Transcriber's Notes.
This file was derived from scanned images. With the exception of
two typographical errors that were corrected, the original text and
copies of the included illustrations are presented. The smaller graphic
shown as Figure 2 was processed to remove moire effects when viewed at smaller
sizes. Clicking on the Figures will allow the user to view a copy of the
original scanned image from which it was derived.
Typographical Errors Corrected:
Page 589, Paragraph 2: Tepalcatapec => Tepalcatepec
Page 592, Paragraph 2: ground color => ground-color
In the original, Figure 1 had the notation X5 after the caption to let the
reader know that the image was enlarged by a factor of five. As the illustration
in the HTML page will display at different sizes based on the resolution of the
monitor and the size the browser window, it was decided to remove that notation
and the viewer would derive the measurements from the body of the paper.
[587]

University of Kansas Publications
Museum of Natural History
Vol. 10, No. 9, pp. 587-598, 2 figs.
 May 2, 1960
May 2, 1960
A New Subspecies of Lizard,
Cnemidophorus sacki, from Michoacán, México
University of Kansas
Lawrence
1960
[588]
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Robert W. Wilson
Volume 10, No. 9, pp. 587-598, 2 figs.
Published May 2, 1960
University of Kansas
Lawrence, Kansas
PRINTED IN
THE STATE PRINTING PLANT
TOPEKA, KANSAS
1960
28-2494
[589]
A New Subspecies of Lizard,
Cnemidophorus sacki, from Michoacán, México
BY
WILLIAM E. DUELLMAN
The systematic status of the populations of lizards assignable to
Cnemidophorus sacki in western México (Sonora southward to
Jalisco) has been reviewed in detail by Zweifel (1959, Bull. Amer.
Mus. Nat. Hist, 117 (2):57-116), who stated that the use of the
specific name sacki for the western populations rests upon a reasonable,
but as yet unproved, assumption that intergradation occurs.
Although Zweifel examined specimens of the nominal subspecies
from the upper Balsas Basin, he did not study specimens from the
intervening area—the Tepalcatepec Valley in Michoacán.
Field studies and the examination of large series of preserved
specimens of Cnemidophorus from the Tepalcatepec Valley show
the presence of Cnemidophorus sacki in the region between the
ranges of C. sacki sacki in the upper Balsas Basin and C. sacki
occidentalis in Jalisco and southern Nayarit. Furthermore, the
populations of sacki inhabiting the Tepalcatepec Valley have characters
of scutellation and coloration that distinguish them from
other described subspecies of sacki. In recognition of the important
contributions to the systematics of the genus Cnemidophorus made
by Dr. Richard G. Zweifel, I propose that the subspecies of Cnemidophorus
sacki in the Tepalcatepec Valley be named as follows:
Cnemidophorus sacki zweifeli new subspecies
Holotype.—University of Michigan Museum of Zoology No. 119542, from
Capirio, Michoacán, México (185 meters), an adult male, one of a series
collected on June 13, 1958, by William E. Duellman, Jerome B. Tulecke, and
John Wellman. Original number WED 12310.
Paratopotypes.—UMMZ Nos. 119536-119541 and 119543-119550.
Diagnosis.—A race of Cnemidophorus sacki characterized by large size
(more than 130 mm. snout-vent length in males), approximately 106 dorsal
granules around the midbody, about 41 femoral pores, and a dorsal color-pattern
in adult males consisting of lateral and dorsolateral rows of spots,
paravertebral rows fused with middorsal light green area at least anteriorly,
and pink throat having a median light blue spot or transverse band.
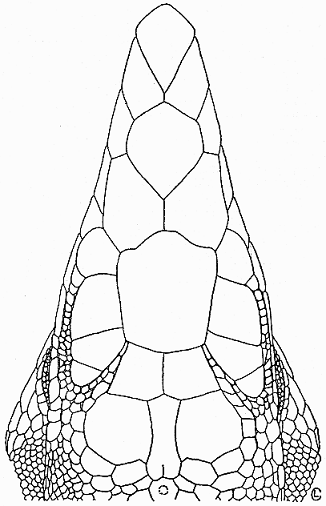
Description of Holotype.—Snout-vent length, 128 mm.; tail length, 252
mm.; tail/body ratio, 1.97; scutellation typical of sacki—four supraoculars,
enlarged postantebrachials, and enlarged mesoptychials; 107 granules around
midbody (excluding enlarged ventrals); 45 femoral pores; three enlarged
preanal scales; supraorbital semicircle-series extending anteriorly to posterior
edge of frontal (Fig. 1).
[590]
 Click on image to view larger sized version.
Fig. 1.
Click on image to view larger sized version.
Fig. 1. Top view of the head of the holotype of
Cnemidophorus sacki
zweifeli (UMMZ 119542) showing scutellation.
[591]
Top of head and nape dusty brown; tip of rostral and lateral edges of
superciliaries dark cream-color; upper labials and sides of head anterior to
eyes cream-color, mottled with blue; lower labials and postocular region pale
blue; mental, postmental, and sublabials cream-color. Upper surfaces of
forelimbs dull bluish gray, spotted with pale greenish yellow; dorsal surfaces
of proximal one-fourth of tail light brownish gray turning to pale orange-brown
posteriorly; lateral surfaces of tail bluish gray anteriorly and creamy brown
posteriorly. Nuchal region light bluish gray; flanks dark gray; dorsal ground-color
dark brown, somewhat paler posteriorly. Body having a row of cream-colored
spots in place of a lateral stripe, and another row in place of dorsolateral
stripe; dorsally, large diffuse tan or light green spots partially fused and
tending to form irregular transverse markings. Chin to posterior end of sublabials
pale pink, bordered posteriorly by bluish white area, and then pink.
Mesoptychials, under surfaces of hind limbs, and belly cream-colored; anterior
edges of belly scales dark blue; lateral two rows of ventrals on posterior two-thirds
of body dark blue having light blue or cream-colored spots. Under
surfaces of forelimbs bluish cream; ventral surface of tail cream-colored.
Variation in Size and Scutellation.—The largest male has a snout-vent
length of 132 mm., the largest female, 114 mm., and the smallest juvenile, 34
mm. The number of dorsal granules at the midbody varies from 91 to 117
(106.2 ± 0.43); the ratio of the number of granules between the paravertebral
stripes to the number of granules around the body (PV/GAB) varies from
0.064 to 0.157 (0.097 ±0.007); the number of femoral pores varies from
32 to 49 (41.1 ± 0.20). Usually there are only three enlarged preanals, but
18 specimens have a somewhat enlarged scale anterior to the normal complement
of three. In 15 specimens the supraorbital semicircle-series terminate
short of the posterior edge of the frontal; in the others the series reach the
frontal.
Variation in Coloration.—The coloration of juveniles and subadults varies
little; large adults vary considerably especially in the amount of diffusion of
the light green middorsal area. In some individuals the vertebral pale area
does not include the paravertebral spots; in other individuals the pale area
includes not only the paravertebral rows, but, at least anteriorly, the dorsolateral
rows. In large males of about equal size (and collected at the same
time) there is considerable variation in the amount of blue on the belly. In
a few of the males the belly is white with only the anterior edge of each scale
blue; in some only the lateral rows of ventrals on the posterior two-thirds of
the body are blue; in others all of the posterior two-thirds of the belly is blue.
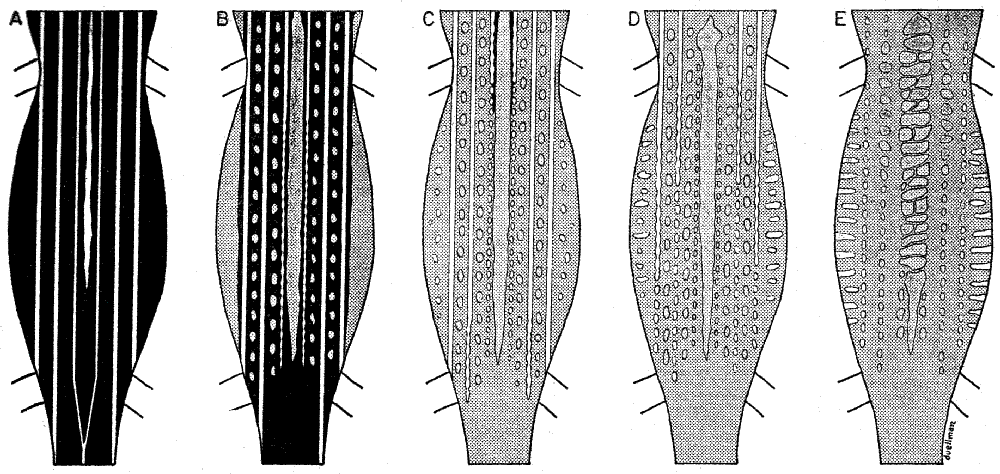
Ontogenetic Change in Color Pattern.—The metamorphosis of
color pattern in Cnemidophorus sacki zweifeli results in the dorsal
ground-color becoming paler with age, the replacement of the
stripes by spots, and finally in large males the suffusion of these
spots.
A single hatchling (UMMZ 114732) is available; this specimen
has a prominent umbilical scar and a snout-vent length of 34 mm.
The top of the head is olive brown; the dorsal surfaces of the limbs
are dark brown with cream mottling; the dorsal ground-color is
brownish black; this is paler on the lower flanks. The lateral and
dorsolateral stripes are cream-colored; the paravertebral stripes are
[592]
white. There is a faint, diffuse vertebral stripe anteriorly (Fig. 2 A).
The throat and undersides of the limbs and tail are cream-colored;
the belly is bluish white. In life the stripes were pale yellowish
green, and the tip of the tail was pink.
In larger individuals the dorsal ground-color is dark brown; the
lower flanks are grayish tan. Light brown diffuse spots are present
in the lateral and dorsolateral dark fields. The tan vertebral stripe
is diffuse and nearly fills the paravertebral dark fields; the paravertebral
stripes are faint posteriorly; throughout their length they are
scalloped—the beginning of their fragmentation into spots (Fig.
2 B).
In subadults (± 80 mm. snout-vent length) the paravertebral
stripes are fragmented into spots posteriorly. Also, the dorsolateral
stripes in some individuals are fragmented posteriorly. The dorsolateral
dark fields are somewhat paler than the lateral dark fields.
Cream-colored spots are present on the flanks. The mottling on
the thighs tends towards the formation of light spots (Fig. 2 C).
In small adults (± 100 mm. snout-vent length) the paravertebral
stripes are entirely fragmented into spots. The lateral and dorsolateral
stripes are broken into spots posteriorly. The middorsal pale
area (formed by the suffusion of the vertebral stripe) and paravertebral
and dorsolateral rows of spots are pale green. The cream-colored
spots on the flanks are expanded to form vertical bars
(Fig. 2 D).
Large adult males (± 120 mm. snout-vent length) have all of
the stripes fragmented into spots. The diffuse middorsal area is
expanded and encloses the paravertebral rows of spots. The pale
spots present in the dark fields in smaller individuals are either
absent or fused with spots resulting from the fragmentation of the
stripes (Fig. 2 E).
Sexual dimorphism.—Males attain a larger size (known maximum
snout-vent length of 132 mm., as compared with 114 mm. in females).
Males have larger but not more numerous, femoral pores,
blue bellies, and pink and blue throats, whereas females are unicolor
creamy white ventrally. The more nearly complete metamorphosis
of color pattern exhibited by adult males probably is
correlated with their large adult size. Large females retain complete
lateral and dorsolateral stripes. The jowls of breeding males
are swollen.
[593]
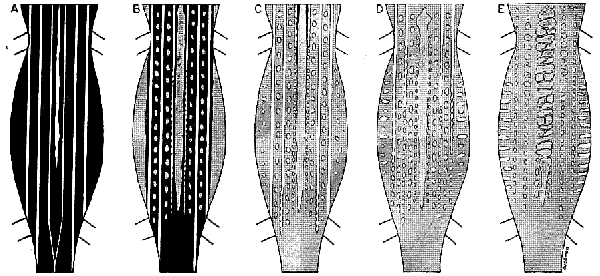
Click on image to view larger sized version. |
Fig. 2. Diagrammatic representation of ontogenetic change in color pattern in Cnemidophorus sacki zweifeli: A—hatchling, 34
mm. snout-vent length; B—juvenile, 55 mm. snout-vent length; C—subadult male, 80 mm. snout-vent length; D—small adult male,
100 mm. snout-vent length; E—large adult male, 120 mm. snout-vent length.
[594]
Geographic variation.—No noticeable geographic variation in
this subspecies is evident in the series from the Tepalcatepec
Valley. However, lizards from eastern Michoacán (Chinapa, 6
km. N of Tafetan, 6 km. S of Tzitzio, and 19 km. S of Tzitzio) differ
slightly from those from the Tepalcatepec Valley; the eastern
specimens have fewer dorsal granules and femoral pores, and a
higher ratio of dorsal granules between the paravertebral stripes
to the number of granules around the body (see Tables 1-3). No
large adult males are present in the eastern series; the subadults
and small adult males have color patterns like lizards of similar
size from the Tepalcatepec Valley. The largest male from the east
has a snout-vent length of 110 mm., rows of pale spots, and no trace
of brown and tan cross-bars. Specimens of Cnemidophorus sacki
sacki of equal size from Guerrero, Morelos, and Puebla in the upper
Balsas Basin have a tan dorsum with dark brown cross-bars. The
localities in eastern Michoacán are intermediate geographically
between the Tepalcatepec Valley and the known range of the
nominal subspecies in the upper Balsas Basin. In characters of
scutellation specimens from the east are intermediate between C.
sacki sacki and C. sacki zweifeli in the Tepalcatepec Valley. However,
in coloration the lizards from the east are like those from the
Tepalcatepec Valley, but differ distinctly from the nominal subspecies.
Therefore, the eastern series is referred to the subspecies
C. sacki zweifeli.
Comparisons.—Four other species of Cnemidophorus occur in
the Tepalcatepec Valley with Cnemidophorus sacki zweifeli. Of
these, C. calidipes has a maximum snout-vent length of 79 mm., 66
to 86 dorsal granules, and a light brown dorsum with pale blue
spots and vertical bars; C. communis communis has a maximum
snout-vent length of 135 mm., 105 to 144 dorsal granules, and a
greenish tan dorsum with yellow spots; C. deppei infernalis has a
maximum snout-vent length of 84 mm., 91 to 120 dorsal granules,
and a striped pattern throughout life; and C. lineatissimus exoristus
has a maximum snout-vent length of 98 mm., 108 to 135 dorsal
granules, and a middorsal yellow stripe and vertical bars on the
flanks. Both calidipes and communis are like sacki in possessing
four enlarged supraoculars and enlarged postantebrachials, whereas
deppei and lineatissimus have three enlarged supraoculars and
granular postantebrachials. Juveniles of calidipes and sacki are
alike in coloration but different in the extent of the supraorbital
semicircle-series. In calidipes the supraorbital semicircle-series usually
are complete, whereas in sacki the series never extended anterior
to the posterior edge of the frontal.
[595]
Table 1.—Variation in the Number of Dorsal Granules
in Three Subspecies of Cnemidophorus sacki
| Key for Table: |
| No. = Number of Specimens | SD = Standard Deviation |
| SE = Standard Error | C/V = Coefficient of Variation |
| Population | No. | Range | Mean | SD | SE | C/V |
sacki sacki | | | | | | |
| Entire Sample | 106 | 88-105 | 96.3 | 4.16 | 0.40 | 3.92 |
| Puebla: Tehuitzingo | 22 | 88-103 | 95.1 | 4.18 | 0.89 | 4.39 |
| Guerrero: Chilpancingo | 23 | 90-105 | 95.8 | 3.86 | 0.80 | 4.02 | |
| Guerrero: Mexcala | 22 | 90-102 | 96.5 | 3.56 | 0.76 | 3.69 | |
| Morelos | 39 | 89-105 | 97.2 | 4.56 | 0.73 | 4.69 | |
| | | | | | | |
| sacki zweifeli | | | | | | |
| Entire Sample | 191 | 91-117 | 106.2 | 5.98 | 0.43 | 3.13 | |
| Michoacán: Tafetan | 21 | 91-116 | 101.4 | 8.04 | 1.75 | 7.92 | |
| Michoacán: Apatzingán | 170 | 95-117 | 106.8 | 5.42 | 1.32 | 3.19 | |
| | | | | | | |
| sacki occidentalis[A] | 62 | 97-118 | 106.3 | 4.72 | 0.60 | 7.61 |
Table 2.—Variation in the Number of Femoral Pores
in Three Subspecies of Cnemidophorus sacki
| Key for Table: |
| No. = Number of Specimens | SD = Standard Deviation |
| SE = Standard Error | C/V = Coefficient of Variation |
| Population | No. | Range | Mean | SD | SE | C/V |
sacki sacki | | | | | | |
| Entire Sample | 106 | 32-44 | 36.2 | 2.42 | 0.25 | 2.28 |
| Puebla: Tehuitzingo | 22 | 33-41 | 36.7 | 2.36 | 0.50 | 6.43 |
| Guerrero: Chilpancingo | 23 | 32-39 | 35.7 | 2.15 | 0.45 | 6.02 |
| Guerrero: Mexcala | 22 | 33-44 | 37.5 | 2.59 | 0.55 | 6.91 |
| Morelos | 39 | 32-40 | 35.4 | 2.19 | 0.35 | 6.18 |
sacki zweifeli | | | | | | |
| Entire Sample | 189 | 32-49 | 41.1 | 2.77 | 0.20 | 1.46 |
| Michoacán: Tafetan | 19 | 33-43 | 38.1 | 2.61 | 0.60 | 6.85 |
| Michoacán: Apatzingán | 170 | 2-49 | 41.4 | 2.58 | 0.62 | 1.52 |
sacki occidentalis | 67 | 32-45 | 38.8 | 2.46 | 0.30 | 3.67 |
[596]
Table 3.—Ratio of Number of Granules Separating Paravertebral
Stripes to Granules Abound Midbody (PV/GAB) in Three Subspecies of Cnemidophorus sacki
| Key for Table: |
| No. = Number of Specimens | SD = Standard Deviation |
| SE = Standard Error | C/V = Coefficient of Variation |
| Population | No. | Range | Mean | SD | SE | C/V |
sacki sacki | | | | | | |
| Entire Sample | 72 | 0.101-0.205 | 0.154 | 0.052 | 0.006 | 0.073 |
| Puebla: Tehuitzingo | 12 | 0.140-0.205 | 0.169 | 0.052 | 0.015 | 0.433 |
| Guerrero: Chilpancingo | 16 | 0.120-0.192 | 0.157 | 0.021 | 0.005 | 0.131 |
| Guerrero: Mexcala | 21 | 0.142-0.180 | 0.159 | 0.031 | 0.007 | 0.147 |
| Morelos | 23 | 0.101-0.180 | 0.137 | 0.023 | 0.005 | 0.100 |
| | | | | | | |
| sacki zweifeli | | | | | | |
| Entire Sample | 105 | 0.064-0.157 | 0.097 | 0.070 | 0.007 | 0.067 |
| Michoacán: Tafetan | 21 | 0.103-0.157 | 0.128 | 0.039 | 0.009 | 0.186 |
| Michoacán: Apatzingán | 84 | 0.064-0.126 | 0.089 | 0.051 | 0.005 | 0.060 |
| | | | | | | |
| sacki occidentalis | 50 | 0.086-0.183 | 0.130 | 0.021 | 0.003 | 0.042 |
From the geographically adjacent populations of sacki (sacki and
occidentalis), zweifeli differs in coloration and scutellation (see
Tables I-III). Cnemidophorus sacki sacki has a dorsal pattern in
adult males of dark brown cross-bars on a tan ground-color. Both
occidentalis and zweifeli have variable, diffuse color patterns in
large adults, but zweifeli differs from occidentalis in having a blue
spot on the pink throat.
Ecology.—In the arid Tepalcatepec Valley Cnemidophorus sacki
zweifeli lives at elevations of 160 to 1300 meters. In the lower parts
of the valley the lizards live primarily in open scrub forests, characterized
by deciduous trees offering only partial shade from the
sun, especially during the prolonged dry season. Common trees in
this scrub forest are Acacia cymbispina, Cercidium plurifoliolatum,
Mimosa distachya, and Prosopis juliflora.
During the dry season (November through May) adult males
apparently aestivate; several large series collected in the winter
include only subadults and females. This absence of males is
corroborated by personal observations in the Tepalcatepec Valley
in April and May. In the summer rainy season the lizards are active
in the morning and again in the late afternoon; only Cnemidophorus
calidipes is active during the heat of the midday. In some
[597]
areas of the scrub forest Cnemidophorus sacki zweifeli is found in
association with Cnemidophorus communis communis. Throughout
the scrub forest C. sacki zweifeli occurs with C. deppei infernalis.
In some of the more dense scrub forest, where C. sacki zweifeli
is not so abundant as in the more open forest, it has been taken
with C. lineatissimus exoristus. In the open Acacia-Cercidium associations
on the valley floor C. sacki zweifeli occurs with C. calidipes.
This subspecies is not restricted to the scrub forest. On the
lower slopes of the Cordillera Volcanica in Michoacán C. sacki
zweifeli has been collected in open pine-oak forest near Zirimicuaro
and Ziracuaretiro.
Distribution.—Cnemidophorus sacki zweifeli inhabits the valley
of the Río Tepalcatepec in Michoacán and probably extreme southwestern
Jalisco, and the western part of the Balsas Basin in
Michoacán. No specimens have been seen from extreme western
Guerrero, but C. s. zweifeli may occur there.
Specimens examined.—Catalogue numbers are preceded by abbreviations of
the name of the institution as listed in the acknowledgements.
Cnemidophorus sacki occidentalis, 22 specimens, as follows: Jalisco: 8
km. E of Ameca, UMMZ 102045; 7 km. SE of Ameca, UMMZ 102046; Autlán,
UMMZ 102044, 102219-21; 7 km. ESE of El Arenal, UMMZ 114736 (2);
San Gabriel, UMMZ 102040, 102042-3. Michoacán: 2 km. ESE of Jiquilpan,
UMMZ 117557 (3). Nayarit: Ixtlán del Río, UMMZ 104747; San José de la
Conde, UMMZ 102047 (4); 5 km. N of Santa Isabel, UMMZ 102048-50.
Cnemidophorus sacki sacki, 108 specimens, as follows: Guerrero: Chilpancingo,
UMMZ 72426 (8), 73937 (7), 88422 (4); 8 km. W of Chilpancingo,
UMMZ 119144 (4); 5 km. N of Iguala, UMMZ 114712 (11); 15 km. N of
Iguala, UMMZ 99039; Mexcala, UMMZ 114711 (10); 8 km. S of Taxco,
UMMZ 114709 (2). Morelos: Amacuzac, UMMZ 114716; 3 km. S of Cuautla,
UMMZ 99031 (9), 99917 (11): Río Cuautla, UMMZ 99038 (6); 5 km.
S of Temixco, UMMZ 114718 (12); Puebla: 5 km. SE of Izúcar de Matamoros,
UMMZ 112650 (4); 13 km. SE of Izúcar de Matamoros, UMMZ 117497 (2);
3 km. NW of Tehuitzingo, UMMZ 114714 (10); 1 km. N of Teyuca, UMMZ
114713 (6).
Cnemidophorus sacki zweifeli, 207 specimens, as follows: Michoacán:
Apatzingán, CNHM 36966-8, 38969, 38971 (18), 38972 (50), UIMNH
36772-7, USNM 135967-8, 135970, 135972-3; 4 km. E of Apatzingán, UMMZ
85412; 6.5 km. E of Apatzingán, UMMZ 114731 (5); 5 km. W of Apatzingán,
KU 29289-90, 29293-7; 10 km. W of Apatzingán, UMMZ 114730 (4); 12.3
km. S of Apatzingán, UMMZ 112647; 16 km. S of Apatzingán, KU 29298;
14 km. SSW of Apatzingán, KU 29735, 29746, 29750-2; 10 km. W of Buenavista,
UMMZ 114719 (3); Capirio, UMMZ 112643, 114722 (2), 114733,
119536-50; 4 km. N of Capirio, UMMZ 112644 (3), 112645; 5.6 km. N of
Capirio, UMMZ 114732; 2 km. S of Charapendo, UMMZ 112639 (12);
Chinapa, UMMZ 119556 (2); Jazmin, UMMZ 114725 (2); between La
Playa and Volcán Jorullo, UMMZ 104748 (2); Limoncito, UMMZ 119552
(3); 14 km. S of Lombardia, KU 29299-301, 29303, 29305-11; 6 km. SW
of Nueva Italia, UMMZ 112640 (2); 2.7 km. S of Nueva Italia, UMMZ
112641 (4); 5 km. N of Nueva Italia, UMMZ 114721; Río Marquez, 10 km.
S of Lombardia, UMMZ 112642, 112646; Río Marquez, 13 km. SE of Nueva
[598]
Italia, UMMZ 114726; 6 km. N of Tafetan, UMMZ 119555 (18); 14.5 km.
E of Tepalcatepec, UMMZ 114720 (2); 6 km. S of Tzitzio, UMMZ 99199,
99200 (2); 19 km. S of Tzitzio, UMMZ 99154; Volcán Jorullo, UMMZ 104449
(4), 104750; Ziracuaretiro, UMMZ 114724; 3 km. NW of Zirimicuaro, UMMZ
114723.
Acknowledgments.—For the loan of specimens under their care I am indebted
to Doris M. Cochran, United States National Museum (USNM); Norman
Hartweg, University of Michigan Museum of Zoology (UMMZ); Robert
F. Inger, Chicago Natural History Museum (CNHM); and Hobart M. Smith,
University of Illinois Museum of Natural History (UIMNH). I thank Ann
S. Duellman, Richard E. Etheridge, Fred G. Thompson, Jerome B. Tulecke,
and John Wellman for their assistance in the field, Lorna Cordonnier for the
drawing reproduced as Figure 1, and Richard G. Zweifel for helpful suggestions
and criticism. Field work in México was made possible by grants from
the Penrose Fund of the American Philosophical Society and the Bache Fund
of the National Academy of Sciences in co-operation with the Museum of
Zoology of the University of Michigan.
Transmitted February 2, 1960.

28-2494