, and the two halves are drawn from under the basal edges of the walls, and thus come to invest their outer surfaces. The basal membrane, and the whole cementing apparatus, which is much simpler than in Balanus, has been fully described (p. 135) in the Introduction.
Opercular Valves.—In the sessile cirripedes hitherto examined, the four opercular valves are inclined towards each other, and nearly fill up the orifice of the shell, being united to the walls by a more or less narrow circular border of membrane; this membrane being attached rather low down to the sheath. In Coronula the opercular membrane is stretched like the skin of a drum, almost horizontally across the top of the shell; it is, however, generally attached to the sheath rather lower down at the carinal than at the rostral end: hence the animal’s body, as remarked by Burmeister, is attached almost horizontally; but this we shall see, under Xenobalanus and Tubicinella, cannot be considered a character of much importance. The valves are quite small compared with the opercular membrane, and certainly are of little functional importance. The scuta in C. balænaris and diadema (Pl. 15, fig. 3 b) are sub-triangular; but the under or growing surface is elongated and arched. These valves stand almost at the rostral end of the orifice, instead of on each side of it. In C. balænaris the terga are small, with the under surface oval: in C. diadema they are either quite aborted, or are represented by a barely visible plate of shell, parallel and close to the tergal margin of the scutum. The aperture leading into the sack is formed by a nearly medial slit of considerable length, furnished with irregularly protuberant, inwardly inclined lips. These lips are formed by the development of an inner fold or crest of membrane, which can be just detected in most Cirripedes: the lips include a double fold of corium, and are covered by a delicate tunic, continuous with that lining the sack, and homologically continuous[Pg 408] with the opercular membrane. The opercular membrane is very thick, tough, and yellowish; it is, in parts, finely plicated in lines radiating from the apices of the valves: these plicæ consist of membrane in an altered condition, being harder, more horny, and of a browner yellow; the plicæ are large at the rostral end of the scuta, and projecting beyond these valves, they afford attachment to the rostral depressor muscles. Rims of similarly modified membrane (Pl. 15, fig. 2 b) connect the scuta and terga together. The rims and plicæ are occasionally moulted together with the opercular membrane. In almost all hitherto described Balanidæ, a new opercular membrane is formed at each period of exuviation, and as soon as formed, the old one is generally moulted, together with the other membranes of the body: the case is very different in Coronula, in which it is evident, from the lines of growth on the valves and sheath, that a new membrane is formed only at rather long intervals, and that it is formed in some degree extensible, so as to allow of some growth in the shell. Two or even three of these membranes are retained at the same time, one over the other; and thus, by their joint thickness, they afford protection to the included animal’s body, and compensate for the smallness of the opercular valves. In a large specimen of C. balænaris, two inches in diameter, there had been formed, since its existence as a very young shell, not more than eight opercular membranes, whereas the other membranes must have been moulted within this same period at least thirty times. In a young specimen of this species, having the orifice of the shell only two tenths of an inch in diameter, I found the opercular membrane, as usual, double. This membrane is not furnished with spines, nor is it penetrated by tubuli as in most other genera. The tissue left after the opercular valves have been dissolved in acid, presents no tubuli, or any other recognisable character, and is not divided (as is usual) into layers.
Muscles of the Sack.—These muscles differ considerably from those of other sessile cirripedes hitherto described. There are the usual five (or properly six, the two tergal muscles being here, as elsewhere, confluent) bundles of fasciæ; but they hardly can be called bundles, they are so[Pg 409] much spread out and thin. In C. balænaris, each rostral muscle consists, in different specimens, of from three to five principal fasciæ; these, at their upper ends, are attached to the cartilaginous plicæ at the rostral extremities of the scuta; at their lower ends, they do not reach (as in all the previous genera) to the basal membrane, but after converging, they diverge again into a little fan of fibres, which are firmly attached to the corium low down on the sides of the sack. The lateral depressores of the scuta consist each of about three fasciæ, and they terminate downwards like the rostral muscles. The tergal depressores are spread out into a thin sheet; upwards they reach to the basal edges of the lips of the sack-aperture, and downwards they curve a little towards the rostral and opposite end of the shell, and extend nearly to the basal membrane. These tergal muscles include two fasciæ, larger than the others, which extend rather further, both upwards and downwards, than the other fasciæ. But the most novel character in these several muscles is that in their lower portions they do not exhibit transverse striæ, thus showing a tendency to become involuntary as in pedunculated cirripedes. This circumstance, and their feebleness, is easily accounted for by the thick unyielding nature of the opercular membrane, and the feebly developed character of the opercular valves. In C. diadema, the tergal muscles are much spread out, having, as in C. balænaris, a larger fascia on each side; but the lateral depressores of the scuta form a well defined nearly cylindrical bundle; the rostral pair are extremely weak and spread out: I could perceive only feeble transverse striæ on some of these muscles, and on others there was not the least trace of striæ. I may add that the adductor scutorum muscle is well developed, as are the eight pairs of muscles which unite the animal’s body to the under surfaces of the scuta. The action of the adductor scutorum serves to close the sack-aperture, but towards the carinal end necessarily with very small force; protection in this part can only be afforded by the protuberant, valvular lips, and by the dorsal surfaces of the inwardly curled cirri, with their tufts of bristles.
Mouth.—The mouth (Pl. 26, figs. 3, 4) is much elongated[Pg 410] transversely, and does not differ essentially from that of most other Balanidæ. The labrum is notched and not in the least bullate, though in C. diadema there is a slight prominence outside close beneath the notch. The palpi are of large size, with their bristly apices touching each other. The mandibles are very strong, with from four to five main teeth, which are remarkable by presenting only rudiments of being laterally double; but between the second and third, and between the third and fourth main teeth, there is a small intermediate tooth,—these I have not met with in any sessile cirripede hitherto described. The maxillæ are small, with the two upper spines remarkably strong. The outer maxillæ are on their inner faces bilobed. Between these organs there is a minute projection, or mentum, flattened in the longitudinal axis of the body; I have not noticed this in any previous Cirripede.
The Cirri are short and extremely much flattened: the three anterior pairs have their rami unequal in length by two or three segments; the posterior edges of their pedicels are fringed by tufts of extremely fine hairs. The pedicel of the first cirrus is very broad; its rami are short, with the segments very broad. The rami of the second and third cirri are short, with the segments protuberant in front and thickly clothed with spines; the terminal segments have some short, thick, claw-like spines. The three posterior pairs have protuberant segments, each supporting three or four pairs of short, strong, main spines, with a small intermediate tuft: the dorsal tuft is small.
The prosoma is of large size. The stomach is large, without cæca, but with some internal longitudinal plaits (in C. balænaris at least), showing a tendency to the formation of cæca. In the stomach of C. balænaris I found a considerable quantity of a conferva, too much, I think, to have got in accidentally.
Generative system.—The vesiculæ seminales are large; and at their broad blunt ends, in C. balænaris, four separate vasa deferentia enter, of which fact I have seen no other instance. The ovarian tubes do not extend up the sides of the sack, but lie at the bottom, over the basal membrane; in C. balænaris they send six short ray-like prolongations[Pg 411] into the six sutures; in C. diadema they send similar prolongations into the sutures, and fill up, as I believe, the six chambers (Pl. 16, fig. 7, v) lying between the radii and alæ: I have examined only one specimen of C. diadema in spirits, and this had the ovarian tubes in an early state of development, when they can with difficulty be distinguished from the pulpy corium; the orange-coloured masses, however, which filled the six cavities, resembled the layer which certainly consisted of undeveloped ovarian tubes and cæca, lying over the basal membrane. The ova are wonderfully numerous; their length is 3/400ths of an inch. The larvæ have been noticed in the anatomical introduction.
Branchiæ.—These are immensely developed, covering almost four fifths of the area of each side of the sack. Each consists of two nearly equal folds, attached vertically to the carinal end of the sack, and transversely across the upper end extending to the animal’s body. The outline of the free part is rounded. Both folds of both branchiæ are deeply plicated on both sides; hence the superficies of the whole is very great. We shall find that this structure is common to the three following allied genera, but with these exceptions, I have observed double branchiæ only in one species of one other genus, namely, in Chthamalus dentatus.
Attachment.—The shells adhere with remarkable strength to the whale’s skin. Having, until recently, examined only separated specimens, and observing portions of the whale’s skin adhering to the outside, and solidly filling up the cavities on the under side, I did not doubt that the shell had the power of forming, by its own action, a deep cavity in the skin of the whale. Inspection of the basal outline of the walls of the shell (Pl. 16, fig. 5) of C. balænaris, will show how singularly unfitted its structure is for any burrowing process; and I was led to speculate on the possibility of the pupa being able to bury itself deeply in the skin, but rejected this view as opposed to what is known of the habits and structure of the pupæ of other Cirripedes. Having now examined several specimens of C. diadema adhering, in a group, to a large piece of skin, in Mr. Stutchbury’s collection, it has become evident that the[Pg 412] attachment is as much owing to the upward growth of the whale’s skin, as to the downward growth of the Coronula. In Pl. 15, fig. 4, a vertical section is given of the whale’s skin, taken through the place whence a shell of C. diadema has been removed; consequently we here see nothing but the whale’s skin: the upper black layer is the dark horn-coloured epidermis, forming the general surface of the whale’s body, and resting on an orange-coloured fibrous layer, which is lightly shaded in the drawing. The two horns in the section are two of the eighteen projections, formed entirely of the dark epidermic layer, which fill up the eighteen flattened cavities produced by the folding of the walls. Outside the horns we see the section of a circular furrow, in which the circumferential margin of the shell was lodged; and between the horns, there is the central hollow, within which, when lined with shell, the cirripede’s body was included. The circular furrow is formed in main part by the epidermic layer being thinner there than on either side, and partly by the orange-coloured, underlying fibrous layer curving a little downwards, from having apparently yielded to the pressure of the circumferential margin of the shell. With respect to the cause of the thinness of the epidermis under the circular furrow, I do not know how much to attribute to mere mechanical compression or stretching, and how much to the pressure of the shell, having checked[124] its formation. In the case of very young and small shells, it is hardly possible that their pressure can have in any way influenced the formation of new epidermic layers under the thick old layers; and we must believe, at least in these cases, that the whole effect is mechanical, the sharp basal edges of the shell having indented the epidermis; but this is not more surprising than that the radicle of a plant should penetrate hard ground. Whether the indented epidermis in the circular furrow becomes ruptured, I am not sure; ragged layers may commonly be observed outside the shell, but it is very possible that these may be the ends of layers of epidermis which have been preserved by the covering of the shell, whereas, on the[Pg 413] surrounding parts of the whale’s body, these same layers have been removed by disintegration. To return to the section,—the outline of the boss of orange-coloured fibrous tissue, under the central hollow, clearly shows that it must have been formed by its own upward growth, for it stands above the general surrounding level of the corresponding layer. This same conclusion is still more obvious with respect to the eighteen flattened prominent horns, formed of the dark epidermis; the manner in which the epidermis has been forced, moulded, and packed into the eighteen flattened and curved cavities of the shell, so as to adhere to them with considerable tenacity, is extremely curious. The prominence of these horns is so great that it appears to me quite impossible to account for them, excepting by a special formation of epidermis beneath each cavity. The basal membrane of the Coronula, which lies at the bottom of the central hollow, adheres by its own cementing apparatus; and when the larva first attaches itself, this adhesion must be very important, as it allows the basal edges of the shell, during their slow downward growth, to press firmly on the whale’s skin, and so slowly indent it with the circular furrow. The final cause, probably, of the cavities on the under side of the shell in this genus, formed by the singularly convoluted parietes, is to allow of the upward growth into them of the epidermis of the whale, thus securing a firm attachment and allowing the shell to exert a strong downward pressure, and thus effect its partial imbedment, and protection from the enormous force of the waves to which it must be exposed.
[124] Formerly I was inclined to believe, as stated in my former volume on the Lepadidæ, that the cement injured the true skin of the supporting animal, but this, at least in such cases as the present, I do not now at all believe.
With respect to C. balænaris, I have seen only specimens, preserved on shrunk and twisted whale’s skin, with the underlying fibrous layer not preserved; but the cavities in the shell were filled by horns of epidermis, exactly as in C. diadema. There is, however, this difference in the attachment of the two species, that in C. balænaris, owing to its depressed form, the circumference of the shell indents the whale’s skin, not vertically downwards, as in C. diadema, but very obliquely outwards; and, consequently, buries itself much more completely, but less deeply, under a folded and apparently ruptured flap of the epidermis. In young[Pg 414] specimens, of the size of a shilling, the entire shell, with the exception of the operculum, is thus covered up and protected whilst young and tender.
Geographical Distribution.—The genus is found wherever whales occur, that is, from the Arctic to the Equatorial regions, in both hemispheres. It is asserted that sometimes as many as a couple of hundred specimens will adhere to a single whale. Coronula barbara, a form closely allied to C. diadema, existed during the Red Crag period; and Bronn has described some fossil specimens from Italy.
Affinities.—In the wonderfully convoluted shell,—in the parietal tubes not being either filled up by calcareous layers or being crossed by calcareous septa,—in the outer lamina of the shell between the longitudinal septa near the basis being imperfect, Coronula differs from all the foregoing genera; in the two latter respects it agrees with the three following allied genera, viz., Platylepas, Tubicinella, and Xenobalanus. The equal size of all six compartments of the shell, has been observed in very few genera besides Coronula. In a new opercular membrane not being formed at each exuviation, and in two or three of these membranes being persistent, and in their being attached high up the sheath, this genus agrees with the three following genera alone. In the valves tending to be rudimentary, and in the protuberant lips of the sack-aperture, we have a close alliance with Xenobalanus. The muscles of the sack being spread out, and tending to lose their transversely striated character, are great peculiarities in Coronula, Tubicinella, and Xenobalanus. The simplicity of the cement-ducts is a remarkable character, observed in two of the following genera, but not in Tubicinella. The double branchiæ is a peculiarity common to all four genera. Neither the mouth nor cirri offer any new characters of much importance in Coronula or in the three allied genera: the lower teeth of the mandibles not being laterally double, but having small intermediate teeth, is the newest feature in the mouth.
LEPAS BALÆNARIS. Gmelin. Systema Naturæ (1789).
---- ------ Chemnitz. Conch., vol. 8, Tab. 99, fig. 845, 846 (1785).
BALANUS ------ Bruguière. Encyclop. Method., Pl. 164, fig. 13-18 (1789).
CORONULA ------ Lamarck. Annales du Museum, vol. 1, Tab. 30, fig. 2-4 (1802).
-------- ------ Chenu. Illust. Conch., Plate, fig. 1 and 4.
-------- ------ De Blainville. Dict. des Scien. Nat., 1818 and 1824, Tab. 117, fig. 3, 3 a.
Shell much depressed, with longitudinal flattened ribs, having simple edges; orifice rounded-hexagonal: radii very thick, almost equalling the shell in thickness: opercular valves four.
Hab.—Attached to whales in the Southern Ocean.
Having described, under the genus, in so much detail the structure of the whole shell, it will be sufficient here to point out the characters by which this species differs from the others.
General Appearance.—The shell is generally much depressed, though sometimes, in large specimens, from the turning in of the basal edges, a considerable degree of convexity is acquired. The radii are moderately wide, and give a star-like appearance to the shell. The surface of the whole upper part of the shell is smooth; the broad and much flattened ribs (i. e. the terminal, transverse loops of the folded walls) of which each compartment is formed, generally divide at a short distance from the apex. The close sutures separating these ribs are straight, and not finely serrated, by which character alone this species can at once be discriminated from the others. The surface, when closely examined, is found to be finely striated longitudinally, and is transversely crossed, chiefly in the lower part, by minutely beaded growth-ridges. The orifice is rounded-hexagonal, and is small compared with the whole shell; it is also smaller than the basal edge of the internal cavity of the shell; consequently the whole basal edge cannot be seen through the orifice from one point of view,—the operculum of course having been removed. An unusually large specimen was 2.75 of an inch in[Pg 416] diameter; this individual was one of the more convex varieties, and yet its entire thickness, from top to base, was only .9 of an inch.
Operculum.—Having described, under the genus, the general structure of the opercular valve, I will here only add a few details on their shape. They are small compared with the whole extent of the opercular membrane. The scuta (Pl. 15, fig. 2 b, scutum to the left) stand near each other at the rostral end of the aperture leading into the sack, with their two rostral ends united by the yellowish-brown, longitudinally plicated, horny membrane, described under the genus. The terga (fig. 2 b) stand a little apart from the scuta, on the sides of nearly the middle of the sack-aperture. The lips are protuberant and moderately developed all round. The scuta are elongated, and a little curved; including the upper imbedded portion, they are almost sub-triangular: but the under growing surface is much elongated, nearly flat, with the two ends of nearly the same width, truncated and rounded. The Tergum, in rather small specimens is, if the upper imbedded portion be included, sub-triangular, with the growing surface oval, and between one third and one half of the length of the scutum; but in large specimens, the tergum becomes style-formed, lying parallel to the tergal margin of the scutum, with the growing surface proportionally much smaller, and not above one fifth of the length of the growing surface of the scutum. Hence we see some tendency in the tergum to become rudimentary, as it is in the next species. The brown, horny, plicated substance in which the terga are imbedded, extends considerably beyond the valves themselves.
Structure of the Shell and Radii.—I have already so fully discussed this subject, that I will here only enumerate the points in which this species differs from C. diadema and reginæ:—Firstly, the more symmetrically folded walls (Pl. 16, fig. 5), new folds arising on both sides of all six sutures. Secondly, the inner ends of the folded walls, which surround the internal cavity, being almost square, but with their angles rounded; their inner ends descend some little way beneath the basal edge of the sheath, as low, or lower, than the circumference of the shell. Thirdly, the external lines of junction between the transverse terminal loops being smooth or not serrated. Fourthly, these loops being flatter towards the outside, and being internally filled by septa (Pl. 15, fig. 2 a); tubes being thus formed larger than the proper parietal pores. Sixthly, the sutural edges of the compound radii (Pl. 16, fig. 3) being very much broader in the inner and upper part of each compartment than in the outer and lower part; for in the inner and upper part they stretch from the outside of the shell to the sheath, so that the alæ rest on them, and hence no large open cavity is left between the radii and alæ; in the inner and upper part, also, the radii extend down nearly to the basal edges of the folded walls. The septa, of which the radii are formed, stand further apart than in C. diadema and reginæ. Seventhly, the alæ are only 1/5th or 1/6th of the thickness of the radii, whereas in the two other species they are very much thicker, being nearly as thick as the radii: the alæ are also here (fig. 3, a′) squarer than in those two species, that is, their basal margins are not so short compared with their upper margins; their edges present, also, a slightly different structure. Eighthly, the basal edge of the[Pg 417] sheath does not here project freely, and does not descend quite so close to the basal membrane.
Mouth.—The palpi are furnished with a row, extending along the whole basal exterior margin, of very long upwardly pointing spines; on their upper edges there is a brush of small spines. The edge of the labrum is clothed with extremely fine spines, and is furnished with a few minute teeth. The mandibles have four teeth, and the lower end is broadly rounded and coarsely spinose: the second and third teeth are double at their points, and between the second and third, and again between the third and fourth, there is a small intermediate tooth.
Regarding the Cirri, I have nothing especial to remark.
Geographical Distribution.—I have examined nine sets of specimens, having localities attached to them: three from off New South Wales; two off the Cape of Good Hope; two from the west coast of South America; and two marked only “South Sea.” Hence I am led to conclude that this species is confined to the southern hemisphere, or if it extends into the northern hemisphere, it is probably only in the Pacific Ocean. I do not believe (though so stated in some works) that this species occurs on the shores of Europe. It seems often to be associated with Tubicinella. Some specimens thus associated, sent by Mr. Bennett to Professor Owen, were said to have been attached to the Balæna australis.
LEPAS DIADEMA. Linn. Systema Naturæ, 1767.
---- ---- Chemnitz. Conch., vol. 8, Tab. 99, figs. 843, 844.
BALANUS DIADEMA. Bruguière. Encyclop. Method., n. 164, fig. 13, 14 (1789).
CORONULA ------ De Blainville. Dict. des Sc. Nat. (1824), Tab. 117, fig. 4.
-------- ------ Leach. Encyclop. Brit. Suppl., vol. iii, 1824.
-------- ------ Chenu. Illust. Conch., Plate, fig. 3.
-------- ------ Burmeister. Beiträge zur Naturgeschichte der Rankenfüsser, 1834, Tab. 2, fig. 1-14, 18.
Shell crown-shaped, with longitudinal convex ribs, having their edges crenated; orifice hexagonal: radii moderately thick, very broad: terga absent or rudimentary.
Hab.—Attached to whales, in the Arctic Seas; United States and Great Britain; Gulf-Stream, Atlantic Ocean; New Zealand(?).
General Appearance.—As previously remarked, owing to the fulness of the generic description, minute details on structure, excepting those[Pg 418] characteristic of the present species, need not here be given. The crown-like shape of the shell is well expressed by its name of Diadema, but the crown tends to pass into a cylinder. The radii are extremely broad. The orifice is large, and neatly hexagonal: when the operculum is removed the whole inside of the cup-formed shell can be seen at once, for the flat membranous basis is much smaller than the orifice. The under side of the shell is deeply concave. The outside of each compartment is formed by broad, rounded, and somewhat prominent, rarely divided, ribs (i. e. the transverse ends of the folded walls); these ribs are closely united together by finely serrated lines of junction (Pl. 16, fig. 1, f). Their surfaces outside are delicately striated longitudinally, and plainly crossed (more plainly than in the foregoing and the next species) by irregular, transverse ridges, especially in the lower part of the shell. The largest specimen which I have seen was two and a half inches in diameter and two in height.
Scuta.—These are placed close together at the rostral end of the orifice, and are imbedded in the brownish, tough, longitudinally plicated, horny substance, which extends far beyond both ends of the valves. In outline (fig. 3 b) they are mitre-shaped, or rounded and sub-triangular, a little curved, and more or less elongated, being most so in young specimens; they are, however, less elongated and rather more massive than in C. balænaris. Terga,—these seem entirely absent in most specimens; but in one (fig. 3 a) I found a rudiment, namely, a short thin plate of shell, barely visible to the naked eye, extending parallel and near to the tergal margin of the scutum. The lips of the aperture of the sack are prominent, and highly so towards the carinal end.
Structure of Shell and Radii.—Owing to the shell not spreading much at the base, new folds in the walls are much seldomer formed, and therefore the external longitudinal ribs (i. e. the terminal transversely elongated loops), are much seldomer divided, than in C. balænaris or reginæ; even rather large specimens sometimes having only the original eighteen folds.[125] When new folds are formed, only one is formed on one, viz., the rostral, side of each suture, instead of on both sides, as in C. balænaris. The inner ends of the folded walls, surrounding the basal membrane, are narrow, instead of being almost square, as in C. balænaris. The lower edge of the sheath, which projects freely, descends almost to the level of the basal edges of the walls. The outer ends of the folded walls, forming the transverse loops, are internally filled up solidly by calcareous matter, instead of by septa forming tubes, as in C. balænaris. The radii (Pl. 16, fig. 2) are a little thicker in the lower and outer than in the upper and inner part of each compartment; in the middle, they do not reach the sheath by[Pg 419] about half the thickness of the compartment, and consequently they are separated from the plates (c′′ in fig. 1 and 7) on which the alæ rest, by large chambers (v in fig. 7), which extend up to nearly the apices of the compartments: the extent, however, to which the upper ends of these chambers have been solidly filled up, varies a little. The sinuous plates forming the main portion of the compound radii are rather thinner and closer together than in C. balænaris. The alæ are thick, being thickest in the middle part, and there equal the radii in thickness; their lower margins are very short compared with their upper margins, hence they are almost wedge-formed.
[125] These should be counted on the under side of the shell, for if counted from the outside, the number would be thought to be twenty-four, as on the side of each compartment bordering the radius, a half-fold has the appearance of being a whole fold, so that in appearance six folds are added to the real number of the folds. This caution is necessary whatever the number of real folds may be, that is, whether or not the original eighteen folds have been increased by subsequent plications. These remarks, also, are applicable to the other species.
Mouth.—The teeth and fine hairs on the labrum are sometimes obscure, and sometimes plain: close outside the bottom of the medial notch, there is a small hard prominence. The palpi are broad; on their basal exterior margins there is a short row of spines, which do not equal in length the width of the palpi, and therefore are not so long as in C. balænaris. The mandibles have five main teeth, of which the second and third show only an obscure rudiment of being double; between these two teeth, and between the third and fourth tooth, there is a small intermediate tooth: the inferior angle is narrow, rounded, and spinose. The extremity of the apodeme of the maxillæ is expanded.
Cirri.—I have only to remark, that the pedicel of the first cirrus is extremely broad, and that the rami are set on in an unusually crooked manner: the basal segment of the shorter and broader ramus of this cirrus has its dorsal surface produced into a plate fringed with very fine hairs.
Geographical Distribution.—I have received only four specimens with certain localities attached to them: namely, the Arctic Seas of Scandinavia; the coast of the United States, and of Britain; and the Gulf-Stream. There is also a specimen in the British Museum, sent by Mr. Stephenson, mingled with shells of Mollusca from New Zealand; but a Coronula, procured from a floating whale in the early part of the outward voyage, might so easily be sent home with specimens subsequently collected in another county, that I do not as yet fully admit that this species is an inhabitant of the Southern Pacific Ocean: I am less willing to admit this, from suspecting that C. reginæ in the Pacific, replaces the C. diadema of our Northern Seas.
Shell globulo-conical or depressed, with longitudinal, much flattened ribs, having their edges crenated, and their surfaces striated and granulated; orifice hexagonal: radii thin, not exceeding one fifth of the thickness of a compartment: terga absent.
Hab.—Attached to whales, Pacific Ocean; Mus. Cuming, Stutchbury, and Darwin.
I have seen eight specimens of this form—two in Mr. Cuming’s collection, one of which was procured at Iquique in Peru, the other from an unknown locality; one was given me in Chile, and was stated to have come from the Pacific; two others are in Mr. Stutchbury’s collection, from unknown but distinct localities, and three in a group in the British Museum. These eight specimens, of which six were collected separately, all strikingly agree in general aspect and in details of structure, so that I can easily recognise the shell, and can at once pick out a single compartment, when mingled with those of the two foregoing species; nevertheless, the differences are so small, that I at first hesitated whether to name the species; but, upon reflection, I am convinced that it is distinct. Coronula reginæ is much more closely related to C. diadema than to C. balænaris, and I suspect that it replaces in the Pacific the former of these two species.
General Appearance; shell conical, straight-sided; some specimens being nearly as much depressed as C. balænaris, and considerably more depressed than any variety of C. diadema; other specimens being globulo-conical, but rather less globular than the ordinary form of C. diadema. I may here remark, that shape is of more value in this genus, in which the shell is attached to the yielding skin of whales, than in those genera in which it adheres to rocks. The orifice is neatly hexagonal, and the whole internal cavity, when the opercular membrane is removed, can be seen from one point of view, owing to the contraction of the lower part of the cavity and small size of the basal membrane. The surface of the shell is smooth, but has, even up to the summit, a peculiar frosted appearance, different from that of the foregoing species, caused by the longitudinal striæ being rather more distinct, and being crossed by beaded, very delicate transverse lines of growth. The ends of the transverse loops, forming the exterior surfaces of the compartments, are much flattened, even more so than in C. balænaris. The lines of junction between the loops, are finely serrated, as in C. diadema; and internally they are solidly filled up, instead of being formed into a set of tubes by longitudinal septa, as in C. balænaris. The under side of the shell, with its folded walls, presents an appearance intermediate between the variable appearance of this part in the above two species.
The sutural edges of the radii (Pl. 16, fig. 4) offer by far the most remarkable character, in their thinness from top to bottom; for they hardly exceed one fifth of the thickness of the compartment, measured from the external surface to the base of the sheath; hence a very large cavity is left between the radii and alæ: in the thickness of the radii the three species already described form a series, C. reginæ at one[Pg 421] end and C. balænaris at the other. The sinuous plates, forming the lower part of the radius, are coarser and stand rather further apart than in C. diadema. The alæ are thick, and have the same outline, being narrow at their basal margins and broad at top, as in C. diadema, with their sutural edges similarly constructed: the basal edge of the sheath likewise projects freely.
Operculum.—This resembles most closely that of C. diadema. There are no rudiments of terga. The scuta cannot be distinguished from those of C. diadema.
Mouth.—The labrum has a row of inwardly curved little teeth along the whole crest, and these I did not notice in C. diadema: there is only a trace of the prominence on the outside at the bottom of the central notch. The hairs on the basal exterior margin of the palpi are moderately long. The mandibles have five teeth. In the Cirri, the first pair resembles in its peculiar structure those of C. diadema. In the sixth pair, however, the segments support only three pairs of main spines: but the specimen was not very large, and probably in old specimens there would have been four pairs.
Summary.—The present species differs from both the foregoing only in its rather more conical and straight-sided outline, smooth, frosted surface, and in the narrowness of the sutural edges of the radii, and consequent large size of the chambers between the radii and alæ. It resembles C. diadema, as far as the shell is concerned, in the external ribs or transverse loops having their lines of junction serrated and in being solidly filled up—in the shape of the orifice and of the internal cavity of the shell—in the shape and structure of the alæ—and in the basal edge of the sheath being free. It comes nearer to C. diadema than to C. balænaris in the structure of the radii. It differs from C. diadema, and comes nearer to C. balænaris, in the external ribs being flattened, instead of being convex, and in the lines of growth being very delicate. But as it resembles C. diadema in the several foregoing characters of its shell, in the opercular valves, in all parts of the mouth (excepting the labrum), and in the cirri, it is very much more nearly related to that species than to C. balænaris.
CORONULITES DIADEMA (?) Parkinson, Organic Remains (1811), vol. 3, p. 240, Pl. 16, fig. 19.
Shell (probably) crown-shaped, with longitudinal convex ribs, having their edges crenated, and their surfaces rugged, both externally and internally, with transverse ridges: radii moderately thick; the spaces between the radii and the alæ solidly filled up.
Fossil in Red Crag, (Bawdsey and Sutton); Mus. S. Wood and Geological Society.
The species here described, though near to C. diadema and easily confounded with it, I have no doubt is distinct. I owe to the kindness of the Rev. Mr. Image an examination of the original specimen figured by Parkinson; and in Mr. Stutchbury’s collection there is a similar and more perfect specimen; both of these resemble C. diadema in general form, but have been too much worn to be positively identified. The following description is drawn up from some compartments collected by Mr. Searles Wood, belonging certainly to three, and probably to four individuals, one of which was young; as these specimens agree in all essential respects, I feel pretty confident that the characters, by which the present species differs from C. diadema, are of specific value.
Structure of Shell.—The longitudinal ribs on each compartment (i. e. the terminal transverse loops), are convex and prominent, as in C. diadema, but they are crossed by more prominent ridges of growth than even in the roughest varieties of that species, so that the surface of the shell is more rugged. In the three previous species, the surface of the wall entirely round the cavities occupied by the whale’s skin, is striated only by longitudinal very fine lines; but here, the outer portion, or that (fig. 6) formed by each transverse loop, is crossed by transverse ridges of growth, like, but less prominent than, those on the external surface of the shell. The minute teeth, along the lines of junction between the transverse loops, are here less regular, and can hardly be said to exist; for the two edges are locked together by what may be more strictly called minute zig-zag ridges than teeth. The exact number of the circumferential plications in the walls is variable in the same manner as in the three foregoing species. The sutural edges of the radii are about as thick as, or rather thicker than, in C. diadema. As in this latter species, and in C. reginæ, each ala rests, not on the internal surface (as in C. balænaris, and in all other Balanidæ) of the radius, but on a special plate; but in C. barbara, instead of a deep chamber, running up to the apex of the compartment, being left between the radius and ala, this part is filled up almost entirely by solid shell. Although the extent to which this chamber is filled up varies a little, and although its depth varies a little in C. diadema, yet there is a marked difference between the specimens of this latter species, in which the chamber is most filled up, and those of C. barbara, in which it is least filled up. The alæ are thick, as in C. diadema, and their sutural edges have a central ridge, sending off on both sides sinuous ridges. The basal margins of the alæ are not short compared with their upper margins, and therefore the whole ala is not wedge-formed; and in this rather important respect C. barbara resembles C. balænaris (Pl. 16, fig. 3), and differs from C. diadema[Pg 423] (fig. 2). The lower edge of the sheath does not seem to have projected freely,—in this respect, also, resembling C. balænaris. From the basal margin of the alæ not being narrow, and from the inner ends of the folded walls being, as it would appear, also broad, I have little doubt that the cavity in which the animal’s body was lodged, resembled in shape that in C. balænaris, the membranous basis being larger than the orifice of the shell.
Opercular valves unknown.
Summary.—This species is more nearly related to C. diadema than to the others; but in some points, just specified, it resembles C. balænaris. The characters by which it differs from all the species are, firstly, the more prominent transverse ridges on the external surface of the shell, and more especially on the surfaces bounding the outer sides of the cavities occupied by the epidermis of the whale. Secondly, the character of the teeth, or rather ridges, along the lines of junction between the transverse loops. And, thirdly, the spaces between the radii and the plates on which the alæ rest, being solidly filled up.
The Coronula bifida is an Italian tertiary species, so named by Bronn, in his “Italiens Tertiär-Gebilde” (1831), p. 126. It is very possible that this may be identical with C. barbara, but Bronn does not seem to have been aware of the absolute necessity of giving minute details in his descriptions of fossil cirripedes. The chief character of C. bifida is thus given:—“Eine tiefe Furche oder Spalte theilt die Längenrippe von oben herab bis zur Hälfte, welche bei der sonst ähnlichen C. diadema entweder ganz fehlt, oder nur zuweilen kurz angedeutet ist.” Had it been stated that the longitudinal ribs were divided from the middle down to the base, instead of from the top to the middle, the description would have been intelligible to me, though the character thus afforded would not have been of specific value, as this dividing of the ribs occasionally occurs in all four species, and is produced by the formation of new folds to the walls.
PLATYLEPAS. J. E. Gray. Annals of Philosophy, (new series), vol. x (1825).
CORONULA. De Blainville. Dict. des Sciences Nat. (1824).
COLUMELLINA. Bivona (fide Philippi). Nuovi generi, &c. di Molluschi, Palermo (1832).
Compartments six, each bilobed and inwardly produced, so as to form six midribs, which support the outwardly convex, membranous basis.
Distribution, throughout the tropical and warmer temperate seas. Imbedded in turtles, sea-snakes and manatee.
This small genus, consisting of three species (though I have named only two), is a very natural one, yet closely allied to Coronula. Most authors have united these genera, but in doing so they destroy two very natural little groups. Platylepas, moreover, when all its characters are considered, has as good a claim to be generically separated from Coronula, as has Tubicinella,—a genus universally admitted. I shall presently recur to this subject.
General Appearance.—The most remarkable character, and which gives a peculiar aspect to the whole shell, is that each of the six compartments has a medial fissure or fold; the shell thus consisting of twelve lobes. Each fold extends from the summit to the base of the shell, and is produced inwards in the shape of a midrib (Pl. 17, fig. 1 a). The membranous basis is supported by the basal edges of the six midribs, and is thus rendered convex. As the midrib on the rostrum would have interfered, if fully developed, with that part of the animal’s thorax which I have called the prosoma, it is of less size than the other midribs; and from this same cause the whole rostrum is pushed a little on one side, generally to the left, so that the shell is not perfectly symmetrical. The midrib of the carina is less than the lateral midribs. The orifice is oval, and is more or less plainly indented by six angular points,—the worn-down summits of the six midribs. The narrow elongated opercular valves stretch from[Pg 425] one end of the orifice to the other. The rostrum is much broader than the carina: the lateral compartments are but little broader than the carino-lateral compartments. The radii are narrow, though variable in width. The general shape of the shell varies much in the same species, being either much depressed, or rather steeply conical; either circular, or more commonly oval. The (so called) epidermis is usually persistent in the lower part of the shell. The shell itself is white. The basal diameter of the largest specimen of P. bissexlobata was nearly three-quarters of an inch.
Structure of the Parietes, Radii, &c.—The walls of the shell in P. bissexlobata are permeated by minute pores, and have exactly the same structure as in Coronula; the pores being completed by the union of ledges on the outer sides of the longitudinal septa, and the latter have little knobs formed on them at each period of growth. In P. decorata the walls are solid. Each compartment, as already stated, has a deep nearly medial furrow, the sides of which are closely pressed together, thus forming the midribs, and thus differing from the furrows in Coronula, which not being pressed together, form cavities on the under side of the shell. The inner longitudinal margin of each midrib is thickly coated by a layer of shell, and is thus rendered much more prominent than it would have been if formed exclusively by the inward folding of the wall. The upper part of each midrib forms a slight, longitudinal ridge (fig. 1 d) on the sheath, having become encased by the sheath during its downward growth. Owing to the small size of the midrib of the rostrum, there is scarcely any ridge on the sheath of this compartment. The sheath does not descend half-way down the walls in P. bissexlobata, but further down in P. decorata; but perhaps the thickened inner margins of the midribs may be considered as the downward prolongations of the sheath, and on this view the sheath descends actually to the basal membrane. The radii are narrow: their edges have simple septa, with the interspaces filled up solidly. The alæ have their sutural edges smooth.
The Basis is membranous, and more or less convex. This convexity is rendered possible by the support afforded[Pg 426] by the basal edges of the six midribs. The so-called epidermis or membrane which covers the shell, is prolonged in six flaps under the midribs; and hence the proper basal membrane has six deep indentations, receiving the above flaps. The cementing apparatus is constructed on the same simple type as in Coronula, as has been described in the Introduction.
Scuta and Terga.—These are narrow and long, stretching the whole length of the orifice of the shell. The two valves touch each other, but are not properly articulated together: their upper surfaces scale off in layers: they resemble each other very closely in shape (fig. 1 c), being simply oblong. The membrane covering the valves supports some very minute spines, 2/1000th of an inch in length. When a portion of valve is dissolved in acid it is seen to be penetrated by very numerous tubuli. The opercular membrane is rather thick: in P. decorata I found it occasionally double; in this species it supports some very minute spines; it is attached about one third down the sheath, of which the upper part is not marked (as in Balanus) by regular concentric rings. The aperture between the valves, leading into the sack, has protuberant lips, as in Coronula and Tubicinella. The normal five bundles of muscles surrounding the sack, differently from in the two genera just mentioned, do not thin out and spread out downwards, but retaining their full thickness, reach the basal membrane, to which they are attached.
Mouth: the parts present no difference from those in Coronula; between the outer maxillæ we have the same small flattened mentum-like projection; and in the mandibles, the same small intermediate teeth between the second and third, and between the third and fourth main teeth, as in that genus. The main teeth here are more plainly double laterally. There are teeth on each side of the central notch of the labrum. The cirri differ little from those of Coronula, but the segments are not nearly so much flattened and broad, and the second and third pairs are not so short. The segments of the sixth pair support four pairs of main spines, with a few minute intermediate spines, but with no intermediate tuft.
Branchiæ: these consist on each side (at least in P. decorata) of a double fold, much less plicated than in Coronula; the fold nearest to the animal’s body is rounder, and not quite so large as the outer fold.
Range and Habits.—The P. bissexlobata ranges from the Mediterranean and west coast of Africa to the West Indies, to the north-east coast of Australia, and, judging from the name given by Chenu, to California: the second species, P. decorata, inhabits the Pacific Ocean; and the third imperfectly known species, was deeply buried in the skin of a sea-snake, off Borneo. The P. bissexlobata is attached both to turtles and to the skin of the manatee: some specimens in the British Museum, attached to the latter, from the west coast of Africa, were entirely buried in the skin, with the exception of the operculum. The specimens of P. decorata, which I obtained at the Galapagos Archipelago, were buried about half their depth in the softer parts of the skin of the green turtle. I do not doubt that the imbedment is effected in the same manner as in the case of Chelonobia.
Affinities.—This genus, in general structure and habits, approaches closely to Coronula; but it differs from all the species of that genus in the following respects, which appears to me fully to justify its generic separation:—The outline is often oval, instead of circular; the compartments are not of equal sizes and shapes; each compartment has here only one inward furrow or fold, and this has its sides pressed closely together, and is encased, but not obliterated, by the sheath; the rostrum is not quite medial; and the radii are narrow and nearly solid. The opercular valves, again, are here more developed, and their microscopical structure is different. The opercular membrane is less constantly double; and, what is much more important, the muscles of the sack are not here spread out, and show no tendency to become rudimentary and lose their transverse striæ. The cirri differ only in not being so broad and flattened. The branchiæ are not quite so much developed. Finally, I may add, that in several of the above respects in which Platylepas differs from Coronula, it approaches ordinary sessile cirripedes.
CORONULA BISSEXLOBATA. De Blainville. Dict. des Sciences Naturelles, tom. 32, (1824), Tab. 117, fig. 1.
PLATYLEPAS PULCHRA. J. E. Gray (!). Annals of Philosophy, (new series), vol. 10, (1825).
COLUMELLINA BISSEXLOBATA. Bivona (fide Philippi). Nuovi generi di Mollusch. (1832), Tab. 3, fig. 1.[126]
CORONULA CALIFORNIENSIS. Chenu (!). Illust. Conch., Tab. 1, fig. 4.
[126] This memoir was published in the ‘Effemeridi Scientifiche e Litt. per la Sicilia,’ according to the ‘Bibliographia Zoologiæ et Geologiæ,’ by Agassiz and Strickland.
Shell with the transverse lines of growth conspicuous: parietes permeated by pores; sheath descending barely half-way down the parietes.
Hab.—Mediterranean, attached to turtles. River Gambia, attached to manatee. Honduras, attached to manatee. Moreton Bay, lat. 27° S., Australia, apparently attached to the dugong of that coast. California(?) Mus. Brit., Stutchbury, and Cuming.
General Appearance.—Shell generally much depressed, and broadly oval or circular; sometimes steeply conical. Orifice oval, generally not large. Surface rather plainly marked by closely approximate lines of growth, which with the lobed outline gives to the whole an elegantly sculptured appearance: occasionally the longitudinal ridges formed by the parietal septa are distinct. Basal diameter of largest specimen three quarters of an inch.
Structure of Shell.—I have nothing material to add to the generic description. The midribs are not so prominent as in the following species, and hence the basal membrane is less convex. The origin of the midrib as a fold is very plain. The sheath descends barely half-way down the walls, and is a little hollow on its under margin, on each side of the midrib. Beneath the sheath the parietes are finely ribbed (fig. 1 d), but to a variable degree. I may here remark, that in the specimens taken from manatee, on the coast of Africa and at Honduras, the internal ribs extended further up and were plainer, and the opercular valves seemed to be a little narrower than in the other specimens, so that I at first suspected that they were specifically distinct, but I could make out no other than these small and variable points of difference.
Scuta: oblong (fig. 1 c), about twice as long as broad, with the[Pg 429] rostral end rounded, rather narrower than the other end, and curled a little inwards. Terga, of nearly the same shape and nearly as long as the scuta: the carinal end is rather more pointed than the scutal end of the valve, and when viewed internally, the growing surface of this end is seen to be bluntly pointed. In both valves the upper layers of shell usually scale off.
Mouth and Cirri. The only differences, compared with the following species, that I perceived, were that the hairs at the end of the palpi were here rather longer, and the fourth tooth in the mandibles rather less distinct. The rami of the first cirrus are very unequal in length.
Shell with fine longitudinal ridges, ornamented in the lower part by minute beads; parietes not porose: basal membrane equalling in convexity the shell.
Hab.—Pacific Ocean; Galapagos Archipelago; Lord Hood’s Island, Low Archipelago; Mus. Cuming, Darwin.
General Appearance and Structure.—Shell oval, with the orifice large. The walls are thick, and of less height from top to base than in the last species. The surface (and this is the chief external character) is marked by fine longitudinal ridges, each of which, when closely examined, is found to be double (fig. 2 b), and near the basis, where not abraded, to be ornamented with minute beads on each side: these beads are largest on those ridges which border the sutures. The parietes are not permeated by pores; on their inner surfaces there are a few rather prominent but short ridges, instead of the many finer ridges, as in P. bissexlobata. The sheath is of great thickness; in young specimens layers scale off its summit, all round the shell, as in Tubicinella; it is only slightly hollowed out at its lower margin; it descends more than half-way down the walls. The midribs, formed by the folded walls, are more prominent than in the last species, and descend lower down; hence the basal membrane is more convex than in the last species, for it projects downwards as much as the shell projects upwards. In medium-sized specimens, the midribs project inwards to a distance exceeding half the breadth of a compartment. A less proportional length of the midrib, in an inward direction, is formed by the inward folding of the wall, and a greater length by the thickening of its inner longitudinal margin, than in the last species. The basal edge of the inner end of the midrib is smooth.
Dimensions.—The average size of the numerous specimens which I obtained at the Galapagos Islands was about .2 of an inch in diameter, and I found none larger; but Mr. Cuming’s specimens from Lord Hood’s Island are half an inch in diameter.
Opercular Valves.—These closely resemble those of the former[Pg 430] species, with the exception that the carinal end of the growing or under surface of the Tergum is much squarer.
Cirri.—The only difference which I could perceive was, that the rami of the first pair were not quite so unequal in length.
Hab.—Imbedded in the skin of a sea-snake, taken off Borneo.
I am indebted to Dr. Gray for a single specimen of this supposed species, but as it is very young and imperfect, wanting the opercular valves and cirri, I do not choose to name it. The shell presents all the usual characters of the genus; the rostrum, I may remark, being pushed to the left side. The parietes are permeated by pores of considerable size, which shows that the species is distinct from P. decorata. On the inner basal surfaces of the walls, there are two or three very distinct ridges on each side of the midrib; and this fact, together with the size of the parietal pores, makes me suspect that it is not an immature specimen of P. bissexlobata.
TUBICINELLA. Lamarck. Annales du Museum, tom. 1 (1802), Tab. 30, fig. 1.
CORONULA. De Blainville. Dict. des Sciences Naturelles, (1824).
Compartments six, of equal sizes; shell sub-cylindrical, wider at the top than at the basis, belted by several large transverse ridges.
Hab.—Southern Pacific Ocean, Western South America, New South Wales, Cape of Good Hope; imbedded in whales, and often associated with Coronula balænaris.
This genus is closely allied to Coronula, and perhaps De Blainville was right in uniting them. But Coronula, as it now stands, is so natural a genus, that it seems a pity that a[Pg 431] form so entirely different in general aspect, as Tubicinella, should be forced into it. The main difference between these genera consists in the walls being here not folded, in the simpler radii, and in the general shape of the shell and of the included animal’s body; but there are many other minor points of difference. The most novel character in Tubicinella consists in the shell being lined almost close down to the basis by the opercular membrane. In the opercular membrane thus forming a long tube, and in the general shape of the animal’s body, we shall presently see that Tubicinella is closely related to Xenobalanus. In numerous other respects Tubicinella is almost equally allied to the latter genus, to Platylepas, and to Coronula. Finally, several points of structure indicate that Tubicinella may be considered as a Coronula, with the shell much simplified in structure.
LEPAS TRACHEALIS. Shaw. Nat. Miscell. (1789-1813), vol. 17 (1806?) tab. 726.
---- TRACHEÆFORMIS. Wood. General Conch. (1815), tab. 4, fig. 1-3.
TUBICINELLA TRACHEALIS. J. E. Gray. Annals of Philosophy, (new series), vol. 10, (1825.)
---------- MAJOR ET MINUS. Lamarck.[127] Annales du Mus. Nat., tom. 1 (1802), Tab. 30, fig. 1-2.
---------- BALÆNARUM. Lamarck. Animaux sans Vertèbres, (1818).
---------- ---------- Chenu. Illust. Conch. (Plate).
---------- ---------- Sowerby. Genera of Recent and Fossil Shells (Plate).
---------- LAMARCKII. Leach. Encyclop. Brit. Suppl., vol. 3 (1824), Pl. 57.
CORONULA TUBICINELLA. De Blainville. Dict. des Sciences Nat., tom. 32, Pl. 117, fig. 5 (1824).
[127] It may be observed that I have here broken through the great law of priority; for it appears to me too grossly incorrect to retain the specific name either of major or minus in a genus including a single species. Lamarck himself seems to have been of this opinion, by giving, in 1818, the new specific name of balænarum; but Shaw’s name of trachealis has the clear right to priority, if major or minus be rejected.
General Appearance.—Shell elongated, sub-cylindrical, with the upper end rather wider than the lower, and therefore widening in a direction the reverse of that usual with sessile cirripedes. The shell is often a little bent to one or the other side: it is surrounded by from two or three to about ten very prominent, strong, blunt ridges or belts, placed at rather irregular distances from each other. The surface is finely striated longitudinally. The six compartments are of nearly equal sizes and shapes. In full-grown specimens the parietes are not wider at the base, and often they are even a little narrower, than at the summit of the shell: in young specimens the parietes do widen a little downwards. The radii are narrow; but in young specimens they are proportionably much broader (Pl. 17, fig. 3 b) than in old specimens. The whole compartment, including the radius and wall, is always a little wider at the summit than at the base, in accordance with the shape of the whole shell. The operculum consists of four nearly equal-sized, similar valves, projecting above the upper end of the shell, which is always broken and jagged: the valves are united to the sheath by a very thick, much folded membrane. The aperture leading into the sack is bordered by very prominent lips, projecting above the opercular valves; the latter have their upper layers always scaled off. The shell is imbedded in the whale’s skin up to the level of its operculum. The largest specimen which I have seen was barely one inch (.95) in diameter at the summit, and 1.5 in length; the longest specimen which I have seen scarcely attained a length of one inch and three quarters.
Structure of the Shell.—The parietes are thin, and if the sheath (which extends to near the basis) be removed, they are rendered extremely thin. They are formed by an outer and inner lamina, united by fine longitudinal septa, projecting at the basis beyond the laminæ. The pores thus produced (which in a transverse section are oblong in outline), run up to the summit of the shell, and are not filled up by shelly matter; but I presume that the included tubular threads of corium are protected, at the broken upper end of the shell, by transverse membranous septa. The outer lamina of shell, as in Coronula, is formed, though obscurely, by the union of ledges projecting from the longitudinal septa. The circular prominent belts, surrounding the shell, are formed by the longitudinal septa, at certain, irregular and rather distant periods, growing outwards; the wall at each belt being increased to nearly twice its thickness in other parts. At each belt the threads of corium within the parietal pores lend off minute branches to supply the thickened wall. These belts, which continuously surround the shell, correspond (as is best seen in young specimens), with the little knobs or beads, which, in Coronula (Pl. 16, fig. 4), rise separately, and not quite regularly, on the longitudinal parietal septa, and which, I believe, are formed at every successive period of growth; here they are much larger, stand in straight transverse rows, become confluent, and are formed only at occasional intervals. The whole external surface of the shell is covered by membrane, stronger and more persistent than is usual with most cirripedes.
Internally the sheath extends almost to the basis of the much elon[Pg 433]gated shell, and terminates in a slight shoulder: it is divided as in common Balanidæ, and differently from in Coronula, into zones of growth, but these are very broad; at the upper end of the shell, which, as will hereafter be explained, is always breaking away, the sheath readily yields along the oblique planes, which separate the zones of growth and dip outwards: a similar but less strongly marked structure occurs in Platylepas, and in no other genus. We shall presently see that the sheath presents a much more anomalous character, in being lined down to its basal edge by the innermost and last-formed layers of the opercular membrane.
Radii.—The radii are narrow. The belts which surround the shell are prolonged, with slightly diminished prominence, across the radii, their formation being simply due to the radii being here thicker than in other parts. It can be seen more plainly in Tubicinella, than in other Balanidæ, that the membrane externally investing the shell, splits along the radii during the diametric growth of the shell, and is continually repaired and added to along these lines by new longitudinal slips of membrane. The radius consists (as usual) of an inner and outer lamina, which latter does not extend quite to the line of suture—a slightly gaping fissure being thus left. The two laminæ are connected by septa, which are not denticulated, but near the outer lamina bi- or tri-furcate, and the ends of the branches thus formed spread out, forming a sort of outer scalloped lamina, in advance of the true outer lamina. The fine threads of corium running between these septa, do not spring, as in all common cirripedes, from a fillet of corium occupying the actual suture, but from two nearly circular threads of corium occupying two tubes, which run along the line of junction between the radius and the compartment whence it springs. In Coronula alone we have a nearly similar structure; for the fine threads of corium occupying the proper radius, spring from a single very minute tube (Pl. 16, fig. 7, d′), occupying the same position with the two tubes in Tubicinella. I may further add, that the structure of the proper radius in Coronula is precisely the same as here just described, but being on so very minute a scale, I did not there describe it so carefully as I have here done.
Alæ: these are only remarkable from their extreme thinness; for they are not thicker than the inner lamina of the radius. Their sutural edges are quite smooth. Forming part of the sheath, they extend down close to the basis of the shell; where, instead of, as in general, ending abruptly in a rectangular shoulder, they slope off into their own compartment.
Basis.—The basal membrane is complicated, owing to the shell, when full-grown, barely, or not at all, increasing in diameter, and, in consequence, membrane after membrane, each with its own cement-ducts attached to it, are thrown down one nearly over the other. In the Introduction (p. 143—Pl. 28, fig. 3), I have fully described the cementing apparatus, which is very curious from one of the ducts always having a loop with two spurs projecting from it. The basal membrane does not equal in diameter the base of the shell, for the membrane externally covering the walls is inflected inwards all round for a considerable width, and is then united to the basal membrane: in[Pg 434] Coronula, the basal membrane extends only under the internal cavity of the shell, and not under the folded walls, and therefore presents a somewhat analogous structure.
Opercular Valves.—The scuta and terga are nearly of the same size and shape: they are mitre-formed, and higher than broad. They do not fill up the orifice of the shell. The scutum is a little larger than the tergum, and rather less symmetrical, the rostral corner of the valve being a little produced. There is no hollow or crest for the adductor muscle, which is small. In the tergum there is no trace of a spur. The two valves are not articulated together, but standing close to each other are united, as well as the scutum to the scutum, and tergum to the tergum, by thick, brown, tough, yet soft membrane, in layers continuous with, but differing in appearance from, the surrounding opercular membrane. The layers of shell, forming the valves, are thick, and only the three or four lower layers are usually preserved, the upper ones having symmetrically scaled off, leaving snow-white surfaces. Owing to the thickness of the successive shelly layers, and to the circumstance of each new layer being but very little larger than the last, the scaling off of the old upper layers is a quite necessary process; for otherwise the orifice into the sack would have been encumbered and almost closed by four long, slightly tapering points, prolonged upwards from the basal layers that form the four existing valves. The same scaling off process takes place in Platylepas, and amongst pedunculated cirripedes in Lithotrya. Microscopical examination does not exhibit any fine spines on the membrane investing the valves, or any tubuli in the shelly layers after their dissolution in acid: in this respect the valves resemble those of Coronula. The summits of the valves project freely for about a third of their own height, above the level of the membrane by which they are surrounded. The orifice leading into the sack is bordered by very protuberant lips, standing up even considerably above the upper freely projecting portions of the valves.
The Opercular Membrane, connecting the valves and the top of the shell, is thick and tough, and deeply folded in concentric wrinkles. As in Coronula, it consists of two or three separate membranes (each composed of many laminæ) one over the other, united to successive shelly layers of the opercular valves. As the upper shelly layer scales off, the membrane attached to it is likewise thrown off. The innermost laminæ of the last-formed opercular membrane extend down, closely attached to the sheath, to its basal edge, and therefore nearly to the basis of the shell; the outer and older laminæ, all closely attached one within the other and to the sheath, extend to a less and less distance downwards; consequently, the animal’s body is enclosed in a tube, thinning out downwards, formed by the laminæ of the successive opercular membranes, surrounded outside by the shell: only in the following genus, Xenobalanus, shall we meet with a nearly analogous structure. As the shell of Tubicinella increases in diameter, from the growth of the radii, the opercular membrane lining the sheath is necessarily split along the six lines of suture, in the same manner as is the membrane externally investing the shell; in a like manner, also, it is repaired and added to by new longitudinal slips of membrane. Of[Pg 435] this structure, in the opercular membrane, I have seen no other instance; for in most genera the old opercular membrane is moulted, and a new and larger one formed at each period of growth; in Coronula, in which the opercular membrane is likewise for a time persistent, it does not run far down the inside of the shell, and each new membrane is formed large and extensible, so as to allow, without splitting, of some increase in the diameter of the shell. The opercular membrane at the summit of the shell, in Tubicinella, is folded in concentric lines, and so deeply, that the basal edges of the opercular valves are generally hidden: this folding arises partly from each last deposited and innermost membrane being originally formed slightly folded, but chiefly from the rapid downward growth of the shell, and the consequent downward movement of the whole animal’s body, together with the opercular valves to which the body is attached, and this necessarily tends to wrinkle and fold the opercular membrane. Owing to the opercular membrane extending far down inside the shell, and being firmly attached to the sheath, as the upper part of the shell breaks away and disintegrates (which we shall presently see is constantly taking place), small particles of shell are left adherent to the circumferential and folded parts of the opercular membrane; and this at first much perplexed me.
Muscles of the Sack: these extend down almost to the base of the shell, but in the lower part they spread out and become thin and very irregular, not even corresponding on the opposite sides of the body. The fasciæ in the upper part show very distinct transverse striæ, but lower down these become either obscure or entirely deficient. In all these characters the muscles of Tubicinella and Coronula resemble each other. The rostral depressor muscles of the scuta consist each of four small bundles of fasciæ; the lateral depressores run not quite straight down, but in a curved course towards the carinal end of the sack: the tergal depressores are proportionally smaller than in ordinary sessile cirripedes, but they project and form two crests (with some fasciæ between them), which support the Branchiæ. The membrane lining the sack, I may here mention, is unusually strong.
Branchiæ.[128]—These are enormously developed; the two together covering two thirds of the area of the sack. Each consists of two folds, both deeply plicated. They are attached longitudinally to the two crests, including and formed by the muscles running from the terga to near the basis of the shell. The branchiæ are likewise attached transversely to the sack, under the basal margins of the terga.
[128] These have been described by Professor Owen in the second volume of the ‘Descriptive Catalogue of the Museum of the Royal College of Surgeons.’
Mouth.—The labrum is very finely hirsute, without teeth; the palpi have a short row of moderately long spines along their exterior basal margins. The mandibles have four rather narrow, sharp teeth, which (excepting the first) have double points: between the second and third, and again between the third and fourth teeth, there is a single small intermediate tooth: the inferior angle is irregularly pectinated. The[Pg 436] maxillæ are small; there is a small notch beneath the two upper great spines, and a second notch near the inferior angle. Between the outer maxillæ, there is a square-topped mentum. Hence we see that the mouth in all its few peculiarities, resembles that of Coronula.
Cirri.—These are short, with short and broad segments protuberant in front. The pedicel of the first cirrus is very broad, and exteriorly clothed with fine hairs: its rami are slightly unequal in length. The second and third cirri are very short. The three posterior pairs are remarkable from the pairs of main spines being placed so close one under the other, and in an oblique direction, that at first they appear to form a single crowded transverse row: the dorsal tufts are rather large.
Body.—The body is remarkable from its nearly vertical position, and from the much elongated pyramidal form of the prosoma, extending down nearly to the bottom of the sack. The membrane investing the prosoma, presents a few circular folds, falsely appearing like articulations. The œsophagus enters the stomach rather obliquely. With respect to the generative system, I have only to remark, that the vesiculæ seminales are of great length, and convoluted to a remarkable degree. The ovarian cæca, form a thick layer at the bottom of the sack; they do not appear to extend up the shell round the sack. The only other point, which I shall here mention, is that beneath the basal articulation of the first cirrus, there is a longitudinal swelling, ending in a freely projecting point, .06 of an inch in length; at first, I thought, that we here had a rudiment of a filamentary appendage like those found in several Lepadidæ; but closer examination showed an orifice at the apex, leading into the acoustic meatus, in which the singular, wrinkled, heart-shaped acoustic vesicle, mentioned in the Introduction, hangs suspended. Alongside the freely depending point, with an orifice at its end, there is a smaller upward projecting point, without any orifice, but hollow within and lined by corium; I believe it opens internally into the acoustic meatus.
Attachment and general Growth of Shell.—All the specimens which I have seen have been attached in groups. They are buried up to the level of the operculum in the whale’s skin; and their summits, I suspect, lie even beneath the general surface of the body of the whale. It is certain that the shells grow much at their basal ends. As in the case of Coronula, the flat membranous basis does not actually penetrate the skin; but the general pressure of the whole group of shells seems to push inwards the skin of the whale, and directly beneath each shell the formation of new epidermis is apparently checked. Between the shells, however, though close together, the epidermis continues to be formed, and is pushed upwards between them, in the same manner as it is forced into the flattened cavities on the under side of the shell of Coronula.
The manner in which a full-grown shell assumes its ordinary shape, at first appears very perplexing:[129] it has to change from a cylinder, at[Pg 437] first probably not much above the 1/50th of an inch in diameter, to a cylinder nearly one inch in diameter: and this is not effected by the growth of the radii, for the radii never reach the basis, and the basis of course has to increase in diameter like the rest of the cylindrical shell. The radii serve only to keep the summit of the shell wider than the basis, which is the natural shape of this species; and in large-sized specimens, this purpose is sometimes aided by the parietes during their downward growth decreasing slightly in width. In ordinary nearly full-sized specimens, the parietes are of the same width at the top and bottom, but in some large-sized specimens, as just stated, they even become narrower towards the bottom; as they grow only at the bottom, one does not at first see how they can ever increase in width, or how the older shells can have acquired their present diameter. But an examination of young specimens, from .1 to .3 of an inch in diameter, at once serves to show how the shell attains its full size and shape: for here the parietes are all found to increase downwards sensibly in width, though at a much slower ratio than in other sessile cirripedes; in larger, but not full-grown specimens, a similar increase can by care be detected: hence by long-continued growth at the base of the shell, with the removal of the upper part, a young Tubicinella of small diameter will be converted into an old one of large diameter, retaining during all the time its sub-cylindrical form, with its summit rather broader than its base. With respect to the removal of the upper part of the shell, this seems almost constantly going on, for the summit of every specimen invariably had a freshly broken aspect. The peculiar structure of the sheath, which is the strongest part of the shell, namely, its division into oblique layers, separable by a slight force, doubtless is subservient to the repeated breakage of the summit. In some species of Tetraclita and of Balanus, gradual disintegration of the upper part of the shell is a necessary element in the growth of the animal, in order that the orifice may increase in size, and here we have mechanical breakage equally necessary.
[129] I am indebted to Dr. J. E. Gray for calling my attention to this subject, and explaining to me several points.
Some curious results follow from the peculiar growth of Tubicinella just described. At Plate 17, fig. 3 b, we have a careful drawing of a lateral compartment, together with its radius, (which latter does not here concern us), taken from a shell .2 of an inch in diameter. The two protracted dotted lines show the form which this compartment would have assumed, if it had continued growing downwards at the same rate of increase in width as hitherto. But the increase in width always seems to become less and less as the shell grows older; hence the dotted lines, representing the wall after long-continued growth, ought to have been drawn diverging or widening still more slowly than they do. The lateral compartment in fig. 3 a is the exact size of the compartment of a large specimen nearly one inch in diameter; in this specimen the parietes, far from increasing in width downwards, had commenced, as is represented, decreasing. Compartments of all intermediate sizes between those figured at 3 a and 3 b can easily be shown in different specimens. From these facts we may safely infer, that if the whole growth of the compartment 3 a had been preserved, instead of its upper end having been continually chipped away, it would have[Pg 438] had even a more tapering form than that represented by the whole and dotted lines in the two figures, and would have exceeded six inches in length! this of course being also the length of the whole shell. The young Tubicinella, of which 3 b is a compartment, was imbedded in the whale’s skin nearly up to the level of its operculum; if it had lived, it would no doubt have grown to the length just specified, viz., above six inches, but as all the growth is at the lower end, the bottom of the shell, it might be thought, would necessarily have become buried in the whale’s skin to this same depth; and the summit of the shell, on this same view, would have been buried to a depth by as much less as the height or length of the old shell itself, namely, by about one inch and a half less than the six inches. As far as I can judge from an examination of several large groups of full-grown specimens, preserved in their imbedded condition, the summits of the shells seem always to lie a little beneath the surrounding level of the whale’s skin, but not nearly to the extent here just inferred. Nor can I believe that the epidermis of the whale had ceased being formed under these specimens, whilst it had gone on being formed all round them, to the thickness of between four and five inches, and that it had subsequently disintegrated to this same thickness,—which processes would account for the summit of the shell being still on nearly a level with the surface of the whale. The view which seems to me most probable, is, that the rapid downward growth of the shell, besides indenting the whale’s skin, at the same time slowly pushes the whole shell out of the skin, and thus continually exposes the summit to the wear and breakage which seems to be necessary for its existence. On this view, the very peculiar form of Tubicinella, which is retained during life, namely, the slightly greater width at top than at bottom, is beautifully explained, viz., for the sake of facilitating the protrusion of the shell; for the ordinary conical shape of sessile cirripedes, with the apex upwards, would have rendered the pushing out of an imbedded shell almost impossible; on the other hand, we can see that the likewise very peculiar, concentric, prominent belts may be necessary to prevent too easy protrusion.
Fossil Species.—I do not believe that this genus has hitherto been found fossil. The Tubicinella maxima of Ch. Morren, said to have been found (see Bronn, Index Palæontologicus) in the chalk of Belgium, I have good reason to believe does not really belong to this genus.
XENOBALANUS. Steenstrup. Videnskabelige Meddelelser. Aaret, 1851.
SIPHONICELLA (sine descript.) Darwin. Monograph on the Lepadidæ, p. 156 (1852).
Shell almost rudimentary, star-formed, composed of six compartments, with a long peduncle-formed body rising from the middle: opercular valves none.
Hab.—North Atlantic Ocean, attached to Porpoises; Mus. R. T. Lowe, Steenstrup.
This Cirripede, in appearance the most anomalous of its family, has affinities distinctly pronounced. Four years ago the Rev. R. T. Lowe sent me some specimens, which he had obtained from a porpoise between Madeira and England; and I named them in MS. Siphonicella, from their relationship to Tubicinella,—a fact which I mention only because Sir C. Lyell has alluded to this genus under the above name (without any description), in his anniversary address to the Geological Society, as have I, in my volume (p. 156) on the Lepadidæ. Since that time Professor Steenstrup has described and named the genus, fully recognising its place and affinities, and has most kindly sent me a magnificent group of specimens.
This genus singularly resembles, in general appearance, some of the pedunculated Cirripedes, so much so that in the specimens sent me by Mr. Lowe, in which the almost rudimentary shell was, from disintegration and its deep imbedment, not plainly visible, I did not in the least doubt that I was examining a new genus of Lepadidæ. I may mention, as a proof how truly all the parts and organs are correlated in Cirripedes, that I was at first in despair when I found a species to all appearance pedunculated, with its labrum not bullate, its palpi of large size, its third pair of cirri totally unlike the fourth and succeeding pairs, and with only a single layer of muscles round the peduncle; but when, in addition, I found that there were branchiæ, and that these were double, I felt convinced that I was dissecting a disguised sessile cirripede, and that its true place was near Coronula: soon, I found the imbedded and almost rudimentary shell, of which a mere fragment would equally well have declared the true position and relationship of the whole animal. Though Xenobalanus, in external aspect, is so completely masked, yet in its habits, namely, in living attached on Cetaceans, as in its essential structure, it displays its real affinities. In the course of the following description, it will be seen that in the shell, the affinity is almost equally close to Coronula and Platylepas, but that, considering the[Pg 440] whole animal, the affinity is somewhat closer to Tubicinella. Xenobalanus may indeed be described as a Tubicinella without opercular valves,—with the opercular membrane thickened down to the basis,—and with the shell, excepting the few last-formed basal zones of growth, almost wholly removed by the breakage of its upper end; this remnant of a shell, however, presenting some strong points of resemblance to Coronula.
XENOBALANUS GLOBICIPITIS. Steenstrup. Videnskabelige Meddelelser fra den Naturhist. Forening i Kjöbenhavn, for Aaret, 1851. Tab. 3, fig. 11-15.
General Appearance.—The shell is in an almost rudimentary condition, and appears like a small white irregular star, imbedded up to its top in the skin of the porpoise. Out of this thin, star-shaped shell, a cylindrical, flexible, peduncle-formed body springs, which forms the main part of the animal; it is narrow where coming out of the central cavity of the star, but soon acquires its full diameter; at the upper end it has a reflexed hood, and hence is broader, and this has the appearance of forming a capitulum, like that of a pedunculated Cirripede. This pseudo-capitulum is formed by a membranous reflexed collar or hood, which is very narrow at the lower end of the orifice, close under the mouth, and becomes wider and wider towards the upper and carinal or posterior end of the orifice; hence the lower reflexed edge of the hood is only slightly oblique or even nearly transverse. The orifice leading into the sack is large, and nearly in the same straight line with the peduncle; it is a little hollowed out in the middle at the upper end, and on each side of this medial hollow, there is a small rounded projection or horn, not perforated, but hollow, as may be seen by turning up the hood and looking at its under side. These two little horns curiously bring to mind the ear-like appendages in Conchoderma aurita (Otion), but these latter are perforated, open into the sack, and point outwards. The peduncle-formed body answers, as we shall presently see, to the main part of the shell in Tubicinella, and the hood, as it would appear, to the lips of the sack-aperture, which project between its scuta and terga; of these valves there is not here a trace. The whole surface is smooth, and is formed by rather thin membrane, of an orange colour; but from the colour of the underlying corium, the whole appears of a dark chocolate red, the reflexed hood being rather lighter coloured. It is singular how closely the colour resembles that of some dark varieties of the above-mentioned Conchoderma aurita, and likewise of Anelasma squalicola, both pedunculated Cirripedes,[Pg 441] having oceanic habits, and destitute, to a remarkable degree, like Xenobalanus, of shelly valves.
The largest specimen which I have seen was very nearly two inches in length: in this specimen the star-shaped shell measured, from extreme point to point, nearly a quarter of an inch in diameter, but the internal cavity only about one eighth of an inch. This latter measure gives also the diameter of the peduncle, where coming out of the shell; the diameter just beneath the hood, was in this same specimen rather more than a quarter of an inch, and therefore greater than the diameter of the points of the shell. The depth of the shell from the upper rim to the basal membrane, in one specimen which I measured, was only one twentieth of an inch, and this specimen had its pseudo-peduncle one inch and three quarters in length, consequently thirty-five times as long as the shell was deep.
Structure of Shell.—The almost rudimentary shell (fig. 4 b) consists of a small, thin, six-rayed disc, formed of six compartments, each of which, instead of being outwardly convex, as in ordinary Cirripedes, is deeply bowed inwards. The narrow sutures (s s) separating the six compartments, run along the middle of the six rays, each ray being composed of the bowed ends of the walls of the adjoining compartments. The rays are a little curved towards the carinal end of the shell. It is remarkable that the rostrum is smaller and less deeply folded inwards than the other compartments, and the lateral compartments are a little smaller than the carino-lateral compartments, which is exactly the reverse of what is usually the case. Only about four zones of growth have been preserved in any specimen, and consequently the shell is very nearly of the same diameter at the top and bottom; for the upper end of the shell is rapidly removed, as in Tubicinella, by the scaling off of the upper rims of the sheath, and by the disintegration of the walls. The zones of growth are commonly not piled exactly over each other, but rather obliquely, as represented in fig. 4 b. Each zone projects, forming a prominent, sharp, toothed ridge round the shell. In Coronula (Pl. 16, fig. 6) and its allies, the outer lamina of the wall is formed by the union, a little above the basal margin, of ledges running along the sides of the longitudinal septa. In Xenobalanus (Pl. 17, fig. 4 c) similar ledges are less perfectly joined, and apertures seem always to be left in transverse rows under the transverse toothed ridges, which latter are best seen in fig. 4 b. The apertures, of course, are covered by membrane. The transverse ridges are surmounted by knobs arising from the longitudinal septa; and the knobs themselves are capped by other little heads, which are not represented in the drawing. Owing to these projections, and to the prominence of the transverse ridges and of the longitudinal septa, the external membrane is attached so firmly to the shell that even with the aid of caustic potash it can hardly be separated.
The internal cavity of the shell is small: it approaches a hexagon in shape, with the rostral side very short, and the lateral sides curved inwards. It is lined by a rather thick sheath, which descends very near to the basal membrane; the sheath is divided into very distinct, successive convex zones of growth. The external membrane of the pseudo-[Pg 442]peduncle is attached with remarkable strength to these rib-like zones of the sheath. The alæ (a in fig. 4 b) are represented by mere angular shoulders, received into very slight notches, and placed at the inner ends or entrances of the double walls, or rays as I have called them. With respect to the radii, they also are in an extremely rudimentary condition; but a thin layer of shell, apparently continuous and homologous with the sheath, extends from the sheath along both sides of each ray, and on the rostral side (whence the radius ought to arise), about half-way from the end of the ray, gives rise to a projection or ridge (d, in fig. 4 b) which runs from the top to the base of the shallow shell. From this longitudinal ridge, septa, parallel to the basis, extend to nearly the extremity of the ray or double wall. These represent the radii; but they never grow, so as to increase the diameter of the shell. These radii evidently correspond to the additional or pseudo-radii in Coronula, which in that genus lie between the parallel, as here, and folded parietes. Of the true radius, having the same thickness as the paries, I here saw only traces in an internal, very slight, longitudinal ridge running up the shell, close to the outer extremity of each ray or double wall.
The membranous Basis is united all round to a rather wide flap of membrane which is inflected from the outer surface of the shell. The cement-glands appear to be mere enlargements of the cement-trunks, which latter extend in two nearly straight and parallel lines. From each gland two cement-ducts proceed, one of which runs parallel to the cement-trunk.
The pseudo-peduncle, forming the main part of the animal, has, as far as external appearance is concerned, been sufficiently described. The part forming the hood apparently answers to the protuberant lips of the operculum, and the lower part to the sub-cylindrical shell of Tubicinella; both shell and peduncle in the two genera being wider at top than at bottom. If in imagination we chip away (an action always in progress) the whole upper part of the shell of Tubicinella, leaving only two or three zones of growth at the base, we shall convert it into a Xenobalanus, with every internal part and organ occupying the same relative position: for it should be borne in mind that the shell of Tubicinella is lined close down to the basis by the opercular membrane, and this is strictly comparable with the outer membrane of the pseudo-peduncle of Xenobalanus. The body, as in Tubicinella, is attached in a vertical position, with the longer axis of the thorax and of the much elongated prosoma extending in the direction of the longitudinal axis of the pseudo-peduncle. The point of attachment of the body to the lower part of the hood is much elongated, but presents the usual muscles running to near the base of the labrum and embracing the prosoma. The adductor scutorum muscle is well developed, and is placed close beneath the mouth, where the collar or hood commences folding over: hence, no doubt, the lower end of the large and long orifice into the sack can be closed by the adductor; but the upper end probably can only be blocked up by the outer surfaces of the curled cirri. The scuta, I may add, if such had existed, would certainly have covered the point where the adductor muscle is attached to the exterior membrane.[Pg 443] The sack extends down to within the almost rudimentary shell: the tunic lining it is unusually strong; indeed, in the reflexed hood-like portion, it is as strong as the properly external membrane. The latter seems to be moulted in large strips, and not in a single piece, like the opercular membrane in Balanus: just above the shell, fragments of three or four of these outer coats are retained by their very firm attachment to the sheath. The two layers of corium, lining the outer membrane of the pseudo-peduncle and the inner tunic of the sack, instead of being, as usual, united almost continuously together, stand some little way apart, and are connected by longitudinal septa; hence, in a transverse section, especially of the lower part, the sack is surrounded by an irregular ring of square tubes of corium. The muscles surrounding the sack and imbedded in the inner fold of corium, are very thinly spread out; they branch, and even sometimes cross each other; they are more numerous at the carinal and rostral ends, but certainly cannot be said to form six (or five) bundles, as in all other sessile cirripedes. Some of the fasciæ extend down to the very basis, and some up to the summit, to near the two little horn-like projections. I could not perceive any transverse striæ on these muscles. Altogether, they are very weak, and cannot have much power in moving the whole peduncle-like body.
Branchiæ.—These are largely developed: they are attached to two approximate, longitudinal, fleshy crests, which extend more than half-way down the sack, along the carinal margin. Each branchia is double, the two folds being united where attached in a transverse line across the sack, on a level with the attachment of the body. The inner fold is much smaller than the outer; not extending half so far down the sack, and not extending so far transversely; it is also hardly at all plicated. The larger and plicated fold extends down considerably below the lower end of the prosoma, and altogether fully equals one third of the entire length of the animal, measured from the shell to the summit of the orifice. Both folds are formed of very delicate membrane.
Mouth.—Labrum unusually prominent, as measured from its basal margin to the crest, which is but slightly notched, hairy, and without teeth. Palpi broad, heart-shaped, clothed on their inner sides by a thick brush of spines, which here, as on the other gnathites and cirri, are almost all doubly serrated. On the outer margin of the palpi there are a few longer spines. Mandibles villose, with five teeth, of which the fifth is very small and of irregular shape: the inferior angle is broad and pectinated. There are no intermediate teeth between the second, third, and fourth teeth, as in the three foregoing genera. Had I not known that the lower main mandibular teeth were always laterally double in the Balaninæ, and had I not observed how obscure this structure was in Coronula and Tubicinella, I should have overlooked the merest vestiges of double teeth in the present genus; indeed, in some specimens the teeth seemed to be absolutely single. The maxillæ are villose: their edge exhibits a trace of being notched under the two great upper spines. The outer maxillæ are bilobed, but not very plainly: between these organs there is no little prominent mentum, as in the three previous genera.
Cirri.—The cirri are short, particularly the three anterior pairs. The segments in all are singular, from being so much compressed, so short, and of such great breadth; they are protuberant in front. In the second and third cirri, the broad lateral faces of the segments, with the exception of the posterior face of the posterior ramus, are almost bare of spines. In the three posterior pairs of cirri (Pl. 29, fig. 6), the segments are protuberant in front, and support three pairs of short thick spines, with an intermediate tuft; the dorsal tufts are unusually small: their pedicels are remarkable from the upper segment, and the upper part of the lower segment, being produced into a rounded protuberance, dotted with spines: I have met with a similar structure only in Scalpellum vulgare. In the third pair (fig. 5) there is only a trace of this structure; and in the second pair the anterior margin of the pedicel is straight, and clothed with three tufts of bristles. The pedicel of the first cirrus is very broad, and clasps the mouth.
The cirri and mouth are dark chocolate red, like the outside of the animal and the upper part of the sack. The thorax is redder and paler. The four posterior articulations of the thorax are straight and transverse; the next segment, or that corresponding with the second pair of cirri, is slightly inflected, in the usual way, towards the prosoma. The prosoma is pale coloured, extraordinarily elongated, and bluntly pointed; it extends down (see the dotted outline in Pl. 17, fig. 4 a) about one third of the length of the whole animal. The orifice leading into the acoustic sack forms a freely depending little point beneath the basal articulation of the first cirrus. The stomach in the uppermost part is deeply and closely plaited longitudinally, but has no cæca; it runs down (externally coated, as usual, by the testes) to the lower point of the prosoma, and is then doubled back on itself, so that it is very long.
Generative System.—The probosciformed penis is short and thick, and covered with very minute tufts of bristles: there is no knife-edged projection at its dorsal base. The vesiculæ seminales are much convoluted and of great length. The ovarian tubes form a small sheet within the rudimentary shell, in the normal position, over the basal membrane; and likewise higher up between the two folds of corium surrounding the pseudo-peduncle; they do not, however, appear to occur round the lower part of the peduncle: they extend highest on the rostral and carinal sides, and lowest on the two lateral faces. The ova are wonderfully numerous; they are 15/2000ths of an inch in length: they form, instead of two thin lamellæ, two almost cylindrical packets, which are held together by most feeble membrane. Each packet, in the upper part lies between the two folds of the branchiæ; and in the lower part, is embraced only by the larger outer fold. The two packets of eggs sometimes cohere together at their lower ends.
Imbedment.—The shell is imbedded up to its summit, but the shell is very shallow. The imbedment seems due either to the compression of the epidermis of the porpoise, or to its formation beneath the shell having been checked; the outline of the true skin under the dark-coloured epidermis is not in the least affected. The epidermis fills up the bay-like spaces formed by the inwardly folded walls, and firmly adheres to them.
Affinities.—This genus presents very varied affinities to Tubicinella, Coronula, and Platylepas. To the latter it is more especially allied in the compartments, being singly folded inwards, though the sides of the folds are not here closely pressed together, as in Platylepas: in both these genera the fold is less deep in the rostrum, of which fact, in Platylepas, the final cause is evident, but here there seems no cause, excepting the simple one of affinity. Xenobalanus is further allied to Platylepas, in the lesser size of the inner fold of each branchia, compared with the outer, and in the structure of the cement-glands, and to a certain extent in that of the sheath. To Coronula the special alliance is shown by the remarkable character of the pseudo-radii lying between the parallel and adjoining walls, and in the general character of the cirri: in Coronula we have the terga sometimes quite aborted, and the scuta of small size, thus exhibiting a tendency to the entire absence of opercular valves, so remarkable in Xenobalanus. To Tubicinella, the alliance is still more plainly shown in the external shape of the whole animal, wider at top than at bottom,—in the opercular membrane descending almost to the very base—in the relative positions of the different parts and organs—in the upper end of the shell continually scaling off—in the prominence of the transverse external ridges—in the sheath being divided into successive zones of growth, and being prolonged nearly to the basal membrane—and in the edges of the alæ being smooth. Internally, the resemblance is also plainly shown, in the strength of the internal tunic of the sack—in the branchiæ springing from two approximate fleshy crests—in the freely depending acoustic orifices—in the form of the thorax and prosoma—and consequently of the alimentary canal.
Seeing the state of the almost rudimentary shell, it is not difficult to imagine its total disappearance. Thinking of this, it occurred to me to doubt for a few minutes, whether Anelasma, described in my volume on the Lepadidæ, which presents many points of resemblance with Xenobalanus,—viz., in colour, in the strength of the internal membrane of the sack, in the size of the orifice with its thin membranous margin, in the entire absence of opercular valves, and in epizoic habits,—might not in truth be a sessile cirripede, in an extremely altered condition; for the cirri of Anelasma are in so rudimentary a state, and the mouth in so modified a condition, that the internal characters by which pedunculated cirripedes can be distinguished from the Balaninæ, though not from the Chthamalinæ, are almost lost. But if Anelasma had belonged to either of the two sub-families of the Balanidæ, the sack would have penetrated almost to the bottom of the peduncle; there would have been only a single layer of longitudinal muscles round the peduncle; and there[Pg 446] would not have existed ovigerous fræna. I believe that Anelasma and Xenobalanus are only analogically connected, being no more related together by true affinity, than any other two genera in the Lepadidæ and Balanidæ.
Shell with the rostrum having alæ, but without radii: rostro-lateral compartments without alæ on either side: parietes not porose.
Mouth with the labrum bullate; palpi hardly touching each other: third pair of cirri with the segments resembling those of the fourth pair.
The shell in this sub-family consists of four, six, or eight compartments, with the addition, in Catophragmus, of several whorls of supplemental compartments or scales, like those forming the lower part of the capitulum in Pollicipes. The rostrum has alæ and no radii; in shape and size it resembles the carina. The rostro-lateral compartments are destitute of alæ; in all cases they overlap the adjoining compartments, and have radii (when such are developed) on both sides. In the genus, Pachylasma, however, the shell must be looked to very young, in order to detect this normal structure, for soon the true rostrum and rostro-lateral compartments become blended together, making a compound rostrum, destitute, as in the Balaninæ, of alæ, but furnished with radii. The parietes are never porose, nor furnished with regular symmetrical ribs (representing the longitudinal parietal septa) on their inner surfaces: sometimes, however, on the basal internal edges, there are some irregular depending points. In conformity with the simplicity of the parietes, the Radii (which are seldom much developed) have likewise a simple structure; and often are merely formed by the simple lateral growth of the parietes. The scuta and terga are articulated together more deeply than is usual in the Balaninæ; and the terga never have a long spur.
Looking to the animal’s body. The Labrum is always swollen and bullate, and though hollowed out, is never notched in the middle. The Palpi are rather small, and have not their tips nearly touching each other. The mandibles generally have their lower main teeth laterally single, though sometimes they are double: the inferior angle is always pectinated. The maxillæ are always notched under the upper pair of great spines. Of the Cirri, the third pair much more closely resembles, in external structure and powers of movement, the fourth than the second pair, though sometimes a few of the basal segments, especially on the anterior ramus, are thickly covered with bristles, as on the segments of the second pair. In three species belonging to two genera, there are caudal appendages, resembling those generally present amongst the Lepadidæ, but never found in the Balaninæ. The Branchiæ are generally smaller than in the Balaninæ; and are sometimes quite rudimentary.
In the introductory description (p. 152 and 176) of the Family Balanidæ, I have already discussed the relations of the Chthamalinæ to the Balaninæ, and of the several genera to each other, so that I need not here re-enter on the subject.
CHTHAMALUS. Ranzani. Memoire di Storia Naturale, 1820.
EURAPHIA. Conrad. Journal Acad. Nat. Sc. Philadelphia, vol. 7, 1834.
Compartments six: basis membranous, but sometimes in appearance calcareous, owing to the inflected parietes.
Distribution mundane; attached generally to littoral rocks and shells.
This, the typical genus, is the largest and widest distributed group of the sub-family Chthamalinæ. The founder of the genus apparently did not perceive its essential character; Savigny, however, as is evident from the excellent figure in the great work on Ægypt, perfectly understood[Pg 448] the difference between Chthamalus and Balanus. I was first indebted to Dr. J. E. Gray for explaining to me this difference; but the only published account which I have met with is in a paper by the Rev. R. T. Lowe,[130] in which he states, on the authority of Mr. Clark of Bath, that in Chthamalus the anterior compartment or rostrum has alæ like the posterior compartment or carina, the anterior or rostro-lateral compartments being destitute of alæ. These characters being exactly reversed in Balanus, as I have already explained (p. 176) under the sub-family of Balaninæ.
[130] ‘Zoological Journal,’ vol. 3, p. 76, 1828.
The shell, owing apparently to its containing much animal matter, is particularly subject to disintegration; and when thus much affected it is quite impossible to distinguish the species by external characters. It is, in fact, best to cast on one side external appearance, though when the shells happen to be well preserved, each species has its own peculiar aspect. We have in this genus smooth and plicated, cylindrical and depressed varieties of most of the species. The development of the radii is very apt to vary, and even the compartments often become so completely united and calcified together that the sutures are almost or quite obliterated. A more serious difficulty in discriminating the species, arises from the fact of the opercular valves, not only varying extremely in external appearance in consequence of the greater or less disintegration of their apices, and consequent exposure of their articular ridges and furrows (compare fig. 1 a, 1 b, 1 c, in Pl. 18), but from their truly varying in outline with the varying shape of the shell: this latter circumstance is probably due to the opercular membrane which unites the valves to the shell being very narrow, and in consequence, differences in the shape of the shell affect the opercular valves, in a manner and to a degree to which the Balaninæ are not subject (compare fig. 1 e, 1 f, 1 h, in Pl. 18). The scuta, on the other hand, differ to an unusually slight degree in the different species. In the common Chthamalus stellatus, which abounds on the southern British shores, the whole external aspect of the shell is[Pg 449] often so completely masked, owing to its varying shape, its obliterated sutures, its deeply disintegrated and punctured surface, and by the corroded condition of its opercular valves, that I have found this species, in the collections of naturalists who have attended to cirripedes, arranged actually on the same tablet, mingled with specimens of Balanus balanoides. I have myself several times found it less troublesome to discriminate these two genera by the included animal’s body than by the shell,[131] though the latter, when well preserved and developed, possesses such obviously well-marked characters: the same thing has occurred to me with some of the other species.
[131] It will be found ultimately to save time and trouble, to soak for half an hour in hot caustic potash, a specimen out of each group of shells to be examined, and then well wash and brush the separated valves and compartments; this process has been followed by me with all the species here to be described.
General Appearance.—The shell is generally depressed, but sometimes, when growing in groups, cylindrical. The surface is either smooth or longitudinally folded; and, as already stated, very apt to be deeply disintegrated. The radii, when developed, are narrow, with their summits oblique and rounded; but they are often quite absent, and sometimes even the sutures are almost obliterated. The rostrum and carina are of the same shape and size, and the two lateral compartments on each side are of nearly equal breadths. The orifice is generally sub-rhomboidal, being widest towards the carinal, instead of towards the rostral end, as is usual in Balanus: but in Chthamalus fissus the orifice is narrow and elongated. The opercular valves have their apices generally disintegrated and worn away, and are then seen to be deeply locked together. The colour of the shell is dirty white or dull purplish-red or brown; but in C. intertextus rich violet-purple. The species are small, not often exceeding half an inch in basal diameter, with the exception of C. Hembeli, of which I have seen a specimen two and three quarters of an inch in diameter.
Scuta.—These present no particular character, excepting in most cases the large development of the articular ridge, and sometimes the presence of a furrow above the articular[Pg 450] ridge, of which only traces can rarely be detected in Balanus; hence in these cases, as in C. Hembeli (Pl. 18, fig. 5 a, 5 c) and scabrosus (Pl. 19, fig. 2 a, 2 c), the line of articulation between the scutum and tergum is more complicated than is usual. There is generally a slight pit, sometimes even furnished with small crests, for the lateral scutal depressor muscle: in C. scabrosus (fig. 2 d, q) a part of this muscle is attached to a small pit at the basi-scutal corner of the tergum,—a fact of which I have observed no instance in any other genus. In C. intertextus the terga and scuta are calcified together, without even a suture being visible on their internal faces. In C. Hembeli the valve is externally marked by a few longitudinal furrows.
The Terga, like the scuta, have a prominent articular ridge and deep furrow. In many specimens of C. stellatus and in C. scabrosus the valve is narrow: in C. fissus it is triangular and nearly equilateral. The crests for the depressores muscles are well developed: in C. scabrosus (Pl. 19, fig. 2 d, p) these crests are united into a plate, which, together with the outer lamina of the valve, forms a deep narrow pit: in C. Hembeli the crests are furnished with small sub-crests. The opercular membrane is narrow; it is sometimes furnished with a few minute spines.
Structure of Parietes.—The parietes are solid, and composed of successive layers of shell; the inner surface varies in condition in the same species, being either smooth or marked with branching impressed lines, or mamillated, or often irregularly punctured for the entrance of tubuli. Owing to these tubuli, the walls, when externally disintegrated, often become punctured. In certain depressed varieties of both C. stellatus and scabrosus the walls are supported by irregular depending columns, placed along either one side or both sides of the sutures. In C. intertextus we have the remarkable character of the wall of the shell growing (I presume after a certain age) almost rectangularly inwards, thus forming a rather wide, flat, calcareous rim round the central basal membrane. In C. Hembeli the internal basal edges of the parietes, in one moderately young specimen, were rugged with irregular points, but presented no other remarkable appearance; but in five old and very large spe[Pg 451]cimens, the whole basis was calcareous, being absolutely continuous with the inner lamina of the parietes, showing that the latter had grown flatly inwards all round, and had then become confluent in the middle, so that there was no longer any basal membrane; excepting, no doubt, that which had existed in the younger stage, and which would be preserved in a functionless condition between the surface of attachment and the inflected parietes.
When an opercular valve or compartment is dissolved in acid, layers of tissue are left, and these are seen to be penetrated by tubuli, which enter at the punctures before mentioned on the inside of the shell: these tubuli often stand in groups of three or four together; they are about 1/10000th of an inch in diameter. Besides these irregularly scattered tubuli, there are in the opercular valves of C. antennatus, innumerable smaller parallel tubuli, running to the external investing membrane.
Structure of the Radii and Alæ.—The radii, when developed, are always rather narrow. Their recipient furrows are generally nearly as broad as the radii themselves. Their edges are either quite smooth, as in C. antennatus; or very finely crenated; or, as in C. dentatus and Hembeli (Pl. 18, fig. 3 a, 5 a), so strongly crenated as to make the suture, both externally and internally, toothed: in these two species, the radii are ribbed in transverse lines parallel to the basis, each rib corresponding with one of the projecting and interlocking teeth on the sutures. In C. intertextus, and much less plainly in most specimens of C. scabrosus (Pl. 19, fig. 1 a, 2 a), we have a structure in appearance very different, for the radii here consist of several very oblique plates, (i. e. nearly parallel to the parietes) on both sides of the sutures, which are interfolded or locked together: I believe that this structure is a mere modification of that in C. dentatus and Hembeli, the transverse ridges on the radii of those species being here developed into oblique plates. We shall hereafter meet with a similar structure in the genus Verruca; to which genus, until meeting with these two species of Chthamalus, I had thought that the interfolding sutures had been confined. The alæ have their edges generally finely crenated: during diametric growth (when such takes place), they are rarely[Pg 452] added to above the level of the opercular membrane, and hence their summits are oblique: in C. intertextus, however, the alæ are laterally added to above the opercular membrane, and their recipient furrows are likewise added to, of which fact I have seen no other distinct instance in any genus; hence on both sides of the sutures, in the sheath of this species, the lines of growth are upturned. In some much disintegrated specimens, both of C. stellatus (var. depressus) and of C. antennatus, the radii have been corroded away, and the diametric growth is effected exclusively by the growth of the alæ, which are moreover much exposed, and rendered conspicuous. The sheath descends a moderate distance down the shell. When a shell is boiled in potash, the sutures (excepting when abnormally calcified together, as very often happens with some species) always fall apart, showing that the union is simply by animalised matter.
Basis.—The basis is always membranous; but we have seen, in C. intertextus, that the walls form a flat ledge all round the base, and that in old specimens of C. Hembeli, they grow so far inwards and become so completely confluent, that they might most easily be mistaken for a true calcareous basis. I may add, that in one elongated specimen of C. stellatus from La Plata, the walls had likewise grown rectangularly inwards, forming a flat base, and had then turned upwards in the middle, forming a medial crest, with the edges not quite calcified together. The true basal membrane is very obscurely divided into concentric slips. I observed in several species, attached to the lower surface, an excessively fine network, quadrangular or hexagonal, of yellow vessels, which seemed insensibly to pass into the sheets, discs, and globules of cement, by which the membranous basis adheres to the supporting surface. I saw, in C. antennatus, numerous irregular, bifurcating, and inosculating cement-ducts, of unequal diameter, often crossing each other, and sending off branches ending in points: the older ducts, instead of being solidly filled up with cement, were only divided by septa. I did not succeed, in any species, in discovering the cement-glands.
Mouth.—The labrum is slightly bullate, with the middle portion depressed, but not forming a notch; in some species[Pg 453] it is hairy, and in some pectinated with short spines. The palpi are of moderate size. The mandibles have from three to five main teeth, the number sometimes varying even in the same species: the lower teeth are either plainly double laterally, or very obscurely double, or to all appearance quite single: a rather large lower portion of the mandible is finely pectinated. The maxillæ are always notched under the two or three large upper spines: the notch bears some fine spines: beneath the notch there are some large spines, and at the inferior angle some smaller ones.
Cirri.—The first and second pairs are always very short compared with the four posterior pairs. The rami of the first pair are slightly unequal. The third pair, in length and arrangement of the spines, very closely resembles the three posterior pairs; in C. intertextus, however, the few basal segments, chiefly on the anterior ramus, are thickly clothed with bristles, like the segments of the second cirrus. In C. antennatus (Pl. 29, fig. 2), the anterior ramus of this same third pair is usually (one single specimen being excepted) much elongated, having at least twice as many segments as the posterior ramus, but the number is variable; and these segments, either all, or only the upper ones, instead of having their spines regularly arranged in pairs, in a double row, are surmounted each by a circle of spines: I suspect that these elongated rami of the third cirrus act as antennæ. It can hardly be an accidental coincidence, that certain genera, as Lysmata and Pandalus, amongst the Macrourous Crustaceans, have the same leg (homologically the second thoracic limb) elongated and antenniformed. Certain varieties of C. stellatus and cirratus, also, have the anterior ramus of this same third cirrus considerably elongated. We are thus reminded of the remarkable variability in the numbers of the segments, and in the arrangement of their spines, in the cirri of Tetraclita porosa; in that species, however, it was chiefly the terminal segments of the posterior ramus of the third cirrus which were so highly variable. The three posterior pairs of cirri in Chthamalus support from three to five pairs of main spines on each segment, the number often varying in the same species, with some minute intermediate bristles. The[Pg 454] dorsal surfaces of these segments, in some of the species, are serrated in an upward direction.
Body, &c.—The body does not present any particular character: in C. scabrosus there is a slight ridge running from the base of the first cirrus towards the adductor scutorum muscle: this ridge is clothed with a few hairs; there are also some hairs at the carinal end of the sack. In C. dentatus, also, there are hairs on the outer tunic of the prosoma. In two species which I opened, there were no cæca to the stomach. The ova vary in length from 13 to 14/2000th of an inch in length; they are packed in two lamellæ lying on each side of the animal’s body. The larva just escaped out of the egg, in C. stellatus, scabrosus and dentatus, had a large probosciformed mouth.
Branchiæ.—These present a very singular amount of difference within the limits of the same genus. In C. stellatus and antennatus we have a simple fillet, tapering a little, barely plicated, and about half, or more than half, as long as the sack; in C. scabrosus the branchiæ are entirely aborted, or are perhaps represented by the slight hairy ridge at the carinal end of the sack: in C. dentatus, on the other hand, each branchia consists of two large folds, barely plicated, almost covering the whole side of the sack, so that here the branchiæ are developed to an unusual degree, more than in Balanus, and as in Coronula and its allies: in the same manner as in these latter genera, the outer fold is considerably larger than the inner fold.
Affinities.—C. intertextus is the most distinct species of the genus, as shown by the peculiar radii and alæ, by the scuta and terga being calcified together, by the character of the third pair of cirri, and by the inflected parietes forming a ledge round the membranous basis; but in this latter respect C. intertextus resembles C. Hembeli. C. Hembeli, in its serrated radii, is closely related to C. dentatus; and this latter species differs in the structure of its radii only in degree from certain varieties of C. stellatus. Lastly, C. intertextus, in its peculiar radii, closely resembles C. scabrosus, and this latter species does not differ much from the other species. Hence the genus Chthamalus has no claims to be subdivided into smaller genera.
Range.—The species are found all round the world, from (as far as I have seen) 54°-55° north, to Cape Horn, in 55°-56° south. All the species, of the habits of which I know anything, are littoral; and in many parts of the world are excessively numerous, quite covering large spaces of the coast-rocks, and often coating the coast-shells. Chthamalus dentatus is littoral, like the other species; but it often lives attached on Balanus tintinnabulum and amphitrite, on the bottoms of ships arriving in British ports from the west coast of Africa. I do not know of any instance of more than two species occurring in the same region. Some of the species have large ranges: C. scabrosus extends from the Falkland Islands and Tierra del Fuego to Peru; and C. stellatus has an enormous extension over almost the whole world, excepting the west coast of South America and Australia. I do not believe any species of the genus, owing probably to their littoral habits, have hitherto been found fossil.
LEPAS STELLATA. Poli. Testacea Utriusque Siciliæ (1795), Tab. 5, fig. 18-20.
---- DEPRESSA (var.) Ib., Tab. 5, fig. 12-16.
CHTHAMALUS STELLATUS. Ranzani. Memoire di Storia Naturale (1820), Tab. 3, fig. 21-24.
-------- GLABER (var.) Ranzani. Ib.
-------- STELLATUS. Philippi (!). Enumeratio Mollusc. Siciliæ.
LEPAS PUNCTATUS. Montagu (!). Testacea Britannica (1803).
Shell white or gray, generally much corroded and punctured: radii (when present) narrow, with their sutural edges most finely crenated; tergum with the crests for the depressor muscle depending barely beneath the basal margin.
Var. (a, communis) fig. 1 a, 1 f: Shell conical depressed, upper part corroded, walls folded, sutures moderately plain or obliterated; radii not developed; orifice broadly oval.
Var. (b): Shell elongated, sub-cylindrical; sutures obliterated; surface much corroded; orifice almost circular.
Var. (c, communis) fig. 1 c, 1 e, 1 h: Shell conical, folded, sometimes covered by membrane; radii developed, narrow; orifice sub-hexagonal, toothed.
Var. (d, fragilis) fig. 1 d: Shell conical, smooth, thin; compartments easily separable, pale-coloured; radii developed, narrow; orifice large, toothed, sub-hexagonal.
Var. (e, depressus) fig. 1 b, 1 g, 1 h: Shell much depressed, surface much corroded, smooth: alæ largely exposed, marked by lines of growth: radii not present; parietes on the under side often supported by pillars; orifice sub-hexagonal.
Hab.—Southern shores of England, Ireland, Isle of Man, Mediterranean, Madeira, Cape de Verde Islands, Southern United States (Charlestown), West Indies, Brazil (Bahia), Rio Plata (Guritti Island), Red Sea, Philippine Archipelago, Coast of China, Gulf of Corea, Oregon or Northern California.
This species is very widely distributed and extremely common. On the coast-rocks of the southern shores of England it is, in parts, even more numerous than the Balanus balanoides, with which it often grows mingled. As already stated, it is often confounded in British collections with this species of Balanus, under the name of Balanus punctatus; that Montagu had this Chthamalus in view, when describing his Lepas punctatus, is certain, from his original specimen in the British Museum, but whether this was the case with his predecessor, Pulteney, in the Dorset Catalogue, I do not feel so sure. Chthamalus stellatus varies, as we shall immediately see, extremely in appearance. Some of the varieties, as var. depressus, which in external aspect are especially distinct, I have no doubt are really varieties, but whether this is the case with some of the forms from the more distant localities, is a little more doubtful; but I beg that it may be observed, that I have, in the case of every one of the varieties, and of all the specimens from distant localities, cleaned with potash and most carefully examined the disarticulated valves, and likewise dissected the included animal’s body.
General Appearance.—British specimens are usually conical, and have their walls folded, corroded in their upper parts, with the radii not developed, and the sutures more or less obliterated; in this state the orifice of the shell is entire, and very broadly oval: I have seen specimens in this condition from Madeira, Brazil, and the Gulf of[Pg 457] Corea. The shells when crowded, are rendered cylindrical, and more or less elongated, with the sutures as viewed externally quite obliterated; the surface rugged and much disintegrated; and the orifice nearly circular: in this extreme condition I have seen specimens from England, from near Genoa, and from the mouth of the Plata: the specimens from these last two localities were of remarkable size, being half an inch in diameter, and rather more than half an inch in height. Again, other British specimens (fig. 1 c), though not nearly so common, are rather steeply conical, and have not only their sutures distinct, but narrow radii are plainly developed on apparently both sides of the sutures; in this case the orifice is slightly toothed, and is rather elongated: I have seen specimens in this condition, but with their walls rather more deeply folded, from the Cape de Verde and Philippine Archipelago. Other specimens, from some unknown tropical sea, differed only in the walls being but slightly folded, and being so perfectly preserved as to be externally covered with membrane: this latter circumstance gave the specimens a peculiar appearance. In this condition were some specimens (fig. 1 d) attached to oysters sent to me by Professor Agassiz, from Charlestown; and which differed from all the others that I have seen, in the thinness and smoothness of their compartments, and in the facility with which the compartments separated from each other and from the surface of attachment: hence I have called this form, which, until finding more important differences, I must rank as a variety, fragilis: it has stronger claims than the other varieties to be specifically separated; but I suspect that it has been exposed to some peculiar modifying conditions.
Lastly, we have the variety (fig. 1 b) excellently figured and described by Poli, under the name of L. depressa: this is much depressed, with the walls thin, not folded, with the surface much corroded, with the sutures very distinct, with the radii not at all or barely developed, but with the alæ largely developed, and marked with lines of growth, resulting from the diametric growth of the shell: the orifice is hexagonal, but broadest towards the carinal end: most of the specimens, but not all, have on the under sides of their compartments rugged pillars depending from beneath the alæ, for the purpose, apparently, of supporting the much depressed shell. This much depressed variety attains a larger basal diameter (but not a greater bulk or internal capacity) than any other variety, namely, sometimes three quarters of an inch. The great peculiarities of this variety result, apparently, from its much depressed form, deeply corroded not folded walls, and considerable diametric growth; from the latter cause the alæ are largely developed; as I can find neither internally nor externally any fixed diagnostic character I have not hesitated to rank this form as a variety. Poli found his specimens mingled with the ordinary C. stellatus, on the shores of Sicily; and I collected at St. Jago, in the Cape de Verde Archipelago, some specimens nearly as well characterised, also associated with the common variety.
The shell itself is dirty-white or gray, or brown: in some varieties, however, the white is nearly pure: internally the parietes are generally tinted dull purple. The corium of the sack is dark greenish-black,[Pg 458] with a white edge to the lips lining the aperture between the opercular valves.
Structure of Shell and Radii, &c.—The under side of the parietes is either quite smooth, or marked with slight, branching, depressed lines; or mamillated; or irregularly studded with large pores. I have already alluded to the pillars, depending from the under sides of the alæ in most specimens of var. depressus: these pillars tend to form ridges, parallel to the sides of the compartment, like those we shall presently see in certain specimens of C. scabrosus. The radii are very narrow, when best developed; their edges, when disarticulated, can be seen, when examined by a strong lens, to be finely crenated. The edges of the alæ are likewise very finely crenated.
Scuta.—The outline of these valves varies considerably in specimens taken out of the same group: we have either a nearly equilateral triangle (fig. 1 f) or the tergal margin (1 e) is much shorter than the other margins. There is always a deep depression for the adductor muscle, and a small pit of very variable depth for the lateral depressor. But the tergal margin offers the greatest variability; here we see a very prominent articular ridge or fold, having either a straight edge or a single or double prominence (fig. 1 e-1 h). In specimens in the same group we find considerable variation in these points; but the amount of difference is sometimes so great, that I long hesitated whether to rank some of the varieties as species. The Terga, likewise, vary greatly in shape and width: in some of the commonest varieties (and in var. depressus) the valve is very narrow, with the under surface channelled or concave: in other varieties the valve is much broader and flatter. The spur is but slightly developed. The crests for the depressor muscles barely descend beneath the basal margin of the valve. The articular ridge, in some varieties (as in specimens from Madeira and the Cape de Verde Islands), is extremely prominent and straight (1 f); in others, it is little prominent and deeply sinuous (1 h). This great variability in the articular margins of the scutum and tergum seems to be mainly due to the corrosion to which these valves have been subjected, and their consequently modified growth: in some specimens the articular ridge of the scutum, and in others that of the tergum, has been largely developed, in either case their mutual outlines being greatly affected.
Branchiæ.—These are narrow, hardly at all plicated, elongated, being about half as long as the sack. In a specimen from La Plata, this organ ended rather more abruptly in a point than it did in other specimens.
Mouth.—The crest of the labrum is usually hairy, but in a specimen from Bahia (Brazil) there were some very fine teeth. The palpi vary somewhat in shape, and sometimes have a row of bristles along their basal exterior margins. The mandibles usually have four main teeth, the lowest one being confluent with the inferior pectinated angle: in var. depressus, in the same individual, there were only three teeth on one side of the mouth and five on the other; the lower main teeth are laterally double, but generally one tooth of each pair is so small and obscure as to be perceived with difficulty. In the maxillæ, there are some large spines above the notch, and in the notch some fine ones:[Pg 459] in var. depressus, and in a cylindrical var. from La Plata, there was quite a tuft of small spines above the notch.
Cirri.—The outer surface of the pedicel of the second cirrus bears a tuft of long, fine, plumose hairs: the terminal segments of the rami of this cirrus sometimes (as in the La Plata specimens) support a clump of coarsely pectinated spines. In specimens having six segments in the shorter ramus of the second cirrus, the shorter ramus of the third cirrus had fifteen segments. The two rami of the third cirrus are usually equal in length and in the number of their segments; but in the Brazilian specimen there were fifteen segments in the posterior, and twenty-six in the anterior ramus; in another specimen, fixed on a tropical Perna, there were in the two rami of this third cirrus eighteen and twenty-four segments. In the three posterior pairs of cirri each segment carries either four or five pairs of main spines: the segments vary a little in the degree to which they are elongated, being most elongated in the var. from La Plata, with an elongated shell.
Varieties.—It will have been observed, that the shell, in the specimens from several distant quarters of the world over which this species, as I believe, ranges, differs considerably in external aspect: so do the opercular valves; and so do the parts of the mouth and cirri: but I cannot make out that these differences are coordinated. Thus, var. depressus, which is so entirely different from the others in appearance, differs only internally in the presence of a tuft of fine spines above the notch of the maxillæ; and this character is found in the La Plata variety, which, as far as the shell and opercular valves are concerned, is at the other end of the scale of variation. Again, var. fragilis, from Charlestown, presents, in the animal’s body, hardly any difference. The Brazilian specimens, which in the shell and operculum offer only quite common characters, have the remarkable peculiarity of a considerable difference in the length and number of the segments in the rami of the third cirrus; they, also, have the segments of the sixth cirrus considerably elongated, and the labrum finely toothed. Of these peculiarities one alone, namely, the inequality in the rami of the third cirrus, but in a lesser degree, is common to the specimens adhering to the tropical Perna, which had a shell very unlike the Brazilian variety, but which, on the other hand, differed scarcely in a single character from some other specimens from an unknown tropical sea, in which the rami of the third cirrus were quite equal. The La Plata specimens differ most in internal characters, viz., in the tuft of fine spines above the notch of the maxillæ, in the coarsely pectinated spines on the tips of the second pair of cirri, in having the segments of the sixth cirrus much elongated, and in the apex of the branchiæ being abruptly pointed; yet in the shell and operculum they were identical with certain Mediterranean varieties. From these several facts, I must believe that all the widely distributed forms here grouped together, do really belong to the same species.
Shell conical, generally smooth: when not deeply corroded of a pale dirty flesh-colour: sutures always distinct: radii, when present, with their sutural edges quite smooth.
Hab.—New South Wales, (Moreton Bay, 27° S.; Sydney; Twofold Bay), Van Diemen’s Land (Hobart Town). Attached to littoral rocks and shells; Mus. Brit., Cuming, Darwin.
General Appearance and Structure of Shell.—Shell conical, rather smooth; when not much corroded, of a pale dirty flesh-colour; often covered by membrane; sometimes deeply corroded, extremely rugged, and then of a brown colour; in this condition not much punctured, as generally is the case with C. stellatus. Sutures almost always quite distinct; rarely the shell becomes cylindrical with the sutures obliterated. Orifice moderately elongated, sub-hexagonal. Radii rather narrow, but not so narrow as in C. stellatus, smooth, with their upper margins very oblique: when disarticulated their edges are quite smooth. The edges of the alæ are sometimes crenated, and sometimes not so, being only marked by lines of growth; they are often rather thick. The parietes are usually rather thick, with their internal surfaces smooth, and not mamillated, as is so often the case with C. stellatus. The largest specimens which I have seen, were .6 of an inch in basal diameter.
Opercular Valves.—These are hardly distinguishable from those of C. stellatus. The only very slight difference which I can point out is, that the crests for the tergal depressores are less spread out, and depend rather more beneath the basal margin of the valve; and lastly, that the surface of the tergum, just above these crests, is rather prominent.
Branchiæ: these are oblong; taper but little, and have a broad rounded end: they are scarcely plicated.
Mouth.—The crest of labrum is hairy: the palpi are square, and have no bristles along their basal exterior margins, but long ones at their truncated ends. The mandibles have three or four main teeth apparently single: the inferior coarsely pectinated portion is short. The maxillæ are deeply notched.
Cirri.—The first and second pairs, and portions of the third, are darker coloured than the three posterior pairs. The rami in both of the first two pairs are slightly unequal in length. The third cirrus is much longer than the second: in a specimen in which there were six segments in the shorter ramus of the first and second pairs, there were twenty segments in the posterior and shorter ramus of the third cirrus; and in this same individual there were no less than forty-nine segments in the anterior ramus. In another specimen (Pl. 29, fig. 2) the number of segments in the two rami of the third cirrus, was 20 and 41; in another, the numbers were 18 and 53; in several other specimens the[Pg 461] numbers varied in about these proportions; but in one single specimen the numbers were equal. Not only did the number of segments thus vary in the two rami, but likewise the arrangement of the spines on the segments in the anterior and longer ramus; in some specimens the spines on all the segments were arranged in a single circle, and then the organ had a specially antenniformed appearance: in other specimens, some of the lower segments (in one case thirteen in number) had the spines placed in regular pairs precisely as on the posterior ramus, and as on the three posterior pairs of cirri. Under the genus I have pointed out the resemblance between this structure and that occurring in certain Macrourous Crustaceans. The pedicel of the third cirrus had its spines more crowded and irregular than on the three posterior pairs of cirri. The segments in the latter vary in bearing either three or four pairs of main spines. The whole dorsal surfaces of the lower segments of the several posterior cirri are serrated in an upward direction by short spines, but to a variable degree.
Shell white or gray: sheath and opercular valves generally clothed by fimbriated membrane: tergum, with its basi-carinal angle depending and pointed.
Hab.—Peru, Chile, Chiloe, Northern Chonos Islands. Attached to littoral rocks, and sometimes to littoral shells, often mingled with Chthamalus scabrosus; Mus. Brit., Cuming, Darwin.
General Appearance and Structure of Shell.—Shell dirty white or gray: sometimes tinted pale purple within; irregularly conical, or much depressed, or cylindrical and much elongated. Generally much corroded, sometimes well preserved and covered by membrane. Orifice rather large, of variable shape. Sutures often quite obliterated. Radii when developed narrow, with their sutural edges, as well as those of the alæ, generally very finely crenated, but to a variable degree. The membrane lining the sheath and covering the opercular valves, is remarkable from each zone being fimbriated; for this expression is more correct than to say that the membrane bears a row of spines, though the fimbriæ do approach in character to spines; sometimes, though rarely, the fimbriæ are branched. The largest specimens which I have seen (from Coquimbo and Valparaiso) were half an inch in basal diameter, and some of these were so much elongated as to be one inch in height.
Scuta.—The scuta are rather narrow: they have a somewhat peculiar appearance, from the articular furrow being wide, and from the articular ridge projecting with a uniform curvature: the pit for the lateral depressor muscle has some minute crests, of which I have seen traces in the foregoing species. The Terga vary somewhat in shape:[Pg 462] they have the basi-carinal angle of the valve, where the narrow crests for the depressores are placed, pointed and dependent, and the surface of the valve above these crests is prominent. Altogether the opercular valves have a sufficiently distinct character to be recognised without much difficulty.
Mouth.—The crest of the labrum is not toothed; the palpi have long hairs along the exterior basal margin. The lower main teeth of the mandibles are plainly laterally double. Cirri.—The pedicel of the second cirrus is extremely broad, and on the exterior margin supports a tuft of very long, finely plumose spines: in some specimens each of the lower segments of the anterior ramus of this cirrus bore one or two very large spines, doubly and extremely coarsely pectinated. In two specimens the rami of the third cirrus were of equal length: but in one specimen (from Iquique, Peru), having seven segments in the shorter ramus of the first and second pairs of cirri, the posterior ramus of the third pair had fourteen segments, and the anterior ramus twenty-two segments. The posterior cirri have segments carrying five pairs of main spines: the dorsal surfaces of the lower segments are serrated.
Shell brownish, plicated; orifice twice as long as broad: tergum triangular, equilateral, with the basal and carinal margins slightly protuberant.
Hab.—California, attached to Lottia grandis. Peru(?); Mus. Brit.
General Appearance and Structure of Shell.—Shell globulo-conical, irregular, with the walls much folded. Colour brownish. Sutures generally distinct in young specimens, and nearly obliterated in full-grown individuals. Radii, when present, very narrow. The orifice of the shell, in full-grown specimens which have their summits a little worn, is narrow, much elongated, about twice as long as wide, oval or ovate, with the rostral end the narrowest: the elongation of the orifice offers almost the only character by which this species can be externally recognised. The lateral compartments are rather wider than usual in proportion to the rostro-lateral compartments. Basal diameter of largest specimen .2 of an inch.
Scuta, elongated transversely, with the pit for the adductor muscle bordered on the under side by an adductor ridge rather more prominent than usual in this genus. Terga, triangular, equilateral, with the margins slightly curved and protuberant: the basal margin is regularly and equably curved from one end to the other.
Neither the Mouth nor Cirri offer any peculiar characters distinct from the genus. I may mention, however, that the crest of the labrum is toothed, and that the segments of the posterior pairs of cirri support five pairs of main spines.
The characters by which this species differs from C. stellatus and from the other species, consist almost exclusively in the triangular and equilateral terga, and in the much elongated orifice of the shell; and these differences I believe to be of specific value. I must, however, confess that I have examined one young specimen attached to a Pollicipes elegans, in which the orifice was not nearly so much elongated, and in which the terga were not so equilateral, with the basal margin not quite equably curved, but more protuberant on the scutal than on the carinal side: from an examination, however, of only one specimen, and that a young one, I cannot decide on its specific nature.
CHTHAMALUS DENTATUS. Krauss (!). Die Südafrikanischen Mollusken, 1848, tab. 6, fig. 27.
Shell dirty white or brownish: sutures formed by interlocking teeth: tergum with the carinal margin protuberant.
Hab.—South Africa, Natal; West Africa, Loanda and the Gold Coast; West Indies (?). Attached to ships’ bottoms and to littoral shells, and to Tetraclita serrata, Balanus perforatus, and amphitrite; often attached to Balanus tintinnabulum and amphitrite on ships’ bottoms.
General Appearance and Structure of Shell.—Shell dirty white, pale brown, or gray: conical, moderately depressed: walls either broadly and irregularly folded (fig. 3 a), with the surface corroded, or (when attached to ships’ bottoms and sometimes to other Cirripedes) narrowly and regularly folded (3 b), with the surface well preserved, smooth, and generally covered by thin brown membrane. These latter specimens generally have the shell more steeply conical, with the orifice rather smaller, and the radii broader, than in the first-mentioned specimens, which are attached to coast-rocks and shells, and have had their summits worn down. The sutures in all cases are tolerably distinct, and have their edges toothed and interlocked: the crenations are visible before the compartments are disarticulated, when viewed either internally or externally, but occasionally they are obscure. The radii, when best developed, are rather narrow, and of equal width on both sides of the sutures, with their summits rounded: their surfaces are finely ribbed transversely, each rib corresponding with, or rather forming a point of, the toothed edge. On the under side these teeth usually are a little[Pg 464] hollow or are pitted: the radii, in fact, may be said to be folded, like the parietes, only more symmetrically and narrowly, so that the points in the opposed edges interlock. The edges of alæ are serrated, but more finely than the radii. The parietes are rather thin, with their under surfaces generally smooth. The sheath does not descend far down the shell. The colour of the corium lining the sack and the animal’s body varies considerably, being either almost black or pale purple; and the specimens adhering to ships’ bottoms are internally almost white. Of these latter specimens, I have seen some .6 of an inch in basal diameter: of corroded specimens attached to littoral shells, I have not seen one quite .4 of an inch in basal diameter.
Scuta, with the articular ridge very prominent: the pit for the adductor is deep, and there are generally some distinct, though minute, pits for the lateral depressores. The Terga (fig. 3 c) have the articular ridge very prominent; and the carinal margin is rather more arched and protuberant than in the other species.
Mouth.—The crest of the labrum is toothed: the palpi are short, truncated, with long spines arising from their ends, and along the basal exterior margin. In the mandibles the first tooth is seated rather far from the succeeding teeth: the inferior part is coarsely pectinated: the maxillæ are deeply notched. The tips of second pair of Cirri have many coarsely pectinated spines: the shorter ramus had six segments, whilst the shorter ramus of the third pair had eighteen segments: the segments on the posterior cirri carry five pairs of main spines.
Branchiæ, &c.—These consist of a double fold, the outer one being the largest, and the inner semi-circular, as has been described under the genus. In a young specimen, only one tenth of an inch in basal diameter, the branchiæ consisted of a single fold; in a specimen a little larger, there were two folds, but these were of equal size: ultimately the outer fold increases in size so as to become nearly double the inner fold. The Ovarian tubes are remarkable from their large diameter. I was surprised to observe in the specimen only one tenth of an inch in basal diameter, that the larvæ were ready to escape. On the prosoma, there are some longish hairs arranged in short rows, and some few on the membrane lining the sack, and some on the opercular membrane and valves.
It may be seen in Pl. 18, fig. 3 a, and 3 b, that I have here united two varieties considerably different in external aspect: I have done this without hesitation, inasmuch as there are intermediate forms, and as the differences are analogous with those so commonly occurring in sessile cirripedes; dependent on whether or not the specimens have been exposed to corrosion. I have seen both varieties from Natal, and both from the west coast of Africa; although the smooth, well-preserved, narrowly plicated varieties seem more common in western than in southern Africa. With respect to the range of the species, I have seen a specimen[Pg 465] from the West Indies, but it was the variety which so commonly adheres to ships’ bottoms. This variety often arrives alive in British ports; and I have seen a specimen picked up dead on the beach near Dublin.
EURAPHIA HEMBELI. Conrad. Journal Acad. Nat. Sc. Philadelphia, vol. 7, 1831, Pl. 20, fig. 6.
Shell dull reddish purple: sutures, when not obliterated, formed by interlocking teeth: basis sometimes rendered calcareous by the inflexion of the parietes: scutum with two or three furrows extending down the middle of the valve.
Hab.—California, near S. Diego, according to Conrad. Mus. Brit., Cuming.
I have seen five, old, large specimens, from an unknown locality, with their whole surfaces deeply corroded, and with most of the sutures obliterated; and three separated valves of a young specimen. From these materials, imperfect as they are, I feel no hesitation in identifying this species with the Euraphia Hembeli of Conrad, which is remarkable in several respects, and especially from being gigantic in size, compared to other members of its sub-family.
General Appearance and Structure of Shell.—The young specimen (fig. 5 b, about .7 of an inch in basal diameter) consists only of the carina, and the two lateral compartments; but these, as far as I can judge, resemble the specimen figured and described by Conrad, which was two inches in diameter. Shell depressed, spreading, surface moderately smooth, covered by brownish membrane: shell itself pale dull reddish-purple. The radii are not very narrow, with their summits rounded and very oblique: their edges are toothed, and their external surfaces are transversely ribbed, in correspondence with the interlocking points of the sutures. On the internal surface, the toothed suture is not visible, except near the base of the shell, owing to the overlapping of the alæ. The alæ have oblique summits, which are slightly notched owing to the upturned prominent lines of growth. The parietes are thick; their basal internal surfaces are rugged, with slightly branching ridges.
The old specimens (fig. 5 a) are so much corroded that not a particle of[Pg 466] the external surface is left: one of them which was 2-3/4 of an inch in basal diameter. Shell much depressed, spreading; colour pale purple; orifice large, rhomboidal, with a slight hollow on each side for the corners of the scuta: sutures generally in part or wholly obliterated; where still preserved, the interlocking toothed structure is distinct; the sheath is strongly marked by lines of growth, and is of a dark brown colour. The carina and rostrum are of unusually large size compared with the lateral compartments; and this, as far as I can judge, must have been the case with the younger specimens. That portion of the rostro-lateral compartment which forms part of the sheath, is reduced to a mere ridge. The most remarkable character is, that all these old specimens (of which the smallest measured nearly one inch and a half in diameter) had a flat, wide, calcareous basis, which is absolutely continuous with the inner lamina of the parietes, whereas in the younger specimen there was no appearance of any tendency in the parietes thus to grow inflected. There can be hardly any doubt that in a series of specimens some would be found with the parietes first forming a flat narrow ledge round the true basal membrane (as in the following species); and that in others, this ledge would be wider and wider, till its edges met in the middle, and coalesced into a continuous plate.
Opercular Valves.—I have seen these only in the old corroded specimens (fig. 5 c, 5 d): they are locked together by remarkably strong articular ridges and furrows.
The Scuta have externally two or three impressed lines or narrow furrows, proceeding from the apex to the middle of the basal margin; these can be seen only in one of my specimens, owing to the degree to which the valves have suffered disintegration; but they are mentioned by Conrad. The basal margin is rather short compared with the other two margins. The tergal margin is remarkable from the depth of the upper furrow above the articular ridge, and from the size of the prominence (appearing like the true apex of the valve) above this upper furrow. There is a hollow for the adductor muscle, and traces of crests for both the rostral and lateral depressores. The Terga are generally but little corroded, and hence the dark brown membrane with which they are covered is well preserved; the shelly matter itself is also brown: there is only a trace of this colour in the more corroded scuta: the external surface of the terga is very smooth. The spur is pretty well developed, and is half as wide as the whole valve. The lines of growth are upturned along the carinal margin. The articular ridges and furrows are much developed. The crests for the depressor muscles are extremely strong; they depend beneath the basal margin, and are remarkable (fig. 5 c) from being furnished each with fine sub-crests.
Animal’s body, unknown.
Shell, when well preserved, violet-purple: sutures, when not obliterated, formed by oblique interfolding laminæ: basis membranous, but surrounded by a ledge formed by the inflected basal edges of the parietes: scutum and tergum completely calcified together.
Hab.—Philippine Archipelago; Mus. Cuming.
General Appearance and Structure.—Shell depressed, with a large diamond-shaped orifice. Colour beautiful violet-purple, but externally much obscured by disintegration, causing the shell to be ashy gray. Walls smooth or slightly folded. Sutures generally quite, or almost quite obliterated; but when well preserved, they differ remarkably in appearance from those in the foregoing species; for the radii externally here consist of oblique plates or laminæ arising on both sides of the sutures, standing nearly parallel to the parietes, and interfolding with each other. These laminæ are rather plainly marked by lines of growth. Essentially the radii do not differ much from those in C. dentatus and Hembeli; we have but to produce obliquely upwards the transverse and interlocking ribs on their radii, and so convert them into laminæ. During the diametric growth of the shell, the sutural edges of the alæ are added to, in the usual manner, by upturned lines of growth; and, in addition, the recipient furrows of the alæ are similarly added to, so that the lines of growth are upturned, and alæ appear to have been developed on both sides of the sutures in the same way as the radii appear to have been developed on both sides in many Chthamalinæ, though rarely in the Balaninæ. The inside of the shell is beautifully coloured rich violet; it is punctured with small holes as so often is the case with C. stellatus. In every specimen (all full-grown) which I opened, the inner basal edges of the parietes were inflected rectangularly inwards, forming a smooth-edged ledge all round the basal membrane, which, in proportion to the width of this ledge, was by so much reduced in diameter. The largest specimen which I have seen was .35 of an inch in basal diameter.
Opercular valves.—The scuta and terga, in all the specimens which I have seen, were firmly calcified together; in some, a trace of a suture could be seen externally, but hardly a trace internally. In one specimen, there were vestiges of some impressed lines on the scutum, in exactly the same position in which such occur in C. Hembeli. The scutum is rather narrow. The basal margin of the tergum is either straight, or depends a little on the scutal side, thus producing a small spur: the crests for the depressor muscles are strongly marked, and depend beneath the basal margin.
Mouth.—The labrum is strongly toothed: the palpi have long hairs along the exterior basal margin: the mandibles have only three main teeth, and the inferior coarsely pectinated portion is short; the maxillæ deeply notched. Cirri: the first and second pairs have their rami slightly unequal in length; the third pair differs from the same pair in the other species of the genus, in having some few of the basal segments on the anterior ramus thickly clothed with spines, so as to be brush-like: there is even a trace of a similar structure in the lowest segments of the posterior ramus. In the three posterior pairs of cirri the segments are much elongated, and support four pairs of spines.
Branchiæ.—Unknown.
Shell (when well preserved) dull purplish-brown: sutures formed by oblique interfolding though rarely well developed: tergum with a deep narrow pit, at the basi-carinal angle, for the depressor muscle.
Hab.—Peru, Chile, Tierra del Fuego, Falkland Islands. Very common; attached to littoral rocks and shells, and often associated with Balanus flosculus, and sometimes with Chthamalus cirratus; Mus. Brit., Cuming, W. Dunker, Darwin.
General Appearance and Structure.—Shell generally depressed; when growing crowded, sometimes cylindrical: colour dingy purplish-brown, but when much corroded, dirty gray; very young shells are very dark green, owing to the corium, which is of this colour, being seen through the valves. Surface generally rugged, from irregular slight longitudinal folds, and from the transverse overlapping tile-like lines of growth; but sometimes the surface is nearly smooth, with very slight longitudinal folds, these being gray coloured, the intermediate parts being pale dingy purple, the shell thus becoming striped. Orifice rhomboidal, passing into trigonal, owing to the great width of the carinal end. Sutures generally very distinct, rarely obliterated in the cylindrical varieties. Radii narrow, generally exposing much of the alæ, which are plainly marked by lines of growth: the radii themselves, when well developed, which is not often the case, consist of small laminæ or ridges, placed on both sides of the sutures, and interfolded or interlocked together: usually only a trace of this structure is exhibited, but occasionally, along some or along all the sutures (as in the specimen figured), the laminæ of the radii interfold, as plainly as in C. intertextus. The alæ differ slightly from the alæ of the other species, in not forming so much of a rectangular projection, the lower margin running with a gentle curve into the parietes. The internal surface of the parietes is either smooth, or near the basal margin is roughened with depending points: in some[Pg 469] specimens from the Falkland Islands, both edges of each suture were inflected, forming a double ridge, with roughened edges, resting on the basal membrane, and supporting the shell. I must mention that in my notes made at these Islands, I remark that the basal membrane seemed sometimes to be surrounded by a calcareous rim; none of the specimens brought home are thus characterised; but bearing in mind the affinity of this species to C. intertextus, no doubt we have here an indication of the shelly ledge surrounding the basis, as described under that species. The largest specimens which I have seen are a quarter of an inch in basal diameter.
The Opercular Valves generally have their summits much worn down. The scuta are elongated in the line of the longer axis of the orifice; the articular ridge is very prominent, and is placed in the middle of the tergal margin. The terga are very narrow, as in some varieties of C. stellatus: they are remarkable in two respects, namely, in the depressor muscle being attached to a plate, formed apparently by the union of the usual crests, parallel to the outer lamina of the valve itself, a deep narrow cavity (fig. 2 d, p) being thus formed; and secondly, in the far more extraordinary circumstance of the existence of a small pit (q) at the extreme basi-scutal corner of the valve, in which about half of the scutal lateral depressor muscle is attached: I have observed no other instance in any cirripede of the partial attachment of a muscle properly belonging to one valve to another valve. The figures of the valves 2 b, 2 c, 2 d, are from specimens most unusually perfect, with the upper portion not worn away; the ordinary appearance of the valves as seen from above, is given in fig. 2 a; at fig. 2 d, a view is given of the tergum seen from vertically beneath, showing the cavity for its own depressor muscle, and for part of the lateral depressor muscle of the scutum.
Mouth.—The crest of the labrum is hairy, without teeth: the palpi have long spines at the end, but none along the inferior margin: the mandibles have either four or five graduated teeth, the lower ones of which are plainly double laterally: the maxillæ have a very sinuous edge. Cirri: the first and second pairs are very short: on the four posterior pairs the segments support either four or five pairs of main spines, with the small intermediate spines rather larger than usual.
Branchiæ.—None; but where they ought to occur, there are two very slight ridges clothed with hairs, about the 1/100th of an inch in length. On the prosoma, there is a slight ridge, extending from the base of the first cirrus towards the adductor scutorum muscle, also clothed with hairs; this unusual character of the prosoma being hairy is common to C. dentatus.
This species is the commonest cirripede on the shores of the Falkland Islands: many of the specimens are there crowded together, and rendered elongated and cylindrical, with the walls very thin, and the sutures often obliterated; as the opercular membrane is very narrow, the opercular valves are much influenced both in their outline and in their[Pg 470] crests and articulations, by the varying form of the shell: I have even seen specimens with the scutum and tergum on one side twice as large as on the other side.
[132] Χαμαι [Greek:Chamai], on the ground, and σιφων [Greek:siphôn], a tube.
Compartments four, with the sutures often much obliterated: basis membranous.
Distribution, Australia, China (?). Attached to littoral shells and rocks.
The two species united under this genus agree in having only four compartments, and in these having a strong tendency to become confluent; but they resemble each other hardly in any other respect, more than do all the species of the present sub-family. Chamæsipho columna differs from all the other Chthamalinæ in the structure of its second pair of cirri, and C. scutelliformis differs from all in its shell,—namely, in the small size of the rostrum, with its alæ but little developed, and in the very peculiar apertures in the three other compartments. Hence this genus can hardly be considered a very natural one, though I could not introduce the two present species into Chthamalus, or into any other genus, without doing still more violence to the principles of classification followed throughout this work. Chamæsipho bears nearly the same relation to Chthamalus, as Tetraclita and Elminius do to Balanus.
LEPAS COLUMNA. Spengler. Skrifter Naturhist. Selbskabet, b. 1, (1790), Tab. 6, fig. 6.
Sutures, excepting during early youth, generally obliterated both externally and internally: tergum with small pits for the attachment of the depressor muscle.
Hab.—New South Wales, Tasmania, New Zealand; extremely common; attached to littoral shells and rocks; often associated with Chthamalus antennatus and Elminius modestus; and in New Zealand, often thickly coating Elminius plicatus.
I have identified, with some doubt, the present species with the Lepas columna of Spengler, obtained from Otaheite, as no description is given by him of the opercular valves, and more especially as Spengler’s specimens were an inch in height and seven lines in width, which is much larger than any of the many specimens seen by me: from Spengler’s clear description of the structure of his shell, it evidently belongs to the present genus.
General Appearance and Structure of Shell.—The sutures separating the four compartments are generally, excepting at an early age, quite obliterated, both internally and externally, the shell in this case consisting of a single piece, with its summit and opercular valves always much worn. Occasionally the sutures are preserved, and then the four compartments are seen to be of nearly equal sizes. The orifice is always broadly oval, with the carinal end the broadest; and it often approaches closely to circular. The upper part of the shell is frequently steeply conical, with the lower part spreading and folded; sometimes deeply folded. Very young shells are apt to be remarkably smooth. The radii appear never to be developed: the inside of the shell is smooth. The colour is either blackish-green (this being the tint of the corium lining the sack), with the upper part gray from disintegration; or the lower part is brown, from the investing membrane, the shell itself being pale coloured. This species seems particularly liable to grow crowded together; often covering rocks and shells with a honey-combed layer. The basal diameter of some of the largest specimens was .3 of a inch. In Mr. Cuming’s collection, however, there is an Australian specimen .55 in diameter and .3 in height, which is, moreover, remarkable from the projecting, extremely rugged, overlapping, dark-coloured layers of growth, which surround the lower part of the shell: we have seen that Spengler’s specimens, said to have come from Otaheite, are even broader and considerably higher.
Scuta: from the disintegration which the valves have undergone, the scuta and terga are externally seen to be locked together by a deeply sinuous articulation. The Scuta have a wide articular furrow and a very prominent articular ridge; but the exact outline of these parts varies greatly, very much as in the genus Chthamalus.
The Terga are very narrow, with the under surface channelled: the attachment of the depressor muscle offers the only peculiarity,—the muscle being attached to four or five little pits, placed transversely to the longer axis of the valve; the septa between these little pits evidently answering to the crests as usually developed.
Mouth.—The crest of the labrum is hairy, and slightly bullate.[Pg 472] The palpi are rather small, with long spines at their ends. The mandibles have four (sometimes five) teeth, with the inferior portion pectinated. The maxillæ are notched.
Cirri.—The first pair presents no remarkable character. The second and third pairs are subject to extreme variation, as in Chthamalus antennatus and Tetraclita porosa. In all the specimens, the anterior ramus of the second cirrus is short, with all the segments thickly covered with bristles; it is the posterior ramus which varies so much in relative length and in the arrangement of the bristles; but in no case are all the segments clothed with bristles as on the anterior ramus, and as is normally the case with all sessile cirripedes. In some specimens from New Zealand, the anterior ramus having only five segments, the posterior ramus was twice as long, having sixteen segments, with the bristles arranged in circles, but standing rather thicker together on the basal segments. In a Tasmanian specimen, the posterior ramus was only a little longer than the anterior ramus, and the spines were arranged in regular pairs (as on the three posterior pairs of cirri) on all the segments, excepting the few basal ones, on which they were more crowded: a nearly similar arrangement occurred in some other specimens from unknown localities, excepting that the rami were of nearly equal length. In the third cirrus, in all the specimens, the anterior ramus has three or four of its basal segments much broader than the upper segments, and thickly clothed with spines (as is the case with one species of Chthamalus, viz. C. intertextus) all the other segments having regular pairs of spines. The posterior ramus of the third cirrus varies in being either much longer than, or only equal in length to, the anterior ramus; in the former case (in the New Zealand specimen) the spines were arranged in circles, giving an antenniformed structure to the ramus; and in the latter case they were arranged in regular pairs. In Mr. Cuming’s great Australian specimen, there was a further peculiarity, in the presence on the posterior cirri of a tuft of intermediate spines between the main pairs; and, in there being on those segments, which are thickly covered with spines, certain very large spines, doubly pectinated, with the pectinations elbowed, closely like the spines met with on the cirri in certain species of Pollicipes. Finally, the segments in the three posterior pairs of cirri support five or six pairs of main spines; the dorsal surfaces of the segment are rough and hairy.
Branchiæ, rudimentary; consisting of a small, simple, tongue-formed fold, projecting about 1/100th of an inch. Ova, 16/2000ths of an inch in length.
Rostrum very small, elongated, triangular: lateral compartments, each with an aperture, and carina with two similar apertures, all four leading into shelly tubular columns.
Hab.—Attached to Pollicipes mitella, probably from the seas of China; Mus. Brit.
General Appearance.—This very singular shell would not, without some examination, be thought to be a cirripede. From the symmetrical position of the four apertures, with the diamond-shaped orifice in the middle, with the sutures on each side of the rostrum, and from its depressed and circular form, this shell bears some resemblance to the perforated species of Scutella. Shell much depressed, generally nearly circular, with the basal margin highly sinuous and even sometimes almost branched. Surface slightly irregular, marked by fine lines of growth, and covered by brown membrane. Of the four compartments, the Rostrum is very narrow, triangular, and comes up to the orifice almost in a point: it is rather depressed, that is, it lies rather below the level of the other compartments: the straight sutures separating it from the lateral compartments are distinct in the upper part, though always obliterated in old shells in the lower part: these sutures are generally far plainer than those separating the lateral compartment from the carina, which in most cases are obliterated and calcified together, excepting close to the orifice. The alæ of the rostrum are not externally visible, and there are no radii to any of the compartments. The Carina is twice as broad as the rostrum, and is furnished with alæ of the usual shape, which are, to a certain extent, externally visible. The Lateral Compartments are broad, being broader than the carina; they are both penetrated, down to the surface of attachment, by a hole or rather tube,—the two holes standing opposite the rostral end of the operculum: the carina is penetrated by two rather smaller but similar holes. It is these four holes which give to this cirripede its very singular aspect: they are rather smaller than the orifice of the shell; they are oval, with their longer axes placed in the direction of the ray of the circular shell: their manner of formation will be immediately explained. The orifice is neatly diamond-shaped and broad: it is rather small compared with the whole shell, and is closed by the operculum, which is seated near the summit of the shell. Basal diameter of largest specimen 2/10 of an inch; few, however, attained this size, and perfect larvæ were included in much smaller specimens.
Structure of Shell.—The rostrum (fig. 4 c) is remarkable from its small size, and from the plainness of its sutures, in comparison with those separating the other compartments, and this is exactly the reverse of what I should have expected in a compartment tending to become rudimentary. Not only is the rostrum small, but the alæ project to an unusually small degree, and gradually slope away into the lower part of the parietes. These peculiarities are even more strongly marked in very young shells: thus in one of the size of a pin’s head, the rostrum consisted of a minute parallelogram, without, as far as I could see, any alæ, and was only one fourth of the size of the carina,—this latter compartment being only half as wide as the lateral compartments. The carina, at this early period, had quite distinct alæ.
The tubular prolongations from the four external holes are of course very conspicuous on the under side of the shell (4 b); and their struc[Pg 474]ture is there plainly seen. In extremely young shells the holes are not present; but very soon, at four points of the circumference, namely, two in the carina and one in each lateral compartment, the basal edge becomes indented, and during growth more and more deeply indented; at last the horns or points of the bays, thus formed in the circumference of the shell, grow inwards and meet, the four indentations being thus converted into four rings or holes; as the shell is added to, at its circumference, these come to stand further and further from the exterior margin; and as the shell at the same time rises above the surface of attachment, the holes are added to at their basal edges, and are thus converted into shelly tubes, generally freely open at the bottom as well as at the top. Sometimes these tubes are closed at the bottom, and this is usually caused by their sides having been added to in a spiral direction. A somewhat sinuous double ridge or fissure, leading from the tubes or holes to the exterior border, can always be perceived on the under side of the shell. Occasionally, though rarely, in very old shells, a second series of holes is formed outside the first four holes, and often a tendency to this may be perceived in the just-mentioned fissures expanding a little at their outer ends, thus forming four new circumferential indentations. The purpose of this peculiar structure, apparently, is to give support to the much depressed and thin shell. In Chthamalus stellatus and scabrosus, we have seen a slight indication of a similar structure, in the formation on the under side of the shell, but confined to the lines of sutures, of obscurely tubular pillars: we have also something analogous in the singular midribs, in Platylepas, causing the membranous basis to be convex. I need only further add, that the parietes, in Chamæsipho, are rather thin, and are composed of translucent shell, punctured for the entrance of tubuli, with the punctures often arranged in straight lines.
Opercular Valves (fig. 4 d).—These are attached by a narrow opercular membrane to the sheath, but little beneath the summit of the shell. The Scuta are considerably arched or convex: the articular ridge is very prominent, and there is a thick strong adductor ridge. The Terga have a short, rather broad, rounded spur, placed very nearly in the middle of the valve: the crests for the tergal depressores are moderately developed, and are simple.
Mouth.—The crest of the labrum is hairy, and is much hollowed out. The palpi are small and narrow, with long bristles at their apices. The mandibles have four or five teeth, with the lower part pectinated. The maxillæ are notched. Of the Cirri, the second pair is short, and all the segments are thickly clothed with bristles. The third pair in the arrangement of the bristles resembles the four posterior pairs. The segments in these pairs are elongated, and support four pairs of main spines.
I was not able to observe any Branchiæ.
[133] Παχυς [Greek: Pachus] thick, and ελασμα [Greek: elasma], a valve.
CHTHAMALUS. Philippi. Enumeratio Mollusc. Siciliæ.
Compartments, when the shell is very young, eight; when mature, either six, or in appearance only four owing to the close union of the lateral compartments: basis calcareous.
Distribution, Mediterranean, and New South Wales; deep water.
The two species here included form a very natural genus, though, as far as the shell alone is concerned, at first sight there is an unusual amount of difference between them. This genus offers an instance of a case, far from uncommon in nature, though so unfortunate for the systematist, in which the most obvious and useful characters of a group are completely masked. When I first examined Pachylasma giganteum,[134] I did not doubt that it was a Balanus; and when I first looked at P. aurantiacum, I thought, from there being in appearance only four compartments, that it was an Elminius; in neither case, from the absence of alæ to the rostrum, did I even suspect that the species belonged to the sub-family of the Chthamalinæ. But when I examined the included animal’s body, I found, in both species, the labrum bullate, not notched, with the palpi small, and the mandibles with their lower teeth not laterally double. Again I found in the third pair of cirri only the basal segments thickly clothed with spines; and lastly, there were caudal appendages. Now these characters are pre-eminently those of the Chthamalinæ; in fact, they are those met with in the typical genus Octomeris, with the exception of the presence of caudal appendages, and these occur in Catophragmus,—a genus standing next to Octomeris, and in no other genus of sessile cirripedes. Moreover, if we look to the shell of Pachylasma, the absence of pores in the parietes, or at least[Pg 476] of symmetrical longitudinal ribs on their inner surfaces, and the peculiar character of the narrow radii, hardly differing in structure from the parietes, are characters which are rare in the Balaninæ, but universal in the Chthamalinæ. Hence, taking the whole organisation of the two species of Pachylasma, it is certain that they must be ranked amongst the Chthamalinæ, though the leading character of the group, namely, the rostrum being furnished, like the carina, with alæ, here fails. Owing to this conviction, I examined very young individuals of P. giganteum, and in specimens only the 1/100th of an inch in height, I was interested by finding eight separate compartments, with the rostrum having distinct alæ; hence, at this early age, as far as the shell is concerned, this species may be said to be an Octomeris; and we have seen that this likewise holds good with the included animal’s body; as the young shell increases in size the minute rostrum and rostro-lateral compartments blend together (Pl. 19, fig. 5 b), without even traces of sutures being left.
[134] Dr. Philippi called this species a Chthamalus; giving this generic name from an examination only of the separated valves in a fossil condition.
With respect to Pachylasma aurantiacum, I have no doubt that at an early age it would possess a perfectly distinct rostrum with alæ; for in the one specimen which I have seen, the compound rostrum is divided by sutures, faintly visible, both externally and internally (Pl. 20, fig. 1 a, 1 b), into three compartments (B, A, B), of which the middle one, or true rostrum, still shows, in the manner in which it underlaps the little rostro-lateral compartments, vestiges of alæ. I may remark, that we have here the same structure as in Chelonobia, formerly described, with the following differences, that here the sutures pass through the outer lamina of the parietes, so that, as seen externally, the separation of the three compartments is much more perfect than in Chelonobia; on the other hand, internally, the separation is less distinct, as the two rudimentary rostro-lateral compartments do not form part of the sheath. Pachylasma aurantiacum is further remarkable, from the two lateral compartments (C, D) on each side, tending to blend together, being only separated by sutures not more distinct, externally, than those separating the compound rostrum, but more distinct internally, for they run up the sheath. From these facts it follows, that[Pg 477] this species, viewed outside, without particular care, would be said to consist, like an Elminius or Chamæsipho, of only four compartments; if the sheath alone were looked at, there would be said to be six compartments; but when the walls, especially their basal edges, are carefully examined, either internally or externally, the eight compartments can be plainly distinguished.
With respect to the affinities of this genus, we have seen that both in the included animal’s body, and, at an early age, in the shell, it is extremely close to Octomeris: in the presence of caudal appendages, and in the basis being calcareous, we have a clear affinity with Catophragmus; in the two lateral compartments of P. aurantiacum, tending to become blended together, we have some relationship exhibited to Chamæsipho, as is likewise shown in the structure of the second and third pairs of cirri. If the genera of the Chthamalinæ were ranged in a circle, Pachylasma would be the point of contact with the Balaninæ. I must repeat, that it is extremely unfortunate that when the shell alone of P. giganteum is examined, it is hardly possible to separate this genus from Balanus.
CHTHAMALUS GIGANTEUS. Philippi (!). Enum. Mollusc. Siciliæ, 1836.
Shell and operculum dirty white: carino-lateral and lateral compartments furnished with similar alæ.
Hab.—Mediterranean; Sicily; deep water; often attached to the Millepora aspera, and sometimes associated with B. tulipiformis. Fossil in the tertiary beds, near Messina. Mus. Brit., Cuming, Stutchbury, Lyell, Philippi.
I owe to the kindness of Dr. Philippi, authentic specimens of his Chthamalus giganteus, from the tertiary beds of Messina: had this distinguished naturalist seen recent specimens, or a fossil one with all the valves united, he would no doubt have perceived that this species cannot be classed with Chthamalus. I am also indebted to Sir Charles Lyell for some magnificent specimens, which he himself collected near Messina.
General Appearance.—Shell conical, rugged, irregular, with the lines of growth plain; colour dead dirty white. Orifice large, diamond-shaped, narrow towards the carinal end; notched. Radii narrow, barely distinct from the parietes; in the same individual sometimes absent, and sometimes forming a mere ribbon, confined to the lower edge of a compartment. The diametric growth is effected by the alæ, which seen externally are broad, and strongly marked by lines of growth. Basal diameter of largest recent specimen 1.15; height of highest compartment (a carina) 1.4; and width at base of widest rostrum .85 of an inch. Amongst the fossil specimens, height of highest compartment (a carina) 1.8, width of the same one inch; width of widest rostrum 1.1 of a inch. One of these carinæ, a little below the middle point, was actually .3 of an inch in thickness, from which circumstance I have given the generic name of Pachylasma.
Scuta, triangular, but the width varies a little: growth-ridges prominent, sinuous, with a few slight furrows radiating from the apex of the valve. Internally, the articular ridge is not very prominent; nor is there a deep depression for the adductor muscle, and none for the lateral depressor muscles. The upper part of the valve projects freely.
Terga, broad, broader than the scuta: the growth-ridges are prominent, and angularly upturned close along the scutal margin; the carinal half of the valve is smooth, with the faintest traces of longitudinal striæ. A portion of the valve, nearly half of its entire width on the scutal side, is slightly depressed below the general level, and depends slightly beneath the basal margin on the carinal side: this evidently forms the spur. The carinal margin is nearly straight, with the lines of growth upturned along it. Internally, the articular ridge in the upper part is extremely prominent: the crests for the depressor muscles are very prominent, and depend beneath the basal margin like a comb: they extend over nearly half the basal margin, and the muscle, in a corresponding manner, is unusually spread out.
Structure of Shell and Radii: the compartments are attached to each other less strongly than in any other cirripede which I have examined, so that when dried specimens are soaked in spirits of wine they generally fall to pieces with a touch. In full and half-grown specimens the carino-lateral compartments are nearly as broad as the lateral compartments; in very young specimens, about 1/20th of an inch in basal diameter, they are proportionally much narrower. The walls are strong even in young specimens; in old ones they attain a thickness I have scarcely seen equalled except in Chelonobia. Their internal surfaces are smooth, as is the basal internal margin in young specimens, but in old specimens it is roughened with short, blunt ridges and little points. The Radii are often absent; when present they are very narrow, and consist merely of a ribbon-like portion, formed by obliquely upturned layers of growth, more prominent than on the parietes. The Alæ are very largely developed; they are added to, during the diametric growth of the shell, in a regular sweep all the way down to the basal margin, and consequently they do not form a rectangular shoulder as is usual; externally they are plainly marked by lines of growth: they[Pg 479] are added to a little above the line of attachment of the opercular membrane; their summits are very oblique. The sheath has its basal edge slightly hollowed out.
Basis, solid, calcareous, very irregular, and of variable thickness.
I have, under the Genus, alluded to the structure of the rostrum: in one shell, the basal diameter of which barely exceeded 1/20th of an inch; the compound rostrum (being 5/200ths of an inch in height), had its basal margin (being 9/200ths of an inch in width) rendered deeply sinuous (see Pl. 19, fig. 5 b) by two indentations, corresponding with and caused by two notches at the top of the valve. These two notches extended down barely 1/100th of an inch (strictly 4/1000ths) from the summit and then disappeared; so that when the shell was under 1/100th of an inch in height, (only one distinct zone of growth having been formed), the now compound rostrum consisted of three separate compartments; and there were eight compartments altogether. Of the above three little compartments, the middle one, or true rostrum, had large alæ, which could be most distinctly seen, extending on both sides, under the little rudimentary rostro-lateral compartments. These latter overlapped the compartments on both sides of them, as in all the Chthamalinæ.
Mouth: labrum bullate, with no central notch; nor is the inner fold of the labrum, forming the supra-œsophageal cavity, thickened, as in Balanus: minute muscles run from this inner fold straight back to the cavity formed by the outer bullate fold: the crest of the labrum is hairy, with a row of the minutest bead-like points or teeth. Palpi, small, broad, placed almost parallel to the sides of the mouth, with their apices not nearly touching each other. Mandibles, with three large nearly equal-sized single teeth; the whole inferior angle strongly pectinated. Maxillæ, small, with a broad, square notch beneath the two or three great upper spines.
Cirri: first pair short, with the rami equal in length. Second pair, with the anterior ramus having broader segments than those of the posterior ramus, and with all the segments, except the few uppermost, thickly covered with spines; the posterior ramus has rather less than half the segments thickly covered. Third pair, very slightly shorter than the sixth pair; anterior ramus with the lower segments, less than half of the whole in number, thickly covered with spines; posterior ramus with only the lowest segments, about one fifth of the entire number, thickly covered; the other segments of these two rami, and the upper segments of the posterior ramus of the second pair, closely resemble in the regular arrangement of their spines in pairs, the three posterior pairs of cirri. The pedicel of the third pair supports numerous, irregularly scattered bristles. The segments of the sixth pair bear four or five pairs of main spines, with a few intermediate spines.
Caudal Appendages.—Multiarticulate, narrow, tapering, situated on each side of the anus: each segment has two little tufts of spines on each side of its upper edge. These appendages are about one third longer than the pedicel of the sixth pair of cirri: in a specimen, in which the rami of the sixth pair had twenty-three segments, the caudal appendages had nineteen segments.
Penis short, hairy, finely-ringed, with no projecting point at its dorsal basis. Branchiæ moderately large, nearly circular, not much plicated.
Fossil Specimens.—With respect to the Sicilian specimens sent me by Dr. Philippi and Sir C. Lyell, I can see no difference whatever from the recent specimens, excepting in their greater size and thickness; it must, however, be borne in mind that I have seen only half-a-dozen recent shells. The one fossil scutum which I have seen is rather broader than is usual with the recent, but I have seen one nearly as broad. This species seems to have been extremely common when the beds at Messina were deposited, and probably it attained a larger size than it does at present. The compartments are always found separated, which is easily understood, by the facility with which, as above stated, recent specimens fall to pieces.
Shell tinged with orange; viewed externally seems formed of only four compartments, owing to the carino-lateral and lateral compartments on each side being separated only by an obscure fissure.
Hab.—New South Wales; apparently from deep water, attached to sandstone.[135]
[135] I am indebted to Mr. Bowerbank for this unique and interesting species, which I have deposited in the British Museum.
General Appearance.—Shell conical, smooth, with a tinge of orange colour; orifice large, deeply notched, sub-triangular. Viewed externally, the compartments in appearance are only four; but on close examination, the lateral compartments are seen to be divided by a very fine fissure into two nearly equal compartments. The rostrum is broad and flat, and when carefully examined, it also is seen to be divided by two fine fissures into three compartments; of these the middle one, or true rostrum, is a very little broader than the rostro-lateral compartment on each side; hence, on careful examination, the shell is found to consist of eight compartments. The carina is much compressed. The radii (in my one specimen) are not developed. The carina, and the two lateral compartments alone, have alæ; for the carino-lateral compartments are too closely joined to the lateral compartments, and the true rostrum is too closely joined to the rostro-lateral compartments, to have their alæ developed. The four alæ which are developed, are very broad, widely exposed, and marked externally by lines of growth. The basal diameter of my one, apparently old specimen, is one inch.
Structure of Shell, &c.—The inner surface (fig. 1 b) of the rather thick parietes is smooth, except close to the base, where it is roughened by a few irregular points. The sheath descends low down, and has its lower edge slightly free. The sutures are much plainer on the internal than on the external surface, and can be here plainly seen to be eight in number. That portion of the carino-lateral compartment, which helps to form the sheath, is narrow, though the whole compartment is of very nearly equal width with the lateral compartment. It is a singular fact, that no portion of the rostro-lateral compartment helps to form the sheath; for the alæ of the lateral compartments, overlap the whole upper part of the rostro-lateral compartments, and abut against the true rostrum. Hence, when the sheath alone is examined, the number of the compartments appears only six. In a section the true rostrum can be seen to underlie the rostro-lateral compartments, and thus exhibits vestiges of alæ. The fact of the rudimentary rostro-lateral compartments not forming a part of the sheath offers a marked difference from Chelonobia, which otherwise has this part of the shell very similarly constructed. The diametric growth of the shell, which seems to be considerable, is effected by the four large alæ of the carina and of the lateral compartment on each side. The sutural edges of the alæ are added to in a regularly inclined line down to the basis. The basis is calcareous, and not very thin.
Both opercular valves (fig. 1 c, 1 d) closely resemble those of the last species.
The Scuta are remarkably narrow and elongated: the external surface is slightly furrowed longitudinally, the prominent lines of growth are much wrinkled. The articular ridge is blunt, and not very prominent. The upper part of the valve is reflexed, and a considerable portion must have projected freely. The Terga are nearly twice as broad as the scuta: the carinal half of the valve is very smooth, and is coloured beautiful reddish-orange. The spur is tolerably distinct. The articular ridge is not very prominent.
The Mouth resembles that of the last species. The mandibles have three sharp teeth, with the inferior part narrow and pectinated with long spines: the edge, also, between the upper main teeth, is pectinated with short spines, which latter often have their summits crenated. The maxillæ are notched; the inferior corner is produced into a small step-formed projection. The outer maxillæ are more pointed than is usual.
The Cirri most closely resemble in every detail those of P. giganteum; I can point out only one slight difference, namely, that the lower segments, in the posterior ramus of the third cirrus, which are thickly clothed with spines, are more numerous in proportion to the upper segments with the spines arranged in regular pairs (being as 4 to 14), than in P. giganteum, in which only one fifth (or 4 to 20) are so clothed.
Caudal Appendages.—These are very small, not being more than one fifth of the length of the pedicel of the sixth cirrus: their segments are indistinct, and they support a very few coarse spines. The rami of the sixth cirrus, in the one specimen, had twenty-five segments, whilst each caudal appendage had only five. Hence the caudal appendages are far less developed than in P. giganteum.
OCTOMERIS. G. B. Sowerby. Zoological Journal, vol. 2, p. 244, July, 1825.
Compartments eight: radii with their edges crenated: basis membranous.
Distribution, Cape of Good Hope; Philippine Archipelago.
The two species of this genus differ considerably in external appearance, though not in essential character. Both, as the name expresses, have eight compartments: the carino-lateral pair are rather narrower than the lateral. The basis is membranous. The radii are narrow, and are distinctly crenated on both sides of the sutures with the teeth neatly interlocking; but these teeth can hardly be distinguished in the large, corroded specimens of O. angulosa. The crenated structure of the radii is identical with that described under Chthamalus dentatus and Hembeli; if, indeed, we were to add carino-lateral compartments to the shells of these two species, they would belong to Octomeris. I have seen only a few specimens of either species of Octomeris, and none preserved in spirits; and therefore I know nothing of the anatomy of the softer parts: I was not able to make out distinctly any branchiæ. The cirri differ considerably in the two species, in nearly the same way as in the two species of Chamæsipho. In the structure of the second pair of cirri, and in the tendency of the basal margin of the parietes to form bay-like indentations, Octomeris angulosa shows some special affinity to Chamæsipho scutelliformis. Under Pachylasma, I stated that that genus was closely related to Octomeris; and I have just alluded to the close affinity of the latter to the division of the genus Chthamalus, which has crenated radii.
OCTOMERIS ANGULOSA. G. B. Sowerby. Zoological Journal, vol. 2, July, 1825. And Genera of Recent and Fossil Shells, Plate.
-------- STUTCHBURII. J. E. Gray. Annals of Philosophy, new series, vol. 10, August, 1825.
-------- AUGUBRA (?) Chenu. Illust. Conch., Tab. 4, fig. 2.
Shell dirty white, rugged and massive: alæ thick, with their sutural edges coarsely crenated.
Hab.—Algoa Bay, Cape of Good Hope. Attached to littoral rocks; often associated with Balanus Capensis and Chthamalus dentatus; Mus. Brit., Cuming, Stutchbury, Bowerbank.
General Appearance and Structure of Shell.—Shell extremely rugged, irregular, massive, generally much corroded, steeply conical or even sub-cylindrical: orifice large, broad, rhomboidal, of nearly equal breadth at both ends. Colour dirty white, often slightly tinted yellow from the investing membrane, and from thin layers of punctured membrane alternating with the laminæ of shell. The parietes, in old specimens, have very irregular longitudinal ridges, or rather plates projecting out, sometimes much branched, and generally curved inwards so as to meet each other, thus forming round the basal margin a circle of cylindrical apertures. In old large specimens the radii are not developed, and till the compartments are disarticulated there is no trace of the toothed structure of their sutural edges: in this condition the sutures exist as deep, rugged, narrow fissures. In younger shells, the radii, though narrow, are distinct, and have their surfaces transversely ribbed, and their edges toothed and interlocked with the teeth of the recipient furrow. Some of the specimens present a curiously deceptive resemblance to Elminius plicatus. Basal diameter of largest specimen, one inch and a quarter; height one inch.
The parietes are remarkably thick, but the compartments separate easily: I have, however, seen one instance in which they were partially calcified together. Their internal surfaces are very smooth. The sheath does not descend low. The alæ project rectangularly; they have thick edges, and these are coarsely crenated in transverse lines. Of the radii sufficient has been said.
Scuta (fig. 2 b): these, in all the specimens seen by me but one, have been deeply corroded, and their outline, as in Chthamalus, considerably modified: in the one specimen well preserved, the exterior growth-ridges were extremely prominent. The articular ridge does not project much, nor is the articular furrow very deep. There are more or less distinct crests for the lateral depressor muscles. The Terga are rather[Pg 484] narrow, with a small rounded spur, moderately distinct. The tips of the crests for the depressor muscle barely depend beneath the basal margin.
Mouth: labrum considerably bullate, with the crest hairy and furnished with a few most minute teeth. Palpi small, with their tips not nearly touching each other. Mandibles with four teeth, of which the lower ones are laterally double: inferior pectinated portion small. The maxillæ are notched, but their outline differed on opposite sides of the individual examined. Outer maxillæ bilobed on their inner face.
Cirri.—The first pair has the rami unequal in length by about three segments, having six segments on one and nine on the other ramus. In the second pair the anterior ramus is remarkably short and extremely broad; the five segments of which it is composed being thickly covered with bristles, some of which are very coarsely and doubly pectinated: the posterior ramus is nearly twice as long as the anterior ramus, and has eleven segments; of these, excepting the two basal segments which are rather thickly covered with spines, the others resemble in the arrangement of their spines the four posterior pairs of cirri, with the exception that there is a single transverse row of rather long spines along the upper and inner lateral edge of each segment. The segments in all the four posterior pairs of cirri resemble each other in bearing each four pairs of main spines (of which the two lower pairs are short), with a tuft of small intermediate spines.
The penis is remarkable from its very small size and shortness, not being more than once and a half as long as the pedicel of the sixth cirrus.
Shell reddish-brown, depressed, thin, finely furrowed longitudinally: tergum with the basal margin having a slight angular bend.
Hab.—Philippine Archipelago; rare; Mus. Cuming.
General Appearance and Structure.—Shell circular, much depressed; coloured brown, with an orange-red tint; surface regularly and narrowly furrowed in longitudinal lines, the intermediate rounded ridges projecting at the basal margin in finger-like points. The eight compartments, when disarticulated, are of nearly equal sizes; the carino-lateral and rostro-lateral compartments being rather smaller than the others. The radii are neatly toothed; but with the exception of these teeth, which are equally developed on both sides of the sutures, and which closely interlock in the lower part of the shell, but stand a little apart in the upper part, the radii can hardly be said to be developed. The alæ do not form a rectangular shoulder, the lower margin being regularly curved into the parietes; the upper margin is only slightly[Pg 485] oblique; the external surface is furrowed by lines of growth. The shell increases a little in diameter, chiefly by the growth of the alæ. The parietes are not very thick, with the inner surface smooth, but with punctures placed in rows for the entrance of the tubuli: the laminæ of shell alternate with layers of yellow finely punctured membrane. Basis formed of thin membrane, in concentric slips. Basal diameter of largest specimen, .6 of an inch.
Scuta broad, with the lines of growth few in number, but extremely prominent, so as to form folds: basal margin with a very slight angular bend: articular ridge not very prominent. Terga, with a narrow slip along the scutal margin, having the lines or folds of growth so prominent as almost to form a series of small transverse pits: the basal margin of this portion forms an angle with the rest of the basal margin: no spur can be said to exist: the crests for the depressor muscle depend a little beneath the basal margin: articular ridge very prominent.
Mouth: labrum and palpi as in the last species. Mandibles with three main teeth, apparently single, of which the lower tooth has its upper edge pectinated: the inferior part of the mandible is pectinated as usual. The maxillæ have two notches, one beneath the two upper large spines, and the other in the middle, separating some thicker and thinner spines.
Cirri: first and second pairs short, with the anterior rami in each longer by about two segments than the posterior rami; all the segments thickly covered with bristles. The third cirrus is much longer than the second cirrus, with the posterior ramus longer than the anterior ramus; on the latter the three basal segments, and on the posterior ramus the two basal segments are thickly covered with bristles; the other segments have bristles arranged as on the three posterior pairs of cirri, namely, each segment has four pairs of main spines, of which the two lower pairs are short.
The unarticulated support, whence the articulated portion of the penis arises, is unusually long, equalling the pedicel of the sixth cirrus.
CATOPHRAGMUS. G. B. Sowerby. Genera of Recent and Fossil Shells. Plate.
Interior compartments eight, with several exterior whorls of small supplemental compartments: basis either membranous or calcareous.
Distribution, West Indies and Australia. Attached to littoral shells and rocks.
This genus is very remarkable amongst sessile cirripedes, from the eight normal compartments of the shell being[Pg 486] surrounded by several whorls of supplemental compartments or scales: these are arranged symmetrically, and decrease in size but increase in number towards the circumference and basal margin. A well preserved specimen has a very elegant appearance, like certain compound flowers, which when half open are surrounded by imbricated and graduated scales. The Chthamalinæ, in the structure of the mouth and cirri, and to a certain extent in that of the shell, fill up the interval between the Balaninæ and Lepadidæ; and Catophragmus forms, in a very remarkable manner, the transitional link, for it is impossible not to be struck with the resemblance of its shell with the capitulum of Pollicipes. In Pollicipes, at least in certain species, the scuta and terga are articulated together—the carina, rostrum, and three pairs of latera, making altogether eight inner valves, are considerably larger than those in the outer whorls—the arrangement of the latter, their manner of growth and union,—all are as in Catophragmus. If we, in imagination, unite some of the characters found in the different species of Pollicipes, and then make the peduncle so short (and it sometimes is very short in P. mitella) that the valves of the capitulum should touch the surface of attachment, it would be impossible to point out a single external character by which the two genera in these two distinct families could be distinguished: but the more important differences in the arrangement and nature of the muscles which are attached either to the opercular valves or surround the inside of the peduncle, would yet remain.
Although all the valves of the shell, even the eight in the innermost whorl, are very thin, yet from their number in the successive whorls, and from each being concave inwards, so as to form a cavity or tube into which the corium enters, the total thickness of the sides of the shell is very considerable. Both of the species of Catophragmus occurred mingled, in the one case with Tetraclita porosa and in the other with T. purpurascens; now the walls of these shells, we know, are very thick, and are permeated by several rows of pores, occupied by threads of corium; seeing this, we may be permitted to believe, that the several exterior whorls of valves in Catophragmus, between which[Pg 487] the corium is prolonged for some way upwards, are of service to the animal, by thickening its shell, in an analogous, but not homologous, manner, as in Tetraclita.
Considering the whole structure, external and internal, of Catophragmus, with the one great exception of the exterior whorls of valves, there is hardly a single generic character by which it can be separated from Octomeris and Pachylasma; indeed, I am not quite sure that it would not have been better to have run these three genera together.
Of the two species, I will first describe C. polymerus, and not the C. imbricatus of Sowerby, inasmuch as I have plenty of excellent specimens of the former, whereas the original specimens of C. imbricatus, in the British Museum, consist of one old and not perfect shell, without the opercular valves or the included animal’s body; and the other, though quite perfect, far from mature. As far as these materials allow of minute comparison, the whole shell, with the exception of the basis, and the opercular valves agree very closely in the two species, whereas the included animal’s body differs more than is usual in nearly related species;—thus, C. imbricatus has caudal appendages, of which there is no trace in C. polymerus, and I have seen only one other instance in which this organ was absent in one species (Scalpellum villosum) and present in the other species of the same genus. Under these circumstances it will be most convenient first to describe in detail C. polymerus, and then only indicate the points of difference in C. imbricatus.
Basis membranous: caudal appendages none.
Hab.—New South Wales (Twofold Bay), Mus. Darwin; Swan River (?), Mus. Cuming. Attached to littoral rocks and shells, and associated with Tetraclita purpurascens, Balanus nigrescens, Chthamalus antennatus, Chamæsipho columna.
General Appearance.—Shell nearly circular, moderately or slightly depressed; colour gray. The eight normal compartments of the inner whorl are two or three times as large as those in the second whorl; the other smaller compartments or scales graduate very regu[Pg 488]larly in size, to mere beads, at the extreme basal edge, and are arranged symmetrically. The general aspect of the shell depends chiefly on the degree to which the surface has been disintegrated, and differs greatly in the two extreme states. When well preserved, the general appearance is very elegant; the scales are all neatly imbricated; they terminate upwards in points, but with their extreme tips generally broken; from laterally overlapping each other, their external surfaces (as seen, when corroded, in fig. 4 d, or when slightly exposed after the continued growth of their basal margins, in fig. 4 a) become longitudinally keeled, often with a secondary ridge or shoulder on one side; they are, also, crossed by rather conspicuous and regular lines of growth, or more strictly, former lines of union between the several valves: the orifice of the shell in this perfect condition is sub-rhomboidal and notched, and the scuta are united to the terga by nearly straight sutures. On the other hand, when the shell has been considerably corroded, and this seems to be the more common condition, the appearance is not elegant: the scales in the successive whorls are not imbricated, but owing to their upper parts having been worn down, they present a tesselated surface (4 d), with the tesseræ graduated in size, and of a peculiar shape, namely, a rectangle, with a more or less broad square projection on the exterior side, together often with a large square notch on one or both corners, caused by their laterally overlapping each other. Some of the shells are so deeply corroded, that no portion of the original surface is preserved, excepting the lowermost bead-like scales; and the whole shell has so rugged an aspect, that the successive whorls of the worn-down valves might easily be overlooked. In the corroded specimens, the orifice approaches to circular in outline, and is large and nearly entire: the scuta and terga are deeply interlocked together. The largest specimen which I have seen was one inch and a quarter in basal diameter.
Structure of the Shell.—The eight inner normal compartments are not thicker than the outer valves, and are far thinner than in ordinary sessile cirripedes. They are arranged as in Octomeris. The lateral compartments are broader than the rostro-lateral and carino-lateral compartments. The shelly laminæ, of which these compartments and the opercular valves are composed, alternate with yellow membranous layers, exactly as is the case with Octomeris. The lateral edges of the compartments, beneath the alæ, in the six compartments having alæ, bend inwards, especially just above the basis, so that the compartments in their lower parts (fig. 4 c) stand much more separate than is usual. The basal edge of each is irregularly toothed. The sheath presents no particular character. The eight compartments have longitudinal shoulders and ribs on their external surfaces, caused by the lateral overlapping and pressure of the exterior scales. They are also marked by slight, transverse or oblique calcareous ridges, caused by the attachment of the membrane, by which they are united to the smaller compartments or scales outside them. A new line of attachment, and consequently a new ridge is formed, lower and lower down at each period of growth, as the shell is added to at the basis,—in the same manner as new ridges are added to the lower edge of the sheath at each period of growth. The[Pg 489] shell, excepting in old worn-down specimens, increases largely by diametric growth: during the diametric growth, the outer scales must be laterally separated a little from each other, and probably they are laterally added to; but there are no distinct lines of suture, or rows of smaller scales, corresponding with the sutures between the eight inner compartments. The alæ do not project much; their edges, as well as the shoulders into which they fit, are generally irregularly crenated: they are added to during diametric growth above the line of attachment of the opercular membrane. There is no appearance of radii; but as the eight inner compartments are added to laterally, and are often crenated, on the edges which correspond with the radii in other Cirripedes, such edges must be considered as radii.
With respect to the scales in the several outer whorls, they resemble each other except in size, and the outermost scales are reduced to mere transversely elongated beads. Their basal edges are concave inwards, being bent like the ridge of a house; hence sub-triangular spaces or tubes, lined by the corium, run up between the scales. When perfectly preserved, the outline of each scale is a much elongated triangle, but usually, from their summits having been worn off, the outline is nearly that of a parallelogram. Their basal edges are dentated, and their upper parts, both on the outside and inside, are marked, where joined to the other compartments, by slight calcareous ridges: outside, there are longitudinal shoulders (Pl. 20, fig. 4 b), caused by the lateral overlapping of the adjoining scales; these are best seen in section in corroded specimens (fig. 4 d): on the inside there are, also, in the upper part, slight medial longitudinal ridges, caused by the sutures, which the scales have covered.
In large old specimens there are ten, or even more, whorls of compartments, but it is scarcely possible to count them with any accuracy. The first whorl consists of the eight large inner compartments, though, homologically, it is doubtful whether the rostro-lateral compartments in any sessile cirripede really belong to the same whorl with the others. The second whorl consists of eight smaller pieces, covering the eight sutures in the first whorl (see the tracing of the basal edges of all the compartments and valves in a very perfect specimen, Pl. 20, fig. 4 c). The third whorl, in large and perfect specimens, consists of twice the number, or sixteen, still smaller scales, corresponding with the sixteen sutures of the second whorl; but sometimes there are less than sixteen pieces, owing to some of the scales being large enough to cover two adjoining sutures as well as the intermediate portion of the compartments of the first whorl. In the fourth whorl, instead of there being, even in the most perfect specimens, twice sixteen, or thirty-two pieces, there are only twenty-four; this being caused by single pieces (placed alternately with two pieces) being broad enough to cover two sutures as well as the intermediate portion of the compartment of the third whorl. In the succeeding whorls this same arrangement seems to be the usual one, so that in the fifth whorl, instead of there being, in the most perfect specimens, twice twenty-four, or forty-eight pieces, that is, twice the number in the last whorl, there are only thirty-six scales, or once and a half as many scales.
The Basis is thin and membranous; it firmly adheres to the surface of attachment.
Scuta, nearly flat: the articular ridge is very prominent, and there is a deep articular furrow both above and below; but the precise outline of the ridge and furrows varies: there is a pit for the adductor muscle, but no crests or marks for the other muscles. The Terga are remarkable from the extreme prominence of the articular ridge and depth of the articular furrow. The basal margin viewed internally seems straight, so that there appears to be no spur; but viewed externally, when the crests for the depressor muscles are seen to depend considerably beneath the true basal margin, a slight, very broad spur may be perceived to exist. These depending crests for the muscles are rather thin, but they extend over half the basal margin of the valve.
Mouth.—The labrum is very bullate, being as long in its longitudinal axis as the rest of the mouth: crest hairy, with some very minute teeth. Palpi truncated, with their apices not nearly touching each other; thickly clothed with spines. Mandibles with three large single teeth, of which the lower one has a single fine tooth at its upper basal edge, showing a tendency to become pectinated; inferior part short, coarsely pectinated. Maxillæ notched, with a slight second notch and slight double prominence in the lower part.
Cirri.—First and second pairs short, with the rami in each unequal in length by about four segments; on both rami in the second cirrus, and in the shorter ramus of the first cirrus, there are some coarsely pectinated spines. All four posterior cirri are alike; the segments bear five pairs of strong spines, with a large intermediate tuft of fine spines: the dorsal tuft is also large, consisting of short thick, and long finer spines. There is no vestige of caudal appendages, though present in the succeeding species.
Branchiæ moderately large, in area equalling the prosoma; surface not plicated.
CATOPHRAGMUS IMBRICATUS. G. B. Sowerby. Genera of Recent and Fossil Shells, Plate.
Basis calcareous: caudal appendages present.
Hab.—Antigua, West Indies, attached to a Tetraclita porosa.
As stated under the genus, this species is known from two specimens in the British Museum, one of which is full-sized, being three quarters of an inch in diameter, but is destitute of the outermost whorls, of the basis, opercular valves, and animal’s body; the other is perfect, but very young, being barely two tenths of an inch in basal diameter. As far as the characters can be made out from these materials, I can perceive[Pg 491] no difference from C. polymerus in the shell, excepting that in the small specimen of C. imbricatus, there would appear to exist fewer whorls. The opercular valves are likewise closely similar: in the scuta, however, of the young specimen in the present species, the articular ridge seems to be a little broader, but this is so variable a character that no confidence can be placed in it: these valves, moreover, externally have a broad furrow along the middle, running from the apex to the basal margin, which is not the case with the scutum of C. polymerus; but then I have often seen, in young specimens of Balanus, a similar furrow, which is quite absent in full-grown specimens. When we come to the basis we find a good diagnostic character, for here it is calcareous: it is rather thin, solid, and white; towards the outside it is pitted with small cavities, corresponding with the small teeth on the basal edges of the compartments. The latter adhere firmly to the basis. The central internal surface is covered by an irregular network of imbedded cement-ducts, some of which bifurcate. From the description here given, it will be seen that any figure would have been superfluous, the last species having been so well illustrated.
Mouth.—I can point out no difference, excepting that the palpi are here more oval or less truncated at their ends; and that the lower corner of the maxillæ seems to be more prominent. In the Cirri, the rami of the first and second pairs are nearly equal in length: none of the spines are coarsely pectinated. In the four posterior pairs of cirri, instead of a tuft of small spines on each segment between the pairs of main spines, there are only a few minute intermediate spines: the dorsal tufts are also here smaller, but are, as in the last species, composed of short thick, and longer thinner spines.
Caudal Appendages.—This is the only species of sessile cirripede, with the exception of the two species of Pachylasma, which possesses these organs: they are situated on each side of the anus in the usual position: they are minute, equalling in length only the lower segment of the pedicel of the sixth cirrus: in a specimen in which the rami of the sixth cirrus had eighteen or twenty segments, these appendages consisted of only three tapering segments, supporting a few thick spines.
Branchiæ.—I believe I discovered these, consisting of two minute pouches, placed at the carinal end of the sack: if this observation be correct, this species differs from C. polymerus in the much smaller size of these organs.
The following species of fossil Balanidæ are given in that most useful work, the ‘Enumerator Palæontologicus’ in Bronn’s ‘Gesichte der Natur:’ it has appeared to me that a few words on each species, might hereafter save others the trouble of searching through several works.
Tubicinella maxima of Morren, said to have been found in the Chalk: this would have been a wonderful fact, considering that no true sessile cirripede has hitherto been found in this formation, and that it implies the existence of Cetacea at this period; but I have been informed that the fossil in question is not a Cirripede.
Diadema bifidum = Coronula bifida of Bronn, in his ‘Italiens Tertiär-Gebilde’ (1831), p. 126 (no Plate). Without a much fuller description I can form no judgment on this species.
Diadema vulgare is probably the Coronula barbara, a Crag fossil described by me.
Pyrgoma undata, Michelotti, in ‘Bull. Soc. Geolog.’ tom. x, p. 141, a mere name without any description: probably it is a synonym of
Pyrgoma sulcatum, Philippi, ‘Enum. Mollusc. Siciliæ,’ which is a synonym of Pyrgoma Anglicum of the present work; found recent and fossil.
Acasta Montagui is probably the extinct Acasta undulata described by me.
Chthamalus giganteus of Philippi, is the Pachylasma giganteum of this work; found recent and fossil.
Chthamalus stellatus, said by Philippi to be found fossil in Sicily; such may be the case, but the littoral habits of the species do not render it very probable.
Balanus carbonarius of Petzholdt, found in the Carboniferous formation! but I have given my reasons, in my ‘Monograph on the Fossil Lepadidæ,’ p. 5, for disbelieving that this is a Balanus, or even a Cirripede.
Balanus ostrearum appears to be a mere name by Conrad, published by Morton in his ‘Synopsis of the Organic Remains of the Cretaceous Group,’ 1834, Appendix, p. 8.
Balanus peregrinus is briefly described and poorly figured, without the opercular valves, by Morton, in his ‘Synopsis,’ ut supra, p. 72, Pl. 10, fig. 5: this Eocene species apparently resembles the Eocene B. unguiformis of Sowerby, described in this work; but quite indispensable details of structure for identification are not given. Another figure of this shell is given in the ‘American Journal of Science’ (N. S.), vol. i, Pl. 2, fig. 6.
Balanus circinnatus, communis, and pustula, of Defrance, as well as all the other species named by him, are described so imperfectly, that the descriptions are of no value whatever, every description being applicable to every species: I must add, that the B. communis is not the B. communis of British authors, a name applied to several forms.
Balanus Finchii is briefly described and figured, but without the opercular valves, by Isaac Lea, in his ‘Contributions to Geology,’ 1833, p. 211, Pl. 6, fig. 222. I do not think that I have seen this species.
Balanus Holgeri, Geinitz, ‘Grundriss der Versteinerungen,’ tab. ix, fig. 19. No opercular valves are given; this species cannot be even approximately recognised.
Balanus proteus, Conrad, ‘Fossil Shells of Miocene Formation of U. States,’[Pg 493] p. 77, Pl. 44 (in ‘Journal Acad. Nat. Sc.,’ Phil., vol. vii, p. 134). I cannot recognise this species; it resembles B. porcatus; but as the radii are rather narrow, and apparently with slightly oblique summits, it may be B. concavus; the opercular valves are not figured.
Balanus sagittata is merely a provisional name without any description, given in a paper on the Crag by S. Woodward, in the ‘London and Edin. Philosoph. Magazine, Brewster, Taylor, and Phillips,’ vol. vii, July-December, 1835, p. 354.
Balanus sublævis, J. de C. Sowerby, in ‘Geolog. Trans.,’ 2d series, vol. v, Pl. 25, fig. 3. Plate extremely imperfect; description extremely short and useless; a species from India not to be recognised.
Balanus balanoides; the species thus named by Ranzani, and found by Philippi in Sicily, certainly is not the true Lepas balanoides of Linn., but may be B. amphitrite of this work. In British collections of Crag specimens I have found the B. dolosus (nov. spec.) thus named: I much doubt whether the truly littoral B. balanoides of Linnæus has been found fossil.
B. costatus of Montagu, a synonym of B. sulcatus of Bruguière, and of B. porcatus of this work; found fossil and recent. This species was originally described by Linnæus under the name of Lepas balanus. But the fossil Lepas balanus of Brocchi is a different species; if it be the same with the recent Lepas balanus of Poli, then it is the B. perforatus of the present work. Again, the B. sulcatus of Bronn, in his ‘Lethæa Geognostica’ (tab. 36, fig. 14), is quite different from the B. sulcatus of Bruguière (i. e., B. porcatus of the present work), as is at once obvious from the oblique summits of the radii. Lastly, B. tesselatus of Sowerby is a synonym of B. porcatus, sulcatus, costatus, and Lepas balanus.
B. punctatus of Montagu is a synonym of Chthamalus stellatus, see remarks on that species: the name of B. punctatus is often applied by British authors to varieties of B. balanoides, see remarks on that species.
B. rugosus of Morris’s ‘Catalogue’ is a synonym of B. crenatus; found recent and fossil.
B. sulcatus of Bruguière, a synonym of B. porcatus; found recent and fossil: see remarks under B. costatus.
B. tintinnabulum of Linn., found recent and fossil; but no trust whatever ought to be placed in the identifications of this species given in several works; thus the Lepas tintinnabulum of Brocchi is distinct. The B. crassus of Sowerby is the true B. tintinnabulum. The B. fasciatus of Dujardin (perhaps only a MS. name) probably is also this species. The B. crispatus is only a variety of B. tintinnabulum.
B. dentiformis, Defrance: see B. circinnatus.
B. ornatus, Münster, ‘Beiträge zur Petrifact.,’ B. 3, p. 29, tab. vi (1840). No opercular valves or details of structure of the several species of Balanus named by Münster, are given, and consequently none can be recognised with certainty.
B. concavus of Bronn, fully described in this work, with the synonyms given; recent and fossil.
B. crassus of Sowerby, a synonym for B. tintinnabulum.
B. latiradiatus, Münster, probably a synonym of B. tintinnabulum, see remarks under B. ornatus.
B. pectinarius, Bronn (‘Italiens Tertiär-Gebilde,’ p. 128), does not appear to me fully enough described to be recognised.
B. pictus of Münster, possibly a synonym of B. amphitrite: see remarks under B. ornatus and balanoides.
B. plicarius, Bronn: see remarks under B. pectinarius.
B. porosus of Hausman (according to Münster of Blumenbach). I cannot recognise this species in Münster’s ‘Beiträge.’
B. pustularis of Lamarck, ‘Animaux sans Vertèbres.’ Scarcely one of the[Pg 494] fossil species of Balanus, described by Lamarck, can be recognised; the descriptions are extremely imperfect. There is a figure of B. pustularis in Münster’s ‘Beiträge,’ which makes me think that this may be a synonym of B. concavus of Bronn.
B. pyramidalis, Münster: see remarks under B. ornatus.
B. rhomboicus, Bronn: see remarks under B. plicarius.
B. squamosus, Defrance: see remarks under B. circinnatus.
B. stellaris, Bronn (‘Lethæa Geognostica,’ tab. 36, fig. 13). To this species, Lepas stellaris (I presume a misprint for stellata) of Poli, is given as a synonym; but the Lepas stellata of Poli is a Chthamalus, and this certainly is not the case with B. stellaris. I have received two specimens from the Continent named B. stellaris, but they certainly differed from the form so called by Bronn, for in that, the parietes are said to be porose and the radii very narrow: these foreign specimens I have named B. inclusus. A species described by me as B. corrugatus, resembles in external appearance the B. stellaris of Bronn, but it is mere labour in vain to attempt identifying Balani by their external characters.
B. striatus, Defrance: see remarks under B. circinnatus.
B. tertiarius, Risso, ‘Hist. Nat. de l’Europe Merid.,’ vol. iv. I cannot recognise this species.
B. tesselatus, Sowerby, a synonym of B. porcatus; recent and fossil: see remarks under B. costatus.
B. zonarius, Münster: see remarks under B. ornatus; possibly this is a synonym of B. concavus.
B. amphimorphus, Lamarck: see remarks under B. pustularis.
B. crispatus, var. of B. tintinnabulum.
B. cylindraceus, Lamarck. From Chenu’s ‘Illust. Conch.,’ in which work Lamarck’s original specimens are figured; it appears that this is the B. psittacus of South America, where it is also found fossil, but assuredly Lamarck is quite in error when he states that a variety of this species occurs fossil near Turin. Bronn (in his ‘Italiens Tertiär-Gebilde,’ p. 127) gives as a synonym to the Turin fossil the Lepas tintinnabulum of Brocchi (in the ‘Conchologia Fossile Subapennina,’ t. 2, p. 597), and this probably is correct; and I have hardly any doubt that the Lepas tintinnabulum of Brocchi is the B. concavus of Bronn described in the present work.
B. perforatus of Bruguière is said by Philippi to be found in Sicily: see remarks under B. tulipa.
B. semiplicatus, Lamarck: see remarks under B. pustularis.
B. tulipa of Müller is the B. Hameri of this work, under which name full information on its geological history has been given. The B. tulipa of Poli and Ranzani is the B. tulipiformis of the present work; and this latter species is said by Philippi to occur fossil in Sicily. But there has been so much confusion in the identification of B. tulipiformis, B. Hameri, and B. tintinnabulum, and likewise of B. perforatus (which by Poli was wrongly considered to be L. balanus, Linn.), that doubts must be entertained about which have been really found, until the Sicilian fossils are all carefully examined.
B. ovularis, Lamarck: see remarks under B. pustularis.
B. Uddevallensis, Linnæus, is probably a synonym of the B. Hameri of this work and the B. tulipa of Müller.
[A few other references may be added to those given by Bronn.]
B. miser, stated by Lamarck to be found fossil: see remarks under B. pustularis.
B. patellaris of Lamarck, is stated by Marcel de Serres to be found fossil with several other species of Balanus; thus named without any description in the ‘Annales des Scien. Phys. et Nat. de Lyon,’ tom. i, p. 417.
B. virgatus, delphinus, and crispus, are names given by Defrance, with absolutely worthless descriptions.
B. radiatus is too briefly described by Risso (‘Hist. Nat. de l’Europe Merid.,’ tom. iv, 1826), without a figure, to be recognised; it probably is not the B. radiatus of Spengler, Wood, and other authors.
B. goissopomo, lævis, and radiatus, are mere names without any description, published in a Catalogue by F. Hoeninghaus, in the ‘Jahrbuch für Mineral. Geog.,’ &c., 1831, p. 155.
B. humilis. Conrad, in the ‘American Journal of Science,’ vol. ii (N. S.), p. 400, 1846, has given a short description, with a woodcut, of this species, from the upper Eocene of Florida. The opercular valves are not described, and I doubt whether the species could be recognised.
Cirripedia without a peduncle: scuta and terga, not furnished with depressor muscles, moveable only on one side, on the other side united immoveably with the rostrum and carina into an asymmetrical shell.
The one genus herein contained differs so considerably from all the others in the Order, in the extraordinary unequal development of the two sides of the shell, that I have instituted a Family for its reception. If compelled to place it in one of the foregoing families, I should with much hesitation rank it in the sub-family Chthamalinæ, rather than amongst the Lepadidæ; for it is destitute of a peduncle, and has a shell, though a very different one from that of any true sessile cirripede. In the interfolding sutures which may be considered as representing radii or alæ, in the basis being divided into concentric slips, and in the whole of the basis being attached to the supporting object, this same line of affinity is clearly manifested. On the other hand, in the general shape, manner of growth, and kind of articulation of the scutum and tergum, there is so close an approach to the Lepadidæ, that had I seen these very important valves separately, I should certainly have concluded that they had come from a Pollicipes, allied to certain Cretacean fossil species, as P. fallax and elegans; it likewise, perhaps, deserves notice, that the upward growth of the rostrum, in Verruca nexa, is a peculiarity[Pg 496] found only in the valves of the Lepadidæ. Verruca differs both from the Lepadidæ and Balanidæ in the whole shell or external covering, having no other muscle besides the adductor scutorum. In the characters derived from the animal’s body, Verruca approaches both families; but in the absence of branchiæ, and in the great development of the caudal appendages, perhaps it comes rather the nearest to the Lepadidæ. Whatever affinity there is to the Balanidæ, it is much stronger to the sub-family Chthamalinæ than to the Balaninæ; though the non-bullate labrum, in three of the species, and the great dissimilarity of the third cirrus from the three posterior pairs, at first seems to indicate a closer relationship to the Balaninæ; but the labrum is never notched, as in the latter sub-family, and in V. nexa it is bullate, and supports palpi of only small size. The dissimilarity, also, of the third pair of cirri, compared with the posterior pairs, is hardly greater than in Chthamalus intertextus and Chamæsipho columna, members of the Chthamalinæ, though abnormal in this one respect. Perhaps even a special affinity is evinced between certain species of Chthamalus, as C. intertextus, and certain species of Verruca, as V. nexa, namely, in the interfolding sutures and in the very peculiar, inflected basal margin of the walls. Upon the whole, the affinities of the Verrucidæ are complex, and nearly equally divided between the two great families of Balanidæ and Lepadidæ, or sessile and pedunculated cirripedes.
VERRUCA.[136] Schumacher. Essai d’un Nouveau Syst. Class., 1817.
CLYSIA. Leach. Journal de Physique, tom. 85, July, 1817; Clisia, Leach, Encyclop. Brit. Suppl., vol. 3, 1824; Clitia, G. B. Sowerby, Genera of Recent and Fossil Shells.
CREUSIA. Lamarck. Animaux sans Vertèbres, 1818.
OCHTHOSIA. Ranzani. Memoire di Storia Nat., 1820.
LEPAS ET BALANUS AUCTORUM.
[136] According to Bock, in the ‘Naturforscher’ of 1778, this term was used by Rumph for a Chelonobia, but as it was before the adoption of the binomial nomenclature, according to the Rules, it may be passed over, and does not interfere with the priority of Schumacher.
Distribution, Northern Europe, Mediterranean, Red Sea, Madeira, West Indies, Tierra del Fuego, Chile, Peru.
The shell in this genus is extremely unsymmetrical, not two of the six pieces of which it is composed quite resembling each other. At first it appeared hopelessly difficult to identify, in a homological sense, these six valves, with those of ordinary cirripedes, but the difficulty soon quite vanished. The operculum consists of two moveable valves on one side, namely, a scutum and tergum, but without any moveable valves on the opposed side: the scutum, though remarkable from being much smaller than the tergum, can be easily recognised by giving attachment to the animal’s body and to the adductor scutorum muscle. The four other pieces are articulated together, and form the shell surrounding the sack, in which the animal’s body is enclosed: of these, the two against which the moveable scutum and tergum shut, are smaller, differ greatly in shape, and are articulated together in a different manner from the remaining two pieces; from these facts alone there would be a strong presumption that they were of a different nature. The fixed valve, against which the scutum shuts, is either furnished with a remarkably prominent plate (a in fig. 1 c, s′; compare this with s′ in the reversed shell in fig. 1 e), or is hollowed out, as in V. nexa, for the attachment of the adductor scutorum muscle. Thus it is rendered probable that this fixed valve is a modified scutum; but a surface of attachment for one end of the adductor muscle might, perhaps, have been developed on any other valve, or a scutum might have become fused with a lateral valve of the shell; the shell on this latter view being rendered in idea more symmetrical. But when a very young specimen is carefully examined, it is found that the moveable and fixed scutum, the moveable tergum and its opposed valve or fixed tergum, at the first period of calcification, resemble each other quite closely; but that, as each zone of shell is added, the differences become rapidly greater and greater: hence,[Pg 498] it may be considered as directly proved, that the two fixed valves (S′ and T′ in all the figures in Pl. 21), which are opposed to the moveable valves of the operculum (S and T), consist of an extraordinarily modified scutum and tergum. It has been shown (p. 129), that at the period of the metamorphosis, the two scuta, the two terga, and the carina of the Lepadidæ, commence their growth, under the form of the so-called “primordial valves,” and so differ from all the other valves when such occur: now, in two species of Verruca, I have found closely analogous primordial valves on the apices of both the moveable and fixed scutum and tergum (thus affording strong additional evidence that their nature has been rightly interpreted), and on one of the two remaining valves, namely, that at the posterior or carinal end of the shell. Hence, we may safely infer, that this latter valve, which, though very much more developed on one than on the other side, is so far medial as to curl round and cover the line of opening between the moveable and fixed tergum, is really a carina. The sixth valve differs only very slightly in shape from the carina, and is directly opposed to it; therefore, in accordance with all analogy, it must be the rostrum. Consequently, the shell in Verruca consists of a moveable scutum and tergum, a fixed scutum and tergum, a carina and rostrum, and, as we shall immediately see, a membranous basis—the basis being, as in all sessile cirripedes, the homologue of the peduncle in the Lepadidæ.
The moveable scutum and tergum stand at about right angles with the fixed pair; and as these latter form a part of the wall of the shell, which is always steep on this side, the moveable pair, which close the orifice, are nearly horizontal or parallel to the basis and surface of attachment. Hence, the animal’s body, which is attached between the two scuta, but nearest to the moveable scutum, also, lies nearly parallel to the surface of attachment; and I was consequently at first led to suspect that the basal membrane was one side of the shell in a modified condition; but the presence of the prehensile antennæ of the pupa in nearly the middle of this membrane, and the sheet of cement-tissue on its under side, demonstrate that this mem[Pg 499]brane, though lying on one side of the animal, is the true basis. To make all the parts in Verruca hold the same position as in other cirripedes, relatively to the surface of attachment, we must develope the carina and rostrum equally on both sides of the true longitudinal axis of the shell, and insert the newly-developed portion between the basis and the fixed scutum and tergum, reducing the latter in size, and tilting a little up the moveable scutum and tergum; and by this means the animal’s body would be turned, so that its dorso-ventral longitudinal plane would stand at right angles to the basal membrane.
Extraordinarily great as is the difference between the right and left sides of the whole shell, yet in all the species it seems to be entirely a matter of chance whether it be the right scutum and tergum with the right side of the rostrum and carina, or the left scutum and tergum with the left side of the rostrum and carina, which become abnormally developed. Nor does there seem to be any relation between the side of the operculum to be attached, whether right or left, and the nature of the surface of attachment; for I have seen many specimens adhering to perfectly level surfaces, and to quite cylindrical branches of Laminariæ; and in these cases, however the larva might attach itself, there could be nothing to favour the development of one side more than the other. Although the attached scuta and terga are larger than the moveable pair, yet, owing to the small development of the carina and rostrum on the attached side, the upper or unattached side must be considered as the most developed. In this respect, and in the circumstance of either right or left side being modified, we are reminded of the structure of Pœcilasma Kæmpferi (described in my former volume on the Lepadidæ), in which the valves on the side of the capitulum, nearest to the crab’s body, to which the specimens were attached, were somewhat less developed than those on the opposite side. I may add, that in ordinary Crustaceans, as I am informed by Professor Bell, the unequal development of the thoracic limbs seems quite capriciously to affect either the left or right side of the body.
General Appearance of the Shell.—The shell is in most cases much depressed and irregularly circular; the side formed[Pg 500] by the fixed scutum and tergum is always steeper than the other side: the colour is white or pale brownish, and in V. nexa pale red. The surface is naked. The size is small, rarely exceeding a quarter of an inch in diameter, and the whole shell often appears like a mere scale on the surface of attachment. The most remarkable feature in the external aspect is due to the suture between the rostrum and carina, which is formed by oblique, interlocking plates or folds; as all these plates continue to be added to at their extremities during growth, the upper plates become longer than the lower ones; and the plates on both sides of the suture together form a triangular area, with the broad end uppermost, somewhat like the radius of a sessile cirripede: they act, also, like a radius, for their growth serves to separate these two valves, and so adds to the diameter of the shell. The suture between the rostrum and fixed scutum and that between the carina and fixed tergum are nearly of the same nature, but the former is more conspicuous than the latter; neither are so conspicuous as that between the carina and rostrum: accordingly as the right or left scutum and tergum are moveable, so the suture, second in plainness, (see Pl. 21, fig. 1 a, and 1 d,) is placed to the left or right hand. The fourth suture, between the fixed scutum and tergum, as viewed externally, is straight, and so very obscure that it has been overlooked by some authors, and the shell described as consisting of only three nearly equal pieces, for the fixed scutum and tergum together are about equal in size to the carina or rostrum. The orifice approaches more nearly to an unequal-sided triangle, with the apex broadly truncated, than to any other figure. The operculum fits with remarkable closeness, and is surrounded by a slight rim, formed by the edges of the four other valves.
Moveable Scutum and Tergum.—The scutum (S in 1 b and 5) is narrow and very small, barely equalling half the size of the tergum, and therefore proportionally much smaller than in any other cirripede; in a very young shell, however, (of V. Strömia) less than a pin’s head in size, the scutum equalled the tergum in size. The valve is remarkably thick; it is generally depressed down the middle; but in V. nexa[Pg 501] this part is longitudinally ribbed. The occludent margin is curved. On the tergal margin there are two articular ridges (with a deepish furrow between them), of which the upper one (′ in S, in fig. 1 b, and 5) extends from the apex about half-way down the valve; and the other, or lower articular ridge (′′ in S), generally runs down nearly to the basal margin: an angle, running from the apex to the basi-tergal corner of the valve, appears like a third articular ridge, but cannot properly be considered such. The above two articular ridges interfold with analogous ones on the scutal margin of the tergum, and so lock the valves together. On the under side (fig. 1 f), the surface is bounded along the occludent margin by a slight rim: there is generally a very slight depression for the adductor muscle; but in V. Spengleri there is a straight, short, sharp (Pl. 21, fig. 2), prominent adductor ridge.
The moveable tergum is broad and rhomboidal. Externally a prominent axial ridge (′′′ in T, in fig. 1 b, &c.), which widens downwards, runs from the apex of the valve to the basal point, and there projecting slightly, causes the scutum to be indented; this indentation on the scutum appears like a third articular ridge, lying beneath (′′), S, in fig. 1 b, &c. Above the lower and axial ridge, on the scutal margin of the tergum, there is a middle articular ridge, which locks in, between the lower (′′) and upper articular ridges (′) of the scutum (S). Again above the middle ridge there is an upper and third articular ridge (′), which is either quite distinct, as in fig. 5, T, or more commonly is formed by the occludent margin of the valve, as in T, fig. 1 b. The broad extremity of this upper articular ridge is often produced into a slight projection, or shoulder, and this always underlies the scutum, of which the under and upper surface is indented or furrowed (see fig. 1 f), in order to receive this shoulder. The upper articular ridge of the scutum (′, S, 1 b) locks in between the upper articular ridge or occludent margin (′, T), and the middle ridge (′′, T) of the tergum.
Hence, altogether, there are three articular ridges on the scutal margin of the tergum, the occludent margin being generally counted as one; whereas, on the tergal margin of the[Pg 502] scutum, there are only two ridges, though, as before noticed, an outer indentation, which is developed as a ridge in V. nexa (fig. 5, S), might almost be counted as a third articular ridge.
I may here just remark, that the furrow between the two ridges on the tergal margin of the scutum, resembles the articular furrow in the scutum of the Balanidæ; but it may be doubted whether the resemblance be more than superficial, as this furrow, in the case of Balanidæ, receives the edge itself of the tergum, whereas here it receives only a ridge, proceeding from the apex of the tergum, to a nearly middle point on its scutal margin. Finally, I may add, that the tergum in this genus, in general shape, in growth (presently to be referred to), in the manner in which the upper scutal shoulder is overlapped by the scutum, and in the presence of the axial ridge, presents a very striking resemblance to certain old fossil species of Pollicipes, and to a limited extent to the living species of Lithotrya.
The scutum and tergum being interlocked, move together; they can be firmly shut by the contraction of the long adductor scutorum muscle. Their opening appears partly due to the elasticity of the membranous hinge (representing the opercular membrane), by which they are attached transversely, just beneath the summit of the carina and rostrum. No doubt the protrusion of the cirri effectively aids the act of opening. These valves are not capable (nor, of course, the other valves) of any other movement; for there are no muscles for such movements.
Fixed Scutum and Tergum.—The fixed scutum is larger than the fixed tergum, and therefore has the same proportions as the homologous valves in ordinary cirripedia, but reversed proportions compared with the moveable scutum and tergum. The shape of neither valve can hardly be described. The fixed Scutum (S′ in all the figs.), externally, seems at first to consist of two portions, namely, a curved occludent rim (a), closely resembling the opposed occludent margin (a) of the moveable scutum (S), having in fact undergone very little modification, (as may be best seen in Pl. 21, fig. 1 b); and secondly, of a much modified portion (marked ′′), which resembles in outline and state of surface the rest[Pg 503] of the walls of the shell, and may be called the parietal portion. The rostrum (A) curls round the end of the occludent portion, under an edge (b), evidently answering to the basal margin (b) of the moveable scutum, and is simply united to this portion by membrane, but beyond this part, it is articulated to the parietal portion (′′) of the fixed scutum, by oblique interlocking ridges, like those forming the suture between the rostrum and carina. In V. nexa, however, (fig. 5) the rostrum (A) does not curl round any part of the interlocking fixed scutum (S′), but articulates with it by a straight suture. Internally, the fixed scutum (S′ in figs. 1 e, 1 c, of reversed shells) has a surprisingly large, thin adductor plate (m in fig. 1 c, 1 b), with a rounded outline, projecting nearly parallel to the basis or surface of attachment; the adductor scutorum muscle is attached to its upper surface, and consequently the animal’s body lies between this plate and the moveable scutum. In the fixed scutum of V. nexa, however, there is a deep pit, instead of a plate, for this muscle.
The fixed Tergum, likewise, consists of two portions—a middle and lower, or parietal portion, and a rim or upper portion; the rim consists of two unequal arms, answering to the two upper margins (not merely the edges) of the rhomboidal moveable tergum; the longer rim (x in T′, see fig. 5) answers to the carinal margin (x) of the moveable tergum, and may be called the carinal rim; and the shorter rim (o and ′ in fig. 5) answers to the occludent margin (o and ′) of the moveable valve, and may be called the occludent rim. The carina curls round the end of the carinal rim, under an edge, z (much foreshortened in T′ in fig. 1 b, and best seen in fig. 5), answering to at least a large part of the basal margin (z) of the moveable tergum, and interlocks, by a serrated suture, with the edge of the parietal portion of the valve. Internally (fig. 1 e, less plain in 1 c) there is a transverse ledge, notched in the middle, and sometimes deeply hollow beneath, running across the valve in about the line of the adductor plate of the fixed scutum: this ledge, in fact, marks and is partly caused by, the line of separation between the central or parietal, much modified, and the scarcely modified, upper or rim portion of the valve. The use of this ledge is apparently to give attachment, as does the under[Pg 504] side of the adductor plate of the fixed scutum, to ligamentous fibres, presently to be mentioned, by which the shell is attached to the basal membrane: the carina and rostrum being so much more gently inclined, do not stand in need of a ledge for their attachment.
By comparing the moveable scutum and tergum with the corresponding fixed valves, in all the species, the modification of the latter may be clearly made out to have been effected as follows; and the case appears to me a striking and interesting one. The moveable scutum and tergum lie in the same plane, and are articulated by the means of three ridges on the tergum (including the occludent margin), and by two on the scutum. The fixed scutum and tergum have to be curved, and to be greatly increased in size; and this is brought about, as we shall see, by the large development of a certain small portion of each valve. Comparing first the moveable tergum (T) with the fixed tergum (T′), the umbo of growth matches the umbo of the four margins of the moveable valve, the carinal (x), basal (z), and occludent (o in fig. 5), margins can be identified with certainty in the fixed valve, from their close similarity in shape, their absolute apposition, or correspondence in position. There remains only the scutal or articular margin, with its three articular ridges; of these, the uppermost (′), inasmuch as in most of the species it is hardly distinct from the occludent margin, can, as we have just seen, be clearly identified, and is overlapped, as it normally should be, by the upper tergal corner of the fixed scutum: the second or middle articular ridge, though not so distinct as in the moveable valves, can be plainly recognised (′′), T′, in fig. 1 b, and 5; and it serves its normal function of articulating the two valves together. But when we look in the fixed valve for the third or axial ridge (′′′), we find in its exact place, namely, extending from the umbo to the extreme opposite end of the valve, between the second articular ridge (′′) and the basal margin (z, see fig. 5), only that portion of the valve which I have called the parietal portion; consequently, I do not doubt that this really is the axial ridge largely expanded. So again in comparing the moveable scutum (S) with the fixed scutum (S′); two of the three margins of the former, namely,[Pg 505] the occludent (a, see fig. 1 b) and basal (b), can be identified without a doubt in the fixed valve: the third and tergal margin remains; this should have two articular ridges; of these the upper one, still serving its normal function, can be detected in all the species (′ in fig. 1 b), and can be seen pretty plainly (′ fig. 5) in V. nexa: but of the lower and other articular ridge there is no sign,—excepting indeed the whole parietal portion of the valve, which, from holding an exactly homologous position with the lower articular ridge of the moveable valve, I cannot doubt in this ridge expanded and curiously metamorphosed. Hence, in both fixed scutum and tergum, it is the outermost or lowest of the articular ridges which has been modified and expanded, so as to rest on and be fixed to the surface of attachment. It would appear as if it had resulted from one ridge in each of these valves having been thus used up by expansion (so to express myself), that the suture between the fixed scutum and tergum is more simple than any other suture in the whole shell; and it is owing probably to this straightness, and consequent tendency to weakness, that the valves do not grow along this line, and so do not become separated from each other during growth, as on the three other lines of suture. As it actually is, owing to this suture never being separated, it is even stronger than the others; its edges on the inside (fig. 1 c), I may add, are a little inflected or prominent.
Rostrum and Carina: these valves differ from each other, only in the former (A) being rather the largest, and in being more plainly articulated with the fixed scutum, than is the carina (B) with the fixed tergum. Their umbones stand in their normal places, at the two ends of the orifice leading into the sack, that is, facing the dorso-ventral longitudinal plane of the animal; but they are very unequally developed on the two sides, and hence they rise very obliquely from the surface of attachment. Their summits are nearly square, which is caused by the continued growth on both sides of the oblique plates or ridges, by which they are articulated with the adjoining valves. These plates strikingly resemble, as already stated, the radii in certain species of Chthamalus. Without these articulating plates, the outline of the rostrum and carina would have been triangular, with the apex up[Pg 506]wards. In V. nexa, in which the walls of the shell are almost perpendicular, the rostrum (A, fig. 5) is very peculiar and patelliformed, with the umbo sub-central: this results from the development of a border at the upper end of the valve. In this same species, the basal edges of the rostrum, carina, fixed scutum and tergum, are rectangularly inflected, so as to form a ledge round the basis, as in the case of some few species of Chthamalus,—the ledge appearing like part of the real basis. During the growth of the shell, the upper internal ends of the carina and rostrum are either rendered solid, or a ledge is formed on the inside across their summits, hollow beneath, like the sheath of the Balanidæ, to which solid or hollow ledge the basal margins of the moveable scutum and tergum are attached by a rim of membrane, forming a hinge.
Direction of Growth: Minute Structure of Valves.—The shell grows downwards all round its basal margin. As far as the diametric growth of its upper part is concerned, there may be said to be only three valves, for the fixed scutum and tergum never become, as already stated, separated; on the three other lines of suture, the valves are added to on both sides; and thus the whole upper part of the shell, and the orifice, increases in diameter. The moveable scutum and tergum grow along their basal margins, and along the margins by which they are articulated together; but the scutum in this latter respect, less than the tergum. The summits of the moveable scutum and tergum, during continued growth, become either worn away, or they project freely; in this latter case, an internal ledge is added round the upper end of the fixed scutum and tergum, so as to keep the orifice accurately closed. In V. nexa the rostrum, with its sub-central umbo, is anomalous, as already stated, owing to a broad upper internal border growing in a direction almost directly opposed to the basal growth of the moveable opercular valves.
In young specimens, on the apices of both scuta and both terga, and on the carina, but not on the rostrum, primordial valves may be distinguished, resembling the valves, so called, which first appear (p. 129, Introduction) after the metamorphosis in, the Lepadidæ. In the Verrucidæ, how[Pg 507]ever, they are calcareous; and the minute transverse cylinders, of which they appear to be composed, stand further apart, causing the surface of the primordial valve to be marked with little separate circles, instead of by hexagons.
The shelly matter of which the valves are composed is translucent: it is remarkably destitute of any investing membrane. The under surface is marked with rows of minute approximate pores, parallel to the lines of growth, into which the corium enters: after a portion of shell has been dissolved in acid, these threads of corium are seen to change, a short distance within, into cylinders of yellow chitine, running obliquely through the substance of the valve. These cylinders are about 1/2000th of an inch in diameter, but in parts they are spindle-shaped and twice as thick: they vary in length, about 1/100th of an inch being the average length: these cylinders at their upper ends suddenly contract into a point, more or less long, or are produced into a very fine tortuous tubulus of chitine, imbedded in the shell: I have seen in no other Cirripedes tubuli of this structure. There are other ordinary tubuli, such as occur in the valves of most Cirripedes, about 1/6000th of an inch in diameter, and which sometimes alternate with the above-described thicker cylinders. There are no external spines. From the number and length of the tubuli of both kinds, the tissue left after the action of acid is singularly complicated.
Basis.—The basal membrane is thin, and is divided, but not very plainly, into concentric slips, marking the successive increments of growth. In the middle of it, in two young specimens, I found with great difficulty the pupal prehensile antennæ: they were of small size, measuring from the extreme edge of the main or second segment to the end of the disc, only 27/6000ths of an inch: the disc appeared narrow (as in Pollicipes and Scalpellum), with a single spine at the proximate end: the ultimate segment, placed as usual at about right angles to the disc, bore two groups of shorter and longer spines, but I could not count how many. The antennæ were enveloped in a mass of cement of a yellow colour, resembling in all its characters the cement of other Cirripedes. In only one case, I believe I saw bifurcating cement-ducts, of extreme[Pg 508] tenuity, viz. 1/15,000th of an inch in diameter. The sheet of cement on the whole under side of the basal membrane, not rarely shows a very irregular reticulated structure. For convenience sake, it will be best to defer the discussion on the very anomalous, though slight, powers of excavation which this genus possesses, and which I must attribute to the effects of some substance secreted probably by the cement-organs. I will here only mention, that the specimens which have excavated a depression, are less firmly attached than those, which have not acted on their support; and that, in the former case, the basal membrane, for a considerable space in the middle, becomes quite detached.
Animal’s Body.—The body is much flattened and, owing to the little development of one side of the shell, lies parallel to the surface of attachment. The prosoma is but little protuberant. The articulations of the thorax are unusually straight and transverse. The Mouth is also much flattened: it is placed rather distantly from the adductor scutorum muscle, owing to the production of the lower margin of the labrum. The Labrum is not notched, or even hollowed out in the middle, or (excepting in V. nexa) bullate; its crest is surmounted by about eight (more numerous in V. nexa) little teeth, or by some fine bristles. The Palpi are of moderate size, with their tips nearly meeting; they are slightly curved, and have bristles only on their outer sides and extremities: they are apparently capable of being lifted up and down by a muscle attached to them, just outside the rounded swelling on each side of the labrum to which they are articulated: in V. nexa, however, the palpi are very small and narrow, and their tips do not nearly meet. In this genus, therefore, we find the swollen state of the labrum and the size of the palpi—characters generally invariable and of high classificatory importance—variable. The mandibles have three upper main teeth, with two or three minute lower teeth, or, in V. nexa, with the lower part pectinated with small spines: in V. Strömia, I have seen traces of the second tooth being laterally double—a character of some importance. The Maxillæ have a notch under the upper pair of large spines, with the lower part bearing, as usual, a double row of bristles, and forming a large step-formed projection: these organs are furnished[Pg 509] with the usual apodeme and muscles. The Outer Maxillæ are prominent, and deeply lobed on their inner surfaces, the two lobes being clothed with bristles.
Cirri.—The first pair are attached, as usual, on each side of the mouth, and stand some way apart from the five posterior pairs. The second and third pairs differ considerably in structure from the three posterior pairs, which are much elongated. The first pair (excepting in V. nexa) is short, with the two rami slightly unequal in length, and with the segments thickly clothed, as usual, with spines. The second pair is remarkable from the posterior ramus being more than twice as long, and containing thrice as many segments, as the anterior ramus, which is barely as long as the shorter ramus of the first pair: the segments in the anterior ramus of the second pair (only five in number in a full-sized specimen) are broader and more protuberant in front, and more thickly clothed with spines (the terminal spines being doubly pectinated), than are the segments on the posterior ramus; on the latter, the uppermost segments have their bristles arranged in front in simple pairs, with the dorsal spines long, the lower segments being more thickly clothed with bristles, owing to the development of lateral rows. The third pair resembles in every respect the second pair, except in being a little longer, and in the bristles on the posterior ramus being less crowded, more resembling the arrangement of those on the posterior cirri. In V. nexa, however, there is not so great an inequality in length or dissimilarity in structure in the two rami of the second cirrus, and only a very slight difference of any kind in the two rami of the third pair. Fourth, fifth, and sixth pairs have numerous elongated segments, bearing four or three pairs of long slender spines in front, with a single minute bristle between each pair, and with two or three slender spines in the dorsal tuft.
There is a considerable amount of variation in the proportional length, and in the number of the segments, of the several cirri in V. Strömia; in some specimens the two rami of the fourth pair were unequal in length; in some, nearly all the cirri on the lower or attached side were shorter than those on the upper side.
Caudal Appendages.—These are of most unusual length, sometimes even exceeding those of Ibla quadrivalvis, which surpasses, in this respect, all other cirripedes. They arise on each side and over the anus. They consist of numerous (sometimes as many as twenty-four), unequal, cylindrical, thin segments, bearing, at their upper ends, a circle of long and very slender spines. They sometimes equal two thirds or even four fifths of the length of the sixth cirrus; but their length, and the number of their segments, (sometimes imperfectly divided), varies much in different specimens of the same species, and sometimes even on opposite sides of the same individual. In some very young shells, as big as a pin’s head, the caudal appendages were proportionally extremely short, and consisted of only two or three segments. No muscles enter these organs; and when the animal is taken out of its sack, they project straight out behind, instead of being curled in, like the cirri.
Anatomical Structure.—The animal’s body is attached to the two scuta by the adductor scutorum, and by the other usual muscles running towards the mouth, and surrounding the prosoma. The whole external covering or shell has no other muscles; Verruca thus differing from the Balanidæ and Lepadidæ; but the shell is attached all round, near its circumference, to the basal membrane, by a band of very short fibres, appearing like muscles, but really ligamentous, as determined for me by Professor Quekett. Branchiæ are entirely absent. The alimentary canal presents all the usual characters, but in the prosoma is rather abruptly bent back on itself. The orifices of the two olfactory pouches are not at all prominent; they are placed directly under the outer maxillæ, (homologically in their middle segment), just above a small, medial, tongue-like apodeme. The orifices of the acoustic sacks appeared to be in their usual position beneath the basal articulations of the first pair of cirri. The vesiculæ seminales occupy their usual position in the prosoma; they are not much convoluted; they unite before entering the penis. The probosciformed penis is imperfectly ringed; it is thick and short, and tapers much more abruptly than is usual; it supports a few very thin hairs. The ovarian cæca are[Pg 511] spread over the basal membrane, at the bottom of the sack; hence they in fact lie almost on one side of the animal: they consist of two main trunks, proceeding out of the animal’s body at the rostral end of the sack, which then branch and inosculate. In specimens of V. Strömia collected by Mr. Peach for me, in Cornwall, during the first week of April, there were included two ovigerous lamellæ, placed transversely across the rostral and the carinal end of the sack: the lamellæ were .11 of an inch in length; they appeared loose and not attached, as in the Lepadidæ, to any ovigerous fræna. The ova, in their earliest age, have one end much pointed, and are 8/1000ths of an inch in length; they become blunter and increase a little in size before the larvæ burst forth. The larvæ, both during their earliest stage and after the first moult, have been excellently figured and described[137] by Mr. C. Spence Bate: they present no particular characters distinct from the larvæ of other Cirripedes. I will only further add, that the structure of the prehensile antennæ still adherent to the basal membrane, indicates that the larva in its last stage,—that is the locomotive pupa,—has a normal character.
[137] ‘Annals and Mag. of Nat. Hist.,’ 1851, Pl. 7, fig. 8-10.
Affinities.—These have been sufficiently discussed under the family; I need here only remark that all the species, with the exception of V. nexa, are intimately allied together.
Range—Habits—Geological History.—The genus Verruca ranges, being represented by four species, from Iceland to Cape Horn. The species that is found in Tierra del Fuego extends up the west coast to Peru. Our northern form, V. Strömia, (if I may trust a specimen in the British Museum, apparently ticketed in an authentic manner), occurs also in the Red Sea; and this is the only locality in the eastern hemisphere whence I have seen this genus. The species seem generally to live in rather deep water: I procured V. lævigata from nineteen fathoms, on the east coast of Patagonia: V. Strömia is found, according to information given me by Professor Forbes, on the British shores, between five and fifty fathoms, and on the steep shore off Mull, in ninety fathoms; but Mr. Thompson assures me[Pg 512] that he once saw it adhering to tidal rocks and likewise to some floating bark. Generally the species are attached to living organic bodies, especially shells of Mollusca and of Cirripedes, to Gorgoniæ, and Laminariæ; less frequently to rocks. We shall immediately see that it has slight powers of excavation. This genus is geologically older than any true sessile cirripede or member of the Balanidæ: V. Strömia is found in the Glacial Deposits and in the Red and Coralline Crag of England: another species (in a state not to be identified) occurs in the ancient Tertiary formations of Patagonia; and another in the Chalk of England and Belgium. The fact of this Family ascending to a Secondary epoch accords, in an interesting manner, with its affinities; inasmuch as though in appearance a sessile cirripede, it is almost equally related to the Lepadidæ and Balanidæ, and is more nearly related to the Lepadidæ than to the Balaninæ, or typical members of the Balanidæ: of the latter, none have hitherto been found in any Secondary deposit, whereas the Lepadidæ culminated during the Cretacean period.
My attention was called to this subject by Mr. Hancock, whose excellent researches on the boring of Mollusca are well known. Verruca Strömia, when attached to shells destitute of an epidermis, excavates, as he informed me, a slight depression, deepest in the middle; but when the epidermis is present no effect whatever is produced. We shall presently see that the central depression is in some degree distinct from that of the circumference. I have since found Mr. Hancock’s observations strictly applicable to V. lævigata, V. Spengleri, and to an ancient tertiary species from Patagonia. From having found that the following cirripedes, viz., Lithotrya, Alcippe, and Cryptophialus, all form their deep excavations by mechanical means, and from having read the above-mentioned memoirs by Mr. Hancock on the boring of mollusca, I was strongly impressed with the idea that the action in Verruca would likewise prove mechanical: but from the following facts I[Pg 513] have come to the conclusion that the excavation must be due to a solvent, probably poured out from the cement-ducts, which debouch on the under side of the basal membrane.
In the first place, an epidermis, as just stated, perfectly preserves the shells of the various species of mollusca and certain cirripedes, to which I have seen Verruca attached: this is well shown by comparing the effect produced on the same shell in parts covered by the epidermis and in parts whence it has been abraded; or where the shell of the Verruca had fixed itself, whilst very young, within a crack in the epidermis, and had subsequently, by its growth, turned up the edges, and had then acted on the underlying shell; whereas the specimens attached to the sound epidermis had not produced the smallest effect. Again, I have seen an epidermis-covered mussel-shell encrusted by a hard nullipora, on which V. lævigata was attached; and here the calcareous nullipora, under the middle of the basal membrane, was entirely corroded away, whilst the underlying epidermis and the shell beneath it, were not in the least affected. The protection afforded by the epidermis is still more strikingly shown by contrasting shells with very sharp prominent ridges, when thus invested and when naked, to which Verrucæ have been attached: I have given a figure (Pl. 21, fig. 6) of a piece of an invested Venus, from the surface of which a V. Spengleri had been just removed; on the other hand, I have seen a Peruvian Discina in which even sharper ridges, covered by epidermis, were left absolutely untouched, although projecting deeply into the shell of an attached V. lævigata. I have seen several specimens of this latter Verruca (which has the power of corroding naked shell as deeply as its congeners), attached to the membrane-covered variety of Balanus lævis, the shell of which was thus perfectly preserved: now this membrane is little more than the 1/2000th of an inch in thickness; it is not hard, and so brittle that it generally separates with the Verruca, leaving the underlying shell of the B. lævis with its lines of growth glossy and perfect: it appears to me impossible that a membrane so thin and brittle could resist an action, if mechanical, which has worn away from twenty to forty times as great a[Pg 514] thickness of hard shell; but the thinnest film of any matter on which acid does not act, as of grease in certain forms of printing, will perfectly preserve the underlying substance, and as I have ascertained by putting on a drop of acid, is the case with this membrane. I have removed several scores of shells of V. Strömia from the stems of Laminariæ, and when the latter were washed and slightly dried, generally not the least effect could be seen, except that the spots where the shell had adhered were glossy from the still adherent basal membrane: yet the stems of Laminariæ are far from hard. In some cases, however, the attachment of the Verruca seemed to have produced a very slight depression on the Laminaria, but this, I think, may be safely attributed to the growth of the surrounding surface; for I have seen exactly the same effect produced by the attachment of the discs of the antennæ of a Lepas, and these discs, with their long spines, could not possibly produce any excavation; nor is Lepas or its pupa in any case a burrowing animal. Again, I have seen a few specimens of Verruca attached to Gorgoniæ, and they had not acted in the least on the horny axis. I have examined numerous specimens of V. Strömia attached to three pieces of slate-rock, and to one piece of red sandstone, all from different localities, and no effect whatever had been produced; yet the slate-rock, especially in one instance, was soft. Mr. Bate, to whom I am indebted for some of these specimens, also informs me that he could discover no impressions on the slate-rocks, whence specimens of the Verruca had been removed. On the other hand, I have had two specimens of limestone, with attached Verrucæ, one coarse and very impure, and the other hard and marble-like; and in both cases there was a distinct central slight cavity, including loose gritty matter. The loose particles evidently resulted from the unequal action either of a solvent or of some mechanical power on the rock, for it is improbable in the highest degree that the shells should have fixed themselves exactly over small collections of loose particles, even if such could possibly have remained on projecting surfaces of sea-washed rocks.
The above facts seem to indicate pretty plainly that the[Pg 515] excavation of the support does not depend on its hardness, but on its containing calcareous matter, liable to be acted on by some solvent: but as this view, considering what we know of Lithotrya and of the two other burrowing genera of cirripedes to be hereafter described, appears improbable, I will add a few additional observations. I most carefully examined the shell and basal membrane of Verruca, and likewise the tissues left after the dissolution of the shell in acid, and could detect no structure at all fitted for boring; and what appears more important, there was no apparent difference in the state of the specimens which had and had not excavated a hollow; and this, I think, would certainly have been the case (as in Lithotrya) if the action had been mechanical. It is not easy to ascertain, owing to the small effect at any time produced, at how early an age Verruca begins to act on its support; but I found two sets of specimens only 1/20th of an inch in basal diameter, which had certainly commenced. The ribbed shell, (Pl. 21, fig. 6), especially the middle rib, shows, in a somewhat exaggerated degree, the typical form of the excavation; it may be here seen that the excavation is of the same depth for some little distance from the circumference towards the centre, but that in the middle it suddenly becomes deeper. I have seen several specimens with a central hollow, without any, or with scarcely any, marginal depression, and likewise the reversed case. These several facts show that the central excavation cannot be due to an equable action, prolonged during the whole growth of the shell, having thus affected the middle more than the circumferential parts, for in this case the excavations would have sloped into each other. In specimens which have not at all acted on their support, the whole basal membrane is firmly attached, as in all ordinary cirripedes, to the supporting surface; but in those which have acted, the middle portion of the basal membrane is quite unattached, and the circumferential portion is, I think, less firmly attached than is usual; but between these two portions, there is a circular zone strongly cemented to the supporting surface, and which alone keeps the shell in its place. Now, on the mechanical theory, to account for the circumferential hollow, the basal edges of[Pg 516] the shell together with the circumference of the basal membrane must be subjected to movement, but the shell is united to the basal membrane by corium and by transparent structureless chitine (both of which may be left out of question) and by a circle of short fibres, which adhere at their lower ends to the firmly cemented circular zone, and by their upper ends to the shell; and these fibres have been very carefully examined by Professor Quekett, and pronounced to be not muscular, but exclusively ligamentous, and therefore incapable of moving the edge of the shell. The basal membrane over the central hollow is, as stated, quite loose: its lower surface, formed by a reticulated layer of horny cement-tissue, shows no signs of abrasion, and the membrane is so brittle and tender, that in specimens which have been once dried and then well soaked, it almost invariably cracks when the shell is removed, owing to its mere adhesion to the delicate inner tunics of the sack; yet on the mechanical theory, the wearing of the central hollow must have been caused by the action of this middle portion of the basal membrane,[138] which, it may be repeated, is destitute of muscles. From the presence of the prehensile pupal antennæ, enveloped in cement, nearly in the centre of the basal membrane, it is certain that this spot was originally attached to the supporting surface, and has since been detached from it; as, moreover, the central hollow goes on increasing in diameter with the growth of the shell, it is certain that the inner edge of the firmly attached circular zone of basal membrane must likewise continually go on becoming detached: it may, then, be asked by what force can the basal membrane, seeing that it is united to its own shell above only by fibres of ligament near the circumference, be continually torn away from the underlying support, to which[Pg 517] it is strongly cemented? On the other hand, on the theory of a solvent slowly poured out from the cement-ducts, its separation from its support is simply explained. It might be supposed that the calcareous matter, when dissolved, would not be able to escape from the central hollow, owing to the basal membrane being so firmly cemented all round it; but the attachment is by a reticulated layer of cement; and I infer that it must be permeated by open passages, from the fact of the hollow being often filled, in dried specimens, by a bubble of air, instead of the basal membrane being pressed closely down into the hollow, as would have been the case had the hollow been hermetically sealed up. I have seen a few instances in which the bottom of the central hollow was occupied, (as was remarked to me by Mr. Hancock), by a little chalky and gritty matter; and in the case of one of the specimens of calcareous rock, before alluded to, by coarse grains and oxide of iron; this seems quite compatible with a solvent acting more readily on certain parts of the rock or shell than on other and less soluble parts or particles.
[138] Mr. Hancock suggests to me that the basal membrane, on the mechanical theory, need not itself move; the motion of epithelial scales, were they transferred into cutting agents, might be supposed to be sufficient. But of such scales, though I used very high powers, I could see no trace; and their presence on the under side of the layer of cement seems hardly possible. Moreover, according to Von Siebold (‘Anatomie Comparée,’ tom. 1, p. 412), ciliary action has not been observed in any Crustacean, or indeed any Articulate animal. This same statement is likewise made in Annals and Mag. of Nat. Hist. 1854, p. 136, by Dr. T. Williams.
The greatest depth of the central hollow, in any specimen seen by me, even measuring from the top of a rib in the case of a ribbed shell, to the deepest point, was only 1/50th of an inch; but considering how much depressed the shell of Verruca is, I have no doubt that this small gain of space is of service to the animal: we must suppose the loose middle portion of the basal membrane is stretched slightly, or splits and is repaired, so as to fit the hollow. With respect to the even much slighter circumferential excavation, it barely equals in depth the thickness of the extreme edges of the walls; it must, I presume, give strength to the shell when laterally pushed; but it certainly appeared to me that the individuals which had excavated a depression for themselves, could be pried vertically up much more easily than those which had not acted on their support. Finally, we must suppose that the hypothetical solvent is poured out of the cement-ducts at the extreme circumference of the basal membrane, which is almost loose and destitute of cement, so as to slightly corrode outwards and downwards the calcareous support; the action here then stops, and this rim[Pg 518] of basal membrane becomes, after a new rim has been formed and as the shell grows outwards, firmly cemented down to the now slightly excavated surface of attachment; but during all the time the solvent goes on acting in the middle, and continues, during the whole growth of the shell, to encroach on and dissolve the supporting surface from under the inner edges of the previously cemented down, circular zone of basal membrane. I have discussed this subject at considerable length, as it appears to me an interesting one. In this case we have the action of ciliæ[139] and of respiratory currents, to which in the case of Mollusca so much has been attributed, entirely eliminated. It is, also, an interesting fact, that within the same Order we should have some Cirripedes boring by simply mechanical means, and others by a chemical solvent.[140]
[140] The solvent may be carbonic acid gas, as suggested by Mr. C. S. Bate in the case of Mollusca (‘Report of British Association,’ 1849, p. 73), but here, under the basal membrane, we cannot have the respiratory currents, or the ciliary action (see note, supra), as likewise suggested by Mr. Bate.
LEPAS STRÖMIA. O. Müller. Zoolog. Dan. Prod., No. 3025, 1776.
---- ------ Ib. Zoolog. Dan., vol. 3, Tab. 94, 1789.
---- STRIATA. Pennant. British Zoology, vol. 4, Tab. 38, fig. 7, 1777.
DIE WARZENFORMIGE MEEREICHEL. Spengler. Schriften der Berlin. Gesell., 1 B., Tab. 5, fig. 1-3, 1780.
LEPAS VERRUCA. Spengler. Skrifter af Naturhist. Selskabet, 1 B., 1790.
---- ------ ET STRÖMIA. Gmelin. Syst. Nat., 1789.
BALANUS VERRUCA. Bruguière. Encyclop. Meth., 1789; Clisia verrucosa, Deshayes, in Tab.
------ INTERTEXTUS. Pulteney. Catalogue of Shells of Dorsetshire, 1799.
LEPAS STRIATUS. Montagu. Test. Brit., 1803.
---- VERRUCA. Wood’s General Conchology, Pl. 9, fig. 5, 1815.
VERRUCA STRÖMII. Schumacher. Essai d’un Nouveau Syst. Class., 1817.
CREUSIA STRÖMIA ET VERRUCA. Lamarck. Animaux sans Vertèbres, 1818.
OCHTHOSIA STROEMIA. Ranzani. Memoire di Storia Nat., 1820.
CLISIA STRIATA. Leach. Encyclop. Brit. Suppl., vol. 3 (sine descript.), 1824.
CLITIA VERRUCA. G. B. Sowerby. Genera of Recent and Fossil Shells, Plate.
VERRUCA STRÖMII. J. E. Gray. Annals of Philosophy (new series), vol. 10, Aug., 1825.
Moveable scutum, with the lower articular ridge not half as broad as the short upper articular ridge: shell generally ribbed longitudinally.
Var., with the shell not longitudinally ribbed.
Hab.—Shores of Great Britain and Ireland, Shetland Islands; and, according to various authors, Denmark, Iceland, and shores of northern Europe. Red Sea, Brit. Mus. Attached to shells, laminariæ, rocks, crabs, and floating bark, from low tidal mark to fifty or ninety fathoms.
Fossil in Glacial deposits of Scotland, Mus. Lyell; Red Crag (Walton, Essex), Coralline Crag (Sutton), Mus. S. V. Wood.
I have given so full a description of the genus that little remains to be said under the species. Generally the whole shell is covered (independently of the interfolding, oblique, articulating plates) by narrow, longitudinal ridges or folds; and by this character alone the ordinary variety of V. Strömia can be distinguished (as far as I have seen) from all the other species. The shell is white or dirty yellowish-brown. The scutum has the lower articular ridge on its tergal margin very narrow (but somewhat variable in width), appearing like a mere slight shoulder, against which the longitudinal axial ridge of the tergum abuts: it is not half as wide as the short, upper articular ridge. On the under side there is a very slight depression for the adductor scutorum muscle. There is considerable variation in the degree to which the transverse ledge on the under side of the fixed tergum projects, and therefore in the depth of the hollow thus formed. The specimens with the right-side, and those with the left-side opercular valves moveable, are apparently about equally numerous.
The specimen in the British Museum, from the Red Sea, was attached to a Gorgonia, and was in the same box with a Pyrgoma—circumstances favouring the correctness of the[Pg 520] locality—but I am much surprised from the general distribution of the species, that V. Strömia should occur in so distant and isolated an area. After careful examination, I can discover no constant difference between the Red Sea and British specimens.
The specimens from the Crag have not their moveable opercular valves, which offer much more important diagnostic characters than the shell; but as far as the latter is concerned, no difference whatever can be perceived from V. Strömia.
VERRUCA LÆVIGATA. G. B. Sowerby. Genera of Recent and Fossil Shells, Plate.
Moveable scutum, with the lower articular ridge broader than the short upper articular ridge; moveable tergum broader than high, with the upper articular ridge produced into a point.
Hab.—Tierra del Fuego; Eastern Patagonia, nineteen fathoms; Chile; Peru; Mus. Brit., Cuming, Stutchbury, Darwin: attached to shells, and often to Balanus lævis and psittacus.
I can point out no difference in the shell between this species and V. Strömia, excepting that its walls seem invariably to be smooth, which is rarely the case with V. Strömia; perhaps also the oblique interfolding articular plates between the several compartments are here more prominent. It appears that specimens with the left side uppermost, and therefore with the left opercular valves moveable, are considerably more common than those with the right valves moveable. The moveable scutum and tergum are articulated together by much more prominent articular ridges than in V. Strömia, and the two valves together are broader in proportion to their height,—the height being measured from the apex to the basal margin. In the scutum the lower articular ridge is considerably broader than the short upper ridge. In the tergum, the[Pg 521] basi-carinal corner is more rectangular, and the whole valve is nearly square: owing to the deep furrow receiving the lower articular ridge of the scutum, the axial ridge of the tergum is proportionally narrower but more prominent than in V. Strömia; the uppermost ridge (formed by the occludent margin of the valve) projects, especially when viewed on the under side (fig. 3 b), as a moderately sharp point.
In the mouth, the lower teeth of the mandibles are more distinct than in V. Strömia; the lower part of the edge of the maxilla is very prominent. In the second and third pairs of cirri the terminal spines on the shorter rami are coarsely pectinated; on the sixth pair there are only three pairs of main spines on each segment; but these several points, according to the analogy of other species, I should expect to be variable.
This species is alluded to by Bruguière, in the ‘Encyclopédie Méthodique,’ but was confounded by him with the V. Strömia of Europe.
Moveable scutum, with a sharp, straight, medial adductor ridge: fixed scutum not larger than the fixed tergum.
Hab.—Madeira, Mus. Lowe; attached to shells.
It would appear that the present species does not attain quite so large a size as the more northern V. Strömia; the walls are not longitudinally ribbed as is usual with this latter species. The proportional sizes of the compartments seem to be somewhat different; the fixed scutum is either equal to or even smaller than the fixed tergum, instead of being larger, as in V. Strömia; but in young individuals the proportions are reversed. In several specimens the fixed scutum and tergum together were larger than the carina. The rounded adductor plate of the fixed scutum is extremely large. The lines of growth, especially on the moveable opercular valves, are rather more plainly crenated than in V. Strömia. In the moveable scutum the lower articular ridge on the tergal margin varies a little in size, and is[Pg 522] sometimes larger than in V. Strömia (but never so large as in V. lævigata), and is placed more in the middle of the tergal margin: but by far the most important character by which this species can be distinguished from all the others, is the presence, on the under side of the moveable scutum, of a straight, prominent adductor ridge, which runs up to and even under the apex of the valve, for it is there slightly hollowed out. In the moveable tergum, owing to the medial position of the lower articular ridge of the scutum, the middle of the scutal margin is more hollowed out, and the axial ridge narrower, than in V. Strömia.
In the animal’s body the only difference which I could perceive was that the shorter rami of the second and third pairs of cirri were not so short, compared either to the other cirri or to the longer rami of these same cirri. In the second cirrus, in a moderately-sized specimen, the segments were six and thirteen in number in the two rami, and in the third cirrus, seven and fifteen.
Had it not been for the specimen in the British Museum of V. Strömia, from the Red Sea, I should have concluded, from geographical considerations, that V. Spengleri probably was the species found in the Mediterranean, and noticed by Spengler (‘Schriften der Berl. Gesell.,’ 1 B., 1780), as a small variety of the northern V. Strömia; and likewise that it was the Creusia echinoides of Risso (‘Hist. Nat. Product. de l’Europe,’ tom. 4, p. 382, 1826), which is certainly a Verruca, but not described with sufficient minuteness to be recognised.
Shell reddish: moveable scutum, with three strongly prominent longitudinal ridges, besides the articular ridges: fixed scutum larger than the carina, with no distinct adductor plate.
Hab.—West Indies, Mus. Brit.; attached to a Gorgonia.
This species differs considerably from all the others in the genus. The shell is brownish-red, tinted yellow: it is[Pg 523] not at all depressed like the former species, but the walls are almost perpendicular or even overhang their bases, and the summit of the shell consequently is broad. This form may be in part, but only in part, due to the attachment on the thin branches of the Gorgonia. The umbones of the compartments are remarkably prominent and sharp. Although the parietes are nearly smooth, yet from being so steep, they are little seen, and owing to the very prominent but rounded ribs by which the compartments and opercular valves are articulated together, the whole shell has a strongly ribbed appearance. The diameter of the largest specimen was .2 of an inch.
The rostrum (A, fig. 5) is patelliformed, with the umbo of growth sub-central, but rather above the middle point; hence this valve, differently from the carina, and differently from the rostrum of the other species, grows not only at its basal margin, and on both sides where opposed to the carina and fixed scutum, but also along its upper margin where opposed to the basal edges of the moveable scutum and tergum: owing to the perpendicularity of this valve, the upper part forms a ledge almost parallel to the orifice of the shell. The carina (B) is of unusually small size, being about only half the size of the rostrum, and scarcely exceeding in size the fixed tergum. The fixed scutum (S′) is large, larger even than the carina; it is oblong, and its shape is more simple than in the other species; this is chiefly owing to the rostrum articulating with the whole of that margin (b) which answers to the basal margin of the moveable valve; whereas in the other species (fig. 1 b) it curls beyond this margin, and articulates with the very protuberant, so-called, parietal portion of the valve. Three or four rounded prominent longitudinal ribs, exactly like the homologous ribs on the moveable scutum, run from the apex of the fixed scutum to the basal margin, and their extremities form the teeth by which it articulates, as just stated, with the rostrum. Its upper articular ridge (′) is more prominent, and placed much lower down in the suture between it and the fixed tergum, than in the foregoing species. The ledge (o) by which the orifice is kept neatly closed, is here more distinct than in V. Strömia: this ledge is neces[Pg 524]sary, as well as in the case of the fixed tergum, owing to the altered shape of the summits of the moveable scutum and tergum, due to their corrosion and to their coming to project freely. But the most remarkable character of the fixed scutum is, that on the under side there is no great adductor plate, but a rounded hollow with its lower edge only slightly prominent; the absence of the adductor plate, which is present in all the other species of the genus, is no doubt due to the under side of this valve being inclined even outwards, and so standing in some degree opposed to the moveable valve; thus affording on its under surface a place for the attachment of the lower end of the adductor scutorum muscle; whereas in the other species this muscle could not possibly have been attached, without the aid of an adductor plate, to the under side of the much depressed and sloping fixed valve. The fixed tergum (T′) is a little more simple in form than the corresponding valve in the other species; the two arms, answering to the occludent and carinal margins of the moveable tergum, are more nearly equal in length: the internal transverse ledge, separating these rims or margins from the parietal portion of the valve, is but little developed.
All four valves forming the shell are remarkable from having, when full-grown, but not whilst young, their basal edges abruptly inflected inwards, thus forming a ledge all round the basal membrane, as in Chthamalus intertextus and Hembeli.
Moveable Scutum.—This is slightly larger in proportion to the tergum than in the foregoing species: it is chiefly remarkable from the presence of three prominent longitudinal ridges on the main part of the valve, like the two articular ridges on the tergal margin; of these latter, the lower one extends down to about the middle of the tergal margin. The moveable tergum is rhomboidal, with the whole carinal portion marked only by lines of growth: it is only remarkable by the upper of the three articular ridges on the scutal margin being unusually distinct from the occludent margin.
With respect to the animal’s body, its several peculiarities have already been pointed out under the genus. The[Pg 525] labrum is decidedly bullate, triangular in section, with a row of minute bead-like teeth on the crest; the palpi are very narrow and short, and do not nearly touch each other: this variation in the structure of the labrum and in the size of the palpi, is very remarkable, considering how important, in a classificatory point of view, these parts are in all other Cirripedes. In the mandibles there are either two or three main teeth, with the whole lower part of the organ pectinated with sharp spines. Cirri: the first pair is not short; in the individual examined, the two rami had eleven and twelve segments. In the second pair, the shorter ramus was two thirds of the length of the longer ramus, the segments being in number ten and fifteen; in the arrangement of the spines this second pair resembles its homologue in the three other species. In the third pair, the two rami are very nearly equal in length, having sixteen and eighteen segments; and the segments of the anterior ramus are only a little thicker and more thickly clothed with spines than those of the posterior ramus. The remaining cirri and the caudal appendages are as in the other species.
VERRUCA PRISCA. Bosquet. Monographie des Crustacés fossiles du Terrain Crét. de Limbourg, Tab. 1, fig. 1-6; 1853.
Shell smooth: moveable scutum, with the lower articular ridge somewhat broader than the upper articular ridge.
Fossil—‘Système Senonien et Maestrichtien,’ Belgium, Mus. Bosquet; in Chalk, Norwich, Mus. J. de C. Sowerby.
M. Bosquet has admirably figured and described the several separated valves belonging to this species, and I owe to his great kindness an examination of some of them. In Mr. J. de C. Sowerby’s collection, also, there is a single specimen, attached to a Mollusc, with the four valves of the shell united together, but without the two moveable opercular valves; it cannot be positively asserted that this is the same species with that of M. Bosquet, but such probably is[Pg 526] the case. This is the species to which I alluded in the Introduction to my ‘Monograph on Fossil Lepadidæ.’ It is an interesting species, from being the only known Secondary one, but in itself it is a very poorly characterised form, and I can point out no important character in the shell by which it can be recognised. The rostrum and carina, which are of nearly equal sizes, are locked together by the usual interfolding plates, and likewise to the fixed scutum and tergum; but these latter plates seem to have been less developed in M. Bosquet’s specimen than in the English. The fixed scutum has a large adductor plate, which seems to have been chipped in M. Bosquet’s specimen; this valve and the fixed tergum in all essential respects resemble the same valves in V. Strömia. The surface of the shell is very smooth.
The moveable scutum has its occludent margin considerably arched: the lower articular ridge is broader than the upper ridge, in which respects it resembles the same valve in V. lævigata, but the whole valve is not so broad as in that species. There is no adductor ridge on the under surface. The moveable tergum has its upper articular ridge narrow, and slightly produced into a point on the scutal margin: in this latter respect this species also resembles V. lævigata, but the whole valve is not so broad in proportion to its height.
Cirripedia having a flexible peduncle, provided with muscles: scuta and terga, when present, not furnished with depressor muscles: other valves, when present, not united into an immoveable ring.
This Family has been fully treated of in my former volume, published by the Ray Society,[141] and I should here[Pg 527] only have alluded to its existence, had it not been for the genus Alcippe, which differs in so many important characters from the other members of the Lepadidæ, that formerly I did not even suspect that it could belong to this Family, and therefore deferred its examination. The genus Alcippe was discovered, well described and illustrated, in 1849, by Mr. Hancock; to whose very great kindness I am indebted for permission to dissect and examine his entire stock of this truly remarkable Cirripede. In the classification of the whole class I have not felt so much doubt, as whether I ought to institute a family for the reception of this genus. Alcippe differs from all other Cirripedes (putting on one side for the instant, the males and complemental males of Ibla and Scalpellum) in the very singular fact of being destitute of a rectum and anus;—in the three segments of the thorax, which usually support the second, third, and fourth pairs of cirri, being without any appendages;—in the fifth and sixth pairs of cirri having their inner or posterior rami metamorphosed into very singular roughened cushions or buttons, which apparently serve to triturate the food;—in the caudal appendages being muscular, and being used conjointly with the cirri;—and lastly, in the pupa having a lesser number of segments in its abdomen and caudal appendages than in (as far as I have seen) any other Cirripede. It will be thought that these characters are amply sufficient to justify the placing Alcippe in a separate family, more especially when the close general resemblance in the animal’s body in most of the other members of the Balanidæ, Verrucidæ, and Lepadidæ, is borne in mind. On the other hand, the males and complemental males of Scalpellum and Ibla must indisputably be considered as members of the Lepadidæ; yet the male of Scalpellum vulgare and ornatum has no stomach, anus, or mouth, which is a far more abnormal structure than the absence only of the anus in Alcippe: the cirri, also, in these same males, differ from the ordinary cirripedial type decidedly more than in Alcippe. Again, in the male of Ibla, all the cirri, excepting the fifth and sixth pairs, are aborted, and these two pairs are usually only uniramous; here, then, we have a decided resemblance to Alcippe. Hence, if we might assume that[Pg 528] the female Alcippe had partially assumed characters confined to the males of the other genera, it would assuredly stand amongst the Lepadidæ. Independently of this comparison with the foregoing males, the affinities of Alcippe are so special to several genera amongst the Lepadidæ, that it seems unnatural to force it out of the position which it well occupies between Ibla and Anelasma, and place it in another family by itself: thus, in being bisexual, and in the general character of its very curious males, Alcippe shows an affinity to Ibla and Scalpellum; and to the former of these genera it is related in several particulars, such as in the body being lodged within the peduncle, and in the structure of the larval antennæ, &c.: to Anelasma and Alepas it is allied in the general character, and to a certain extent in the muscles, of the capitulum; Anelasma, also, has all its cirri to a certain degree rudimentary, and Alepas cornuta has the inner rami of the fifth and sixth pairs of cirri,—namely, the very same rami which are so curiously modified in Alcippe,—small, destitute of muscles, and functionless for their proper purpose: to Anelasma and Lithotrya it is allied in the peculiarity of the lower end of the peduncle becoming elongated by growth, and in being imbedded; and to Lithotrya by its powers of excavation and manner of attachment. Now, I believe it generally holds good that when a form is really distinct from another group, its affinities are general, or only in a slight degree special to the members of that group. Nor, indeed, can it be asserted that Alcippe differs much more, somewhat more it certainly does, from the other genera, than does Anelasma, with its more singular mouth, spineless rudimentary cirri, and fimbriated peduncle; and I have never regretted having included this genus amongst the Lepadidæ. Hence, after much consideration, I have resolved to consider Alcippe as one of the Lepadidæ, though so curiously modified,[142] and[Pg 529] having characters confined to the males of the other genera. Perhaps I have been in some degree influenced by the difficulty of finding external characters by which to separate Alcippe as a family from the other Lepadidæ.
[141] The fossil species have been described in a separate Monograph published by the Palæontographical Society. Since its publication, M. Bosquet has produced an excellent memoir, containing descriptions, with the most beautiful illustrations, of several new Cretacean species of Pollicipes and Scalpellum. The memoir is entitled a ‘Monographie des Crustacés Fossiles du Terrain Crétacé, du D. de Limbourg.’
[142] Adrien de Jussieu, in his ‘Memoir on the Malpighiaceæ,’ ‘Archives du Museum,’ tom. 3, p. 86, when speaking of the characters afforded by the degraded flowers, which in certain genera are borne together with ordinary flowers, makes the following observations bearing on the question here discussed, viz., whether or not to include Alcippe amongst the Lepadidæ. “Ces exemples peut-être aideront à comprendre comment à des genres d’une organisation assez compliquée, viennent quelquefois s’en rattacher d’autres d’une organisation beaucoup trop simple en apparence, membres appauvris et dégradés d’une même famille, qui lui appartiennent sans la représenter; comment le type, s’y présente comme effacé, ne conservant plus pour se laisser reconnaître que quelque trait isolé, mais caractéristique, dont la valeur, essentiellement ordinale, peut être ainsi constatée.” Under the point of view, so strongly and admirably insisted on lately by Milne Edwards (‘Annales des Sciences Nat.,’ 3d series, tom. 17), of describing types without regarding whether the different members blend together on their confines, perhaps Alcippe should be raised to the rank of a Family: I feel quite unable to decide how properly to act.
But we shall presently find, when we come to Cryptophialus, that all the above difficulties, great as they are, are greatly enhanced, for Cryptophialus is certainly allied in a very direct and curious manner (in decided opposition to the remarks just made on special affinities) to Alcippe, and yet in all the more important parts of its organisation, and in its metamorphosis, it differs so fundamentally, that I have felt myself obliged to form not merely a Family, but a distinct Order for its reception.
ALCIPPE. Hancock. Annals and Mag. of Nat. Hist., vol. 4, 1849, Pl. 8, 9.
Fem.—Capitulum without valves, with the orifice spinose: peduncle with the basal end added to during growth; its rostral surface depressed and covered by a horny disc: capitulum and peduncle imbedded in a cavity excavated in the shells of molluscs.
Labrum very large, with a row of long hairs on each side: palpi rudimentary: mandible one-toothed: second, third, and fourth cirri absent: fifth and sixth cirri with the posterior ramus represented by a button-like body: caudal appendages four jointed, muscular: anus none.
Males,—several, adhering to the upper end of the horny disc of the female: capitulum naked, transparent, elongated, with a small orifice at the end: peduncle lobed, with the lower end extending far beyond the pupal antennæ: eye, testis, and vesicula seminalis single; probosciformed penis very long: mouth, stomach, thorax, abdomen, and cirri none.
Hab.—North-eastern shores of England, fifteen to twenty fathoms, imbedded in dead shells of Fusus antiquus and Buccinum undatum (A. Hancock); south-eastern shores, off the Eddystone, Lighthouse (C. S. Bate).
I may premise that after the sketch of the leading peculiarities of Alcippe, and after the discussion on its affinities, just given under the Family, I think it would be superfluous to institute a full generic description, separately from the following detailed account of this most anomalous cirripede.
General Appearance.—The whole animal is from .2 to .3 of an inch in length, of a soft texture, colourless or yellowish, and lives concealed in a cavity of its own formation in the shells of certain Gasteropods. This cavity communicates with the water by a narrow fissure-like orifice (Pl. 22, fig. 4), broadest at the posterior end, where the cirri are exserted; narrow, closed, and generally curved at the other (a) end: the two sides of the fissure (b) are commonly bordered by a calcareous inorganic deposit: the walls of the cavity are worn so thin over the peduncle, at the narrow end of the fissure, that the orange-coloured ovaria can generally be seen through the shell of the mollusc, and hence there is here a distinct fan-shaped stain (fig. 3) on the surface. The animal consists of a compressed capitulum, without valves, and of a sort of peduncle depressed on its rostral face, and covered with a broad, oval, thin, horny disc. We must remember that in the Lepadidæ the peduncle does not essentially differ from the capitulum, being only the flexible lower or anterior end of the animal, and is separated from the capitulum only by shape, and generally by the direction of the lines of growth. The disc, when most regular (fig. 1, H), lies in a plane at right angles to the sides of the capitulum, and almost in a line with the orifice leading into the sack; but the peduncle is often very irregular (fig. 2), and the disc comes even to occupy a position[Pg 531] nearly parallel to one or the other side of the capitulum. On the carinal side, the capitulum is generally separated from the peduncle by a rather deep fold (f, in the section fig. 5), but this depends in some degree upon the state of distension of the mass of ovarian cæca. I have given a drawing, fig. 1 (partly taken from Mr. Hancock), of a very regular individual, and of an extremely distorted specimen (fig. 2). The distortion, I believe, is generally caused by the animal, during its excavation, breaking into some old cavity.
External Structure.—The orifice leading into the sack is about one third of the total length of the animal: its edges or lips are thickened, horny, and brownish: at the lower end, exactly where the orifice ends, the lips are formed, from being deeply notched, into two sharp projections (a, figs. 1, 5, 6), unlike anything occurring in any other Cirripede. The external membrane (c, fig. 6) of the lip supports an irregular but nearly straight band of sharp, thick spines of chitine, about 1/1000th of an inch in length, together with a few hairs: at the carinal or upper end of the orifice the spines are largest and most numerous; at the other and lower end, they decrease in size; and on the two projections (a, fig. 6), and on the adjoining parts of the external membrane, they graduate into the small dentated points which cover the whole surface of the animal. The inner tunic of the sack (b), on each side along the upper half of the orifice, is remarkable from having a moderately broad, curved band of short, sharp spines, not quite so thick as those on the external surface, closely adpressed together and pointing upwards, like the javelins of an ancient phalanx, thus probably preventing the ingress of any intruding animal. This band of spines curves at the upper end, conformably with the shape of the orifice. The inner tunic of the sack in this upper part is yellowish, and, what is very unusual, is thicker than the external membrane. A little way down, within the orifice, and more especially in front of an elegant row of hairs on the two sides of the great labrum, there is a band of very fine but stiff hairs (5/1000ths of an inch in length), pointing upwards, and making together with those on the labrum a hedge, barring ingress into the sack.
The external membrane over the whole animal, excepting the horny disc which covers the rostral face of the peduncle, is very thin and transparent; it is periodically and often moulted, as may be inferred from the many old lines of junction round the edges of the horny disc: it is irregularly and pretty thickly (but not so thickly as in fig. 7) studded with star-headed, minute points, from 2 to 5/10,000ths of an inch in diameter, composed of hard chitine, seated on a short footstall, and this on a circular, yellowish, slightly thickened disc of the general investing membrane, appearing like a halo surrounding each little point. These points are directed obliquely upwards. There are none on the horny disc, though particularly numerous close to its margin. Their state varies much: just after a moult, when newly formed, the spines are regularly star-headed, with quite sharp rays, from two to six in number, with some of them occasionally bifid; but these points or rays soon become blunted, and ultimately half the star is worn away, so that the appearance then presented is that of a crescent with a few blunt points on its convex side. At each exuviation, the thickened membrane of the orifice with its strong external spines (the condition of which also varies according to the period elapsed since the last moult), and of course the whole internal tunic of the sack, with its spines and hairs, are all moulted, together with the external membrane and the little star-shaped points. In most specimens a barely distinguishable band or bar of yellowish, slightly thickened membrane, runs from the points (a), at the lower end of the orifice, for some way obliquely downwards; and at the lower end of this bar the weak adductor scutorum muscle (having transverse striæ) is attached. This bar is often strengthened by a prominent external fold of membrane, but yet it is so flexible, and as it is united only to the lower end of the orifice, I can hardly believe that it can, by means of the adductor muscle attached to its opposite extremity, have much power in closing the orifice. I believe that this muscle acts simply in narrowing the whole animal, so as to favour its movement within the cavity in which it is imbedded. Owing to this position of the adductor muscle, and its consequent little power in[Pg 533] closing the orifice, we can understand the necessity for the defence afforded by the bands of spines and hairs on the inner tunic of the sack and on the labrum, which do not occur in other Cirripedes.
Horny disc.—The general shape of the disc, its irregularity and position, have been already described. It never extends, as remarked by Mr. Hancock, to the extreme lower point of the peduncle; upwards it reaches to a little below the lower end of the orifice. It consists of successive layers of membrane, either moderately thick and opaque, or only a little thicker than the general membrane of the body, but never furnished with the little sharp points; it increases in size, in like manner as the calcareous valves of other Cirripedes, the undermost and last formed layer extending beyond the others, with its edge united, till the next exuviation, to the general membrane of the body. The disc is attached, at its upper end, apparently in the usual way, by cement, to the roof of the cavity of the shell in which it is imbedded; but the lower parts of the disc are also slightly and partially attached, chiefly along the lines of growth or exuviation; and this, I suspect, is effected by an inorganic calcareous deposit; anyhow I could not perceive here any cement or cement-ducts. Beyond the circumference of the disc the whole animal lies free in its cavity. The lines of growth in the middle part of the disc are generally obliterated by the decay of the older and outer layers. These lines, though of course ordinarily conformable with the general outline of the disc, are not always so, for the disc sometimes becomes during growth slightly changed in form, and the animal, consequently, slightly changed in position; sometimes either one or the other side or the upper end of the disc is left deserted by the new layers of the growing disc; these being formed on the deserted side of less size or extension, instead of larger size, as they normally should be all round the disc.
The upper end of the disc is always produced into a projection of not regular shape, but generally hollowed out or embayed in front (fig. 1), and almost always hollowed out on the two sides. This projection stands directly over the adductor muscle (b in fig. 5), and on the exterior surface[Pg 534] is generally convex, being concave on the under side for the attachment of several muscles presently to be described. The horny layers are in this part usually thicker than elsewhere. The disc is thus upwardly produced, owing apparently to the fissure which leads into the cavity of the shell of the mollusc becoming, during the process of excavation, considerably longer than is necessary,—that is longer than the orifice leading into the sack; and consequently, for the protection of the imbedded animal, the lower and narrow end of the fissure is closed on its under side by this upward production of the horny disc, formed of layers of membrane of unusual thickness. A deposition, also, of lime, hereafter to be mentioned, gives further protection.
In the bays on each side of the upward production of the horny disc, and likewise a little lower down on its edges, and therefore somewhat protected by lying within the narrow, pointed, lower end of the fissure in the shell of the mollusc, the short-lived Males (Pl. 22, fig. 1, m) are attached often in groups of two, three, or more together.
It may be asked to what part, in other Cirripedes, does the horny disc answer? Not considering the upward prolongation, which has been developed for a special purpose, the disc is irregularly circular,—is added to all round,—serves for the attachment of the whole animal to the supporting surface,—is covered on the under surface by a conformable and parallel mass of ovarian cæca, and the latter by the inner tunic of the sack; therefore in every character, and in its relation to the other parts of the animal, the disc answers to the end of the peduncle, or to the basal cup in Lithotrya, or still more closely to the basis in sessile cirripedes, with the important exception that it lies in a line with the longitudinal axis of the whole animal instead of at right angles to this axis. We know that all ordinary cirripedes become first permanently attached in their pupal state by their antennæ, which are seated on the ventral or rostral surface, near to the anterior end of the body; and that from the young cirripede, after the act of metamorphosis, being turned vertically upwards, and from the extreme anterior, now lower, end of the body not being rapidly developed, the surface cemented down, or the basis,[Pg 535] encroaches almost equally on the dorsal, lateral, and ventral surfaces. But if we were to suppose the extreme anterior point of the body to be rapidly developed, the surface of attachment or basis, without it grew still more rapidly, could not possibly reach the dorsal surface, and would, consequently, be confined to the ventral or rostral surface. I have not seen the young of the ordinary or female Alcippe soon after its metamorphosis, but in the male the development of the extreme anterior end of the body is extraordinarily rapid, and from analogy we may fairly conclude that this is likewise the case with the female. Hence I believe that the horny disc answers to the cemented down, lower end of the peduncle, in other members of the Lepadidæ, and to the basis in the Balanidæ, and that it is confined to the ventral or rostral surface, owing to the anterior or lower end of the body having been rapidly developed. To make all the parts, internal and external, of Alcippe, correspond with those of other cirripedes, the main circular part of the horny disc must be turned up at nearly right angles to its present position (the dorsal or carinal integuments, to the right-hand in fig. 5, being shortened), and then we should have a peduncle, certainly very short and broad, but holding its proper relative position.[143]
[143] In the genus Lithotrya, as long as the animal continues to bore into the rock, the calcareous discs by which it is attached in its cavity, stand, as in Alcippe (Pl. 8, fig. 2, 2 a′, Darwin’s ‘Monograph on the Lepadidæ’), parallel to the longitudinal axis, but as soon as the animal ceases to bore, and the discs become converted into a cup, they occupy a normal position at right angles to the peduncle. According to Reinhardt, these discs, in Lithotrya, are situated on the carinal or dorsal surface of the peduncle, at which statement I now feel considerable surprise, as undoubtedly the pupa must first permanently attach itself by its prehensile antennæ on its ventral or rostral surface. In Anelasma I failed to discover any cement or cement-ducts; but I am now strongly inclined to believe, considering that the extreme lower or anterior end goes on growing, that the surface of attachment will be found to occur, as in Alcippe, on the rostral surface, a little way below the orifice.
Sack and its Muscles.—I have already described the curious phalanx of spines, the long fine hairs, and thickened condition of the inner tunic of the sack along the sides of the orifice. This inner tunic is a reflexion from that enveloping the body of the animal, in the usual manner, as may be seen in the section (Pl. 22, fig. 5). Between the external membrane and the inner tunic of the sack (e), there[Pg 536] is of course the usual double fold of corium, these two folds being united by minute, transverse, ligamentous fibres, branched at the two ends, as in other Lepadidæ. Imbedded in the corium there are numerous, longitudinal, striæ-less muscles, which do not run quite up to the orifice, but to an oblique line beneath it. Externally to these muscles there are, as in the other Lepadidæ, fine transverse muscles, confined to the middle part of the animal, and running from the carinal margin more than half way round both sides. Attached to the upper notched or folded end of the orifice (g, fig. 5, above the upper ends of the longitudinal muscles), there is a fan of rather strong, striæ-less muscles, expanding downwards, with their lower extremities attached to the outer membrane of the capitulum; these muscles apparently serve to open the orifice: there is a somewhat analogous muscle in Lithotrya, but in no other member of the Family: in Cryptophialus, however, there is a closely similar muscle. Owing to the action of these several muscles, the tissues forming the capitulum and peduncle are, according to Mr. Hancock, highly contractile.
I have stated that the under surface of the upper produced end of the horny disc is concave, and serves for the attachment of several muscles. Of these some run to the basal margin of the great labrum, and no doubt, as usual, move the whole mouth; others, as usual, run to the skin between the labrum and the lower end of the orifice,—i. e. in fig. 5, between the lower end of the row of fine hairs (see fig. 11), which shows where the basal margin of the labrum is situated, and the lower side of the point (a), where the orifice terminates: others run obliquely on both sides towards the point of attachment of the small adductor scutorum muscle (b, fig. 5): others, of considerable strength, and these are more peculiar, run and are attached to the lower end of the orifice (a), and serve apparently to draw up the orifice from within the fissure-like cavity, in which it lies lodged: others, again, extend transversely on both sides, close beneath the inner tunic of the sack, a little beyond the line whence the ovigerous fræna or branchiæ arise. These transverse muscles lie within the longitudinal muscles, and therefore are quite different from[Pg 537] the exterior transverse muscles, which are situated more towards the carinal portion of the peduncle and capitulum, and which are common to most Lepadidæ. The internal transverse muscles, and those running to the lower end of the orifice, are peculiar, but we shall hereafter meet with them even more developed in Cryptophialus.
Along the medial carinal line there is, between the two layers of corium, the usual circulatory channel. On each side of this line, on the inside of the sack, there are generally some slight irregular swellings, and sometimes a large extent of the inner surface is irregularly carunculated with little knobs. The sack (e in fig. 5) extends down almost to the basal point of the peduncle (d), more especially when the ovarian cæca are not gorged with ova.
Branchiæ, or ovigerous Fræna.—Within the sack, on each side of the body, rising not far from the ends of the adductor muscle (b), there is a large fillet or fold; the two occupy so exactly the position of the ovigerous fræna that I cannot doubt such is their nature, though, as happens in the case of some species of Pollicipes, they are destitute of their proper glands, and so do not serve for the attachment of the ovigerous lamellæ; this attachment probably is not required, owing to the protected situation which the lamellæ hold in the sack, under the animal’s body, and over the ovarian cæca. From the unusually large size of these so-called fræna, I cannot doubt that they serve as branchiæ, equally well with the plicated folds of membrane, believed to be homologous with the fræna, in the Balanidæ, which have by every one been considered as branchiæ. The fræna are broad and truncated at their upper ends; their margins are sinuous, and their outer surfaces papillose; they run longitudinally down the sack, narrowing as they extend, almost to the basal point of the peduncle, and hence are of considerable length; they are hidden in the section (fig. 5) by the medial, somewhat protuberant mass (c) of ovarian cæca, and partly by the (i) prosoma.
Body.—The body is constructed on the usual type, and indeed does not differ greatly from that of Ibla. The labrum is very large, its lower or basal margin is separated by an unusual space (capable of being contracted or folded)[Pg 538] from the lower end of the orifice of the sack; hence the labrum and whole mouth is placed quite remarkably near the upper (or carinal) end of the orifice. This upper end of the orifice, I may remind the reader, is homologically the posterior end of the general covering or carapace, and all that portion of the whole animal (as the sectional figure, 5, stands) below the lower margin of the labrum, on the rostral or ventral surface, is formed by the three anterior segments of the head. The main part of the body, carrying the mouth, is formed by the great development of that segment of the thorax which bears the first pair of cirri (h), here closely adpressed, as usual, to the sides of the mouth. The lower portion of this segment forms the prosoma (i), and has the characteristic outline, but is not much developed. On each side of the prosoma an oval space of membrane is yellowish and is thickened, and so gives support to this part of the body. The five succeeding thoracic segments, which ought to carry the five succeeding and posterior pairs of cirri, are together of very small size (as in Ibla), in comparison with either the prosoma, or the whole anterior part of the animal. The segment (k) which should have borne the second pair of cirri, is considerably longer than the following segments, and is at the same time less distinct, owing to an oval convex shield of thickened membrane on the sides, not extending the whole length of the segment, thus causing two transverse creases, which, when the thorax is contracted, appear like two additional segments. Had this segment borne cirri, they would have stood, as in Ibla, at a considerable distance from the first pair. The segments (l, m) which should have borne the third and fourth pairs of cirri are like each other, except that the former is rather the longest. The membrane covering all the thorax is surprisingly thin; and at the articulations, which are straight and transverse, is deeply folded, so that the thorax must be highly extensible, to a degree which I have not seen equalled in any Cirripede except in the males of Scalpellum vulgare and ornatum. The thorax is represented as somewhat extended in fig. 5. This part of the thorax is amply furnished with striated and striæ-less muscles for its retraction and protrusion, and for lateral movements. The segment (m) which should[Pg 539] have borne the fourth pair of cirri, at first sight falsely appears like the terminal segment of the thorax: in one monstrous specimen it bore a single cirrus, showing (if there had been any doubt) that it was a true segment. The three terminal pairs of articulated appendages, form together a brush; they consist of the fifth and sixth pairs of cirri and of the caudal appendages: my reasons for considering the last-named organs as of caudal origin will be given hereafter. A moderately careful inspection, especially of the ventral surface, will show that the fifth pair of cirri are borne on a small segment (n, fig. 5, but plainer in fig. 13), which is quite distinct from, but partially concealed by, that which ought to bear (and did bear in the monstrous case) the fourth pair of cirri: this segment is oblique, and cannot be traced distinctly all round the dorsal surface. The segment (o, fig. 13) bearing the sixth pair is much less distinct, and can only be seen by a longitudinal section, or when the cirri are a little separated, but it certainly exists,[144] as is likewise shown by the presence of small apodemes dipping in amongst the muscles, between this and the last segment. The posterior or caudal appendages are closely approximated; they are not separated by any fold from the sixth thoracic segment; but appear as if they were articulated on the dorsal surface of the sixth pair of cirri, in exactly the manner usual in the other Lepadidæ. The segments bearing the fifth and sixth pairs of cirri are highly oblique to the preceding segments, and consequently the cirri, which they support, instead of projecting inwards, lie like a brush in a line with the longitudinal axis of the main part of the thorax. The membrane forming the two small oblique terminal segments of the thorax is strengthened by irregularly shaped plates of thicker and yellowish membrane.
[144] In the middle, the fold is slightly prominent and pointed, and being most finely villose, I for some time looked at this projection as a rudiment of the probosciformed penis.
Mouth.—The mouth is constructed on the strictly normal type of the Family, but is peculiar in every part; it is remarkable from being situated so near the upper (or posterior) end of the capitulum, this being caused by the great[Pg 540] length of the labrum, and of the space of body between the latter and the lower end of the orifice. The labrum is a very singular part of the mouth from its vast size and outline: in fig. 8, we have a front view of the mouth, of which the whole upper pointed part consists of the labrum, and h h is the first pair of cirri; in fig. 11, we have a lateral view of the labrum, with the surrounding thin membrane of the body, a—a, still adhering to its edges; h is the first cirrus on the near side; m the mandible, a little distorted in order to show its tooth, marking the position of the transverse crest of the labrum and of the orifice of the œsophagus; b b is the medial longitudinal ridge of the labrum. In the Balaninæ the labrum forms a mere rim to the back of the mouth, consisting of an inner fold running down the œsophagus, and of an outer fold, both close together: in the Lepadidæ the folds are separated, the outer one being swollen or bullate; and here this structure is carried to even a greater extreme than in Ibla and its allies. The distance between the transverse crest over the œsophagus and the blunt projecting point on the summit of the medial ridge, b b, equals twice the longitudinal diameter of the rest of the mouth. The lower margin on each side of the labrum is produced into two projections (fig. 11), the longer one curling round to a point beneath the jaws, with its extremity imbedded as an apodeme. Another very peculiar character in the labrum, prominently noticed by Mr. Hancock, is caused by a longitudinal row, on each side, of closely approximate, long, very finely pointed hairs, which, as already stated, are fronted on the opposed internal surface of the sack by an irregular band of still finer hairs. The surface of the labrum is partially covered by minute toothed scales, and these, seen on the longitudinal medial ridge, b b, give it a finely denticulated structure. At each end of the transverse crest which overhangs the œsophagus, there is a knob, such as occurs on the labrum of every Cirripede. United to these two knobs, which are formed of thick and yellowish membrane, and springing from the adjoining sides of the mandibles, there are two swellings formed of thin membrane (fig. 8), which occupy the exact position of the palpi, and may be considered as these organs in a rudimentary condition and destitute of bristles.
The mandibles are simpler than in any other Cirripede; they are minute; they consist of an oblong plate, with only one very strong tooth at the upper end: the face towards the labrum is swollen: beneath the upper free part there is a small, sub-triangular piece of thickened membrane, let in and forming part of the general outer surface of the mouth, and representing the large square plate found in other Cirripedes. The maxillæ (fig. 15) are smaller but broader than the mandibles; they have an upper tooth and a smaller lower one, lying not quite in the same plane with the upper one, but nearer the mandibles. The apodeme (fig. 8, 15) is of remarkable length, extending beneath the basal fold of the mouth: it does not arise from the ridge or outer edge of the maxilla, but a little on one side, from the face directed towards the mandible. Between maxillæ and mandibles there is a very singular prominent fold of membrane (fig. 8, 15), which resembles, but probably falsely, the supposed rudimentary palpus attached to the mandible. Altogether the maxillæ differ considerably from the same part in other Cirripedes. In structure they seem adapted to assume the function of mandibles; but they do not stand directly over the œsophagus. The outer maxillæ (fig. 8) appear like a minute, deeply notched lower lip: each consists of a simple, oblong, rounded plate, with a few small bristles at its upper end. The basal fold of the mouth in front, beneath the outer maxillæ is distinct, and runs in a line with the basal articulation of the first pair of cirri. In the rudimentary palpi, minute and little developed outer maxillæ; and in the inner maxillæ, taking the function of the mandibles, the mouth of Alcippe presents some resemblance with that of Anelasma.
Cirri.—These consist of the first, fifth, and sixth pairs: the other pairs are absent, except in one monstrous specimen, in which there was a fourth cirrus quite like the fifth. First pair, fig. 14, these are seated on each side of the mouth in the usual position. They are formed of very thin and flexible membrane. The pedicel, as usual, consists of two segments, the upper one is short and not very distinct; but when viewed on the inner side can be seen to have[Pg 542] the ordinary structure: both segments are destitute of bristles. There are two short rami, being about one third of the length of the pedicel: they are directed either in the line of the pedicel, or more commonly posteriorly, that is towards the other cirri, and therefore in an unusual direction. The anterior ramus is generally rather longer and thinner (as is commonly the case with other Cirripedes) than the posterior ramus; but there is some variation in this respect. On neither ramus is there any trace of the ordinary articulations: both are thickly clothed with fine bristles, which are singular from being thickened in their lower parts, and plumose, like a feather. These cirri have some resemblance, as remarked by Mr. Hancock, to a pair of pincers; but they cannot act as such; they serve, I believe, as brushes. Delicate muscles, transversely striated, enter and are attached within both rami and within both segments of the pedicel, on the usual type, showing that these organs (if there had been any doubt) are truly cirri.
The fifth and sixth pairs of cirri (fig. 13, n′, o′) are almost exactly alike: they are of very small size: each cirrus consists of four segments: the lower or basal segment is broad, with a few minute bristles scattered on its inner surface: the second segment is also broad, but shorter, with a few, generally hooked bristles, in two short irregular rows, in the upper part: these two segments answer to the two segments of the pedicel of ordinary cirri. The third segment is thinner and longer than the second; it bears two or three longitudinal rows of bristles, most of which are neatly hooked at the point; its upper end is surrounded with a circle of bristles. The fourth and terminal segment is short, thin, and simple, with only a few bristles at the apex. These two upper segments are bent a little inwards; they answer to one of the two normal rami of ordinary cirri. The third segment does not stand exactly on the middle of the summit of the second segment,—the posterior corner of the latter being occupied by a very curious, convex, oblong, rather hard (especially in the lower part), protuberant cushion (as called by Mr. Hancock) or button (fig. 9, c′), transversely wrinkled by fine, distinctly crenated ridges. This button presents a considerably dif[Pg 543]ferent appearance according to the point of view, fig. 9, 10: on one of its sides it projects beyond the outline of the second segment, whence it arises; on the other side it is prolonged, as a smooth ridge, on the top of the second segment, embracing to a certain extent the base of the third segment. On the face opposite to that which has been drawn (fig. 9) as most characteristic, it is seen to be somewhat constricted round its base; this constriction, representing, I believe, an articulation. When viewed directly in front (fig. 10) its outline is oval, passing into shield-shaped. Its longitudinal axis is 3/1000ths of an inch in length; but it varies a little in shape and size. I shall presently assign my reasons for believing that these buttons are the posterior or inner rami of the fifth and sixth pairs of cirri in a rudimentary and much modified condition.
Caudal Appendages.—These (fig. 13, p) are placed close together, being articulated between the bases of the sixth pair of cirri, the lines of junction being internally marked by minute apodemes. They consist of four segments, resembling in every respect those forming the cirri, with the important exception that there is not a vestige of the button on the summit of the second segment; the segments are not so thick as those of the cirri, and the terminal segment is smaller.
Muscles and Functions of the Cirri.—For their size, the cirri and caudal appendages have voluntary muscles of remarkable strength, attached within their basal segments, and springing from the dorsal and ventral surfaces of the so-called third and fourth (l, m) thoracic segments. Other muscles, rising from within the basal segment of each limb, run to the second segment, and from that to the third segment. I could not distinctly make out whether any entered the terminal segment. I have seen no other instance of muscles entering the caudal appendages, but as in the pupa they are so furnished, we here have only an embryonic character preserved. I may remark that the fifth and sixth cirri, consisting of two large basal and two thinner terminal segments, is likewise an embryonic character. From the position of the cirri, the four hard protuberant buttons or cushions, tend to oppose each other at a common point;[Pg 544] and the caudal appendages fill up a gap behind, between the cirri of the sixth pair. I at first thought, with Mr. Hancock, that these buttons served to catch the prey; but, reflecting on their convexity and hardness, they appear very badly adapted for this purpose; it would, in fact, be a marvellous feat to secure, in the dark, any moving object between four balls. On the other hand, this very convexity, the hardness, and especially the crenated ridges, and the powerful muscles (which from the first surprised me), are all well explained, if we suppose the prey, being secured by the terminal segments, to be triturated between these four balls: any part which escaped upwards would, moreover, be retained in a sort of cage, formed by the inwardly inflected terminal segments with their hooked spines. This view of the very curious and unparalleled use made of a modified portion, not of the haunch, but of an upper part of the two posterior pairs of thoracic limbs, is in some degree confirmed by finding that Cryptophialus, which has apparently analogous habits, requires its food to be triturated, though in this case it is effected by very different means, namely, by four beautifully toothed discs, with brushes of hairs, developed within the lower end of the œsophagus.
The prey, when caught, would probably at once be carried by the movement of the articulated thorax to the mouth (itself moveable), and being there secured by the mouth in front, the caudal appendages behind, the tips of the cirri above, and the broad pedicels of the first pair on the two sides, it would be triturated by the four crenated buttons, and would then be forced down the œsophagus by the action of the simple jaws. I looked in vain in several specimens for any object within the stomach. I believe, that when the specimens are first taken, all half digested food is ejected by the mouth. Whether we may thus account for the extremely foul condition of the rami of the first cirri in all the many specimens examined by me, I know not; but that these rami, which are thickly clothed with fine plumose hairs, and are furnished with delicate muscles, act as brushes, so as to clean the orifice of the sack, I can hardly doubt.
Homologies.—I have as yet, to a certain extent, assumed[Pg 545] that I have correctly named the different parts; and a few remarks on this head may be desirable, considering the absence of certain cirri, the singular condition of the others, the close general resemblance of the cirri and caudal appendages, and the fact of the latter being furnished with muscles. The only cause for any doubt regarding the thoracic segments is the shield of thick membrane on that segment (k, fig. 5), which ought to have borne the second pair of cirri, causing two transverse wrinkles (not distinguishable, however, on the ventral surface), and sometimes making the segment appear as if it consisted of three segments: if it did consist of three, as there can be no doubt about the nature of the first pair of cirri, (not in a more rudimentary condition than in Anelasma) or about the segment whence this first pair arises, the two terminal oblique segments, with their appendages, would be abdominal instead of thoracic: but this is improbable, inasmuch as the abdomen is unusually little developed in the pupa (as presently to be shown), and more especially from the circumstance of a monstrous cirrus, identical in structure with the two succeeding pairs, having been borne on a segment (m), which, in any case must be considered as thoracic, for it is well known how very rarely thoracic and abdominal limbs resemble each other. I cannot myself feel hardly any doubt on the nature of these three pairs of appendages; for, in the first place, the posterior appendages are articulated on and between the bases of the adjoining pair, exactly as the undoubted caudal appendages are articulated in all other members of the family on the sixth pair of cirri or terminal thoracic appendages. Secondly, we see in the male of the allied genus Ibla, the very same appendages preserved as in Alcippe, namely, the caudal, and the fifth and sixth pairs of cirri, which latter, moreover, are generally uniramous. Thirdly and lastly, in the likewise allied Alepas cornuta, we have the posterior rami of these same fifth and sixth pairs of cirri in a rudimentary condition, and resembling in every respect the caudal appendages. Assuming, then, that the several appendages in Alcippe have been rightly denominated, we have to consider the nature of their segments: in all cirripedes, the pedicels of the cirri[Pg 546] consist of two segments, of which the lower one (as here) is longer than the upper one, and both (as here) considerably thicker than the segments of the rami: in all cirripedial pupæ, likewise, the thick pedicels of the limbs consist of two segments, and each ramus, also (as here), of two segments: now, with these coincidences, and bearing in mind that in Alcippe the two upper segments do not arise from the exact middle of the summit of the second segment, but from rather its anterior side,—bearing, also, in mind the case just cited of Alepas cornuta with the posterior rami of these very same cirri rudimentary,—we may, I think, safely conclude that here in Alcippe the two lower segments form the pedicel; the two upper segments, the anterior ramus; and that the button-like protuberance is the posterior ramus in a modified condition. As the caudal appendages in none of the Lepadidæ, either in the mature state or in the pupa, have two rami, we can satisfactorily understand the absence of any trace of the button-like protuberance on the top of the second segment.[145]
[145] I almost wish I could persuade myself that I had taken an erroneous view of the thoracic segments, and therefore that the three pairs of terminal appendages were all abdominal, for then Alcippe would come into much closer relationship with Cryptophialus; though even in that case it would form a distinct family from it: but I cannot alter my opinion.
Alimentary Canal.—The œsophagus runs down from the mouth, beneath and nearly parallel to the straight row of hairs on the two sides of the labrum: it is surrounded by the usual muscles: at the lower end it bends down, and expanding a little, like a bell, enters the stomach. The stomach is of considerable size and fills the main part of the body, bulging out under the mouth, and prolonged as far as about the middle of that segment (l), which ought to have borne the third pair of cirri; here the stomach terminates in a blunt rounded point. The tissue surrounding the stomach, and keeping it in its proper place, can be traced to the posterior end of the thorax, but there is no rectum or anus. I am prepared to assert positively that this is the case,[146] for I made repeated longitudinal sections of the whole[Pg 547] thorax in two planes, and I subsequently cleaned the outer tissues with boiling potash, and then, when as transparent as a sheet of glass, I examined every part, and certainly there is no rectum (which in every case is formed of chitine, and so is not acted on by potash) nor an anal orifice. Singular as this fact is, it is not so improbable as it at first appears; inasmuch as I have shown, in my former volume, that the Lepadidæ can reject half digested food by their mouth, and secondly, that the final stage of digestion appears to take place in the upper part of the stomach. In the male of this very species, as we shall immediately see, there certainly is no mouth or stomach, and apparently no rectum or anus; so is it likewise with the males of Scalpellum vulgare and ornatum: in Proteolepas, there is a mouth and an œsophagus, but no stomach, rectum, or anus. There are, I believe, no other known instances, in the whole great class of Crustacea, of the absence of an anus.
[146] I may venture to remark that I succeeded in every attempt, which I made, in seeing plainly the œsophagus, and the acoustic and olfactory orifices and sacks, which, according to all analogy, would be of much smaller size, and far more difficult to discover, than the rectum and anus. I may mention that, according to Mr. Newport (‘Annals of Nat. Hist.,’ 1849, p. 277), the larvæ of certain parasitic Hymenoptera have a stomach without any anus. No crustacean, according to Milne Edwards, is destitute of this orifice.
The stomach, in Alcippe, is much corrugated, so as to be deeply pitted; but there are no regular cæca. The enveloping hepatic layer is thick, brownish, pulpy, and formed of pellets of cellular matter, not distinctly arranged in lines as is general; there is the usual delicate muscular layer. The stomach was in every case empty, and I did not notice the separated epithelial coat, so generally found in other cirripedes.
Organs of Sense.—I failed in discovering the eye, which I have no doubt exits, as it is conspicuous in the pupa and in the male. The olfactory pouches are seated rather laterally under the maxillæ, as in Ibla. As in this same genus, the acoustic sack is seated remarkably low down (fig. 5), at a very considerable distance beneath the basal articulation of the first cirrus: the orifice is seated on a slight prominence: the acoustic vesicle, I believe, is sub-cylindrical, with irregular projections. I did not make out anything distinctly on the nervous system.
Female Generative System.—The animal we have thus far described is exclusively female: when a longitudinal[Pg 548] section of the thorax is made, and the stomach removed, it can be most plainly seen that there are no vesiculæ seminales or testes. Mr. Hancock has remarked on the absence of the usual probosciformed penis. The male of Alcippe will be subsequently described in detail. The female organs differ in no respect from those of other members of the family, excepting in so far that the layer or mass formed by the ovarian cæca (c) does not lie transversely to the longitudinal axis of the whole animal, but longitudinally under the horny disc. The ovigerous fræna are largely developed, but serve, as previously stated, as branchiæ, and not for their proper function of giving attachment to the ovigerous lamellæ. The ovigerous lamella is single, and nearly corresponds, in size and shape (as would ensue from the manner of its formation) to the under side of the horny disc. The ova are broadly oval, and rather above 1/100th of an inch in length.
Metamorphoses.—The larva in the first stage has been fully described and figured by Mr. Hancock: it differs in no essential respect from other larvæ of the family. Mr. Hancock overlooked the inferior minute antennæ. With respect to the larvæ in the last stage, or pupa, I obtained several specimens attached to the disc of the female, and which were on the point of being developed into males; and another specimen identical in all respects, but attached independently to the shell of the mollusc, and which, therefore, I have every reason to suppose, would have been developed into a female. In any case these pupæ may be conveniently here described (Pl. 23, fig. 16.) They are .025 of an inch in length; they are of the usual shape, with the anterior end not very blunt and the postero-ventral surface somewhat produced. The whole carapace or shell is very thin and smooth. There are six pairs of thoracic natatory legs, situated far back towards the posterior end of the body; each leg has the usual articulations, and the two rami their usual long but not plumose spines; the presence of the legs deserves notice, considering the rudimentary and modified state of their homologues in the mature animal. The abdomen differs considerably from the same part, as far as I have seen, in other pupæ; it consists[Pg 549] of only a single almost globular (fig. 17, q) segment, instead of three segments; and the two caudal appendages (r) are very long, and are composed each of only a single segment (instead of two), carrying at its tip two short spines. There are two purple eyes, 4/3000ths of an inch in diameter, which, after having been dried and then soaked, could be seen to be compound; they are fixed in the usual manner to two rather short apodemes, which latter have their usual origin. But the pupa has a very unusual appearance owing to the presence of a single dark purple eye, half the diameter of the two larger eyes, situated behind and above the latter, and quite disconnected with the apodemes; this is the eye of the mature animal, which, for some reason, is here developed earlier than usual. The prehensile antennæ are remarkable from being seated very close to the anterior extremity: owing to this, the articulation of the second or main segment with the basal segment, is hardly at all oblique. The whole pupa is of exactly the same length as the pupa of Ibla quadrivalvis, and so are the antennæ, (see p. 286 of my volume on the Lepadidæ), viz., 32/6000th of an inch, but the second segment is narrower, (being only 8/6000ths in breadth in the broadest part), and is longer in proportion, for the disc which forms part of the total length is only 4/6000ths in length, whereas in Ibla it was 8/6000ths; the disc is here hoof-shaped, as in Ibla. The ultimate segment is remarkably short and narrow, (being only 3/20000ths in width, and less than half the size of that in Ibla); it carries (I believe) three terminal spines, and is not notched. Altogether the antennæ more nearly resemble those of Ibla than of any other genus in the family. From the position of the antennæ, and from the length of the second segment, the pupa, when cemented by the disc or third segment, to the supporting surface, adheres, with its posterior end almost vertically upwards. With respect to the young cirripede within the pupa, I could only observe that its anterior end was formed into a blunt point.
Powers of Excavation; Inorganic Deposit of Calcareous Matter; Attachment.—Alcippe, according to Mr. Hancock, attacks only dead shells of the Fusus and Buccinum, and always on their inner sides, especially on the columella. The excavations, in the specimen which I examined, were so[Pg 550] numerous as almost to touch, and sometimes to run into each other, the included animal being thus rendered distorted. The orifices are directed with respect to the shell indifferently upwards or downwards. From the shape and size of the cavity corresponding to that of the included animal, there can be no doubt, as stated by Mr. Hancock, that Alcippe forms its own cavity. That the action is mechanical I think may safely be inferred from the whole outer membrane being studded with minute, star-headed points of hard chitine, which rise from halo-like little discs of thickened membrane, which latter are well adapted to allow the underlying adherent muscular layer to act on the points, and thus on the surrounding shell. Consequently the points generally show signs of severe attrition, but they are periodically and often replaced, at each exuviation, by new and much sharper points. There are no points on the permanently attached layers of the horny disc, but it particularly deserves attention, that the renewable membrane always extends beyond the circumference of the disc, and is there most thickly studded with the points. We have met, in Lithotrya, with a precisely analogous fact in the extension of the periodically moulted membrane of the peduncle, furnished with star-headed points of chitine, and in addition with minute calcareous beads (which, however, seem soon worn away), beyond the calcareous discs, by which this cirripede is attached in its cavity. We need not feel much surprise at points of chitine being hard enough to wear away shell, when we consider what work the jaws of insects, likewise formed of chitine, will effect.
With respect to the first commencement of the excavation, the pupa, owing to the position of its prehensile antennæ, fixes itself with its posterior end almost vertically upwards; and the young cirripede, after its metamorphosis, from the greater length of the ventral integuments formed round the eye-apodemes, must be thrown backwards into nearly the position represented in Pl. 22, fig. 12, b. I have not seen a young female at this early age, but I have traced the development of several males, and have found that the lower end of the peduncle, (i. e. what was the anterior end of the pupa), grows at quite a remarkable rate, so as[Pg 551] very soon to form a great bag extending beyond the attached prehensile antennæ. Now if we suppose an analogous structure in the female or ordinary Alcippe, and the supposition is quite allowable, we shall almost immediately have the anterior or lower end of the young cirripede, just in advance of its antennæ, pressing against the surface of the shell of the mollusc; and if armed with triturating points, as we have every reason to believe it is, it would wear for itself a cavity. The horny disc on the ventral surface of this protuberant anterior end of the young animal will, we may assume, soon become cemented to the near side of the cavity just supposed to have been excavated. And the whole animal, by further slight changes in direction, namely, by working down more and more obliquely, will take, as shown at (c), its final position. As the whole surface of the animal, with the exception of the horny disc, is provided with triturating points, the animal, when once imbedded, can and does increase its cavity at both ends in length, in depth, and all round the edges of the horny disc,—in short, in every direction excepting directly over the horny disc. I believe, as already explained, that the young Alcippe, (b, diagram), first bores obliquely into the shell; and whatever amount of downward extension the horny disc attains before the young cirripede assumes its proper position, with its ventral surface upwards and parallel to the inner surface of the shell of the mollusc, that amount determines the thickness of the plate of shell hereafter to be left unabraded over the horny disc, as the latter continues to extend in circumference. This plate of shell over the horny disc is so thin, that, as mentioned at the commencement, the colour of the ovaria is seen through; and until I reflected on the following considerations, I was much surprised how the instinct of the animal could so neatly guide it not to grind too deeply, and yet to grind till only a very thin plate of shell was left over its horny disc: these considerations are, that whatever thickness was first given to this plate of shell, when the animal was very young and first assumed its ultimate position, that thickness would in most cases be always retained, owing to the flatness of the disc, and to the membrane armed with triturating points protruding very slightly be[Pg 552]yond and above the horny disc, only just enough to wear away the surrounding shell to the thickness necessary to allow of the formation of each new zone of disc; as the disc itself is not armed, it subsequently has no power of wearing away the plate of shell above it. Thus the horny disc, besides giving support and attachment to the peduncle, is of this peculiar service that it seems to guide, (somewhat like the wood-part in a plane), the rasping powers of the lower extreme margin of the peduncle.
I may here observe that certain radiating and often punctured lines, mentioned and figured by Mr. Hancock, which help to render the thin plate of shell over the peduncle conspicuous (fig. 3), are formed by the burrows of an excessively minute annelid, the punctures being apparently the exit orifices: I imagine that these annelids find it difficult to commence their burrows on the smooth surface of the shell, and that they congregate at these particular spots and thence burrow in radiating lines, owing to their having taken advantage of the little cliff-like edges, at the narrow and disused ends of the fissures leading into the cavities occupied by the Alcippe, where alone they would not be disturbed by the action of the cirri, when first they commenced making their little burrows in the shell.
The fissure leading into the cavity is required to be broad at the posterior end, in order that the cirri may be there freely exserted out of the sack; and narrow in other parts, to prevent, as it would appear, anything injurious getting in between the animal’s body and the cavity in the shell of the mollusc. As the fissure is increased in length by attrition at the broad posterior end, which end during growth becomes broader and broader, the lower part of the fissure has to be narrowed, and this is effected in a very singular manner, namely, by advantage being taken of the strong tendency, which triturated shell with animal matter, has to set into a solid shelly mass, although constantly agitated.[147] Mr.[Pg 553] Hancock noticed this edging of hard shelly matter, and naturally thought it was a secretion. Lines of deposition (Pl. 22, fig. 4, b), parallel to the edges of the furrow can often be perceived in it: its thickness and extension vary much: I have seen it on one side alone of the orifice: it is, of course, never found at the broad end where the process of enlargement goes on. The peculiar worn surface with which it irregularly thins away downwards, on the sides of the cavity, made me (together with the apparent impossibility of such a secretion proceeding from an animal wholly invested by a chitine membrane) suspect it to be inorganic; and this view is certainly correct, for when a fragment is dissolved in acid, a considerable residuum is left of bits of membrane, rubbish, and, in one instance, even of the remnants of a foreign animal, apparently an annelid. We have here all the circumstances favorable for inorganic deposits of this nature, namely, finely triturated shell and chitine or animal matter, produced by the excavation of the chamber, sea-water, and movement.
[147] I have given some remarkable cases in my volume on ‘Volcanic Islands,’ (p. 49), in which limestone, having almost the hardness and specific gravity of marble, has been thus deposited. Almost every coral-reef offers similar examples. The curious substance described by Mr. Horner and Sir David Brewster, (‘Philosoph. Transact.,’ 1836, p. 65), which is formed during the manufactory of cloth, offers another example of the strong tendency which lime and animal matter have to unite. Lately, Dr. Horsford, in ‘Silliman’s North American Journal,’ Jan. 1853, has discussed the chemical theory in an analogous case on the coast of Florida; he attributes the aggregation to the formation of a hydrate of lime through the action of the animal matter. Mr. G. B. Sowerby, Junr., has described a case very analogous to that of Alcippe, (‘Proceedings of Zoolog. Soc., Mollusca,’ Pl. 5, fig. 4, p. 162, 1850), namely, that of Pholas calva, in which a tube is formed of inorganic calcareous matter, serving to narrow the entrance.
From the manner of growth of the animal, the fissure leading into the cavity in the shell becomes much longer than the orifice leading into the sack, and to prevent the body being unnecessarily exposed, the upward projection of the disc, already described, is formed under the narrow and disused end of the fissure; moreover, the two rims of the inorganic calcareous deposit sometimes here approach so closely, as almost or actually to touch each other; and between them, as remarked by Mr. Hancock, there is usually a little accumulation of grains of sand. This narrow end of the fissure is generally curled either to the right or left hand; and I can only account for this fact by supposing that, whilst the cirripede is young, and has not a[Pg 554] large horny disc attached to the cavity, it cannot keep its body straight during the long-continued boring process.
The animal is attached by its horny disc to the thin shelly roof over the peduncle, and likewise to the under side of the narrow end of the fissure, but is elsewhere quite free. I carefully examined the disc in many specimens, but could not see any cement-ducts: I believe I saw layers of cement at the upper end of the disc, but it is not easy to discriminate between this substance and the yellowish, somewhat disintegrated, layers of the horny disc. The pupa certainly becomes attached by ordinary cement, so that the attachment in early life, at least, is normal. In some full-grown specimens, I found the lower parts of the horny disc attached, along the edges of the layers, to the roof of shell; and as I looked here in vain with the highest powers for cement-ducts, or for cement, it appears to me probable that the rough edges of these layers were united to the roof by a thin layer of the inorganic calcareous deposit. The animal, from its very protected situation, certainly requires to be less firmly cemented than other cirripedes; and even in Lithotrya, which is less deeply imbedded than Alcippe, the cementing apparatus was feebly developed. From the length of the pupal antennæ, cemented by their terminal segments, the position of the young cirripede (Pl. 22, fig. 12) can be changed to a considerable extent, like a ship swinging at her moorings, but in order to assume its final position, the animal must, I think, travel like Lithotrya, but to a much less extent, by a short succession of overlapping horny discs,—the old discs being partially deserted, each new one extending beyond the last-formed one: even in the case of the mature animal, we have seen that, under certain circumstances, it changes, to a certain extent, its position; portions of the old disc being deserted and attached to the roof of a deserted portion of the cavity.
Affinities.—In the preliminary remarks under the Family, I have discussed this subject almost sufficiently: I will here only remark, that the genus, though so abnormal, yet stands naturally between Ibla and Anelasma, having clear affinities, on the one side, through and beyond Anelasma to Alepas; and on the other side, beyond Ibla to Scalpellum,[Pg 555] and so to Lithotrya. Moreover, it is very distinctly related to Cryptophialus in the succeeding Order.
On every specimen of the female Alcippe, which I carefully examined, I found some minute parasites (or epizoons) attached to the lateral edges of the upper part of the horny disc, and therefore lying within the narrow end of the fissure leading into the chamber excavated in the shell of the Buccinum. Although having had some experience in the very anomalous forms which male cirripedes assume, yet when I first casually inspected these parasites under a weak lens, from their transparency, their elongated and lobed body, including an internal folded up organ, I actually threw them away, thinking that they were probably Bryozoa. Subsequently, a more careful inspection immediately showed the cemented prehensile antennæ, and their cirripedial nature was demonstrated. I soon found specimens with the perfect still adherent exuviæ of the locomotive pupa, undistinguishable from the pupa already described as probably belonging to the female Alcippe. But as this latter fact, may perhaps be doubted, I must show that there is other evidence sufficient to prove that these cirripedial parasites are the males of the female Alcippe. Of the females, I inspected many specimens, and all certainly were without external male organs; and in the four or five specimens which I rigidly examined, there were no testes or vesiculæ seminales, the latter being in all hermaphrodite and male cirripedes so conspicuous. On the other hand, I examined at least thirty specimens of the parasite, and they were all exclusively males, for all had a probosciformed penis, and the greater number had their vesiculæ seminales filled with spermatozoa, and hence were ready to perform the act of impregnation, but undoubtedly they contained no ova. It would, then, be very strange, if these two cirripedes of opposite sexes, thus attached together, were not sexually related. Wonderfully different as the parasite is from the female Alcippe, yet, in one very important character it is related to Alcippe, and to[Pg 556] no other member of the Family, namely, in the sack extending down to the extreme lower point of the peduncle; the male organs, I may add, occupying an analogous position with the peculiar position of the female organs in Alcippe. The lateral lobes of the peduncle in the parasite seem to represent the sides of the broad depressed peduncle in Alcippe; and in both the peduncle grows at its lower end—a very rare circumstance—observed only in two genera in this Family, namely, in Anelasma, and in a slight degree in Lithotrya. Besides these points of resemblance between Alcippe and its parasite, which are striking, considering their external utter dissemblance, the affinities of both point, judging from certain small characters, in the same direction, namely, towards Ibla and Alepas. Finally, then, I think, we may confidently admit that this parasite or epizoon is the male of the female Alcippe: indeed, considering the facts given in my former volume, on Ibla and Scalpellum, I have, perhaps, here discussed the question at unnecessary length.
The males are generally attached, as already stated, to the two hollowed out sides of the upward prolongation of the horny disc; they adhere by means of little patches of cement, proceeding from the terminal segments of their antennæ, to the overlapping edges of the few later-formed zones of the disc; hence, they lie protected, within the narrow end and a little under the edges of the fissure leading into the cavity in which the female is lodged. In some specimens, however, the males are attached rather lower down on the disc, and are not confined exclusively to its upper margin, so that they live fairly under the roof of shell which covers the main part of the disc: but they are never attached very low down, so as to lie far from the lower end of the orifice leading into the sack of the female. I have two or three times seen as many as three males on each side, but sometimes there is only one on each side, or none on one side. A large distorted specimen actually had twelve males, and two pupæ on the point of undergoing their final metamorphosis, all fourteen attached on one side, and all evidently must have been alive together! Another specimen had nearly the same number, a few on one side, and the rest on the other side.
The male immediately, after the exuviation of the pupal carapace, 25/1000th of an inch in length, is only 23/1000th of an inch long, but ultimately it becomes, chiefly from the growth of the lower end of the peduncle, nearly twice this length; for the largest specimen which I have seen, that figured, was 45/1000th of an inch long (i. e. under 1/20th of an inch), and 1/100th of an inch in breadth across the peduncle, beneath the lateral lobes. The whole external membrane of the animal (as well as the internal membrane of the sack), is very thin, quite structureless, and as transparent as glass; hence, even the spermatozoa, within the vesicula seminalis, can be seen from the outside. The whole structure of the animal is very simple. The ventral surface can be at once recognised by the attachment of the antennæ (fig. 19, a), and these organs mark the point which was the anterior end of the male, just at the period of its metamorphosis, and before the lower end of the peduncle had grown. These antennæ have already been fully described; they are conspicuous from being composed of membrane, rather thicker than that investing the body of the male, and which external membrane can be traced entering these organs, and appearing like cement-ducts; but within these tubular prolongations of the outer membrane, I could obscurely see the real cement-ducts.
The part answering to the capitulum is much flattened and elongated; it widens but little from the upper to the lower end, where it blends with the carinal or dorsal surface (the under surface in fig. 19) of the lobed peduncle. At the upper end there is a small orifice, and close to this, on the ventral or rostral side, there is a thin, apparently double projection (i, fig. 19) or flap of membrane, one flap lying exactly over the other. The whole length of this capitulum probably corresponds with that small portion of the capitulum in the female, between the upward prolongation of the horny disc and the lower end of the orifice; and the two broad flattened projections in the male, probably answer to the two sharp narrow points (a, fig. 1, Pl. 22) in the female. The peduncle has two lateral lobes (h, g, fig. 19), and, whilst young, what may be called a third and medial lobe, but this soon increases largely by growth, and forms the main part of the peduncle. The lateral lobes are intimately connected with the ventral surface; they tend to lie in a plane, at right angles to the[Pg 558] compressed capitulum, but owing to the excessive thinness and flexibility of the whole external membrane, it is difficult to ascertain the relative position of the different parts. Moreover, owing to the pupa being so much flattened, these lobes are necessarily formed folded up; and, I believe, it depends on the position, with respect to surrounding objects, which the male ultimately holds, whether the lobes ever assume, their apparently normal position, in a plane at right angles to the sides of the pupa; owing, also, to the form of the pupa, the two lobes seem generally to be actually formed of unequal sizes, that formed in the dorsal region of the pupa being the largest. I believe that these lobes correspond with the lateral margins of the upper end of the peduncle of the female, which margins project laterally beyond the sides of the capitulum. The lower lobe, or end of the peduncle, is depressed in the same plane with the lobes; it is of variable length; when first formed it hardly extends beyond the basal articulation of the prehensile antennæ. Commonly it does not lie quite in a straight line of the capitulum; and I have seen specimens in which it stood at nearly right angles to the capitulum and to what was the ventral surface of the pupa; this irregularity in the relative position and sizes of the different parts of the peduncle, no doubt, to a considerable extent, depends on the form of surface to which the male becomes attached, just in the same way as we have seen that the peduncle of the female becomes altered in shape during the excavation of the chamber in which it is lodged.
I feel some difficulty on one point: in the pupa the single eye of the future male can be clearly distinguished, and it lies some way from the anterior end of the body; but in two males, which certainly had just moulted, and in which none of the internal organs were as yet developed, the eye lay close to the anterior end, directly over the basal articulation of the antennæ. I suspect this is somehow caused by the great change of form which supervenes, during the metamorphosis, at this anterior end of the body; the extremely compressed body of the pupa having to become depressed and lobed in the young male. I have given a figure of a young male, just as it appeared (Pl. 23, fig. 18), somewhat distorted from lying on a flat surface; c, being the eye.
The sack extends, in a very remarkable manner, down to[Pg 559] the lower end of the peduncle, the whole inside of the animal being thus freely open to the water. In the upper part, the sack forms a mere narrow tube; it does not appear to have been formed in the same manner as in all other cirripedes, namely, surrounding the thorax and natatory legs of the pupa, but in an abnormal position, along the dorsal surface, above the sack and thorax of the pupa: a transparent line, where the new narrow sack is in process of formation, is the first indication of the coming metamorphosis. The sack in the capitulum of the male is not central, but lies near the dorsal surface; the ventral interspace, between the outside and the sack, is occupied by oblique fibres (l, fig. 19), which may be striæ-less muscles, but I suspect are ligamentous fibres, giving support to the whole projecting capitulum. These fibres enter a little way within the lobed peduncle; they are probably homologous with the strong muscles, which run from beneath the upper end of the horny disc of the female to the lower end of the orifice leading into the sack. Round the lobed peduncle,[148] there are two bands (e, f) of thin muscular fasciæ, slightly oblique to each other, and attached at the ends to the outer membrane; they are evidently homologous with the external transverse muscles, which are best developed round the same part in the female. Some of these muscles present a singular chain-like appearance, from being strangled at intervals: they act probably in aiding the long probosciformed penis to protrude itself out of the sack. I could not detect any longitudinal muscles, and the lower part of the peduncle seems destitute of muscles of any kind.
[148] I believe I saw in one specimen, most delicate transverse muscular fibres round the lower part of the elongated capitulum.
The internal structure of the animal is very simple. Within the lower end of the peduncle there is a dark purple eye (c), under the 1/1000th of an inch in diameter, a testis (d) and a (b) vesicula seminalis. These organs falsely appear as if suspended in the middle of the peduncle, but they are really attached, I believe within a separate partition, to the ventral surface, occupying the same position as the mass of ovarian cæca in the female. The eye lies on the line of junction between the testis and the vesicula seminalis, and[Pg 560] on their ventral side. The testis is rounded, and consists of a mass of cells, on an average 1/5000th of an inch in diameter. The vesicula seminalis varies extremely in condition, being either a mere rather broad vessel, enlarged where it joins the testis, or a bag fully as large as the testis itself, and distended with spermatozoa, all arranged parallel to its longer axis. There was an evident relation between the size of the vesicula seminalis and that of the testis, the number of the cells in the latter decreasing as the mass of the spermatozoa increased: there was also an evident relation between the age of the male and the state of these organs; younger and more opaque individuals, having their testes of large size; and older specimens, with the lower end of the peduncle arrived at its full dimensions, having the vesicula distended. Some few old specimens had evidently discharged their spermatozoa. By dissection I more than once distinctly traced the vesicula seminalis entering the broad lower end of the penis. The membrane, forming the vesicula, is ringed, and I presume is, as in other cirripedes, contractile, so as to expel the spermatozoa. The probosciformed penis (m) is of extraordinary length: it is plainly ringed, or rather articulated, in this respect resembling that organ in Ibla and Alepas; it tapers gradually, and terminates (as usual) with a brush of fine bristles; it is furnished with delicate voluntary muscles, arising from the body round its basis, and extending no doubt up to the apex, but too fine to be traced all the way. Its broad lower end is attached in a slight depression, on the ventral side of the sack, a little above the point of attachment of the pupal antennæ. According to all analogy, the spot whence the penis springs must be considered as representing the thorax and abdomen; and the outer membrane of the penis is here, as on this view it should be, reflexed and is continuous with that lining the sack. Ordinarily the penis lies coiled up in complicated folds, appearing like a large intestinal worm, and fills the lobed part of the peduncle, which apparently serves for no other purpose than its reception. In one case in which I dissected out the penis, I found it in its contracted state; 41/1000th of an inch in length, equal to that of the entire capitulum and peduncle; in a specimen, in which the penis had been naturally exserted, the part which[Pg 561] protruded (m) was by itself rather longer than the whole animal; and as this specimen had been placed in spirits of wine, the organ no doubt was contracted; hence I think it probable that the probosciformed penis, when fully stretched out, would equal twice the length of the entire animal.
There must be a nervous system; and there must likewise be a gland (homologous with the ovaria) for secreting the cement; but I could not distinguish parts so small. Certainly there is no mouth, or stomach, or thorax, or limbs of any kind, or abdomen.
It is obvious that these males must be very short-lived: they perform their masculine functions and then perish. We have seen, however, that after the act of metamorphosis they do grow a little, and I have reason to suspect that this is effected, as with other Cirripedes, by moulting. The growth must be absolutely dependent on the store of nutriment laid up within the pupa. The young male, immediately after the exuviation of the integuments, thorax, natatory legs, abdomen, and eyes of the pupa, consists of a pulpy cellular mass, without any internal organs as yet formed.
Judging from the different sizes of the females which included perfectly developed ova, I infer that they must breed more than once during their lives; and therefore, that successive sets of males, as in the genus Scalpellum, must become attached to them. I was not, however, able to discover the prehensile antennæ or other remains of the old males adherent to the females; a circumstance which I presume is accounted for by their attachment being weak. Considering the very small size of the male, it is not surprising that so many,—in one case fourteen,—are required to impregnate the numerous ova of a single female. How the males know the proper period when the ova, lying in a sheet at the very base of the sack of the female, are ready for impregnation, I cannot say, without it be that they perceive the moulting of the external membrane, close to the edge of which they are attached; for this moulting would indicate the period when the ovigerous lamella came to the surface of the sack, and the ova would then be soon ready for impregnation. From the position in which the males are[Pg 562] attached, and from the extraordinary length of the probosciformed penis, capable of voluntary movements, I have no doubt the males can insert the tip of this organ within the lower edge of the orifice of the sack, and there discharge the spermatozoa, which, by their own movements, must pass down the sides of the sack of the female till they reach their proper destination. The position of the males, with respect to the female’s body, is almost exactly the same as that occupied by the complemental males of Scalpellum Peronii and villosum; the lower and narrow end of the fissure, worn in the gasteropod shell, here affording that protection to the males, which the edges of the opposed scuta afford to the complemental males of the above two species of Scalpellum. We cannot doubt that these latter males aid in the impregnation of the ova of the hermaphrodites, but they are not furnished with a very long penis, probably for the very reason that they are complemental males, and therefore not so absolutely necessary for the impregnation of the ova as are the males of Alcippe.
I have, in my former volume, expressed my astonishment at the extent to which abortion had been carried in the male Ibla; but it has been carried much further in the male Alcippe. In Ibla, the thorax is reduced to a mere flap, and only two pairs of cirri exist in a most useless and rudimentary state, but there is a well organised mouth, stomach, and anus. In the males of Scalpellum vulgare, ornatum, and rutilum there is no mouth or stomach, but there is a thorax with four pairs of minute, modified cirri, and a large abdominal lobe. Here, in the male Alcippe, all these negatives are united, we have no mouth, no stomach, no thorax, no cirri, no abdomen! The archetype crustacean consists of twenty-one segments; of these the seventeen anterior segments can be clearly made out in the archetype Cirripede: now, in the male Alcippe, the first three segments are largely developed, forming all that is externally visible, but the remaining fourteen segments are absolutely aborted, but in idea may be considered as forming the membranous depression whence the probosciformed penis springs; for this organ normally arises at the extremity of the seventeenth segment. To show the wonderful[Pg 563] diversity of nature, even in the same sub-class, I may be permitted to remark, that whilst in Alcippe only the three anterior segments are developed, the fourteen succeeding segments being rudimentary, in Proteolepas (hereafter to be described) these fourteen segments are all largely developed, whilst the three anterior segments are quite aborted, being represented only by a thin envelope to the two threads by which this Cirripede is attached to the supporting object.[149]
Cirripedia, having a flask-shaped carapace; body consisting of one cephalic, seven thoracic, and three abdominal segments; the latter bearing three pairs of cirri; the thoracic segments without limbs; mouth with the labrum greatly produced, and capable of independent movements; œsophagus armed with teeth at its lower end: larva, firstly egg-like, without external limbs or an eye; lastly binocular, without thoracic legs, but with abdominal appendages.
I feel compelled to form an Order for the one genus and species, namely, Cryptophialus minutus, to be here described. We must, I conceive, attribute much greater value, in classification, to internal parts and organs, at least where such are not known to vary, than to external structure. Now in Cryptophialus, the body consists of eight segments, of which the first two are not developed in any cirripede hitherto described. Of the eight, the seven posterior or thoracic segments are quite free, or detached from the carapace, and do not bear any appendages; whereas in all the foregoing cirripedes of the order Thoracica there are (at least in the normal sex) six pairs of cirri; Alcippe alone must be excepted, in which there are only two pairs. Again, in the Thoracica there are no abdominal appendages, excepting the terminal or caudal, whereas in Cryptophialus the abdomen bears three pairs of biramous cirri. In the crest of[Pg 564] the labrum, being produced into a special, lancet-formed organ, articulated at its base and capable of movement, and in the palpi projecting straight upwards, we have a great difference from all other cirripedes; and these organs, we have every reason to believe, possess a high classificatory importance. The œsophagus in Cryptophialus, where it enters the stomach, is armed with teeth and hairs, moved by muscles, forming a beautiful structure, of which we have not a trace in any other cirripede. Lastly, and perhaps most importantly of all, the metamorphosis is different; for the early larval stages are passed under an egg-like condition within the sack of the parent; and the pupa differs from the pupæ of all other cirripedes, in not having natatory thoracic limbs, and is therefore only able to crawl about by the aid of its great prehensile antennæ.
Thus far the evidence is decisive in favour of Cryptophialus being placed in a separate Order; but if we were to trust to the characters derived from the external covering or carapace,—and such characters are of high importance, as we may safely infer from the natural arrangement of the foregoing families which depends on the structure of the carapace,—we should place Cryptophialus close to Alcippe amongst the Lepadidæ. These genera agree in their burrowing habits,—in their attachment by a horny rostral disc,—in the external membrane being covered with triturating points,—in the spinose and notched orifice, with an external lateral bar on each side, and in the inner tunic of the sack being protected by hairs and spines. They agree to a considerable extent in shape, and in the peculiar arrangement of the muscles of the whole external covering of the animal: they agree, also, in their manner of growth, and in the sack extending down to their basal extremity. Some of these resemblances may possibly be analogical, and due to similarity of habits, and not to affinity; and we must attribute to mere similarity in function, a certain amount of resemblance in their labrums, for this part is essentially different in the two genera; and to the same cause, the resemblance between the brush formed by the two pairs of thoracic cirri and caudal appendages at the end of part of the thorax in Alcippe, with the three pairs of abdominal cirri at the end of the[Pg 565] whole thorax in Cryptophialus. I allude to this latter resemblance, for it was owing to it, and to the similarity in the habits of Cryptophialus and Alcippe, that I stated, in the introduction to my former volume on the Lepadidæ, that the two genera would probably fall into the same order. In the structure of all the parts of the mouth and of the cirri, in the digestive organs and in the metamorphoses, Cryptophialus is not more closely related to Alcippe than to any other genus whatever amongst the Lepadidæ.
Nevertheless I am confirmed in the view that the external resemblances between these two genera are due to some real affinity, and are not merely analogical, by a very remarkable fact,—namely, that Alcippe and Cryptophialus are both bisexual, and have males, several in number, attached exactly in the same position, and which males are so closely similar that, considered by themselves, they might absolutely be almost classed as species of the same genus! For they agree in the absence of all internal organs and parts, excepting the single testis, vesicula seminalis, and immensely long probosciformed penis; and they agree, also in manner of growth, in the arrangement of the muscles, and even in shape. The whole case seems to me very singular, and, as far as my knowledge extends, unique: we have two animals, of which the females, if classed by their external parts (homologically consisting of the three anterior segments of the head), would be placed alongside each other in the same family; but when classed by the whole rest of their organisation, certainly must be ranked in distinct orders;[150] yet the males of these very same animals might almost stand in the same genus. If it be asked[Pg 566] why the position of Cryptophialus in the system should not be determined by the male, instead of by the female, the answer would be that the male is here abnormal and rudimentary in its whole structure; and I believe systematists are agreed that less perfect parts (and therefore a less perfect whole) offer less valuable characters than the more perfect parts or whole. We see this conclusion plainly verified in the case of the hermaphrodite Scalpellum vulgare and Ibla quadrivalvis, for there can be no doubt where these species should be arranged, yet if we attempted to place them by their complemental males, we should utterly fail: exactly in the same manner, if Ibla Cumingii and Scalpellum ornatum were ranked by their males, they would be quite misplaced. So again, if we were to attempt to class the six species of Scalpellum by their males and complemental males, undoubtedly the first three and last three species of the genus would have to stand in distinct orders! Hence we may reject the males as a foundation for classification, but no doubt they serve to show that the resemblances in the carapaces of Alcippe and Cryptophialus, are not merely analogical or functional, but evince a true affinity, though these genera differ so greatly, in mouth, body, œsophagus, cirri, and especially in their metamorphoses.
[150] M. Milne Edwards would, perhaps, in accordance with the profound views lately propounded by him on classification, consider Cryptophialus as an extremely modified, and, to a certain extent, degraded member or satellite of the type of the Lepadidæ; but I do not myself feel able to draw a line of distinction between the being a very abnormal member of one group, and belonging to a distinct group. I may add that I have several times tried to persuade myself, with no success, into the belief that I have somehow misunderstood the homologies of the thoracic segments and cirri of Alcippe and Cryptophialus; for if this were so, the two genera could be brought into much closer relationship; but with any conceivable amount of error on my part, there remains the great difference in the metamorphosis, not to mention the palpable differences in the cirri, the parts of the mouth, and in the whole course of the alimentary canal.
Hab.—Chonos Archipelago, Southern Chile; imbedded in the Concholepas Peruviana.
General Appearance.[151]—This, the smallest known cirripede, is flask-shaped and compressed, with a small orifice, on a more or less produced neck, placed at one corner: one of the narrow sides of this carapace is somewhat flattened or[Pg 567] depressed, with its superior edge prolonged a little upwards; by this side, the animal is attached to the cavity in the shell, within which it is lodged. There is here no distinction between a peduncle and capitulum, that is between the lower or anterior, and the upper or posterior end of the animal, as seen externally. The small orifice is toothed and hairy: it is coloured purple, as is likewise the projecting labrum; the rest of the animal being tinted only by the muscles and internal parts seen through the outer integuments. The largest specimen (fig. 2) which I have measured did not quite attain the length of one tenth of an inch. This cirripede inhabits, in vast numbers, the shells of the living Concholepas Peruviana, amongst the Chonos islands; the whole outside of the shell being sometimes completely drilled by its cavities, almost touching each other, as happens in the case of Alcippe with the shells of Buccinum. The oval aperture leading into the shell-cavity, in full-sized specimens, is between (2-3)/100 of an inch in length: it is generally surrounded by a narrow, internal, calcareous rim, which apparently has the same inorganic origin, as in Alcippe. The toothed orifice of the carapace leading into the sack, fills up the orifice of the shell-cavity; but it can be voluntarily withdrawn a little: when opened, and the animal is in action, the lancet-formed, moveable crest of the labrum, and the abdominal cirri, are exserted.
[151] I am greatly indebted to Dr. Hooker, for having several years ago, when I examined this my first cirripede, aided me in many ways, and shown me how to dissect the more difficult parts, and for having made for me several very correct drawings, which, with some subsequent alterations, are now engraved.
Integuments.—The external membrane is colourless, thin, but strong; it is studded with minute bifid, trifid, and quadrifid points of hard chitine, which are the agents of excavation: they are directed upwards, except towards the lower end, where they are directed from the disc or surface of attachment. These points beneath the orifice, and on each side close along the lateral bar, are larger than elsewhere. There are no points on the disc or surface of attachment, which is formed of somewhat thickened, yellowish membrane, and is not moulted, like the rest of the external membrane, but is formed of successive layers extending beyond each other; the lines of growth, however, being obscure, and only occasionally distinguishable. The disc is oval, not extending to the lower end of the animal, and with the upper edge thinning out and produced upwards (fig. 1). The animal, during[Pg 568] its growth, moves a little downwards in its cavity, by means of the new layers of the attached disc being formed, not symmetrically with respect to the old layers, but beyond them or at a greater depth in the shell; hence when the animal is removed quite perfect, by the Concholepas being dissolved in acid, the upper and deserted margin of the disc or surface of attachment generally projects as a free edge, but in a tattered and worn condition. In full grown specimens, which have ceased burrowing downwards, nearly the whole disc, though occupying the same position relatively to the animal, becomes in fact deserted, and is lined by membrane continuous with, and like that, investing the rest of the body, but furnished only with simple blunt points, instead of with the sharp bifid and trifid triturating points.
The orifice is formed on each side by a toothed rim of hard chitine (fig. 3, 4), which can be opened and shut, owing to its being flexible at the rostral end, and folded inwards at the carinal or posterior end. The teeth vary in arrangement and sharpness: generally they form, taking a rim on one side (fig. 3), two prominences or groups of points at the rostral or anterior end, always separated by a broad notch, from the bottom of which the lateral bar extends downwards, from the posterior, larger and less regularly toothed half of the rim. Of these teeth the larger ones project nearly straight up, and the smaller and lower teeth outwards, graduating into the smaller teeth, just below the rim, which again graduate into the minute points, studded over the whole surface. These outer teeth probably serve to prevent any creature crawling into the cavity, between the shell and animal. Scattered bristles rise from all over the rim. The lateral bar, above alluded to, consists of the general membrane of the body, thickened, hardened, rendered elastic, coloured yellow, and apparently formed into a fold: where attached to the under side of the rim, at the above-mentioned notch, the bar is thinner and more flexible than elsewhere: it runs half way down the animal, first straight and then curved towards and closely approaching the disc or surface of attachment. At the lower end, the bar, or more strictly the thickened membranous margin of the bar (b′,[Pg 569] fig. 3), expands into an oblong, slightly rigid plate, studded with from three or four to ten or twelve points, which have their ends expanded and truncated, or even slightly bifid. The extremity of this plate projects freely from the general surface of the body. We shall afterwards refer to the use of this extremely peculiar bar and plate.
The two rims forming the orifice cannot be quite closed; but ingress by any foreign object into the sack is beautifully prevented by an internal membranous lip on each side (d′, fig. 3, 4), and by a third inwardly folded lip (d′) at the posterior and broad end of the orifice. These three lips can be brought together, and form a valve. The lateral lips are very narrow at the mouth or rostral end of the orifice, where the hairy lancet-shaped crest of the labrum closes the orifice, and largely expand towards the posterior end: they are produced from the inner tunic of the sack: they appear formed of the finest hairs, placed parallel and approximate, but when examined under the highest powers, these hairs (for they still appear such) are found to be united by delicate membrane, which has its extreme edge fimbriated. The third, or posterior and inwardly folded lip, differs in being composed of much coarser, flattened hairs, which are united towards their bases, and are free at their extremities, where they are serrated or coarsely plumose on both sides.
The sack extends down to the lower end of the animal. It is lined by delicate membrane. At the orifice on each side, a little posteriorly to the lateral external bars, this inner membrane is strengthened by a pair of thin yellowish bars (c′, fig. 3), which run parallel to the straight portions of the external bars. These inner bars at their lower ends become pointed and die out: at their upper ends, and close to the rim, they are broader, but more flexible, and so transparent as hardly to be distinguished from the rest of the membrane. At the rostral end of the orifice, in a medial line, this same inner tunic of the sack is thickened for a short space downwards, so as to form a fifth bar (c′′); which separating from the inner tunic, runs inwards between the outer and inner membranes of the carapace (i. e. between b and c, fig. 3, 5), behind the mouth, as far down as opposite to the lower end of the œsophagus, and[Pg 570] there becoming thinner and ligamentous, gives attachment to some powerful muscles.
At each exuviation, the external membrane with the dentated hardened orifice, the lateral bars, the inner tunic of the sack with its bars, are all moulted, together with the usual integuments of the animal’s body. New and sharp triturating points are thus periodically formed for the work of excavation. The whole animal increases during growth in every direction, and therefore, at its lower or basal end, as was the case with Alcippe. The disc or surface of attachment, is added to by new underlying layers, extending beyond the old layers at the lower end and on the sides, but not at the upper end, where, as in the case of the calcareous discs of Lithotrya, the old layers are deserted and worn away. I saw what I believed to be little globules or patches of cement; but I was not able to discover any cement-ducts.
Muscles of Sack and Orifice.—The animal is surrounded by rather strong longitudinal muscles, not running up close to the orifice: these muscles exhibited, to my surprise, distinct traces of transverse striæ: there are no external transverse muscles, as in all the Lepadidæ. Attached to both sides of the inward fold or hinge, at the posterior end of the orifice, some striated or voluntary muscles run for a short distance downwards, diverging like a fan: their contraction would cause the dentated rim to open: a strictly homologous muscle occurs only in Alcippe. At the opposite end of the orifice, a remarkably powerful voluntary muscle is attached to the ligamentous bar above described as proceeding from the rostro-medial end of the toothed rim (c′′); and at its lower expanded end, it is attached under rather above the middle of the disc: this muscle corresponds with a similar one in Alcippe, and with some much weaker muscles in other Lepadidæ. Its action would be to draw down within the shell-cavity the whole dentated rim, and likewise to close the orifice; and here, I believe, come into use the lateral elastic horny bars with their curious basal projecting plates, furnished with expanded points, for much friction would thus be caused by, yet some play be allowed for, the several movements; the elasticity of the bar bringing up the dentated orifice, when[Pg 571] the powerful muscles attached to the rostral end of the latter became relaxed. Round the space where the just-mentioned muscles are attached to the horny disc, a sheet of other muscles radiate, a few on both sides obliquely upwards, but the greater number transversely and within the first-described longitudinal muscles; they extend on both sides about half round the animal. There are similar muscles in Alcippe, but not extending so far round the animal. Their action must be to draw the whole carapace towards the surface of attachment; the action of the longitudinal muscles being to shorten it; the orifice supported by the lateral horny bars, serving as the fulcrum for the contraction of the longitudinal muscles. I could not see any adductor scutorum muscle, although I looked particularly under the expanded plates at the ends of the lateral external horny bars.
Body.—This is laterally compressed: it is widest and thickest at the upper end, and thence tapers to the lower or posterior end. The last three or four thoracic segments are bent under the anterior segments, giving the whole something of the appearance of certain crustaceans, divested of their legs. The somewhat conical mouth, with its singular labrum, is very large. The body consists of eight segments. The first segment (fig. 5, 1), or that succeeding the mouth, is the seventh or last cephalic segment of the archetype crustacean; it is the largest of all eight segments; it is joined by its dorsal surface to the carapace or external covering of the animal, and the membrane with which it is invested is prolonged upwards and downwards (c, c, fig. 5), and so forms the inner tunic of the sack. The succeeding seven segments are thoracic; they are free, and are destitute of limbs; the articulations separating them are transverse. The first and second thoracic segments give rise, on their medial dorsal surfaces, each to a remarkable tapering curved appendage, presently to be described. At the end of the last thoracic segment, there is a minute abdomen, bearing three pairs of biramous cirri.
The Mouth consists of three pairs of organs, namely, the outer maxillæ, maxillæ, mandibles with their palpi, and of a great and very curious labrum. These organs, by the fusion (as in other cirripedes) of their lower segments, form a large,[Pg 572] somewhat conical, projecting mouth, which is separated on the ventral surface from the rest of the body by a distinct fold or articulation, where the muscles proceeding to the above gnathites are attached. The labrum (e, fig. 5, 9) is of large size; and the crest close over the opening of the œsophagus is produced into a great, lancet-shaped, moveable organ, wholly unlike anything occurring in any other cirripede: it is coloured purple, and is thickly fringed in the upper part by very fine hairs: it is bowed a little backwards from the mouth: the base, which rather overhangs the œsophagus, is a little contracted, and is transversely marked by an articulation: two small, parallel, voluntary muscles (with transverse striæ) are attached at their lower ends close beneath the articulation, and extend about one third up the organ: their contraction would serve to erect it; and their relaxation would, apparently, allow it to fall backwards on a little knob (e′, fig. 5) behind. This little knob resembles a similar projection in many of the Lepadidæ. As the labrum is formed of similar membrane with that of the succeeding segment of the body, its limit downward, beneath the knob, can be told only by a small apodeme which projects inwards, at a little distance within the line where the membrane of the body is reflexed upwards (c), so as to form the inner tunic of the sack.
The mandibles, palpi, and maxillæ, all project more than is usual. The Palpi (f, fig. 9, 5) are narrow, flattened, and taper a little; they support a few long bristles on their tips, and on one of their sides. In every other cirripede (in which the palpi are developed) they are directed transversely across the mouth, one towards the other, and are for a considerable space united to the labrum: here they project straight up, and seem to rise exteriorly to the bases of the mandibles; they are, however, united to the basal lateral edges of the labrum, and when the latter is torn from the rest of the mouth, the palpi separate with it. I could not distinguish the knob on which, in every other cirripede, the palpi are articulated. The Mandibles (fig. 8) have an upper, lower, and middle tooth, with some finer intermediate points and hairs. The Maxillæ (fig. 7) are narrowed in at their spinose edge, where there are three large spines and several finer[Pg 573] bristles, together forming a flattened brush: this organ is remarkable from the apodeme (a) being bent into the shape of a scythe, with the terminal or blade-portion a little expanded, and directed backwards and inwards. The Outer maxillæ are sub-triangular in outline, with several bristles on their summits and along their outer surfaces.
Segments of Body.—I have stated that the mouth is succeeded by eight segments. As in all the cirripedes hitherto described, the body consists of only six segments, the number eight at first seems very improbable, and therefore I may be permitted to state that both Dr. Hooker and myself, when first examining this animal, and having no notion whatever regarding its homologies or the structure of other cirripedes, came to the conclusion, judging only from external appearances, that is, from the transverse folds, and from the lines of movement when the body was bent by a needle, that there were really eight segments. I have since carefully looked to this point: when the outer membrane is cleaned and examined, the four posterior segments are very plain, owing to a dorsal medial line, being alternately either thickened and coloured yellowish, or thin and colourless: the four anterior segments are less plain, but yet the membrane on the dorsal surface, on the line of each fold or articulation, does present some difference, from being destitute of the fine, transverse, toothed scales which occur on other parts. But I lay most stress on the fact, that all these eight articulations were used for the attachment of muscles. Hence I conclude that the eight segments are real; and we shall see, in the next order, that the very same eight segments are as plain in Proteolepas, as in the larva of an insect or as in an annelid. There is good reason to believe that the general covering or carapace consists in all cirripedes of the three anterior segments, and that the mouth (judging from its appendages) also consists of three segments, consequently the first segment of the body in Cryptophialus must be the seventh or last cephalic segment, and the seven next free segments must be the normal seven thoracic segments.
The first segment of the body (i. e. last cephalic, fig. 5, 1) is, as stated, the largest, and is attached dorsally to the[Pg 574] carapace: its ventral surface is flattened, and is formed of somewhat thickened membrane: on each side, a little below the articulation separating this segment from the mouth, there is a small blunt projection, with the free part only 1/500th of an inch in length. Each of these appendages bears four or five bristles on one side near the summit, and a few on the other side, lower down: from their position I believe them to be rudiments of a first pair of maxillipeds (tetartognathites of Milne Edwards), of which no trace occurs in any other cirripede. The differences between this segment and the seven succeeding segments, is of interest, as offering some confirmation of the belief, lately disputed by some naturalists, that the cephalic and thoracic segments in the class Crustacea, do differ in their nature,—a conclusion which we shall see further confirmed under Proteolepas. The second and third (i. e. first and second thoracic) segments (2, 3, fig. 5) are the next largest, and are remarkable from supporting singular appendages, already alluded to. The sides of the second segment are formed of thickened yellowish membrane. The fifth and sixth segments are the smallest, and mark the point of chief flexure of the body. The eighth segment is a little elongated, formed of thicker membrane than the other segments, and dorsally is indented by the anus.
The singular tapering appendages (fig. 5, i, k) arising dorsally from the second and third segments, differ from each other only by the upper one being smaller, less curled, and perhaps rather smoother. When first examining this animal, not knowing that it was female, and not finding a probosciformed penis, I concluded that these organs were of this nature,—an excusable mistake, considering their almost ringed structure, their somewhat constricted bases, the direction of their curvature, and their position in the midst of the ova within the sack. On careful examination, however, these appendages are seen not to be truly ringed or articulated, but are covered with transverse lines of scales, hirsute on their edges; these scales being less distinct, or even quite absent on the smooth upper portion; they do not include any muscles; they are imperforate at the apex, which is not furnished with bristles (as seems[Pg 575] always to be the case with the probosciformed penis); and, lastly, they are lined by corium, but are not occupied by any vessel, gland, or organ of any kind. The only function which I can assign to these appendages, is that of aiding the retention of the ova within the sack: for the ova, when first produced, are aggregated round them; at this period I several times observed long, somewhat curled, very thin fibres, not tapering like hairs, adhering to both appendages, the nature of which fibres I cannot explain. In very many cirripedes there seems a strong tendency to the production of tapering, filamentary appendages, somewhat like the two (i, k) here described,—namely, at the bottom of the sack in some Balaninæ, at the bases of the anterior cirri in Lepas and in some other genera, and on the dorsal surface of the prosoma in certain species of Pollicipes; in this latter case some of these appendages were covered by scales; and the prosoma whence they arose answers to the third segment of the body in Cryptophialus, or that supporting the lower and larger appendage. Appendages of this nature, in several cirripedes, serve for the lodgment of the testes, but in some cases they are of no apparent use, excepting, perhaps, in aiding respiration by the expansion of corium thus exposed, and this partially may be their function here, for there are no proper branchiæ.
Cirri.—There are three pairs, together forming a nearly straight brush, of considerable length, projecting in a line with the last thoracic segment. Each cirrus is biramous. In a moderately large specimen there were twenty segments in one of the longest rami. Each segment (fig. 14, a) is strengthened by a dorsal or posterior shield of thickened yellowish membrane, from the upper edge of which a single smooth spine projects; the anterior surface is likewise strengthened on the two edges by thickened membrane, and supports two pairs of long spines, which are plumose, or rather hirsute, on both sides. In the lower segments of both rami of the several cirri, the inner spine of each pair is considerably shorter than the outer spine,—evidently in relation to the little power of divergence of the two rami. All the cirri resemble each other, excepting that the rami of the anterior pair, are rather shorter than those[Pg 576] of the other cirri, but the dorsal spines on their segments are longer. In all ordinary cirripedes the two rami are equally supported by the upper segment of the pedicel, which latter is very short compared with the lower segment, and is separated from it by a transverse articulation. Here (fig. 13) the exterior or anterior ramus is articulated on the pedicel, almost like a branch, in front of the other ramus, which seems more directly continuous with the pedicel. The upper segment of the latter is separated from the lower segment, both being of about the same size, by a very oblique articulation. On the front surfaces of the pedicels there are two or three pairs of spines.
Abdomen.—The three cirri on one side are separated from their opposite pairs by a prominent, longitudinal fold, formed of thickened yellowish membrane, which, when foreshortened by being viewed dorsally, looks like a style projecting immediately beneath the anus. The transverse folds separating the three pairs, are exceedingly slight. The inner basal edges of the pedicels of the cirri project slightly inwards as apodemes, giving, I believe, attachment to some muscles. The anus lies between the posterior pair of cirri, and deeply indents the last thoracic segment. The transverse folds separating the three pairs of cirri, little developed as they are, must, according to all analogy, be considered as representing three segments of the body, and as we have accounted for seven cephalic and seven thoracic segments, we must conclude that these are three abdominal segments. We know, moreover, that the abdomen in the pupæ of the Thoracica, with a single exception, does actually consist of three segments.
Movements of the Mouth, Thorax, and Cirri, &c.—Judging from the fact of the lancet-shaped appendage of the labrum being more or less exserted in dead specimens out of the shell-cavity, and from the analogy of other cirripedes, I do not doubt that the whole mouth can be considerably raised and depressed; we have seen, also, that the rostral end of the toothed orifice of the sack can be pulled down, which would aid in exposing the mouth and labrum. The well-articulated thorax, by the aid of the muscles attached to every segment, can certainly be doubled up and contracted,[Pg 577] so that the cirri with their pedicels (coloured purple like the exposed labrum) can be wholly protruded out of the sack and shell-cavity. The three cirri no doubt can be separated a little from each other, both transversely and longitudinally; and according to analogy, the two rami of each cirrus can likewise be separated: there are, also, muscles for moving the two segments of the pedicel of each cirrus; and other muscles run up the many segments of the rami. We have seen that the great lancet-formed appendage of the labrum, laterally fringed with fine hairs, can be erected; and I do not doubt that the prey when entangled by the expanded cirri, is borne against this appendage, and is then, by the retraction of the thorax, dragged down its smooth surface to the mouth, where it is seized by the mandibles and maxillæ, which lie like a trap at the bottom of an inclined and moveable plane.
Alimentary Canal.—The œsophagus is long; it runs backwards from the mouth and then downwards; at its lower end, where it enters the stomach, the part, which in other cirripedes is expanded and bell-shaped, is modified in a most singular and quite peculiar manner; for the lower part of the œsophagus, after widening a little, becomes converted into what appears at first like a square box, 8/1000ths of an inch across. This box is really deeply folded (see diagram, Pl. 24, fig. 11) into six longitudinal ridges and hollows: two of these hollows, facing each other, are wider than the others, and when the organ is dissected out of the body, it generally lies (fig. 10) with one of these faces exactly over the other, the narrower lateral folds being thus hidden, and the whole consequently appearing like a simple square box with concave sides. But when a section is made, or the lower open end is turned upwards, we see that the organ is six-rayed and elongated, with the longer axis standing parallel to the flattened sides of the animal’s body. The edges of the folds are formed of yellowish, thickened membrane, with a sinuous or beaded outline, which serve to strengthen the organ. Internally, the two broad concave sides are armed, in their upper inwardly prominent (fig. 12) part, each with a disc, 2/1000ths of an inch in diameter, crowded with short, thick, brownish, inwardly projecting teeth. The[Pg 578] two discs stand exactly opposed to each other. The bases of the teeth, seen from the outside (Pl. 24, fig. 10), seem like brown little circles, with a smaller circle within. The disc obscurely appears to be formed by the confluence of two smaller discs which lie, I believe, at a very small angle to each other: beneath each of these half discs there is a longitudinal band of very fine hairs, the two bands uniting into one, lower down within the organ. The internal longitudinal edges, also, of the four lateral smaller folds are likewise clothed with fine hairs; hence we have six parallel longitudinal rows of very fine but stiff hairs, or eight, if the united bands on the two broader faces under the discs be counted each as two. These bands of hairs, and the opposed discs, armed with very strong teeth, can be separated and brought together with force, by the action of strong constrictor and diverging muscles. Hence any prey carried down the œsophagus, before entering the stomach, would have to pass, as it were through a mill, and be subjected to a severe trituration by the discs of teeth, and immediately beneath to a brushing by the six longitudinal bands of hairs. This curious and unique structure answers, I believe, the same purpose as the four convex, hardish, crenated buttons on the posterior thoracic cirri in Alcippe, which are likewise unique in that genus. I observed that in some specimens the teeth had been worn quite blunt, but the teeth and hairs are periodically moulted and renewed, together with the whole œsophagus.
The stomach is broadest at the upper end, and extends from a little beneath the mouth down to the fifth segment of the body, where it becomes narrow. It presented an irregularly contracted appearance, and was covered by a pulpy hepatic layer. The rectum is of rather large diameter; it extends from the middle of the fifth segment to the end of the eighth segment of the body, or seventh of the thorax, where the large anus is situated, lying between the posterior abdominal cirri, and partly hollowed out in this seventh segment. The rectum, as in all other cirripedes, is periodically moulted. The food is of a bright green colour, as if of a confervoid nature; the triturating and brushing action of the œsophagus seems to roll this matter into pellets,[Pg 579] which apparently retain this form until finally expelled as excrement: certainly the excrement is in pellets, and I have several times seen pellets within the stomach.
Organs of Generation.—The specimens as yet described are exclusively female, there being certainly no testes or vesiculæ seminales. As in every specimen collected (early in January) there were within the sack either nearly mature eggs, or young larvæ, it was the worst period for seeing the ovarian cæca, and I have failed to discover them in the specimens now long kept in spirits of wine; but I cannot doubt they would be found, between the inner and outer tunics of the carapace or general covering, near the disc. I have stated in my original notes, made when the specimens were alive, that the ova are at first perfectly detached; but some appearances make me believe that I overlooked (as might easily happen) the often excessively fine membrane which in other cirripedes unites the ova together, and so forms the ovigerous lamellæ. The ova are much less numerous than in other genera, varying from only nineteen to about sixty. In the same individual all the eggs were always in the same state of development.
Metamorphoses.—The true ova, in their earliest condition, when in the sack, are ovate (Pl. 24, fig. 15), orange-coloured, quite smooth, and barely 10/1000th of an inch in their longer axes. They soon become broader at one end than at the other; and by degrees the narrow posterior pointed end becomes developed into a slightly club-shaped, almost transparent (fig. 16) horn, and the broader anterior end, into two rather longer horns. The length of the oval part, not measuring the horns, is nearly the same as in the primary true egg condition. There is as yet no trace of internal organs, the whole contents consisting of pulpy granular matter. How far the above changes are effected by moulting, either of the whole or of part of the integuments of the egg-like body, I cannot say; but the pulpy matter within the ovum, even in its earliest stage, was included within an inner envelope or case.
In the next distinct stage (there being, however, slighter intermediate changes) the posterior horn has shrunk, and become converted into a bluntly-pointed conical termina[Pg 580]tion for the whole body (fig. 17), whereas the two anterior horns have approached each other on the future ventral surface, and have increased considerably in length and thickness, and contain within them the prehensile antennæ, which can be externally seen, and which I dissected out of these horn-like cases. The oval part of the egg-like larva (for I hardly know what to call it) is now very slightly shrunk, being hardly more than 9/1000ths of an inch in length. At this stage, these bodies adhere by the tips of their anterior horns, containing the antennæ in process of formation, to the inner tunic of the sack, and likewise in little groups one to another: as the included prehensile antennæ ultimately become attached by cement (proceeding, no doubt, as usual, from a modified portion of the ovarian tubes), it seems probable that some cement may at this early period be excreted, but I could not make out the exact means of attachment. The egg-like larvæ are, also, aggregated round the tapering curled dorsal appendages of the second and third segments of the body, and it is possible that at this, or at an earlier period, these appendages may act like the ovigerous fræna in the Lepadidæ, and serve to retain the egg-like larvæ within the sack.
We come now to the last larval, or pupal condition, before the final metamorphosis into the mature animal; the changes above described have been, at least to a great degree, if not absolutely gradual; but the pupa suddenly appears perfectly developed, from the moulting of the last-described horned, egg-like larva. It is now a free animal crawling about the sack of its female parent. It has increased a little in length, as compared with the oval part of the egg-like larva in its second stage, namely, from a little above 9/1000ths to 16/1000ths of an inch: from the position of the prehensile antennæ in the two states, I have no doubt that this increase of size is entirely due to the anterior part of the pupa being doubled up whilst within the egg-like larval envelope. The pupa in shape (fig. 18) somewhat resembles a coffin, and is far less laterally compressed than other pupæ, and hence can easily be placed either on its dorsal or ventral surface. The prehensile antennæ are of large size: when the animal was alive, they were concealed under and partially included within,[Pg 581] the front part of the carapace or shell, which in this condition was not so much truncated as in the drawing given (fig. 18) of a specimen lying on its back, with its antennæ protruded. Some specimens formerly examined for me by Dr. Hooker, had their antennæ and whole ventral surface forced outwards, apparently from the endosmose of the spirits of wine. The whole dorsal surface, and the overlapping sides of the carapace are elegantly punctured, and are formed of a rather brittle substance, here and there supporting, especially at the front end, some fine and rather long bristles,—which latter I have not seen on the pupæ of other cirripedes. The ventral surface is very narrow towards the posterior end of the animal; it is formed of thin, structureless membrane. On this surface, close to the posterior end, there is a minute orifice, through which three pairs of bristles are protruded, attached to (as I believe) the rudimentary abdomen; the bristles apparently cannot be withdrawn.
The antennæ, (fig. 18) as stated, are of large size compared to the whole animal: they resemble, in all essential respects, the same organ in other cirripedes. The ultimate segment is unusually thick;[152] it is terminated by five bristles, one of which is longer than the others, and stands rather separated from them. The disc-segment is large, nearly circular, with the broad edges transparent and membranous; on its posterior edge there is a single small spine. The second or main segment, counting from the base, has a single spine on its upper margin, close beneath the spine on the disc; it is articulated to the disc-segment, a little way from the disc itself,—which is a peculiarity I have not elsewhere noticed. The basal segment is thick and not so short as usual. These organs are furnished with powerful muscles. They are generally protruded alternately; and by the adhesion of the sucker-like disc, the animal drags itself along. The sucker-disc has great play, and when observing specimens alive, I compared its action to that of[Pg 582] a wrist-joint. The antennæ, when retracted within the carapace, lie parallel to each other.
[152] As I have given, in my former volume on the Lepadidæ, p. 286, so many measurements of the antennæ, I may here add those of Cryptophialus,—the length from the end of the disc to the end of the second segment, (formerly called by me, erroneously, the basal,) is 26/6000ths of an inch; the greatest width of the second segment, 9/6000ths; the length of the little ultimate segment, 3/6000ths, and its width under 3/20000ths of an inch.
Posteriorly to the antenna, I distinctly saw the apodemes to which the eyes are attached: I was not able to distinguish any middle fork to the apodeme, which consequently does not resemble a UU, but U. The eyes are dark purple, and, as usual, compound: in one specimen I counted twelve ocelli within the common spherical envelope.
I could not distinguish any thorax, and certainly there is no mouth; nor, from analogy, could the latter be expected, excepting as forming part of the young cirripede: there are no natatory legs, which the pupæ in all other cirripedes possess. Of the three postero-ventral pairs of bristles, the most posterior or dorsal pair, differs from the other two pairs in being considerably smaller, and in being mounted on elongated pedicels: the two anterior pairs of bristles are strong: the three pairs are articulated, one behind the other, on a small body, apparently enclosed in a minute sack, and certainly attached all round by membrane to the internal edges of the orifice, through which the bristles are protruded. These bristles, when the pupa was alive, were often moved, and served apparently to steady the body during the alternate protraction and retraction of the prehensile antennæ. From the fact of the pupa of other cirripedes having an abdomen, formed of three segments, placed exactly in the same position as the minute body here supporting the three pairs of spines, I believe this body to be the abdomen. In other cirripedes only the posterior segment of the abdomen bears spines, which are supported on little limbs or pedicels, namely, the caudal appendages, the other segments being naked. But as the mature Cryptophialus, unlike other cirripedes, has abdominal cirri, the presence of spines on the corresponding abdominal segments in the pupa, is explained and rendered probable: there can, I think, be little doubt that the small terminal pair of spines, supported on elongated pedicels or limbs, answers to the caudal appendages found in many cirripedes.
The whole course of the metamorphosis is very peculiar. The gradual changes in the egg-like larvæ (for I suppose they must be called larvæ) from a simple oval egg, to pointed[Pg 583] oval, to oval with three horns, and lastly to oval with the two anterior horns larger, and the posterior horn reduced to a mere point, seems to me very curious; and offers, as far as I know, a unique case. It is interesting to reflect how perfect a series, in the development of an animal, we have, in different members of the Articulata,—from an ordinary egg, in which all the changes go on unperceived, and whence a perfect animal is matured,—to an egg-like larva which undergoes the changes just described, and which turns into a pupa that does not eat or increase in size,—to a larva which eats and increases in size, but undergoes only one great change, as in most insects,—to a larva undergoing several great changes, as in the case of ordinary cirripedes, before its final metamorphosis into the mature animal. The first larval condition of other cirripedes, in which there is a single eye, three pairs of thoracic limbs, and a much elongated pointed body, covered by a prolongation of the carapace, is here not fully developed or matured; but this stage is, I think, clearly and very curiously indicated by the posterior horn of the egg-like larva, which we may suppose represents the posterior pointed end of the body, for it disappears in the succeeding stage, just as it does in the second larval condition of other cirripedes. In the first stage of ordinary cirripedial larvæ, the anterior horns are always present, serving, as in the case of these egg-like bodies, to enclose and protect the antennæ during their formation. The second egg-like stage answers to the second larval condition of ordinary cirripedes, as described (and figured, Pl. 30, fig. 1) in the introduction to the Balanidæ. The third or pupal state is fully developed in all cases.
Finally, the pupa of Cryptophialus is peculiar in its punctured, hairy surface, and in its shape, which, in being so much more depressed than usual, retains an earlier larval condition; but its chief and highly remarkable character consists in the entire absence of natatory legs; and, in consequence, instead of there being a large sack within the carapace, with an elongated orifice on the ventral surface, there is only a quite minute orifice at the extreme posterior end of the animal, through which the bristles, borne apparently on all three segments of the minute abdomen, are protruded.
The pupæ of the male and female are exactly alike in all their general characters, and probably in every point of detail; but my later and more minute observations were made only on pupæ, which, from their place of attachment, would certainly have turned into males. As these pupæ, without any further metamorphosis, were developed into males, we may, I think, safely infer that such is the case with the females: and, consequently, that the whole course of the metamorphosis has been, in this cirripede, seen and described. During this whole course, no food could possibly have been obtained, for the pupa is destitute of a mouth or organs of prehension, and the stock of cellular matter, enclosed within the ovum, has been sufficient for all the above changes, and for the final metamorphosis. We shall, moreover, immediately see, in the case of the male, that the stock of cellular matter has also sufficed for the development of testes, spermatozoa, and a wonderfully elongated probosciformed penis.
By throwing pieces of the perforated shell of a Concholepas into acid, I examined several scores of specimens of the Cryptophialus, and on all, with the exception of a few young individuals, males were attached. They were attached by cement, proceeding in the usual manner from the prehensile antennæ, outside, to the edges of the upper half of the disc formed of the thicker not-moulted membrane, by which the female adheres in her chamber: hence the males are included in the upper part of the same cavity with the female, into which they must have crawled as pupæ. I found from one or two up, in one case, to seven males, attached to the same female; four or five being the most usual number. In the early part of January, when all my specimens were taken, many of the males had not shed their pupal integuments, and of those that had, the majority were immature, a few only having spermatozoa: all the females had within their sacks, either ova including almost[Pg 585] perfect pupæ, or fully developed pupæ: we may, consequently, conclude that these young males were maturing in order to impregnate the next set of eggs.
The male, immediately after its metamorphosis from its pupal condition, which has been fully described, is almost globular, but slightly bilobed, and is formed of strong, structureless, transparent membrane, including a mass of cellular matter, apparently without any included organs: it is attached by about the middle, between the anterior and posterior lobes, by the not-moulted prehensile antennæ. When the male is mature, its greatest length, measured from the posterior end, where the orifice is seated, to the anterior and blunter end, is about 13/1000 of an inch, and therefore rather less than the pupa, which was 16/1000ths in length. Relatively to a full grown female, the male slightly exceeds half the diameter of the toothed orifice leading into her sack, see (z) fig. 1, Pl. 23. In the mature condition, (fig. 19), one lobe, namely, the upper or posterior, has become more pointed, and is terminated by a minute orifice, 8/6000ths of an inch in diameter. This orifice is formed by a rim of thickened brownish membrane, which, on what was the ventral surface, has a few very minute, but strong, sometimes bifid spines;—in this one character, the male resembling the female. The other and lower (homologically anterior) end or lobe is broader, and contains a mass of cellular matter, which, from its close resemblance in appearance and position to similar matter within the male Alcippe, I have no doubt forms the contents of the testis. In one single specimen, I succeeded in isolating a vesicula seminalis of small size, containing perfectly distinct spermatozoa. Across the middle, between the two lobes, close under the outer integument, there is a broad layer of rather strong transverse muscular fasciæ. I did not observe any eye, the presence of which I should have expected from analogy. Internally there is no mouth, thorax, cirri, or other organs, excepting the testis and vesicula seminalis just mentioned, and an immensely elongated probosciformed penis, coiled up and filling the rest of the inside of the sack down to the testis, which latter occupies the whole anterior, and generally lower end of the animal. This penis is plainly articulated, and includes fine transversely-striated muscles: no[Pg 586] doubt it can be protruded through the minute orifice, and voluntarily moved about. Out of a male, 12/1000ths of an inch in length, I dissected a penis, which, when not stretched, measured 50/1000ths of an inch in length; when a portion was pulled between two needles, it could be stretched to apparently three times its former length, and I should think that this organ could be extended by the animal to, perhaps, even the 100/1000ths of an inch,—that is, to between eight and nine times its own entire length! The use of this enormously elongated penis obviously is, that the spermatozoa of these males, which are so extremely small in size, compared to the female, should all be conveyed within the sack, and none be lost. It should be borne in mind, that the whole male, including every part, is scarcely larger than a single ovum, of which sometimes sixty have to be impregnated by only two or three males. In a full-grown female, the distance from one of the attached males to the middle of the orifice leading into the sack, is about the 5/100ths of an inch, equal to the length of the coiled up, not-extended penis: the further distance from the orifice of the sack to an ovum lying at the bottom of the sack, would be almost 10/100ths of an inch, so that the spermatozoa have to pass a distance of 15/100ths of an inch from the testis of the male to the lower ova. I believe two thirds of this distance would be passed safely along the probosciformed penis.
The resemblance between the male of Cryptophialus and of Alcippe is truly surprising; and is the more wonderful, considering the great dissimilarity of their pupæ. Hardly any characters can be pointed out in which these males differ, excepting such as might have been thought of only specific value, namely the relative proportions of the different parts, and mere external shape. The peduncle growing a little after the metamorphosis, in the male of Alcippe, and the prolongation of its capitulum with the included oblique ligamentous fibres, are the greatest differences. Having fully remarked, under Alcippe, on the wonderfully rudimentary condition of these males, destitute as they are of so many parts and organs, I will here say nothing further on these singular creatures, destined to discharge their spermatozoa, die, and be succeeded by a fresh set of short-lived male successors.
Cirripedia, with the carapace reduced to two separate threads, serving for attachment: body consisting of one cephalic, seven thoracic, and three abdominal segments, all destitute of cirri. Mouth suctorial, with the mandibles and maxillæ placed back to back, enclosed in a hood, formed by the union of the labrum and palpi. Metamorphoses unknown.
The characters above given fully justify, I think, the formation of this order; though it contains only one species, the Proteolepas bivincta. The mere external appearance (Pl. 25, fig. 7), so wonderfully different from that of every other cirripede, would by itself prompt to this same conclusion. At first sight the Proteolepas, if of fresh-water origin, might even have been mistaken for the larva of some insect, fastened by two threads to its prey. The entire absence of the three anterior segments of the head and therefore of the carapace, or, speaking strictly, the mere rudiment of these parts, forming an envelope to the two cement-ducts,—the absence of a stomach, rectum, and anus,—the entire absence of thoracic and abdominal appendages or cirri,—the absence of a probosciformed penis,—are all negative characters, which might ensue from degradation, so common with parasites; and which might, therefore, have been esteemed of not high classificatory value. But the suctorial mouth, with the palpi and labrum united into a hood, and with the mandibles and maxillæ reversed or turned back to back, so as to be utterly incapable of prehension, is a type of structure not hitherto met with, I believe, in any other animal, and cannot be explained away by degradation. The formation of the ova within the segments of the body, a peculiarity confined to this one cirripede, evidently results from the non-development of the anterior part of the head, within which the ova are usually formed; but the compound structure of the vesicula seminalis is a peculiarity[Pg 588] which cannot be thus explained. Proteolepas has no particular affinity to any other cirripede; it resembles, indeed, Cryptophialus in one important point, but only in one point, namely, in the number of the segments of its body. It is really beautiful to see how the homologies of the archetype cirripede, as deduced from the metamorphoses of other cirripedes, are plainly illustrated during the maturity of this degraded creature, and are demonstrated to be identical with those of the archetype Crustacean. I was at first inclined to rank Proteolepas in one division, and all other cirripedes in another division of equal value; but as it may be inferred from the characters of the prehensile antennæ, that the pupa did not differ much, if at all in any important character, from the pupæ of other cirripedes, I have thought the three orders, which I have instituted, would be the most natural arrangement. As any one looking at the drawing given of Proteolepas, might very naturally feel inclined to protest against its being ranked as a cirripede, I must reurge the importance of the pupal antennæ being constituted on the common type, for from their structure, by the law of correlation, that of the whole pupa may be inferred; and even still more I must insist on the importance of the one great character of the antennæ being cemented to the surface of attachment by matter proceeding, as we shall see, in a modified state, from the great ovarian sack. The structure, also, of the mouth (to a certain extent), the segmentation of the body, though in appearance so peculiar, the hermaphrodite condition, the single penis, the absence of oviducts, all accord with, and taken together demonstrate, its cirripedial nature.
Hab.—Parasitic within the sack of the Alepas cornuta, from St. Vincent’s, West Indies, Brit. Mus.
General Appearance.[153]—The entire animal, as already remarked, curiously resembles, at the first glance, the larva of some insect. It is rounded, but somewhat compressed, and tapers gently towards the posterior end. It lies curved in an arc, the ventral surface being concave, and the dorsal convex, but a little flattened dorsally at the anterior and blunter end. Its length, if straightened, would rather exceed one fifth of an inch. The body consists of eleven segments, which, excepting the three terminal, are conspicuously plain. The first segment is surmounted by a rather small mouth, which any one would, assuredly, at first consider as the entire head, though he would in vain search here for eyes, antennæ, or other parts of the three anterior cephalic segments. On the dorsal surface, low down on the second segment of the body, two, quite flexible, thin, but strong, flattened threads arise, which terminate in a pair of prehensile antennæ, having the usual cirripedial structure. From the penultimate or disc segment of these antennæ, cement has been excreted, by which the antennæ are firmly cemented low down to the rostral end of the sack of the cirripede, the Alepas cornuta, on which it is parasitic: hence Proteolepas lies with its back downwards, and with its ventral concave surface fitting the convex body of the Alepas: its mouth lies under the middle of the soft prosoma of the latter cirripede, which I cannot doubt that it lacerates and sucks.
[153] I may be permitted to premise, that though I procured only a single specimen, yet perceiving its very singular nature, I took such care and length of time in the dissection, and repeated every observation so many times that I think reliance may be placed on the description here given. Fortunately I had acquired, from dissecting many much smaller specimens of various cirripedes, all the advantage which full experience could give me, when I commenced on Proteolepas.
Mouth.—The mouth is suctorial, and is constructed on a different plan from that in any other cirripede, or, indeed, in any[Pg 590] other, as far as I know, articulated animal. It is narrower, in both a longitudinal and transverse plane, than the first segment of the body, and is distinctly separated from it. The lower part on the ventral side, is protuberant and rounded. The summit is square, and is formed by the crest of the labrum, with two large palpi (d, fig. 3), having nearly the usual form amongst the Balaninæ, and pointing towards each other, but differently from in any other Cirripede, they are united for their whole length to the labrum, and by their extremities to each other. These parts together thus form an arch or hood, within which stand the other gnathites. The palpi are roughened by groups of very minute spines. At their bases they can be obscurely seen to be separated from the rest of the mouth by an oblique joining or articulation. The back of the mouth is formed entirely of the labrum, which becomes narrow towards its base: it is, from top to bottom, 20/1000ths of an inch in height. Within the hood formed by the palpi and labrum, a pair (c, fig. 3), of very singular, compounded, mandibular organs project freely, straight up, with their convex outer edges placed parallel and close together, and their teeth pointing directly from each other, so that they stand in a reversed position compared with the jaws of all other cirripedes, and are absolutely incapable of prehension.
This compounded organ is singularly small compared with the palpi and labrum: it is narrow, being about 5/3000ths of an inch in width, but is produced upwards, so that a considerable length projects freely, and the rounded, properly external, margin can be traced down for a length of about 20/3000ths of an inch. In a lateral view of the mouth, the extreme tip of the mandibular organ could sometimes be seen just projecting out of the hood. The mandibular organ, when separated and carefully examined, presents the appearance, represented from a camera drawing, in (Pl. 24, fig. 2): we here see three groups of teeth; of these the lower set (c) consists of blunter teeth, placed more transversely, and easily separated from the others, and altogether clearly appears like a distinct organ. I do not feel nearly so sure regarding the other two sets; my first impression was strongly that they were distinct organs, closely united laterally to[Pg 591]gether,—one (a) probably representing the mandibles, and formed into a single large tooth; the other (b) formed of three teeth, and probably representing the outer maxillæ; the first-mentioned set of teeth, which seemed to me to arise from between the other two sets, being the inner maxillæ. If this view (and it must be remembered how excessively minute the parts are) be not, as I now suspect, correct, we must suppose that the outer maxillæ are aborted, and we have seen some tendency towards this in other cirripedes; the compounded organ being formed only of the mandibles (having on this view four teeth) and the inner maxillæ. As far as the mandibles are concerned, their existence, I may remark, is plainly shown by the presence of the palpi, which in all cases belong to and form part of the mandibles. The ventral surface of the mouth, immediately beneath the free portion of the compounded mandibular organ, consists of a triangular projection, but I could see no appearances to make me suppose that this part represented the outer maxillæ. The compound organ—in general shape, and in the oblique manner in which the front part is cut off and terminates in ligamentous apodemes, to which muscles are attached,—presents an unmistakable likeness to a mandible. It is hollow within, and muscles appear to extend some way up, perhaps to the transversely toothed portion, which represents, as I believe, the inner maxillæ: these two groups of teeth, anyhow, seemed to have some power of sliding over each other, and altered their positions during the course of dissection. On each side of the mouth, there is a muscle attached by its lower end to the basal edge of the labrum, and two others, one above the other, attached by their lower ends to about the middle of the labrum; these muscles, which are distinctly striated or voluntary, I infer, from analogy, run up to the ligamentous apodemes of the compound mandibles. There appeared to be other more delicate muscles attached to the basal articulation of the mouth on the ventral face, and these, I presume, would run to the supposed inner maxillæ.
The mouth in forming a prominence separated by a distinct articulation from the body, and in the union of the palpi and labrum (though here carried to excess), is con[Pg 592]structed so far on the cirripedial type; but how are we to account for the extraordinary reversed position of the united mandibles and maxillæ, with their backs almost touching each other, and their toothed edges twisted round so as to face outwards in a manner unexampled, I believe, in any other articulate animal? It might, perhaps, be at first suspected, that the compounded mandible had not really been twisted round, but that the teeth had been abnormally developed on the outer convex margin: this view, however, certainly cannot be admitted, for the properly outer convex margin can be traced running far down the mouth, in a manner utterly inexplicable, if this were really the inner side; and equally inexplicable on this view would be the position of the ligamentous apodemes. Hence I cannot doubt that this compounded mandibular organ has really rotated on its axis; and if the course of development could be followed, I suspect that the twisting would be seen to be effected as follows: we know in all cirripedes that the outer and inner maxillæ, and to a certain extent the mandibles, instead of facing each other, are directed towards the labrum; they therefore have already been twisted round a quarter of a circle, as may be seen in the diagram (Pl. 24, fig. 4), copied from the mouth of Ibla. Now let us drive inwards the front of the mouth, along a narrow medial line; these organs would then (fig. 5) be compelled to turn round a quarter of a circle more, and so face directly outwards. In this process, the integument between the lower and outer part of the mandible and the base of the palpus, which normally are in close contact, would have to be greatly stretched. By a movement of this order, the mandibles would come to stand posteriorly or exteriorly to the other gnathites; and as far as I could make out (previously to my having any theory) the large single toothed portion of the compound organ which most resembles a mandible, did really stand outside the other toothed portion.
With respect to the action of this singularly constructed mouth; if its ventral and oblique surface were applied to any yielding object, as the adjoining soft prosoma of the Alepas, the compound mandibles would be worked within an absolutely closed chamber. The action of these mandibles[Pg 593] would be to make a transverse slit, and subsequently to serve as a grapnel to keep the mouth closely adpressed to its prey: the other teeth might act in keeping the wound open. When the mouth was thus closely adpressed over a wound, the great power of shortening the whole body which the animal possesses (the œsophagus being closed), would, by the subsequent action of the elasticity of the outer membrane, almost certainly create suction, and thus cause the nutritious juices of the Alepas to flow into the body of the parasite. Hence I have called the mouth suctorial.
Body.—This, as already stated, consists of eleven segments, of which the three posterior (abdominal) smaller segments can hardly be distinguished, without dissection, as separate from each other. The body is mainly occupied by a vast ovarian sack (e, e, fig. 7), filled by innumerable ova: and the three posterior segments by small testes and their vesiculæ seminales (i): but I shall return to the internal anatomy. The outer membrane, lined by delicate corium, is thin, transparent, elastic, and covered by groups of excessively minute blunt little points. The segments can be plainly distinguished by their outlines, especially on the ventral surface; but they are rendered unmistakably distinct by the attachment of the muscles; they can also be perceived when the external membrane is perfectly cleaned, by yellowish lines. The muscular system is highly symmetrical and simple: along all eleven segments, there is a narrow, medial, ventral and dorsal clear space; on both sides of which space there is a band of longitudinal muscles, which, though encroaching on the two sides, and rather largely on the dorso-lateral sides, may be called the ventral and dorsal muscles. These muscles are striæ-less, which is the case with the homologous posterior thoracic muscles in some other cirripedes: on the dorsal surface (lower surface in fig. 7) they are more spread out, and consist, on each side of the medial line, of four ribbons: this seems to be the case on the ventral side, but the ribbons are here much more confluent: in the seventh and eighth segments, the ribbons become broader; but in the ninth, tenth, and eleventh, or three posterior segments, they become much narrower, and some of the fasciæ disappear, so that these muscles can hardly[Pg 594] be seen from the outside. Each separate ribbon expands a little at its two ends, which are attached to the articulations separating the successive segments: I carefully observed that they did not pass over at either end to the adjoining segments: hence their action must be either simply to shorten and arch each segment separately; or when acting together, to shorten the whole body, or perhaps the ventral or dorsal surface by itself.
In the first segment, and in the three posterior segments, these longitudinal muscles alone occur; but on the seven segments, from the second to the eighth inclusive, there are other oblique latero-ventral muscles. These muscles lie within the longitudinal muscles, and adhere pretty firmly to the coat (e, e, fig. 7) of the great ovarian sack. At their ventral extremities they are attached, near the anterior margin of each segment, beneath the point of attachment of the longitudinal fasciæ, and thence they run posteriorly in an oblique line to the anterior margin of the next succeeding segment, where they are attached: so that these muscles run obliquely from segment to segment. The first of these oblique muscles, lying chiefly within the second segment of the body, is thinner and longer than the others: those within the third and fourth segments are short: those within the fifth and succeeding segments extend, at their dorsal (or lower in fig. 7) extremities, as far as the outer dorsal longitudinal fasciæ: those within the seventh segment are broad and short, and cross the longitudinal muscles at only a small angle. In the eighth segment, there is an oblique lateral muscle, like that in the seventh segment, running from the ventral surface towards the dorsal surface; but there is in addition a second oblique lateral muscle, rising from the dorsal surface, and running towards the ventral surface. This muscle does not occur in the other segments, but in the fourth segment, at the dorsal end of the oblique latero-ventral muscle, there may be seen a small branch of fibres, at right angles, which seems to represent a muscle homologous with that just mentioned in the eighth segment: obscure traces, moreover, of similar fibres, can be detected in some of the other segments: had these oblique latero-dorsal muscles been as fully developed in the seven anterior segments of the body, as on[Pg 595] the eighth segment, the whole muscular system would have been perfectly symmetrical. The oblique latero-ventral muscle in the sixth segment is distinctly striated transversely; but this is not the case with most of the other muscles, if with any of them; I cannot account for this difference. The muscles of the gnathites are the only other voluntary muscles in the animal’s body.
Homologies of the Body.—It will hereafter be, I think, clearly shown, that when the shell and integuments of the pupa of Proteolepas are shed, no carapace or general covering for the body is formed; the three anterior segments of the head, the backward prolongation of which (as has been elsewhere explained) certainly forms the carapace of ordinary cirripedes, being here almost absolutely aborted. In every cirripede the mouth is formed of three pairs of gnathites, which, no one will doubt, rise from the fourth, fifth, and sixth segments of the head: here in Proteolepas, the mouth, even on the view of the mandibular organ on each side being compounded of only two gnathites, sufficiently resembles the ordinary cirripedial type to make it very probable, that if examined in the earliest stage of its development, three pairs of gnathites would be discovered. In accordance with this conclusion, the segment succeeding the mouth (i. e., the first segment of the body in fig. 7) homologically is the seventh, or last cephalic segment. The succeeding seven segments, of course, are the seven thoracic segments, and the three posterior segments are abdominal; the latter are not developed in ordinary cirripedes when mature, but are present during their pupal condition. Now this conclusion, which is, in fact, deduced from what we know of the front part of the head in other cirripedes, both larval and mature, appears to me most satisfactorily confirmed by the differences in the muscular system of the segments in Proteolepas. In no other way, I believe, can it be explained, why the last cephalic segment and the three abdominal segments should differ from the seven thoracic segments, in the entire absence of the oblique lateral muscles. The abdominal segments, moreover, differ a little in shape, in the indistinctness of their articulation, in the thinness of the longitudinal muscles, and even in their contents. With respect to the two threads[Pg 596] enclosing the cement-ducts, which spring from the second segment of the body (or first of the thorax), and which terminate within the prehensile antennæ of the pupa, we shall hereafter see that their apparently most anomalous position, and even the flattened shape of the dorsal surface of the two anterior segments of the body, all accord perfectly with the homologies just given.
Alimentary Canal.—The œsophagus is thin, and for a cirripede short, for it extends only half-way down the first segment (i. e. last cephalic) of the body; the lower end, which is slightly dilated, nearly touches the anterior end of the great ovarian sack. At its upper end, it is surrounded by delicate, striæ-less constrictor muscles; and there are others radiating outwards, evidently serving to open it: the lower part of the œsophagus, differently from other cirripedes, is destitute of muscles, and is only coated by a thin layer of corium, which would serve to produce a new œsophagus at each exuviation. Strange as the fact may be, I am prepared to assert that there is no stomach, rectum, or anus. As I was able to trace so distinctly the œsophagus, and likewise the generally far smaller orifice and ducts of the male generative organs, I consider it quite impossible that I could have missed the stomach. The rectum and anus are absent in Alcippe: and the absence of a stomach is here in some degree the less surprising, as the structure of the mouth shows that Proteolepas must live on the already elaborated fluids of the Alepas, to which, being a cirripede, it is allied. It is of some importance to observe, that the œsophagus is fitted with muscles simply for shutting and opening it, the wave-like swallowing action of which other cirripedes are capable, being, apparently, here impossible; but the contraction of the body and its subsequent expansion, the œsophagus being opened, would allow the blood of its prey to flow inwards.
The nervous system must be much atrophied, for I could not detect it, and the small size of the animal is not sufficient to account for this: I wish I could have seen this system, for then I should almost certainly have beheld an articulate animal without a trace of a supra-œsophageal ganglion. There is no eye, but such could hardly be expected, as the[Pg 597] anterior cephalic segments are aborted. There are no branchiæ. I may state that within the abdomen, along the dorsal surface, there was either a lacuna or a delicate vessel, apparently of a circulatory nature, of considerable diameter, which, near the extreme posterior end of the body, gave out branches.
Female Reproductive Organs.—The eight anterior segments of the body, with the exception of a small space at the two ends, are occupied by an immense (e, e), opaque, ovarian sack. The tissue forming it is delicate, and presents a peculiar cellular aspect: it is slightly attached to the corium on the ventral surface of the body, and to the oblique latero-ventral muscles. Internally, at the anterior end, it is thickly coated by cellular matter, the cells varying from 4/6000ths to less than 1/6000th of an inch in diameter, becoming in parts confluent, and the whole forming a dark orange-coloured mass. In the more central parts of the sack this cellular matter became aggregated into little pellets, which, in proceeding towards the posterior end of the sack, gradually increased in size, from about (4 to 6)/1000ths of an inch in diameter, and at last appeared as almost mature and perfect ova of a broadly oval figure. Their size, as we see, is small, and their number almost infinite. I carefully examined all round this ovarian sack, and could detect no oviducts; nor from analogy could they be expected: I have no doubt that the ova burst forth by the rupture, probably, of the posterior end of the sack and of the overlying corium; and that they accumulate beneath the external membrane of the body, until this is moulted, the rupture beneath being in the meantime healed, when they are freed, or perhaps temporarily protected in the old moulted envelope of the body.
On each side, within the first two segments of the body, and projecting a little before the great ovarian sack (e), two gut-formed organs (f) may be seen, even from the outside, owing to their opacity and dark colour. They lie near the external surface; the first pair of latero-ventral oblique muscles passing between them and the ovarian sack. They are formed of a branching, grape-like mass of opaque, orange-coloured cells. They are intimately united, at their pos[Pg 598]terior extremities, to the ovarian sack, and I believe open into it; but I cannot say that I demonstrated this. From their absolute identity in structure, and similarity in position, namely, on each side of the lower end of the œsophagus, no doubt is left on my mind that these bodies answer to the true ovaria, which are situated within the body of other cirripedes; and that the ovarian sack answers to the inosculating and branching ovarian tubes and cæca, which fill the peduncle, or cover the basis in other cirripedes, but here, from the absence of these parts, necessarily occupying the body.
Male Organs.—The whole surface of the ovarian sack, the space before it, even to within the lower parts of the mouth, the posterior half of the last thoracic segment, and especially the whole three abdominal segments, are completely netted by branching delicate vessels or ducts terminating in spherical glands about 1/2000th of an inch in diameter. These little glands include a brownish pulpy centre, and sufficiently resemble the testes of other cirripedes in appearance, position, and connecting ducts, to make me believe that such is their nature. I may remark that in the more central parts of the abdomen the glands and ducts seemed to be in process of formation by the confluence of cellular matter, and in some other cirripedes I have suspected that the testes are periodically renewed, or at least redeveloped from an undistinguishable condition. Within the posterior half of the abdomen, some of the ducts become thicker and unite, others joining in laterally, so as together to make a dark chord, 7/2000ths of an inch in diameter. Until dissecting this chord, I thought it was a single vesicula seminalis, but it separated into several rather thick ducts or vesiculæ. I was not able to remove from within them the contained matter, but it appeared very finely and longitudinally flocculent, like spermatozoa not quite matured. In accordance with the immature state of the contents of the ovarian sack, in all probability these ducts would hereafter have become greatly enlarged, and have formed a compound vesicula seminalis of considerable size. The dark chord, formed by their union, contracts as it enters the rudimentary penis, and terminates in a very minute orifice on[Pg 599] its apex. The penis consists of a papilla, only 3/4000ths of an inch in length, situated on the extreme point of the abdomen, but rather towards the ventral surface.
Metamorphosis.—In accordance with the general law of the correlation of parts, it may be inferred, from the description and measurements of the pupal antennæ immediately to be given, that this abnormal creature was developed within a pupa of the same general structure, and of about the size, as the pupæ whence Scalpellum, Alcippe, and many other cirripedes are developed. As the ova are of remarkably small size, indeed I have seen no others quite so small, it is certain that the larvæ, as in the case with all other cirripedes, excepting Cryptophialus, must undergo several metamorphoses, and increase much in size, before attaining their pupal condition.
Attachment.—The animal is attached, as already stated, to the sack of the Alepas by two threads, rising close together from the medio-dorsal line, near the posterior end of the second segment of the body. These threads are attached likewise close together at their further ends, by the antennæ, into which they enter. They are flattened and strong, yet quite flexible, with a somewhat sinuous surface: they were, in this specimen, 42/1000ths of an inch in length, and a little above 3/1000ths in diameter: where joined to the thoracic segment they were a little contracted. Their structure in this specimen could be made out (Pl. 24, fig. 1) with perfect distinctness. Their transparent outer tunic (e, fig. 1) is 1/2000ths of an inch in thickness, and is continuous with that (d) enveloping the whole body, but is abruptly and considerably thicker than this membrane; and hence a very slight collar is formed outside, round the line of junction of each thread with the body. The delicate corium (c) lining the external membrane of the body runs, at least someway, down these threads. It was likewise indisputably evident that the membrane (b), for I separated it by dissection, forming the great ovarian sack, together with the cellular contents of this sack (a), entered and extended down both threads. It should, also, be particularly observed, that the coarsely cellular matter within the ovarian sack, immediately that it entered the tube formed by the membrane of the ovarian sack, suddenly changed[Pg 600] its appearance into a homogeneous, stiff, pulpy matter, which retained the same appearance all down the threads to within the antennæ. This finer matter readily separated from the coarser cellular matter within the sack, but was not divided from it by any septum or membrane. Some way within the threads, the corium, the membrane of the ovarian sack, and the contents appear (e′), as seen from the outside, to become, and perhaps really are, blended together. These threads could not have been originally formed of their present length, and must therefore have been added to during the growth of the animal; but from their entering the not-moulted antennæ, and from the animal being permanently attached by them, they cannot have grown, by means of the moulting of their integuments; hence I conclude that at each period of growth and exuviation they have been added to only at their upper ends, where there is a sort of collar, or line of growth; and where, I may remark, the lining corium is alone well developed. We shall presently see the bearing of these remarks.
These threads contract to about half their former diameter as they enter the old prehensile antennæ of the pupa, within which they are firmly attached. Each thread, with its three tunics apparently blended together, can be traced to the extremity of the disc-segment (g), where the included matter seems to have burst forth. The whole disc and the terminal segment of both antennæ are enveloped, close together, in cement, formed into two almost separate little capsules, by which they adhere very firmly to the integuments of the Alepas. The cement required to be removed before the antennæ could be plainly seen. The cement presented all the usual characters, namely, its homogeneous laminated structure and its yellowish colour. The cement in the case of the male Ibla, which is parasitic within the sack of the female Ibla, affects the corium and fibrous matter beneath the chitine-tunic, and causes them to adhere together, and thus prevents the male from being cast off each time that the inner tunic of the sack of the female is moulted: exactly so has the cement of the Proteolepas affected the integuments of the Alepas. The only difference between ordinary cement-ducts and the two threads here[Pg 601] described is, that the ducts, in both cases formed by the prolongation of the coat of an ovarian receptacle, are here protected by a thick outer membrane, lined, at least in the upper part, by corium; whereas, in the Lepadidæ the two ducts are included within the peduncle, and are therefore protected by one common membrane, lined of course by corium; and this membrane, we shall presently see, is homologous with that separately investing the two threads.
The antennæ differ remarkably little, considering the anomalous character of the mature animal, from the same organ in other genera; they come nearest, perhaps, to the antennæ of Ibla. The length of the disc (g, fig. 1) and great succeeding segment (f) together is 40/6000ths of an inch. The lower segment has its basal articulation only slightly oblique, showing that, as in Alcippe and Ibla, it was probably articulated near the anterior end of the pupal shell: it is of nearly the same width throughout.[154] The disc (g) is remarkable from its great proportional length; it is hoof-shaped, with the outer side rather protuberant, and the end pointed. The ultimate segment (h) is of moderate size: as in Ibla, it has a shoulder or notch on its inner side near its end, bearing two long spines; and probably there were originally three or four spines on the square broad upper end, but these have been broken. This segment is articulated unusually near to the end of the disc.
[154] As I have given the measurements of the antennæ in so many genera, I will give these: second (f, fig. 1) segment, 24/6000ths of an inch in length, and (8-9)/6000ths in width. Disc, 16/6000ths in length, and 8/6000ths in width. Ultimate segment, 6/6000ths (?) in length, and 10/20,000ths (?) in breadth.
The foregoing remarks on the two threads by which Proteolepas is attached, are, independently of their relation to this individual animal, of considerable interest. In my volume on the Lepadidæ, I have stated, after repeated and rigorous examinations (for I was well aware how singular the facts were), that in Conchoderma aurita and in some other genera, the cement-ducts, which entered the pupal antennæ, could be traced till they joined a gland, the coat of which gland was absolutely continuous with the coats of the adjoining and continuous ovarian tubes, of which it was only a modified portion; and what[Pg 602] was still more remarkable, that the matter within the gland was continuous with, and differed only from, the cellular matter within the ovarian tubes and cæca (from which ova were in the act of formation), by being more homogeneous and more coherent. Furthermore, I have shown, that in Ibla an ovarian tube, becomes by a very small change, namely, by a double flexure and slight thickening of its coat, converted into a gland, and thus acquires the power of affecting the cellular ovarian matter and changing it into cement. Now, in Proteolepas, the great ovarian sack replaces the ovarian tubes and cæca; and we here see the very same relations even still more plainly; for the coat of the ovarian sack is indisputably continuous with that investing or forming the two cement-ducts within the two threads; and immediately that the coarse cellular matter, which within the ovarian sack is being converted into ova, enters the upper contracted end of the cement-duct, by some power, we must suppose, inherent in its coat, it is converted into cement, which debouches with all its usual properties through the pupal antennæ. I may venture to reaffirm that nothing could be plainer than this structure, or be in more striking conformity with my previous observations, given in the introduction to the Lepadidæ.
I can hardly express the perplexity which I felt when I first examined Proteolepas, and when I naturally mistook the mouth for the entire head, for I saw, as I thought, the antennæ in direct connection with the second segment of the body, posteriorly to the mouth! It was quite as monstrous and incredible an inversion of the laws of nature, as those fabulous half-human monsters, with an eye seated in the middle of their stomachs. After a time, I perceived that the following considerations removed all difficulty, and brought Proteolepas into the type of other cirripedes.
Firstly: in ordinary cirripedes, the two cement-ducts can be traced up from the cemented antennæ to the glands, formed by a part of the ovarian branching cæca; and the latter can be traced to where they enter, as two simple tubes, the body of the animal, at a medio-dorsal point, a[Pg 603] little anteriorly to the prosoma, or second thoracic segment.[155] From what is actually seen in the complemental male of Scalpellum Peronii, and from what may be inferred from the structure of these parts in the pupæ of all cirripedes, there can be no doubt that if the ovarian cæca were in any case not developed, the cement-ducts would enter the body at the spot where the two simple ovarian tubes, which serve to unite the ovarian cæca with the true ovaria, do enter. Now if we look at the drawing (Pl. 25, fig. 7) of Proteolepas, we shall see that the cement-ducts enter the body at a medio-dorsal point, a little anteriorly to the second thoracic segment, and therefore in the normal position.
[155] This may be partially seen in the section, fig. 1, of Balanus, on the same plate (25) with the figure of Proteolepas; here (bearing in mind that Balanus is a much modified form) (z) shows the pupal antennæ, within which, whilst young, the cement-ducts are included, and are directly continuous with the layer of branching ovarian cæca (g), which are prolonged up to the ovaria as a pair of simple tubes (only one being here represented), entering the body above the upper margin of the prosoma (c). The prosoma of Balanus, I may add, answers to the segment 2 t in fig. 7 of Proteolepas; (e) the mouth in Balanus, of course corresponding with (m) the mouth of Proteolepas; the segment 1 c and 2 t of the latter, are in Balanus aborted or confluent, at least on the ventral surface; and, lastly, the whole great shell of Balanus, the sack with its muscles and the branchiæ, and the opercular valves with their muscles, are all represented in Proteolepas merely by the outer membrane of the two threads (g), which enter the pupal antennæ!
Secondly: the external membrane of the two threads, investing the two cement-ducts, it should be remembered, is not moulted, and is added to during growth (being lined internally by corium), only round the upper, collar-like edge.
Thirdly: the external covering or carapace of every young cirripede, at the period of its metamorphosis, enters, at its lower end, the cemented antennæ, in the form of two short tubular prolongations, by which alone, at first, the cirripede adheres to the surface of attachment; within these prolongations the cement-ducts are included. I have, moreover, seen instances, as in Conchoderma aurita and in the male of Ibla and Alcippe, in which these tubular prolongations, lined internally by corium, were increased a little in length, so as to form a trouser-like termination to the peduncle. That the forked extremity should be a little more developed, and so be converted into a pair of short tubular threads, cannot be considered as very improbable.
Fourthly: in the male Ibla the capitulum is so much atrophied that it does not enclose the thorax or mouth, but still an elongated support or peduncle is left. But it would be no very violent assumption to imagine the peduncle, which does not essentially differ from the capitulum, to become likewise rudimental,—to grow smaller and smaller, and shorter and shorter, till the merest remnant was left at the spot where it entered the cemented antennæ. And in the last paragraph it has been shown that it would be no violent assumption to imagine this lower end of the peduncle, where it enters the antennæ, developed into two short thread-like prolongations.
Lastly: it is certain, from the existence of the prehensile antennæ, that Proteolepas was developed within a pupa, probably differing in no very essential respect from the pupæ of other cirripedes. Therefore, in accordance with all analogy, we may believe that the position[156] of the young Proteolepas (probably much coiled up, with a deep fold close under the mouth) within the pupa, the general form and structure of the latter, and the course of the[Pg 605] cement-ducts, did not essentially differ from the imaginary figure given, Pl. 25, fig. 6. Now, at the period of the metamorphosis, let us imagine that no general covering or carapace was formed, except a small portion on the ventral surface, round the cemented antennæ. Let us further suppose this remnant to be specially developed (as in the case of some cirripedes) into a short trouser-like prolongation, entering the antennæ; and subsequently, in accordance with the almost universal laws of growth in cirripedes, that this portion was never moulted, but continued to be added to, during growth, only at its upper end. By this means we should produce every leading peculiarity of the Proteolepas bivincta. As this parasite lives within the sack of another cirripede, and is protected by the capitulum of the latter, we can understand, in accordance with the usual admirable economy of nature, the absence of any general covering for its body. We can now, also, understand the structure and manner of growth of the two threads by which it is bound to its prey; and the connection, at first so strange and perplexing, between the old pupal antennæ and the second segment of the thorax. I am convinced that no other explanation than that here given, will accord with the relations of the several parts and organs of Proteolepas. Consequently, I fully believe that we here see an articulate animal in which the whole of the three anterior segments of the head have been, during the act of metamorphosis, absolutely aborted, with the exception of a mere rudiment on the ventral surface, near the anterior end, round the old antennæ, and which rudiment has been specially developed as a covering for the two cement-ducts. As the pupal antennæ are, homologically, the second pair of antennæ, we may further infer that this modified remnant of the carapace, investing the two threads, belongs to the third cephalic segment.
[156] Any one who has not specially attended to the metamorphoses of ordinary cirripedes, who looks at the imaginary figure of the young Proteolepas, will feel much surprise at the relative positions of the parts; for the mouth and the first and even second segments of the body stand posteriorly (i. e. above in the figure) to the succeeding segments of the body, in relation to the carapace of the pupa; but this is only in accordance with the remarkable change in position (as explained in the introduction, p. 123, pl. 30, fig. 2), amounting almost to inversion, which the whole thorax of every young cirripede undergoes within the pupa, whilst the anterior cephalic portions and general covering are developed conformably with the pupal carapace, whence it arises that the dorsal surface of that part of the thorax immediately succeeding the mouth becomes attached to the ventral internal surface of the carapace. I believe that the peculiar flattened dorsal outline of the first two segments of the body of Proteolepas is due to these parts having been formed in contact (as represented in pl. 25, fig. 6) with the straight ventral surface of the carapace of the pupa. To place the young Proteolepas, and at the same time the carapace of the pupa, with all the parts in proper homological sequence, it would be necessary to seize the posterior end of the abdomen (a), and pull till the dorsal surfaces of the first and second segments of the body, separated from the ventral internal surface of the carapace, and stood posteriorly (i. e. above in figure) to the mouth, which latter would thus also have to rotate a quarter of a circle, so that the orifice would come to be directed outwards. Then every part would stand, in accordance with the archetype crustacean structure, in due order; but the three confluent anterior cephalic segments, forming the front part and carapace of the pupa, would, as in the case of all cirripedes, be of disproportionately large size in relation to the rest of the body.
Crustacea ex anteriore capitis parte defixa, cæmento in hunc usum ex ovariorum portione ad id specialiter modificatâ emisso. Archetypus è segmentis 17 compositus, quorum 3 priora magna, in carapacem sæpissimè conformata, quæ non omnino exuitur et varios motus efficit: antennæ nullæ: oculi rudimentarii: os prominens, formatum è labro, palpis, mandibulis, et duobus maxillarum paribus, quæ omnia partim confluunt: thorax ad superficiem internam sternalem carapacis affixus, plerumque cum membrorum captantium biramorum, multiarticulorum paribus 6: abdomen plerumque rudimentarium: branchiæ, si quæ adsunt, ad inferiora carapacis latera affixæ: plerumque bisexualia; in unisexualibus, mares fœminis parasitice inserti: penis unicus, plerumque probosciformis, ad posteriorem abdominis extremitatem situs: oviductus nulli: metamorphoses multiplices.
Cirripedia quibus pro carapace est aut capitulum pedunculatum, aut testa operculata cum basi. Corpus è 6 thoracicis segmentis, ferè cum 6 cirrorum paribus, constat. Abdomen rudimentarium, sed sæpe cum appendiculis caudalibus. Oris labrum motus proprios non efficit. Larva primò monocula cum 3 crurum paribus, postremò binocula cum 6 crurum thoracicorum paribus.
Cirripedia sine pedunculo: scuta et terga musculis depressoribus instructa: reliquæ testæ valvæ inter se immobiliter conjunctæ.
Rostrum cum radiis, sed sine alis; valvæ testæ laterales omnes, ex uno latere alis, ex altero radiis instructæ: parietes ferè aut porosi aut ad interiorem superficiem longitudinaliter costati.
Scutum et tergum inter se articulata aut mutuò interclusa: branchiarum unaquæque ex unicâ plicâ constat.
1. Genus—BALANUS, Auctorum. (Darwin, ‘Balanidæ,’ p. 177.)
Valvæ testæ 6; basis calcarea aut membranacea; valvæ operculares subtriangulares.
2. Sub-Genus—ACASTA, Leach. (Darwin, ‘Balanidæ,’ p. 302.)
Valvæ testæ 6; parietes et basis non porosa; basis calcarea, cyathiformis, non elongata. Spongiis, aut rarò Isidis cortici, affixa.
3. Genus—TETRACLITA, Schumacher. (Darwin, ‘Balanidæ,’ p. 321.)
Valvæ testæ 4; interdum inter se externè confluentes: parietes poris perforati, multis plerumque seriebus; basis plana, irregularis, calcarea aut membranacea.
4. Genus—ELMINIUS, Leach. (Darwin, ‘Balanidæ,’ p. 345.)
Valvæ testæ 4; parietes non porosi; basis membranacea.
5. Genus—PYRGOMA, Leach. (Darwin, ‘Balanidæ,’ p. 354.)
Valvæ testæ in unam confluentes; basis cyathiformis aut subcylindrica, coraliis affixa.
6. Sub-Genus—CREUSIA, Leach. (Darwin, ‘Balanidæ,’ p. 375.)
Valvæ testæ 4, radiis instructæ; basis cyathiformis, coraliis affixa.
7. Genus—CHELONOBIA, Leach. (Darwin, ‘Balanidæ,’ p. 382.)
Valvæ testæ admodum crassæ, 6; sed ex iis, valva rostralis intùs è tribus valvis rudimentariis conjunctis constat; basis membranacea; scuta angusta, tergis cristâ articulari corneâ conjuncta.
Scutum et tergum (ubi ambo adsunt) non inter se articulata; basis membranacea; parietes sæpe profundè plicati, laminâ exteriore, ad basin versâ, plerumque imperfectâ; branchiarum unaquæque e duobus plicis constat. Testa vertebratis vivis affixa.
8. Genus—CORONULA, Lamarck. (Darwin, ‘Balanidæ,’ p. 397.)
Valvæ testæ 6; æquali latitudine; parietes tenues, profundè plicati, plicis cavitates infrà solùm apertas efficientibus; valvæ operculares orificio testæ multo minores. Cetaceis affixa.
9. Genus—PLATYLEPAS, J. E. Gray. (Darwin, ‘Balanidæ,’ p. 424.)
Valvæ testæ 6; unaquæque bilobata et intùs producta, ita ut 6 medias costas longitudinales efficiant, quæ basin membranaceam extrorsùs convexam sustinent.
10. Genus—TUBICINELLA, Lamarck. (Darwin, ‘Balanidæ,’ p. 430.)
Valvæ testæ 6, æquali latitudine; testa subcylindrica, orificio latiore quam basis, pluribus cristis transversis virgata. Cetaceis affixa.
11. Genus—XENOBALANUS, Steenstrup. (Darwin, ‘Balanidæ,’ p. 438.)
Testa pæne rudimentaria, stelliformis, è valvis 6 formata, è quarum medio corpus longum pedunculiforme exoritur; valvæ operculares absunt. Cetaceis affixus.
Rostrum cum alis sed sine radiis; valvæ rostro-laterales utrinque sine alis; parietes non porosi.
12. Genus—CHTHAMALUS, Ranzani. (Darwin, ‘Balanidæ,’ p. 447.)
Valvæ testæ 6; basis membranacea, sed interdum ad speciem calcarea, ideò quod parietes inflectuntur.
13. Genus—CHAMÆSIPHO, Darwin. (‘Balanidæ’ p. 470.)
Valvæ testæ 4, suturis sæpe admodùm obliteratis; basis membranacea.
14. Genus—PACHYLASMA, Darwin. (‘Balanidæ,’ p. 475.)
Valvæ testæ, conchâ recenter natâ, 8; adultâ aut 6, aut ad speciem 4, ideò quod valvæ laterales arctè conjunguntur; basis calcarea.
15. Genus—OCTOMERIS, G. B. Sowerby. (Darwin, ‘Balanidæ,’ p. 482.)
Valvæ testæ 8; radii marginibus crenatis; basis membranacea.
16. Genus—CATOPHRAGMUS, G. B. Sowerby. (Darwin, ‘Balanidæ,’ p. 485.)
Valvæ testæ interiores 8, cum pluribus exterioribus parvarum supplementalium valvarum verticillis; basis aut membranacea aut calcarea.
Cirripedia sine pedunculo; scuta et terga, musculis depressoribus non instructa, ex uno latere tantum mobilia, ex altero cum carinâ et rostro in testam asymmetricam immobiliter conjuncta.
1. Genus—VERRUCA, Schumacher. (Darwin, ‘Balanidæ,’ p. 496.)
Cirripedia pedunculo flexili, musculis instructo; scuta et terga (si qua adsunt) musculis depressoribus non instructa; reliquæ valvæ (si quæ adsunt) in annulum immobilem non conjunctæ.
1. Genus—LEPAS, Linn. (Darwin, ‘Lepadidæ,’ p. 67.)
Valvæ 5, approximatæ; carina sursùm inter terga extensa, deorsùm aut furcâ infossâ aut disco externo terminata; scuta subtriangula, umbonibus ad angulum rostralem positis.
2. Genus—PŒCILASMA, Darwin. (‘Lepadidæ,’ p. 99.)
Valvæ 3, 5, aut 7, approximatæ; carina solùm ad basales apices ter[Pg 609]gorum extensa, termino basali aut truncato aut in discum profunde infossum producto; scuta pænè ovalia, umbonibus ad angulum rostralem positis.
3. Genus—DICHELASPIS, Darwin. (‘Lepadidæ,’ p. 115.)
Valvæ 5, quæ ferè pro septem haberi possent, scuto in segmenta planè duo, ad angulum autem rostralem conjuncta, diviso; carina plerumque sursum inter terga extensa, deorsum aut disco infosso aut furcâ aut calyce terminata.
4. Genus—OXYNASPIS, Darwin. (‘Lepadidæ,’ p. 133.)
Valvæ 5, approximatæ; scutorum umbones in medio marginis occludentis positi; carina rectangulè flexa, sursùm inter terga extensa, termino basali simpliciter concavo.
5. Genus—CONCHODERMA, Olfers. (Darwin, ‘Lepadidæ,’ p. 136.)
Valvæ 2 ad 5, minutæ, inter se remotæ; scuta bi- aut tri-lobata, umbonibus in medio marginis occludentis positis; carina arcuata, terminis utrinque pæne similibus.
6. Genus—ALEPAS, S. Rang. (Darwin, ‘Lepadidæ,’ p. 156.)
Capitulum aut sine valvis, aut scutis corneis, pæne abditis.
7. Genus—ANELASMA, Darwin. (‘Lepadidæ,’ p. 169.)
Capitulum sine valvis; aperturâ amplâ; pedunculus fimbriatus, sub-globosus, infossus.
8. Genus—ALCIPPE, Hancock. (Darwin, ‘Balanidæ,’ p. 529.)
(Fœm.) Capitulum sine valvis, aperturâ spinosâ; pedunculus ad basalem extremitatem crescit; superficie rostrali depressâ et disco corneo tectâ; capitulum et pedunculus in cavitate conduntur, ab ipso cirripedio formatâ.
9. Genus—IBLA, Leach. (Darwin, ‘Lepadidæ,’ p. 180.)
(Herm. et Fœm.) Valvæ 4, corneæ; pedunculus spinis corneis, persistentibus vestitus.
10. Genus—SCALPELLUM, Leach. (Darwin, ‘Lepadidæ,’ p. 215, et ‘Lepad. Foss.’ p. 13.)
(Herm. et Fœm.) Valvis 12 ad 15; lateribus verticilli inferioris quatuor vel sex, lineis incrementi plerumque convergentibus; sub-rostrum rarissime adest; pedunculo squamifero, rarissime nudo.
CHARACTERES VALVARUM IN SPECIEBUS FOSSILIBUS.
Carina angusta, introrsùm arcuata, ab apice ad marginem basalem paululum dilatata; carinæ parietes valde inflexi, costis manifestis a tecto plerumque disjuncti; in multis speciebus intra-parietibus instructa; intraparietes nonnunquam supernè producti ultra umbonem carinæ, qui fit inde subcentralis; carinæ parietum lineæ incrementi perobliquæ.[Pg 610] Scuta plerumque subconvexa et tenuia, trapezoidea; marginibus tergalibus lateralibusque angulo insigni disjunctis.
11. Genus—POLLICIPES, Leach. (Darwin, ‘Lepadidæ,’ p. 293, et ‘Lepad. Foss.’ p. 42.)
Valvæ ab 18 usque ad 100 et amplius; lateribus verticilli inferioris multis; lineis incrementi deorsùm ordinatis; subrostrum semper adest; pedunculus squamiferus.
CHARACTERES VALVARUM IN SPECIEBUS FOSSILIBUS.
Carina ab apice ad marginem basalem multum dilatata, apice plerumque liberè prominente; carinæ parietes à tecto non distincté separati, lineis incrementi parietum parum obliquis. Scuta plerumque subsolida, convexa, subtrigonalia, margine tergo-laterali plus minusve eminente, sed non angulo in margines duos discreto.
12. Genus—LITHOTRYA, G. B. Sowerby. (Darwin, ‘Lepadidæ,’ p. 332.)
Valvæ 8, si inter eas parvum (sæpe rudimentale) rostrum et duo parva latera numerentur; incrementi lineis concinnè crenatis; pedunculus squamis calcareis parvis vestitus, in verticillis superioribus crenatis, aut calyci basali calcareo aut discorum ordini affixus.
13. Genus—LORICULA, G. B. Sowerby, jun. (Darwin, ‘Lepad. Foss.’ p. 81.)
Capitulo decem (fortasse) valvis instructo. Pedunculo seriebus decem squamarum lævium calcarearum instructo; sex lateralibus multum transversè elongatis; quatuor terminalibus angustis; secundùm pedunculi margines rostralem et carinalem decurrit sutura medialis recta, squamis non intersecantibus.
Cirripedia quibus carapax lageni-formis est; corpus ex 1 cephalico, 7 thoracicis, 3 abdominalibus segmentis constat; quorum abdominalia tribus cirrorum paribus muniuntur; thoracica membris carent. Oris labrum longè producitur et motus proprios efficit; œsophagi inferior extremitas dentibus munitur. Larva, primò ovöides, sine externis membris, sine oculo; postremò binocula, thoracicis cruribus nullis.
1. Genus—CRYPTOPHIALUS, Darwin. (‘Balanidæ,’ p. 566.)
Cirripedia quibus carapax ad duo separata fila (quæ defigendo inserviunt) redactus est. Corpus ex 1 cephalico, 7 thoracicis, 3 abdominalibus segmentis constat, quæ omnia cirris carent. Os suctorium, mandibuli et maxillæ (dorsis inter se appositis) cucullo includuntur, qui è labro et palpis confluentibus formatur. Metamorphoses incognitæ.
1. Genus—PROTEOLEPAS, Darwin. (‘Balanidæ,’ p. 589.)
[157] In hâc portione Synopsis, Auctorum nomina referunt solummodo ad species.
Sectio A.
Parietes et basis et radii poris perforati.
Sectio B.
Parietes et basis interdum poris perforati; radii nunquam; testæ axis rostro-carinalis elongatus; basis cymbiformis, Gorgoniis et Milleporis affixa.
Sectio C.
Parietes et basis semper poris perforati; radii nunquam.
Sectio D.
Parietes semper poris perforati; basis et radii nunquam.
Sectio E.
Basis membranacea.
Sectio F.
Parietes et radii nunquam, basis interdum, poris perforata; basis interdum admodum tenuis, adeo ut vix distingui possit.
1. Balanus tintinnabulum, Linn. (Darwin, ‘Balanidæ,’ p. 194, Tab. 1 et 2, fig. 1.)
B. testâ à roseâ ad atropurpuream variante, sæpe longitudinaliter virgatâ et costatâ. Orificio plerumque integro, interdum dentato. Scuti cristâ articulari latâ et reflexâ. Tergi margine basali plerumque in contrariis calcaris partibus rectam lineam formante.
Hab.—In tepidis et torridis ubique maribus. Foss. in Europâ.
2. Balanus tulipiformis, Ellis. (Darwin, ‘Balanidæ,’ p. 204, Tab. 2, fig. 2.)
B. testâ, obscurè roseâ, interdum purpurascente; orificio dentato. Scuto externè admodum lævi membranâ tecto. Tergi musculorum depressorum cristis distinctis.
Hab.—In Europâ meridionali et Madeirâ.
3. Balanus psittacus, Molina. (Darwin, ‘Balanidæ,’ p. 206, Tab. 2, fig. 3.)
B. testâ pallidè roseâ, sordidâ; orificio hexagonali. Scuti cristâ articulari minimâ, quæ cum adductoris cristâ, admodum prominente, confluit; musculo depressore laterali tubulari cavitate locato, quæ sursùm ad valvæ apicem extendit. Tergi apice producto, aculeato, purpureo; calcare ab angulo basi-scutali propiùs quam suâ ipsius latitudine distante.
Hab. et foss. in Americâ meridionali.
4. Balanus Capensis, Ellis. (Darwin, ‘Balanidæ,’ p. 209, Tab. 2, fig. 4.)
B. testâ colore lucidè-roseo, umbratâ et sæpe longitudinaliter virgatâ. Scuto ut in B. psittaco. Tergi apice producto aculeato, albo; calcare ab angulo basi-scutali suâ ipsius latitudine distante.
Hab.—In Africâ meridionali.
5. Balanus nigrescens, Lamarck. (Darwin, ‘Balanidæ,’ p. 210, Tab. 2, fig. 5.)
B. testâ cinereâ, pallidè-cæruleo aut atro-cæruleo, aut albo tinctâ. Scuti cristâ articulari parvâ, deorsùm in parvum, acutum aculeum desinente; adductoris cristâ prominente. Tergi apice producto, aculeato.
Hab.—In Australiâ.
6. Balanus decorus, Darwin. (‘Balanidæ,’ p. 212, Tab. 2, fig. 6.)
B. parietibus pallidè-roseis: radiis aliquanto intensioribus. Scuti cristâ articulari parvâ. Tergi sulco longitudinali admodum tenui et aperto; marginibus basalibus utrinque ad calcar declivibus.
Hab.—In Novâ Zealandiâ.
7. Balanus vinaceus, Darwin. (‘Balanidæ,’ p. 213, Tab. 2, fig. 7.)
B. testâ fusco-purpureâ; parietum laminâ interiore cancellatâ. Scuto longitudinaliter et tenuiter striato. Tergi sulco longitudinali tenui et aperto: marginibus basalibus utrinque ad calcar declivibus.
Hab.—In occidentali littore Americæ merid.
8. Balanus Ajax, Darwin. (‘Balanidæ,’ p. 214, Tab. 3, fig. 1.)
B. testâ globoso-conicâ, sæpe in axe rostro-carinali elongatâ, pallidè-roseâ, lævi, admodum crassâ: tubis parietalibus prope marginem basalem, cylindricis et minimis. Scuti cristâ articulari latâ, reflexâ.
Hab.—In Arch. Philippino.
9. Balanus stultus, Darwin. (‘Balanidæ,’ p. 216, Tab. 3, fig. 2.)
B. parietibus et basi porosis: testâ albâ aut purpurâ leviter tinctâ. Scuti margine basali in medio prominente. Tergi sulco longitudinali supernè clauso; calcare angulo basi-scutali non approximato.
Hab.—In Indiâ occident. et orient.
10. Balanus calceolus, Pallas. (Darwin, ‘Balanidæ,’ p. 218, Tab. 3, fig. 3.)
B. parietibus et basi porosis. Scuto musculi depressoris lateralis fossâ parvâ, profundâ.
Hab.—In Africâ occidentali et Indiâ. Foss. in Angliâ.
11. Balanus galeatus, Linn. (Darwin, ‘Balanidæ,’ p. 220, Tab. 3, fig. 4.)
B. parietibus non porosis: basi porosa. Tergi apice, propter cristæ articularis magnitudinem, quadrato.
Hab.—In Americâ sept. et Indiâ occident.
12. Balanus cymbiformis, Darwin. (‘Balanidæ,’ p. 221, Tab. 3, fig. 5.)
B. parietibus et basi non porosis. Scuti et tergi cristis articularibus minimis. Tergo lato, pæne æquilaterali.
Hab.—In Indiâ.
13. Balanus navicula, Darwin. (‘Balanidæ,’ p. 221, Tab. 3, fig. 6.)
B. parietibus et basi non porosis: valvis testæ carino-lateralibus admodùm angustis, latitudine à vertice ad imum pæne æquâ. Radiorum marginibus suturalibus lævibus. Scuto externè longitudinaliter striato.
Hab.—In Indiâ.
14. Balanus trigonus, Darwin. (‘Balanidæ,’ p. 223, Tab. 3, fig. 7.)
B. parietibus costatis, purpureo-rubris, maculatis: orificio lato, trigonali, vix dentato. Scuto crasso, fossularum seriebus longitudinalibus 1 ad 6. Tergo sine sulco longitudinali; calcare truncato ad plenùm 1/3 valvæ latitudine.
Hab.—In Arch. Indiæ Orient.: Australiâ: Californiâ: Peruviâ.
15. Balanus spongicola, Brown. (Darwin, ‘Balanidæ,’ p. 225, Tab. 4, fig. 1.)
B. parietibus plerumque lævibus, interdum longitudinaliter plicatis; roseis; orificio dentato: scuto longitudinaliter striato: tergum, apice producto, sine sulco longitudinali; calcare truncato 1/3 valvæ latitudine.
Hab.—In Europâ meridionali et mediâ, Africâ meridionali; var. in Indiâ occidentali. Foss. in Angliâ, in “Miocenâ Formatione.”
16. Balanus lævis, Bruguière. (Darwin, ‘Balanidæ,’ p. 227, fig. 2.)
B. testâ aut fuscâ membranâ testâ, aut nudâ et albâ, aut pallide-[Pg 614]purpureâ: orificio parvo: radiis minimis: scuto 1 aut 2 sulcis longitudinalibus profundis.
Hab.—In littore occidentali Americæ utriusque.
17. Balanus perforatus, Bruguière. (Darwin, ‘Balanidæ,’ p. 231, Tab. 4, fig. 3, et Tab. 5, fig. 1.)
B. testâ sordidâ, pallidè-purpureâ aut albâ aut cinereâ, lævi aut propter corrosionem longitudinaliter tenuiter costatâ; vaginâ purpureâ; orificio plerumque parvo: radiis plerumque angustis aut nullis: scuto introrsus cristâ brevi minutâ sub cristam adductoris prominentem proximè et parallelè positâ: tergi apice aliquantum producto.
Hab.—In Europâ meridionali et mediâ; in Africâ occidentali.
18. Balanus concavus, Bronn. (Darwin, ‘Balanidæ’ p. 235, Tab. 4, fig. 4.)
B. testâ, albo cum roseo aut obscurè purpureo longitudinaliter striatâ; interdum pure albâ: scuto longitudinaliter tenuiter striato; internè adductoris cristâ admodum aut modicè prominente.
Hab.—In Peruviâ, Californiâ, Arch. Philippino, Australiâ. Et foss. in Europâ, Americâ septent. et meridionali.
19. Balanus amphitrite, Darwin. (‘Balanidæ,’ p. 240, Tab. 5, fig. 2.)
B. testâ purpureo aut roseo longitudinaliter striatâ; striis interdum confluentibus; interdum pure albâ: scutum internè adductoris cristâ prominente.
Hab.—In tepidis et torridis ubique maribus.
20. Balanus pœcilus, Darwin. (‘Balanidæ,’ p. 246, Tab. 5, fig. 3.)
B. testâ obscurè rubrâ albo maculatâ scuto internè adductoris cristâ nullâ tergi calcare præcisè truncato, ferè 1/3 valvæ latitudine.
Hab.—In littore occident. Americæ merid.
21. Balanus eburneus, A. Gould. (Darwin, ‘Balanidæ,’ p. 248, Tab. 5, fig. 4.)
B. testâ flavescente albâ: scuto longitudinaliter tenuiter striato: tergi calcare truncato, margine basi-carinali ferè admodùm excavato; margine carinali supernè prominente.
Hab.—In Americâ septent. et Indiâ occident.
22. Balanus improvisus, Darwin. (‘Balanidæ,’ p. 250, Tab. 6, fig. 1.)
B. testâ albâ: radiis angustis, marginibus superioribus lævibus, leniter arcuatis, admodùm obliquis: tergi sulco longitudinali; calcaris, termino rotundato.
Hab.—In Europâ, et Americæ utriusque littoribus orient. et occident.
23. Balanus nubilus, Darwin. (‘Balanidæ,’ p. 253, Tab. 6, fig. 2.)
B. testâ albâ, irregulari: basi alicubi imperfecte porosâ: scuti cristâ articulari minutâ; adductoris cristâ prominente, fossam profundam[Pg 615] musculo depressori præbente: tergo introrsus purpureâ maculâ majore notato; apice producto, purpureo.
Hab.—In Californiâ.
24. Balanus corrugatus, Darwin. (‘Balanidæ,’ p. 254, Tab. 6, fig. 3.)
B. testâ albâ longitudinaliter plicatâ: radiis angustis: scuto internè sine adductoris cristâ.
Foss. in Italiâ.
25. Balanus porcatus, Da Costa. (Darwin, ‘Balanidæ,’ p. 256, Tab. 6, fig. 4.)
B. testâ albâ, plerumque longitudinaliter acutè costatâ: radiorum marginibus superioribus pæne basi parallelis: scuto longitudinaliter striato; tergi apice producto, purpureo.
Hab. et foss. in Europâ, et Americâ septent. et regionibus Arcticis.
26. Balanus patellaris, Spengler. (Darwin, ‘Balanidæ,’ p. 259, Tab. 6, fig. 5.)
B. testâ depressâ fuscâ, plerumque obscuro-violaceo longitudinaliter striatâ: radiorum marginibus superioribus (in adultis speciminibus) rotundatis, superficie sulcis tenuibus basi parallelis: basi interdùm poris imperfectis perforatâ: scuto internè cum adductoris cristâ.
Hab.—In Indiâ et Arch. Philippino.
27. Balanus crenatus, Bruguière. (Darwin, ‘Balanidæ,’ p. 261, Tab. 6, fig. 6.)
B. testâ albâ: radiorum marginibus superioribus obliquis, asperis, rectis: scuto sine adductoris cristâ: tergi calcare rotundato.
Hab. et foss. in Europâ, et Americâ septent. et regionibus Arcticis, et Indiâ occident. et Africâ meridionali.
28. Balanus glandula, Darwin. (‘Balanidæ,’ p. 265, Tab. 7, fig. 1.)
B. testâ albâ; parietum laminâ internâ longitudinaliter et fortiter costatâ, poris imperfectis et minutis, interdùm alicubi nullis: radiis angustis marginibus superioribus rotundatis: scuto cum adductoris cristâ: tergi calcare truncato, rotundato.
Hab.—In Californiâ, et Oceano Pacifico meridion.
29. Balanus balanoides, Linn. (Darwin, ‘Balanidæ,’ p. 267, Tab. 7, fig. 2.)
B. parietibus aut solidis, aut cancellatis, aut (rarò) unicâ pororum serie formatis: tergi calcare obtuso aut acuto.
Hab.—In Europâ et Americâ septent. et regionibus Arcticis.
30. Balanus cariosus, Pallas. (Darwin, ‘Balanidæ,’ p. 273, Tab. 7, fig. 3.)
B. parietibus crassis, pluribus seriebus pororum inæqualium formatis; tergo angusto, apice rostrato, calcare acuminato.
Hab.—In littore occident. Americæ septent. et Freto Behringi et Insulis Kuriliis.
31. Balanus declivis, Darwin. (‘Balanidæ,’ p. 275, Tab. 7, fig. 4.)
B. parietibus solidis: rostro, carinâ et valvis carino-lateralibus pæne duplo longiore, basi igitur obliquâ: tergi calcare truncato, 1/2 valvæ latitudine.
Hab.—In Indiâ occident.
32. Balanus Hameri, Ascanius. (Darwin, ‘Balanidæ,’ p. 277, Tab. 7, fig. 5.)
B. testâ albâ: radiorum marginibus superioribus obliquis, lævibus, arcuatis; aciebus suturalibus lævibus: basi solidâ: scuto longitudinaliter, debiliter striato: tergi calcare angusto.
Hab. et foss. in Europâ septent. et Americâ septent.
33. Balanus amaryllis, Darwin. (‘Balanidæ,’ p. 279, Tab. 7, fig. 6.)
B. testâ subroseo-purpureo striatâ, aut obnubilatâ; interdùm purè albâ: radiis angustis, marginibus superioribus obliquis lævibus, arcuatis: basi porosâ: scuto longitudinaliter planè striato: tergi calcare angusto.
Hab.—In Indiâ, et Arch. Indiæ orient. et Australiâ septent.
34. Balanus allium, Darwin. (‘Balanidæ,’ p. 281, Tab. 7, fig. 7.)
B. testâ pallidè purpuratâ, radiis latis, marginibus superioribus non obliquis: basi porosâ: scuto lineis incrementi crenatis: tergi calcare admodum brevi, truncato, 1/2 valvæ latitudine.
Hab.—In Australiâ septent.
35. Balanus cepa, Darwin. (‘Balanidæ,’ p. 283, Tab. 7, fig. 8.)
B. testâ sordidè rubro-purpureâ, abruptè conicâ: radiis angustis: basi obscurè porosâ: scuto lineis incrementi crenatis: tergi calcare truncato, dimidiâ valvæ latitudine, et infrà angulum basi-scutalem dependente, usque ad 1/2 sui ipsius latitudinem.
Hab.—In Japaniâ.
36. Balanus quadrivittatus, Darwin. (‘Balanidæ,’ p. 284, Tab. 8, fig. 1.)
B. testâ abruptè conicâ, quatuor vittis longitudinalibus transversè positis: radiorum marginibus superioribus obliquis: basi tenui, solidâ: scuto, lineis incrementi lævibus; musculi depressoris lateralis fossâ distinctâ nullâ: terga ut in B. cepâ.
Hab.—In Archipel. Indiæ orient.
37. Balanus terebratus, Darwin. (‘Balanidæ,’ p. 285, Tab. 8, fig. 2.)
B. testâ albâ, costis longitudinalibus fortibus, margine basali in longos aculeos producto: basi concavâ non porosâ, lineis radiantibus fortiter costatâ; spatio intercostali foraminibus minutis rotundatis sæpe duplici serie, cribrosè perforato.
Hab.—(?)
38. Balanus vestitus, Darwin. (‘Balanidæ,’ p. 286, Tab. 8, fig. 3.)
B. testâ subroseo-purpureâ aut albâ, membranâ aurantiacâ vestitâ:[Pg 617] radiorum loco, meris fissuris: basi solidâ: scuto, cristâ adductoris acutâ curvatâ; musculi depressoris cristæ adsunt: tergi calcare brevi, truncato, 1/3 valvæ latitudine.
Hab.—In Australiâ et Novâ Zealandiâ.
39. Balanus imperator, Darwin. (‘Balanidæ,’ p. 288, Tab. 8, fig. 4.)
B. testâ internè imperatorio-purpureâ: parietibus crassis margine interno basali spinis et cristis irregularibus aspero: radiis angustis: basi admodùm tenui, solidâ: scuto cum musculorum depressorum rostralium et lateralium cristis: tergi calcare ad finem rotundato.
Hab.—In Australiâ.
40. Balanus flosculus, Darwin. (‘Balanidæ,’ p. 290, Tab. 8, fig. 5.)
B. testâ purpureâ aut sordide-albâ, parietum margine basali spinis et cristis irregularibus asperâ: radiis angustis aut nullis: basi quam tenuissimâ, ad speciem nullâ: scuto cum musculi depressoris lateralis cristis: tergo admodùm angusto, calcare acuminato.
Hab.—In Americæ merid. littore occid.
41. Balanus bisulcatus, Darwin. (‘Balanidæ,’ p. 293, Tab. 8, fig. 6.)
B. radiorum marginibus superioribus obliquis, lævibus; aciebus suturalibus lævibus: basi poris magnis perforatâ: scuto angusto, sulcis longitudinalibus 2 ad 4: tergi calcare brevissimo dimidiâ valvæ latitudine.
Foss. in Angliâ, et Galliâ et Belgio. “Miocenâ Formatione.”
42. Balanus dolosus, Darwin. (‘Balanidæ,’ p. 295, Tab. 8, fig. 7.)
B. radiorum marginibus superioribus obliquis lævibus: aciebus suturalibus item lævibus: basi poris magnis perforatâ: tergi calcare non admodum brevi, 1/3 valvæ latitudine.
Foss. in Angliâ. “Pliocenâ, et miocenâ Formationibus.”
43. Balanus unguiformis, J. de C. Sowerby. (Darwin, ‘Balanidæ,’ p. 296, Tab. 8, fig. 8.)
B. parietibus tenuibus, interdum poris perforatis: radiorum marginibus superioribus obliquis; aciebus suturalibus tenuissimè crenatis: basi solidâ: tergi calcare angusto, obtuso.
Foss. in Angliâ et Belgio. “Eocenâ Formatione.”
44. Balanus varians, Darwin. (‘Balanidæ,’ p. 298, Tab. 8, fig. 9.)
B. parietibus modicè crassis; radiorum marginibus superioribus perobliquis; aciebus suturalibus pæne lævibus aut tenuiter crenatis: basi tenuiter porosâ: tergi calcare, parvo, angusto, obtuso.
Foss. in Patagoniâ.
45. Balanus inclusus, Darwin. (‘Balanidæ,’ p. 299, Tab. 8, fig. 10.)
B. testâ rufo-fuscâ: radiis latis, marginibus superioribus aut non obliquis aut modicè; aciebus suturalibus cum septis planè denticulatis: basi porosâ: scuto sine adductoris cristâ: tergi calcare subangusto.
Foss. in Angliâ et Germaniâ. “Miocenâ Formatione.”
1. Acasta spongites, Poli. (Darwin, ‘Balanidæ,’ p. 308, Tab. 9, fig. 1.)
A. parietibus carino-lateralibus ferè 1/6 parietum lateralium latitudine: superficie internâ parietum plerumque debiliter costatâ: scuti cristâ articulari ad extremitatem inferiorem abruptè præcisâ: tergi calcare truncato rotundato, ferè 1/3 valvæ latitudine.
Hab.—In Europâ et Africâ meridion.
2. Acasta sulcata, Lamarck. (Darwin, ‘Balanidæ,’ p. 310, Tab. 9, fig. 2.)
A. parietibus carino-lateralibus ferè 1/6 parietum lateralium latitudine: parietum superficie internâ plerumque fortiter costatâ: basis acie fortiter crenatâ: testæ orificio sub-exiguo: tergi calcare plerumque truncato, pæne 1/2 valvæ latitudine.
Hab.—In Australiâ.
3. Acasta cyathus, Darwin. (‘Balanidæ,’ p. 312, Tab. 9, fig. 3.)
A. parietibus carino-lateralibus ferè 1/4 parietum lateralium latitudine: radiis latioribus quam parietes: basi pæne planâ, exiguâ: tergi calcare truncato, 1/2 valvæ latitudine.
Hab.—In Madeirâ et Indiâ occid.
4. Acasta undulata, Darwin. (‘Balanidæ,’ p. 313, Tab. 9, fig. 4.)
A. testâ, ad speciem, ut in “A. spongites,” sed majore: scuto externe striis longitudinalibus, sæpe binis, signato, sulcis intermediis latioribus: tergi calcare, pæne 1/2 valvæ latitudine.
Foss. in Angliâ. “Miocenâ Formatione.”
5. Acasta glans, Lamarck. (Darwin, ‘Balanidæ,’ p. 314, Tab. 9, fig. 5.)
A. parietibus internè per-lævibus, marginibus lateralibus uniuscujusque testæ valvæ intùs prominentibus: basis acie rarò crenatâ, sed 6 dentibus introrsùs prominentibus instructâ: scuto longitudinaliter fortiter striato.
Hab.—In Australiâ.
6. Acasta lævigata, J. E. Gray. (Darwin, ‘Balanidæ,’ p. 315, Tab. 9, fig. 6.)
A. parietibus intùs per-lævibus, marginibus lateralibus uniuscujusque testæ valvæ internùs prominentibus: basis acie fortiter crenatâ, et 6 dentibus introrsùs prominentibus instructâ: scuto aut longitudinaliter tenuiter striato aut lævi.
Hab.—In Mari Rubro et Archipel. Philippino.
7. Acasta fenestrata, Darwin. (‘Balanidæ,’ p. 316, Tab. 9, fig. 7.)
A. testâ subrufâ, foraminibus magnis membranâ-tectis inter suturas suprà basin proximis, 6; parietibus carino-lateralibus 1/2 parietum latera[Pg 619]lium latitudine: intùs, parietibus et basis acie lævibus: tergi cristâ articulari brevi prominente; calcare acuminato.
Hab.—In Archipel. Philippino.
8. Acasta purpurata, Darwin. (‘Balanidæ,’ p. 318, Tab. 9, fig. 8.)
A. testâ obscurè cæruleo-purpureâ, foraminibus parvis, membrana-tectis inter suturas suprà basin proximis, 6: tergi cristâ articulari per-brevi prominente; calcare admodum lato rotundato.
Hab.—In Sumatrâ et Archipel. Philippino.
9. Acasta sporillus, Darwin. (‘Balanidæ,’ p. 319, Tab. 9, fig. 9.)
A. testâ purpureo-fuscâ, parietibus intùs fortiter costatis et reticulatis: testæ valvis carino-lateralibus angustissimis, ad basin non extendentibus.
Hab.—In Sooloo Insulis, in Archipelago Indico orient.
1. Tetraclita porosa, Linn. (Darwin, ‘Balanidæ,’ p. 328, Tab. 10, fig. 1.)
T. radiis (si qui rarò adsunt) angustis; etiam suturis sæpe nullis: testâ abruptè conicâ, superficie plerumque corrosâ, stalactitiferam speciem exhibente.
Hab.—In torridis et tepidis ubique maribus.
2. Tetraclita serrata, Darwin. (‘Balanidæ,’ p. 333, Tab. 10, fig. 2.)
T. testâ obscurè viridi-griseâ, costis longitudinalibus serratis angustis: radiis nullis: scuti cristis (articulari et adductoris) cavitatem usque ad apicem valvæ extendentem formantibus.
Hab.—In Africâ merid.
3. Tetraclita rosea, Krauss. (Darwin, ‘Balanidæ,’ p. 335, Tab. 10, fig. 3.)
T. testâ sordidè albâ, subroseo tinctâ; parietibus è serie unicâ magnarum tubarum formatis: radiis plerumque angustis: tergi calcare sub-brevi, lato.
Hab.—In Australiâ, et Africâ meridion.
4. Tetraclita purpurascens, W. Wood. (Darwin, ‘Balanidæ,’ p. 337, Tab. 11, fig. 1.)
T. testâ depressâ, pallidè purpureâ aut sordidè albâ, superficie longitudinaliter costatâ, aut corrosâ et granulatâ: radiis, aut etiam suturis, nullis, aut radiis planè formatis et latis, marginibus superioribus basi parallelibus: basi membranaceâ: scuto transversè elongato: tergo exiguo, calcare per-brevi, rotundato.
Hab.—In Australiâ.
5. Tetraclita costata, Darwin. (‘Balanidæ,’ p. 339, Tab. 11, fig. 2.)
T. testâ depressâ sub-albâ, plerumque costis longitudinalibus admodùm prominentibus, 10: radiis latis, marginibus superioribus basi[Pg 620] parallelis: basi calcareâ: scuto externè longitudinaliter striato: tergi calcare brevi rotundato.
Hab.—In Archipel. Philippino.
6. Tetraclita vitiata, Darwin. (‘Balanidæ,’ p. 340, Tab. 11, fig. 3.)
T. testâ albâ, parte superiore plerumque roseo tinctâ; superficie irregulari; tubis parietalibus admodum irregularibus; radiis modicè latis, marginibus superioribus obliquis; alarum aciebus suturalibus percrassis crenatis: tergi calcare cum angulo basi-scutali non juncto; calcaris termino æquabiliter rotundato.
Hab.—In Australiâ septentr. et orient. et Archipel. Philippino.
7. Tetraclita cœrulescens, Spengler. (Darwin, ‘Balanidæ,’ p. 342, Tab. 11, fig. 4.)
T. testæ parte superiore viridi-cæruleo tinctâ, longitudinaliter costatâ; radiis modice latis, marginibus superioribus obliquis: scuto adductoris cristâ exiguâ, articulari admodùm prominente; ambabus ita conjunctis ut cavitatem parvam subcylindricam efficiant: tergi calcare cum angulo basi-scutali non juncto.
Hab.—In Archipel. Philippino et Oceano Pacifico.
8. Tetraclita radiata, De Blainville. (Darwin, ‘Balanidæ,’ p. 343, Tab. 11, fig. 5.)
T. testâ albâ costis longitudinalibus approximatis multis; radiis latis, marginibus superioribus leviter obliquis, intus porosis: tergi cristâ articulari suprà modum prominente; calcare cum angulo basi-scutali non juncto.
Hab.—In Indiâ occid. et Australiâ: sæpe navigiis suffixa.
1. Elminius Kingii, J. E. Gray. (Darwin, ‘Balanidæ,’ p. 348, Tab. 11, fig. 6.)
E. testâ lævi; griseâ aut sordidè albâ; radiis latis, marginibus et aciebus lævibus: scuto, sine adductoris cristâ: tergi calcare ab angulo basi-scutali distincto: scuto et tergo interdum inter se calcifactis.
Hab.—In Americâ merid.
2. Elminius modestus, Darwin. (‘Balanidæ,’ p. 350, Tab. 12, fig. 1.)
E. testâ longitudinaliter plicatâ, subviridi aut albâ; radiis modicè latis, marginibus et aciebus lævibus: scuto sine adductoris cristâ: tergo angusto, calcare cum angulo basi-scutali confluente.
Hab.—In Australiâ.
3. Elminius plicatus, J. E. Gray. (Darwin, ‘Balanidæ,’ p. 351, Tab. 12, fig. 2.)
E. testâ longitudinaliter profundè plicatâ, corrosâ, alicubi aurantiacâ; radiis per-angustis, aciebus sinuosis, leviter dentatis: scuto cum adductoris cristâ.
Hab.—In Novâ Zealandiâ.
4. Elminius simplex, Darwin. (‘Balanidæ,’ p. 353, Tab. 12, fig. 3.)
E. testâ longitudinaliter costatâ, sordidè albâ; radiis angustissimis, marginibus et aciebus lævibus: scuto cum adductoris cristâ.
Hab.—In Australiâ.
1. Pyrgoma Anglicum, Leach. (Darwin, ‘Balanidæ,’ p. 360, Tab. 12, fig. 4.)
P. testâ abruptè conicâ, purpureo-rubrâ; orificio ovato, angusto; basi porosâ, plerumque è coralio exsertâ: scuto et tergo subtriangularibus.
Hab.—In Magnâ Britanniâ et Europâ meridionali, et Insulis “Cape de Verde.”
2. Pyrgoma Stokesii, Darwin. (‘Balanidæ,’ p. 361, Tab. 12, fig. 6.)
P. testâ modicè conicâ, pallidè purpureo-rubrâ; orificio ovato; basi non-porosâ, in coralium profunde conditâ: scuto et tergo subtriangularibus.
Hab.—In Indiâ occid.
3. Pyrgoma cancellatum, Leach. (Darwin, ‘Balanidæ,’ p. 362, Tab. 12, fig. 5.)
P. testâ circumferentiâ plerumque lobatâ: scuto elongato, adductoris cristâ longè infrà marginem basalem descendente, et ad rostralem finem in acumen quadratum producto: tergi calcare valvæ partem superiorem longitudine quadruplo superante.
Hab.—In Archip. Indico. orient. (?)
4. Pyrgoma conjugatum, Darwin. (‘Balanidæ,’ p. 364, Tab. 12, fig. 7.)
P. testâ pæne planâ, cristis approximatis radiantibus: scuto et tergo inter se sine suturâ calcifactis; scuto, adductoris cristâ infrà marginem basalem descendente, et ad terminum rostralem in acumen producto; tergi calcare superioris valvæ partis longitudinem ferè æquante.
Hab.—In Mari Rubro.
5. Pyrgoma grande, G. B. Sowerby, jun. (Darwin, ‘Balanidæ,’ p. 365, Tab. 13, fig. 1.)
P. testâ modicè convexâ, pæne lævi: scuto et tergo inter se sine suturâ calcifactis; scuto limbo occludente parvo instructo, adductoris cristâ infrà marginem basalem descendente: tergo quadrato, calcare nullo.
Hab.—In Archip. Indico orient.
6. Pyrgoma milleporæ, Darwin. (‘Balanidæ,’ p. 367, Tab. 13, fig. 2.)
P. testæ orificio angustè-ovato; vaginâ obscurè purpureâ: scuto multùm elongato: tergo triangulo, convexo, calcare nullo.
Hab.—In Archip. Philippino.
7. Pyrgoma dentatum, Darwin. (‘Balanidæ,’ p. 369, Tab. 13, fig. 3.)
P. scuto multùm elongato, cum prominentiâ articulari dentiformi:[Pg 622] tergo convexo, irregulariter triangulo, interdum cum calcare imperfecto, et in superficie internâ dente introrsùs prominente; scuto et tergo limbo occludente instructis.
Hab.—In Mari Rubro.
8. Pyrgoma crenatum, G. B. Sowerby. (Darwin, ‘Balanidæ,’ p. 370. Tab. 13, fig. 4.)
P. scuto multùm elongato, adductoris cristâ infra marginem basalem reflexum descendente: tergi calcare lato depresso; scuto et tergo limbo occludente instructis.
Hab.—In Archip. Ind. orient et Philippino.
9. Pyrgoma monticulariæ, J. E. Gray. (Darwin, ‘Balanidæ,’ p. 372. Tab. 13, fig. 5.)
P. testâ irregulari, margine exteriore aspero; orificio minuto rotundo: scuto et tergo multum elongatis, inter se sine suturâ calcifactis, limbo occludente lato instructis.
Hab.—In Archip. Indico orient.
1. Creusia spinulosa, Leach, (Darwin, ‘Balanidæ,’ p. 376, Tab. 13, fig. 6, et Tab. 14, fig. 6.)
Hab.—In Maribus torridis.
1. Chelonobia testudinaria, Linn. (Darwin, ‘Balanidæ,’ p. 392, Tab. 14, fig. 1 et fig. 5, Tab. 15, fig. 1).
Ch. testâ conicâ, depressâ, gravi; radiis subangustis, depressis plerumque utrinque scrobiculatis.
Hab.—In tepidis et torridis ubique Maribus; testudinibus affixa.
2. Chelonobia caretta, Spengler. (Darwin, ‘Balanidæ,’ p. 394, Tab. 14, fig. 2.)
Ch. testâ globoso-convexâ, crassissimâ, gravissimâ; parte superiore tritâ, substriatâ; radiis aut non formatis aut admodum angustis; parietibus sine cavitatibus sursùm inter septa interrupta radiantia extendentibus.
Hab.—In Africâ occid. et Australiâ septent.; testudinibus affixa.
3. Chelonobia patula, Ranzani. (Darwin, ‘Balanidæ,’ p. 396, Tab. 14, fig. 3).
Ch. testâ abruptè conicâ, admodum lævi et tenui; orificio magno, ferè dimidium diametri basalis testæ superante; radiis latis, lævibus, leviter depressis.
Hab.—Mediterraneo; Africâ occid.; Indiâ occid.; Australiâ Crustaceis et molluscarum testis lævibus affixa.
1. Coronula balænaris, Linn. (Darwin, ‘Balanidæ,’ p. 415, Tab. 15, fig. 2, Tab. 16, fig. 3 et 5.)
B. testâ multùm depressâ, costis longitudinalibus planatis, aciebus earum simplicibus; orificio hexagono-rotundato; radiis testæ ipsius crassitudinem pænè æquantibus; valvis opercularibus, 4.
Hab.—In Oceano meridionali.
2. Coronula diadema, Linn. (Darwin, ‘Balanidæ,’ p. 417, Tab. 15, fig. 3.)
C. testâ coroniformi, costis longitudinalibus convexis, aciebus earum crenatis; orificio hexagono; radiis modicè crassis, admodum latis; tergis nullis aut rudimentariis.
Hab.—In Oceano septentrionali.
3. Coronula reginæ, Darwin. (‘Balanidæ,’ p. 419, Tab. 15, fig. 5.)
C. testâ globoso-conicâ aut depressâ, costis longitudinalibus multùm planatis, aciebus earum crenatis, superficie striatâ, granulatâ; orificio hexagono; radiis tenuibus 1/5 testæ valvæ crassitudinem non superantibus; tergis nullis.
Hab.—In Oceano Pacifico.
4. Coronula barbara, Darwin. (‘Balanidæ,’ p. 421, Tab. 15, fig. 6).
C. testâ (probabiliter) coroniformi, costis longitudinalibus convexis, aciebus earum crenatis, superficie internâ et externâ cristis transversis asperâ; radiis modicè crassis; spatio inter radios et alas solidè impleto.
Foss.—In Angliâ, “Red Crag Formatione.”
1. Platylepas bissexlobata, De Blainville. (Darwin, ‘Balanidæ,’ p. 428, Tab. 17, fig. 1.)
P. testâ lineis transversis incrementi conspicuis; parietibus porosis; vaginâ ad dimidium parietum vix descendente.
Hab.—In Mediterraneo; Africâ occid.; Indiâ occid.; Australiâ septentrionali.
2. Platylepas decorata, Darwin. (‘Balanidæ,’ p. 429, Tab. 17, fig. 2.)
P. testâ costis longitudinalibus tenuibus, parte earum inferiore globulis minutis decoratâ; parietibus non porosis; membranâ basali pari cum testâ convexitate.
Hab.—In Oceano Pacifico.
1. Tubicinella trachealis, Shaw. (Darwin, ‘Balanidæ,’ p. 431, Tab. 17, fig. 3).
Hab.—In Oceano Pacifico meridionali.
1. Xenobalanus globicipitis, Steenstrup. (Darwin, ‘Balanidæ,’ p. 440, Tab. 17, fig. 4.)
Hab.—In Oceano Atlantico septentrionali.
1. Chthamalus stellatus, Poli. (Darwin, ‘Balanidæ,’ p. 455, Tab. 18, fig. 1.)
Ch. testâ albâ aut griseâ, plerumque multùm corrosâ et punctâ; radiis (si qui adsunt) angustis, aciebus suturalibus tenuissimè crenatis; tergo, musculi depressoris cristis infrà marginem basalem vix dependentibus.
Hab.—In Europâ; Americâ sept.; Braziliâ; Mari Rubro; Chinâ; Californiâ septent.
2. Chthamalus antennatus, Darwin. (‘Balanidæ,’ p. 460, Tab. 18, fig. 2.)
Ch. testâ conicâ, plerumque lævi; nisi profundè corrosâ, pallidè sordidè carneâ; suturis semper distinctis; radiis, si qui adsunt, aciebus suturalibus per-lævibus.
Hab.—In Australiâ.
3. Chthamalus cirratus, Darwin. (‘Balanidæ,’ p. 461, Tab. 18, fig. 4.)
Ch. testâ albâ aut griseâ; vaginâ et valvis opercularibus membranâ fimbriatâ plerumque vestitis; tergo, angulo basi-carinali dependente et acuminato.
Hab.—In Chiliâ, Peruviâ.
4. Chthamalus fissus, Darwin. (‘Balanidæ,’ p. 462, Tab. 18, fig. 6.)
Ch. testâ subfuscâ, plicatâ; orificio longitudine duplo majore quam latitudo; tergo triangulo, æquilaterali, marginibus basalibus et carinalibus leviter prominentibus.
Hab.—In Californiâ.
5. Chthamalus dentatus, Krauss. (Darwin, ‘Balanidæ,’ p. 463, Tab. 18, fig. 3).
Ch. testâ sordidè albâ, aut subfuscâ; suturis, dentibus inter se concurrentibus formatis; tergi margine carinali prominente.
Hab.—In Africâ.
6. Chthamalus Hembeli, Conrad. (Darwin, ‘Balanidæ,’ p. 465, Tab. 18, fig. 5.)
Ch. testâ obscurè rubro-purpureâ; suturis aut obliteratis aut dentibus inter se concurrentibus formatis; basi interdum per parietum inflectionem calcareâ factâ; scuto, sulcis duobus vel tribus ad mediam valvam deorsùm extendentibus.
Hab.—In Californiâ.
7. Chthamalus intertextus, Darwin. (‘Balanidæ,’ p. 467, Tab. 19, fig. 1.)
Ch. testâ (si benè conservetur) violaceo-purpureâ; suturis aut obli[Pg 625]teratis aut ex interplicatis laminis obliquis formatis; basi membranaceâ, sed cingente projecturâ ex parietum marginibus basalibus inflexis formatâ: scuto et tergo inter se prorsus calcifactis.
Hab.—In Archip. Philippino.
8. Chthamalus scabrosus, Darwin. (‘Balanidæ,’ p. 468, Tab. 19, fig. 2.)
Ch. testâ (si benè conservetur) obscurè purpureo-fuscâ; suturis ex interplicatis laminis obliquis (sed raro benè) formatis; tergo, cum musculi depressoris fossâ profundâ angustâ, ad angulum basi-carinalem.
Hab.—In Americæ merid. littore merid. et occid.
1. Chamæsipho columna, Spengler. (Darwin, ‘Balanidæ,’ p. 470, Tab. 19, fig. 3.)
Ch. suturis, nisi in recenter natis, plerumque et internè et externè obliteratis; tergo, cum depressoris musculi fossis parvis.
Hab.—In Australiâ.
2. Chamæsipho scutelliformis, Darwin. (‘Balanidæ,’ p. 472, Tab. 19, fig. 4.)
Ch. rostro per-exiguo, elongato, triangulo: testæ valvarum lateralium unâquæque cum foramine, et carinâ cum duobus similibus foraminibus; quæ 4 in columnas tubulares testaceas conducunt.
Hab.—In Chinâ (?)
1. Pachylasma giganteum, Philippi. (Darwin, ‘Balanidæ,’ p. 477, Tab. 19, fig. 5.)
P. testâ et operculo sordide albis; testæ valvâ laterali et carino-laterali alis similibus instructâ.
Hab.—In Mari Mediterraneo.
2. Pachylasma aurantiacum, Darwin. (‘Balanidæ,’ p. 480, Tab. 20, fig. 1.)
P. testâ aurantiaco tinctâ; ad speciem externam, quasi è 4 solùm testæ valvis formatâ, ideò quòd laterales, et carino-laterales valvæ utrinque obscurâ fissurâ solùm separantur.
Hab.—In Australiâ.
1. Octomeris angulosa, G. B. Sowerby. (Darwin, ‘Balanidæ,’ p. 483, Tab. 20, fig. 2.)
O. testâ sordidè albâ, asperâ, solidâ; alis crassis, aciebus suturalibus obtusè crenatis.
Hab.—In Africâ merid.
2. Octomeris brunnea, Darwin. (‘Balanidæ,’ p. 484, Tab. 20, fig. 3.)
O. testâ rubro-fuscâ depressâ, tenui, longitudinaliter sulcis tenuibus signatâ; tergi margine basali leviter in angulum flexâ.
Hab.—In Archip. Philippino.
1. Catophragmus polymerus, Darwin. (‘Balanidæ,’ p. 487, Tab. 20, fig. 4.)
C. basi membranaceâ: sine appendicibus caudalibus.
Hab.—In Australiâ.
2. Catophragmus imbricatus, G. B. Sowerby. (Darwin, ‘Balanidæ,’ p. 490.)
C. basi calcareâ: cum appendicibus caudalibus.
Hab.—In Indiâ occident.
1. Verruca Strömia, Müller. (Darwin, ‘Balanidæ,’ p. 518, Tab. 21, fig. 1.)
V. scuto mobili, cristâ articulari inferiore dimidiam brevis cristæ articularis superioris latitudinem non æquante: testâ plerumque longitudinaliter sulcatâ.
Hab.—In Europâ; Mari Rubro.
2. Verruca lævigata, G. B. Sowerby. (Darwin, ‘Balanidæ,’ p. 520, Tab. 21, fig. 3.)
V. scuto mobili, cristâ articulari inferiore latiore quam brevis crista articularis superior; tergo mobili, latitudine majore quam altitudo, cristâ articulari superiore in terminum subacutum producto.
Hab.—In Americâ merid.
3. Verruca Spengleri, Darwin. (‘Balanidæ,’ p. 521, Tab. 21, fig. 2.)
Scuto mobili, cum adductoris musculi cristâ acutâ, rectâ, mediali; scuto immobili non majore quam immobile tergum.
Hab.—In Insulâ Madeirâ.
4. Verruca nexa, Darwin. (‘Balanidæ,’ p. 522, Tab. 21, fig. 5.)
V. testâ sub-rubrâ; scuto mobili cum cristis externis longitudinalibus fortiter prominentibus, præter cristas articulares; scuto immobili majore quam carina, adductoris musculi cristâ vel laminâ distinctâ nullâ.
Hab.—In Indiâ occid.
5. Verruca prisca, Bosquet. (Darwin, ‘Balanidæ,’ p. 525, Tab. 21, fig. 4.)
V. testâ lævi; scuti mobilis cristâ articulari inferiore aliquanto latiore quam superior.
Foss.—In Cretaceâ Formatione.
1. Lepas anatifera, Linn. (Darwin, ‘Lepadidæ,’ p. 73, Tab. 1, fig. 1, var.)
L. valvis aut lævibus aut delicate striatis; è duobus scutis, dextro[Pg 627] solùm dente interno umbonali instructo; pedunculi parte superiore fuscâ.
Hab.—In Oceano, pæne ubique.
2. Lepas Hillii, Leach. (Darwin, ‘Lepadidæ,’ p. 77, Tab. 1, fig. 2.)
L. valvis lævibus; scutorum dentibus internis umbonalibus nullis; carinâ à cæteris valvis, furcâ etiam a scutorum basali margine, paululum distante; pedunculi parte superiore aut pallidâ aut aurantiacâ.
Hab.—In Oceano, pæne ubique.
3. Lepas anserifera, Linn. (Darwin, ‘Lepadidæ,’ p. 81, Tab. 1, fig. 4.)
L. valvis approximatis leviter sulcatis (tergis præcipuè); scuto dextro dente forti interno umbonali, lævo aut dente exiguo, aut merâ cristâ instructo; margine occludente arcuato, prominente; pedunculi parte superiore aurantiacâ.
Hab.—In Oceano, pæne ubique.
4. Lepas pectinata, Spengler. (Darwin, ‘Lepadidæ,’ p. 85, Tab. 1, fig. 3.)
L. valvis tenuibus, crassè sulcatis, sæpe pectinatis; scutorum cristâ prominente ab umbone ad apicem, juxta marginem occludentem, pertinente; furcæ carinalis cruribus inter angulos 135° et 180° divergentibus.
Hab.—In Oceano Atlantico, sept. et meridionali.
5. Lepas australis, Darwin. (‘Lepadidæ,’ p. 89, Tab. 1, fig. 5.)
L. valvis glabris, tenuibus, fragilibus; scutorum dentibus umbonalibus utrinque internis; carinæ parte superiore latâ, planâ, suprà furcam valdè constrictâ; furcæ cruribus latis, planis, tenuibus, acuminatis, intermedio margine non reflexo.
Hab.—In Oceano meridionali.
6. Lepas fascicularis, Ellis. (Darwin, ‘Lepadidæ,’ p. 92, Tab. 1, fig. 6.)
L. valvis glabris, tenuibus, pellucidis; carinâ rectangulè flexâ, parte inferiore in discum planum oblongum expansâ.
Hab.—In Oceano torrido et tepido, pæne ubique.
1. Pœcilasma Kæmpferi, Darwin. (‘Lepadidæ,’ p. 102, Tab. 2, fig. 1.)
P. valvis 5; carinæ basi truncatâ et cristatâ: scutorum dentibus internis umbonalibus fortibus: tergorum acumine basali truncato, margini occludenti pæne parallelo.
Hab. in Japaniâ.
2. Pœcilasma aurantia, Darwin. (‘Lepadidæ,’ p. 105, Tab. 2, fig. 2.)
P. valvis 5; carinæ basi truncatâ: scutis ovatis, margine basali per-brevi, dentibus parvis, internis, umbonalibus instructo: tergorum acumine basali perobliquè truncato.
Hab.—In Insulâ Madeirâ.
3. Pœcilasma crassa, J. E. Gray. (Darwin ‘Lapadidæ,’ p. 107, Tab. 2, fig. 3.)
P. valvis 5; carinæ termino basali in discum parvum infossum producto: scutis convexis, dentibus internis umbonalibus nullis: tergis pæne rudimentalibus, vix carinâ latioribus.
Hab.—In Insulâ Madeirâ.
4. Pœcilasma fissa, Darwin. (‘Lepadidæ,’ p. 109, Tab. 2, fig. 4.)
P. valvis 7; scuto utroque è duobus juxtapositis segmentis formato; segmento altero intus dentato: tergis brevibus, ter aut quater carinâ latioribus: carinæ termino basali in discum parvum angustum infossum producto.
Hab.—In Archipel. Philippino.
5. Pœcilasma eburnea, Hinds. (Darwin, ‘Lepadidæ,’ p. 112, Tab. 2, fig. 5.)
P. valvis 3; scutis acuminatis, ovatis; ad pedunculum pæne transversè spectantibus; dentibus internis umbonalibus fortibus: tergis nullis carinæ termino basali in discum amplum oblongum infossum producto.
Hab.—In Novâ Guineâ.
1. Dichelaspis Warwickii, J. E. Gray. (Darwin, ‘Lepadidæ,’ p. 120, Tab. 2, fig. 6.)
D. scutorum segmento basali duplo latiore quam segmentum occludens: tergorum parte inferiore paulò, latiore quam occludens scutorum segmentum.
Hab.—In Archipel. Indico orientali.
2. Dichelaspis Grayii, Darwin. (‘Lepadidæ,’ p. 123, Tab. 2, fig. 9.)
D. scutorum segmento basali angustiore quam segmentum occludens; longitudine pæne dimidiâ: tergis bipenniformibus, margine crenato, spinâ posticâ, manubrio angustiore quam occludens scutorum segmentum.
Hab.—In Oceano torrido, aut Indico, aut Pacifico.
3. Dichelaspis pellucida, Darwin. (‘Lepadidæ,’ p. 125, Tab. 2, fig. 7.)
D. valvarum singularum acuminibus superioribus et inferioribus vix intersecantibus: scutorum segmento basali multo angustiore quam segmentum occludens; longitudine ferè dimidia: tergis bipenniformibus, margine integro, manubrii acumine ad carinam flexo.
Hab.—In Oceano Indico.
4. Dichelaspis Lowei, Darwin. (‘Lepadidæ,’ p. 128. Tab. 2, fig. 8.)
D. scutorum segmento basali angustiore quam occludens segmentum, longitudine ferè 4/5: tergorum parte inferiori duplo latiore quam occludens scutorum segmentum.
Hab.—In Insulâ Madeirâ.
5. Dichelaspis orthogonia, Darwin. (‘Lepadidæ,’ p. 130, Tab. 2, fig. 10.)
D. scutorum basali segmento angustiore quam occludens segmentum; longitudine ferè dimidiâ; duorum segmentorum junctione calcareâ: tergorum prominentiis marginalibus inæqualibus 5: carinâ deorsum in parvo calyce lunato terminatâ.
Hab.—(?)
1. Oxynaspis celata, Darwin. (‘Lepadidæ,’ p. 134, Tab. 3, fig. 1.)
Hab.—Insulâ Madeirâ.
1. Conchoderma aurita, Linn. (Darwin, ‘Lepadidæ,’ p. 141, Tab. 3, fig. 4.)
C. capitulo duobus tubularibus quasi-auribus instructo, pone terga rudimentalia (sæpe nulla) positis: scutis bilobatis: carinâ nullâ, aut omnino rudimentali: pedunculo longo, a capitulo distincte separato.
Hab.—In Oceano, pænè ubique.
2. Conchoderma virgata, Spengler. (Darwin, ‘Lepadidæ,’ p. 146, Tab. 3, fig. 2.)
C. scutis trilobatis: tergis intùs concavis, apicibus introrsùm leviter curvatis: carinâ modicâ, leviter curvatâ: pedunculo in capitulum coalescente.
Hab.—In Oceano, pænè ubique.
3. Conchoderma Hunteri, Owen. (Darwin, ‘Lepadidæ,’ p. 153, Tab. 3, fig. 3.)
C. valvis angustis: scutis trilobatis, prominentiâ laterali non latiore quam inferior: tergorum parte superiore pæne rectangulè secundùm aperturæ marginem flexâ: carinâ valde arcuatâ: pedunculo brevi, in capitulum coalescente.
Hab.—In Oceano torrido, aut Indico, aut Pacifico.
1. Alepas minuta, Philippi. (Darwin, ‘Lepadidæ,’ p. 160, Tab. 3, fig. 5.)
A. aperturâ non prominente, capituli longitudinis vix tertiam partem æquante: scutis corneis, pæne absconditis: longitudine totâ ad quartam unciæ partem.
Hab.—In Mari Mediterraneo.
2. Alepas parasita, S. Rang. (Darwin, ‘Lepadidæ,’ p. 163.)
A. aperturâ non prominente, capituli longitudinis 2/3 æquante: scutis corneis: longitudine totâ ad 2 uncias.
Hab.—In Mari Mediterraneo et Oceano Atlantico septentrionale.
3. Alepas cornuta, Darwin. (‘Lepadidæ,’ p. 165, Tab. 3, fig. 6.)
A. aperturâ parvâ, leviter prominente: scutis nullis: capitulo ple[Pg 630]rumque tribus, parvis, compressis eminentiis secundum carinalem marginem instructo.
Hab.—In Insulis Indicis occidentalibus.
4. Alepas tubulosa, Quoy et Gaimard. (Darwin, ‘Lepadidæ,’ p. 169.)
A. aperturâ parvâ prominente et tubulosâ: scutis et prominentiis secundùm marginem carinalem, nullis.
Hab.—In Novâ Zealandiâ.
1. Anelasma squalicola, Lovén. (Darwin, ‘Lepadidæ,’ p. 170, Tab. 5, fig. 1-7.)
Hab.—In Mari septentrionali, squalis affixa.
1. Alcippe lampas, Hancock. (Darwin, ‘Balanidæ,’ p. 530, Tab. 22, et Tab. 23, fig. 16-19.)
Hab.—Littoribus Anglicis, molluscis condita.
1. Ibla Cumingii, Darwin. (‘Lepadidæ,’ p. 183, Tab. 4, fig. 8.)
I. (fœm.) valvarum marginibus lateralibus, et superficie interiore, cæruleis; pedunculi spinis plerumque annulis cæruleo-fuscis.
Hab.—In Archipel. Philippino, et littore Burmah.
2. Ibla quadrivalvis, Cuvier. (Darwin, ‘Lepadidæ,’ p. 203, Tab. 4, fig. 9.)
I. (Herm.) valvis et pedunculi spinis sub-flavis; basali tergorum angulo, introrsùm spectanti, hebete, quia margo carinalis inferior longiùs quam mango scutalis prominet.
Hab.—In Australiâ meridionali.
Sectio † Carinæ umbone sub-centrali.
Sectio †† Carinæ umbone ad apicem posito.
1. Scalpellum vulgare, Leach. (Darwin, ‘Lepadidæ,’ p. 222, Tab. 5, fig. 15.)
S. (Herm.) valvis 14, si rostrum pæne rudimentale includatur; sub-carinâ nullâ; lateribus superioribus inæqualiter ovatis.
Hab.—In maribus Europæ.
2. Scalpellum magnum, Darwin. (‘Lepadidæ Fossiles,’ p. 18, Tab. 1, fig. 1.)
S. valvis probabiliter 14, sub-carinâ nullâ; laterum carinalium et rostralium umbonibus liberè (sicut cornua) prominentibus, dimidiam seu tertiam partem longitudinis valvarum æquantibus.
Foss.—In “Coralline Crag Formatione,” Angliâ.
3. Scalpellum ornatum, J. E. Gray. (Darwin, ‘Lepadidæ,’ p. 244, Tab. 6, fig. 1.)
S. (Fœm.) valvis 14, sub-rufis; sub-carinâ nullâ; lateribus superioribus quadranti-formibus, arcu crenâ profundâ notato.
Hab.—In Africâ meridionali.
4. Scalpellum rostratum, Darwin. (‘Lepadidæ,’ p. 259, Tab. 6, fig. 7.)
S. (Herm.) valvis 15; rostro per-magno; sub-carinâ præsente; laterum paribus quatuor; pari superiore pentagono.
Hab.—In Archipel. Philippino.
5. Scalpellum Peronii, J. E. Gray. (Darwin, ‘Lepadidæ,’ p. 264, Tab. 6, fig. 6.)
S. (Herm.) valvis 13; sub-carinâ præsente; laterum paribus tribus; pari superiore multùm elongato; pedunculi squamis calcareis nullis.
Hab.—In Australiâ.
6. Scalpellum Darwinianum, Bosquet. (‘Monograph. Crust. Foss. de Limbourg,’ Tab. 4, fig. 1-5.)
S. carinâ ad angulum 115° flexâ, umbone centrali, costis tenuibus ab umbone utrinque radiantibus; carinæ tectis et superioris et inferioris partis rectis, planis, lævibus.
Foss.—In Cretaceâ superiore Formatione.
7. Scalpellum Hagenovianum, Bosquet. (Idem, Tab. 4, fig. 13-16.)
S. carinæ umbone ad 1/6 totius ab apice longitudinis posito; carinæ tecto plano, utrinque sulcato; parietum et intra-parietum confluentium parte superiore solummodò cristis instructo ab umbone radiantibus.
Foss.—In Cretaceâ superiore Formatione.
8. Scalpellum radiatum, Bosquet.[158] (Idem, Tab. 4, fig. 17-18.)
[158] Scalpelli 4 aliæ species fossiles, ad secundam generis sectionem pertinentes, à Dom. Bosquet descriptæ et nominatæ sunt. Jam cum equidem, ubi valvæ disjunctæ reperiuntur, nomina specifica non nisi carinis (et interdùm scutis) tribuere soleam, S. pulchellum, Bosq., hic inserere non possum, cujus scuti aut carinæ fragmenta valdè imperfecta solùm reperta sunt. Scalpellum gracile, S. pygmæum, et S. radiatum, Bosq., omnia Scalpello maximo et ejus varietatibus tam propè associantur, ut, cum ipse specimina non viderim, characteres breves diagnosticos hic exhibere non audeam. Veruntamen, cum Dom. Bosquet se hujus rei penitus peritum se probaverit, longissimè abest ut in dubium vocare velim quin hæ species reverà distinctæ existant.
S. scuti superficie cristis tectâ, inter se aliquantùm distantibus, ab umbone radiantibus; scuti umbone propè medium marginis occludentis posito; termino superiore truncato, lato, non acuminato.
Foss.—In Cretaceâ superiore Formatione.
9. Scalpellum rutilum, Darwin. (‘Lepadidæ,’ p. 253, Tab. 6, fig. 2.)
S. (Fœm. an Herm.) valvis 14 sub-rufis; sub-carinâ nullâ; carinæ[Pg 632] tecto plano, utrinque cristâ rotundatâ instructo, margine basali truncato. Lateribus superioribus latitudine duplo longioribus.
Hab.—(?)
10. Scalpellum villosum, Leach. (Darwin, ‘Lepadidæ,’ p. 274, Tab. 6, fig. 8.)
S. (Herm.) valvis 14; sub-rostro et sub-carinâ præsentibus; carinâ pæne rectâ; laterum paribus tribus, pari superiore triangulo.
Hab.—In maribus orientalibus.
11. Scalpellum quadratum, Dixon. (Darwin, ‘Lepad. Foss.’ p. 22, Tab. 1, fig. 3.)
S. tecto parietibusque carinæ planis, lævibus, simplicibus, margine basali fere rotundato. Lateribus superioribus quinque-lateralibus, lævibus.
Foss.—In “Eocenâ Formatione” Anglicâ.
12. Scalpellum fossula, Darwin. (‘Lepad. Foss.’ p. 24, Tab. 1, fig. 4.)
S. carinâ intra-parietibus instructâ; tecto utrinque costis magnis, tumidis, superne planatis, marginato; margine basali obtusè acuminato. Lateribus superioribus quinque-lateralibus, costis duabus modicis ab apice ad marginem basalem continuatis.
Foss.—In Cretaceâ superiore Formatione.
13. Scalpellum maximum, J. Sowerby. (Darwin, ‘Lepad. Foss.’ p. 26, Tab. 2, fig. 1-10.)
S. carinâ intra-parietibus instructâ; tecto subangulari vel subcarinato; margine basali rectangulariter acuto; totâ valvâ plus minusve introrsùm arcuatâ, sed margine interno ferè-recto; tecto transversè plus minusve convexo; superficie pænè lævi, striis paucis obsoletis longitudinalibus elevatis; tectum, parietes, et intra-parietes inter se costis plus minusve prominentibus separantur.
Var. typicum, (Tab. 2, fig. 1, 4, 5, 8.)
S. carinâ introrsùm leviter arcuatâ, latitudine valvæ altitudinem superante; tecto transversè leniter arcuato; parietibus intra-parietibusque angustis, superficie pæne lævi.
Var. cylindraceum, (Tab. 2, fig. 2.)
S. parte superiore carinæ liberè prominente, parte interiore intra-parietibus rotundatis, inflexis, itâ repletâ, ut pæne cylindrica fiat; superficie externâ lævi, tecto parietibusque pæne confluentibus.
Var. sulcatum, (Tab. 2, fig. 3.)
S. carinâ introrsùm valde arcuatâ, sub-carinatâ; valvæ latitudine circà dimidium altitudinis æquante, tecto transversè præruptè arcuato; parietibus intra-parietibusque latiusculis. Apice solidè repleto, liberè paululùm prominente; superficie externâ striis paucis, rotundatis, ad alterum vel utrumque latus costarum duarum tectum et parietes separantium.
Foss.—In Cretaceâ a superiore Formatione.
14. Scapellum lineatum, Darwin. (‘Lepad. Foss.’ p. 35, Tab. 2, fig. 11, 12.)
S. superficie totâ carinæ lineis tenuibus, rotundatis, longitudinalibus, proximis, microscopicis obtectâ; cristæ centralis costâ crassiore; costis duabus vel tribus tectum et parietes separantibus; latitudine valvæ circa dimidium altitudinis æquante; intra-parietibus latiusculis, nullâ costâ conspicuâ a parietibus separatis; apice solidè repleto, aliquantulum liberè prominente.
Foss.—In Cretaceâ inferiore Formatione.
15. Scapellum hastatum, Darwin. (‘Lepad. Foss.’ p. 37, Tab. 2, fig. 13.)
S. carinâ intra-parietibus, intrà paululùm positis, instructâ; valvâ totâ introrsùm valdè arcuatâ, margine interno non recto; margine basali acuto, lanceolato; valvâ tenui, lævi, tecto transversè leniter arcuato; parietibus à tecto vix disjunctis.
Foss.—In “Grey Chalk, Dover.”
16. Scalpellum angustum, Dixon. (Darwin, ‘Lepad. Foss.’ p. 37, Tab. 1, fig. 2.)
S. carinâ angustâ, introrsùm valdè arcuatâ; tecto à parietibus rectangulè inflexis costâ, (ut videtur) disjuncto; intra-parietibus usque ad dimidium valvæ pertinentibus, deinde obliquè et abruptè truncatis; margine basali acutè cuspidato.
Foss.—In Cretaceâ Formatione.
17. Scalpellum quadricarinatum, Reuss. (Darwin, ‘Lepad. Foss.’ p. 38.)
S. carinâ intra-parietibus latis (ut videtur) instructâ; tecto transversè plano, lævi, costâ prominente utrinque marginato; margine basali abruptè truncato.
Foss.—In “Untern Pläner-Kalke, Bohemia.”
18. Scalpellum trilineatum, Darwin. (‘Lepad. Foss.’ p. 38, Tab. 1, fig. 5.)
S. carinæ tecto transversè leniter arcuato, subcarinato, costâ centrali et costis duabus lateralibus, rotundatis, tumidis; parietibus angustis leviter concavis, rectangulè inflexis.
Foss.—In “Grey Chalk, Dover.”
19. Scalpellum simplex, Darwin. (‘Lepad. Foss.’ p. 39, Tab. 1, fig. 9.)
S. carinâ lævi; parietibus angustissimis, rectangulè inflexis; tecto subcarinato, transversè mediocriter arcuato; margine basali rectangulè acuminato.
Foss.—In “Lower Greensand.”
20. Scapellum arcuatum, Darwin. (‘Lepad. Foss.’ p. 40, Tab. 1, fig. 7.)
S. valvarum lineis angustis elevatis ab apice radiantibus: carinæ tecto transversè leniter arcuato, et parietibus rectangulè inflexis, leniter concavis, lævibus.
Foss.—In “Gault.”
21. Scalpellum solidulum, Steenstrup. (Darwin, ‘Lepad. Foss.’ p. 42, Tab. 1, fig. 8.)
S. valvarum lineis latiusculis elevatis ab apice radiantibus. Carinæ parte superiori liberè prominente, et cristâ centrali, internâ, longitudinali instructâ.
Foss.—In Cretaceâ superiore Formatione; Scaniâ.
22. Scalpellum tuberculatum, Darwin. (‘Lepad. Foss.’ p. 43, Tab. 1, fig. 10.)
S. valvarum lineis tenuibus, tuberculatis, elevatis, ab apice radiantibus: carinæ tecto transversè leniter arcuato, et parietibus striatis: scuti umbone prope in medio marginis occludentis posito, costis duobus ab umbone ad angulum basi-lateralem, et ad basalis marginis medium decurrentibus.
Foss.—In Cretaceâ Formatione.
23. Scalpellum semiporcatum, Darwin. (‘Lepad. Foss.’ p. 44, Tab. 1, fig. 6.)
S. carinâ ignotâ: scuti costis duobus ab umbone ad angulum basi-lateralem et ad marginis basalis medium decurrentibus; superficie inter hanc costam et marginem occludentem lineis tenuibus, longitudinalibus, elevatis instructâ.
Foss.—In Cretaceâ superiore Formatione; Scaniâ.
24. Scalpellum (?) cretæ, Steenstrup. (Darwin, ‘Lepad. Foss.’ p. 45, Tab. 1, fig. 11.)
S. valvis lævibus tenuissimis: scuti umbone propè medium marginis occludentis posito; costis tribus obscuris ab umbone ad angulos tergo-lateralem et basi-lateralem, et ad medium marginis basalis decurrentibus: carinæ apice et margine basali acutis; distincti parietes absunt.
Foss.—In Cretaceâ Formatione.
†. Scuta, aut lævia aut lineis incrementi tenuibus solùm notata.
A. Scuta, costâ ab apice ad centrum marginis basalis non decurrente.
B. Scuta, costâ, nonnunquam subobsoletâ, ab apice ad centrum marginis basalis decurrente.
††. Scuta, aut longitudinaliter aut transversè (i. e. secundum lineas incrementi) costata.
1. Pollicipes cornucopia, Leach. (Darwin, ‘Lepadidæ,’ p. 298, Tab. 7, fig. 1.)
P. capitulo, valvarum duobus aut pluribus sub rostro verticillis instructo: valvis albis, aut glaucis: pedunculo, squamarum densis verticillis symmetricè dispositis.
Hab.—In Europâ.
2. Pollicipes elegans, Lesson. (Darwin, ‘Lepadidæ,’ p. 304.)
P. capitulo, valvarum duobus aut pluribus sub rostro verticillis instructo: valvis et pedunculi squamis rufo-aurantiacis: squamarum verticillis densis symmetricè dispositis.
Hab.—In Peruviâ et Mexico.
3. Pollicipes polymerus, G. B. Sowerby. (Darwin, ‘Lepadidæ,’ p. 307, Tab. 7, fig. 2.)
P. capitulo, valvarum duobus, tribus, aut pluribus sub rostro verticillis instructo; valvis sub-fuscis; lateribus à supremo ad infimum gradatim quoad magnitudinem positis; carinæ margine basali (introrsùm spectanti) ad medium excavato; pedunculi squamarum verticillis densis, symmetricè dispositis.
Hab.—In Californiâ et Oceano Pacifico meridionali.
4. Pollicipes spinosus, Quoy. (Darwin, ‘Lepadidæ,’ p. 324, Tab. 7, fig. 4.)
P. capitulo valvarum uno aut pluribus sub rostro verticillis instructo: laterum pari superiore vix inferioribus longiore: membranâ valvas tegente (post desiccationem) subfuscâ flavescente: pedunculi squamis inæqualibus, non symmetricis: verticillis longiusculè distantibus.
Hab.—Novâ Zealandiâ.
5. Pollicipes sertus, Darwin. (‘Lepadidæ,’ p. 327, Tab. 7, fig. 5.)
P. capitulo valvarum uno aut pluribus sub rostro verticillis instructo: laterum pari superiore vix inferioribus longiore: membranâ valvas tegente (post desiccationem) fusco rufescente obscuro: rostro dimidiam carinæ longitudinem æquante, superficiei internæ altitudine latitudinem plus duplo superante: pedunculi squamis inæqualibus, non symmetricis: verticillis longiusculè distantibus.
Hab.—Novâ Zealandiâ.
6. Pollicipes concinnus, J. Morris. (Darwin, ‘Lepad. Foss.’ p. 50, Tab. 3, fig. 1.)
P. scutis pæne quadratis, margine basali propè rostrum subconcavo, segmento tergo-laterali, è lineis incrementi, ut videtur, reflexis formato, lato, rotundato et prominente: tergis latis, pæne quadratis: carinæ margine basali, ut videtur, acuto.
Foss.—In ‘Oxford Clay.’
7. Pollicipes ooliticus, Buckman. (Darwin, ‘Lepad. Foss.’ p. 51, Tab. 3, fig. 2.)
P. scutis triangulis; superficie undulatâ; margine basali rectangulè ad marginem rectum tergo-lateralem posito; segmentum tergo-laterale è lineis reflexis incrementi formatum deest. Carinâ pæne rectâ, semi-cylindricâ, margine basali quadrato.
Foss.—In ‘Stonesfield Slate.’
8. Pollicipes Nilssonii, Steenstrup. (Darwin, ‘Lepad.’ Foss. p. 52, Tab. 3, fig. 11.)
P. scutis triangulis, planis: margine basali cum margine occludente[Pg 636] angulum pæne rectum, cum margine recto tergo-laterali, angulum aliquanto minorem formante: deest segmentum tergo-laterale, è lineis incrementi reflexis formatum. Carinâ introrsùs admodùm arcuatâ, crassâ; marginis basalis mucrone obtuso.
Foss.—In Cretaceâ superiore Formatione; Scaniâ.
9. Pollicipes Hausmanni, Koch. (Darwin, ‘Lepad. Foss.’ p. 53, Tab. 3, fig. 3.)
P. scutis subtriangulis, angulo basi-laterali valde rotundato; apice producto; margine basali cum margine occludente angulum pæne rectum formante; internâ apicis superficie prominente, margineque tergali sulcato.
Foss.—In ‘Hilsthon,’ Germaniâ.
10. Pollicipes politus, Darwin. (‘Lepad.’ Foss. p. 54, Tab. 3, fig. 4.)
P. scutis ferè rhombicis, lævissimis; margine basali cum margine occludente angulum recto majorem formante; margine occludente projecturâ parietali, lineari, minutâ instructo; internâ apicis superficie concavâ.
Foss.—In ‘Gault.’ (?)
11. Pollicipes elongatus, Steenstrup. (Darwin, ‘Lepad. Foss.’ p. 55, Tab. 3, fig. 5.)
P. scutis pæne quadratis; margine occludente et parte inferiori marginis tergo-lateralis rectangulè ad marginem basalem positis; apice obtuso.
Foss.—In Cretaceâ Formatione albâ. Daniâ.
12. Pollicipes acuminatus, Darwin. (‘Lepad. Foss.’ p. 56, Tab. 3, fig. 6.)
P. scutis elongatis, triangulis; margine basali cum margine occludente angulum recto longè minorem formante; internâ apicis superficie concavâ.
Foss.—In Cretaceâ Formatione.
13. Pollicipes Angelini, Darwin. (‘Lepad. Foss.’ p. 56, Tab. 3, fig. 7.)
P. scutis elongatis, triangulis, margine basali propè angulum rostralem in prominentiam obliquè rotundatam producto; internâ apicis superficie prominente, margine occludente sulcato.
Foss.—In Cretaceâ superiore Formatione.
14. Pollicipes reflexus, J. Sowerby. (Darwin, ‘Lepad. Foss.’ p. 58, Tab. 3, fig. 8.)
P. scutis tenuibus, subovalibus; margine basali cum margine occludente angulum recto longè majorem formante; costâ obsoletâ rotundatâ ab apice ad angulum basi-lateralem decurrente, valvam in duas ferè æquales partes separante. Carinâ lineari, transversè abruptè arcuatâ; margine basali multum producto, apice truncato. Lateribus superioribus sub-pentagonis.
Foss.—In Angliâ: ‘Eocenâ Formatione.’
15. Pollicipes carinatus, Philippi. (Darwin, ‘Lepad. Foss.’ p. 60, Tab. 3, fig. 9.)
P. scutis crassiusculis ad formam trianguli æquianguli accedentibus; margine occludente externè costâ, humili firmato; costâ firmâ ab apice ad centrum marginis basalis decurrente; margine basali recto; angulo basi-laterali truncato, brevi; segmentum tergo-laterale ex lineis incrementi reflexis formatum, deest. Carinâ externè valdè carinatâ cum sulco laterali ad utrumque latus.
Foss.—In Form. Tertiariâ, Siciliâ.
16. Pollicipes glaber, Roemer. (Darwin, ‘Lepad. Foss.’ p. 61, Tab. 3, fig. 10.)
P. scutis subtenuibus, latiusculis, ad formam trianguli æquianguli accedentibus; margine basali non prorsus recto; tergo-laterali segmento e zonis incrementi reflexis formato, ubi (latissimum), reliquæ valvæ dimidium æquante: margine tergali apicis intùs sulcato. Carinæ margine basali obtusè acuminato. Lateribus superioribus triangulis, tertiam partem longitudinis tergorum æquantibus. Lateribus anticis inferioribus singulis costâ propè terminum marginis basalis decurrente.
Foss.—In Cretaceâ, super. et inferiore Formatione.
17. Pollicipes unguis, J. Sowerby. (Darwin, ‘Lepad. Foss.’ p. 64, Tab. 4, fig. 1.)
P. scutis incognitis, verisimiliter ut in P. glabro. Margine basali carinæ obtusè acuminato, sub-rotundato. Lateribus superioribus elongatis triangularibus, dimidium longitudinis tergorum superantibus. Lateribus anticis inferioribus costâ subcentrali instructis.
Foss.—In ‘Gault.’
18. Pollicipes validus, Steenstrup. (Darwin, ‘Lepad. Foss.’ p. 68, Tab. 4, figs. 2 and 3.)
P. scutis crassis, angustis; margine occludente externè costâ rotundatâ forti firmato; intus prominentiâ rostrali infra marginem rectum basalam dependente; costâ ab apice ad marginem basalem propiùs ad rostralem quam ad basi-lateralem angulum accedente. Carinâ lævissimâ, transversè semicylindricâ; parte superiori liberè prominente, internè aut planâ aut cristâ centrali instructâ.
Foss.—In Cretaceâ sup. Formatione, Scaniâ et Maestricht.
19. Pollicipes Darwinianus, Bosquet. (Monographie Crust. Foss. de Limbourg, Tab. 1, fig. 8-16.)
P. scuti marginibus basali et occludente angulum solum 35° formantibus; intùs prominentiâ rostrali infra marginem basalem dependente; costâ ab apice ad marginem basalam propemodùm ad angulum rostralem accedente. Tergo pæne pentagono, intùs prominentiâ à medio marginis scutalis surgente.
Foss.—In Cretaceâ superiore Formatione.
20. Pollicipes dorsatus, Steenstrup. (Darwin, ‘Lepad. Foss.’ p. 69, Tab. 4, fig. 4.)
P. scutis crassis; ad formam trianguli æquianguli accedentibus; margine occludente externè costâ rotundatâ firmato; angulo basi-laterali latè truncato, dimidium longitudinis marginis basalis veri, æquante; tergo-laterali segmento ex lineis incrementi reflexis formato, angustissimo. Tergorum costâ rectâ, latâ, proclivi, ab apice ad angulum basalem decurrente; angulo basali scutum versùs, obliquè truncato.
Foss.—In Cretaceâ superiore Formatione, Daniâ.
21. Pollicipes mitella, Linn. (Darwin, ‘Lepadidæ,’ p. 316, Tab. 7, fig. 3.)
P. capitulo valvarum unico sub rostro verticillo instructo: laterum pari superiore (introrsum spectanti) inferiorum magnitudinem ter aut quater superante: lateribus inferioribus utrinque obtegentibus: pedunculi squamarum verticillis densis, symmetricè dispositis.
Hab.—In Insulis Indicis orientalibus.
22. Pollicipes striatus, Darwin. (‘Lepad. Foss.’ p. 70, Tab. 4, fig. 5.)
P. valvis longitudinaliter striatis: scutis sub-triangulis, margine tergo-laterali valdè arcuato et prominente; costâ obscurâ, rotundatâ, ab apice ad angulum basi-lateralem decurrente; internâ apicis superficie solidè repletâ, sine sulco manifesto.
Foss.—In Cretaceâ superiore Formatione, Angliâ.
23. Pollicipes semilatus, Darwin. (‘Lepad. Foss.’ p. 72, Tab. 4, fig. 6.)
P. valvis longitudinaliter et transversè costatis: scutorum margine basali brevi, recto, cum margine occludente angulum rectum formante; costâ parietali tenuissimâ ab apice, ad angulum prominentem basi-lateralem decurrente; hæc valvam in duas partes inæquales dividit, è quibus portio tergo-lateralis latior est.
Foss.—In Cretaceâ Formatione.
24. Pollicipes rigidus, J. Sowerby. (Darwin, ‘Lepad. Foss.’ p. 73, Tab. 4, fig. 7.)
P. valvis transversè costatis: scutorum margine basali recto, cum margine occludente angulum recto majorem formante; costâ angustissimâ, parietali, ab apice ad angulum basi-lateralem decurrente: tergis, costâ curvâ, parietali, ad angulum basalem decurrente instructis; apice basali in prominentiam parvam terminante, lateribus prominentiæ parallelis.
Foss.—In ‘Gault.’
25. Pollicipes fallax, Darwin. (‘Lepad. Foss.’ p. 75, Tab. 4, fig. 8.)
P. valvis transversè costatis: scutis margine basali non recto, angulum pæne rectum cum margine occludente formante; costâ, parie[Pg 639]tibus obliquis, ab apice ad angulum basi-lateralem decurrente: tergis, costâ curvatâ, parietibus obliquis, ad angulum basalem latum, rotundatum, decurrente.
Foss.—In Cretaceâ superiore Formatione, Angliâ, Scaniâ, Hanoveriâ.
26. Pollicipes elegans, Darwin. (‘Lepad. Foss.’ p. 76, Tab. 4, fig. 9.)
P. valvis longitudinaliter et transversè striatis: scutorum margine basali recto, cum margine occludente angulum recto paulo majorem formante; costâ parietali, latiore quam pro solitâ incrementorum linearum latitudine, ab apice ad angulum basi-lateralem decurrente: tergorum costâ parietali, rectâ, ad apicem basalem, acuminatum decurrente.
Foss.—In Cretaceâ superiore Formatione, Scaniâ, Daniâ.
[Species aliquot quas, scutis incognitis, auctores a valvis insignibus nominârunt, hìc pro tempore solummodo iterum describuntur.]
27. Pollicipes Bronnii, F. Roemer. (Darwin, ‘Lepad. Foss.’ p. 77, Tab. 4, fig. 10.)
P. carinâ lævi, subcarinatâ, margine basali arcuato et turgido; totâ valvâ vel extrorsùm arcuatâ vel pæne rectâ; internè, costis duabus elevatis ad utrumque latus partis superioris liberè prominentis.
Foss.—In ‘Upper Greensand,’ et ‘Hils-conglomerat.’
28. Pollicipes planulatus, J. Morris. (Darwin, ‘Lepad. Foss.’ p. 27, Tab. 4, fig. 11.)
P. tergis subrhombicis, latis, lævibus, apice basali latè truncato, latitudine dimidium longitudinis marginis occludentis æquante; apice basali truncato angulum rectum cum margine scutali formante; parte superiori marginis scutalis cuspidem latam, rotundatam, leviter prominentem formante.
Foss.—In ‘Oxford Clay.’
1. Lithotrya dorsalis, G. B. Sowerby. (Darwin, ‘Lepadidæ,’ p. 351, Tab. 8, fig. 1 a′.)
L. scutis terga angustè obtegentibus: carinâ intùs concavâ; rostro, duorum aut trium squamarum subjacentium latitudinem æquante: lateribus, squamarum quinque subjacentium longitudinem æquantibus, superficie internâ angustè ellipticâ: pedunculi squamis superioribus verticillum secundum minus duplo superantibus.
Hab.—Insulis Indicis occidentalibus.
2. Lithotrya cauta, Darwin. (‘Lepadidæ,’ p. 356, Tab. 8, fig. 3.)
L. scutis terga amplè obtegentibus: carinâ intus concavâ: rostro, squamarum subjacentium latitudinem vix æquante: lateribus, squamas subjacentes sesquitertio superantibus, superficie internâ latè ellipticâ: pedunculi squamis superioribus verticillum secundum pæne quadruplo superantibus.
Hab.—In Australiâ.
3. Lithotrya Nicobarica, Reinhardt. (Darwin, ‘Lepadidæ,’ p. 359, Tab. 8, fig. 2.)
L. scutis terga angustè obtegentibus: carinæ cristâ internâ tenui in parte superiore positâ: rostro conspicuo, squamarum sex subjacentium latitudinem æquante: lateribus, superficie internâ triangulâ squamarum septem subjacentium latitudinem æquantibus.
Hab.—In Insulis Nicobaricis et Timor.
4. Lithotrya rhodiopus, J. E. Gray. (Darwin, ‘Lepadidæ,’ p. 363, Tab. 8, fig. 4.)
L. scutis terga amplè obtegentibus: carinæ cristâ internâ tenui, in parte superiore positâ: lateribus, superficie internâ symmetricè et latè ovatâ, carinæ latitudinis plus quam tertiam partem æquantibus: tergorum basali apice tenui, et angulo carinali producto: rostro et pedunculo ignotis.
Hab.—(?)
5. Lithotrya truncata, Quoy. (Darwin, ‘Lepadidæ,’ p. 366, Tab. 9, fig. 1.)
L. scutis in profundam tergorum plicam insertis: carinæ cristâ centrali prominente et rotundatâ in parte superiore: rostro et lateribus rudimentalibus, carinæ latitudinis quindecimam fere partem æquantibus.
Hab.—In Arch. Philippino et Mari Pacifico.
6. Lithotrya Valentiana, J. E. Gray. (Darwin ‘Lepadidæ,’ p. 371, Tab. 8. fig. 5.)
L. scutis in profundam tergorum plicam insertis: tergorum opposito superiore margine, plicâ alterâ æquè profundâ instructo: carinæ cristâ prominente centrali, marginibus quadratis, in parte superiore: rostro rudimentali: lateribus et pedunculo ignotis.
Hab.—In Mari Rubro.
1. Loricula pulchella, G. B. Sowerby, jun. (Darwin, ‘Lepad. Foss.’ p. 81, Tab. 5.)
Foss.—In Cretaceâ inferiore Formatione.
1. Cryptophialus minutus, Darwin. (‘Balanidæ,’ p. 566, Tab. 23 et 24.)
Hab.—In littore occidentali Americæ meridionalis.
1. Proteolepas bivincta, Darwin. (‘Balanidæ,’ p. 589, Tab. 24 et 25.)
Hab.—In insulis Indicis occidentalibus.
N.B.—Letters of reference the same throughout the Plate.
N.B.—The same letters of reference are used for the parts of the mouth throughout this plate. All the organs here represented have been cleaned by boiling in potash, and consist exclusively of the external membrane.
N.B. The names in italics are synonyms or doubtful species.
Plate I.
George Sowerby.
Plate II.
George Sowerby.
Plate III.
George Sowerby.
Plate IV.
George Sowerby.
Plate V.
George Sowerby.
Plate VI.
George Sowerby.
Plate VII.
George Sowerby.
Plate VIII.
George Sowerby.
Plate IX.
George Sowerby.
Plate X.
George Sowerby.
Plate XII.
Plate XIII.
Plate XIV.
George Sowerby.
CREUSIA : CHELONOBIA .
Plate XVI.
George Sowerby.
Plate XVIII.
George Sowerby.
Plate XXII.
George Sowerby.
Plate XXIII.
ALCIPPE .
George Sowerby.
Plate XXIV.
George Sowerby.
Plate XXVI.
George Sowerby.
MOUTH:THORAX.
Plate XXVII.
George Sowerby.
NERVOUS SYSTEM.
Plate XXVIII.
George Sowerby.
CEMENTING APPARATUS.
Plate XXIX.
CIRRI.
George Sowerby.
LARVÆ, FIRST STAGES.
Plate XXX.
George Sowerby.
LARVÆ, LAST STAGES.
Obvious misprints have been fixed. Archaic and unusual words, spellings and styling have been maintained when they are used consistently. Spelling has been standardized within the book where one spelling was predominant, even if it is not in common usage. Inconsistent spelling and hyphenation has been maintained when there was not a predominant spelling. Details of the changes follow.
No attempt has been made to correct the capitalization or standardize the italicization of biological names—they are left as in the original.
Hovering over a Greek word will display its transliteration.
All of Darwin’s errata have been applied to the text, except the one on page 255. It was not applied because the wording is not obvious.
This book was published in two volumes, of which this is the second. The first volume was released as Project Gutenberg ebook #31558, available at http://www.gutenberg.org/ebooks/31558. Links to the first book have not been created in this volume. The word “Lepadidæ” has not been cross referenced to the chapter titled “Lepadidæ” because with the exception of that chapter, the use of this word is a reference either to volume 1 of this work, or to Darwin’s monograph on fossil Lepadidæ.
Details of the Changes
Page vii, footnote 1: du terrain Crétacé du D. de Limbourg
In the original book: du terrain Crétacée du D. de Limbourg
Page 4: the shell, without the operculum being removed,
In the original book: the shell, without the operculum be removed,
Page 5, footnote 3: by some authors as “intersticia.”
In the original book: by some authors as “intersticia”.
Page 7: carrying two multiarticulated rami
In the original book: carrying two multi-articulated rami
Page 8: interjection (!)
In the original book: interjection [!]
Page 9: biramous, multiarticulated limbs
In the original book: biramous, multi-articulated limbs
Page 11: knowledge of the class
In the original book: knowlege of the class
Page 12: first and second pairs of antennæ
In the original book: first and second pairs of attennæ
Page 20: what has happened to one organ
In the original book: what has happenend to one organ
Page 30: ORDER I.—THORACICA.
In the original book: I.—ORDER THORACICA.
Page 33. The table of contents has been completely restructured, to make the entries match (or compromise with) the titles in the text.
Page 34. Inserted missing title (which appeared in the TOC):
Structure of Shell.
Page 37: alæ in all cases (as is obvious in Pachylasma)
In the original book: alæ in all cases (as is obvious in Pachylasma)
Page 40: the upper lateral valves alternate
In the original book: the upper Lateral valves alternate
Page 45: owing to its midrib, is generally thrust
In the original book: owing to its mid-rib, is generally thrust
Page 45. Inserted missing title (which appeared in the TOC):
Structure of the Radii.
and left the original paragraph title as it was:
Radius.
Page 47: and of the recipient furrow in Octomeris (Pl.
In the original book: and of the recipient furrow in Octomeris (Pl.
Page 47: Structure of the Alæ.
In the original book: Alæ.
Page 48: Structure of the Sheath.
In the original book: Sheath.
Page 49: Structure of the Basis.
In the original book: Basis.
Page 51: Structure of the Opercular Valves (Scuta and Terga).
In the original book: Opercular Valves.
Page 51: scutum, S, being left articulated to the tergum, T.
In the original book: scutum, S, being left articulated to the tergum, T.
Page 61: Muscles of Sack.
In the original book: Sack; muscles of, &c.
Page 62: can only act as depressores;
In the original book: can only act as depressors;
Page 65: Balanidæ are the ovigerous fræna of the Lepadidæ
In the original book: Balanidæ are the ovigerous fræna of the Lepadidæ
Page 65. Inserted title from the TOC:
Thorax and Body.
and demoted the existing title:
Parts of the body included within the shell or carapace.
to the head of the next paragraph.
Page 67: and p. 440, (foot-note), consider the articulated
In the original book: and p. 440, foot-note), consider the articulated
Page 68: Muscular System.
Attachment of the Body to the Shell.
In the original book: Attachment of the Body to the Shell. Muscular System.
Page 76: there is no trace of any labrum
In the original book: there is no trace of any labium
Page 77: (Pl. 26, figs. 3, 4, c1;
In the original book: (Pl. 26, figs. 3, 4, c 1;
Page 78: pairs of gnathites, which latter have only their
In the original book: pairs of gnathites; which latter have only their
Page 79: thickened membrane (fig. 3, c1)
In the original book: thickened membrane (fig. 3, c 1)
Page 79: thickened membrane (c2), the basal
In the original book: thickened membrane (c 2), the basal
Page 80: piece of membrane (c1) represents
In the original book: piece of membrane (c 1) represents
Page 81: middle segment (fig. 4, a1) of each maxilla
In the original book: middle segment (fig. 4, a 1) of each maxilla
Page 81: this latter (a2) is separated from
In the original book: this latter (a 2) is separated from
Page 81: There are always six pairs; each biramous
In the original book: There are always six pair; each biramous
Page 83: they carry as many as ten pairs in a
In the original book: they carry as many as ten pair in a
Page 84: in the sub-genus Acasta, in which, differently
In the original book: in the sub-genns Acasta, in which, differently
Page 85: are developed into strong, downwardly curved teeth
In the original book: are developed into strong, downwardly-curved teeth
Page 89: The nerves (Pl. 27, fig. 1, r5, r6)
In the original book: The nerves (Pl. 27, fig. 1, r 5, r 6)
Page 89: outer larger pair (r1) entering the first pair
In the original book: outer larger pair (r 1) entering the first pair
Page 93. Inserted missing title (which appeared in the TOC):
Eyes and Vision.
Page 97: Male Organs of Generation.
In the original book: Reproductive System.
Male Organs.
Page 98: B. Perforatus and in the Chthamalus, that
In the original book: B. Perforatus and in the Chthamalus, that
Page 99: including the prosoma: in Pachylasma and in
In the original book: including the prosoma: in Pachylasma and in
Page 100: Female Organs of Generation.
In the original book: Female Organs.
was a paragraph title.
Page 100:
distinctly in Balanus, Tetraclita, and Coronula, the
In the original book:
distinctly in Balanus, Tetraclita, and Coronula, the
Page 103: Larva, First Stage.
In the original book this was a paragraph title.
Page 103: the larva (Pl. 29, fig. 9)
In the original book: the larva (Pl. 29, (fig. 9)
Page 104: to the eye, we see, in Scalpellum vulgare,
In the original book: to the eye, we see, in Scalpellam vulgare,
Page 104. Applied Darwin’s errata: In Balanus galeatus, in the immature
In the original book: In Balanus elongatus, in the immature
Page 105. Applied Darwin’s errata: Lepadidæ; but in Balanus galeatus
In the original book: Lepadidæ; but in Balanus elongatus
Page 106: the large second pair remains thus
In the original book: the large second pair remain thus
Page 110: Larva in the Last or Pupal Stage.
In the original book: Larva in the third or Pupal Stage.
Page 120: though enclosed fairly within the carapace
In the original book: though inclosed fairly within the carapace
Page 121: these latter may possibly be multiplied into the
In the original book: these latter may possible be multiplied into the
Page 126. Inserted missing title (which appeared in the TOC):
Act of Metamorphosis.
Page 132: separation of the three cephalic segments.
In the original book: separation of the three caphalic segments.
Page 152: Affinities, Classification, Variation.
In the original book: Affinities, Value of Characters, Variation.
Page 163: in Southern Patagonia, and near Guayaquil.
In the original book: in Southern Patagonia, and near Guyaquil.
Page 168. The first two appearances of the text: (1 to 5). In the original book: (1 to 5.)
Page 169: (3.) SECOND PROVINCE … Balanus … pœcilus*
In the original book: poecilus*
Page 170: (3.) SECOND PROVINCE … Tetraclita porosa north and south (4 and 5 and W. Indies).
In the original book: Tetraclita porosa north and south 4 and 5 and W. Indies).
Page 170: (4.) THIRD PROVINCE … Tetraclita … cœrulescens
In the original book: coerulescens
Page 172: species of Balanus abounds in individuals
In the original book: species of Balanns abounds in individuals
Page 172: specimens, said to have come from the eocene
In the original book: specimens, said to have come form the eocene
Page 174. Applied Darwin’s errata to the table. Added concavus and Hammeri to C (Red crag) and Verruca to D (Corraline crag) and corrected totals.
Page 184: with inwardly projecting filamentary appendages.
In the original book: with inwardly projecting filimentary appendages.
Page 195: BALANUS TULIPA. Bruguière. Encyclop. Meth.,
In the original book: BALANUS TULIPA. Bruguiere. Encyclop. Meth.,
Page 196: on ships’ bottoms arriving from West Africa
In the original book: on ships’-bottoms arriving from West Africa
Page 198: Galapagos Archipelago, of a uniform grayish-blue.
In the original book: Galapagos Archipelago, of a uniform greyish-blue.
Page 200: coast of America, from Monterey, in lat. 37° N.
In the original book: coast of America, from Monterez, in lat. 37° N.
Page 206: Zoolog. Journal,
In the original book: Zoolog. Zournal,
Page 211: longitudinally ribbed: colour ashy-gray tinged
In the original book: longitudinally ribbed: colour ashy-grey tinged
Page 211: along a line running from the apex
In the original book: along a line runing from the apex
Page 213: septa are united together, making a network
In the original book: septa are united together, making a net-work
Page 216: Parietes and base porose: shell white, or faintly
In the original book: Parietea and base porose: shell white, or faintly
Page 223: California; Sydney; New Zealand.
In the original book: California; Sidney; New Zealand.
Page 233: B. nubilus and cariosus
In the original book: B. nubibus and cariosus
Page 234: specimen adhering to the floating Lepas Hillii.
In the original book: specimen adhering to the floating Lepas Hilii.
Page 235: Bronn. Italiens Tertiär-Gebilde (1831)
In the original book: Bronn. Italiens Tertiar-Gebilde (1831)
Page 235: var. c. Lamarck. Animaux sans Vertèbres
In the original book: var. c. Lamarck. Animaux sans Vertebres
Page 235. Applied Darwin’s errata. Added fossil locality:
Red Crag (Sutton) Mus. S. Wood.
Pages 240-241. Italicized the abbreviation “var.” in each of the varieties.
Page 240: lower part uniform blueish-gray,
In the original book: lower part uniform blueish-grey,
Page 241: Var. (9) (an. spec.?) cirratus:
In the original book: Var. (9) (an. spec.?) cirratus:
(See also note above on abbreviation “var.”)
Page 244: part of the shell was uniformly blueish-gray.
In the original book: part of the shell was uniformly blueish-grey.
Page 246: 20. BALANUS PŒCILUS.
In the original book: 21. BALANUS PŒCILUS.
Page 248: 21. BALANUS EBURNEUS.
In the original book: 22. BALANUS EBURNEUS.
Page 248: General Appearance.—Shell conical, or almost
In the original book: General Appearance—Shell conical, or almost
Page 250: Southern Patagonia, Guayaquil, West Colombia;
In the original book: Southern Patagonia, Guyaquil, West Colombia;
Page 255. Did not apply Darwin’s errata because desired wording is not obvious.
Darwin’s errata: six lines from the top, Bal. crenatus, I have now seen a single Red Crag
specimen .5 of inch in basal diameter.
Page 263: their sutural edges are rather thick and
In the original book: their suteral edges are rather thick and
Page 264: vessels are thickly encrusted with this species,
In the original book: vessels are thickly incrusted with this species,
Page 264: enormous range and capability of resisting
In the original book: enormous range and cabability of resisting
Page 265: sent me by Krantz from the miocene formation
In the original book: sent me by Krantz from the miocence formation
Page 271: Range, Habitats, &c.—This species is
In the original book: Range, Habits, &c.—This species is
Page 278. Applied Darwin’s errata. Added fossil locality:
Red Crag (Sutton) Mus. S. Wood.
Page 279: brought from Barbados, in the West Indies,
In the original book: brought from Barbadoes, in the West Indies,
Page 282: coral-inhabiting sub-genus Creusia
In the original book: coral-inhabiting genus Creusia
Page 288: or more strictly “imperial purple;”
In the original book: or more strictly “imperial purple”;
Page 292: less prominent ridges than even B. flosculus
In the original book: less prominent ridges than even var. flosculus
Page 293. Applied Darwin’s errata: Rauville, dans le Cotentin,
In the original book: Rauville, dans le Cotantin,
Page 294: the crests for the tergal depressor muscles are
In the original book: the crests for the tergal depressores muscles are
Page 297: In both varieties, the parietes
In the original book: In both varietes, the parietes
Page 297: or deeply folded so as to be strongly ribbed
In the original book: or deeply folded so as be strongly ribbed
Page 300. Applied Darwin’s errata: internally, without parietal pores.
In the original book: internally, without parieted pores.
Page 309: for the depressor muscles are feebly
In the original book: for the depressores muscles are feebly
Page 310: Sydney, Port Fairy,
In the original book: Sidney, Port Fairy,
Page 311, footnote 104: Sydney, which I fully
In the original book: Sidney, which I fully
Page 314: crests for the depressor muscles are very feebly
In the original book: crests for the depressores muscles are very feebly
Page 331: The width of the valve and of the spur,
In the original book: Te width of the valve and of the spur,
Page 335: Die Südafrikanischen Mollusken
In the original book: Die Sudafrikanischen Mollusken
Page 337: judging from some dried specimens,
In the original book: judging from some dryed specimens,
Page 338: rostral or lateral depressor muscles
In the original book: rostral or lateral depressores muscles
Page 347: form of a network, or of separate tubes
In the original book: form of a net-work, or of separate tubes
Page 353, footnote 113: I have seen two specimens of
In the original book: I have seen too specimens of
Page 357: forms a mere border to the occludent ledge;
In the original book: forms a mere border to the occludent lodge;
Page 357: between this genus and Creusia, it is probable
In the original book: between this genus and Creusia, it is probable
Page 369: almost obliterated by the encrusting coral.
In the original book: almost obliterated by the incrusting coral.
Page 376: Under these circumstances I have thought it best
In the original book: Under these circustances I have thought it best
Page 376: 1. CREUSIA SPINULOSA. Pl. 13,
In the original book: CREUSIA SPINULOSA. Pl. 13,
Page 376: CREUSIA SPINULEUSE. De Blainville. Dict. Sc. Nat.,
In the original book: CREUSIE SPINULEUSE. De Blainville. Dict. Sc. Nat.,
(de Blainville spells it CREUSIA.)
Page 383: In Chelonobia, the parietes are remarkably thick;
In the original book: In Chelonobia, the parieties are remarkably thick;
Page 405: (see a black dot (d´) in section, fig. 7)
In the original book: (see a black dot (d´) in section 7)
Page 416:the alæ are only 1/5th or 1/6th of the thickness
In the original book: the alæ are only 1-5th or 1-6th of the thickness
Page 423: Eine tiefe Furche oder Spalte
In the original book: Eine tiefe Furcle oder Spalte
Page 423: von oben herab bis zur Hälfte
In the original book: von oben herab bis zur Halfte
Page 424:9. Genus—PLATYLEPAS. Pl. 17, fig. 1 a-2 b.
In the original book: 9. Genus—PLATYLEPAS. Pl. 17, fig. 1 and 2.
Page 424: the whole rostrum is pushed a little on one side
In the original book: the whole rostum is pushed a little on one side
Page 431: In numerous other respects Tubicinella
In the original book: In numerous other respects Tubinicella
Page 431: 1. TUBICINELLA TRACHEALIS.
In the original book: 4. TUBICINELLA TRACHEALIS.
Page 431, footnote 127: the specific name either of major or minus
In the original book: the specific name either of minor or majus
Page 432: the reverse of that usual with sessile cirripedes
In the original book: the reverse of that usual with sessile-cirripedes
Page 440: 1. XENOBALANUS GLOBICIPITIS.
In the original book: XENOBALANUS GLOBICIPITIS.
Pages 455-6. Note that under species Chthamalus stellatus, both varieties (a) and (c) have the name: communis. The original book reads that way.
Page 465: 6. CHTHAMALUS HEMBELI. Pl. 18,
In the original book: 6. CHTHAMALUS HEMBELI. Tab. 18,
Page 467: disintegration, causing the shell to be ashy gray.
In the original book: disintegration, causing the shell to be ashy grey.
Page 471: from the projecting, extremely rugged, overlapping,
In the original book: from the projecting, extremely rugged, over-lapping,
Page 475: and these occur in Catophragmus
In the original book: and these occur Catophragmus
Page 487: moderately or slightly depressed; colour gray.
In the original book: moderately or slightly depressed; colour grey.
Page 488: as seen, when corroded, in fig. 4 d,
In the original book: as seen, when corrroded, in fig. 4 d,
Page 492: Diadema bifidum … ‘Italiens Tertiär-Gebilde’
In the original book: ‘Italiens Tertiar-Gebilde’
Page 492: Balanus ostrearum … Organic Remains of the Cretaceous Group,’
In the original book: Organic Remains of the Cretacean Group,’
Page 493: B. costatus … is a synonym of B. porcatus
In the original book: is a synonym of B. porcatns
Page 493: B. ornatus, Münster, ‘Beiträge zur Petrifact.,’
In the original book: B. ornatus, Münster, ‘Beitrage zur Petrifact.,’
Page 493: B. pectinarius, Bronn (‘Italiens Tertiär-Gebilde,’
In the original book: B. pectinarius, Bronn (‘Italiens Tertiar-Gebilde,’
Page 493: B. porosus … this species in Münster’s ‘Beiträge.’
In the original book: this species in Münster’s ‘Beitrage.’
Page 494: B. pustularis … in Münster’s ‘Beiträge,’
In the original book: in Münster’s ‘Beitrage,’
Page 494: B. cylindraceus … ‘Italiens Tertiär-Gebilde,’
In the original book: ‘Italiens Tertiar-Gebilde,’
Page 494: B. Uddevallensis, Linnæus,
In the original book: B. Uddewallensis, Linnæus,
Page 496: members of the Chthamalinæ, though abnormal in
In the original book: members of the Chthamalinæ, though abnormal in
Page 503: under an edge, z (much foreshortened in T′
In the original book: under an edge, z (much fore-shortened in T′
Page 504: the umbo of growth matches the umbo of the four
In the original book: the umbo of growth matches the umbo: of the four
Page 517: for themselves, could be pried vertically up
In the original book: for themselves, could be prised vertically up
Page 518: BALANUS VERRUCA. Bruguière. Encyclop. Meth., 1789;
In the original book: BALANUS VERUCA. Bruguière. Encyclop. Meth., 1789;
Page 526: The moveable tergum has its upper articular ridge
In the original book: The moveable tergum has its upper articular ridge
Page 529, footnote 141: Crustacés fossiles du terrain Crétacé
In the original book: Crustacés fossiles du terrain Crétacée
Page 529, footnote 142:‘Archives du Muséum’
exemples
comment à des
reconnaître
In the original book: ‘Archives du Museum’
examples
comment a des
reconnaitre
Page 530 title: FEMALE
In the original book: FEMALE
Page 538: or the whole anterior part of the animal
In the original book: or the whole anterior part of theanimal
Page 552: although constantly agitated.
In the original book: although constanty agitated.
Page 553: a considerable residuum is left
In the original book: a considerable residium is left
Page 562: down the sides of the sack of the female till
In the original book: down the sides of the sack of female till
Page 562: mere flap, and only two pairs of cirri exist
In the original book: mere flap, and only two pair of cirri exist
Page 562: thorax with four pairs of minute, modified cirri,
In the original book: thorax with four pair of minute, modified cirri,
Page 563: ORDER II.—ABDOMINALIA.
In the original book: ORDER II.—CIRRIPEDIA ABDOMINALIA.
Page 564: close to Alcippe amongst the Lepadidæ
In the original book: close to Alcippe amonst the Lepadidæ
Page 566 title: FEMALE
In the original book: FEMALE
Page 581: whole dorsal surface, and the overlapping sides
In the original book: whole dorsal surface, and the over-lapping sides
Page 582, footnote 152: antennæ, I may here add those of Cryptophialus,
In the original book: antennæ, I may here add those of Crystophialus,
Page 583: to enclose and protect the antennæ
In the original book: to inclose and protect the antennæ
Page 587: compound structure of the vesicula seminalis
In the original book: compound structure of the visicula seminalis
Page 588: so peculiar, the hermaphrodite condition,
In the original book: so peculiar, the hermaphodite condition,
Page 596: the two threads enclosing the cement-ducts
In the original book: the two threads inclosing the cement-ducts
Page 601: one common membrane, lined of course by corium;
In the original book: one common membrane, lined of course by coriumi;
Page 604: it does not enclose the thorax or mouth
In the original book: it does not inclose the thorax or mouth
Page 606: Sub-Familia 1. BALANINÆ.
In the original book: Sub-familia 1. BALANINÆ.
Page 607: 2. Sub-Genus—ACASTA
In the original book: 2. Sub-genus—ACASTA
Page 607: 6. Sub-Genus—CREUSIA
In the original book: 6. Sub-genus—CREUSIA
Page 607, 9.: quæ basin membranaceam extrorsùs
In the original book: quæ basin membranacean extrorsùs
Page 608: Sub-Familia 2. CHTHAMALINÆ.
In the original book: Sub-familia 2. CHTHAMALINÆ.
Page 611: ORDO I. THORACICA.
In the original book: ORDER THORACICA.
Page 611: Sub-Fam. BALANINÆ.
In the original book: Sub-fam. BALANINÆ.
Page 613, 12.: Tergo lato, pæne æquilaterali.
In the original book: Tergo lato, pœne æquilaterali.
Page 614, 17.: Tab. 4, fig. 3,
In the original book: Tab. 4, fig. 4,
Page 614, 18.: Americâ septent. et meridionali.
In the original book: Americâ septent, et meridionali.
Page 614, 22.: superioribus lævibus, leniter arcuatis,
In the original book: superioribus lævibusleniter arcuatis,
Page 615, 27.: et Indiâ occident. et Africâ meridionali.
In the original book: et Indiâ occident, et Africâ meridionali.
Page 617, 42.: basi poris magnis perforatâ:
In the original book: basi poris maguis perforatâ:
Page 618: Sub-Genus—ACASTA
In the original book: Subgenus—ACASTA
Page 618, 1.: scuti cristâ articulari
In the original book: scuti cris â articulari
Page 619, 4.: longitudinaliter costatâ, aut corrosâ
In the original book: longitudinaliter costatá, aut corrosâ
Page 622, 2.: et Australiâ septent.; testudinibus affixa.
In the original book: et Australiâ septent; testudinibus affixa.
Page 622: 3. Chelonobia patula, Ranzani. (Darwin,
In the original book: 3. Chelonobia patula, Ranzani. (Drawin,
Page 624: 1. Xenobalanus globicipitis, Steenstrup.
In the original book: 1. Xenobalanis globicipitis, Steenstrup.
Page 624: Sub-Fam. CHTHAMALINÆ.
In the original book, this title was missing.
Page 634: 11. Genus—POLLICIPES.
In the original book: 11.—Genus—POLLICIPES.
Page 637: 18. Pollicipes validus … Foss.— … Scaniâ et Maestricht.
In the original book: Scaniâ et Mäestricht.
Page 638: [Sectio ††.]
In the original book: [†† Scuta, aut longitudinaliter aut transversè (i. e. secundum lineas
incrementi) costata.]
Page 640: ORDO II. ABDOMINALIA.
In the original book: ORDER II. ABDOMINALIA.
Page 640: ORDO III. APODA.
In the original book: ORDER III. APODA.
Page 641: PLATE 1.
BALANUS TINTINNABULUM.
In the original book: PLATE 1.
Genus—BALANUS.
Page 641. In plates 1 and 2, the word “Fig.” appeared on a line by itself above the figure numbers. It has been moved to be more consistent with the other plate descriptions.
Page 642, Plate 3, 2 c and 2 d: scutum, internal and external views.
In the original book: scutum and tergum, internal views.
Page 643, Plate 5, 2 d: Stutchburii.
In the original book: Stutchburi.
Page 643, Plate 5, 2 i: Stutchburii.
In the original book: Stutchburi.
Page 643, Plate 5, 2 m, 2 n, and 2 o: Stutchburii.
In the original book: Stutchburi.
Page 644, Plate 9: Sub-Genus—ACASTA.
In the original book: Sub-genus—ACASTA.
Page 647, Plate 14: Genera—CREUSIA AND CHELONOBIA.
In the original book: Genera—CREUSIA, CHELONOBIA.
Page 650, Plate 16: Of the letters in italics, …
In the original book, this sentence was not aligned differently from the surrounding items.
Page 650, Plate 16: open beneath, and filled up by the epidermis
In the original book: open beneath, and filled up by the epiderdermis
Page 651, Plate 17, 2 a: A, rostrum;
In the original book: A, rostum;
Page 653, Plate 19, 4 b, 4 c, 4 d: Chamæsipho scutelliformis,
In the original book: Chamæsipho columna,
Page 657, Plate 22, 8: appear like a bilobed lower lip;
In the original book: appear like a bi-lobed lower lip;
Page 658, Plate 23. Title inserted to match plate:
Genera—ALCIPPE AND CRYPTOPHIALUS.
Page 660, Plate 24. Title inserted to match plate:
Genera—CRYPTOPHIALUS AND PROTEOLEPAS.
Page 662, Plate 25. Title inserted to match plate:
Genera—PROTEOLEPAS AND BALANUS.
Page 664, Plate 26: STRUCTURE OF THE MOUTH AND THORAX.
In the original book: STRUCTURE OF THE MOUTH, &C.
Pages 664-6, Plate 26. The original book enclosed all the letters of reference in parentheses for this Plate and Plate 29 only. The parentheses have been removed for consistency with the descriptions of all the other plates, without noting the individual changes.
Page 669, Plate 28, 4 a: k, t, … the corresponding branch
In the original book: the corrresponding branch
Page 669, Plate 28, 4 b: and that (gh) represented in fig. 4 a;
In the original book: and that (g h) represented in fig. 4 a;
Page 669, Plate 29: CIRRI AND LARVÆ, FIRST STAGES.
In the original book: CIRRI: LARVA, FIRST STAGE.
Pages 669-71, Plate 29. The original book enclosed all the letters of reference in parentheses for this Plate and Plate 26 only. The parentheses have been removed for consistency with the descriptions of all the other plates, without noting the individual changes.
Page 670, Plate 29: letters apply to all the figures, 1 to 6.
In the original book: letters apply to all the figures, 1 to 5.
Page 674. In Darwin’s errata on Page 174: total in the Coralline Crag ten,
In the original book: total in the Coralline Crag nine,
Page 675: Adna … Anglica, 360.
In the original book: anglica, 360.
Page 675: Baer, Von, on morphological differentiation, 19.
In the original book: Baer, Von, on morphological differentation, 19.
Page 676: d’Orbignii, 195.
In the original book: D’Orbignii, 195.
Page 676. The index has been changed due to the errata on pages 104 and 105 changing the references from Balanus elongatus to Balanus galeatus.
Page 676: (Balanus) intertextus, 518.
In the original book: intertextus, 518.
Page 677: (Balanus tintinnabulum) varieties of, 201.
In the original book, the “varieties” listing was at the same level of indentation as the line before it.
Page 677: (Balanus) tulipiformis, 204.
In the original book: tulipiformis, 204.
Page 677: Chamæsipho, genus, 470.
In the original book: Chamœsipho, genus, 470.
Page 678: Conopea … ovata, 218.
In the original book: ovala, 218.
Page 678: Creusia rayonnante, 362.
In the original book: Creusie rayonnante, 362.
Page 679: Daracia … monticulariæ, 372.
In the original book: monitculariœ, 372.
Page 680: Lepas … fistulosus, 231.
In the original book: fistulosa, 231.
Page 681: Male organs … of Cryptophialus minutus, 584.
In the original book: of Crytophialus minutus, 584.
Page 681. Index entries for Ochthosia and Octomeris were put in alphabetical order.
Page 681: Otolithes absent in Crustacea, 97.
In the original book: Otolites absent in Crustacea, 97.
Page 681: Penis, probosciformed, of wonderful length
In the original book: Penis, pubosciformed, of wonderful length
Page 683: Stomopoda, affinities to cirripedes, 19.
In the original book: Stomapoda, affinities to cirripedes, 19.
Page 683. Index entry for Tetraclita was put in proper alphabetical order.
Page 684. Index entry for Verrucidæ was put in proper alphabetical order.
Pages 685-714. The titles of the plates have been changed. In the original book, the word plate was abbreviated, i. e. “Pl. II.” whereas in this ebook it is “Plate II.” Also, the individual figure captions have been changed. In the original book, they were just the figure number, e. g. “1 a”, whereas in this book, the plate number and labels are included, e. g. “Plate II., figure 1 a.”
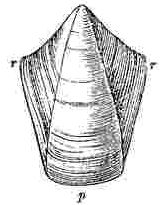
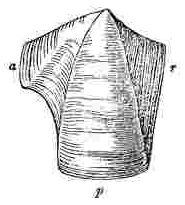
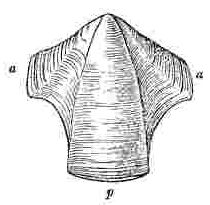
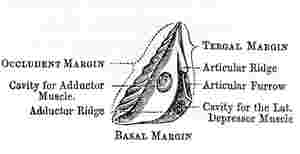
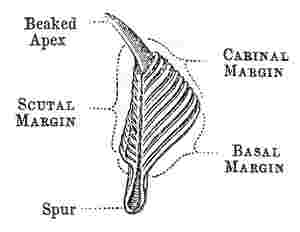
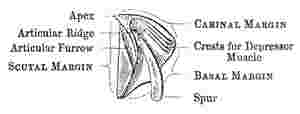
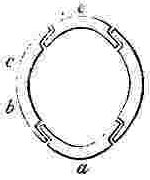
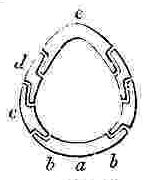
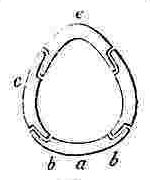


















































")





































































































































































































































































































































.")
.")
.")
.")








































































































































.")
.")
.")






